Antioxidant and free radical-scavenging activities of phenolic extracts of olive fruits Vasileios Ziogas a , Georgia Tanou b , Athanassios Molassiotis a, * , Grigorios Diamantidis b , Miltiadis Vasilakakis a a Laboratory of Pomology, School of Agriculture, Aristotle University of Thessaloniki, University Campus, GR-54124 Thessaloniki, Greece b Laboratory of Agricultural Chemistry, School of Agriculture, Aristotle University of Thessaloniki, University Campus, GR-54124 Thessaloniki, Greece article info Article history: Received 25 June 2009 Received in revised form 22 September 2009 Accepted 24 November 2009 Keywords: Altitude Antioxidant activity Anti-radical activity DNA nicking Genotype Maturation Olive fruit Phenols abstract The antioxidant ability of phenolic extracts of olive fruits during maturity in Chondrolia and Amfissis cul- tivars grown at 10 m and 300 m altitude showed that altitude affected phenol content, antioxidant and Fe 2+ /Fe 3+ reduction/binding ability. The hydroxyl radical, peroxyl radical and peroxynitrite-induced DNA nicking assays have been used to evaluate the anti-radical activity of the extracts. Results showed that the ability of olive extracts to prevent radical-mediated DNA damage arises from the triple synergis- tic action of the genotype, the altitude and the maturation stage. Ó 2009 Elsevier Ltd. All rights reserved. 1. Introduction Olea europaea is an emblematic species and one of the most widespread and economic important fruit trees in the Mediterra- nean basin. Olive and its different products are basic ingredients of the Mediterranean diet, which provide numerous health benefits (Blekas, Vassilakis, Harizanis, Tsimidou, & Boskou, 2002). The health-promoting properties of the Mediterranean diet have been largely attributed to the antioxidant and free radical-scavenging activity of polyphenols contained in the dietary components, espe- cially in olive fruit and oil (Kountouri, Mylona, Kaliora, & Andriko- poulos, 2007). During the ripening process remarkable changes have been observed in the phenolic contents (Amiot, Fleuriet, & Macheix, 1986; Bouaziz, Chamkha, & Sayadi, 2004; Esti, Cinquanta, & La Notte, 1998; Romero, Brenes, García, & Garrido, 2002). Cultivar and altitude are also important factors with regard to the phenolic content (Amiot, Fleuriet, & Macheix, 1989; Blekas et al., 2002; Jemai, Bouaziz, & Sayadi, 2009; Vinha et al., 2005). Cur- rently, olive culture spreads gradually in many non-Mediterranean regions with variable altitudes (Gutierrez, Ponti, & Cossu, 2009). In addition, climate warming, low rainfall and soil salinity are becom- ing the principal environmental problems for the olive industry (Jemai et al., 2009) and will drive olive tree cultivation to frost-free northern and high altitude areas (Gutierrez et al., 2009; Moriondo, Stefanini, & Bindi, 2008). Although the antioxidant activity of oil ol- ive phenolics has been under constant research for many years, the properties of table olives phenolics have not been studied to an equal extent. Additionally, there are no reports about the antioxi- dant profile of field-grown olive cultivars of Greek origin. The pres- ent study was designated to investigate the influence of altitude on the antioxidant and the free radical-scavenging potential of pheno- lic extracts of two Greek table olive cultivars, at different stages of maturation. 2. Materials and methods 2.1. Plant and sample material Two cultivars of table olive (Olea europaea L.) of Greek origin, namely Amfissis (Konservolia) and Chondrolia, were used in this study. Samples from both cultivars were collected from two olive groves at different altitudes. The first one was located in the experimental farm of the Aristotle University of Thessaloniki (Thessaloniki, Greece) at 10 m altitude, and the second in the vicinity of the city of Poligiros (Chalkidiki, Greece) at 300 m alti- tude. In each grove, five irrigated and healthy trees of the same age (15 years old) were used. The olive fruits (500 g in each 0308-8146/$ - see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.foodchem.2009.11.058 * Corresponding author. Tel./fax: +30 2310 998882. E-mail address: [email protected] (A. Molassiotis). Food Chemistry 120 (2010) 1097–1103 Contents lists available at ScienceDirect Food Chemistry journal homepage: www.elsevier.com/locate/foodchem

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Food Chemistry 120 (2010) 1097–1103
Contents lists available at ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Antioxidant and free radical-scavenging activities of phenolic extracts of olive fruits
Vasileios Ziogas a, Georgia Tanou b, Athanassios Molassiotis a,*, Grigorios Diamantidis b,Miltiadis Vasilakakis a
a Laboratory of Pomology, School of Agriculture, Aristotle University of Thessaloniki, University Campus, GR-54124 Thessaloniki, Greeceb Laboratory of Agricultural Chemistry, School of Agriculture, Aristotle University of Thessaloniki, University Campus, GR-54124 Thessaloniki, Greece
a r t i c l e i n f o a b s t r a c t
Article history:Received 25 June 2009Received in revised form 22 September2009Accepted 24 November 2009
Keywords:AltitudeAntioxidant activityAnti-radical activityDNA nickingGenotypeMaturationOlive fruitPhenols
0308-8146/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.foodchem.2009.11.058
* Corresponding author. Tel./fax: +30 2310 998882E-mail address: [email protected] (A. Molassi
The antioxidant ability of phenolic extracts of olive fruits during maturity in Chondrolia and Amfissis cul-tivars grown at 10 m and 300 m altitude showed that altitude affected phenol content, antioxidant andFe2+/Fe3+ reduction/binding ability. The hydroxyl radical, peroxyl radical and peroxynitrite-inducedDNA nicking assays have been used to evaluate the anti-radical activity of the extracts. Results showedthat the ability of olive extracts to prevent radical-mediated DNA damage arises from the triple synergis-tic action of the genotype, the altitude and the maturation stage.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Olea europaea is an emblematic species and one of the mostwidespread and economic important fruit trees in the Mediterra-nean basin. Olive and its different products are basic ingredientsof the Mediterranean diet, which provide numerous health benefits(Blekas, Vassilakis, Harizanis, Tsimidou, & Boskou, 2002). Thehealth-promoting properties of the Mediterranean diet have beenlargely attributed to the antioxidant and free radical-scavengingactivity of polyphenols contained in the dietary components, espe-cially in olive fruit and oil (Kountouri, Mylona, Kaliora, & Andriko-poulos, 2007). During the ripening process remarkable changeshave been observed in the phenolic contents (Amiot, Fleuriet, &Macheix, 1986; Bouaziz, Chamkha, & Sayadi, 2004; Esti, Cinquanta,& La Notte, 1998; Romero, Brenes, García, & Garrido, 2002).
