Antagonistic regulation of convergent extension movements in Xenopus by Wnt/b-catenin and Wnt/Ca 21 signaling Michael Ku ¨hl a, * , Karin Geis b , Laird C. Sheldahl c , Tobias Pukrop b , Randall T. Moon c , Doris Wedlich b,d a Abteilung Entwicklungsbiochemie, Junior Group SFB 271, Universita ¨t Go ¨ttingen, Humboldtallee 23, D-37073 Go ¨ttingen, Germany b Abteilung Biochemie, Universita ¨t Ulm, Albert-Einstein-Allee 11, D-89081 Ulm, Germany c Howard Hughes Medical Institute, Department of Pharmacology, and Center for Developmental Biology, University of Washington School of Medicine, Seattle, WA 98195-7530, USA d Universita ¨t Karlsruhe, Zoologisches Institut II, Kaiserstraße 12, D-76131 Karlsruhe, Germany Received 8 March 2001; received in revised form 26 April 2001; accepted 3 May 2001 Abstract Convergent extension movements are the main driving force of Xenopus gastrulation. A fine-tuned regulation of cadherin-mediated cell– cell adhesion is thought to be required for this process. Members of the Wnt family of extracellular glycoproteins have been shown to modulate cadherin-mediated cell–cell adhesion, convergent extension movements, and cell differentiation. Here we show that endogenous Wnt/b-catenin signaling activity is essential for convergent extension movements due to its effect on gene expression rather than on cadherins. Our data also suggest that XLEF-1 rather than XTCF-3 is required for convergent extension movements and that XLEF-1 functions in this context in the Wnt/b-catenin pathway to regulate Xnr-3. In contrast, activation of the Wnt/Ca 21 pathway blocks convergent extension movements, with potential regulation of the Wnt/b-catenin pathway at two different levels. PKC, activated by the Wnt/Ca 21 pathway, blocks the Wnt/b-catenin pathway upstream of b-catenin and phosphorylates Dishevelled. CamKII, also activated by the Wnt/Ca 21 pathway, inhibits the Wnt/b-catenin signaling cascade downstream of b-catenin. Thus, an opposing cross-talk of two distinct Wnt signaling cascades regulates convergent extension movements in Xenopus. q 2001 Elsevier Science Ireland Ltd. All rights reserved. 1. Introduction Gastrulation in vertebrates is a complex merging of temporally and spatially coordinated cell and tissue move- ments that leads to a reorganized embryo composed of three germ layers. In Xenopus, gastrulation is governed by region-specific movements: (i) epiboly of the animal cap, (ii) migration of head mesoderm on the blastocoel roof, (iii) convergent extension of the involuting marginal zone (Keller, 1986), and (iv) vegetal rotation (Winklbauer and Schu ¨rfeld, 1999). Migration of head mesoderm requires cell-substrate adhesion to fibronectin deposited on the blas- tocoel roof (Winklbauer, 1994). For epiboly and convergent extension that are based on radial and lateral cell intercala- tions, regulation of cell–cell adhesion is essential. Cell–cell adhesion in the early embryo is mainly mediated by cadher- ins, calcium-dependent cell–cell adhesion molecules (reviewed in Ku ¨hl and Wedlich, 1996). Misexpression of cadherin mutants affects cell adhesion and gastrulation movements (Lee and Gumbiner, 1995; Broders and Thiery, 1995; Ku ¨hl et al., 1996; Ku ¨hl and Wedlich, 1996; Torres et al., 1996). Several attempts were made to decipher how signals involved in mesoderm induction and patterning also control cell movements. All of these studies end up with the observation that different types of cell movements are differentially regulated by growth factors (Howard and Smith, 1993). However, the detailed molecular basis for these control mechanisms has remained obscure. Several lines of evidence suggest that members of the Wnt family are involved in regulating convergent exten- sion. Wnt genes encode for glycoproteins that are involved in cell fate decisions and cell differentiation. A role for Wnt proteins has also been shown in regulating cadherin- mediated cell adhesion and cytoskeletal reorganization (Wodarz and Nusse, 1998). In PC-12 cells, transfection of Wnt-1 enhances cell adhesion by increasing the steady state levels of b-catenin, a known cytoplasmic interactor of cadherins (Bradley et al., 1993). Xwnt-5A, instead, leads to a decrease in cell adhesion when overexpressed in Xenopus Mechanisms of Development 106 (2001) 61–76 0925-4773/01/$ - see front matter q 2001 Elsevier Science Ireland Ltd. All rights reserved. PII: S0925-4773(01)00416-6 www.elsevier.com/locate/modo * Corresponding author. Tel.: 149-551-399857; fax: 149-551-3912198. E-mail address: [email protected] (M. Ku ¨hl).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Antagonistic regulation of convergent extension movements in Xenopusby Wnt/b-catenin and Wnt/Ca21 signaling
Michael KuÈhla,*, Karin Geisb, Laird C. Sheldahlc, Tobias Pukropb,Randall T. Moonc, Doris Wedlichb,d
aAbteilung Entwicklungsbiochemie, Junior Group SFB 271, UniversitaÈt GoÈttingen, Humboldtallee 23, D-37073 GoÈttingen, GermanybAbteilung Biochemie, UniversitaÈt Ulm, Albert-Einstein-Allee 11, D-89081 Ulm, Germany
