Antioxidant and antimutagenic properties of the monoterpene indole alkaloid psychollatine and the crude foliar extract of Psychotria umbellata Vell. Variluska Fragoso a,1 , Naı ´la Cannes do Nascimento a,1 , Dinara Jaqueline Moura b , Ana Catarina Romano e Silva b , Marc Franc ßois Richter c , Jenifer Saffi b,d, * , Arthur Germano Fett-Neto a a Departamento de Bota ˆnica/Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, RS, Brazil b Departamento de Biofı ´sica/Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, RS, Brazil c Laborato ´ rio de Farmacocine ´tica, Centro de Pesquisa em Cie ˆncias Me ´dicas, Universidade Luterana do Brazil (ULBRA), Canoas, RS, Brazil d Laborato ´ rio de Gene ´tica Toxicolo ´ gica, Universidade Luterana do Brazil (ULBRA), Avenida Farroupilha 8001, Pre ´dio 01 Sala 122, Bairro Sa ˜o Jose ´, Canoas 92425-900, RS, Brazil Received 4 October 2007; accepted 9 November 2007 Available online 19 November 2007 Abstract Psychollatine is a monoterpene indole alkaloid produced and accumulated by Psychotria umbellata Vell. (Rubiaceae) leaves in rela- tively high amounts (approximately 3% of the dry weight). The alkaloid has been shown to display opioid-like analgesic, anxiolytic, anti- depressive and antipsychotic activities in rodents. In vitro assays suggested a protective role for this molecule in plant oxidative stress responses. This work reports antioxidant properties of psychollatine and the crude foliar extract from P. umbellata in strains of Saccha- romyces cerevisiae proficient and deficient in antioxidant defenses exposed to H 2 O 2 and paraquat. The antimutagenic activity of P. umbellata and its main alkaloid were assayed in S. cerevisiae N123 strain in presence of H 2 O 2 . Moreover, the antioxidant capacity of these substances on the hydroxyl radical (OH ) was investigated, using the hypoxanthine/xanthine oxidase assay. Psychollatine and the crude foliar extract of P. umbellata showed protective effect against oxidative stress in yeast, acting both as antioxidant and antimu- tagenic agents. Ó 2007 Elsevier Ltd. All rights reserved. Keywords: Psychollatine; Psychotria umbellata; Oxidative stress; Antimutagenicity; Saccharomyces cerevisiae 1. Introduction Monoterpene indole alkaloids (MIAs) represent a large and diverse class of plant alkaloids whose members occur mainly in Apocynaceae, Rubiaceae and Loganiaceae (O’Connor and Maresh, 2006). Ethnobotanical and chemotaxonomical studies on species of Psychotria, one of the largest and more complex genus from Rubiaceae, resulted in the discovery of a set of novel bioactive MIAs, some of them with clear pharmaceutical potential (Pasquali et al., 2006). Psychotria umbellata Vell. (Smith and Downs, 1956) is a shrub that reaches 1–3 m in height and grows in the tropi- cal and subtropical forests of Brazil; its southern limit is the restricted region of the northernmost Atlantic Forest rem- nants of the state of Rio Grande do Sul. The species accu- mulates relatively high amounts of an unusual MIA called 0887-2333/$ - see front matter Ó 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.tiv.2007.11.010 * Corresponding author. Address: Laborato ´rio de Gene ´tica Toxicolo ´g- ica, Universidade Luterana do Brazil (ULBRA), Avenida Farroupilha 8001, Pre ´dio 01 Sala 122, Bairro Sa ˜o Jose ´, Canoas 92425-900, RS, Brazil. Tel.: +55 51 34774000x2774; fax: +55 51 34779214. E-mail address: jenifer.saffi@ulbra.br (J. Saffi). 1 These authors contributed equally to this work. www.elsevier.com/locate/toxinvit Available online at www.sciencedirect.com Toxicology in Vitro 22 (2008) 559–566

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Available online at www.sciencedirect.com
www.elsevier.com/locate/toxinvit
Toxicology in Vitro 22 (2008) 559–566
Antioxidant and antimutagenic properties of the monoterpeneindole alkaloid psychollatine and the crude foliar extract
of Psychotria umbellata Vell.
Variluska Fragoso a,1, Naıla Cannes do Nascimento a,1, Dinara Jaqueline Moura b,Ana Catarina Romano e Silva b, Marc Franc�ois Richter c, Jenifer Saffi b,d,*,
Arthur Germano Fett-Neto a
a Departamento de Botanica/Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, RS, Brazilb Departamento de Biofısica/Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, RS, Brazil
c Laboratorio de Farmacocinetica, Centro de Pesquisa em Ciencias Medicas, Universidade Luterana do Brazil (ULBRA), Canoas, RS, Brazild Laboratorio de Genetica Toxicologica, Universidade Luterana do Brazil (ULBRA), Avenida Farroupilha 8001, Predio 01 Sala 122,
Bairro Sao Jose, Canoas 92425-900, RS, Brazil
Received 4 October 2007; accepted 9 November 2007Available online 19 November 2007
Abstract
Psychollatine is a monoterpene indole alkaloid produced and accumulated by Psychotria umbellata Vell. (Rubiaceae) leaves in rela-tively high amounts (approximately 3% of the dry weight). The alkaloid has been shown to display opioid-like analgesic, anxiolytic, anti-depressive and antipsychotic activities in rodents. In vitro assays suggested a protective role for this molecule in plant oxidative stressresponses. This work reports antioxidant properties of psychollatine and the crude foliar extract from P. umbellata in strains of Saccha-
romyces cerevisiae proficient and deficient in antioxidant defenses exposed to H2O2 and paraquat. The antimutagenic activity of P.
