Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activity of Newcastle disease virus strains AF2240 and V4-UPM in murine myelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.2011.11.001 ARTICLE IN PRESS G Model LR-4452; No. of Pages 12 Leukemia Research xxx (2011) xxx–xxx Contents lists available at SciVerse ScienceDirect Leukemia Research jo ur nal homep age: www.elsevier.com/locate/leukres Anti-leukemic activity of Newcastle disease virus strains AF2240 and V4-UPM in murine myelomonocytic leukemia in vivo Aied M. Alabsi a,d , Rola Ali a , Aini Ideris b , Abdul Rahman Omar c , Mohd Hair Bejo c , Khatijah Yusoff d , Abdul Manaf Ali a,∗ a Department of Biotechnology, University Sultan Zainal Abidin (UniSZA), Terengganu, Malaysia b Department of Veterinary Clinical Studies, Universiti Putra Malaysia, Serdang, Malaysia c Department of Veterinary Pathology and Microbiology, Universiti Putra Malaysia, Serdang, Malaysia d Department of Molecular and Cell Biology, Universiti Putra Malaysia, Serdang, Malaysia a r t i c l e i n f o Article history: Received 10 September 2011 Received in revised form 31 October 2011 Accepted 1 November 2011 Available online xxx Keywords: NDV AF2240 NDV V4-UPM Cytotoxicity Apoptosis Leukemia BALB/c mice a b s t r a c t Newcastle disease virus (NDV) is a member of the Paramyxoviridae that has caused severe economic losses in poultry industry worldwide. Several strains of NDV were reported to induce cytolysis to can- cerous cell lines. It has prompted much interest as anticancer agent because it can replicate up to 10,000 times better in human cancer cells than in most normal cells. In this study, two NDV strains, viserotropic–velogenic strain AF2240 and lentogenic strain V4-UPM, showed cytolytic activity and apo- ptosis induction against Mouse myelomoncytic leukemia (WEHI 3B). The cytolytic effects of NDV strains were determined using microtetrazolium (MTT) assay. The cytolytic dose – fifty percent (CD 50 ) were 2 and 8 HAU for AF2240 and V4-UPM strains, respectively. Cells treated with NDV strains showed apop- totic features compared to the untreated cells under fluorescence microscope. NDV induced activation of caspase-3 and DNA laddering in agarose gel electrophoresis which confirmed the apoptosis. The anti- leukemic activity of both strains was evaluated on myelomoncytic leukemia BALB/c mice. The results indicated that both NDV strains significantly decreased liver and spleen weights. It also decreased blasts cell percentage in blood, bone marrow and spleen smears of treated mice (p < 0.05). Histopathologi- cal studies for spleen and liver confirmed the hematological results of blood and bone marrow. From the results obtained, the exposure to both NDV stains AF2240 and V4-UPM showed similar results for Ara-c. In conclusion NDV strains AF2240 and V4-UPM can affect WEHI 3B leukemia cells in vitro and in vivo. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction Leukemia is a type of cancer caused by the unregulated prolifer- ation of immature blood cells derived from mutant hematopieotic stem cell. The malignant cells arise out of the arrest of normal blood cell maturation. The malignant cells are effectively trapped at the early stage of differentiation [1,2]. Chemotherapy is considered the main curative treatment for Leukemia; it is characterized by a relatively low efficacy and high toxicity for the patient [3]. Virother- apy, a type of immunotherapy, has been known for more than 70 years. Some viruses such as adenovirus, herpes simplex virus (HSV), reovirus, rabies virus, poliovirus, measles virus, vesicular ∗ Corresponding author at: Department of Biotechnology, University Sultan Zainal Abidin (UniSZA), Kampus Kota, Jalan Sultan Mahmud, Terengganu 43300, Malaysia. Tel.: +60 192723453; fax: +60 192723453. E-mail address: aied [email protected] (A.M. Ali). stomatitis virus, hepatitis A virus and Newcastle disease virus (NDV) have the ability to destroy cancer cells. These viruses can be used without any genetic manipulation or undergo genetic engi- neering for increasing selectivity in animal models and human clinical trials [4–7]. One important attribute to consider virus as a good anti-cancer agent is that the virus must selectively infect, replicate in, and destroy human tumor cells, but should not initiate a symptom-free illness or only cause mild, well- characterized human disease [8,9]. NDV is used as an antineoplastic and immunostimulatory agent in clinical tumor therapy and has been used as an anticancer agent for more than 30 years. The per- ception that NDV can replicate up to 10,000 times better in human cancer cells than in most normal cells, has prompted much interest in this virus as a potential anticancer agent [10]. NDV causes fatal disease in birds, but, in humans, it is generally not very virulent, causing only mild flu-like symptoms, conjunctivitis and laryngi- tis [4,10–14]. The murine WEHI-3B cell line was first established in 1969 and showed characteristics of myelomonocytic leukemia 0145-2126/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.leukres.2011.11.001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

G
L
Am
AKa
b
c
d
a
ARRAA
KNNCALB
1
asbttra7(
AT
0d
ARTICLE IN PRESS Model
R-4452; No. of Pages 12
Leukemia Research xxx (2011) xxx– xxx
Contents lists available at SciVerse ScienceDirect
Leukemia Research
jo ur nal homep age: www.elsev ier .com/ locate / leukres
nti-leukemic activity of Newcastle disease virus strains AF2240 and V4-UPM inurine myelomonocytic leukemia in vivo
ied M. Alabsia,d, Rola Alia, Aini Iderisb, Abdul Rahman Omarc, Mohd Hair Bejoc,hatijah Yusoffd, Abdul Manaf Alia,∗
Department of Biotechnology, University Sultan Zainal Abidin (UniSZA), Terengganu, MalaysiaDepartment of Veterinary Clinical Studies, Universiti Putra Malaysia, Serdang, MalaysiaDepartment of Veterinary Pathology and Microbiology, Universiti Putra Malaysia, Serdang, MalaysiaDepartment of Molecular and Cell Biology, Universiti Putra Malaysia, Serdang, Malaysia
r t i c l e i n f o
rticle history:eceived 10 September 2011eceived in revised form 31 October 2011ccepted 1 November 2011vailable online xxx
eywords:DV AF2240DV V4-UPMytotoxicitypoptosiseukemia
a b s t r a c t
Newcastle disease virus (NDV) is a member of the Paramyxoviridae that has caused severe economiclosses in poultry industry worldwide. Several strains of NDV were reported to induce cytolysis to can-cerous cell lines. It has prompted much interest as anticancer agent because it can replicate up to10,000 times better in human cancer cells than in most normal cells. In this study, two NDV strains,viserotropic–velogenic strain AF2240 and lentogenic strain V4-UPM, showed cytolytic activity and apo-ptosis induction against Mouse myelomoncytic leukemia (WEHI 3B). The cytolytic effects of NDV strainswere determined using microtetrazolium (MTT) assay. The cytolytic dose – fifty percent (CD50) were 2and 8 HAU for AF2240 and V4-UPM strains, respectively. Cells treated with NDV strains showed apop-totic features compared to the untreated cells under fluorescence microscope. NDV induced activationof caspase-3 and DNA laddering in agarose gel electrophoresis which confirmed the apoptosis. The anti-leukemic activity of both strains was evaluated on myelomoncytic leukemia BALB/c mice. The results
ALB/c mice indicated that both NDV strains significantly decreased liver and spleen weights. It also decreased blastscell percentage in blood, bone marrow and spleen smears of treated mice (p < 0.05). Histopathologi-cal studies for spleen and liver confirmed the hematological results of blood and bone marrow. Fromthe results obtained, the exposure to both NDV stains AF2240 and V4-UPM showed similar results forAra-c. In conclusion NDV strains AF2240 and V4-UPM can affect WEHI 3B leukemia cells in vitro andin vivo.
