Anti-HuD–Induced Neuronal Apoptosis Underlying Paraneoplastic Gut Dysmotility ROBERTO DE GIORGIO,* MONICA BOVARA,* GIOVANNI BARBARA,* MARCO CANOSSA, ‡ GIOVANNI SARNELLI, § FABRIZIO DE PONTI, ‡ VINCENZO STANGHELLINI,* MARCELLO TONINI, SILVIA CAPPELLO, ‡ ELEONORA PAGNOTTA, ¶ EDUARDO NOBILE– ORAZIO, # and ROBERTO CORINALDESI* Departments of *Internal Medicine and Gastroenterology, ‡ Pharmacology, and ¶ Biochemistry, University of Bologna, Bologna; § Department of Clinical and Experimental Medicine-Hepatogastroenterology, University “Federico II,” University of Naples, Naples; Department of Physiological and Pharmacological Sciences, University of Pavia, Pavia; and # Department of Neurological Sciences, University of Milan, Milan, Italy Background & Aims: The role of autoimmunity under- lying paraneoplastic gut dysmotility remains unset- tled. Because anti-Hu antibodies may impair enteric neuronal function, we tested whether anti-HuD–posi- tive sera from patients with paraneoplastic gut dys- motility or commercial anti-HuD antibodies activated the apoptotic cascade in a neuroblastoma cell line and cultured myenteric neurons. Methods: Anti-HuD antibodies from patients with severe paraneoplastic gut dysmotility were characterized by immunofluores- cence and immunoblot. SH-Sy5Y neuroblasts and cultured myenteric neurons were exposed to sera containing anti-HuD antibodies or 2 commercial anti- HuD antibodies. Cells were processed for terminal deoxynucleotidyl transferase–mediated deoxyuridine triphosphate nick-end labeling (TUNEL) technique to evaluate apoptosis. Immunofluorescence was used to identify activated caspase-3 and apaf-1, along with microtubule-associated protein 2. Results: In SH-Sy5Y cells, the percentage of TUNEL-positive nuclei observed after exposure to anti-HuD–positive sera (32% 7%) or anti-HuD antibodies (23% 2%) was significantly greater than that of control sera or fetal calf serum (P < 0.001). The time-course analysis showed a significantly greater number of apoptotic neuroblastoma cells evoked by the 2 commercial anti-HuD antibodies at 24, 48, and 72 hours versus controls. The number of TUNEL-positive myenteric neurons exposed to anti-HuD antibodies (60% 14%) was significantly greater than that of fetal calf serum (7% 2%; P < 0.001). Apaf-1 and caspase-3 immunolabeling showed intense cytoplasmic staining in a significantly greater proportion of cells exposed to anti-HuD–positive sera or to commercial anti-HuD antibodies compared with controls. Conclusions: Anti-HuD antibodies evoked neuro- nal apoptosis that may contribute to enteric nervous sys- tem impairment underlying paraneoplastic gut dysmotility. Apaf-1 activation suggests participation of a mitochondria- dependent apoptotic pathway. I nflammatory neuropathies affecting the enteric ner- vous system may be idiopathic in origin 1–3 or, more commonly, secondary to a variety of diseases encompass- ing neurologic 4 or infective 5 disorders and paraneoplastic syndromes. 6 –12 In the latter case, a remote, often uniden- tified cancer (such as small cell lung carcinoma or carci- noid tumor) triggers an autoimmune response against the neoplasm that may involve the whole gastrointestinal tract innervation, thus leading to severe dysmotility. 6 –14 Patients with paraneoplastic dysmotility often have one or more types of circulating antineuronal autoanti- bodies detectable in their sera. These autoantibodies include type 1 antineuronal nuclear antibodies (also de- fined as anti-Hu on the basis of their molecular target), type 2 antineuronal nuclear antibodies (or anti-Ri), anti– Purkinje cell cytoplasmic antibodies (anti-Yo), N-type voltage-gated calcium channel antibodies, P/Q-type cal- cium channel antibodies, and ganglionic- and muscle- type nicotinic acetylcholine receptor antibodies. 15–17 In patients with gut motor dysfunction related to small cell lung carcinoma, one of the most commonly identified antineuronal antibodies is anti-Hu, an immu- noglobulin (Ig) G that recognizes a group of proteins with molecular weights in the range of 35– 40 kilodal- tons expressed by both neurons and neoplastic cells. 15–17 Hu antigens are 4 nervous system–specific RNA-binding proteins (identified as HuD, HuC, HuR, and Hel-N1) that share sequence homology with the embryonic-lethal Abbreviations used in this paper: BSA, bovine serum albumin; BSA- PBST, bovine serum albumin in phosphate-buffered saline with Triton X-100; DMEM, Dulbecco’s modified Eagle medium; FCS, fetal calf serum; FITC, fluorescein isothiocyanate; MAP, microtubule-associated protein; TUNEL, terminal deoxynucleotidyl transferase–mediated de- oxyuridine triphosphate nick-end labeling. © 2003 by the American Gastroenterological Association 0016-5085/03/$30.00 doi:10.1016/S0016-5085(03)00664-4 GASTROENTEROLOGY 2003;125:70 –79

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Anti-HuD–Induced Neuronal Apoptosis UnderlyingParaneoplastic Gut Dysmotility
ROBERTO DE GIORGIO,* MONICA BOVARA,* GIOVANNI BARBARA,* MARCO CANOSSA,‡
GIOVANNI SARNELLI,§ FABRIZIO DE PONTI,‡ VINCENZO STANGHELLINI,* MARCELLO TONINI,�
SILVIA CAPPELLO,‡ ELEONORA PAGNOTTA,¶ EDUARDO NOBILE–ORAZIO,# andROBERTO CORINALDESI*Departments of *Internal Medicine and Gastroenterology, ‡Pharmacology, and ¶Biochemistry, University of Bologna, Bologna; §Department ofClinical and Experimental Medicine-Hepatogastroenterology, University “Federico II,” University of Naples, Naples; �Department ofPhysiological and Pharmacological Sciences, University of Pavia, Pavia; and #Department of Neurological Sciences, University of Milan,Milan, Italy
Background & Aims: The role of autoimmunity under-lying paraneoplastic gut dysmotility remains unset-tled. Because anti-Hu antibodies may impair entericneuronal function, we tested whether anti-HuD–posi-tive sera from patients with paraneoplastic gut dys-motility or commercial anti-HuD antibodies activatedthe apoptotic cascade in a neuroblastoma cell lineand cultured myenteric neurons. Methods: Anti-HuDantibodies from patients with severe paraneoplasticgut dysmotility were characterized by immunofluores-cence and immunoblot. SH-Sy5Y neuroblasts andcultured myenteric neurons were exposed to seracontaining anti-HuD antibodies or 2 commercial anti-HuD antibodies. Cells were processed for terminaldeoxynucleotidyl transferase–mediated deoxyuridinetriphosphate nick-end labeling (TUNEL) technique toevaluate apoptosis. Immunofluorescence was used toidentify activated caspase-3 and apaf-1, along withmicrotubule-associated protein 2. Results: In SH-Sy5Ycells, the percentage of TUNEL-positive nuclei observedafter exposure to anti-HuD–positive sera (32% � 7%) oranti-HuD antibodies (23% � 2%) was significantly greaterthan that of control sera or fetal calf serum (P < 0.001).The time-course analysis showed a significantly greaternumber of apoptotic neuroblastoma cells evoked by the 2commercial anti-HuD antibodies at 24, 48, and 72 hoursversus controls. The number of TUNEL-positive myentericneurons exposed to anti-HuD antibodies (60% � 14%) wassignificantly greater than that of fetal calf serum (7% �2%; P < 0.001). Apaf-1 and caspase-3 immunolabelingshowed intense cytoplasmic staining in a significantlygreater proportion of cells exposed to anti-HuD–positivesera or to commercial anti-HuD antibodies compared withcontrols. Conclusions: Anti-HuD antibodies evoked neuro-nal apoptosis that may contribute to enteric nervous sys-tem impairment underlying paraneoplastic gut dysmotility.Apaf-1 activation suggests participation of a mitochondria-dependent apoptotic pathway.
