PDFlib PLOP: PDF Linearization, Optimization, Protection Page inserted by evaluation version www.pdflib.com – [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PDFlib PLOP: PDF Linearization, Optimization, Protection
Page inserted by evaluation versionwww.pdflib.com – [email protected]

Antigen-Specific Tolerance Induced by Autoimmunization with Photoinactivated
Syngeneic Effector Cells" LILIANE LAROCHE,b*' RICHARD L. EDELSON, MARITZA PEREZ, AND CAROLE L. BERGERb
bDepartrnents of Dermatology and Pathology Columbia University
New York, New York 10032 and
Department of Dermatology Yale University
New Haven, Connecticut 06510
INTRODUCTION
Clinically relevant immunosuppression, induced by cytotoxic drugs, lymphoid irradiation, anti-lymphocyte globulin and thoracic duct drainage, unavoidably in- troduces nonspecific immunologic effects along with the more desirable inhibition of the particular T cell clones responsible for deleterious reaction^.'-^ A more attractive immunosuppressive therapy would preferentially target those antigen- specific T cells committed to participate in the pathologic process. Such effector T cells have membrane receptors specific to the antigen in association with major histocompatibility complex molecules (reviewed in REF. 4). These T cell antigen receptors are distinctive protein heterodimers which may themselves serve as clone-specific immunogens under certain circumstances5 and are, therefore, po- tential targets for directed therapy.
The induction of tolerance by anti-receptor autoimmunity has been reported in several in uivo experimental models. Anderson et al . have demonstrated that specific unresponsiveness to transplantation antigens may be induced by injection of alloantigen specific blasts generated in vitro.6 These results have been extended to vaccination with inactivated effector T cell clones which mediate autoimmune disease.' In these systems, effector cells alone were not immunogenic and re- quired modification or inactivation in vitro prior to infusion into recipients. Meth- ods that potentiated immunogenicity of these T cells included the use of complete Freund's adjuvant or X-irradiati~n.'-~ These protocols led to specific tolerance in naive immunocompetent recipients. Unfortunately, the requirement for clonal
a L.L. was a recipient of an Association pour la Recherche contre le Cancer fellowship. M.P. was a National Institutes of Health (NIH) postdoctoral trainee (5-T-32-ARO). This work was supported by grants from the NIH (CA 20499 and CA 39700), the Matheson Foundation, the Dermatology Foundation (6-41497) and Therakos, Inc.
Present address: Department of Dermatology, HBpital Avicenne, University of Paris XIII, 125, rue de Stalingrad, 93000 Bobigny, France.
113

114 ANNALS NEW YORK ACADEMY OF SCIENCES
expansion of the specific population or the use of adjuvants severely limits their clinical applicability.
A method that may provide a more clinically acceptable means for immune manipulation has been used in the treatment of disseminated cutaneous T cell lymphoma. It consists of the readministration of 8-methoxypsoralen (8-MOP) ultraviolet A (UVA) light-treated autologous pheresis-enriched leukocytes and resulted in profound amelioration and prolonged attenuation or disappearance of the disease process. lo The long-term clinical improvement, seen in several pa- tients, suggested a host response to distinct antigens expressed on reinfused photomodified lymphocytes, possibly rendered more immunogenic by the proce- dure. It is noteworthy that these clinical responses were achieved in the absence of in uitro manipulations of the patients' cells and were associated with quite limited adverse effects. Based on this clinical trial, we have developed animal models to determine: 1) whether 8-MOP-UVA inactivation of effector T cells provides a means of inducing an autoregulatory host response and 2) whether an immunocompetent host can select a markedly expanded relevant effector T cell population from an unfractionated leukocyte pool. In this context, we asked whether photoinactivated antigen-specific autologous cells can be used as immu- nogens to render a naive, immunocompetent individual unresponsive to subse- quent exposure to the relevant antigen.
We report here that intravenous injections of sheep red blood cell (SRBC) reactive cells from an unfractionated spleen pool photoinactivated in uitro can induce a state of selected unresponsiveness to SRBC and suppress an already existing immunity to that antigen. Identical pretreatment of splenocytes from unsensitized mice did not have this effect, indicating that light and photoinactived drug alone are ineffective in this system.
MATERIALS AND METHODS
Mice
Male BALB/c adult mice (6-8 weeks of age) were obtained from the Jackson Laboratory (Bar Harbor, ME).