umbellata and its main alkaloid were assayed in S. cerevisiae N123 strain in presence of H2O2. Moreover, the antioxidant capacity ofthese substances on the hydroxyl radical (OH�) was investigated, using the hypoxanthine/xanthine oxidase assay. Psychollatine andthe crude foliar extract of P. umbellata showed protective effect against oxidative stress in yeast, acting both as antioxidant and antimu-tagenic agents.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Psychollatine; Psychotria umbellata; Oxidative stress; Antimutagenicity; Saccharomyces cerevisiae
1. Introduction
Monoterpene indole alkaloids (MIAs) represent a largeand diverse class of plant alkaloids whose members occurmainly in Apocynaceae, Rubiaceae and Loganiaceae
0887-2333/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.tiv.2007.11.010
* Corresponding author. Address: Laboratorio de Genetica Toxicolog-ica, Universidade Luterana do Brazil (ULBRA), Avenida Farroupilha8001, Predio 01 Sala 122, Bairro Sao Jose, Canoas 92425-900, RS, Brazil.Tel.: +55 51 34774000x2774; fax: +55 51 34779214.
E-mail address: [email protected] (J. Saffi).1 These authors contributed equally to this work.
(O’Connor and Maresh, 2006). Ethnobotanical andchemotaxonomical studies on species of Psychotria, oneof the largest and more complex genus from Rubiaceae,resulted in the discovery of a set of novel bioactive MIAs,some of them with clear pharmaceutical potential (Pasqualiet al., 2006).
Psychotria umbellata Vell. (Smith and Downs, 1956) is ashrub that reaches 1–3 m in height and grows in the tropi-cal and subtropical forests of Brazil; its southern limit is therestricted region of the northernmost Atlantic Forest rem-nants of the state of Rio Grande do Sul. The species accu-mulates relatively high amounts of an unusual MIA called

Fig. 1. Chemical structure of the monoterpene indole alkaloid psycholl-atine. Glc – glucose residue.
560 V. Fragoso et al. / Toxicology in Vitro 22 (2008) 559–566
psychollatine (Fig. 1), which has been shown to have anopioid-like analgesic effect in a dose-dependent manner,as well as anxiolytic, antidepressive, antipsychotic andamnesic effects, by modulating different neurotransmittersystems, including NMDA, opioid and serotonergic 5-HT2A/C receptors (Both et al., 2005,2006; Pasquali et al.,2006).
Psychollatine is restricted to shoots and its accumulationseems to be regulated by developmental stage. The depen-dence on differentiated shoots for psychollatine biosynthe-sis could be observed in plants regenerated by somaticembryogenesis. Psychollatine was absent in rhizogenic callibut, in shoots of differentiated plants derived from thesecalli, the alkaloid was found in amounts equivalent to thoseobserved in field grown individuals (Paranhos et al., 2005).
The most common source of free radicals of the cell inaerobic eukaryotic organisms is the respiratory chain ofmitochondria, since 2–5% of the total oxygen undergoesan incomplete reduction that generates reactive oxygenspecies (ROS) including superoxide anion (O��2 ), hydrogenperoxide (H2O2) and hydroxyl radicals (OH�) (Boveris,1998). In plants, another important source of ROS is thechloroplast (Edreva, 2005). ROS can attack important bio-logical macromolecules like DNA, proteins and lipid mem-branes. ROS toxicity, due to protein oxidation or to theproduction of lipid and carbohydrate derivatives that canreact with protein functional groups, has been involved indevelopment and progression of several diseases, such ascancer (Wiseman and Halliwell, 1996; Stadtman and Ber-lett, 1997; Halliwell and Gutteridge, 1998; Cyrne et al.,2003) Thus, the identification of new effective antioxidantsis a topic of interest; several plant extracts or secondarymetabolites have shown potential to protect against oxi-dant-induced damage (Borek, 2001; Zhang et al., 2006).Few reports have been published on antioxidant activitiesof the crude extracts or compounds isolated from Psycho-
tria (Saha et al., 2004), especially of MIAs displaying suchproperty. The potential protective role of psychollatineagainst oxidative stress was suggested by in vitro assaysthat showed action against singlet oxygen, superoxideand peroxide (Paranhos, J.T., Fragoso, V., Fett-Neto,A.G., unpublished data).
Considering the pharmacological properties of psycholl-atine, the present study was undertaken with a biologicalmodel (baker’s yeast) to obtain information on the antiox-
idant mechanism(s), antimutagenic properties and toxicityof the alkaloid to eukaryotic cells. The properties of puri-fied psychollatine and crude foliar extracts of P. umbellata
were investigated both in wild type and in mutant yeaststrains defective in endogenous antioxidant defenseproteins.
2. Materials and methods
2.1. Chemicals
Psychollatine was isolated from leaves of P. umbellata
(Both et al., 2002). Crude extracts were prepared with1 mg of dry leaves from P. umbellata extracted in 5 mLmethanol (HPLC grade), mixed and sonicated for 30 min.The extracts were centrifuged at 5000g for 10 min andthe supernatant was recovered and then dried for 5 h inSpeedVac not exceeding 35 �C, preventing possible degra-dation. The oxidant reagents hydrogen peroxide (H2O2)and paraquat (methyl viologen), amino acids, nitrogenatedbases, hypoxanthine, xanthine oxidase and salicylic acidwere purchased from Sigma (St. Louis, MO, USA). Yeastextract, bacto-peptone, bacto-agar and yeast nitrogen basewere obtained from Difco Laboratories (Detroit, MI). Allother reagents were of analytical grade.
For cells treatment, stock solutions of the crude extractand alkaloid were prepared immediately prior to use. In themutagenicity and antimutagenicity assays, dimethylsulfox-ide (DMSO) was used as solvent. The appropriate concen-trations were obtained by dilution of stock solution insterile distilled water and the final concentration of DMSOin the incubation mixture never exceeded 0.2% (v/v).