Inflammatory neuropathies affecting the enteric ner-vous system may be idiopathic in origin1–3 or, more
commonly, secondary to a variety of diseases encompass-ing neurologic4 or infective5 disorders and paraneoplasticsyndromes.6–12 In the latter case, a remote, often uniden-tified cancer (such as small cell lung carcinoma or carci-noid tumor) triggers an autoimmune response againstthe neoplasm that may involve the whole gastrointestinaltract innervation, thus leading to severe dysmotility.6–14
Patients with paraneoplastic dysmotility often haveone or more types of circulating antineuronal autoanti-bodies detectable in their sera. These autoantibodiesinclude type 1 antineuronal nuclear antibodies (also de-fined as anti-Hu on the basis of their molecular target),type 2 antineuronal nuclear antibodies (or anti-Ri), anti–Purkinje cell cytoplasmic antibodies (anti-Yo), N-typevoltage-gated calcium channel antibodies, P/Q-type cal-cium channel antibodies, and ganglionic- and muscle-type nicotinic acetylcholine receptor antibodies.15–17
In patients with gut motor dysfunction related tosmall cell lung carcinoma, one of the most commonlyidentified antineuronal antibodies is anti-Hu, an immu-noglobulin (Ig) G that recognizes a group of proteinswith molecular weights in the range of 35–40 kilodal-tons expressed by both neurons and neoplastic cells.15–17
Hu antigens are 4 nervous system–specific RNA-bindingproteins (identified as HuD, HuC, HuR, and Hel-N1)that share sequence homology with the embryonic-lethal
Abbreviations used in this paper: BSA, bovine serum albumin; BSA-PBST, bovine serum albumin in phosphate-buffered saline with TritonX-100; DMEM, Dulbecco’s modified Eagle medium; FCS, fetal calfserum; FITC, fluorescein isothiocyanate; MAP, microtubule-associatedprotein; TUNEL, terminal deoxynucleotidyl transferase–mediated de-oxyuridine triphosphate nick-end labeling.
© 2003 by the American Gastroenterological Association0016-5085/03/$30.00
doi:10.1016/S0016-5085(03)00664-4
GASTROENTEROLOGY 2003;125:70–79

abnormal vision RNA-binding protein of Drosophila.Whereas HuR is ubiquitously expressed in proliferatingcells, HuC, HuD, and Hel-N1 proteins are identifiedexclusively in neurons,18,19 where they exert a crucial rolein neuronal development and survival.20–22
Although the identification of circulating anti-Hu inpatients with paraneoplastic dysmotility is important fordiagnosis, the role exerted by these autoantibodies inevoking enteric neuron dysfunction, and thus gastroin-testinal dysmotility, is still unclear. Preliminary in vitroevidence in an animal model suggests that anti-Hu an-tibodies may impair the ascending reflex pathways ofperistalsis, thus providing the basis for occurrence of gutdysmotility.23 Because it is known that enteric neurode-generation may occur in patients with paraneoplasticdysmotility,6–11,13 the present study was undertaken togenerate new insights into the mechanisms throughwhich anti-Hu antibodies evoke neuronal damage. Weused a neuroblastoma cell line and guinea pig culturedmyenteric neurons to assess whether sera with provenanti-HuD autoantibodies from patients with small celllung carcinoma–related gut paraneoplastic syndromecould activate apoptosis. In addition, to further confirmthat the HuD autoantibodies were responsible for neu-ronal alterations, we tested whether pure, specific anti-HuD antibodies, obtained from a commercial source,induced the same alterations.
Patients and MethodsPatients and Characterization of Anti-HuDAntibodies
Five selected patients (2 women and 3 men; age range,55–64 years) with an established diagnosis of paraneoplasticsyndrome with gastrointestinal involvement and concomitantcentral or peripheral nervous system dysfunction were studied(Table 1). Patients were investigated at or referred to theDepartment of Internal Medicine and Gastroenterology of theUniversity of Bologna and the Department of Neurology of theUniversity of Milan, and all gave informed consent.
The paraneoplastic syndrome was related to an occult, pre-viously unidentified small cell lung carcinoma in all patients.The neoplasm was detected during follow-up and was proven
histologically either by biopsy material or postmortem analy-sis. Sera from healthy subjects (blood donors) (2 men and 2women; age range, 50–65 years) served as controls.
In each patient, anti-HuD antibodies were identified andcharacterized by indirect immunofluorescence (using either ratcerebellar cortex or gastrointestinal tract) (for titer range, seeTable 1)24 and immunoblots25 of recombinant paraneoplasticproteins or human neuronal protein extracts (amphiphysin,CDR62, Nova-1, and HuD).24,25 The latter analysis provedthat all tested sera were HuD positive. Control sera from blooddonors tested negative to all of the aforementioned autoanti-bodies as expected.
Cell Cultures
SH-Sy5Y neuroblasts and 7-day-old myenteric neuronswere exposed to anti-HuD–positive sera or specific anti-HuDantibodies for 12 hours (for activated apaf-1 and caspase-3immunohistochemistry) or 48 hours (to identify apoptosis byterminal deoxynucleotidyl transferase–mediated deoxyuridinetriphosphate nick-end labeling [TUNEL] technique).
SH-Sy5Y cells. The human neuroblastoma cell clonewas maintained in 100-cm2 dishes in a 1:1 mixture of Dul-becco’s modified Eagle medium (DMEM) and Ham’s F-12containing 15% fetal calf serum (FCS), penicillin (100 IU/mL),streptomycin (100 �g/mL), and nonessential amino acids (100�g/mL) at 37°C in a 5% CO2 atmosphere. The cells weredetached every 2 days with trypsin/ethylenediaminetetraaceticacid (0.05% trypsin and 0.53 mmol/L ethylenediaminetet-raacetic acid) followed by centrifugation, resuspension, andfinally seeding in new plates. All reagents were obtained fromGibco BRL Laboratories (Gaithersburg, MD). Passages neverexceeded 30.
Cultured myenteric neurons. The experiments wereperformed at the University of Bologna according to theEuropean Union Directive 86/609 on the care and use ofexperimental animals. The protocol was approved by the in-stitutional ethics committee at the University of Bologna.Guinea pigs (Charles River, Como, Italy) of both sexes (bodywt, 300–350 g) were used. Myenteric neuron cultures wereprepared from freshly isolated guinea pig small intestine ac-cording to well-standardized protocols.26–28 Briefly, the lon-gitudinal muscle and myenteric plexus were stripped from thecircular muscle and mucosa and digested in an enzymaticsolution containing 1 mg/mL protease and 1.25 mg/mL col-lagenase (both from Sigma Chemical Co., St Louis, MO). After
Table 1. Clinical Features and Anti-HuD Titers (Highest Dilutions) of Patients With Paraneoplastic Syndromes
Patient no. Age (yr) SexAnti-HuD
titer Gastrointestinal involvementCentral/peripheral nervous system
involvement
1 55 M 1:10,000 Intestinal pseudo-obstruction —2 58 F 1:51,000 Constipation with megacolon Encephalomyelitis3 61 M 1:52,200 Intestinal pseudo-obstruction Subacute sensory-motor neuropathy4 64 M 1:20,000 Gastroparesis —5 58 F 1:12,800 Intestinal pseudo-obstruction —
—, absence.
July 2003 ANTI-HUD EVOKED APOPTOSIS IN ENTERIC NEURONS 71

a 30-minute incubation at 37°C, the suspension was placed onice and centrifuged at 200g. Ganglia were picked up andplated in culture dishes, where they adhered to the coverglassbottom. After a few days, neurons started growing in network-like structures reminiscent of the ganglionated plexuses. Theculture medium was changed every 2 days and consisted ofmedium 199 enriched with 10% FCS and 50 ng/mL of 7snerve growth factor. Antibiotics (penicillin, 100 IU/mL; strep-tomycin, 100 �g/mL; gentamicin, 50 �g/mL) were added tothe medium; glucose concentration was elevated to 30mmol/L. All reagents were obtained from Gibco BRL Labora-tories. The culture chambers were kept in an incubator at 37°Cand continuously gassed with a mixture of 95% O2 and 5%CO2. Experiments were performed after 7–10 days of plating.