Antigens and Immunization
Sheep and chicken red blood cells (SRBC, CRBC) were collected and stored in Alsever's solution at 4°C. The RBC were washed 3 times before use and suspended at the appropriate concentration in sterile pyrogen-free phosphate- buffered saline (PBS). Donor mice were intravenously primed with lo6 RBC, a dose which had previously been shown to induce a T cell response to the exclu- sion of a demonstrable B cell response." This dose of antigen was also used for the induction of delayed-type hypersensitivity (DTH).
Assay for the DTH Reaction in Sensitized Mice
DTH was measured as described previously." Preliminary experiments were performed to determine the optimal regimen for demonstrating the DTH reaction to RBC. The challenging dose was shown to be subinflammatory in an unprimed

LAROCHE el a/.: PHOTOINACTIVATED CELLS 115
animal.I2 The peak of the reaction was determined by a kinetic curve. Four days after immunization with RBC, the mice were challenged with 2.5 x lox SRBC or 0.25 x lo8 CRBC suspended in 50 pI of PBS or saline, injected subcutaneously in the left hind footpad. Footpad swelling was measured 24 hours later with a dial gauge caliper. The results were expressed as the increased thickness of the anti- gen-injected footpad compared to the saline control. In each experiment, a group of nonimmune mice was tested and the footpad swelling was used as a negative control.
Preparation of Cell Suspensions
Spleens were removed from naive or immune mice 4 days after sensitization. Single cell suspensions were prepared by pressing the spleens through a wire mesh into PBS. Red cells were lysed with 0.83% ammonium chloride. The cells were washed twice and adjusted to the appropriate concentration in PBS. Viabil- ity of the cell suspension was determined by trypan blue dye exclusion and found to be at least 90% in all experiments.
In Vitro Photoactivation of 8-MOP by UVA Light
8-MOP (Elder Pharmaceutical) was dissolved in 100% ethanol and subse- quently in PBS. Purity and concentration of the drug was verified by UV spectros- copy and high-pressure liquid chromatography (personal data). Optimal condi- tions for the 8-MOP photoactivation had been previously defined by inhibition of the lymphocyte proliferative response to mitogen and alloantigen. l3 Spleen cells were incubated in uitro with 100 ng/ml of 8-MOP for 20 minutes in foil-wrapped tubes to allow equilibrium to be established between the psoralen solution and the cells. Cells were deposited into Petri dishes (Falcon) and placed on the irradiation surface. The irradiation unit consisted of six black light fluorescent bulbs (40 BL, Sylvania) emitting broad band UVA energy (320-400 nm). Emission in the UVB range was filtered out by a sheet of window glass. The light dose was monitored by a photometer (International Light, IL 700 A). Spleen cells received a light dose of 1 joule/cm2 and were washed twice in PBS before injection.
Treatment of Naive Adult Mice
In uitro photoinactivated spleen cells (2 to 5 x lo7) from either immune or naive donors were injected into the tail vein of naive adult syngeneic recipients every 7 to 10 days for 8 weeks. Following this treatment, recipient mice were tested for a DTH response.
Transfer of DTH
Twenty-four hours after the DTH reaction, mice were sacrificed, the spleens removed and single cell suspensions prepared. The cells were washed and trans- ferred intravenously into syngeneic recipients. Immediately after cell transfer, mice were challenged in the hind footpad with 2.5 x lox SRBC.

116 ANNALS NEW YORK ACADEMY OF SCIENCES
Statistical Analysis
The statistical significance of the data was determined using the Student t test. A p value of less than 0.05 was considered significant.