cHoward Hughes Medical Institute, Department of Pharmacology, and Center for Developmental Biology, University of Washington School of Medicine,
Seattle, WA 98195-7530, USAdUniversitaÈt Karlsruhe, Zoologisches Institut II, Kaiserstraûe 12, D-76131 Karlsruhe, Germany
Received 8 March 2001; received in revised form 26 April 2001; accepted 3 May 2001
Abstract
Convergent extension movements are the main driving force of Xenopus gastrulation. A ®ne-tuned regulation of cadherin-mediated cell±
cell adhesion is thought to be required for this process. Members of the Wnt family of extracellular glycoproteins have been shown to
modulate cadherin-mediated cell±cell adhesion, convergent extension movements, and cell differentiation. Here we show that endogenous
Wnt/b-catenin signaling activity is essential for convergent extension movements due to its effect on gene expression rather than on
cadherins. Our data also suggest that XLEF-1 rather than XTCF-3 is required for convergent extension movements and that XLEF-1
functions in this context in the Wnt/b-catenin pathway to regulate Xnr-3. In contrast, activation of the Wnt/Ca21 pathway blocks convergent
extension movements, with potential regulation of the Wnt/b-catenin pathway at two different levels. PKC, activated by the Wnt/Ca21
pathway, blocks the Wnt/b-catenin pathway upstream of b-catenin and phosphorylates Dishevelled. CamKII, also activated by the Wnt/Ca21
pathway, inhibits the Wnt/b-catenin signaling cascade downstream of b-catenin. Thus, an opposing cross-talk of two distinct Wnt signaling
cascades regulates convergent extension movements in Xenopus. q 2001 Elsevier Science Ireland Ltd. All rights reserved.
1. Introduction
Gastrulation in vertebrates is a complex merging of
temporally and spatially coordinated cell and tissue move-
ments that leads to a reorganized embryo composed of three
germ layers. In Xenopus, gastrulation is governed by
region-speci®c movements: (i) epiboly of the animal cap,
(ii) migration of head mesoderm on the blastocoel roof, (iii)
convergent extension of the involuting marginal zone
(Keller, 1986), and (iv) vegetal rotation (Winklbauer and
SchuÈrfeld, 1999). Migration of head mesoderm requires
cell-substrate adhesion to ®bronectin deposited on the blas-
tocoel roof (Winklbauer, 1994). For epiboly and convergent
extension that are based on radial and lateral cell intercala-
tions, regulation of cell±cell adhesion is essential. Cell±cell
adhesion in the early embryo is mainly mediated by cadher-
ins, calcium-dependent cell±cell adhesion molecules
(reviewed in KuÈhl and Wedlich, 1996). Misexpression of
cadherin mutants affects cell adhesion and gastrulation
movements (Lee and Gumbiner, 1995; Broders and Thiery,
1995; KuÈhl et al., 1996; KuÈhl and Wedlich, 1996; Torres et
al., 1996). Several attempts were made to decipher how
signals involved in mesoderm induction and patterning
also control cell movements. All of these studies end up
with the observation that different types of cell movements
are differentially regulated by growth factors (Howard and
Smith, 1993). However, the detailed molecular basis for
these control mechanisms has remained obscure.
Several lines of evidence suggest that members of the
Wnt family are involved in regulating convergent exten-
sion. Wnt genes encode for glycoproteins that are involved
in cell fate decisions and cell differentiation. A role for
Wnt proteins has also been shown in regulating cadherin-
mediated cell adhesion and cytoskeletal reorganization
(Wodarz and Nusse, 1998). In PC-12 cells, transfection
of Wnt-1 enhances cell adhesion by increasing the steady
state levels of b-catenin, a known cytoplasmic interactor of
cadherins (Bradley et al., 1993). Xwnt-5A, instead, leads to
a decrease in cell adhesion when overexpressed in Xenopus
Mechanisms of Development 106 (2001) 61±76
0925-4773/01/$ - see front matter q 2001 Elsevier Science Ireland Ltd. All rights reserved.
PII: S0925-4773(01)00416-6
www.elsevier.com/locate/modo
* Corresponding author. Tel.: 149-551-399857; fax: 149-551-3912198.
E-mail address: [email protected] (M. KuÈhl).

embryos (Torres et al., 1996). Together with other observa-
tions this bias in Wnt activity led to the model of two
functional distinct Wnt classes. Members of the Wnt-1/
Wg class (Wnt-1, -3a, -8a, and -8b) are able to induce
secondary axes formation in Xenopus embryos when
misexpressed on the ventral side. They transform
mammary C57mg cells and they increase cell adhesion.
On the other hand, members of the Wnt-5A class (Wnt-
4, -5A, and -11) do not induce secondary axes, they antag-
onize the function of the Wnt-1/Wg class and they decrease
cell adhesion (Torres et al., 1996). Members of both classes
also differentially affect convergent extension movements
during Xenopus gastrulation. Whereas members of the
Wnt-1/Wg family can enhance convergent extension
movements in the arti®cial animal cap assay (Sokol,
1993), members of the Wnt-5A class negatively in¯uence
convergent extension (Du et al., 1995; Torres et al., 1996;
Slusarski et al., 1997).