Immunofluorescence
SH-Sy5Y cells were grown for 24 hours on poly-L-lysine– coated coverslips and then fixed in 3.7% buffered form-aldehyde for 10 minutes and then in methanol for 20 minutesat room temperature. Fixed cells were incubated for 30 min-utes at room temperature in buffer containing 3% bovineserum albumin (BSA) in phosphate-buffered saline (PBS), pH7.4, with 0.1% Triton X-100 (BSA-PBST). To identify im-munolabeling of patient sera containing anti-Hu antibodies,SH-Sy5Y cells were incubated with 15% of either control ortest serum overnight at 4°C and then with goat anti-humanIgG conjugated with fluorescein isothiocyanate (FITC) (1:64;Sigma Immunochemicals, St. Louis, MO) in BSA-PBST for 1hour at room temperature in the dark. Further immunofluo-rescence studies were performed using an affinity-purified goatpolyclonal antibody raised against the human HuD peptide(Santa Cruz Biotechnology, Santa Cruz, CA; catalogue refer-ence L-20) and a mouse monoclonal anti-human HuC/HuDantibody (Molecular Probes Inc., Eugene, OR), both diluted1:100 and incubated overnight at 4°C. Cells were rinsed withPBS and then exposed to either an affinity-purified rabbitanti-goat IgG FITC conjugated (1:150; Sigma Immunochemi-cals) or an affinity-purified horse anti-mouse IgG FITC conju-gated (1:150; Sigma Immunochemicals) in BSA-PBST for 1hour at room temperature in the dark. Coverslips were rinsedwith PBS and mounted onto glass slides with mountingmedia. Specificity of the immunolabeling was assessed with thefollowing control experiments: (1) substitution of the primaryantibody with normal goat or human sera and (2) incubation ofthe primary antibodies (anti-HuD antibodies) preadsorbedwith a 5-fold excess (according to weight) of the homologousHuD antigen (Santa Cruz Biotechnology) in 50 �L PBS over-night at 4°C.
Cultured myenteric neurons were fixed in 4% paraformal-dehyde solution in PBS for 30 minutes at 4°C and thereafterpretreated with 3% BSA in a 0.1 mol/L PBS solution for 30minutes at room temperature. Cells were processed for single-and double-labeling immunofluorescence. For single labeling,cells were incubated with a rabbit polyclonal anti-microtu-bule–associated protein (MAP)-2 antibody (1:50; ChemiconInternational, Temecula, CA) overnight at 4°C, washed thor-
oughly in PBS, and then incubated with a goat anti-rabbit IgGconjugated with tetramethyl rhodamine isothiocyanate (1:32;Sigma Immunochemicals) in BSA-PBST for 1 hour at roomtemperature in the dark. Cells were washed again in PBS andmounted onto glass slides with mounting media. To establishwhether anti-HuD antibodies actually labeled cultured myen-teric neurons, a double-label immunofluorescence techniquewas performed. Briefly, cells were incubated in a mixturecontaining MAP-2 and anti-HuD antibodies (1:100) over-night at 4°C, followed by 1-hour incubation at room temper-ature with a mixture of goat anti-rabbit tetramethyl rhoda-mine isothiocyanate– conjugated IgG (1:32) and rabbit anti-goat FITC-conjugated IgG (1:150; Sigma Immunochemicals).Coverslips were rinsed with PBS and mounted onto glass slideswith mounting media. Specificity of binding to HuD antigenwas tested as previously described. For double-labeling tech-nique, additional specificity studies were performed by incu-bating the primary antibodies with the HuD peptide (used atthe concentration previously reported) at 4°C overnight beforeincubation with myenteric neurons. In addition, further ap-propriate controls for double-labeling technique were per-formed to determine that the primary antisera do not crossreact when mixed together and that the secondary antibodiesrecognize the appropriate antigen-antibody complexes.12
Evaluation of Apoptosis
The apoptosis induced by patient sera or anti-HuDantibodies was detected using the TUNEL method and thefluorescent dye Hoechst 33342 (Sigma Chemical Co.). Fur-thermore, the apoptotic nature of neuronal cell death wasanalyzed by the immunocytochemical staining with antibodiesdirected against the activated forms of the 2 apoptotic en-zymes, caspase-3 and apaf-1, according to the protocol de-scribed below.
TUNEL method. SH-Sy5Y neuronal cells were grownfor 24 hours on poly-L-lysine– coated coverslips and thenincubated for 48 hours in DMEM containing 15% of eithercontrol or patient sera. SH-Sy5Y neuroblastoma cells were alsoincubated for 24, 48, and 72 hours in DMEM containingcommercial anti-HuD antibodies at several dilutions (rangingfrom 1:100 up to 1:1000). All of these experiments wereperformed in duplicate. Fixed neuroblastoma cells were pro-cessed with either an in situ apoptosis detection kit (NeurotacsII, Trevigen, MD) or the dead-end fluorometric TUNEL sys-tem (Promega, Madison, WI) according to validated proto-cols.29,30 With the Neurotacs II method, the apoptotic nucleiappeared dark brown and could be visualized with phase-contrast light transmission microscopy (Leica Westlar, Germa-ny); with the TUNEL system, which allowed identification ofFITC-labeled DNA fragments, apoptotic cells were visualizedwith a microscope equipped with fluorescent light (LeicaWestlar). As a positive control for the induction of apoptosis,SH-Sy5Y cells were cultured for 12 hours in DMEM contain-ing dopamine (100 mmol/L), a well-known proapoptotic mes-senger for this cell line.31
72 DE GIORGIO ET AL. GASTROENTEROLOGY Vol. 125, No. 1

Nuclear condensation assessed by Hoechst33342 dye. This method was applied to further prove theoccurrence of nuclear condensation as a result of the apoptosisinduced by commercial anti-HuD commercial antibodies or bysera of patients positive for anti-HuD antibodies. Briefly, cellswere cultured for 48 hours in DMEM containing either anti-HuD antibodies (1:100 to 1:1000) or 15% test serum fromeach patient. Control experiments were performed in parallelusing cells cultured for 48 hours in DMEM containing 15% ofeither FCS or control sera. Cells incubated at 37°C withHoechst 33342 dye (10 �g/mL, 15 minutes) were subse-quently fixed as previously described and visualized using afluorescence microscope.
Caspase-3 and Apaf-1 immunocytochemistry.SH-Sy5Y cells were grown for 24 hours on poly-L-lysine–coated coverslips and then cultured for 12 hours in DMEMcontaining 15% of either control or test serum and commercialanti-HuD antibodies at several dilutions (1:100 to 1:1000).Rabbit polyclonal anti-activated caspase-3 (1:50) and apaf-1(1:50) antibodies were applied to fixed SH-Sy5Y cells. Thesecondary antibody used was an affinity-purified goat anti-rabbit FITC-conjugated IgG (1:64; Sigma Immunochemicals)according to the method previously described. Specificity stud-ies included control experiments commonly applied in immu-nohistochemical protocols as previously reported.12
Neuronal apoptotic assays in cultured myentericneurons. Myenteric neurons plated for 7–10 days were ex-posed to culture medium containing either 15% patient sera oranti-HuD antibodies (1:100 for the antibody from Santa CruzBiotechnology and 1:50 for the antibody from MolecularProbes Inc.) for 48 hours. To show neuronal cell death, simul-taneous visualizations of (1) MAP-2, Hoechst 33342 dye, andTUNEL or (2) MAP-2, Hoechst 33342 dye, and caspase-3were performed.
Quantification of Apoptosis
The number of TUNEL-positive neurons along withneurons immunolabeled for activated caspase-3 or apaf-1was counted in microscopic fields and expressed as a per-centage of neurons detectable in the field. At least 3 ran-domly selected fields per dish were blindly counted by 2expert investigators (R.D. and M.B.). In each experiment,one dish per experimental condition (i.e., patient’s serumpositive for HuD or commercial HuD antibodies at differ-ent dilutions/incubation times) was counted and experi-ments were repeated 3 times.
Confocal Microscopy
Slides were analyzed using a laser scanning confocalmicroscope (Bio-Rad MRC 1024; Bio-Rad, Hercules, CA)equipped with a Nikon Eclipse TE300 (Nikon, Turin,Italy). Filter sets included the following: for tetramethylrhodamine isothiocyanate, excitation 568 nm, emission605/32 nm; for FITC, excitation 488 nm, emission 522/35nm; for UV excitation, 351 � 363 nm, emission 522/35
nm. Images were cropped in Adobe Photoshop with mini-mal alteration (minor adjustment to brightness and con-trast) and transferred to dedicated software for constructionof figure sets.