© 2011 Elsevier Ltd. All rights reserved.
. Introduction
Leukemia is a type of cancer caused by the unregulated prolifer-tion of immature blood cells derived from mutant hematopieotictem cell. The malignant cells arise out of the arrest of normallood cell maturation. The malignant cells are effectively trapped athe early stage of differentiation [1,2]. Chemotherapy is consideredhe main curative treatment for Leukemia; it is characterized by aelatively low efficacy and high toxicity for the patient [3]. Virother-
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.2
py, a type of immunotherapy, has been known for more than0 years. Some viruses such as adenovirus, herpes simplex virusHSV), reovirus, rabies virus, poliovirus, measles virus, vesicular
∗ Corresponding author at: Department of Biotechnology, University Sultan Zainalbidin (UniSZA), Kampus Kota, Jalan Sultan Mahmud, Terengganu 43300, Malaysia.el.: +60 192723453; fax: +60 192723453.
E-mail address: aied [email protected] (A.M. Ali).
145-2126/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.leukres.2011.11.001
stomatitis virus, hepatitis A virus and Newcastle disease virus(NDV) have the ability to destroy cancer cells. These viruses can beused without any genetic manipulation or undergo genetic engi-neering for increasing selectivity in animal models and humanclinical trials [4–7]. One important attribute to consider virusas a good anti-cancer agent is that the virus must selectivelyinfect, replicate in, and destroy human tumor cells, but shouldnot initiate a symptom-free illness or only cause mild, well-characterized human disease [8,9]. NDV is used as an antineoplasticand immunostimulatory agent in clinical tumor therapy and hasbeen used as an anticancer agent for more than 30 years. The per-ception that NDV can replicate up to 10,000 times better in humancancer cells than in most normal cells, has prompted much interestin this virus as a potential anticancer agent [10]. NDV causes fatal
of Newcastle disease virus strains AF2240 and V4-UPM in murine011.11.001
disease in birds, but, in humans, it is generally not very virulent,causing only mild flu-like symptoms, conjunctivitis and laryngi-tis [4,10–14]. The murine WEHI-3B cell line was first establishedin 1969 and showed characteristics of myelomonocytic leukemia

ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
2 A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx
Fig. 1. Percentage of cell viability of WEHI 3B treated with NDV AF2240 virus strain after 24, 48, and 72 h. Fifty percent of cytotoxic dose (CD50) after 24 h was 8 HAU whilea
[tBec
smsU(Ulc
altelFmh
Fa
fter 48 and 72 h were 4 and 2 HAU, respectively.
15]. This cell line is a good animal model for detecting differen-iation of leukemia cells because they originally derived from theALB/c mouse [15,16]. The ability of NDV to kill and stop replicatefficiently in human cancer cells was investigated in many miceancer models studies.
There were many studies conducted in the world using differenttrains of NDV on different cancers cell lines in vitro and in vivo. Theost important strains were Strain 73-T, Lytic strain Italians, NDV
train P701 and NDV strain Ulster. Local NDV strains AF2240 and V4PM as oncolytic agent have been studied on breast cancer cell lines
MCF-7 and MDA-231) and brain tumor cell line (DBTRG.5MG and87MG [14,17–21]. However, no study was conducted to use the
ocal NDV strains AF2240 and V4-UPM against WEHI 3B leukemiaell line in vitro and in vivo.
In this study, the in vivo cytolytic activity of NDV strains AF2240nd V4-UPM was investigated by using WEHI 3B a myelomoncyticeukemia cell line, established from BALB/c mice strain. The effect ofhe virus on the hemopioetic system studied by measuring periph-ral blood, bone marrow, spleen and liver cell populations such as
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.
eukemia cells using hematological and histopathological methods.undamental sciences gained from the effects of NDV onto a murineyelomonocytic leukemia cell line will enhance the knowledge on
ow these virus strains affect the activity of these cells.
ig. 2. Percentage of cell viability of WEHI 3B treated with NDV V4-UPM virus strain afterfter 48 and 72 h were 16 and 8 HAU, respectively.
2. Materials and methods
2.1. Propagation and purification of NDV strains AF2240 and V4-UPM
NDV strains were propagated in allantoic fluid of 9–11 days-old embryonatedchicken eggs at 37 ◦C for 48 h. The allantoic fluid was harvested and the presenceof virus was confirmed by the haemaglutination test [22]. NDV strains AF2240andV4-UPM purified as previously described [23,24].
2.2. Murine WEHI-3 leukemia cells
The murine myelomonocytic leukemia cell line WEHI-3B was cultured inDMEM (Sigma, USA) containing 10% fetal bovine serum (FBS), 100 U/ml peni-cillin and 100 �g/ml streptomycin at 37 ◦C in a humidified atmosphere of 5% CO2
in air. Cells were split every 4 days to maintain exponential growth before theexperiments.
2.3. In vitro studies
2.3.1. Assessment for cell viabilityLeukemia cells inhibitions by NDV and Doxorubicin and Arbinocytocine (pos-
itive control) were measured using Microtitration cytotoxicity [25]. About 150 �lcomplete medium were added into of flat-bottom 96-well plate (NunclonTM,
of Newcastle disease virus strains AF2240 and V4-UPM in murine2011.11.001
Denmark) and 50 �l of the 2-folded serial virus dilution were added into the wells.In the last well, 50 �l of PBS were added to instead of the virus, which represented ascontrol. Then 50 �l of 5 × 105 WEHI-3B cells were added to top up the final volumeto 200 �l and the plate was incubated at 37 ◦C in an atmosphere of 5% CO2. Seventy-two hours later, 20 �l of MTT (5 mg/ml) in PBS solution was added to each well
24, 48, and 72 h. Fifty percent of cytotoxic dose (CD50) after 24 h was 64 HAU while

IN PRESSG Model
L
ia Research xxx (2011) xxx– xxx 3
t1FTTt
2d
(rtafai1wsG2t
2
(o7wwQ
2
2
m(Tce
2
dlfdfan
2
iee
22t2lw
2osmcsc
2
hfa(
Fig. 3. Photo-micrographs of fluorescence microscopy examination of WEHI 3B cellstreated with NDV strains AF2240 and V4-UPM at CD50 titer after 72 h staining withacridine orange and propidium iodide (magnification 400×). (A) Untreated WEHI 3B
ARTICLER-4452; No. of Pages 12
A.M. Alabsi et al. / Leukem
hen the plate was further incubated for 4 h. Most of the medium was removed and00 �l of DMSO (dimethyl sulfoxide) was added into the wells to soluble the crystals.inally the absorbance was measured by (ELISA) reader at wavelength of 570 nm.hen graphs of percentage of viable cells versus virus titer (HAU/ml) were plotted.he value of CD50 was determined from the graphs obtained at the concentrationhat cause 50% cell reduction as compared with controls.