Recent observations suggest that the different classes of
Wnt activities activate different cytoplasmic signaling path-
ways. Members of the Wnt-1/Wg class stabilize cytoplas-
mic b-catenin that subsequently can enter the nucleus. Here
b-catenin interacts with transcription factors of the TCF/
LEF family and the heterodimer that functions as a tran-
scriptional activator (KuÈhl and Wedlich, 1997). The so-
called Wnt/b-catenin pathway also involves the phospho-
protein dishevelled and the serine-threonine kinase GSK-
3b, both of which function upstream of b-catenin. Members
of the Wnt-5A class can trigger intracellular Ca21-release
(Slusarski et al., 1997) and activate PKC and CamKII (Shel-
dahl et al., 1999; KuÈhl et al., 2000a,b). Further downstream
components of the Wnt/Ca21 pathway are not known. In
addition, Wnt-11 has been implicated in regulating conver-
gent extension movements through activating jun-N-term-
inal kinase (JNK) (Heisenberg et al., 2000; Tada and Smith,
2000; Wallingford et al., 2000). This Wnt/JNK pathway is
thought to resemble or be part of the planar cell polarity
pathway in Drosophila melanogaster (Shulman et al.,
1998). However, it is not clear whether there is a relation-
ship between the Wnt/Ca21 and planar cell polarity path-
ways.
The multiple proteins involved in Wnt signaling raise the
question of how Wnts regulate convergent extension move-
ments. In the present study we use in vitro `open face Keller
explants', which mimic the dorsal cell movements of
normal gastrulation, to investigate the role of the Wnt/b-
catenin and Wnt/Ca21 pathways in regulating convergent
extension movements. Our data indicate that the Wnt/b-
catenin pathway is absolutely required for proper conver-
gent extension movements due to regulation of Xnr-3
expression as a direct target gene of TCF/LEF factors.
The Wnt/Ca21 pathway inhibits convergent extension
movements via negative regulation of the Wnt/b-catenin
pathway at different levels. Thus, a ®ne-tuned balance of
opposing Wnt signaling effects serves to control convergent
extension movements during Xenopus gastrulation.
2. Results
2.1. Endogenous Wnt/b -catenin signaling is required for
convergent extension
We ®rst con®rmed reports that the Wnt/b-catenin pathway
enhances convergent extension movements in activin-treated
animal caps (Sokol, 1993) and that Wnt-5A blocks these
movements (Torres et al., 1996) (Fig. 1A). We then turned
to the Keller open face explant assay for molecular analyses,
since this assay is preferable to the animal cap assay in terms
of mimicking the normal movements of gastrulation.
Furthermore, Keller open face explants represent a very
sensitive method to study convergent extension movements.
Initial experiments using Keller explants revealed that over-
expressing Xwnt-8 RNA on the dorsal side of the embryo
does not block convergent extension movements whereas
injection of Xwnt-5A RNA does, as previously shown
(Torres et al., 1996; data not shown). Based on these obser-
vations we hypothesized that Xwnt-8-triggered Wnt/b-cate-
nin signaling might be required as a positive regulator of
convergent extension whereas Xwnt-5A-mediated Wnt/
Ca21 signaling might negatively regulate convergent exten-
sion. We therefore analyzed the effects of both Wnt signaling
pathways on convergent extension in greater detail by using
the Keller open face explant assay.
In order to quantify these experiments we established a
system that allows ef®cient scoring of open face Keller
explants even in large numbers, similar to the system used
by Tada and Smith (2000) for scoring the elongation of
animal caps. For this purpose, explants were con®ned to
one of the following groups after elongation process: (0)
no elongation and no constriction, (1) slight constriction
but no elongation, and (2) strong constriction and elongation
(as illustrated in Fig. 1B). In order to compare the results of
differently treated explants we present our data in two ways.
First, we give the data for each injected RNA by histograms
showing the distribution of each class given in percentages
(see Fig. 1C as an example for Wnt-injected explants). In
addition, we calculated the ratio of those explants showing
strong elongation (class 2) to those that show neither
constriction nor elongation (class 0). We found that these
convergent elongation coef®cients of different experiments
fall into four classes if at least 30 explants were performed
per injected RNA. Values of 1, 3, and 10, respectively, were
used to group the results of the Keller explant experiments
(see Fig. 1D). Based on this we considered that a convergent
extension coef®cient of between 3 and 10 de®nes normal
elongation behavior, whereas a value below 1 represents a
strong inhibition of convergent extension.
We ®rst analyzed the requirement of cytoplasmic b-cate-
nin for convergent extension movements. Overexpression of
XBDe349, an extracellular deleted version of XB/U-
cadherin that binds b-catenin with its intracellular domain,
results in a block of convergent extension movements (Fig.
2A). Similarly, injection of LEFDHMG that binds b-catenin
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±7662

but that is not suf®cient to activate downstream events of
Wnt/b-catenin signaling also interferes with convergent
extension (Fig. 2A). We could reverse this effect by coex-
pressing b-catenin but not XB/U-cadherin RNA (Fig. 2B).