Statistical Analysis
The data are expressed as mean � standard error.The percentages of cells positive for TUNEL (neuro-blastoma cells and myenteric neurons), caspase-3, andapaf-1 (SH-Sy5Y) immunoreactivities exposed to sera con-taining anti-HuD, commercial anti-HuD antibody, andcontrol sera/FCS were evaluated by Fisher exact test (Graph-Pad Prism version 3.00 for Windows; GraphPad Software,San Diego, CA). The accepted level of significance was P �0.05.
ResultsAnti-HuD–Positive Sera and Anti-HuDAntibody Immunostaining in SH-Sy5Y Cellsand Cultured Myenteric Neurons
SH-Sy5Y cells exposed to anti-HuD–positive seraor commercially available anti-HuD antibodies (Figure1A and B) showed an intense immunofluorescence in thecytoplasm and around the perinuclear membrane. Incontrast, no immunolabeling was observed in SH-Sy5Ycells exposed to control sera (Figure 1C) or in cells inwhich the anti-HuD antibody was preadsorbed with thehomologous peptide (Figure 1D).
Cultured myenteric neurons were identified byMAP-2 immunoreactivity (Figure 2A and C). Double-labeling experiments showed that HuD immunoreactiv-ity of specific commercially available antibodies (Figure2B) colocalized with MAP-2 staining in the ganglion cellbodies. Anti-HuD immunoreactivity was also detectedaround the nuclei of cultured myenteric neurons.
Specificity experiments showed absence of immunore-activity in myenteric neurons incubated with normalgoat serum or with the primary antibodies (anti-HuD)preadsorbed with the HuD synthetic peptide (Figure2D). Double-labeling specificity was shown by the lackof cross-reactivity between primary antisera.
Anti-HuD–Positive Sera and Anti-HuDAntibodies Evoke Apoptosisin SH-Sy5Y Cells
The incubation of SH-Sy5Y cells with anti-HuD–positive sera induced an intense nuclear fluorescence(TUNEL positivity, Figure 3A). All of the TUNEL-positive nuclei showed nuclear condensation with theHoechst 33342 dye (Figure 3B). Quantitative analysisshowed that the number of TUNEL-positive nuclei in-duced by anti-HuD–positive sera (32% � 7%, n � 5
July 2003 ANTI-HUD EVOKED APOPTOSIS IN ENTERIC NEURONS 73
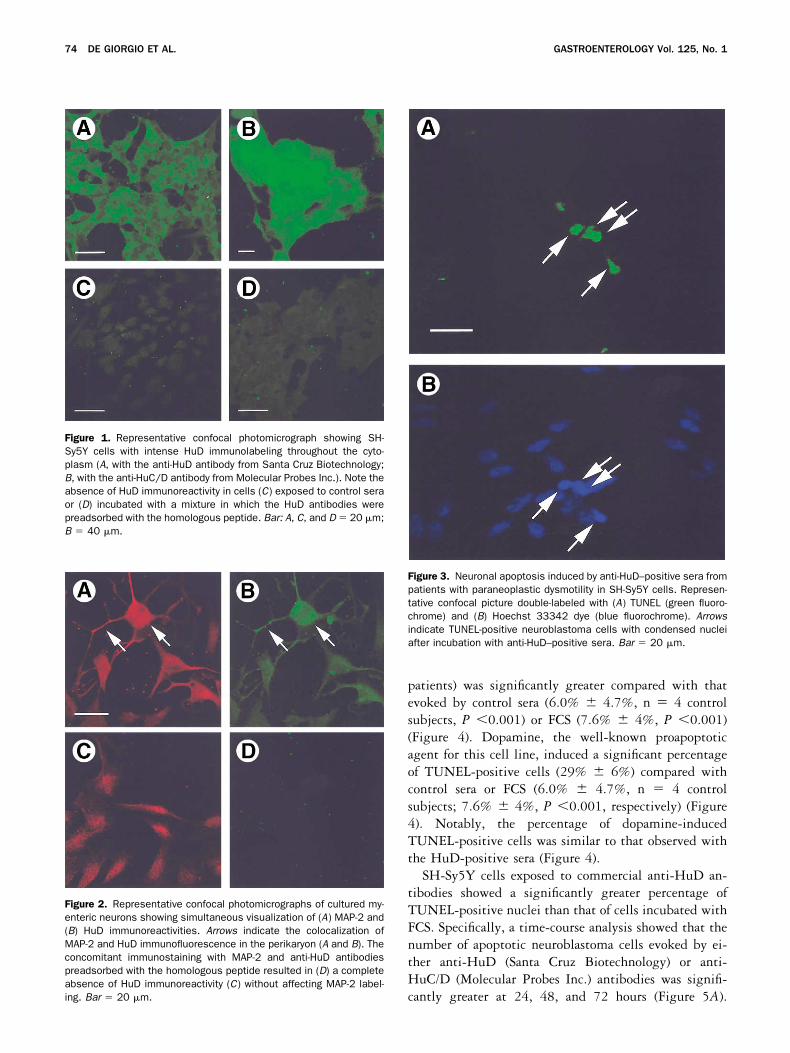
patients) was significantly greater compared with thatevoked by control sera (6.0% � 4.7%, n � 4 controlsubjects, P �0.001) or FCS (7.6% � 4%, P �0.001)(Figure 4). Dopamine, the well-known proapoptoticagent for this cell line, induced a significant percentageof TUNEL-positive cells (29% � 6%) compared withcontrol sera or FCS (6.0% � 4.7%, n � 4 controlsubjects; 7.6% � 4%, P �0.001, respectively) (Figure4). Notably, the percentage of dopamine-inducedTUNEL-positive cells was similar to that observed withthe HuD-positive sera (Figure 4).
SH-Sy5Y cells exposed to commercial anti-HuD an-tibodies showed a significantly greater percentage ofTUNEL-positive nuclei than that of cells incubated withFCS. Specifically, a time-course analysis showed that thenumber of apoptotic neuroblastoma cells evoked by ei-ther anti-HuD (Santa Cruz Biotechnology) or anti-HuC/D (Molecular Probes Inc.) antibodies was signifi-cantly greater at 24, 48, and 72 hours (Figure 5A).
Figure 1. Representative confocal photomicrograph showing SH-Sy5Y cells with intense HuD immunolabeling throughout the cyto-plasm (A, with the anti-HuD antibody from Santa Cruz Biotechnology;B, with the anti-HuC/D antibody from Molecular Probes Inc.). Note theabsence of HuD immunoreactivity in cells (C) exposed to control seraor (D) incubated with a mixture in which the HuD antibodies werepreadsorbed with the homologous peptide. Bar: A, C, and D � 20 �m;B � 40 �m.
Figure 2. Representative confocal photomicrographs of cultured my-enteric neurons showing simultaneous visualization of (A) MAP-2 and(B) HuD immunoreactivities. Arrows indicate the colocalization ofMAP-2 and HuD immunofluorescence in the perikaryon (A and B). Theconcomitant immunostaining with MAP-2 and anti-HuD antibodiespreadsorbed with the homologous peptide resulted in (D) a completeabsence of HuD immunoreactivity (C) without affecting MAP-2 label-ing. Bar � 20 �m.
Figure 3. Neuronal apoptosis induced by anti-HuD–positive sera frompatients with paraneoplastic dysmotility in SH-Sy5Y cells. Represen-tative confocal picture double-labeled with (A) TUNEL (green fluoro-chrome) and (B) Hoechst 33342 dye (blue fluorochrome). Arrowsindicate TUNEL-positive neuroblastoma cells with condensed nucleiafter incubation with anti-HuD–positive sera. Bar � 20 �m.