Cultivar and altitude are also important factors with regard tothe phenolic content (Amiot, Fleuriet, & Macheix, 1989; Blekaset al., 2002; Jemai, Bouaziz, & Sayadi, 2009; Vinha et al., 2005). Cur-rently, olive culture spreads gradually in many non-Mediterraneanregions with variable altitudes (Gutierrez, Ponti, & Cossu, 2009). Inaddition, climate warming, low rainfall and soil salinity are becom-ing the principal environmental problems for the olive industry
ll rights reserved.
.otis).
(Jemai et al., 2009) and will drive olive tree cultivation to frost-freenorthern and high altitude areas (Gutierrez et al., 2009; Moriondo,Stefanini, & Bindi, 2008). Although the antioxidant activity of oil ol-ive phenolics has been under constant research for many years, theproperties of table olives phenolics have not been studied to anequal extent. Additionally, there are no reports about the antioxi-dant profile of field-grown olive cultivars of Greek origin. The pres-ent study was designated to investigate the influence of altitude onthe antioxidant and the free radical-scavenging potential of pheno-lic extracts of two Greek table olive cultivars, at different stages ofmaturation.
2. Materials and methods
2.1. Plant and sample material
Two cultivars of table olive (Olea europaea L.) of Greek origin,namely Amfissis (Konservolia) and Chondrolia, were used in thisstudy. Samples from both cultivars were collected from two olivegroves at different altitudes. The first one was located in theexperimental farm of the Aristotle University of Thessaloniki(Thessaloniki, Greece) at 10 m altitude, and the second in thevicinity of the city of Poligiros (Chalkidiki, Greece) at 300 m alti-tude. In each grove, five irrigated and healthy trees of the sameage (15 years old) were used. The olive fruits (500 g in each

1098 V. Ziogas et al. / Food Chemistry 120 (2010) 1097–1103
harvest) were randomly manually collected from all parts of theolive tree in three distinct stages of maturation as reflected byskin coloration (green, purple, black) and were immediatelystored at �20 �C. The sample collection of the olive fruits (begin-ning of September until end of November) was done according tothe colour of their skin, as this approach was considered to bemore suitable, based on the fact that different coloured fruitsare known to be chemically distinct, particularly with respect tophenolics (Amiot et al., 1986).
2.2. Antioxidant activity of olive phenols
The extraction of olive fruit samples was conducted as previ-ously reported by Blekas et al. (2002). The total phenolic contentof the extracts was determined by the reaction with Folin–Ciocal-teu reagent (Skerget et al., 2005). The total antioxidant activity(FRAP assay) was conducted according to Benzie and Strain(1996). The ability of phenolic extracts to exert antioxidant activitythrough iron chelation and reducing ability was investigated basedon the �OH-mediated deoxy-D-ribose degradation assay (Aruoma,1994).
2.3. Inhibitory effect of phenolic extracts on supercoiled DNA breakageinduced by hydroxyl radical (�OH)
The ability of olive phenolic (100 lV GAE) extracts to protectsupercoiled pBR322 plasmid DNA against �OH generated by Fen-ton’s reagent was estimated with the DNA nicking assay describedby Hu and Kitts (2001). Photographs were scanned by a Bio-Rad(Hercules, CA) imaging calibrated densitometer (GS-800), and theImage J software (National Institutes of Health, NIH; http://rsb.in-fo.nih.gov/ij/) was used to quantify DNA strand breaks, based onthe intensity of the supercoiled DNA. All tests were run in triplicateand averaged.
2.4. Inhibitory effect of phenolic extracts on supercoiled DNA breakageinduced by peroxyl radical
The experiment was conducted in a potassium phosphate buffer(10 mM, pH 7.4). Intact pBR322 plasmid DNA (0.5 lg) was mixedwith 100 lV GAE of phenol extract in phosphate-buffered saline.Two microlitres of 2,20-azobis(2-amidinopropane) dihydrochloride(AAPH) (from a 30 mM solution dissolved in 10 mM potassiumphosphate buffer, pH 7.4, 10 mM NaCl) were added to initiate thereaction (total volume 25 ll) (Lim, Hu, & Kitts, 2001). The mixturewas incubated for 2 h at 37� C before being applied to 0.7% (w/v)agarose gel. DNA bands were separated and analysed as describedabove.
2.5. Inhibitory effect of phenolic extracts on supercoiled DNA breakageinduced by peroxynitrite (ONOO�)
2.5.1. Synthesis of peroxynitriteA solution of 30% (approximately 9 M) H2O2 (0.22 ml) was di-
luted to 0.5 ml with water and chilled to 4 �C in an ice/water mix-ture; 0.4 ml of 5 N NaOH and 0.05 ml of 0.04 M DTPA in 0.05 NNaOH were added with gentle mixing, and then diluted to 1 ml to-tal volume (Uppu & Pryor, 1996). The concentration of H2O2 in thefinal solution was 2 M with the pH at 12.5–13.0. The buffered H2O2
solution (1 ml) was stirred vigorously with an equimolar amountof isoamyl nitrite (0.27 ml) for 10 h at 4 �C in a cold room. Afterincubation, the organic phase was removed and the aqueous phasewas washed six times with two volumes of hexane to eliminate thecontaminating isoamyl alcohol and isoamyl nitrite. The unreactedH2O2 was then removed by passing the aqueous phase through a0.5 � 5 cm column filled with 0.25 g of granular MnO2. The MnO2
column was washed twice with 2 ml each of water and 0.5 N NaOHbefore use. To minimize the dilution of peroxynitrite in the aque-ous phase, the first few microlitres of the peroxynitrite solutioneluting from the MnO2 column were discarded. The peroxynitriteconcentration was assayed by diluting the solution 1000-fold with0.1 N NaOH and then measured at 302 nm (e = 1670 M�1 cm�1).
2.5.2. Assay for ONOO�-induced DNA nickingIn a total volume of 25 ll, the ONOO� was added in a reaction
mixture containing 50 mM sodium phosphate buffer, pH 7.0,10 mM NaCl, 0.1 mM diethylenetriaminepentaacetic acid (DTPA),0.5 lg of intact pBR322 plasmid DNA (Barr & Gedamu, 2003) and100 lV gallic acid equivalents of phenol extract. After incubation(5 min) the DNA bands were separated on a 1.5% (w/v) agarosegel and analysed as described.