2.2. Saccharomyces cerevisiae strains and media
Strains of S. cerevisiae employed in this study aredescribed in Table 1. The antioxidant effects of crudeextract and purified alkaloid on cellular systems were eval-uated using strains of S. cerevisiae deficient in superoxidedismutase and/or catalase.
For mutagenic and antimutagenic assays we used thehaploid strain N123, which is very responsive to H2O2
induced mutagenesis since it shows a low glutathione con-tent (Brendel et al., 1998). Complete liquid medium (YPD)containing 1% yeast extract, 2% bacto-peptone and 2%dextrose was used for routine growth. Minimal medium(MM) contained 0.67% yeast nitrogen base without aminoacids, 2% dextrose, 0.5% ammonium sulfate. Solid mediacontained 2% bacto-agar. Synthetic complete medium(SC) was MM supplemented with appropriate amino acidsand nitrogenated bases (adenine, lysine, histidine, leucine,uracil, methionine and tryptophan) at a concentration of2.0 mg per 100 mL MM. For mutagenic assay SC mediumwas supplemented with 60 lg/mL canavanine. The herbi-cide paraquat, a redox cycling compound, was used toincrease the intracellular flux of superoxide anion (O��2 ).The appropriate concentrations of H2O2 and paraquat

Table 1Strains of Saccharomyces cerevisiae used in this study
Strain Genotype Source
EG103 (SOD-WT) MATa leu2-3 112his3-D1 trp1-289 ura3-52 E. Grallaa
EG118 (sod1D) Like EG103, except sod1::URA3 E. GrallaEG110 (sod2D) Like EG103, except sod2::TRP1 E. GrallaEG133 (sod1D sod2D) Like EG103, except sod1::URA3 and sod2::TRP1 E. GrallaEG223 (ctt1D) Like EG103, except ctt1::TRP1 E. GrallaEG (sod1D ctt1D) Like EG103, except sod1::URA3 and ctt1::TRP1 E. GrallaN123 MATa his1-7 J. Henriquesb
a Department of Chemistry and Biochemistry, University of California, Los Angeles 90024-1569, USA.b Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre 91501-970, Rio Grande do Sul, Brazil.
V. Fragoso et al. / Toxicology in Vitro 22 (2008) 559–566 561
were determined by survival assays, according to the differ-ential sensitivity of each strain. Sub-lethal concentrationsof the oxidants were used for all subsequent experiments.
Stationary phase cultures were obtained by inoculationof an isolated colony in liquid YPD medium. After 72 hincubation at 30 �C with aeration by shaking, the culturescontained (2–3) � 108 cells/ml. Cells in stationary phaseof growth were chosen to be used in this work becausethese resemble cells of multicellular organisms in importantaspects: (1) most energy comes from mitochondrial respira-tion, (2) cells are in the G0 phase, and (3) damage accumu-lates over time (Longo et al., 1996; Cyrne et al., 2003).
2.3. Growth inhibition (central disk assay)
Stationary phase cultures of yeast cells were diluted inPBS (phosphate buffered saline – Na2HPO4, KH2PO4
and KCl; 20 mM; pH 7.4) to an appropriate density of1–2 � 107 cells/mL for the growth inhibition assay. Aninoculation loop of cells from a cell suspension wasstreaked from the center to the border of a Petri dish inone continuous streak; growing concentration of theextracts and alkaloid (100, 250 and 500 lg/mL) wereplaced on a filter-paper disk in the center of the plate andpre-incubated for 4 h at 30 �C. Afterwards, 5 lL of 30%H2O2 or 8 lL of paraquat (500 mM) were placed on thesame filter-paper disk and incubated for 2 days at 30 �C.Impaired growth was measured as mm of growth inhibitionfrom the border of the filter-disk to the beginning of cellgrowth. Values could be within the range from 0 mm (com-plete growth to the filter-disk) to 30 mm (absence of growthto the rim of the Petri dish). Assays were repeated twotimes and plating for each dose was done in duplicate.
2.4. Mutagenicity and antimutagenicity in vivo assay
For the mutagenicity assay, cells in a density of 1–2 � 108 cells/mL were exposed to psychollatine or leafextract concentrations varying from 25 to 250 lg/mL andincubated for 1 h at 30 �C in term-mixer. For the antimuta-genic assay, the same procedure was taken but cells pluspsychollatine or crude foliar extract were also exposed tooxidative mutagenic agent (H2O2) and then the mixturewas further incubated. Suitable aliquots were plated in trip-
licate on solid SC with or without canavanine. Survival wasdetermined on SC (2–5 days at 30 �C) and mutation induc-tion (CAN revertants) on appropriate supplementationmedia (4–7 days at 30 �C). Surviving and revertant colonieswere counted. Forward mutation was measured with thecanavanine resistance assay (CAN1 ? can1) (Whelanet al., 1979) after induction with different treatments.Assays were repeated at least three times and plating foreach dose was done in triplicate.
2.5. Hipoxanthine/xanthine oxidase in vitro assay
The assay employed to determine antioxidant potentialof purified psychollatine and crude foliar extracts of P.
umbellata and hydroxyl radical scavenging was based onthe method described by Owen et al. (1996). Briefly, boththe extract and pure compound were diluted in the assaybuffer (hypoxanthine, Fe(III), EDTA and salicylic acid)at a concentration of 2.0 mg/mL and diluted appropriatelyin assay buffer to a final volume of 1.0 mL giving the spe-cific range, starting from 0.05 mg/mL. A 5.0 lL aliquotof xanthine oxidase (18 mU) dissolved in 3.2 M NH4SO4
was added to initiate the reaction. The sample tubes wereincubated for 3 h at 37 �C, at which time the reactionwas complete. A 30 lL aliquot of the reaction mixturewas analyzed by HPLC. Chromatographic analyses weredone using a gradient based on methanol/water/acetic acidwith a lBondaPak C18 reverse phase column (waters) anddetection at 325 nm. The HPLC equipment had a Waters2695 separation module and 2487 UV detector. Theamount of dihydroxyphenols (2,5-dihydroxibenzoic acidand 2,3-dihydroxibenzoic acid) produced by hydroxyl rad-ical (OH�) attack on salicylic acid was determined fromstandard curves generated with the respective pure dihydr-oxyphenols. Each time point was tested with three repli-cates and the experiment was independently repeated twice.