74 DE GIORGIO ET AL. GASTROENTEROLOGY Vol. 125, No. 1
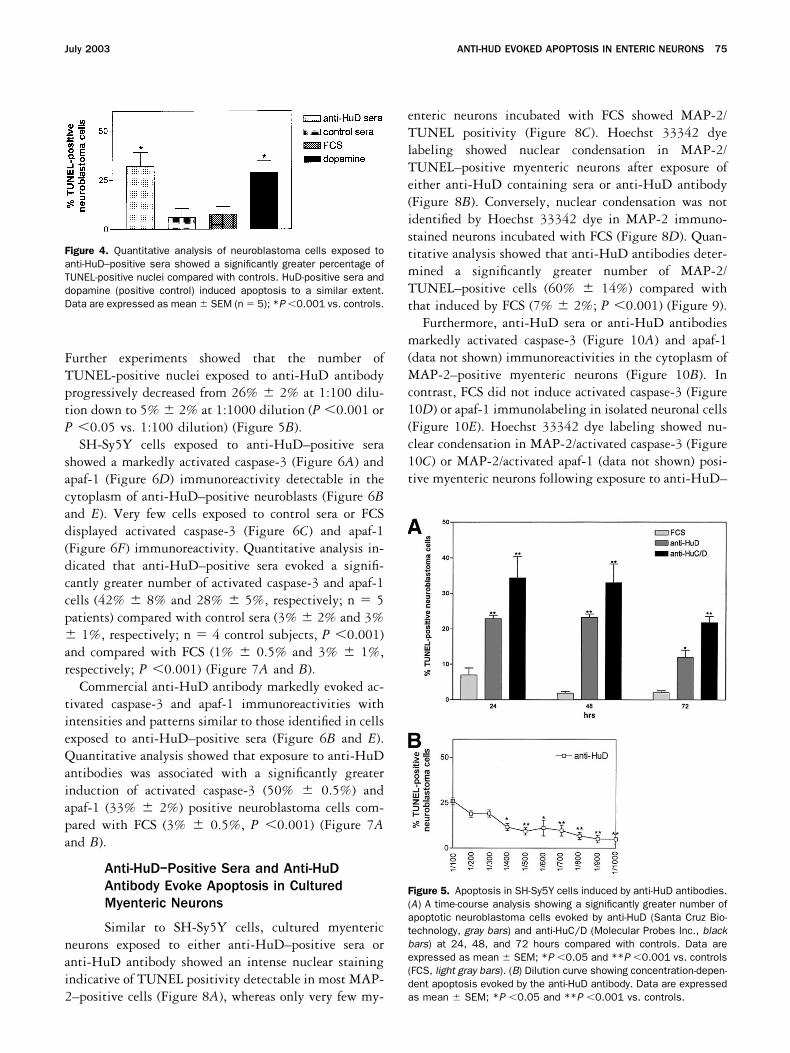
Further experiments showed that the number ofTUNEL-positive nuclei exposed to anti-HuD antibodyprogressively decreased from 26% � 2% at 1:100 dilu-tion down to 5% � 2% at 1:1000 dilution (P �0.001 orP �0.05 vs. 1:100 dilution) (Figure 5B).
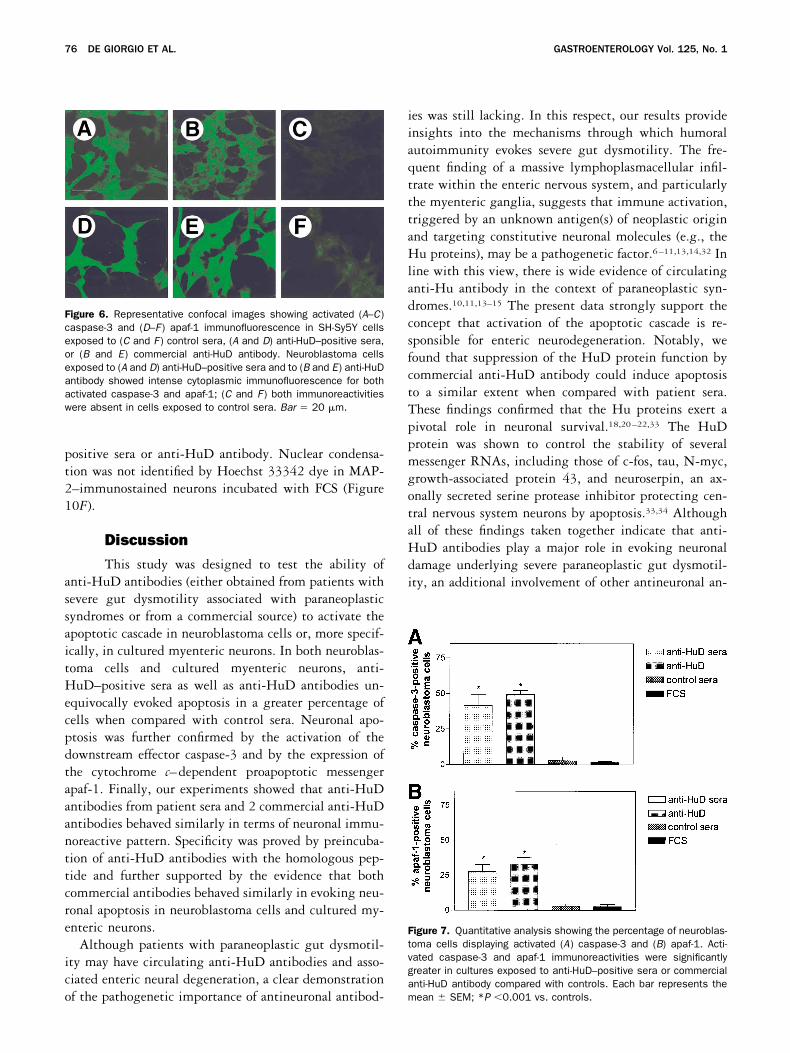
SH-Sy5Y cells exposed to anti-HuD–positive serashowed a markedly activated caspase-3 (Figure 6A) andapaf-1 (Figure 6D) immunoreactivity detectable in thecytoplasm of anti-HuD–positive neuroblasts (Figure 6Band E). Very few cells exposed to control sera or FCSdisplayed activated caspase-3 (Figure 6C) and apaf-1(Figure 6F) immunoreactivity. Quantitative analysis in-dicated that anti-HuD–positive sera evoked a signifi-cantly greater number of activated caspase-3 and apaf-1cells (42% � 8% and 28% � 5%, respectively; n � 5patients) compared with control sera (3% � 2% and 3%� 1%, respectively; n � 4 control subjects, P �0.001)and compared with FCS (1% � 0.5% and 3% � 1%,respectively; P �0.001) (Figure 7A and B).
Commercial anti-HuD antibody markedly evoked ac-tivated caspase-3 and apaf-1 immunoreactivities withintensities and patterns similar to those identified in cellsexposed to anti-HuD–positive sera (Figure 6B and E).Quantitative analysis showed that exposure to anti-HuDantibodies was associated with a significantly greaterinduction of activated caspase-3 (50% � 0.5%) andapaf-1 (33% � 2%) positive neuroblastoma cells com-pared with FCS (3% � 0.5%, P �0.001) (Figure 7Aand B).
Anti-HuD–Positive Sera and Anti-HuDAntibody Evoke Apoptosis in CulturedMyenteric Neurons
Similar to SH-Sy5Y cells, cultured myentericneurons exposed to either anti-HuD–positive sera oranti-HuD antibody showed an intense nuclear stainingindicative of TUNEL positivity detectable in most MAP-2–positive cells (Figure 8A), whereas only very few my-
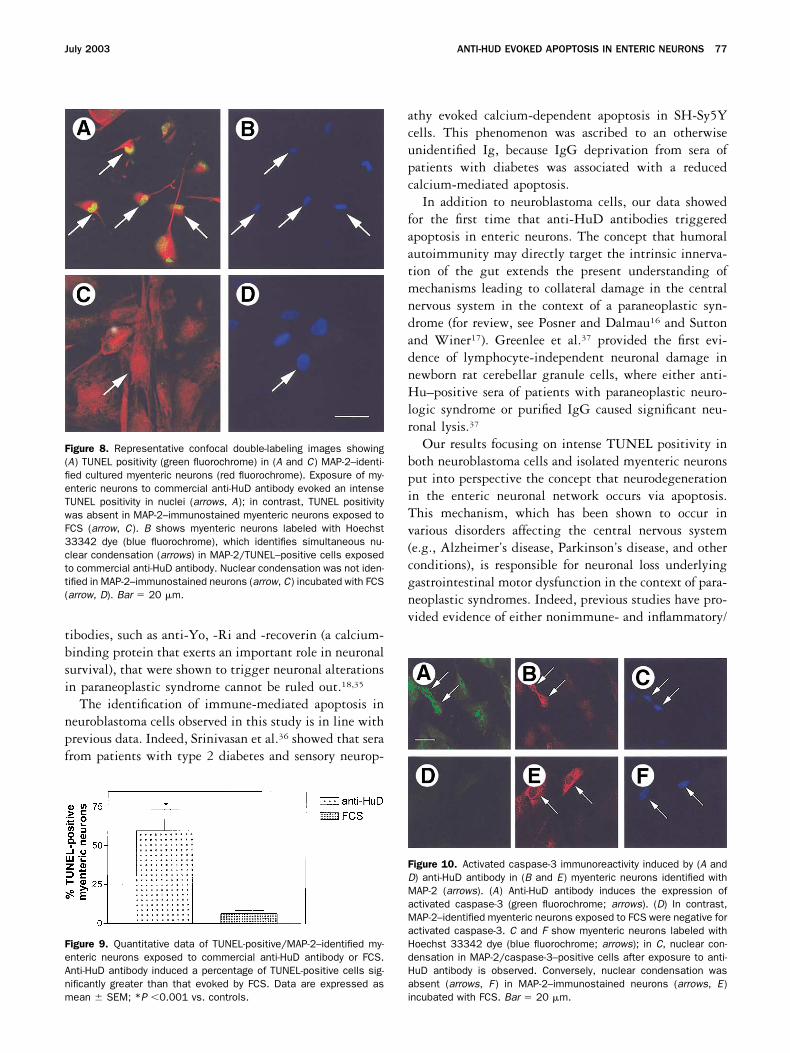
enteric neurons incubated with FCS showed MAP-2/TUNEL positivity (Figure 8C). Hoechst 33342 dyelabeling showed nuclear condensation in MAP-2/TUNEL–positive myenteric neurons after exposure ofeither anti-HuD containing sera or anti-HuD antibody(Figure 8B). Conversely, nuclear condensation was notidentified by Hoechst 33342 dye in MAP-2 immuno-stained neurons incubated with FCS (Figure 8D). Quan-titative analysis showed that anti-HuD antibodies deter-mined a significantly greater number of MAP-2/TUNEL–positive cells (60% � 14%) compared withthat induced by FCS (7% � 2%; P �0.001) (Figure 9).