.3.2. Quantification of apoptosis using propidium iodide and acridine orangeouble staining
WEHI-3B cells were quantified using propidium iodide (PI) and acridine-orangeAO) double staining according to standard procedures and examine under fluo-escence microscope (Lieca attached with Q-Floro Software) [26,27]. Cells werereated with the viruses at CD50 concentration in six-well plate and incubatedt 37 ◦C in 5% CO2 atmosphere and incubated at 37 ◦C in 5% CO2 atmosphereor 24, 48, and 72 h. After incubation period, the cells were washed with PBSnd then incubated with 5 �l of acridine orange (10 �g/ml) and 5 �l propid-um iodide (10 �g/ml) at a ratio of 1:1 in 1 ml of cells and recentrifuged at000 rpm/5 min. After centrifuge, 50 �l of remaining supernatant with pelletas resuspended and 10 �l of was pipetted on slide before putting on cover
lip. Within 30 min, the slide was analyzed using fluorescent microscope (Leica,ermany). Viable, apoptotic and necrotic cells was quantified in a population of00 cells. This assay provides a useful quantitative evaluation and was done threeimes (n = 3).
.3.3. DNA fragmentationCells at a concentration of 5 × 106 cells/ml were seeded into 6-well plate
NunclonTM, Denmark) in 2 ml culture medium with a concentration of CD50 valuef viruses. Some wells were left with no virus to be used as a control. After the2 h of incubation, cells were spun down at 1000 rpm for 10 min and the pelletas washed with PBS twice. The DNA extraction from treated and untreated cellsas carried out according to protocol of a kit for Blood and Cultured Cells fromIAGEN.
.4. In vivo studies
.4.1. Animal groups and cellsForty 8 weeks old BALB/c mice, weighing 17–21 g, were used in this study. The
ice were bred from the Institute of Medical Research (IMR), Selangor, MalaysiaFBSB 2008 20). They were housed in groups in stainless steel mesh-bottomed cages.he mice were fed with clean water and nutrient food in standard conditions ofonstant temperature and humidity. Both male and female mice were used in thexperiments.
.4.2. Virus and arbinocytocine treatmentBALB/c mice were randomly divided into eight groups with five mice each. One
ay before dosing, every mouse was peritoneally inoculated with 1 × 106 WEHI-3Beukemia cells and then the mice were peritoneally inoculated with CD90 resultedrom MTT, 128 and 300 HAU/mouse for AF2240 and V4-UPM respectively, for sevenays continuously. The treatments of mice with the virus were repeated at day 15or booster dose. Ara-C (Cytosine �-d-arabinofuranoside, C1768) (Sigma, USA) at
dose of 20 mg/kg/day [28,29] was used as the positive control and saline as theegative control.
.4.3. Body weight and mortalityThe body weights of the mice were measured at 0, 7, 14, 21, and 28 days after
noculation with WEHI 3B leukemia cells. All mice groups were checked for Mortalityvery day until day 30 after inoculation of WEHI 3B leukemia cells to determine theffect of NDV on the survival rate of mice.
.4.4. Hematology examination
.4.4.1. Total leukocyte sand differential blood count. Blood from the BALB/c miceail vein was collected after inoculation of WEHI-3B leukemia cells for 7, 14, 21 and8 days, respectively. Total white blood cells were counted by hemocytometry and
eukocyte classification in terms of cell morphology was done by Giemsa stainingith blood smear.
.4.4.2. Bone marrow and spleen aspiration smear. Leukocyte classification in termsf cell morphology was done by Wright’s staining with bone marrow smear andpleen aspiration smear. At the end of the study, all mice were dissected and bonearrows were washed out from the femur of sacrificed mice at day 30 after leukemia
ell inoculation. On the other hand, the spleens were removed and weighed thenmall pieces of spleen were removed and squeezed with PBS to make the spleenells suspension.
.4.5. Histopathological study
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.2
At end of the experiment, all mice were dissected and their spleens and livers,eart and kidney were removed, weighed and labeled and fixed in 10% buffered
ormalin to prepare for light microscopy via dehydration of the tissue samples inn ascending series of alcohol for 14 h in an automatic tissue processor machineLeica, Germany). Leukocyte classification in terms of cell morphology and tissue
cells, (B) WEHI 3B cells treated with CD50 (8 HAU) of NDV V4-UPM, (C) WEHI 3B cellstreated with CD50 (2 HAU) of NDV AF2240. Viable cells (V), Secondary apoptosis (S),Necrotic cell (N) and arrows for cell blebbing were observed.
morphology were done by Haematoxylin and Eosin with spleen, liver, heart andkidney sections.
2.5. Statistical analysis
The results were presented as means ± SEM and the difference between groupswas analyzed by one-way ANOVA in SPSS window program version 14.0 (Chicago,SPSS Inc.). p Value at 0.05 was considered as the level of significance. TukeyHSD post-hoc analysis was done following the ANOVA with p value of lessthan 0.05.
3. Results
3.1. In vitro studies
3.1.1. Cytotoxicity of Newcastle disease virus strains AF2240 and
of Newcastle disease virus strains AF2240 and V4-UPM in murine011.11.001
V4-UPMIn this study, the cytolytic effects of Newcastle disease
virus strains AF2240 and V4-UPM on mouse myelomonocyticleukemia (WEHI-3B), normal mouse fibroblast (3T3), mouse spleen

ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
4 A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx
Table 1Cytotoxic effect dose (CD50) of Newcastle disease virus strains (AF2240 and V4-UPM)against WEHI 3B cell after 6, 24, 48 and 72 h.
Hours AF2240 V4-UPM
6 1024 ± 2.0 NO CD50
24 8 ± 0.7 64 ± 0.648 4 ± 0.6 16 ± 0.7
N
lmcoraasmo
Table 2Percentage of cell viability of human lymphocytes, mouse spleen lymphocytes andnormal mouse fibroblasts treated with 2 and 8 HAU of NDV strains AF2240 andV4-UPM, respectively that killed 50% of WEHI 3B cell line.
Strain WEHI 3B Humanlymphocyte
Mouse spleenlymphocyte
3T3
AF2240 (2 HAU) 50% ± 1.0a 97% ± 0.5b 98% ± 0.7b 97% ± 0.8b
V4-UPM (4 HAU) 50% ± 0.9a 98% ± 0.8b 97% ± 0.5b 98% ± 0.4b
Note: Each value represents of the mean of three replication ± SEM. Values with the
F
Frl
72 2 ± 0.2 8 ± 0.2
ote: Each value represents of the mean of three replication ± SEM.
ymphocyte and peripheral blood lymphocyte were determined byeasuring the cytotoxic dose that kill 50% of the cell population as
ompared to the untreated control for various periods using col-rimetric cytotoxicity assay (MTT). The assay for each strain wasepeated three times. The percentage of viable cells was plottedgainst virus titer (HAU) and the CD50 values were determined
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.
fter different periods (Figs. 1 and 2). AF2240 and V4-UPM strainshowed cytolytic effect on WEHI 3B cell lines in dose-dependentanner. The concentrations of AF2240 strain that killed 50% (CD50)
f WEHI 3B cells, compared to untreated cells after 6, 24, 48 and 72 h
ig. 4. Percentage of necrotic, apoptotic and viable of WEHI 3B cells in the population aft
ig. 5. Agarose-gel-electrophoretic patterns showing DNA fragmentation of WEHI 3B cellespectively. From left to right: lane 1, 100 bp DNA markers; lane 2, WEHI 3B cell treated wane 4, untreated WEHI 3B cell; lanes 5, 6 and 7, WEHI 3B cell treated with NDV strains af
different superscript letters are significantly different (p < 0.05) from each other.