RESULTS
Inhibition of the DTH Response to SRBC
The effect of pre-exposure of splenocytes photoinactivated in vitro with 8-MOP and UVA on the induction of the DTH reaction to SRBC in adult naive mice was examined. FIGURE 1 shows that mice challenged with SRBC but not sensitized, did not develop a DTH response (negative controls, Group A). Mice that received a priming dose of SRBC and were subsequently challenged in the footpad developed a normal DTH response (ppsitive controls, Group B). Mice treated with photoinactivated nonimmune splenocytes (Group C) and primed and challenged exhibited a normal DTH reaction similar to that observed in the posi- tive controls (Group B). In contrast, treatment with photoinactivated immune
Group
A
B
C
D
- Treatment and
Photoinact ivated Cell Source
None
None
Naive
Immune
Priming
-
+ + +
Footpad Increase
I
I 1
0 20 40 60 (X 10-*mm 1
FIGURE 1. Induction of unresponsiveness to SRBC in mice treated with photoinactivated SRBC-reactive cells. To measure the DTH reaction, BALB/c mice were primed intrave- nously with 106 SRBC and then challenged 4 days later by the injection of 2.5 x 108 SRBC into the left hind footpad. Footpad swelling was measured 24 hr later with a dial caliper. Group A mice (negative controls) received only a challenging dose of SRBC. Group B mice (positive controls) were sensitized and challenged with SRBC. Photoinactivated naive splenocytes or splenocytes from lo6 SRBC-primed mice were treated in uitro with 100 ng/ml of 8-MOP for 20 min and irradiated with 1 joule/cm2 ultraviolet A light. The cells were washed and 2-5 x lo7 photoinactivated splenocytes from naive donors (Group C) or SRBC- primed donors (Group D) were injected into the tail vein of naive syngeneic recipients every 7 to 10 days for 8 weeks. Two days after the sixth treatment, recipient mice were assayed for the T cell response to SRBC, as measured by the DTH reaction. All groups consisted of 5 mice. The results were expressed as increase in footpad thickness. The bars represent the arithmetic mean +/- the standard error.

LAROCHE et al.: PHOTOINACTIVATED CELLS 117
DONORS * REC I PI ENTS - Treatment ond Previous
Source of Res onse Group Photolnoctivated Celh to 8RBC - Footpad Increase
I None - b 2 None + -
............................... 3 Naive + 4 Immune I- -
I
0 4 8 12 16 20 (X mm)
FIGURE 2. Passive transfer of unresponsiveness to normal recipients. Splenocytes (4 x 10’) were harvested from the same animals presented in FIGURE I * and transferred to naive recipients. Group 1 and 2 mice received splenocytes from the negative and positive control animals respectively (Groups A and B respectively, FIG. I ) . Group 3 received cells from Group C mice that had been pretreated with photoinactivated naive splenocytes and had a normal DTH response to SRBC (FIG. I ) . Group 4 mice were recipients of cells from Group D mice that had been pretreated with antigen-reactive splenocytes and did not develop a DTH reaction when primed and challenged with SRBC. Recipient mice were challenged immediately with 2.5 X los SRBC. Swelling of the footpad was measured 24 hr later.
splenocytes rendered the mice unresponsive to priming and challenge with SRBC (Group D, p <0.001). The suppression of the DTH response in this group of mice exposed to photoinactivated immune splenocytes was comparable to the degree of footpad swelling observed in the negative controls (Group A).
Adoptive Transfer of Unreactivity to SRBC
To determine whether inhibition of the response to SRBC could be transfer- red, splenocytes from the same mice previously demonstrated to be unresponsive to SRBC and the controls (FIG. 1) were adoptively transferred to naive syngeneic recipients (FIG. 2). Immediately after transfer, without priming, recipients were challenged in the footpad with SRBC. As expected, transfer of lymphoid cells from the negative control Group A (FIG. 1) resulted in minimal footpad swelling after challenge (Group 1, FIG. 2). Transfer of cells from mice previously shown to be responsive to SRBC (Group B, FIG. 1) resulted in a positive response to the challenge with SRBC (Group 2, FIG. 2) in the naive recipient mice. When cells from the mice treated with photoinactivated naive cells that demonstrated a DTH reaction to priming and challenge with SRBC (Group C, FIG. 1) were injected into naive recipients, they mediated a positive response to SRBC when challenged (Group 3, FIG. 2). In contrast, mice that had received lymphoid cells from animals rendered unresponsive to SRBC (Group D, FIG. 1) by pre-exposure to 8 MOP- UVA inactivated effector cells were unable to develop a significant DTH reaction ( p <0.001, Group 4, FIG. 2).

118 ANNALS NEW YORK ACADEMY OF SCIENCES
Specificity of Unresponsiveness to SRBC
We next assessed whether unresponsiveness to SRBC was antigen specific using chicken RBC (CRBC) as an unrelated antigen (FIG. 3). Untreated control mice not exposed to SRBC did not develop a DTH reaction when challenged with CRBC (Group A, negative control). Positive control mice were primed and chal- lenged with CRBC and developed a strong DTH response (Group B). In addition, a group of mice primed and challenged to SRBC and responsive to that antigen were subsequently primed and challenged with CRBC. These mice were shown to be normally responsive to the second antigen, CRBC (Group C). Mice which were previously treated with photoinactivated naive splenocytes (Group E) developed a normal DTH response after priming and challenge with CRBC. A group of mice, that had previously been shown to be unresponsive to SRBC after exposure to photoinactivated immune splenocytes, received two additional treatments to maintain unresponsiveness before being assayed for the DTH reaction to CRBC (Group D). The positive DTH response to CRBC, in mice demonstrated to be unresponsive to SRBC, was similar to the response to CRBC elicited in the positive controls. As in Group D, the capacity to respond to CRBC was not inhibited in mice treated with photoinactivated SRBC-immune sulenocvtes but. in this case, the mice were not exposed to SRBC prior to testing f i r the rkponse' to CRBC (Group F).