Furthermore, anti-HuD sera or anti-HuD antibodiesmarkedly activated caspase-3 (Figure 10A) and apaf-1(data not shown) immunoreactivities in the cytoplasm ofMAP-2–positive myenteric neurons (Figure 10B). Incontrast, FCS did not induce activated caspase-3 (Figure10D) or apaf-1 immunolabeling in isolated neuronal cells(Figure 10E). Hoechst 33342 dye labeling showed nu-clear condensation in MAP-2/activated caspase-3 (Figure10C) or MAP-2/activated apaf-1 (data not shown) posi-tive myenteric neurons following exposure to anti-HuD–
Figure 4. Quantitative analysis of neuroblastoma cells exposed toanti-HuD–positive sera showed a significantly greater percentage ofTUNEL-positive nuclei compared with controls. HuD-positive sera anddopamine (positive control) induced apoptosis to a similar extent.Data are expressed as mean � SEM (n � 5); *P �0.001 vs. controls.
Figure 5. Apoptosis in SH-Sy5Y cells induced by anti-HuD antibodies.(A) A time-course analysis showing a significantly greater number ofapoptotic neuroblastoma cells evoked by anti-HuD (Santa Cruz Bio-technology, gray bars) and anti-HuC/D (Molecular Probes Inc., blackbars) at 24, 48, and 72 hours compared with controls. Data areexpressed as mean � SEM; *P �0.05 and **P �0.001 vs. controls(FCS, light gray bars). (B) Dilution curve showing concentration-depen-dent apoptosis evoked by the anti-HuD antibody. Data are expressedas mean � SEM; *P �0.05 and **P �0.001 vs. controls.
July 2003 ANTI-HUD EVOKED APOPTOSIS IN ENTERIC NEURONS 75
positive sera or anti-HuD antibody. Nuclear condensa-tion was not identified by Hoechst 33342 dye in MAP-2–immunostained neurons incubated with FCS (Figure10F).
DiscussionThis study was designed to test the ability of
anti-HuD antibodies (either obtained from patients withsevere gut dysmotility associated with paraneoplasticsyndromes or from a commercial source) to activate theapoptotic cascade in neuroblastoma cells or, more specif-ically, in cultured myenteric neurons. In both neuroblas-toma cells and cultured myenteric neurons, anti-HuD–positive sera as well as anti-HuD antibodies un-equivocally evoked apoptosis in a greater percentage ofcells when compared with control sera. Neuronal apo-ptosis was further confirmed by the activation of thedownstream effector caspase-3 and by the expression ofthe cytochrome c–dependent proapoptotic messengerapaf-1. Finally, our experiments showed that anti-HuDantibodies from patient sera and 2 commercial anti-HuDantibodies behaved similarly in terms of neuronal immu-noreactive pattern. Specificity was proved by preincuba-tion of anti-HuD antibodies with the homologous pep-tide and further supported by the evidence that bothcommercial antibodies behaved similarly in evoking neu-ronal apoptosis in neuroblastoma cells and cultured my-enteric neurons.
Although patients with paraneoplastic gut dysmotil-ity may have circulating anti-HuD antibodies and asso-ciated enteric neural degeneration, a clear demonstrationof the pathogenetic importance of antineuronal antibod-
ies was still lacking. In this respect, our results provideinsights into the mechanisms through which humoralautoimmunity evokes severe gut dysmotility. The fre-quent finding of a massive lymphoplasmacellular infil-trate within the enteric nervous system, and particularlythe myenteric ganglia, suggests that immune activation,triggered by an unknown antigen(s) of neoplastic originand targeting constitutive neuronal molecules (e.g., theHu proteins), may be a pathogenetic factor.6–11,13,14,32 Inline with this view, there is wide evidence of circulatinganti-Hu antibody in the context of paraneoplastic syn-dromes.10,11,13–15 The present data strongly support theconcept that activation of the apoptotic cascade is re-sponsible for enteric neurodegeneration. Notably, wefound that suppression of the HuD protein function bycommercial anti-HuD antibody could induce apoptosisto a similar extent when compared with patient sera.These findings confirmed that the Hu proteins exert apivotal role in neuronal survival.18,20–22,33 The HuDprotein was shown to control the stability of severalmessenger RNAs, including those of c-fos, tau, N-myc,growth-associated protein 43, and neuroserpin, an ax-onally secreted serine protease inhibitor protecting cen-tral nervous system neurons by apoptosis.33,34 Althoughall of these findings taken together indicate that anti-HuD antibodies play a major role in evoking neuronaldamage underlying severe paraneoplastic gut dysmotil-ity, an additional involvement of other antineuronal an-
Figure 6. Representative confocal images showing activated (A–C)caspase-3 and (D–F ) apaf-1 immunofluorescence in SH-Sy5Y cellsexposed to (C and F ) control sera, (A and D) anti-HuD–positive sera,or (B and E) commercial anti-HuD antibody. Neuroblastoma cellsexposed to (A and D) anti-HuD–positive sera and to (B and E) anti-HuDantibody showed intense cytoplasmic immunofluorescence for bothactivated caspase-3 and apaf-1; (C and F ) both immunoreactivitieswere absent in cells exposed to control sera. Bar � 20 �m.
Figure 7. Quantitative analysis showing the percentage of neuroblas-toma cells displaying activated (A) caspase-3 and (B) apaf-1. Acti-vated caspase-3 and apaf-1 immunoreactivities were significantlygreater in cultures exposed to anti-HuD–positive sera or commercialanti-HuD antibody compared with controls. Each bar represents themean � SEM; *P �0.001 vs. controls.
76 DE GIORGIO ET AL. GASTROENTEROLOGY Vol. 125, No. 1
tibodies, such as anti-Yo, -Ri and -recoverin (a calcium-binding protein that exerts an important role in neuronalsurvival), that were shown to trigger neuronal alterationsin paraneoplastic syndrome cannot be ruled out.18,35
The identification of immune-mediated apoptosis inneuroblastoma cells observed in this study is in line withprevious data. Indeed, Srinivasan et al.36 showed that serafrom patients with type 2 diabetes and sensory neurop-
athy evoked calcium-dependent apoptosis in SH-Sy5Ycells. This phenomenon was ascribed to an otherwiseunidentified Ig, because IgG deprivation from sera ofpatients with diabetes was associated with a reducedcalcium-mediated apoptosis.