2.6. Data analysis
All results are the average of three measurements. Means werecompared for significant differences between treatments accordingto Duncan’s multiple range test at p < 0.05 using SPSS r.11.0.0 sta-tistical software (SPSS Inc., Chicago, IL). The correlation coefficients(r) were also calculated.
3. Results and discussion
3.1. Phenol content
The fruits of Chondrolia contained nearly double the amount ofphenols compared to Amfissis at both altitudes during all stagesof maturation (Table 1), indicating that the genotypes tested varygreatly in their capacity to synthesize phenolics. Since fruits fromthe Chondrolia cultivar reached black maturation stage prior tothat of the Amfissis fruit, the above differences could be also re-lated to the agronomic characteristics of the time of fruit ripening.There are also studies showing differences in the phenolic contentof olive fruit and virgin oils from different Italian (Esti et al., 1998)and Portuguese (Vinha et al., 2005) cultivars. Given the good bio-availability of phenolic compounds of Greek table olives (Kounto-uri et al., 2007), data indicate that olive fruit of Chondroliacultivar could be an interesting source of phenolics. A decreasein phenolic contents had been reported with ripening of olivefruits (Amiot et al., 1989). However, the amount of phenols inChondrolia increased at the black stage at both altitudes (Table1). Similarly, Bouaziz et al. (2004) found an increase in phenoliccontent of Chemlali olive cultivar at the last maturation period.In cultivar Amfissis, the phenolic content at green stage of matu-ration had a specific value that decreased at the purple stage atboth altitudes but increased in the black stage of maturation at300 m (Table 1). In contrast to Chondrolia, Amfissis olives in theblack stage of maturation had a higher amount of phenols whengrown at 300 m altitude, compared to olives at 10 m altitude(Table 1), demonstrating that factors related to altitude have a di-verse strong effect on the biosynthetic pathway of phenolics inthe cultivars tested. As reported previously (Bilger, Rolland, &Nybakken, 2007), enhanced UV-B radiation and lower tempera-ture at higher altitudes might contribute to the altitudinal geno-type-dependent difference observed in the phenols of the olivecultivars.
3.2. Total antioxidant activity
Recent health interest in olive phenols is related primarily totheir antioxidant-related activity (Kountouri et al., 2007). The val-ues of the total antioxidant activity of the phenolic extracts accord-
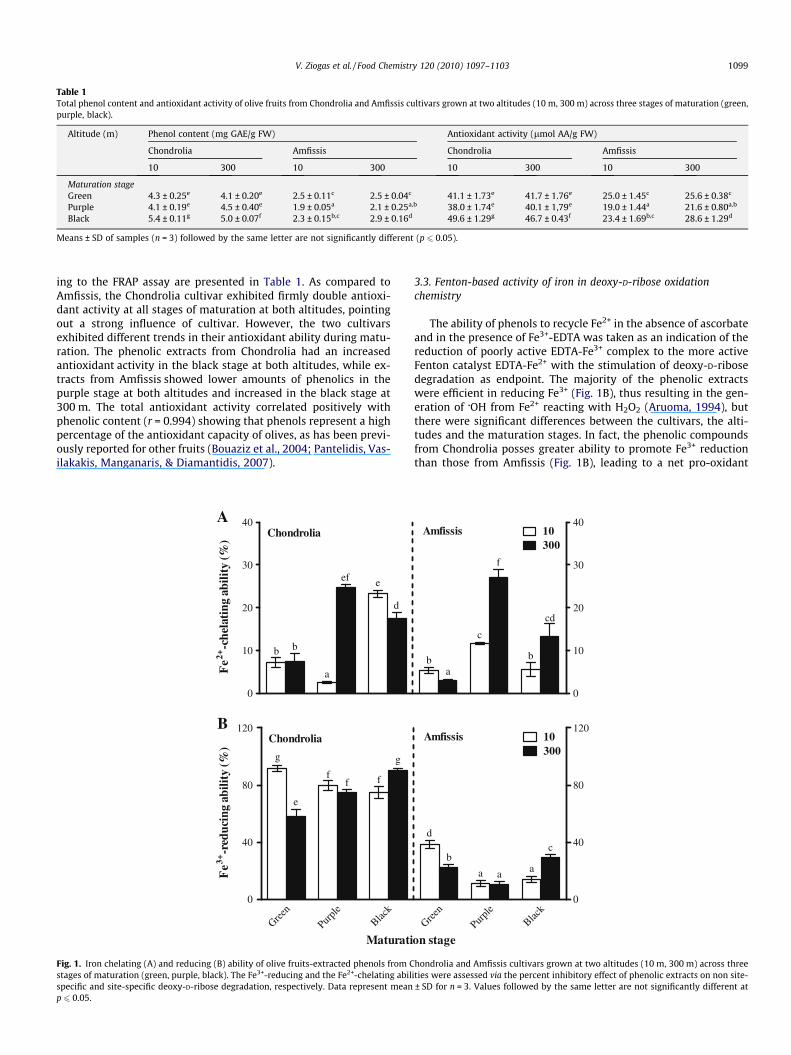
Table 1Total phenol content and antioxidant activity of olive fruits from Chondrolia and Amfissis cultivars grown at two altitudes (10 m, 300 m) across three stages of maturation (green,purple, black).
Altitude (m) Phenol content (mg GAE/g FW) Antioxidant activity (lmol AA/g FW)
Chondrolia Amfissis Chondrolia Amfissis
10 300 10 300 10 300 10 300
Maturation stageGreen 4.3 ± 0.25e 4.1 ± 0.20e 2.5 ± 0.11c 2.5 ± 0.04c 41.1 ± 1.73e 41.7 ± 1.76e 25.0 ± 1.45c 25.6 ± 0.38c
Purple 4.1 ± 0.19e 4.5 ± 0.40e 1.9 ± 0.05a 2.1 ± 0.25a,b 38.0 ± 1.74e 40.1 ± 1,79e 19.0 ± 1.44a 21.6 ± 0.80a,b
Black 5.4 ± 0.11g 5.0 ± 0.07f 2.3 ± 0.15b,c 2.9 ± 0.16d 49.6 ± 1.29g 46.7 ± 0.43f 23.4 ± 1.69b,c 28.6 ± 1.29d
Means ± SD of samples (n = 3) followed by the same letter are not significantly different (p 6 0.05).