Group
A
B
C
D
E
F
-
Treatment * ond Source of Photoinac- tivated Cells
None
None
None
Immune
Naive
Immune
EXPOSURE TO SRBC CRBC Pr/Ch Pr/Ch Footpad Increase -- -/- -/+ I -1- +/+ y h +/+ +/+ I-+ +/+ +/+ -.
-/- +/+ -3 1 ' " ' " '
0 4 8 12 16 20 24 28 ( X lo-* mm)
FIGURE 3. Specificity of unresponsiveness to SRBC. In Group A (negative control), mice were not primed (FY) but only received a challenging (Ch) dose of CRBC. Group B (positive control) was primed and challenged with SRBC before being exposed to CRBC. Group C mice were first primed and challenged with CRBC. Group D mice were rendered unrespon- sive to SRBC by treatment with photoinactivated SRBC-reactive splenocytes. These mice did not develop a DTH reaction when they were primed and challenged with SRBC. They were treated twice more with photoinactivated effector cells and subsequently primed and challenged with CRBC. Group E mice were pretreated with photoinactivated naive spleno- cytes and then primed and challenged with CRBC. Group F mice were pre-exposed to photoinactivated SRBC-reactive splenocytes and then primed and challenged with CRBC. The priming dose in all groups of mice was lo6 CRBC. All groups were challenged in the footpad with 0.25 x lo8 CRBC.
LAROCHE et al.: PHOTOINACTIVATED CELLS 119
First Second Priming Prlmin
with SRBC Treatment with SRIC Group Footpad Increase
- - D - I I
2 + + I -
3 + + + -
0 20 40 60 ( X 10-2 mm 1
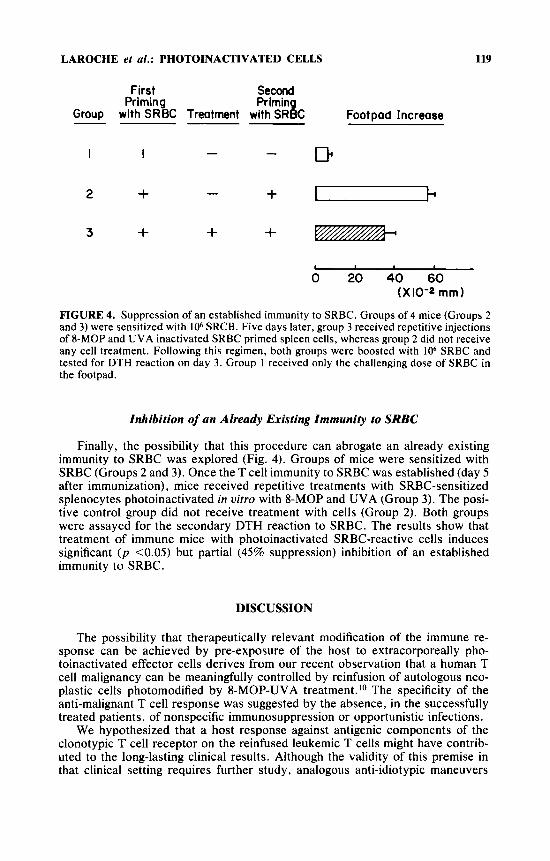
FIGURE 4. Suppression of an established immunity to SRBC. Groups of 4 mice (Groups 2 and 3) were sensitized with lo6 SRCB. Five days later, group 3 received repetitive injections of 8-MOP and UVA inactivated SRBC primed spleen cells, whereas group 2 did not receive any cell treatment. Following this regimen, both groups were boosted with lo6 SRBC and tested for DTH reaction on day 3. Group 1 received only the challenging dose of SRBC in the footpad.