The latter argues that the effect of LEFDHMG on conver-
gent extension is not due to a competition of this mutant
with endogenous cadherins for cytoplasmic b-catenin. To
analyze this effect in more detail we injected RNA encoding
for conductin, which is able to trigger b-catenin degradation
(Behrens et al., 1998). Consistent with the aforementioned
results, conductin disturbs convergent extension and again
this effect could be rescued by overexpressing b-catenin but
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±76 63
Fig. 1. Assay systems to judge convergent extension movements. (A) In animal caps of Xenopus laevis, addition of activin induces elongation of the caps. This
effect is blocked by injecting Xwnt-5A RNA into the animal pole of two-cell stage embryos. In contrast, injection of Xwnt-8 enhances elongation. A
representative experiment is shown. (B) Elongation of open face Keller explants is shown. For quantitative analyses, explants were grouped into different
classes. Normal constriction and strong elongation of the explant was classi®ed as `class 2'. Explants that just show constriction but no elongation were
grouped as `class 1', whereas explants that neither constrict nor elongate were of `class 0'. (C) Evaluation of prolactin-injected explants (control), Xwnt-5A- or
Xwnt-8-injected explants according to classes exempli®ed in (B). Xwnt-5A blocks convergent extension, while Xwnt-8 does not. Classi®cation of explants was
as given in Fig. 1C. Detailed results are given in Table 1. Calculated elongation coef®cients are also indicated. (D) Dividing class 2 (%) through class 0 (%) of a
given experiment results in the elongation coef®cient that allows rapid scoring and comparison of experiments. Values for the elongation coef®cient fall into
different groups, allowing a comparative judgment of different experiments. Each square in (D) represents an elongation coef®cient as given in Table 1. The y-
axis indicates the number n of times a given elongation coef®cient was found in the experiments presented here.

not XB/U-cadherin (Fig. 2C). We next asked whether this
effect of conductin is primarily due to its degrading effect of
cytoplasmic or cadherin-bound b-catenin. To answer this
question we prepared lysates of conductin or control-
injected embryos and separated membrane-bound and cyto-
plasmic b-catenin. Whereas conductin decreases the overall
amount of b-catenin it mainly affects cytoplasmic rather
than cadherin-associated b-catenin (Fig. 2D). In summary,
these data indicate that the positive effect and the require-
ment of Wnt/b-catenin signaling for convergent extension is
due to its stabilizing effect on cytoplasmic b-catenin.
2.2. XLEF-1 and Xnr-3 are required for convergent
extension
As b-catenin interacts with transcription factors of the
LEF/TCF family to regulate target gene expression, this
raises the possibility that nuclear events of Wnt/b-catenin
signaling are involved in regulating convergent extension.
This hypothesis is supported by our injecting an LEF-1
mutant, LEFDbBD, that still binds to DNA but does not
recruit the transcriptional activator b-catenin and thus
behaves as a dominant negative mutant with respect to
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±7664
Fig. 2. Blocking the Wnt/b-catenin pathway disturbs convergent extension movements. (A) Titration of cytoplasmic b-catenin by either a cadherin mutant
(XB-cadDe349) or an LEF-1 mutant (LEF-1DHMG) leads to a block of convergent extension movements. (B) The effect of LEF-1DHMG on convergent
extension movements can be rescued by coinjecting b-catenin but not XB/U-cadherin RNA. (C) Injection of conductin RNA blocks convergent extension
movements. This can be rescued by coinjecting b-catenin but not XB/U-cadherin RNA. Classi®cation of explants was as given in Fig. 1. Detailed results are
given in Table 1. Calculated elongation coef®cients are also indicated above the bar graphs. (D) Western blot analysis to monitor b-catenin expression (arrow
head) in conductin-injected embryos. Total lysates, glycoprotein fractions, or soluble fractions of control or conductin-injected embryos were loaded as
indicated. The equivalent of two embryos per lane was used.

TCF/LEF factors (Behrens et al., 1996; Gradl et al., 1999).
This mutant clearly blocks convergent extension (Fig. 3A),
an effect that cannot be rescued by coinjections of b-catenin
(Table 1). However, the LEFDbBD mutant does not discri-
minate between TCF or LEF members of the HMG-box
transcription factor family, and XLEF-1 (Molenaar et al.,
1998) and XTCF-3 (Molenaar et al., 1996) are both
expressed during gastrulation. Thus the function of either
might be blocked by LEFDbBD. Importantly, XLEF-1 as
well as murine LEF-1 are activators of transcription whereas
XTCF-3 acts as a transcriptional repressor (Pukrop et al.,
2001). Thus it is important to determine whether XLEF-1
and XTCF-3 have distinct roles in regulating convergent
extension. Coinjection of mLEF-1 RNA was able to rescue
the inhibiting effect of LEFDbBD, whereas XTCF-3 was
not, indicating that both transcription factors differ in their
ability to regulate convergent extension movements (Fig.
3A). Consistent with this observation is the result that a
single injection of LEF-1 does not interfere with convergent
extension whereas XTCF-3 is a strong inhibitor of this
process (Fig. 3B).
Three target genes of the Wnt/b-catenin pathway have
been described in the early Xenopus embryo: the TGF-bfamily member Xnr-3 and the closely related and functional
redundant homeobox transcription factors siamois and twin
(Brannon et al., 1997; Laurent et al., 1997; McKendry et al.,
1997). We therefore asked which of these target genes might
be suf®cient to restore proper convergent extension in open
face explants de®cient in Wnt/b-catenin signaling. We
found that Xnr-3, but not siamois, was able to rescue conver-
gent extension movements in LEFDbBD- or conductin-
injected explants (Fig. 3C,D). These data indicate that
LEF-1 but not XTCF-3 is required for regulation of conver-
gent extension movements through Wnt/b-catenin signaling
and that Xnr-3 is likely a key target gene for the regulation
of convergent extension.