V. Ziogas et al. / Food Chemistry 120 (2010) 1097–1103 1099
ing to the FRAP assay are presented in Table 1. As compared toAmfissis, the Chondrolia cultivar exhibited firmly double antioxi-dant activity at all stages of maturation at both altitudes, pointingout a strong influence of cultivar. However, the two cultivarsexhibited different trends in their antioxidant ability during matu-ration. The phenolic extracts from Chondrolia had an increasedantioxidant activity in the black stage at both altitudes, while ex-tracts from Amfissis showed lower amounts of phenolics in thepurple stage at both altitudes and increased in the black stage at300 m. The total antioxidant activity correlated positively withphenolic content (r = 0.994) showing that phenols represent a highpercentage of the antioxidant capacity of olives, as has been previ-ously reported for other fruits (Bouaziz et al., 2004; Pantelidis, Vas-ilakakis, Manganaris, & Diamantidis, 2007).
0
10
20
30
40Chondrolia
b b
a
ef e
d
Fe2+
-che
lati
ng a
bilit
y (%
)
Green
Purple
Black
0
40
80
120Chondrolia
g
e
ff f
g
Maturati
Fe3+
-red
ucin
g ab
ility
(%)
A
B
Fig. 1. Iron chelating (A) and reducing (B) ability of olive fruits-extracted phenols from Cstages of maturation (green, purple, black). The Fe3+-reducing and the Fe2+-chelating abilspecific and site-specific deoxy-D-ribose degradation, respectively. Data represent meanp 6 0.05.
3.3. Fenton-based activity of iron in deoxy-D-ribose oxidationchemistry
The ability of phenols to recycle Fe2+ in the absence of ascorbateand in the presence of Fe3+-EDTA was taken as an indication of thereduction of poorly active EDTA-Fe3+ complex to the more activeFenton catalyst EDTA-Fe2+ with the stimulation of deoxy-D-ribosedegradation as endpoint. The majority of the phenolic extractswere efficient in reducing Fe3+ (Fig. 1B), thus resulting in the gen-eration of �OH from Fe2+ reacting with H2O2 (Aruoma, 1994), butthere were significant differences between the cultivars, the alti-tudes and the maturation stages. In fact, the phenolic compoundsfrom Chondrolia posses greater ability to promote Fe3+ reductionthan those from Amfissis (Fig. 1B), leading to a net pro-oxidant
0
10
20
30
4010300
c
f
b
cd
ba
Amfissis
Green
Purple
Black
0
40
80
12010300
a a a
cd
b
Amfissis
on stage
hondrolia and Amfissis cultivars grown at two altitudes (10 m, 300 m) across threeities were assessed via the percent inhibitory effect of phenolic extracts on non site-± SD for n = 3. Values followed by the same letter are not significantly different at
1100 V. Ziogas et al. / Food Chemistry 120 (2010) 1097–1103
action (Hu, Zhang, & Kitts, 2000; Yen, Duh, & Tsai, 2002). Oxidativebreakdown of deoxy-D-ribose in iron-containing Fenton-type sys-tems as a result of reducing (pro-oxidant) activity of olive-derivedphenols has been reported (Rietjens, Bast, & Haenen, 2007). Recentevidence strongly suggests that the pro-oxidant action of phenolicsrather than their antioxidant activity might be an important mech-anism to exhibit their anticancer and apoptosis properties (Hadiet al., 2007). Interestingly, the Chondrolia which had higher phenolcontent than Amfissis (Table 1), showed also higher reducing abil-ity in converting Fe3+ to Fe2+ (Fig. 1B). A similar shift from antiox-idative effects at low phenol content to pro-oxidant effects at highphenol content has also been observed previously (Cao, Sofic, &Prior, 1997). It was further documented that both cultivars in thegreen stage at 10 m exhibited higher Fe3+-reducing ability com-pared with those at 300 m. However, no differences in the Fe-reducing potential of olive phenols were observed between thetwo altitudes at the purple maturation stage. Finally, olive samplescollected in the black stage at 300 m showed an increase of theirreducing capacity, compared to those obtained at 10 m. It is note-worthy that olive extracts show similar trends in both non site-specific deoxy-D-ribose (conducted at the physiological pH of 7.4)(Fig. 1B) and FRAP (conducted at the low pH of 3.6) (Table 1) as-says, particularly at 300 m altitude (r = 0.973). Since FRAP assayis also based on the ability of the tested extracts to act as reduc-tants in iron redox-linked reactions (Benzie & Strain, 1996), itmay be assumed that the olive fruit-derived phenols possess anti-
10 300 300 1010
Green
23,1309,4166,557
2,3222,027
Purple Bl
Chondrolia
3 4 5 6 721M
A
B
4,361
(bp)
Green
Purple
Black
0.0
0.5
1.0
1.5
2.0Chondrolia
bbc
c
dd
e
Matura
Inte
nsity
of i
ntac
t DN
A
Fig. 2. (A) Inhibitory effect of olive fruits-extracted phenols from Chondrolia and Amfismaturation (green, purple, black) in preventing DNA strand cleavage by �OH generated fpBR322 DNA (no Fenton reagents); lane 2 = Fenton reactants-induced DNA scission (noaltitude, respectively; Lanes 5 and 6 = extracts at the purple stage at 10 m and 300 m alaltitude, respectively. SC, supercoiled DNA; NC, open circular DNA (after a single-strand bmarkers. (B) Densitometric quantification of the intensity (in arbitrary units) of the SC frasignificantly different at p 6 0.05.
oxidant capacity through an electron donation mechanism over awide pH range.