In addition to neuroblastoma cells, our data showedfor the first time that anti-HuD antibodies triggeredapoptosis in enteric neurons. The concept that humoralautoimmunity may directly target the intrinsic innerva-tion of the gut extends the present understanding ofmechanisms leading to collateral damage in the centralnervous system in the context of a paraneoplastic syn-drome (for review, see Posner and Dalmau16 and Suttonand Winer17). Greenlee et al.37 provided the first evi-dence of lymphocyte-independent neuronal damage innewborn rat cerebellar granule cells, where either anti-Hu–positive sera of patients with paraneoplastic neuro-logic syndrome or purified IgG caused significant neu-ronal lysis.37
Our results focusing on intense TUNEL positivity inboth neuroblastoma cells and isolated myenteric neuronsput into perspective the concept that neurodegenerationin the enteric neuronal network occurs via apoptosis.This mechanism, which has been shown to occur invarious disorders affecting the central nervous system(e.g., Alzheimer’s disease, Parkinson’s disease, and otherconditions), is responsible for neuronal loss underlyinggastrointestinal motor dysfunction in the context of para-neoplastic syndromes. Indeed, previous studies have pro-vided evidence of either nonimmune- and inflammatory/
Figure 8. Representative confocal double-labeling images showing(A) TUNEL positivity (green fluorochrome) in (A and C) MAP-2–identi-fied cultured myenteric neurons (red fluorochrome). Exposure of my-enteric neurons to commercial anti-HuD antibody evoked an intenseTUNEL positivity in nuclei (arrows, A); in contrast, TUNEL positivitywas absent in MAP-2–immunostained myenteric neurons exposed toFCS (arrow, C). B shows myenteric neurons labeled with Hoechst33342 dye (blue fluorochrome), which identifies simultaneous nu-clear condensation (arrows) in MAP-2/TUNEL–positive cells exposedto commercial anti-HuD antibody. Nuclear condensation was not iden-tified in MAP-2–immunostained neurons (arrow, C) incubated with FCS(arrow, D). Bar � 20 �m.
Figure 9. Quantitative data of TUNEL-positive/MAP-2–identified my-enteric neurons exposed to commercial anti-HuD antibody or FCS.Anti-HuD antibody induced a percentage of TUNEL-positive cells sig-nificantly greater than that evoked by FCS. Data are expressed asmean � SEM; *P �0.001 vs. controls.
Figure 10. Activated caspase-3 immunoreactivity induced by (A andD) anti-HuD antibody in (B and E) myenteric neurons identified withMAP-2 (arrows). (A) Anti-HuD antibody induces the expression ofactivated caspase-3 (green fluorochrome; arrows). (D) In contrast,MAP-2–identified myenteric neurons exposed to FCS were negative foractivated caspase-3. C and F show myenteric neurons labeled withHoechst 33342 dye (blue fluorochrome; arrows); in C, nuclear con-densation in MAP-2/caspase-3–positive cells after exposure to anti-HuD antibody is observed. Conversely, nuclear condensation wasabsent (arrows, F ) in MAP-2–immunostained neurons (arrows, E)incubated with FCS. Bar � 20 �m.
July 2003 ANTI-HUD EVOKED APOPTOSIS IN ENTERIC NEURONS 77

immune-mediated neurodegeneration in the entericnervous system of patients with severe gut dysmotil-ity.1–6,8,9,11–14,38 The latter mechanism is supported bythe evidence shown in this report that apoptosis is actu-ally triggered by anti-HuD autoantibodies. Enteric gan-glia are actually sheltered by a barrier that resembles theblood-brain barrier, and one may wonder about themechanisms through which circulating antibodies canget into the plexus. Although under normal circum-stances the enteric innervation is preserved from exoge-nous noxae, activation of the immune system may resultin a disruption of the barrier and facilitate the access ofimmunocompetent cells to the myenteric microenviron-ment, as reflected by the histopathologic changes foundin myenteric ganglionitis.1–5,7,10–14 As a result, the al-teration of the blood-enteric barrier makes it plausiblethat even specific neuronal autoantibodies may reachtheir target.
Activation of caspases, namely caspase-3, seems irre-versibly associated with apoptotic cell death.38 This pro-apoptotic messenger is a key signal for the activation ofboth the mitochondria-dependent and the mitochon-dria-independent apoptotic pathways. Therefore, thecaspase-3 activation induced by anti-HuD antibodies inSH-Sy5Y cells and myenteric neurons suggests that ap-optosis may occur in paraneoplastic gastrointestinal neu-rodegenerative diseases. Regarding the mitochondria-dependent apoptotic pathway, a regulatory step forcaspase-3 activation triggered by various noxae occurs atthe level of cytochrome c release,39,40 which initiatescaspase-3 activation via apaf-1, a 130-kilodalton cyto-solic monomer. The complex apaf-1/cytochrome c recruitsprocaspase-9, which ultimately leads to caspase-3 activa-tion and related downstream pathway ending with apo-ptosis.39–41 In this study, we showed that either seracontaining anti-HuD or commercial anti-HuD antibod-ies induced apoptosis through caspase-3 activation bothin neuroblastoma cells and in myenteric neurons. Inaddition, the involvement of apaf-1 suggests that theapoptotic pathway is, at least in part, mitochondriadependent. Indeed, recent data indicate that mitochon-drial dysfunction represents a crucial event in neuronalinjury in the central and peripheral nervous system.38
Irreversible mitochondrial depolarization triggered byexcitatory amino acids, such as glutamate, is associatedwith an initial necrosis followed by a late dorsal rootganglion apoptosis in streptozocin-induced diabetes inrats.36 Although it is likely that apoptosis may be pre-cipitated by these mechanisms, the role played by hu-moral autoimmunity on mitochondrial dysfunction in
the context of gut paraneoplastic syndromes deservesfurther analysis.
In conclusion, the present report, which shows thatanti-HuD antibodies may contribute directly to entericneuron impairment, offers new insights into the patho-physiology of gut dysmotility related to paraneoplasticsyndromes and suggests that neurodegeneration occursvia autoimmune-mediated neuronal apoptosis. The acti-vation of apaf-1 suggests that programmed cell deathinvolves, at least in part, a mitochondria-dependentpathway in neuroblastoma cells and cultured myentericneurons. We believe that the novelty of our results standsmainly in the proof of the concept that circulating an-tineuronal antibodies may indeed commit cultured neu-rons to programmed cell death. Obviously, this does notnecessarily mean that neuronal apoptosis must invariablyoccur in vivo in the presence of circulating antineuronalautoantibodies. A possible scenario is disruption of theenteric microenvironment due to cross-reactivity be-tween tumoral antigens and neuronal proteins, whichmay occur at any gut level. This may explain why not allpatients with paraneoplastic syndrome have gut dysmo-tility and why dysmotility may involve either a specificgut segment or the whole gastrointestinal tract.
References1. De Giorgio R, Barbara G, Stanghellini V, De Ponti F, Salvioli B,
Tonini M, Velio P, Bassotti G, Corinaldesi R. Clinical and morpho-functional features of idiopathic myenteric ganglionitis underlyingsevere intestinal motor dysfunction: a study of three cases. Am JGastroenterol 2002;97:2454–2459.
2. De Giorgio R, Barbara G, Stanghellini V, Cogliandro RF, Arrigoni A,Santini D, Ceccarelli C, Salvioli B, Rossini FP, Corinaldesi R.Idiopathic myenteric ganglionitis underlying intractable vomitingin a young adult. Eur J Gastroenterol Hepatol 2000;12:613–616.
3. Smith VV, Gregson N, Foggensteiner L, Neale G, Milla PJ. Ac-quired intestinal aganglionosis and circulating autoantibodieswithout neoplasia or other neural involvement. Gastroenterology1997;112:1366–1371.
4. Horoupian DS, Kim Y. Encephalomyeloneuropathy with ganglion-itis of the myenteric plexuses in the absence of cancer. AnnNeurol 1982;11:628–632.
5. Tanowitz HB, Kirchhoff LV, Simon D, Morris SA, Weiss LM, WittnerM. Chagas’ disease. Clin Microbiol Rev 1992;5:400–419.
6. Krishnamurthy S, Schuffler MD. Pathology of neuromuscular dis-orders of the small intestine and colon. Gastroenterology 1987;93:610–639.
7. De Giorgio R, Stanghellini V, Barbara G, Corinaldesi R, De Ponti F,Tonini M, Bassotti G, Sternini C. Primary enteric neuropathiesunderlying gastrointestinal motor dysfunction. Scand J Gastroen-terol 2000;35:114–122.