2.3. The ability of Xnr-3 to rescue convergent extension
movements is not due to a dorsalizing activity
As inhibition of Wnt/b-catenin signaling results in ventra-
lized embryos (Behrens et al., 1996) and as Xnr-3 has been
shown to have a modest dorsalizing activity (Smith et al.,
1995) we next analyzed whether the ability of Xnr-3 to rescue
Wnt/b-catenin signaling de®cient explants just re¯ects
different migration behavior of dorsal versus ventral meso-
derm. As expected, injection of conductin resulted in ventra-
lized embryos (Fig. 3E) and in a down-regulation of several
dorsal marker genes (Fig. 3F). Coinjection of siamois led to a
rescue of dorsal marker gene expression (Fig. 3F) although
siamois was unable to restore proper cell migration behavior
in Keller explants (Fig. 3D). Conversely, Xnr-3 was unable to
rescue dorsal marker genes in conductin-injected embryos
(Fig. 3F) despite its ability to rescue convergent extension
(Fig. 3D). The panmesodermal marker Xbra remained unaf-
fected in these experiments (Fig. 3F). This differential ability
of Xnr-3 and siamois to rescue conductin ventralized
embryos is also re¯ected phenotypically (Fig. 3E). Consis-
tent with an inability to rescue dorsal gene expression, coin-
jection of Xnr-3 with conductin failed to rescue dorsal
structures. In contrast, coinjection of siamois with conductin
rescued dorso-anterior structures like cement glands (Fig.
3E). Some of these embryos exhibited a phenotype consistent
with gastrulation defects. Speci®cally, we found that siamois
is unable to rescue Xnr-3 expression in Wnt/b-catenin signal-
ing defective embryos and vice versa. In summary, the Wnt/
b-catenin pathway is required for both dorsal mesoderm
speci®cation and for convergent extension movements. Our
®ndings indicate that these two properties of the Wnt/b-cate-
nin pathways are governed by two different target genes,
siamois and Xnr-3, respectively.
2.4. The effect of Wnt-5A on convergent extension is due to
interference with Wnt/b-catenin signaling
We next con®rmed (Fig. 4A) that injection of Xwnt-5A
RNA into the dorsal marginal zone blocks elongation of
Keller open face explants as previous described (Torres et
al., 1996). Two downstream effectors of Xwnt-5A were
recently described in the early Xenopus embryo that are
activated by intracellular release of calcium ions, protein
kinase C (PKC) and Ca/calmodulin-dependent kinase II
(CamKII) (Sheldahl et al., 1999; KuÈhl et al., 2000a,b). We
®rst asked whether one or both of them were able to pheno-
copy the effect of Xwnt-5A on Keller explants. Treatment of
Keller explants with the PKC activator PMA resulted in an
inhibition of convergent extension movements (Fig. 4A). In
addition, injection of a constitutively active mutant of
CamKII, CamKII T286D, also blocks convergent extension
movements. Supporting a role for CamKII in regulating the
effects of Xwnt-5A on convergent extension, a kinase-dead
version of this kinase, CamKII K42M, was able to partially
reverse the inhibitory effects of Xwnt-5A on Keller explants
(Fig. 4A). These experiments demonstrate that both PKC
and CamKII are likely involved in mediating the Wnt/
Ca21 signal in regulating convergent extension.
Torres et al. (1996) reported that the effects of Xwnt-5A
on convergent extension can be rescued by coexpression of
full length N-cadherin. We therefore asked whether the
effect of Xwnt-5A or its downstream components can be
rescued by overexpressing XB/U-cadherin (MuÈller et al.,
1994) or Xcadherin-11 (Hadeball et al., 1998). We focused
on these cadherins rather than on the neural-speci®c N-
cadherin since both of them are expressed in the involuting
mesoderm during convergent extension. In addition, a role
for these cadherins in convergent extension movements has
been described (KuÈhl et al., 1996; KuÈhl and Wedlich, 1996;
Broders and Thiery, 1995; Lee and Gumbiner, 1995; Hade-
ball et al., 1998). Surprisingly, overexpression of neither
XB/U-cadherin nor Xcadherin 11 was suf®cient to rescue
the block of convergent extension by either Xwnt-5A or
constitutively active CamKII (Fig. 4B).