2.6. Statistics
Statistical analyses of the data were performed usingANOVA one-way and Dunnett’s multiple comparisontest. P-values equal or less than 0.05 were considered signif-icant. Data were expressed as means ± SD (standarddeviation).
562 V. Fragoso et al. / Toxicology in Vitro 22 (2008) 559–566
3. Results
Wild type cells (WT) and isogenic mutant strains of S.
cerevisiae lacking antioxidant defenses (Table 1) were trea-ted with solvent (50% DMSO) and several concentrationsof the alkaloids and crude foliar extract from P. umbellatafor 1 h, during the stationary phase. After 3 days of incuba-tion at 37 �C, all strains showed the same sensitivity to sub-stances as that observed for the WT cells grown in PBS andnone of the substances induced significant effects on yeastgrown in the employed concentrations (data not shown).Hence, we used the non cytotoxic concentration range ofalkaloid and extract from 25 to 500 lg/mL in the followingexperiments in order to investigate the protective activityagainst oxidants in the same strains.
3.1. Central disk assay
To examine the role of psychollatine and P. umbellata
crude foliar extract in cell oxidative stress, yeast cells dur-ing stationary phase were pre-treated with several non-cytotoxic concentrations of the alkaloid or extract for
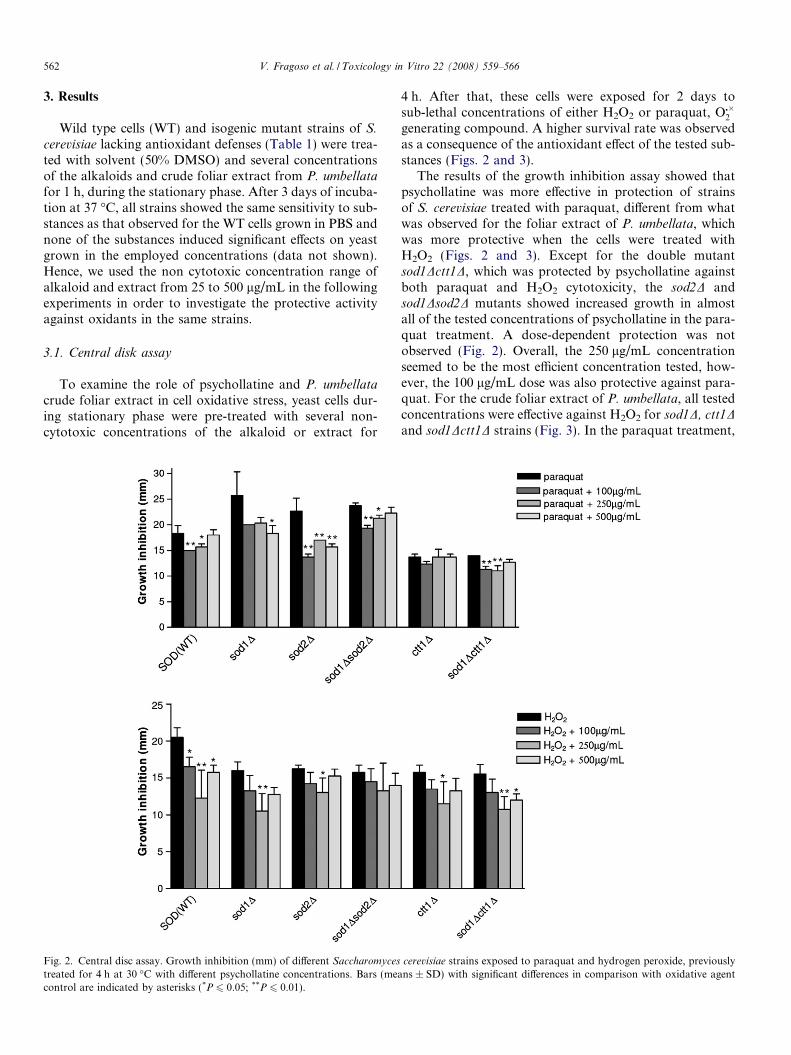
Fig. 2. Central disc assay. Growth inhibition (mm) of different Saccharomyces
treated for 4 h at 30 �C with different psychollatine concentrations. Bars (meacontrol are indicated by asterisks (*P 6 0.05; **P 6 0.01).
4 h. After that, these cells were exposed for 2 days tosub-lethal concentrations of either H2O2 or paraquat, O��2generating compound. A higher survival rate was observedas a consequence of the antioxidant effect of the tested sub-stances (Figs. 2 and 3).