In the deoxy-D-ribose assay, when EDTA is omitted from thesystem, iron ions are equally accessible to both the deoxy-D-riboseand the phenolic extracts under test. In that case some of the ironions bind to the deoxy-D-ribose molecule and the occurring dam-age to the sugar becomes site-specific, since the �OH that areformed, immediately attack the deoxy-D-ribose molecule. Thus, ex-tracts that are able to preferentially chelate Fe2+, rendering the me-tal complex in a less redox-active form, will protect the deoxy-D-ribose against degradation in the presence of Fenton reagents (Aru-oma, 2003). Under these conditions, the phenolics at the greenstage of maturation from both cultivars and altitudes possess a rel-atively weak binding activity for redox-active Fe2+ ions (Fig. 1A)and thus promote iron-dependent �OH generation at a particularsite (Moran, Klucas, Grayer, Abian, & Becana, 1997). Purple stage-derived extracts from both olives grown at 300 m exhibited con-siderably higher Fe2+-binding efficacies than those at 10 m(Fig. 1A), thereby diminishing free Fe2+ and thus preventing �OHformation via Fe2+-catalyzed Fenton chemistry. Iron-coordinationmechanism has also been identified as an important componentin characterizing polyphenol antioxidant activity (Moran et al.,1997). It has been suggested that iron binding to polyphenol com-pounds may result in an iron complex that cannot be reduced bycellular reductants to catalytically generate �OH (Perron, Hodges,Jenkins, & Brumaghim, 2008). Results further showed that the
300
ack
Amfissis
8
NC
NLSC
3 4 5 6 7 8
10 300 300 3001010
Green Purple Black
(m)
Green
Purple
Black
0.0
0.5
1.0
1.5
2.010 m
300 mAmfissis
ab
bc
d
e e
tion stage
sis cultivars grown at two different altitudes (10 m, 300 m) across three stages ofrom a Fenton-like reaction mixture (Fe3+-EDTA/ascorbic acid/H2O2). Lane 1 = intactphenolic extract); Lanes 3 and 4 = extracts at the green stage at 10 m and 300 m
titude, respectively; Lanes 7 and 8 = samples at the black stage at 10 m and 300 mreak); NL, linear DNA (after double-strand break); M, HindIII lambda digest DNA sizection. Data represent mean ± SD for n = 3. Values followed by the same letter are not
V. Ziogas et al. / Food Chemistry 120 (2010) 1097–1103 1101
black stage-derived phenols of Chondrolia at 10 m were moreeffective in chelating Fe2+ than the extracts obtained at 300 m;however the opposite altitude effect was observed in the corre-sponding samples of Amfissis (Fig. 1A). Overall, our findings estab-lish that genetic and environmental factors may strongly affect theFenton-related iron properties of phenols during olive fruitmaturation.
3.4. Anti-free radical specific activities
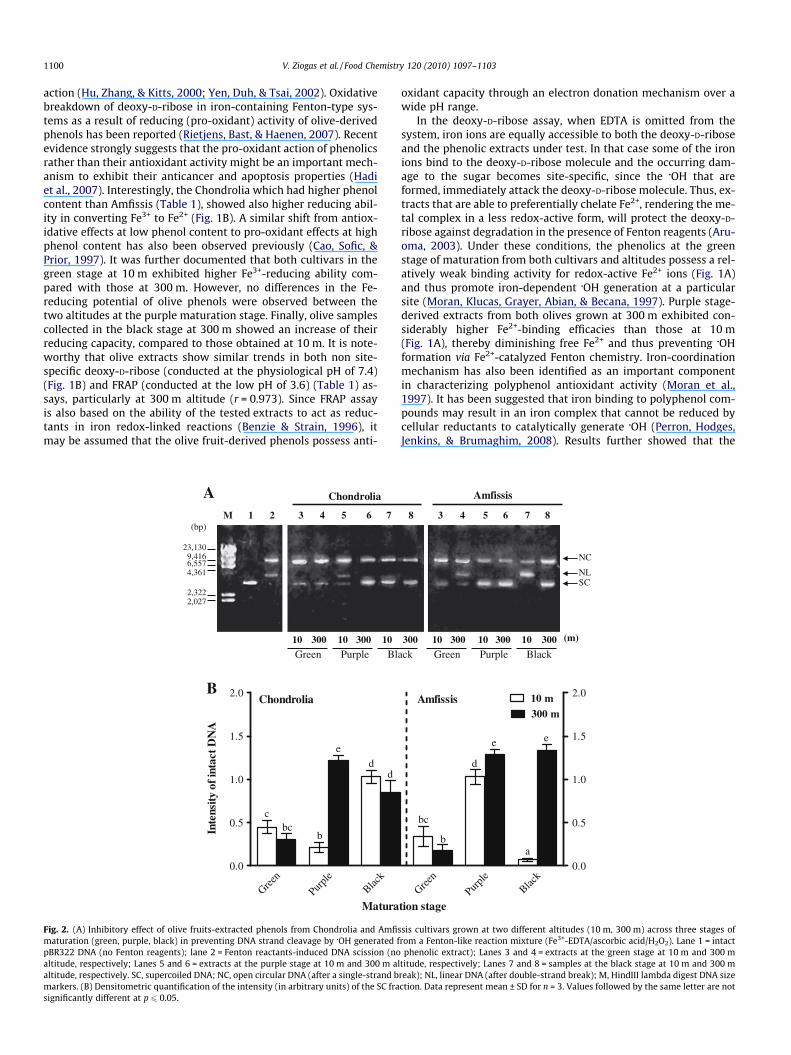
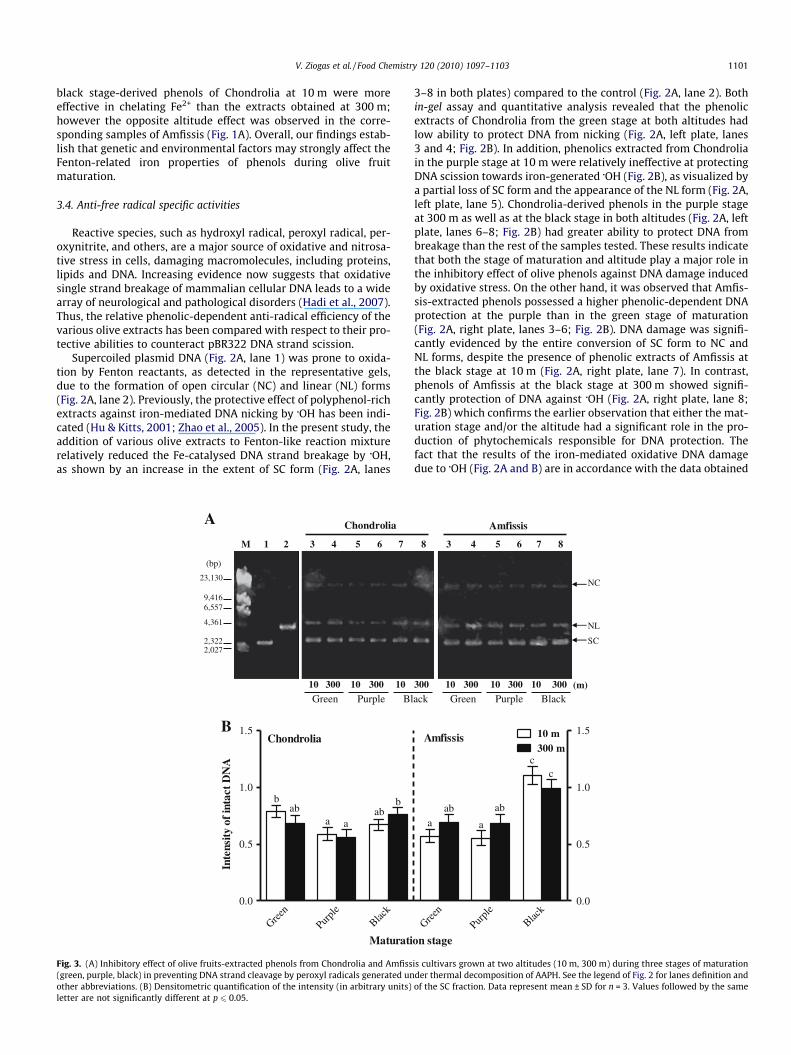
Reactive species, such as hydroxyl radical, peroxyl radical, per-oxynitrite, and others, are a major source of oxidative and nitrosa-tive stress in cells, damaging macromolecules, including proteins,lipids and DNA. Increasing evidence now suggests that oxidativesingle strand breakage of mammalian cellular DNA leads to a widearray of neurological and pathological disorders (Hadi et al., 2007).Thus, the relative phenolic-dependent anti-radical efficiency of thevarious olive extracts has been compared with respect to their pro-tective abilities to counteract pBR322 DNA strand scission.