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±76 65

M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±7666
Fig. 3. Differential ability of downstream factors of the Wnt/b-catenin pathway to rescue b-catenin targeted embryos. (A) LEF-1 but not XTCF-3 was able to
rescue LEF-1DbBD-injected embryos. (B) XTCF-3 inhibits convergent extension while injection of LEF-1 RNA has no effect on convergent extension by
itself. (C) Xnr-3 but not siamois was able to completely rescue LEF-1DbBD-injected open face explants. (D) Xnr-3 but not siamois was able to fully rescue
conductin-injected open face explants. Classi®cation of explants was as given in Fig. 1. Detailed results are given in Table 1. Calculated elongation coef®cients
are also indicated above the bar graphs in panels (A)±(D). (E) Injection of conductin RNA results in ventralized embryos. This effect can be rescued by
injection of siamois but not Xnr-3 RNA. The DAI index indicates the degree of ventralization with 5 representing wild-type embryos and 0 representing
completely ventralized embryos. (F) Marker gene analysis of embryos injected as in Fig. 3E. Conductin down-regulates dorsal marker genes like goosecoid
(gsc), chordin or XFD-1/pintalavis. This effect can be reversed by injecting siamois but not Xnr-3 RNA. Note that Xnr-3 does not upregulate siamois and vice
versa. Signals indicated by a star arise from injected RNAs.

This raises the question of whether the effect of Xwnt-5A
on convergent extension is due to an interaction with the
Wnt/b-catenin pathway rather than its direct interference
with the cadherin-mediated cell adhesion system. To test
this hypothesis we coinjected b-catenin with Xwnt-5A
into dorsal marginal zones and found that these explants
showed a slight rescue in elongation behavior (Fig. 4C).
Xnr-3, the putative downstream target of b-catenin, was
much stronger than b-catenin in restoring convergent exten-
sion when coinjected with Wnt-5A RNA (Fig. 4C). We next
examined whether this rescuing activity of Xnr-3 might be
due to its dorsalizing activity. We ®rst needed to con®rm
that the effect of Xwnt-5A on convergent extension in
Keller explants and in whole embryos (Fig. 4D) is indepen-
dent of cell speci®cation. Xwnt-5A did not change expres-
sion of two dorsal marker genes, goosecoid and chordin
(Fig. 4E). We then found that dorsal injection of Xnr-3
did not change dorsal marker gene expression, either
alone or in combination with Xwnt-5A (Fig. 4E). We
conclude that the ability of Xnr-3 to rescue convergent
extension movements in Xwnt-5A-injected Keller explants
is independent of cell speci®cation and re¯ects the ability of
Xnr-3 to modulate cell migration.
In search for molecular mechanisms underlying this
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±76 67
Table 1
Effect of overexpressing different RNA constructs on convergent extension: detailed dataa
Probe Experiments Explants Classi®cation of explants Coef®cient Judgment
0 1 2
Nic 32 236 26 85 125 4.81 Normal
PPL 3 30 4 1 25 6.25 Normal
Xwnt-8 3 30 4 9 17 4.25 Normal
XBDe349 5 48 23 15 10 0.43 Inhibited
LEFDbBD 10 83 38 23 22 0.58 Inhibited
LEFDbBD 1 b-catenin 3 42 17 13 12 0.71 Inhibited
LEFDbBD 1 LEF 4 53 10 18 25 2.5 Slight rescue
LEFDbBD 1 XTCF-3 3 46 19 21 5 0.26 Inhibited
LEFDbBD 1 siamois 4 49 12 22 15 1.25 Slight rescue
LEFDbBD 1 Xnr-3 5 59 5 17 37 7.4 Rescue
LEFDHMG 11 92 36 36 20 0.56 Inhibited
LEFDHMG 1 b-catenin 4 46 10 20 16 1.6 Slight rescue
LEFDHMG 1 XB/U-cadherin 4 46 25 13 7 0.28 Inhibited
cdt 7 102 59 17 26 0.44 Inhibited
cdt 1 XB/U-cadherin 3 29 20 7 2 0.41 Inhibited
cdt 1 b-catenin 4 46 8 20 18 2.25 Slight rescue
cdt 1 siamois 4 49 18 19 12 0.67 Inhibited
cdt 1 Xnr-3 3 32 1 15 16 16 Rescue
LEF-1 5 27 3 7 17 5.67 Normal
TCF-3 3 43 24 17 2 0.08 Inhibited
Xwnt-5A 9 93 50 25 18 0.36 Inhibited
Xwnt-5A 1 XB/U-cadherin 3 44 23 15 6 0.26 Inhibited
Xwnt-5A 1 b-catenin 3 45 8 17 20 2.5 Slight rescue
Xwnt-5A 1 Xnr-3 3 28 1 13 14 14 Rescue
Xwnt-5A 1 Xcad11 3 30 18 8 4 0.2 Inhibited
Xwnt-5A 1 K42M 5 55 12 16 27 2.3 Slight rescue
CamKII T286D 8 96 54 20 22 0.41 Inhibited
T286D 1 XB/U-cadherin 4 55 30 18 7 0.23 Inhibited
T286D 1 b-catenin 3 40 19 12 9 0.47 Inhibited
T286D 1 Xnr-3 5 59 10 22 29 2.9 Slight rescue
T286D 1 LEF-1 4 51 10 20 21 2.1 Slight rescue
PMA 9 155 60 58 37 0.7 Inhibited
PMA 1 b-catenin 4 39 10 9 20 2.0 Slight rescue
PMA 1 dnGSK 3 62 9 29 23 2.56 Slight rescue
a Number of experiments indicates different batches of embryos that were used to cut explants. Number of explants represents the total number of explants
cut for this study. Only embryo batches that showed proper elongation in uninjected explants were used as control. Classi®cation of explants according to the
rules as presented in Fig. 1 as well as the elongation coef®cient are given. Abbreviations: nic, non-injected control; PPL, preprolactin; XBDe349, extracellular
deleted mutant of XB/U-cadherin; LEFDbBD, N-terminal deleted mutant of LEF lacking the b-catenin binding domain; LEFDHMG, mutant of LEF-1 lacking
the DNA binding domain; cdt, conductin; Xnr-3, Xenopus nodal-related gene 3; Xcad-11, Xenopus cadherin-11; K42M, kinase-dead mutant of CamKII;
CamKII T286D, constitutively active mutant of CamKII; PMA, phorpbol-12-myristate-13 acetate; dnGSK, dominant negative mutant of GSK-3b.