of treatment were 1024, 8, 4 and 2 HAU, respectively, while theconcentrations of V4-UPM strain that killed 50% (CD50) of WEHI 3Bcells compared to untreated cells after 24, 48 and 72 h of treatmentwere 64, 16 and 8 HAU, respectively (Table 1). On the other hand,AF2240 and V4-UPM strains showed low cytotoxic effect (CD50)on normal mouse fibroblast (3T3), mouse spleen lymphocyte and
of Newcastle disease virus strains AF2240 and V4-UPM in murine2011.11.001
peripheral blood lymphocyte cells, which were used as normal cells(Table 2).
er treated with CD50 of NDV strains AF2240 (A) and V4-UPM (B) at different times.
s treated with NDV strain AF2240(A) and V4-UPM (B) at CD50 (2 HAU) and (8 HAU),ith doxorubicin (1 �g/ml); lane 3, WEHI 3B cell treated with doxorubicin (8 �g/ml);ter 24, 48 and 72 h, respectively.

Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activity of Newcastle disease virus strains AF2240 and V4-UPM in murinemyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.2011.11.001
ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx 5
Fig. 6. The effects of Ara-C, NDV strains AF2240 and V4-UPM on body weight (A) and survival rate (B) of BALB/c mice inoculated with WEHI-3B leukemia cells.
Fig. 7. The effects of Ara-C, NDV strains AF2240 and V4-UPM on the total white blood cells count (A) and the percentage of blasts cells (B) in peripheral blood of BALB/c miceinoculated with WEHI-3B leukemia cells.
ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
6 A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx
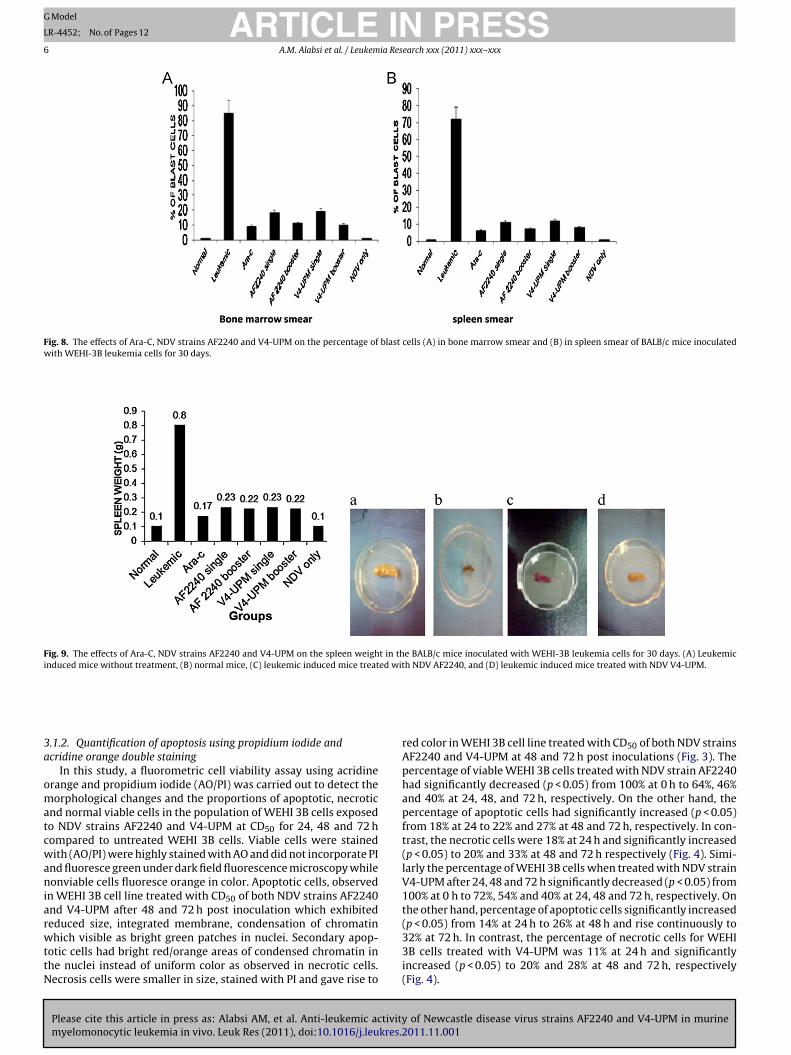
Fig. 8. The effects of Ara-C, NDV strains AF2240 and V4-UPM on the percentage of blast cells (A) in bone marrow smear and (B) in spleen smear of BALB/c mice inoculatedwith WEHI-3B leukemia cells for 30 days.
Fig. 9. The effects of Ara-C, NDV strains AF2240 and V4-UPM on the spleen weight in the BALB/c mice inoculated with WEHI-3B leukemia cells for 30 days. (A) Leukemicinduced mice without treatment, (B) normal mice, (C) leukemic induced mice treated with NDV AF2240, and (D) leukemic induced mice treated with NDV V4-UPM.
3a
omatcwaniarwttN
.1.2. Quantification of apoptosis using propidium iodide andcridine orange double staining
In this study, a fluorometric cell viability assay using acridinerange and propidium iodide (AO/PI) was carried out to detect theorphological changes and the proportions of apoptotic, necrotic
nd normal viable cells in the population of WEHI 3B cells exposedo NDV strains AF2240 and V4-UPM at CD50 for 24, 48 and 72 hompared to untreated WEHI 3B cells. Viable cells were stainedith (AO/PI) were highly stained with AO and did not incorporate PI
nd fluoresce green under dark field fluorescence microscopy whileonviable cells fluoresce orange in color. Apoptotic cells, observed
n WEHI 3B cell line treated with CD50 of both NDV strains AF2240nd V4-UPM after 48 and 72 h post inoculation which exhibitededuced size, integrated membrane, condensation of chromatin
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.
hich visible as bright green patches in nuclei. Secondary apop-otic cells had bright red/orange areas of condensed chromatin inhe nuclei instead of uniform color as observed in necrotic cells.ecrosis cells were smaller in size, stained with PI and gave rise to
red color in WEHI 3B cell line treated with CD50 of both NDV strainsAF2240 and V4-UPM at 48 and 72 h post inoculations (Fig. 3). Thepercentage of viable WEHI 3B cells treated with NDV strain AF2240had significantly decreased (p < 0.05) from 100% at 0 h to 64%, 46%and 40% at 24, 48, and 72 h, respectively. On the other hand, thepercentage of apoptotic cells had significantly increased (p < 0.05)from 18% at 24 to 22% and 27% at 48 and 72 h, respectively. In con-trast, the necrotic cells were 18% at 24 h and significantly increased(p < 0.05) to 20% and 33% at 48 and 72 h respectively (Fig. 4). Simi-larly the percentage of WEHI 3B cells when treated with NDV strainV4-UPM after 24, 48 and 72 h significantly decreased (p < 0.05) from100% at 0 h to 72%, 54% and 40% at 24, 48 and 72 h, respectively. Onthe other hand, percentage of apoptotic cells significantly increased(p < 0.05) from 14% at 24 h to 26% at 48 h and rise continuously to
of Newcastle disease virus strains AF2240 and V4-UPM in murine2011.11.001
32% at 72 h. In contrast, the percentage of necrotic cells for WEHI3B cells treated with V4-UPM was 11% at 24 h and significantlyincreased (p < 0.05) to 20% and 28% at 48 and 72 h, respectively(Fig. 4).

ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx 7
Fig. 10. Morphology of the blood cells smear after staining with Giemsa staining: (A) leukemic induced mice without treatment, (B) normal mice, (C) leukemic inducedmice treated with Ara-C, (D) mice inoculated with NDV only as control, (E) leukemic induced mice treated with NDV AF2240 single dose, (F) leukemic induced mice treatedwith NDV AF2240 booster dose, (G) leukemic induced mice treated with NDV V4-UPM single dose, and (H) leukemic induced mice treated with NDV V4-UPM booster dose(magnification 1000×). Leukemia cells (black arrow).
3
c1tNpa
3
3
awstbt
.1.3. DNA fragmentation assayIn this study, both NDV strains AF2240 and V4-UPM had
aused DNA fragmentation in WEHI 3B cells of multiples of80–200 base pairs that appeared as distinctive ladder-like pat-ern on an agarose gel. Fig. 5 showed that at CD50 value bothDV strains AF2240 and V4-UPM were able to induce apo-tosis by forming clear DNA ladders on agarose gel at 24, 48nd 72 h.
.2. In vivo studies
.2.1. Mice body weight and mortalityThere was no significant difference (p > 0.05) in body weights
mong the mice groups, Fig. 6 showed slight increases in bodyeights of all mice groups with time due to mice feeding. Fig. 6
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.2
howed that 40% of BALB/c mice inoculated with WEHI 3B withoutreatment died before the end of study. In contrast, no mice died inoth NDV strains AF2240 and V4-UPM, and Ara-C groups. The mor-ality was significantly increased in mice group without treatment
compared to mice groups treated with both NDV strains AF2240and V4-UPM and Ara-C. On the other hand, no significant differ-ence was determined between both NDV strains and between bothNDV strains and Ara-C.
3.2.2. Hematological studyAs shown in Figs. 2–4, 7–12 there were significant increases
(p < 0.05) in total white blood cells count, peripheral blood blast,percentages of bone marrow blast cells, spleen blasts cells countand spleen and liver weight of mice group without treatmentcompared to mice groups treated with NDV strains AF2240,V4-UPM and Ara-C, and there was also significant difference(p < 0.05) in total white blood cells count, peripheral blood blast,percentages of bone marrow blast cells, spleen blasts cellscount and spleen and liver weight observed between
of Newcastle disease virus strains AF2240 and V4-UPM in murine011.11.001
mice group treated with single dose compared to micegroup treated with booster dose of both NDV strains. In contrast, no significant difference (p > 0.05) in total whiteblood cells count, peripheral blood blast, percentages of bone
ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
8 A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx
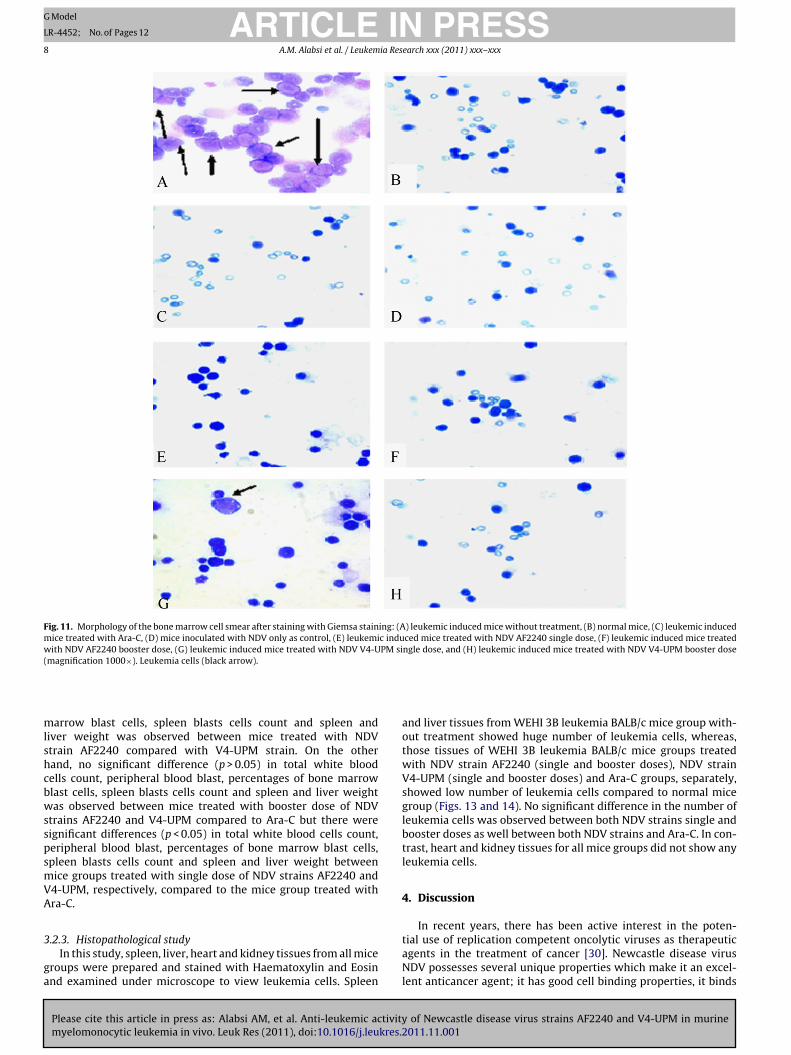
Fig. 11. Morphology of the bone marrow cell smear after staining with Giemsa staining: (A) leukemic induced mice without treatment, (B) normal mice, (C) leukemic inducedmice treated with Ara-C, (D) mice inoculated with NDV only as control, (E) leukemic induced mice treated with NDV AF2240 single dose, (F) leukemic induced mice treatedwith NDV AF2240 booster dose, (G) leukemic induced mice treated with NDV V4-UPM single dose, and (H) leukemic induced mice treated with NDV V4-UPM booster dose(magnification 1000×). Leukemia cells (black arrow).
mlshcbwsspsmVA
3
ga
arrow blast cells, spleen blasts cells count and spleen andiver weight was observed between mice treated with NDVtrain AF2240 compared with V4-UPM strain. On the otherand, no significant difference (p > 0.05) in total white bloodells count, peripheral blood blast, percentages of bone marrowlast cells, spleen blasts cells count and spleen and liver weightas observed between mice treated with booster dose of NDV
trains AF2240 and V4-UPM compared to Ara-C but there wereignificant differences (p < 0.05) in total white blood cells count,eripheral blood blast, percentages of bone marrow blast cells,pleen blasts cells count and spleen and liver weight betweenice groups treated with single dose of NDV strains AF2240 and4-UPM, respectively, compared to the mice group treated withra-C.
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.
.2.3. Histopathological studyIn this study, spleen, liver, heart and kidney tissues from all mice
roups were prepared and stained with Haematoxylin and Eosinnd examined under microscope to view leukemia cells. Spleen
and liver tissues from WEHI 3B leukemia BALB/c mice group with-out treatment showed huge number of leukemia cells, whereas,those tissues of WEHI 3B leukemia BALB/c mice groups treatedwith NDV strain AF2240 (single and booster doses), NDV strainV4-UPM (single and booster doses) and Ara-C groups, separately,showed low number of leukemia cells compared to normal micegroup (Figs. 13 and 14). No significant difference in the number ofleukemia cells was observed between both NDV strains single andbooster doses as well between both NDV strains and Ara-C. In con-trast, heart and kidney tissues for all mice groups did not show anyleukemia cells.