Supercoiled plasmid DNA (Fig. 2A, lane 1) was prone to oxida-tion by Fenton reactants, as detected in the representative gels,due to the formation of open circular (NC) and linear (NL) forms(Fig. 2A, lane 2). Previously, the protective effect of polyphenol-richextracts against iron-mediated DNA nicking by �OH has been indi-cated (Hu & Kitts, 2001; Zhao et al., 2005). In the present study, theaddition of various olive extracts to Fenton-like reaction mixturerelatively reduced the Fe-catalysed DNA strand breakage by �OH,as shown by an increase in the extent of SC form (Fig. 2A, lanes
3 4 5 6 721M
Chondrolia
10 300 300 1010
Green Purple Bl
A
B
23,130
9,4166,557
2,3222,027
4,361
(bp)
Green
Purple
Black
0.0
0.5
1.0
1.5Chondrolia
aaab ab
bb
Maturati
Inte
nsity
of i
ntac
t DN
A
Fig. 3. (A) Inhibitory effect of olive fruits-extracted phenols from Chondrolia and Amfiss(green, purple, black) in preventing DNA strand cleavage by peroxyl radicals generated unother abbreviations. (B) Densitometric quantification of the intensity (in arbitrary units)letter are not significantly different at p 6 0.05.
3–8 in both plates) compared to the control (Fig. 2A, lane 2). Bothin-gel assay and quantitative analysis revealed that the phenolicextracts of Chondrolia from the green stage at both altitudes hadlow ability to protect DNA from nicking (Fig. 2A, left plate, lanes3 and 4; Fig. 2B). In addition, phenolics extracted from Chondroliain the purple stage at 10 m were relatively ineffective at protectingDNA scission towards iron-generated �OH (Fig. 2B), as visualized bya partial loss of SC form and the appearance of the NL form (Fig. 2A,left plate, lane 5). Chondrolia-derived phenols in the purple stageat 300 m as well as at the black stage in both altitudes (Fig. 2A, leftplate, lanes 6–8; Fig. 2B) had greater ability to protect DNA frombreakage than the rest of the samples tested. These results indicatethat both the stage of maturation and altitude play a major role inthe inhibitory effect of olive phenols against DNA damage inducedby oxidative stress. On the other hand, it was observed that Amfis-sis-extracted phenols possessed a higher phenolic-dependent DNAprotection at the purple than in the green stage of maturation(Fig. 2A, right plate, lanes 3–6; Fig. 2B). DNA damage was signifi-cantly evidenced by the entire conversion of SC form to NC andNL forms, despite the presence of phenolic extracts of Amfissis atthe black stage at 10 m (Fig. 2A, right plate, lane 7). In contrast,phenols of Amfissis at the black stage at 300 m showed signifi-cantly protection of DNA against �OH (Fig. 2A, right plate, lane 8;Fig. 2B) which confirms the earlier observation that either the mat-uration stage and/or the altitude had a significant role in the pro-duction of phytochemicals responsible for DNA protection. Thefact that the results of the iron-mediated oxidative DNA damagedue to �OH (Fig. 2A and B) are in accordance with the data obtained
8 3 4 5 6 7 8
10 300 300 3001010
Green Purple Black300
ack
NC
NL
SC
Amfissis
(m)
Green
Purple
Black
0.0
0.5
1.0
1.510 m300 m
Amfissis
a a
ab ab
cc
on stage
is cultivars grown at two altitudes (10 m, 300 m) during three stages of maturationder thermal decomposition of AAPH. See the legend of Fig. 2 for lanes definition andof the SC fraction. Data represent mean ± SD for n = 3. Values followed by the same
1102 V. Ziogas et al. / Food Chemistry 120 (2010) 1097–1103
by the site-specific deoxy-D-ribose assay (Fig. 1A), illustrates thestrength of the present experimental approach and provides proofthat the Fe2+-chelation ability by olive phenols could be consideredas a means of prevention of DNA oxidation. Previous studiesshowed that Fe2+ binding is a key mechanism for DNA damageinhibition by polyphenol compounds (Hadi et al., 2007; Perronet al., 2008). However, the ability of phenols extracts to inhibitDNA damage did not correlate with their Fe3+-reduction capacity,an observation that was also noted by Perron et al. (2008).
The in vitro peroxyl radical scavenging capacity of differentplant phenolics has been reported elsewhere (Hu & Kitts, 2001;Hu et al., 2000). However, the present study demonstrated thestrong effectiveness of olive extracts to inhibit peroxyl radical-in-duced DNA strand breakage. In fact, peroxyl radicals generatedby thermal decomposition of AAPH degrade DNA under our exper-imental conditions (Fig. 3A, lane 2) and the phenol extract of botholive cultivars provided protection against damage by AAPH per-oxyl radical (Fig. 3A, lanes 3–8 in both plates). The degree ofDNA protection against the radical flux generation from AAPHwas higher in Amfissis-extracted phenols at the black stage(Fig. 3A, right plate, lanes 7 and 8; Fig. 3B) whereas there wereno significant differences among the rest of the extracts tested(Fig. 3B).