Inhibition of an Already Existing Immunity to SRBC
Finally, the possibility that this procedure can abrogate an already existing immunity to SRBC was explored (Fig. 4). Groups of mice were sensitized with SRBC (Groups 2 and 3). Once the T cell immunity to SRBC was established (day 5 after immunization), mice received repetitive treatments with SRBC-sensitized splenocytes photoinactivated in vitro with 8-MOP and UVA (Group 3). The posi- tive control group did not receive treatment with cells (Group 2). Both groups were assayed for the secondary DTH reaction to SRBC. The results show that treatment of immune mice with photoinactivated SRBC-reactive cells induces significant ( p <0.05) but partial (45% suppression) inhibition of an established immunity to SRBC.
DISCUSSION
The possibility that therapeutically relevant modification of the immune re- sponse can be achieved by pre-exposure of the host to extracorporeally pho- toinactivated effector cells derives from our recent observation that a human T cell malignancy can be meaningfully controlled by reinfusion of autologous neo- plastic cells photomodified by 8-MOP-UVA treatment. lo The specificity of the anti-malignant T cell response was suggested by the absence, in the successfully treated patients, of nonspecific immunosuppression or opportunistic infections.
We hypothesized that a host response against antigenic components of the clonotypic T cell receptor on the reinfused leukemic T cells might have contrib- uted to the long-lasting clinical results. Although the validity of this premise in that clinical setting requires further study, analogous anti-idiotypic maneuvers

120 ANNALS NEW YORK ACADEMY OF SCIENCES
have been previously demonstrated to control the in uitro and in viuo growth of B cell tumors.13J4
In order to test this concept in an experimental system, it was first necessary to demonstrate that T cell-mediated immune responses could be suppressed by the infusion of antigen-reactive effector cells pretreated with 8-MOP-UVA. In the present study, we provide evidence that a state of specific unresponsiveness, as detected by inhibition of the DTH reaction, was induced by administration of syngeneic photoinactivated effector cells to immunocompetent naive mice. The DTH reaction is a sensitive T cell-mediated in uivo assay in which a very small number of anti-SRBC T helper cells have been reported to initiate an inflamma- tory reaction by recruiting bystander cells to the site of antigen deposition in the skin.l5.I6 The dose of SRBC optimal for DTH induction has been demonstrated to be insufficient to elicit B cell activation.I2 Therefore, the regimen used in our study probably evaluated tolerance to a principally T cell-mediated reaction.
The observed suppression of the T cell response to SRBC resulted from a response to the photoinactivated effector T cells and not to a systemic effect of the drug and/or light, since nonimmune splenocytes treated with 8-MOP-UVA and injected into immunologically naive recipients did not inhibit the DTH re- sponse to SRBC. Therefore, effector cells preserved in an immunogenic fashion are required for the autoregulatory immune response which we observe. We have also previously shown that UVA alone, at the dose used in the studies (1 joule/ cm2), does not alter the T cell response to mi tog en^.'^^^^ Furthermore, in the experiments reported in this paper, the in vitro photoinactivated cells were washed before infusion into mice, thereby removing unbound 8-MOP and any minute residual amounts of free 8-MOP would have been excreted by the recipient within several hours.
The induction of tolerance to SRBC was shown to be antigen specific, since reactivity to an irrelevant antigen (CRBC) was unaltered. The retention of the response to this irrelevant antigen demonstrates that reinfusion of photoinacti- vated effector cells does not result in generalized immune suppression. It is note- worthy that, in these experiments, the recipient mice were exposed to splenocyte pools in which expanded populations of T cells reactive to SRBC were admixed with presumably smaller numbers of lymphocytes belonging to each of a multi- plicity of other unstimulated clones. Therefore, the recipient’s immune system was able to discriminate between antigen-reactive cells and the other unrelated cells and specifically suppress the reactivity to SRBC.
The tolerance to SRBC was shown to be transferrable. Naive unprimed recipi- ents of lymphoid cells from unresponsive mice were unable to respond to a challenge with SRBC, while those given lymphoid cells from donors responsive to SRBC were able to mount a normal response to SRBC. The lack of response to SRBC in the unprimed recipient of cells from tolerant mice raises at least two possibilities. Either the effector cells mediating the T cell response to SRBC were not present in the transferred cell population, or a specific suppressor cell subset was present that prevented expression of the DTH reaction. These two possibili- ties, the role of humoral factors and carry-over of tolerogenic antigen in the antigen presenting cell and the immune lymphocyte populations are currently being studied. However, the transfer of suppression to an unprimed secondary recipient that was fully immunocompetent confirms that generalized immune sup- pression of the recipient was not required for inhibition of the response to SRBC in our system. Moreover, our tolerance-inducing procedure was also shown to be able to inhibit an already existing immunity to the antigen, favoring the hypothesis of specific suppressor cell induction. This hypothesis is further supported by our

LAROCHE et a/.: PHOTOINACTIVATED CELLS 121
results obtained in another experimental model in which an antigen-specific inhib- itory cell population was shown to induce tolerance to skin alloantigens.