4. Discussion
In recent years, there has been active interest in the poten-
of Newcastle disease virus strains AF2240 and V4-UPM in murine2011.11.001
tial use of replication competent oncolytic viruses as therapeuticagents in the treatment of cancer [30]. Newcastle disease virusNDV possesses several unique properties which make it an excel-lent anticancer agent; it has good cell binding properties, it binds

ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx 9
Fig. 12. Morphology of the spleen cells smear after staining with Giemsa staining: (A) leukemic induced mice without treatment, (B) normal mice, (C) leukemic inducedmice treated with Ara-C, (D) mice inoculated with NDV only as control, (E) leukemic induced mice treated with NDV AF2240 single dose, (F) leukemic induced mice treatedwith NDV AF2240 booster dose, (G) leukemic induced mice treated with NDV V4-UPM single dose, and (H) leukemic induced mice treated with NDV V4-UPM booster dose(magnification 1000×). Leukemia cells (black arrow).
sc[lloosNv
Accibabc
pecifically to tumor cells, it replicates selectively in tumor cellytoplasm, it is relatively safe and it can act as an adjuvant31]. NDV has been used on several different human tumor cellines and tumor models world-wide [18]. In this study, Malaysianocal strains of NDV, AF2240 and V4-UPM, were tested as anncolytic agent on the WEHI 3B leukemia cell line. The effectf NDV has not yet been established in leukemia animal modelo in the present study, we examined the cytolytic effect ofDV strains AF2240 and V4-UPM in WEHI-3 cells in vitro and inivo.
The results obtained from this study revealed that NDV strainsF2240 and V4-UPM inhibited proliferation of WEHI-3B leukemiaells in vitro and induced differentiation of WEHI-3B leukemiaells toward macrophage-like cells. A major pathway of NDVnduced cell death is apoptosis. Apoptosis is often characterized
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.2
y cellular shrinking, chromatin condensation and margination,nd ruffling of the plasma membrane with eventual breaking upreakage of the cell into apoptotic bodies [32,33]. Morphologi-al and chemical changes of apoptotic cells can be differentiated
from any another cell death via many methods. In this study,confirmation of WEHI-3B cell apoptosis initiated by NDV-AF2240and V4-UPM was done using the microscopic examination. AO/PIstaining confirms that NDVAF2240 and V4-UPM in cell culturesinduces cell death via the apoptotic pathway. The number of apop-totic cells revealed that in treated WEHI-3B cells the percentageof apoptotic cells proportionally increased with post-inoculationtime.
In this study we reported that both NDV strains stimulate DNAfragmentation characteristic of apoptosis in WEHI-3B cell line at24, 48 and 72 h post-inoculation. DNA ladder formation is observedwhen the extent of oligonucleosomal cleavage is prominent. Inter-nucleosomal cleavage of DNA is likely to be in the later phaseof apoptotic process [34–36]. In most cell types, the biochemicalcharacteristics of apoptotic response include activation of endoge-
of Newcastle disease virus strains AF2240 and V4-UPM in murine011.11.001
nous calcium and magnesium dependent endonucleases, leadingto fragmentation of the chromosomal DNA. Initially, the DNAfragments are large (50–300 kb) but are later digested to oligonu-cleosomal size (multimers of 180–200 bp). The formation of this
ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
10 A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx
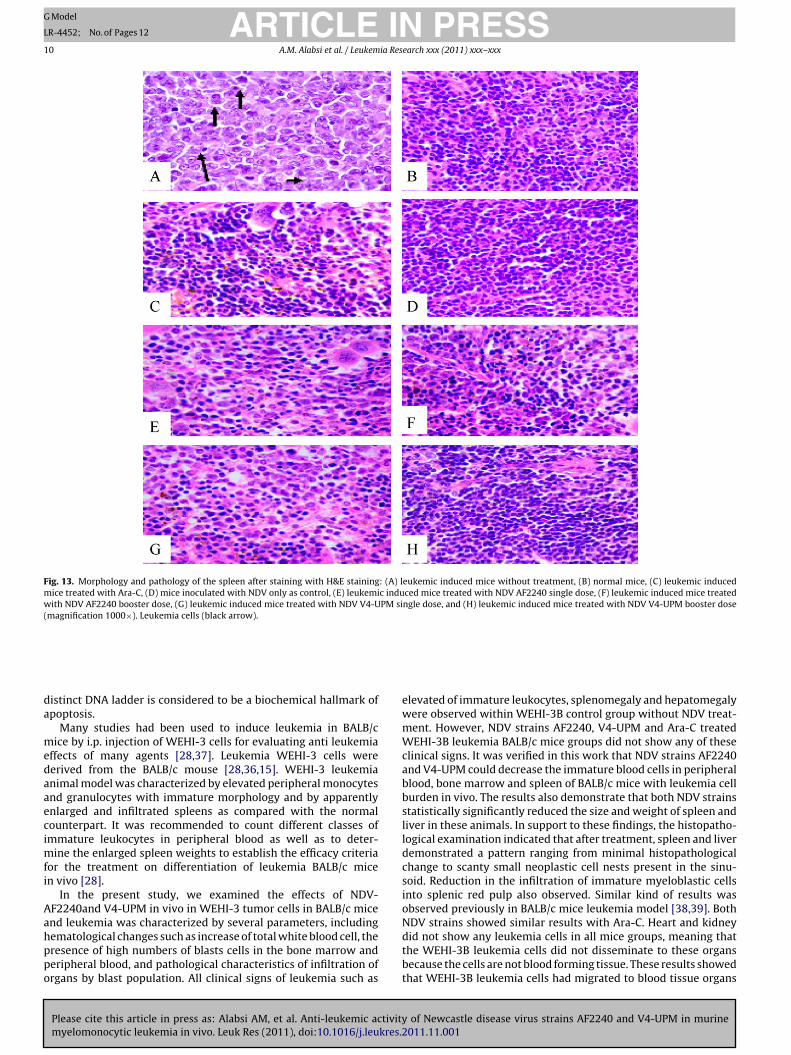
Fig. 13. Morphology and pathology of the spleen after staining with H&E staining: (A) leukemic induced mice without treatment, (B) normal mice, (C) leukemic inducedmice treated with Ara-C, (D) mice inoculated with NDV only as control, (E) leukemic induced mice treated with NDV AF2240 single dose, (F) leukemic induced mice treatedwith NDV AF2240 booster dose, (G) leukemic induced mice treated with NDV V4-UPM single dose, and (H) leukemic induced mice treated with NDV V4-UPM booster dose(magnification 1000×). Leukemia cells (black arrow).
da
medaaecimfi
Aahppo
istinct DNA ladder is considered to be a biochemical hallmark ofpoptosis.
Many studies had been used to induce leukemia in BALB/cice by i.p. injection of WEHI-3 cells for evaluating anti leukemia
ffects of many agents [28,37]. Leukemia WEHI-3 cells wereerived from the BALB/c mouse [28,36,15]. WEHI-3 leukemianimal model was characterized by elevated peripheral monocytesnd granulocytes with immature morphology and by apparentlynlarged and infiltrated spleens as compared with the normalounterpart. It was recommended to count different classes ofmmature leukocytes in peripheral blood as well as to deter-
ine the enlarged spleen weights to establish the efficacy criteriaor the treatment on differentiation of leukemia BALB/c micen vivo [28].