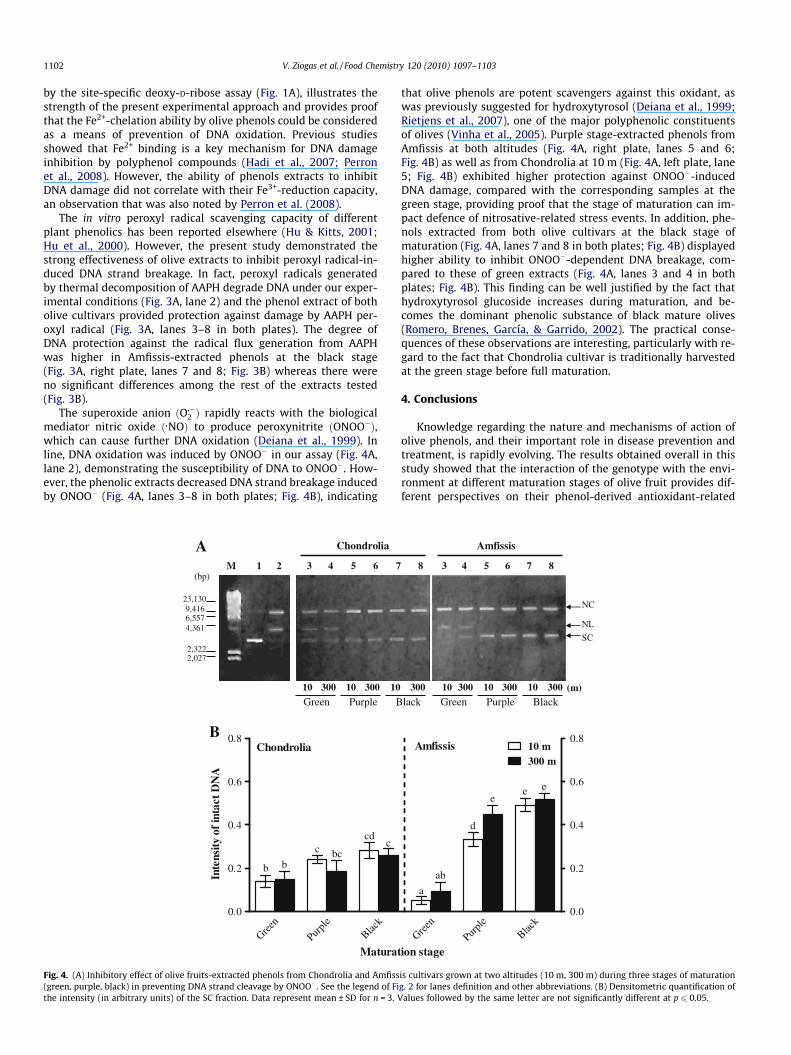
The superoxide anion ðO��2 Þ rapidly reacts with the biologicalmediator nitric oxide ð�NOÞ to produce peroxynitrite ðONOO�Þ,which can cause further DNA oxidation (Deiana et al., 1999). Inline, DNA oxidation was induced by ONOO� in our assay (Fig. 4A,lane 2), demonstrating the susceptibility of DNA to ONOO�. How-ever, the phenolic extracts decreased DNA strand breakage inducedby ONOO� (Fig. 4A, lanes 3–8 in both plates; Fig. 4B), indicating
Chondrolia
3 4 5 6 721M
10 300 300 1010
Green Purple B
A
B
Green
Purple
Black
0.0
0.2
0.4
0.6
0.8Chondrolia
b bbcc
cdc
Maturat
Inte
nsity
of i
ntac
t DN
A
(bp)
23,1309,4166,557
2,3222,027
4,361
Fig. 4. (A) Inhibitory effect of olive fruits-extracted phenols from Chondrolia and Amfiss(green, purple, black) in preventing DNA strand cleavage by ONOO� . See the legend of Fithe intensity (in arbitrary units) of the SC fraction. Data represent mean ± SD for n = 3. V
that olive phenols are potent scavengers against this oxidant, aswas previously suggested for hydroxytyrosol (Deiana et al., 1999;Rietjens et al., 2007), one of the major polyphenolic constituentsof olives (Vinha et al., 2005). Purple stage-extracted phenols fromAmfissis at both altitudes (Fig. 4A, right plate, lanes 5 and 6;Fig. 4B) as well as from Chondrolia at 10 m (Fig. 4A, left plate, lane5; Fig. 4B) exhibited higher protection against ONOO�-inducedDNA damage, compared with the corresponding samples at thegreen stage, providing proof that the stage of maturation can im-pact defence of nitrosative-related stress events. In addition, phe-nols extracted from both olive cultivars at the black stage ofmaturation (Fig. 4A, lanes 7 and 8 in both plates; Fig. 4B) displayedhigher ability to inhibit ONOO�-dependent DNA breakage, com-pared to these of green extracts (Fig. 4A, lanes 3 and 4 in bothplates; Fig. 4B). This finding can be well justified by the fact thathydroxytyrosol glucoside increases during maturation, and be-comes the dominant phenolic substance of black mature olives(Romero, Brenes, García, & Garrido, 2002). The practical conse-quences of these observations are interesting, particularly with re-gard to the fact that Chondrolia cultivar is traditionally harvestedat the green stage before full maturation.
4. Conclusions
Knowledge regarding the nature and mechanisms of action ofolive phenols, and their important role in disease prevention andtreatment, is rapidly evolving. The results obtained overall in thisstudy showed that the interaction of the genotype with the envi-ronment at different maturation stages of olive fruit provides dif-ferent perspectives on their phenol-derived antioxidant-related
8
300
lack
3 4 5 6 7 8
10 300 300 3001010
Green Purple Black
Amfissis
NC
NL
SC
Green
Purple
Black
0.0
0.2
0.4
0.6
0.810 m300 m
Amfissis
a
ab
e ee
d
ion stage
(m)
is cultivars grown at two altitudes (10 m, 300 m) during three stages of maturationg. 2 for lanes definition and other abbreviations. (B) Densitometric quantification ofalues followed by the same letter are not significantly different at p 6 0.05.

V. Ziogas et al. / Food Chemistry 120 (2010) 1097–1103 1103
properties. These results provide evidence about the phenolic-dependent biological function of olive fruits and may influenceconsumer preference and agricultural practice.
References
Amiot, M. J., Fleuriet, A., & Macheix, J. J. (1986). Importance and evolution ofphenolic compounds in olive during growth and maturation. Journal ofAgricultural and Food Chemistry, 34, 823–826.
Amiot, M. J., Fleuriet, A., & Macheix, J. J. (1989). Accumulation of oleuropeinderivatives during olive maturation. Phytochemistry, 28, 67–69.
Aruoma, O. X. (1994). Deoxyribose assay for detecting hydroxyl radicals. Methods inEnzymology, 233, 57–66.