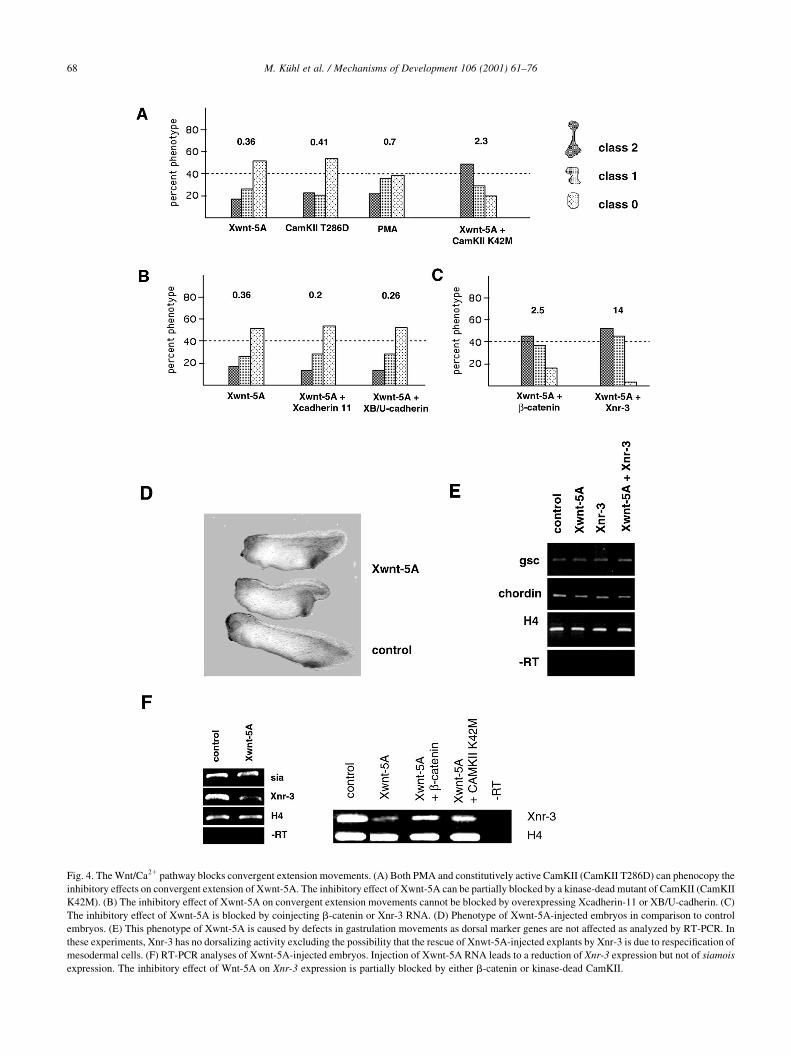
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±7668
Fig. 4. The Wnt/Ca21 pathway blocks convergent extension movements. (A) Both PMA and constitutively active CamKII (CamKII T286D) can phenocopy the
inhibitory effects on convergent extension of Xwnt-5A. The inhibitory effect of Xwnt-5A can be partially blocked by a kinase-dead mutant of CamKII (CamKII
K42M). (B) The inhibitory effect of Xwnt-5A on convergent extension movements cannot be blocked by overexpressing Xcadherin-11 or XB/U-cadherin. (C)
The inhibitory effect of Xwnt-5A is blocked by coinjecting b-catenin or Xnr-3 RNA. (D) Phenotype of Xwnt-5A-injected embryos in comparison to control
embryos. (E) This phenotype of Xwnt-5A is caused by defects in gastrulation movements as dorsal marker genes are not affected as analyzed by RT-PCR. In
these experiments, Xnr-3 has no dorsalizing activity excluding the possibility that the rescue of Xnwt-5A-injected explants by Xnr-3 is due to respeci®cation of
mesodermal cells. (F) RT-PCR analyses of Xwnt-5A-injected embryos. Injection of Xwnt-5A RNA leads to a reduction of Xnr-3 expression but not of siamois
expression. The inhibitory effect of Wnt-5A on Xnr-3 expression is partially blocked by either b-catenin or kinase-dead CamKII.
cross-talk of different Wnt pathways we asked whether
Xwnt-5A alters expression of Wnt/b-catenin target genes.
Most strikingly, Xnr-3 was down-regulated by Xwnt-5A,
while siamois expression was unaffected (Fig. 4F, left
panel). Xnr-3 down-regulation could be rescued by coinject-
ing either kinase-dead CamKII or b-catenin RNA together
with Xwnt-5A (Fig. 4F, right panel). These data strongly
suggest that the inhibitory effect of Wnt/Ca21 signaling is
due to an interaction of this pathway with the Wnt/b-catenin
pathway, which results in a speci®c down-regulation of Xnr-
3, the target gene important for convergent extension.