These experiments demonstrate that it is possible to create a state of specific unresponsiveness to an antigen by pretreating the host with photoinactivated uncloned effector mononuclear cells. Other reported experimental protocols which may mediate similar effects are less clinically feasible, since they rely on isolation and cloning of a known effector cell of an undesirable immune response’ or require inactivation of the reinfused cells with toxic agents such as complete Freund’s a d j ~ v a n t . ~ . ’ ~ In contrast, 8-MOP has been widely used clinically in the photochemotherapy of various skin disorders without significant systemic side effects.19.20 The clinical availability of an 8-MOP-UVA extracorporeal exposure systemlo should allow our tolerance inducing regimen to be readily tested in humans, distinguishing it from the other currently known approaches.
The results of this study have direct implications. First, we have described a new in uiuo model for inducing tolerance and studying the basic mechanisms involved in this phenomenon. Second, it is likely that the principle(s) demon- strated here will apply to other antigens. Therefore, the clinical availability of such a regimen for 8-MOP-UVA activation in extracorporeally routed human blood offers some exciting therapeutic prospects in the control of undesirable cell clones, such as those expanded in graft rejection, allergy, autoimmune diseases, anti-globulin responses elicited by xenogeneic monoclonal antibodies and malig- nant lymphocyte proliferations.
SUMMARY
Development of a protocol that could invoke specific suppression of an unde- sired immune response, while sparing normal immune competence, would be of great clinical value. This report demonstrates that multiple infusions of spleno- cytes sensitized in uiuo to sheep red blood cells (SRBC) and photoinactivated in uitro with 8-methoxypsoralen and ultraviolet A light can render a syngeneic recip- ient selectively unresponsive to subsequent challenge with this antigen. Mice treated in this fashion did not develop a T cell-mediated delayed type hypersensi- tivity (DTH) reaction to SRBC. In contrast, control mice exposed to nonimmune splenocytes pretreated in an identical manner developed a normal DTH response to SRBC, thereby demonstrating that drug and light in the absence of effector T cells were not suppressive. Inhibition of the DTH response was antigen specific, since animals rendered unresponsive to SRBC developed a normal DTH response to chicken red blood cells. Cell transfer experiments demonstrated that unprimed recipients of splenocytes from mice rendered unresponsive to SRBC could not mount a DTH reaction when challenged. Moreover, this procedure can also sup- press established immunity to that antigen. The use of photoinactivated syngeneic antigen-reactive effector cells as immunosuppression agents suggests that this method may be clinically useful in inhibiting pathogenic antigen-specific immuno- logic reactions.
ACKNOWLEDGMENTS
We are indebted to Dr. D. Tripodi for his valuable comments and to Dr. L. Rogozinski for helpful advice during the preparation of the manuscript.

122 ANNALS NEW YORK ACADEMY OF SCIENCES
I . 2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
REFERENCES
WEIGLE, W. 0. 1973. Immunological unresponsiveness. Adv. Immunol. 16: 61-121. SHELLAM, G. R. 1969. Mechanism of induction of immunological tolerance. VI. Toler-
ance induction following thoracic duct drainage or treatment with anti-lymphocyte serum. Immunology 17: 260-280.
SLAVIN, S. , S. STROBER, Z. FUKS & H. KAPLAN. 1977. Induction of specific tissue transplantation tolerance using fractionated total lymphoid irradiation in adult mice: long-term survival of allogeneic bone marrow and skin grafts. J . Exp. Med. 146:
MARRACK, P. & J. KAPPLER. 1987. The T cell receptor. The major histocompatibility complex-restricted antigen receptor on T cells. I. Isolation with monoclonal anti- body. Science 238: 1073-1078.
HASKINS, K., R. KUBO, J. WHITE, M. PIGEON, J. KAPPLER & P. MARRACK. 1983. The major histocompatibility complex-restricted antigen receptor on T cells. I . Isolation with a monoclonal antibody. J. Exp. Med. 157: 1149-1169.