Aruoma, O. (2003). Methodological consideration for characterizing potentialantioxidant actions of bioactive components in plant foods. Mutation Research– Fundamental and Molecular Mechanisms of Mutagenesis, 523–524, 9–20.
Barr, S. D., & Gedamu, L. (2003). Role of peroxidoxins in Leishmania chagasi survival:Evidence of an enzymatic defense against nitrosative stress. Journal of BiologicalChemistry, 278, 10816–10823.
Benzie, I. F. F., & Strain, J. J. (1996). The ferric reducing ability of plasma (FRAP) as ameasure of ‘‘antioxidant power”: The FRAP assay. Analytical Biochemistry, 239,70–76.
Bilger, W., Rolland, M., & Nybakken, L. (2007). UV screening in higher plants inducedby low temperature in the absence of UV-B radiation. Photochemical &Photobiological Sciences, 6, 190–195.
Blekas, G., Vassilakis, C., Harizanis, C., Tsimidou, M., & Boskou, D. (2002). Biophenolsin table olives. Journal of Agricultural and Food Chemistry, 50, 3688–3692.
Bouaziz, M., Chamkha, M., & Sayadi, S. (2004). Comparative study on phenoliccontent and antioxidant activity during maturation of the olive cultivarChemlali from Tunisia. Journal of Agricultural and Food Chemistry, 52,5476–5481.
Cao, G., Sofic, E., & Prior, P. L. (1997). Antioxidant and prooxidant behaviour offlavonoids: Structure–activity relationships. Free Radical Biology and Medicine,22, 749–760.
Deiana, M., Aruoma, O. I., Bianchi, M. D. L. P., Spencer, J. P. E., Kaur, H., Halliwell, B.,et al. (1999). Inhibition of peroxynitrite dependent DNA base modification andtyrosine nitration by the extra virgin olive oil-derived antioxidanthydroxytyrosol. Free Radical Biology and Medicine, 26, 762–769.
Esti, M., Cinquanta, L., & La Notte, E. (1998). Phenolic compounds in different olivevarieties. Journal of Agricultural and Food Chemistry, 46, 32–35.
Gutierrez, A. P., Ponti, L., & Cossu, Q. A. (2009). Effects of climate warming on Oliveand olive fly (Bactrocera oleae (Gmelin)) in California and Italy. Climatic Change,95, 195–217.
Hadi, S. M., Bhat, S. H., Azmi, A. S., Hanif, S., Shamim, U., & Ullah, M. F. (2007).Oxidative breakage of cellular DNA by plant polyphenols: A putativemechanism for anticancer properties. Seminars in Cancer Biology, 17, 370–376.
Hu, C., & Kitts, D. (2001). Evaluation of antioxidant activity of epigallocatehin gallatein biphasic model systems in vitro. Molecular and Cellular Biochemistry, 218,147–155.
Hu, C., Zhang, Y., & Kitts, D. D. (2000). Evaluation of antioxidant and prooxidantactivity of bamboo Phyllostachys nigra var. henonis leaf extract in vitro. Journal ofAgricultural and Food Chemistry, 48, 3170–3176.
Jemai, H., Bouaziz, M., & Sayadi, S. (2009). Phenolic composition, sugar contents andantioxidant activity of Tunisian sweet olive cultivar with regard to fruitripening. Journal of Agricultural and Food Chemistry, 57, 2961–2968.
Kountouri, A. M., Mylona, A., Kaliora, A. C., & Andrikopoulos, N. K. (2007).Bioavailability of the phenolic compounds of the fruits (drupes) of Oleaeuropaea (olives): Impact on plasma antioxidant status in humans.Phytomedicine, 14, 659–667.
Lim, K., Hu, C., & Kitts, D. D. (2001). Antioxidant activity of a Rhus verniciflua stokesethanol extract. Food and Chemical Toxicology, 39, 229–237.
Moran, J., Klucas, R., Grayer, R., Abian, J., & Becana, M. (1997). Complexes of ironwith phenolic compounds from soybean nodules and other legume tissues:Prooxidant and antioxidant properties. Free Radical Biology and Medicine, 22,861–870.
Moriondo, M., Stefanini, F. M., & Bindi, M. (2008). Reproduction of olive tree habitatsuitability for global change impact assessment. Ecological Modelling, 218,95–109.
Pantelidis, G., Vasilakakis, M., Manganaris, G., & Diamantidis, G. (2007). Antioxidantcapacity, phenol, anthocyanin and ascorbic acid contents in raspberries, redcurrants, gooseberries and Cornelian cherries. Food Chemistry, 102, 777–783.
Perron, N. R., Hodges, J. N., Jenkins, M., & Brumaghim, J. L. (2008). Predicting howpolyphenol antioxidants prevent DNA damage by binding to iron. InorganicChemistry, 47, 6153–6161.
Rietjens, S. J., Bast, A., & Haenen, G. R. M. M. (2007). New insights into controversieson the antioxidant potential of the olive oil antioxidant hydroxytyrosol. Journalof Agricultural and Food Chemistry, 55, 7609–7614.
Romero, C., Brenes, M., García, P., & Garrido, A. (2002). Hydroxytyrosol 4-b-D-glucoside, an important phenolic compound in olive fruits and derivedproducts. Journal of Agricultural and Food Chemistry, 50, 3835–3839.
Skerget, M., Kotnik, P., Hadolin, M., Hras, A., Simonic, M., & Knez, Z. (2005). Phenols,proanthocyanidins, flavones and flavonols in some plant materials and theirantioxidant activities. Food Chemistry, 89, 191–198.
Uppu, R. M., & Pryor, W. A. (1996). Synthesis of peroxynitrite in a two-phase systemusing isoamyl nitrite and hydrogen peroxide. Analytical Biochemistry, 236,242–249.
Vinha, A., Ferreres, F., Silva, B., Valentao, P., Goncalves, A., Pereira, J., et al. (2005).Phenolic profiles of Portuguese olive fruits (Olea europaea L): Influences ofcultivar and geographical origin. Food Chemistry, 89, 561–568.
Yen, G. C., Duh, P. D., & Tsai, H. L. (2002). Antioxidant and pro-oxidant properties ofascorbic acid and gallic acid. Food Chemistry, 79, 307–313.
Zhao, C., Dodin, G., Yuan, C., Chen, H., Zheng, R., Jia, Z., et al. (2005). ‘‘In vitro”protection of DNA from Fenton reaction by plant polyphenol verbascoside.Biochimica et Biophysica Acta, 1723, 114–123.
Related Documents