The results of the growth inhibition assay showed thatpsychollatine was more effective in protection of strainsof S. cerevisiae treated with paraquat, different from whatwas observed for the foliar extract of P. umbellata, whichwas more protective when the cells were treated withH2O2 (Figs. 2 and 3). Except for the double mutantsod1Dctt1D, which was protected by psychollatine againstboth paraquat and H2O2 cytotoxicity, the sod2D andsod1Dsod2D mutants showed increased growth in almostall of the tested concentrations of psychollatine in the para-quat treatment. A dose-dependent protection was notobserved (Fig. 2). Overall, the 250 lg/mL concentrationseemed to be the most efficient concentration tested, how-ever, the 100 lg/mL dose was also protective against para-quat. For the crude foliar extract of P. umbellata, all testedconcentrations were effective against H2O2 for sod1D, ctt1Dand sod1Dctt1D strains (Fig. 3). In the paraquat treatment,
cerevisiae strains exposed to paraquat and hydrogen peroxide, previouslyns ± SD) with significant differences in comparison with oxidative agent
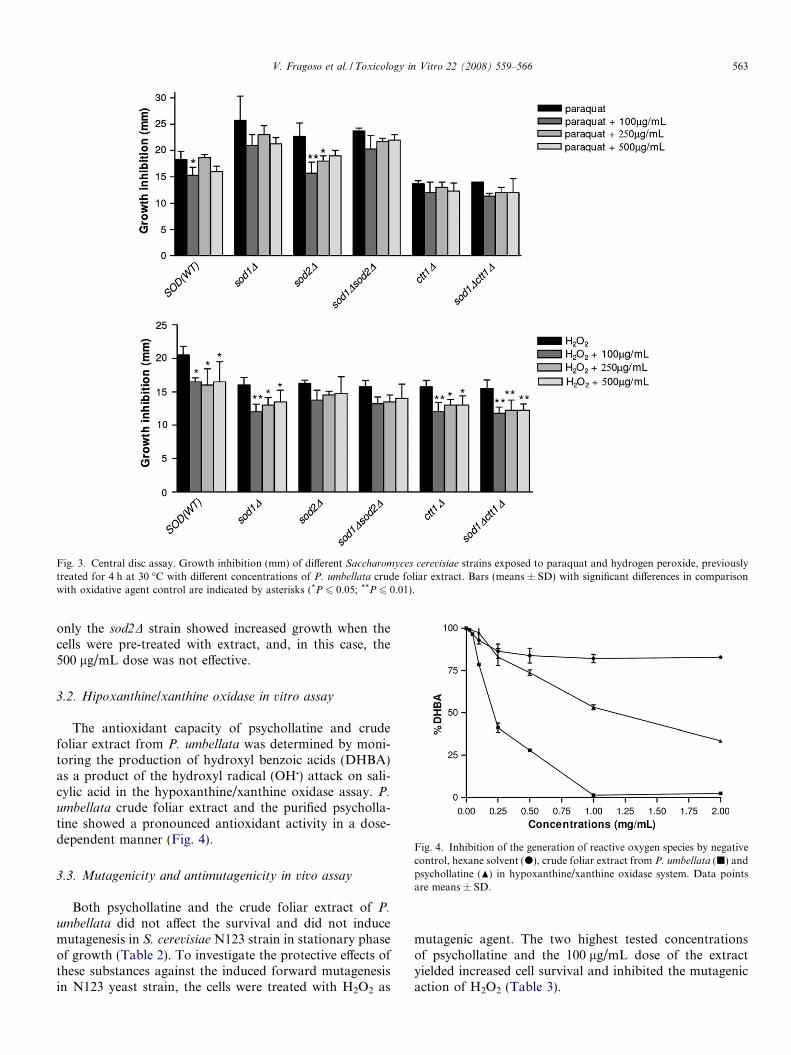
Fig. 3. Central disc assay. Growth inhibition (mm) of different Saccharomyces cerevisiae strains exposed to paraquat and hydrogen peroxide, previouslytreated for 4 h at 30 �C with different concentrations of P. umbellata crude foliar extract. Bars (means ± SD) with significant differences in comparisonwith oxidative agent control are indicated by asterisks (*P 6 0.05; **P 6 0.01).
V. Fragoso et al. / Toxicology in Vitro 22 (2008) 559–566 563
only the sod2D strain showed increased growth when thecells were pre-treated with extract, and, in this case, the500 lg/mL dose was not effective.
Fig. 4. Inhibition of the generation of reactive oxygen species by negative
3.2. Hipoxanthine/xanthine oxidase in vitro assay
The antioxidant capacity of psychollatine and crudefoliar extract from P. umbellata was determined by moni-toring the production of hydroxyl benzoic acids (DHBA)as a product of the hydroxyl radical (OH�) attack on sali-cylic acid in the hypoxanthine/xanthine oxidase assay. P.umbellata crude foliar extract and the purified psycholla-tine showed a pronounced antioxidant activity in a dose-dependent manner (Fig. 4).
control, hexane solvent (d), crude foliar extract from P. umbellata (j) andpsychollatine (N) in hypoxanthine/xanthine oxidase system. Data pointsare means ± SD.
3.3. Mutagenicity and antimutagenicity in vivo assay
Both psychollatine and the crude foliar extract of P.
umbellata did not affect the survival and did not inducemutagenesis in S. cerevisiae N123 strain in stationary phaseof growth (Table 2). To investigate the protective effects ofthese substances against the induced forward mutagenesisin N123 yeast strain, the cells were treated with H2O2 as
mutagenic agent. The two highest tested concentrationsof psychollatine and the 100 lg/mL dose of the extractyielded increased cell survival and inhibited the mutagenicaction of H2O2 (Table 3).