In the present study, we examined the effects of NDV-F2240and V4-UPM in vivo in WEHI-3 tumor cells in BALB/c micend leukemia was characterized by several parameters, including
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.
ematological changes such as increase of total white blood cell, theresence of high numbers of blasts cells in the bone marrow anderipheral blood, and pathological characteristics of infiltration ofrgans by blast population. All clinical signs of leukemia such as
elevated of immature leukocytes, splenomegaly and hepatomegalywere observed within WEHI-3B control group without NDV treat-ment. However, NDV strains AF2240, V4-UPM and Ara-C treatedWEHI-3B leukemia BALB/c mice groups did not show any of theseclinical signs. It was verified in this work that NDV strains AF2240and V4-UPM could decrease the immature blood cells in peripheralblood, bone marrow and spleen of BALB/c mice with leukemia cellburden in vivo. The results also demonstrate that both NDV strainsstatistically significantly reduced the size and weight of spleen andliver in these animals. In support to these findings, the histopatho-logical examination indicated that after treatment, spleen and liverdemonstrated a pattern ranging from minimal histopathologicalchange to scanty small neoplastic cell nests present in the sinu-soid. Reduction in the infiltration of immature myeloblastic cellsinto splenic red pulp also observed. Similar kind of results wasobserved previously in BALB/c mice leukemia model [38,39]. BothNDV strains showed similar results with Ara-C. Heart and kidney
of Newcastle disease virus strains AF2240 and V4-UPM in murine2011.11.001
did not show any leukemia cells in all mice groups, meaning thatthe WEHI-3B leukemia cells did not disseminate to these organsbecause the cells are not blood forming tissue. These results showedthat WEHI-3B leukemia cells had migrated to blood tissue organs
ARTICLE IN PRESSG Model
LR-4452; No. of Pages 12
A.M. Alabsi et al. / Leukemia Research xxx (2011) xxx– xxx 11
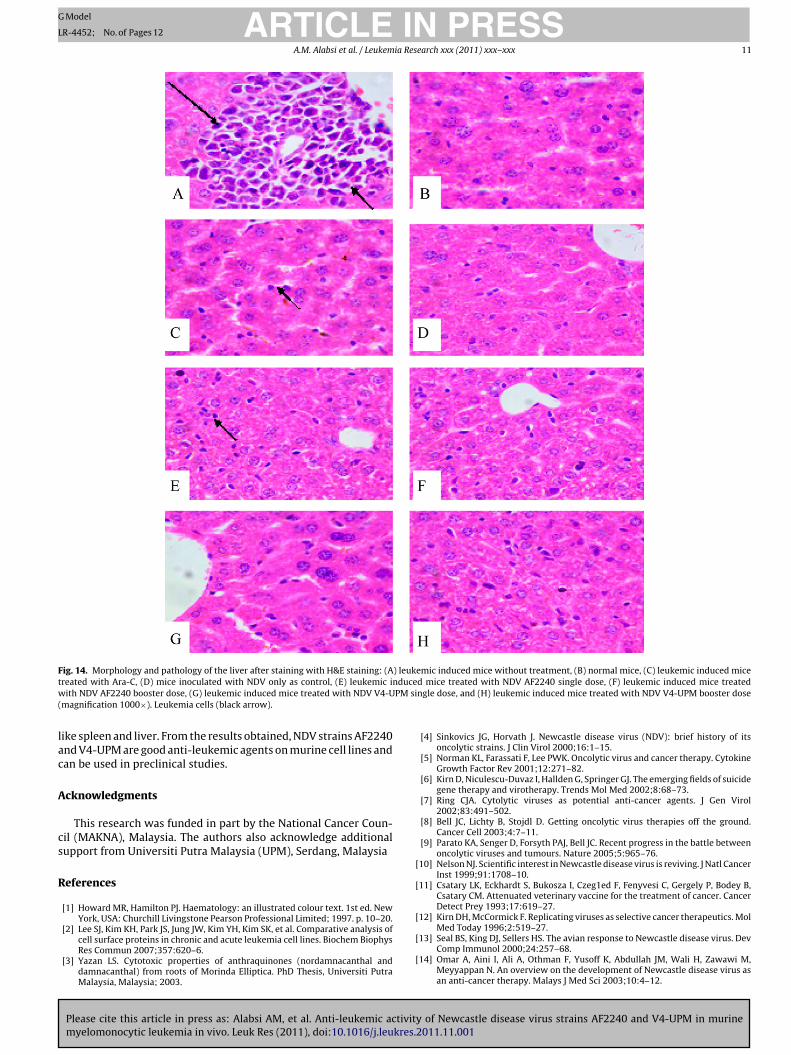
Fig. 14. Morphology and pathology of the liver after staining with H&E staining: (A) leukemic induced mice without treatment, (B) normal mice, (C) leukemic induced micet nducew PM si(
lac
A
cs
R
[
[
[
reated with Ara-C, (D) mice inoculated with NDV only as control, (E) leukemic iith NDV AF2240 booster dose, (G) leukemic induced mice treated with NDV V4-U
magnification 1000×). Leukemia cells (black arrow).
ike spleen and liver. From the results obtained, NDV strains AF2240nd V4-UPM are good anti-leukemic agents on murine cell lines andan be used in preclinical studies.
cknowledgments
This research was funded in part by the National Cancer Coun-il (MAKNA), Malaysia. The authors also acknowledge additionalupport from Universiti Putra Malaysia (UPM), Serdang, Malaysia
eferences
[1] Howard MR, Hamilton PJ. Haematology: an illustrated colour text. 1st ed. NewYork, USA: Churchill Livingstone Pearson Professional Limited; 1997. p. 10–20.
[2] Lee SJ, Kim KH, Park JS, Jung JW, Kim YH, Kim SK, et al. Comparative analysis of
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.2
cell surface proteins in chronic and acute leukemia cell lines. Biochem BiophysRes Commun 2007;357:620–6.
[3] Yazan LS. Cytotoxic properties of anthraquinones (nordamnacanthal anddamnacanthal) from roots of Morinda Elliptica. PhD Thesis, Universiti PutraMalaysia, Malaysia; 2003.
[
[
d mice treated with NDV AF2240 single dose, (F) leukemic induced mice treatedngle dose, and (H) leukemic induced mice treated with NDV V4-UPM booster dose
[4] Sinkovics JG, Horvath J. Newcastle disease virus (NDV): brief history of itsoncolytic strains. J Clin Virol 2000;16:1–15.
[5] Norman KL, Farassati F, Lee PWK. Oncolytic virus and cancer therapy. CytokineGrowth Factor Rev 2001;12:271–82.
[6] Kirn D, Niculescu-Duvaz I, Hallden G, Springer GJ. The emerging fields of suicidegene therapy and virotherapy. Trends Mol Med 2002;8:68–73.
[7] Ring CJA. Cytolytic viruses as potential anti-cancer agents. J Gen Virol2002;83:491–502.
[8] Bell JC, Lichty B, Stojdl D. Getting oncolytic virus therapies off the ground.Cancer Cell 2003;4:7–11.
[9] Parato KA, Senger D, Forsyth PAJ, Bell JC. Recent progress in the battle betweenoncolytic viruses and tumours. Nature 2005;5:965–76.