8. Wood JD. Neuropathy in the brain-in-the-gut. Eur J GastroenterolHepatol 2000;12:597–600.
9. Wood JD, Alpers DH, Andrews PL. Fundamentals of neurogastro-enterology. Gut 1999;45(Suppl 2):ii6–ii16.
10. Schuffler MD, Baird HW, Fleming CR, Bell CE, Bouldin TW, Mal-agelada JR, McGill DB, LeBauer SM, Abrams M, Love J. Intestinalpseudo-obstruction as the presenting manifestation of small-cell
78 DE GIORGIO ET AL. GASTROENTEROLOGY Vol. 125, No. 1

carcinoma of the lung: A paraneoplastic neuropathy of the gas-trointestinal tract. Ann Intern Med 1983;98:129–134.
11. Lennon VA, Sas DF, Busk MF, Scheithauer B, Malagelada JR,Camilleri M, Miller LJ. Enteric neuronal autoantibodies inpseudoobstruction with small-cell lung carcinoma. Gastroenter-ology 1991;100:137–142.
12. Gerl A, Storck M, Schalhorn A, Muller-Hocker J, Jauch KW, Schild-berg FW, Wilmanns W. Paraneoplastic chronic intestinalpseudoobstruction as a rare complication of bronchial carcinoid.Gut 1992;33:1000–1003.
13. Schobinger-Clement S, Gerber HA, Stallmach T. Autoaggressiveinflammation of the myenteric plexus resulting in intestinalpseudoobstruction. Am J Surg Pathol 1999;23:602–606.
14. Lee HR, Lennon VA, Camilleri M, Prather CM. Paraneoplasticgastrointestinal motor dysfunction: clinical and laboratory char-acteristics. Am J Gastroenterol 2001;96:373–379.
15. Kiers L, Altermatt HJ, Lennon VA. Paraneoplastic anti-neuronalnuclear IgG autoantibodies (type I) localize antigen in small celllung carcinoma. Mayo Clin Proc 1991;66:1209–1216.
16. Posner JB, Dalmau JO. Paraneoplastic syndromes affecting thecentral nervous system. Annu Rev Med 1997;48:157–166.
17. Sutton I, Winer JB. The immunopathogenesis of paraneoplasticneurological syndromes. Clin Sci (Lond) 2002;102:475–486.
18. Musunuru K, Darnell RB. Paraneoplastic neurologic disease an-tigens: RNA-binding proteins and signaling proteins in neuronaldegeneration. Annu Rev Neurosci 2001;24:239–262.
19. Lin Z, Gao N, Hu H-Z, Liu S, Gao C, Kim G, Ren C, Xia Y, Peck OC,Wood JD. Immunoreactivity of Hu proteins facilitates identifica-tion of myenteric neurones in guinea-pig small intestine. Neuro-gastroenterol Motil 2002;14:197–204.
20. Okano HJ, Darnell RB. A hierarchy of Hu RNA binding proteins indeveloping and adult neurons. J Neurosci 1997;17:3024–3037.
21. Wakamatsu Y, Weston JA. Sequential expression and role of HuRNA-binding proteins during neurogenesis. Development 1997;124:3449–3460.
22. Peng SS, Chen CY, Xu N, Shyu AB. RNA stabilization by theAU-rich element binding protein, HuR, an ELAV protein. EMBO J1998;17:3461–3470.
23. Caras SD, McCallum HR, Brashear HR, Smith TK. The effect ofhuman antineuronal antibodies on the ascending excitatory reflexand peristalsis in isolated guinea pig ileum. Is the paraneoplasticsyndrome a motor neuron disorder? (abstr) Gastroenterology1996;110:A643.
24. Lennon VA. Anti-Purkinje cell cytoplasmic and neuronal nuclearantibodies aid diagnosis of paraneoplastic autoimmune neuro-logical disorders. J Neurol Neurosurg Psychiatry 1989;52:1438–1439.
25. Voltz RD, Posner JB, Dalmau J, Graus F. Paraneoplastic enceph-alomyelitis: an update of the effects of the anti-Hu immuneresponse on the nervous system and tumour. J Neurol NeurosurgPsychiatry 1997;63:133–136.
26. Van den Berghe P, Tack J, Coulie B, Andrioli A, Bellon E, JanssensJ. Synaptic transmission induces transient Ca2� concentrationchanges in cultured myenteric neurons. Neurogastroenterol Motil2000;12:117–124.
27. Jessen KR, Saffrey MJ, Burnstock G. The enteric nervous systemin tissue culture I: Cell types and their interactions in explants ofthe myenteric and submucous plexuses from guinea pig, rabbitand rat. Brain Res 1983;262:17–35.
28. Hanani M, Xia Y, Wood JD. Myenteric ganglia from the adult
guinea-pig small intestine in tissue culture. NeurogastroenterolMotil 1994;6:103–118.
29. Migheli A, Attanasio A, Lee WH, Bayer SA, Ghetti B. Detection ofapoptosis in weaver cerebellum by electron microscopic in situend-labeling of fragmented DNA. Neurosci Lett 1995;199:53–56.
30. Li X, Traganos F, Melamed MR, Darzynkiewicz Z. Single-stepprocedure for labeling DNA strand breaks with fluores. Cytometry1995;20:172–180.
31. Junn E, Mouradian MM. Apoptotic signaling in dopamine-inducedcell death: the role of oxidative stress, p38 mitogen-activatedprotein kinase, cytochrome c and caspases. J Neurochem 2001;78:374–383.
32. Collins SM. The immunomodulation of enteric neuromuscularfunction: implications for motility and inflammatory disorders.Gastroenterology 1996;111:1683–1699.
33. Perrone-Bizzozero N, Bolognani F. Role of HuD and other RNA-binding proteins in neural development and plasticity. J NeurosciRes 2002;68:121–126.
34. Cuadrado A, Navarro-Yubero C, Furneaux H, Kinter J, Sondereg-ger P, Munoz A. HuD binds to three AU-rich sequences in the3�-UTR of neuroserpin mRNA and promotes the accumulation ofneuroserpin mRNA and protein. Nucleic Acids Res 2002;30:2202–2211.
35. Vincent A, Honnorat J, Antoine JC, Giometto B, Dalmau J, Lang B.Autoimmunity in paraneoplastic neurological disorders. J Neuro-immunol 1998;84:105–109.
36. Srinivasan S, Stevens MJ, Sheng H, Hall KE, Wiley JW. Serumfrom patients with type 2 diabetes with neuropathy inducescomplement-independent, calcium-dependent apoptosis in cul-tured neuronal cells. J Clin Invest 1998;102:1454–1462.
37. Greenlee JE, Parks TN, Jaeckle KA. Type IIa (’anti-Hu’) antineu-ronal antibodies produce destruction of rat cerebellar granuleneurons in vitro. Neurology 1993;43:2049–2054.
38. Hall KE, Wiley JW. Neural injury, repair and adaptation in the GItract. I. New insights into neuronal injury: a cautionary tale. Am JPhysiol 1998;274:G978–G983.
39. Green DR. Apoptotic pathways: the roads to ruin. Cell 1998;94:695–698.
40. Zimmermann KC, Bonzon C, Green DR. The machinery of pro-grammed cell death. Pharmacol Ther 2001;92:57–70.
41. Nijhawan D, Honarpour N, Wang X. Apoptosis in neural develop-ment and disease. Annu Rev Neurosci 2000;23:73–87.
Received October 25, 2002. Accepted April 3, 2003.Address requests for reprints to: Roberto De Giorgio, M.D., Ph.D.,
Department of Internal Medicine and Gastroenterology, St. Orsola-Malpighi Hospital, Via Massarenti, 9, I-40138, Bologna, Italy. e-mail:[email protected]; fax: (39) 051-345864.Supported in part by grants from the Italian Ministry of University,
Research, Science and Technology (COFIN 1999 project no.9905222532 and 9906198133) and National Research Council(CNRC0008_02).The authors thank Dr. Gary M. Mawe (Department of Anatomy and
Neurobiology, University of Vermont, Burlington, VT) for insightful sug-gestions and Silvia Allaria (Department of Neuroscience, University ofMilan, Milan, Italy) for skillful technical assistance in performing theHuD characterization in some of the patients.
July 2003 ANTI-HUD EVOKED APOPTOSIS IN ENTERIC NEURONS 79
Related Documents