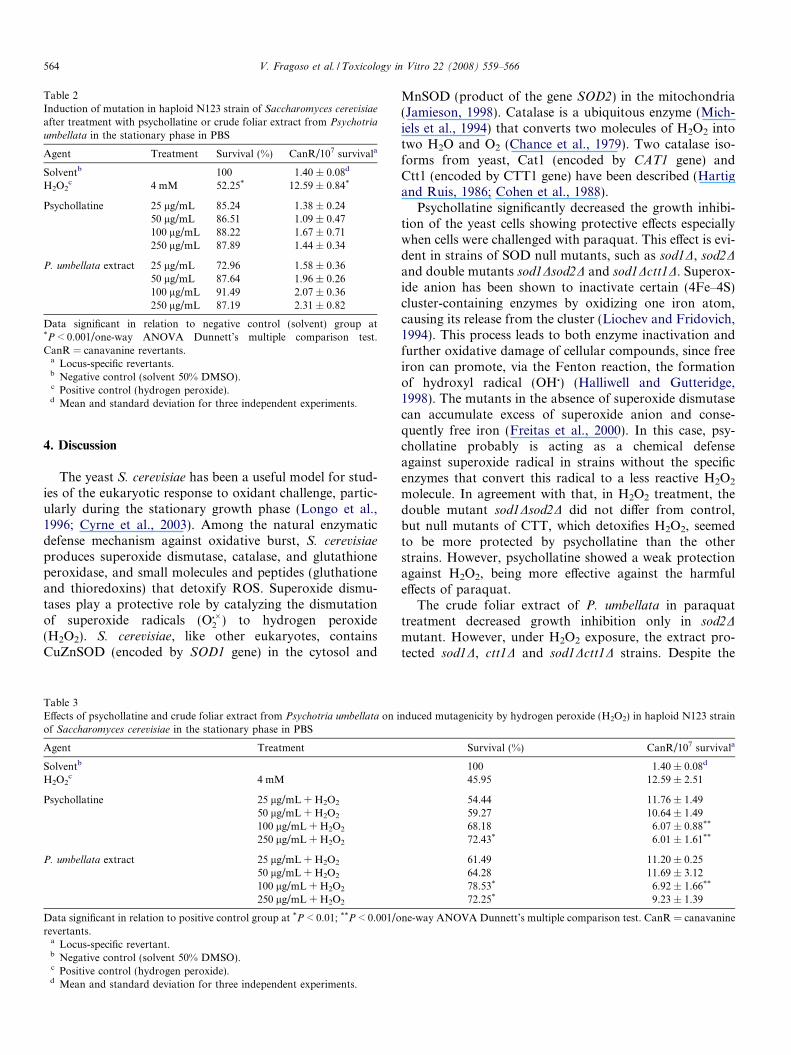
Table 2Induction of mutation in haploid N123 strain of Saccharomyces cerevisiae
after treatment with psychollatine or crude foliar extract from Psychotria
umbellata in the stationary phase in PBS
Agent Treatment Survival (%) CanR/107 survivala
Solventb 100 1.40 ± 0.08d
H2O2c 4 mM 52.25* 12.59 ± 0.84*
Psychollatine 25 lg/mL 85.24 1.38 ± 0.2450 lg/mL 86.51 1.09 ± 0.47100 lg/mL 88.22 1.67 ± 0.71250 lg/mL 87.89 1.44 ± 0.34
P. umbellata extract 25 lg/mL 72.96 1.58 ± 0.3650 lg/mL 87.64 1.96 ± 0.26100 lg/mL 91.49 2.07 ± 0.36250 lg/mL 87.19 2.31 ± 0.82
Data significant in relation to negative control (solvent) group at*P < 0.001/one-way ANOVA Dunnett’s multiple comparison test.CanR = canavanine revertants.
a Locus-specific revertants.b Negative control (solvent 50% DMSO).c Positive control (hydrogen peroxide).d Mean and standard deviation for three independent experiments.
564 V. Fragoso et al. / Toxicology in Vitro 22 (2008) 559–566
4. Discussion
The yeast S. cerevisiae has been a useful model for stud-ies of the eukaryotic response to oxidant challenge, partic-ularly during the stationary growth phase (Longo et al.,1996; Cyrne et al., 2003). Among the natural enzymaticdefense mechanism against oxidative burst, S. cerevisiaeproduces superoxide dismutase, catalase, and glutathioneperoxidase, and small molecules and peptides (gluthationeand thioredoxins) that detoxify ROS. Superoxide dismu-tases play a protective role by catalyzing the dismutationof superoxide radicals (O��2 ) to hydrogen peroxide(H2O2). S. cerevisiae, like other eukaryotes, containsCuZnSOD (encoded by SOD1 gene) in the cytosol and
Table 3Effects of psychollatine and crude foliar extract from Psychotria umbellata on iof Saccharomyces cerevisiae in the stationary phase in PBS
Agent Treatment
Solventb
H2O2c 4 mM
Psychollatine 25 lg/mL + H2O2
50 lg/mL + H2O2
100 lg/mL + H2O2
250 lg/mL + H2O2
P. umbellata extract 25 lg/mL + H2O2
50 lg/mL + H2O2
100 lg/mL + H2O2
250 lg/mL + H2O2
Data significant in relation to positive control group at *P < 0.01; **P < 0.001/orevertants.
a Locus-specific revertant.b Negative control (solvent 50% DMSO).c Positive control (hydrogen peroxide).d Mean and standard deviation for three independent experiments.
MnSOD (product of the gene SOD2) in the mitochondria(Jamieson, 1998). Catalase is a ubiquitous enzyme (Mich-iels et al., 1994) that converts two molecules of H2O2 intotwo H2O and O2 (Chance et al., 1979). Two catalase iso-forms from yeast, Cat1 (encoded by CAT1 gene) andCtt1 (encoded by CTT1 gene) have been described (Hartigand Ruis, 1986; Cohen et al., 1988).
Psychollatine significantly decreased the growth inhibi-tion of the yeast cells showing protective effects especiallywhen cells were challenged with paraquat. This effect is evi-dent in strains of SOD null mutants, such as sod1D, sod2Dand double mutants sod1Dsod2D and sod1Dctt1D. Superox-ide anion has been shown to inactivate certain (4Fe–4S)cluster-containing enzymes by oxidizing one iron atom,causing its release from the cluster (Liochev and Fridovich,1994). This process leads to both enzyme inactivation andfurther oxidative damage of cellular compounds, since freeiron can promote, via the Fenton reaction, the formationof hydroxyl radical (OH�) (Halliwell and Gutteridge,1998). The mutants in the absence of superoxide dismutasecan accumulate excess of superoxide anion and conse-quently free iron (Freitas et al., 2000). In this case, psy-chollatine probably is acting as a chemical defenseagainst superoxide radical in strains without the specificenzymes that convert this radical to a less reactive H2O2
molecule. In agreement with that, in H2O2 treatment, thedouble mutant sod1Dsod2D did not differ from control,but null mutants of CTT, which detoxifies H2O2, seemedto be more protected by psychollatine than the otherstrains. However, psychollatine showed a weak protectionagainst H2O2, being more effective against the harmfuleffects of paraquat.