10] Nelson NJ. Scientific interest in Newcastle disease virus is reviving. J Natl CancerInst 1999;91:1708–10.
11] Csatary LK, Eckhardt S, Bukosza I, Czeg1ed F, Fenyvesi C, Gergely P, Bodey B,Csatary CM. Attenuated veterinary vaccine for the treatment of cancer. CancerDetect Prey 1993;17:619–27.
12] Kirn DH, McCormick F. Replicating viruses as selective cancer therapeutics. MolMed Today 1996;2:519–27.
of Newcastle disease virus strains AF2240 and V4-UPM in murine011.11.001
13] Seal BS, King DJ, Sellers HS. The avian response to Newcastle disease virus. DevComp Immunol 2000;24:257–68.
14] Omar A, Aini I, Ali A, Othman F, Yusoff K, Abdullah JM, Wali H, Zawawi M,Meyyappan N. An overview on the development of Newcastle disease virus asan anti-cancer therapy. Malays J Med Sci 2003;10:4–12.

ING Model
L
1 ia Res
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ARTICLER-4452; No. of Pages 12
2 A.M. Alabsi et al. / Leukem
15] Warner NL, Moore MA, Metcalf D. A transplantable myelomonocytic leukemiain BALB-c mice: cytology, karyotype, and muramidase content. J Natl CancerInst 1969;43:963–82.
16] Hayashi M, Tomida M, Hozumi M. Detection of in vivo differentiation of murineWEHI-3B D+ leukemia cells transfected with the lac-z marker gene using two-color flow cytometry. Leuk Res 1996;2:333–41.
17] Zulkifli MM, Ibrahim R, Ali AM, Aini I, Jaafar H, Hilda SS, Alitheen NB, AbdullahJM. Newcastle diseases virus strain V4UPM displayed oncolytic ability againstexperimental human malignant glioma. Neurol Res 2009;13:3–10.
18] Othman F, Ideris A, Motalleb G, Eshak ZB, Rahmat A. Oncolytic effect of New-castle disease virus AF2240 strain on the MCF-7 breast cancer cell line. YakhtehMed J 2010;12:17–24.
19] Rola A, Aied AM, Ali AM, Ideris A, Omar AR, Yousoff K. Apoptosis induction andcytolytic effects of Newcastle disease virus strain AF2240 on DBTRG. 05 mgbrain tumor cell line. Int J Cancer Res 2011;7, 25–23.
20] Rola A, Aied AM, Ali AM, Ideris A, Omar AR, Yousoff K, Saif-Ali R. Cytolytic effectsand apoptosis induction of Newcastle disease virus strain AF2240 on anaplasticastrocytoma brain tumor cell line. Neurochem Res 2011;36:2051–62.
21] Mustafa Z, Roslan H, Abdullah JM. Targeted therapy for gliomas: the oncolyticvirus applications. In: Abujamra AL, editor. Brain tumors – current and emerg-ing therapeutic strategies. InTech.; 2011., ISBN 978-953-307-588-4.
22] Alexander DJ. Newcastle disease. In: Newcastle disease virus: an avianparamyxovirus. Netherlands: Kluwer Academic Publishers; 1988. p. 1–22.
23] Chambers P, Samson ACR. A new structural protein for Newcastle disease virus.J Gen Virol 1980;50:155–66.
24] Yusoff K, Tan WS, Lau CH, Ng BK, Ibrahim L. Sequence of the haemagglutinin-neuraminidase gene of the Newcastle disease virus oral vaccine strain V4-UPM.Avian Pathol 1996;25:837–44.
25] Mosmann T. Rapid colorimetric assay for cellular growth and survival:application to proliferation and cytotoxicity assays. J Immunol Methods1983;65:55–63.
Please cite this article in press as: Alabsi AM, et al. Anti-leukemic activitymyelomonocytic leukemia in vivo. Leuk Res (2011), doi:10.1016/j.leukres.
26] Mishell BB, Shiiqi SM, Henry C. Selected methods. In: Mishell BB, Shiiqi SM,editors. Cellular immunology. San Francisco, CA, USA: Freeman; 1980. p. 21–2.
27] Guan T, Qin F, Du J, Geng L, Zhang Y, Li M. AICAR inhibits proliferation andinduced S-phase arrest, and promotes apoptosis in CaSki cells. Acta PharmacolSin 2007;28:1984–90.
[
PRESSearch xxx (2011) xxx– xxx
28] He Q, Na X. The effects and mechanisms of a novel 2-aminosteroid onmurine WEHI-3B leukemia cells in vitro and in vivo. Leuk Res 2001;25:455–61.
29] Liu J, Li Y, Tang LJ, Zhang GP, Hu WX. Treatment of lycorine onSCID mice model with human APL cells. Biomed Pharmacother 2007;61:229–34.
30] Schirrmacher V, Haas C, Bonifer R, Ahlert T, Gerhards R, Ertel C. Human tumorcell modification by virus infection: an efficient and safe way to produce cancervaccine with pleiotropic immune stimulatory properties when using Newcastledisease virus. Gene Ther 1999;6:63–73.
31] Schirrmacher V, Ahlert T, Probstle T, Steiner HH, Herold-Mende C, Ger-hand R, et al. Immunization with virus-modified tumor cells. Semin Oncol1998;25:677–96.
32] Cruchten SV, Broeck WV. Morphological and biochemical aspects of apoptosis,oncosis and necrosis. Anat Histol Embryol 2002;31:214–23.
33] Fabian Z, Torocsik B, Kiss K, Csatary LK, Bodey B, Tigyi J, et al. Inductionof apoptosis by a Newcastle disease virus vaccine (MTH-68/H) in PC12 ratphaeochromocytoma cells. Anticancer Res 2001;21:125–35.
34] Cohen GM, Sun XM, Snowden RT, Dinsdale D, Skilleter DN. Key morphologi-cal features of apoptosis may occur in the absence of internucleosomal DNAfragmentation. Biochem J 1992;286:331–4.
35] Oberhammer F, Wilson JW, Dive C, Morris ID, Hickman JA, Wakeling AE, WalkerPR, Sikorska M. Apoptotic death in epithelial cells: cleavage of DNA to 300and/or 50 kb fragments prior to or in the absence of internucleosomal frag-mentation. EMBO J 1993;12:3679–84.
36] Gooch JL, Yee D. Strain-specific differences in formation of apoptotic DNA lad-ders in MCF-7 breast cancer cells. Cancer Lett 1999;144:31–7.
37] Wietrzyk J, Chodynski M, Fitak H, Wojdat E, Kutner A, Opolski A. Antitumorproperties of diastereomeric and geometric analogs of vitamin D3. AnticancerDrugs 2007;18:447–57.
38] Lin JP, Yang JS, Lu CC, Chiang JH, Wu CL, Lin JJ, et al. Rutin inhibits the prolifera-
of Newcastle disease virus strains AF2240 and V4-UPM in murine2011.11.001
tion of murine leukemia WEHI-3 cells in vivo and promotes immune responsein vivo. Leuk Res 2009;33:823–8.
39] Tsou MF, Peng CT, Shih MC, Yang JS, Lu CC, Chiang JH, et al. Benzyl isothiocyanateinhibits murine WEHI-3 leukemia cells in vitro and promotes phagocytosis inBALB/c mice in vivo. Leuk Res 2009;33:1505–11.
Related Documents