ANDERSON, L. C., H. BINZ & H. WIGZELL. 1976. Specific unresponsiveness to trans- plantation antigens induced by auto-immunization with syngeneic antigen-specific T lymphoblasts. Nature 264: 778-780.
BEN NUN, A., A. WEKERLE & I. R. COHEN. 1981. Vaccination against autoimmune encephalomyelitis using attenuated cells of a T lymphocyte line reactive against myelin basic protein. Nature 292: 60-61.
SCHWARTZ, M., D. NOVICK, D. GIVOL & S. FUCHS. 1978. Induction of anti-idiotypic antibodies by immunization with syngeneic spleen cells educated with acetylcholine receptor. Nature 273: 543-545.
MARON, R., R. ZERUBAVEL, A. FRIEDMAN & I. R. COHEN. 1983. T lymphocyte line specific for thyroglobulin produces or vaccinates autoimmune thyroiditis in mice. J , Immunol. W1: 2316-2322.
EDELSON, R. L., C. BERGER, F. GASPARRO, B. JEGASOTHY, P. HEALD, B. WINTROLJB, E . VONDERHEID, R. KNOBLER, K. WOLFF, G. PLEWIG, G. MCKIERNAN, I. CHRIS- TIENSEN, M. OSTER, H. HONIGSMAN, H. WILFORD, E. KOKOSHKA, T. REHLE, M. PEREZ, G. STINGL & L. LAROCHE. 1987. Treatment of cutaneous T cell lymphoma by extracorporeal photochemotherapy. Preliminary results. N. Engl. J. Med. 316
LAGRANGE, P. H., G. B. MACKANESS & T. E. J. MILLER. 1974. Influence of dose and route of antigen injection on the immunologic induction of T cells. J . Exp. Med. 139:
MILON, G., G. MARCHAL, M. SEMAN, P. TRUFFA-BACCHI & V. ZILBERBERG. 1983. Is the delayed-type hypersensitivity observed after a low dose of antigen mediated by helper T cells? J. Immunol. WO 1103-1107.
BERGER, C. L., C. CANTOR, J. WELSH, P. DERVAN, T. BEGLEY, S. GRANT, F. GAS- PARRO & R. L. EDELSON. 1986. Comparison of synthetic psoralen derivates and 8-MOP in the inhibition of lymphocyte proliferation. Ann. N.Y. Acad. Sci. 453:
KROLICK, K. A., P. C. ISAKSON, J. W. UHR & E. S. VITETTA. 1979. BCLI, a murine model for chronic lymphocytic leukemia: use for the surface immunoglobulin idiotype for the detection and treatment of tumor. Immunol. Rev. 48: 81-106.
MILLER, R. A., D. G. MALONEY, R. WARNER & R. LEVY. 1982. Treatment of B-cell lymphoma with monoclonal anti-idiotype antibody. N. End. J. Med. 306: 517-522.
MARCHAL, G., M. SEMAN, G. MILON, P. TRUFFA-BACCHI & V. ZILBERBERG. 1982. Local adoptive transfer of skin delayed-type hypersensitivity initiated by a single T lymphocyte. J. Immunol. U 9 : 954-958.
PEREZ, M., R. EDELSON, L. LAROCHE & C. BERGER. 1989. Inhibition of antiskin allograft immunity by infusions with syngeneic photoinactivated effector lympho- cytes. J. Invest. Dermatol. 92: 669-676.
BINZ, H. & H. WIGZELL. 1979. Induction of specific transplantation tolerance in adult animals. Transplant. Proc. 11: 914-918.
34-48.
297-303.
528-542.
80-90.

LAROCHE er al.: PHOTOINACTIVATED CELLS 123
19. PARRISH, J . A. , T. B. FITZPATRICK, L. TANNENBAUM & M. A. PATHAK. 1974. Photo- chemotherapy of psoriasis with oral methoxsalen and longwave ultraviolet light. N . Engl. J . Med. 291: 1207-1211.
MELSKI, J . W. , L. TANNENBAUM, J . A. PARRISH, T. FITZPATRICK, H. L. BLEICH e t a / . 1977. Oral methoxsalen photochemotherapy for the treatment of psoriasis: a cooper- ative clinical trial. J . Invest. Dermatol. 68: 328-335.
20.
Related Documents