The crude foliar extract of P. umbellata in paraquattreatment decreased growth inhibition only in sod2Dmutant. However, under H2O2 exposure, the extract pro-tected sod1D, ctt1D and sod1Dctt1D strains. Despite the
nduced mutagenicity by hydrogen peroxide (H2O2) in haploid N123 strain
Survival (%) CanR/107 survivala
100 1.40 ± 0.08d
45.95 12.59 ± 2.51
54.44 11.76 ± 1.4959.27 10.64 ± 1.4968.18 6.07 ± 0.88**
72.43* 6.01 ± 1.61**
61.49 11.20 ± 0.2564.28 11.69 ± 3.1278.53* 6.92 ± 1.66**
72.25* 9.23 ± 1.39
ne-way ANOVA Dunnett’s multiple comparison test. CanR = canavanine

V. Fragoso et al. / Toxicology in Vitro 22 (2008) 559–566 565
fact that the extract did not protect the double mutantsod1Dsod2D, for sod1D and sod2D mutants there seems tobe a certain prevalence of extract protection againstH2O2 rather than paraquat, perhaps related to SOD1 andSOD2 subcellular localization and/or extract uptake bycells. The 100 lg/mL dose seemed to be the most efficientconcentration of extract tested, followed by the 250 lg/mL dose.
The capacity of alkaloid and crude extract to scavengeOH� radicals was confirmed in the in vitro hypoxantine/xantine oxidase assay (Fig. 4). The extract showed a nota-ble in vitro OH� scavenging activity and was apparentlymore protective against H2O2 damages than psychollatinein S. cerevisiae (Figs. 2 and 3). It is known that P. umbel-lata leaves have about 2.3 mg of total flavonoids/g of dryweight (Correa, L.R., Fett-Neto, A.G., personal communi-cation), and these substances can improve antioxidantproperties of the extract (Rice-Evans et al., 1996). More-over, it is possible that psychollatine acts more as a O��2scavenger than as a H2O2 scavenger. The possible protec-tive role of psychollatine against ROS may be explainedby the presence of two secondary amines, double bondsand the glucose residue in its structure (Fig. 1) (Gregianiniet al., 2003).
As previously pointed out, DNA can be a target of ROSaction. Oxidative lesions in DNA include base modifica-tions, sugar damage, strand breaks and abasic site forma-tion. Antimutagenic activity of substances derived fromplants may be due to a variety of mechanisms such as inhi-bition of genotoxic effects, signal transduction modulation,antioxidant activity and scavenging of free radicals (DeFl-ora et al., 1999; Mantle et al., 2000). To test the protectingaction of P. umbellata foliar extract and psychollatine onH2O2-induced mutagenesis we used the N123 strain of S.
cerevisiae. Both the extract and alkaloid showed a pro-nounced antimutagenic effect at higher concentrations(Table 3). Interestingly, these same doses have shown tobe efficient against paraquat and H2O2-induced stress inthe central disc assay. The antioxidant activity and OH�
scavenging activity may contribute to the antimutageniceffect. These findings suggest new pharmacological proper-ties for psychollatine and the leaf extract of P. umbellata.
Conflict of Interest Statement
There are no conflicts of interest involved in this study.
Acknowledgments
This study was supported by a Research Grant from theNational Council for Scientific and Technological Devel-opment (CNPq – Conselho Nacional de DesenvolvimentoCientıfico e Tecnologico, Brazil) and student bursariesfrom the Brazilian Federal Agency for Support and Evalu-ation of Graduate Education (CAPES – Coordenac�ao deAperfeic�oamento de Pessoal de Nıvel Superior) and CNPq.We are also grateful to Dr. E. Gralla (Department of
Chemistry and Biochemistry UCLA) and Dr. J.A.P. Henr-iques (Centro de Biotecnologia, UFRGS) for the gift ofyeast strains.
References
Borek, C., 2001. Antioxidant health effects of aged garlic extract. TheJournal of Nutrition 131 (3S), 1010S–1015S.
Both, F.L., Kerber, V.A., Henriques, A.T., Elisabetsky, E., 2002.Analgesic properties of umbellatine from Psychotria umbellata. Phar-maceutical Biology 40, 336–341.
Both, F.L., Meneghini, L., Kerber, V.A., Henriques, A.T., Elisabetsky,E.J., 2005. Psychopharmacological profile of the alkaloid psychollatineas a 5HT2A/C serotonin modulator. Journal of Natural Products 68(3), 374–380.
Both, F.L., Meneghini, L., Kerber, V.A., Henriques, A.T., Elisabetsky,E.J., 2006. Role of glutamate and dopamine receptors in thepsychopharmacological profile of the indole alkaloid psychollatine.Journal of Natural Products 69 (3), 342–345.
Boveris, A., 1998. Biochemistry of free radicals, from electrons to tissues.Medicina 58, 350–356.
Brendel, M., Grey, M., Maris, A., Hietkamp, J., Fesus, Z., Pich, C., Dafre,L., Schmidt, M., Eckardt-Schupp, F., Henriques, J.A.P., 1998. Lowglutathione pools in the original pso3 mutant of Saccharomyces
cerevisiae are responsible for its pleiotropic sensitivity phenotype.Current Genetics 33 (1), 4–9.
Chance, B., Sies, H., Boveris, A., 1979. Hydroperoxide metabolism inmammalian organs. Physiological Reviews 59 (3), 527–605.
Cohen, G., Rapatz, W., Ruis, H., 1988. Sequence of the Saccharomyces
cerevisiae CTA1 gene and amino acid sequence of catalase A derivedfrom it. European Journal of Biochemistry 176 (1), 159–163.
Cyrne, L., Martins, L., Fernandes, L., Marinho, H.S., 2003. Regulation ofantioxidant enzymes gene expression in the yeast Saccharomyces
cerevisiae during stationary phase. Free Radical Biology & Medicine34 (3), 385–393.
DeFlora, S., Bennicelli, C., Bagnasco, M., 1999. Rationale and mecha-nisms of cancer chemoprevention. Recent Results in Cancer Research151, 29–44.
Edreva, A., 2005. Generation and scavenging of reactive oxygen species inchloroplasts: a submolecular approach. Agriculture, Ecosystems andEnvironment 106, 119–133.
Freitas, J.M., Liba, A., Meneghini, R., Valentine, J.S., Gralla, E.B., 2000.Yeast lacking Cu–Zn superoxide dismutase show altered iron homeo-stasis. The Journal Biological Chemistry 275, 11645–11649.
Gregianini, T.S., Da Silveira, V.C., Porto, D.D., Kerber, V.A., Henriques,A.T., Fett-Neto, A.G., 2003. The alkaloid brachycerine is induced byultraviolet radiation and is a singlet oxygen quencher. Photochemistryand Photobiology 78 (5), 470–474.
Halliwell, B., Gutteridge, J.M.C., 1998. Free Radicals in Biology andMedicine, third ed. Oxford University Press, New York, p. 980.
Hartig, A., Ruis, H., 1986. Nucleotide sequence of the Saccharomyces
cerevisiae CTT1 gene and deduced amino-acid sequence of yeastcatalase T. European Journal of Biochemistry 160 (3), 487–490.
Jamieson, D.J., 1998. Oxidative stress responses of the yeast Saccharo-
myces cerevisiae. Yeast 14, 1511–1527.Liochev, S.I., Fridovich, I., 1994. The role of O��2 in the production of
OH�: in vitro and in vivo. Free Radical Biology & Medicine 16 (1), 29–33.
Longo, V.D., Gralla, E.B., Valentine, J.S., 1996. Superoxide dismutaseactivity is essential for stationary phase survival in Saccharomyces
cerevisiae. The Journal of Biological Chemistry 271 (21), 12275–12280.Mantle, D., Lennard, T.W., Pickering, A.T., 2000. Therapeutic applica-
tions of medicinal plants in the treatment of breast cancer: a review oftheir pharmacology, efficacy and tolerability. Adverse Drug Reactionsand Toxicology Reviews 19, 223–240.
Michiels, C., Raes, M., Toussaint, O., Remacle, J., 1994. Importance ofSe-glutathione peroxidase, catalase, and Cu/Zn-SOD for cell survival

566 V. Fragoso et al. / Toxicology in Vitro 22 (2008) 559–566
against oxidative stress. Free Radical Biology & Medicine 17 (3), 235–248.
O’Connor, S.E., Maresh, J.J., 2006. Chemistry and biology of monoter-pene indole alkaloid biosynthesis. Natural Products Reports 23 (4),532–547.
Owen, R.W., Wimonwatwatee, T., Spiegelhalder, B., Bartsch, H., 1996. Ahigh performance liquid chromatography method for quantification ofhydroxyl radical formation by determination of dihydroxy benzoicacids. European Journal of Cancer Prevention 5 (4), 233–240.
Paranhos, J.T., Fragoso, V., Henriques, A.T., Ferreira, A.G., Fett-Neto,A.G., 2005. Regeneration of Psychotria umbellata and production ofthe analgesic indole alkaloid umbellatine. Tree Physiology 25, 251–255.
Pasquali, G., Porto, D.D., Fett-Neto, A.G., 2006. Metabolic engineeringof cell cultures versus whole plant complexity in production ofbioactive monoterpene indole alkaloids: recent progress related to olddilemma. Journal of Bioscience and Bioengineering 101 (4), 287–296.
Rice-Evans, C.A., Miller, N.J., Paganga, G., 1996. Structure-antioxidantactivity relationships of flavonoids and phenolic acids. Free RadicalBiology & Medicine 20, 933–956.
Saha, K., Lajis, N.H., Israf, D.A., Hamzah, A.S., Khozirah, S., Khamis,S., Syahida, A., 2004. Evaluation of antioxidant and nitric oxideinhibitory activities of selected Malaysian medicinal plants. Journal ofEthnopharmacology 92 (2–3), 263–267.
Smith, L.B., Downs, R.J., 1956. Resumo preliminar das Rubiaceas deSanta Catarina. Sellowia 7, 13–86.
Stadtman, E.R., Berlett, B.S., 1997. Reactive oxygen-mediated proteinoxidation in aging and disease. Chemical Research in Toxicology 10,485–494.
Whelan, W.L., Gocke, E., Manney, T.R., 1979. The CAN1 locus ofSaccharomyces cerevisiae: fine-structure analysis and forward mutationrates. Genetics 91, 35–51.
Wiseman, H., Halliwell, B., 1996. Damage to DNA by reactive oxygenand nitrogen species: role in inflammatory disease and progression tocancer. The Biochemical Journal 313, 17–29.
Zhang, W., Guo, Y.W., Gu, Y., 2006. Secondary metabolites from theSouth China Sea invertebrates: chemistry and biological activity.Current Medicinal Chemistry 13 (17), 2041–2090.
Related Documents