2.5. Wnt/Ca21 signaling blocks Wnt/b -catenin signaling at
multiple levels
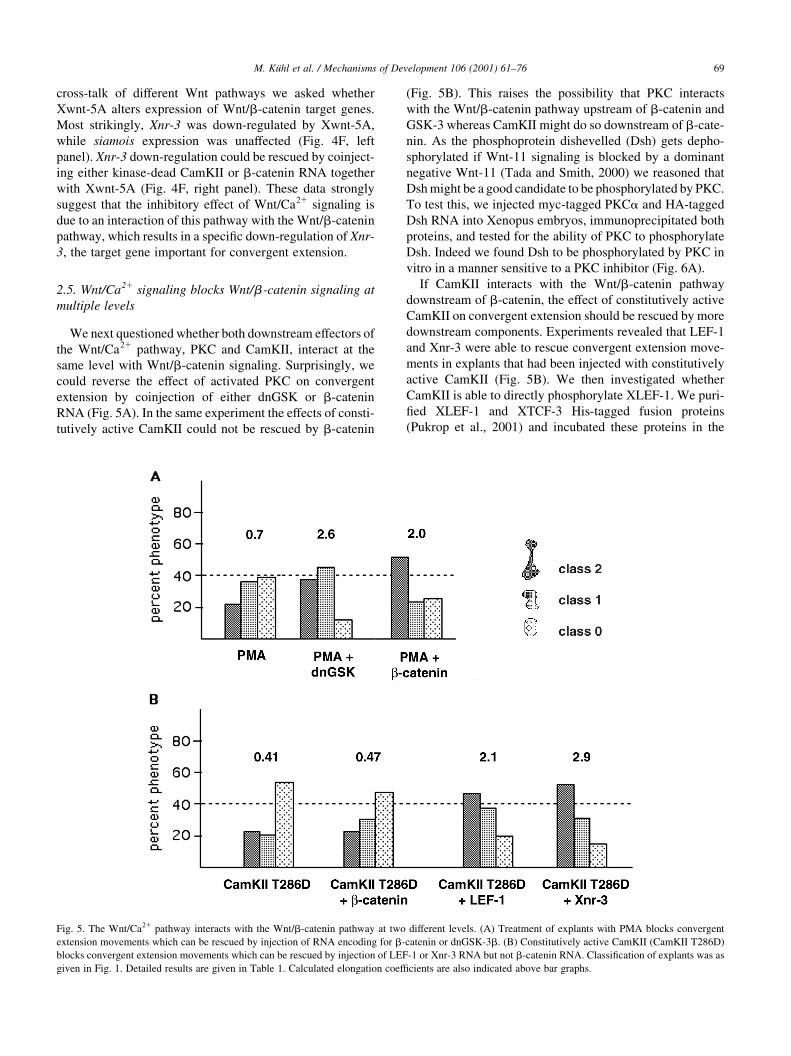
We next questioned whether both downstream effectors of
the Wnt/Ca21 pathway, PKC and CamKII, interact at the
same level with Wnt/b-catenin signaling. Surprisingly, we
could reverse the effect of activated PKC on convergent
extension by coinjection of either dnGSK or b-catenin
RNA (Fig. 5A). In the same experiment the effects of consti-
tutively active CamKII could not be rescued by b-catenin
(Fig. 5B). This raises the possibility that PKC interacts
with the Wnt/b-catenin pathway upstream of b-catenin and
GSK-3 whereas CamKII might do so downstream of b-cate-
nin. As the phosphoprotein dishevelled (Dsh) gets depho-
sphorylated if Wnt-11 signaling is blocked by a dominant
negative Wnt-11 (Tada and Smith, 2000) we reasoned that
Dsh might be a good candidate to be phosphorylated by PKC.
To test this, we injected myc-tagged PKCa and HA-tagged
Dsh RNA into Xenopus embryos, immunoprecipitated both
proteins, and tested for the ability of PKC to phosphorylate
Dsh. Indeed we found Dsh to be phosphorylated by PKC in
vitro in a manner sensitive to a PKC inhibitor (Fig. 6A).
If CamKII interacts with the Wnt/b-catenin pathway
downstream of b-catenin, the effect of constitutively active
CamKII on convergent extension should be rescued by more
downstream components. Experiments revealed that LEF-1
and Xnr-3 were able to rescue convergent extension move-
ments in explants that had been injected with constitutively
active CamKII (Fig. 5B). We then investigated whether
CamKII is able to directly phosphorylate XLEF-1. We puri-
®ed XLEF-1 and XTCF-3 His-tagged fusion proteins
(Pukrop et al., 2001) and incubated these proteins in the
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±76 69
Fig. 5. The Wnt/Ca21 pathway interacts with the Wnt/b-catenin pathway at two different levels. (A) Treatment of explants with PMA blocks convergent
extension movements which can be rescued by injection of RNA encoding for b-catenin or dnGSK-3b. (B) Constitutively active CamKII (CamKII T286D)
blocks convergent extension movements which can be rescued by injection of LEF-1 or Xnr-3 RNA but not b-catenin RNA. Classi®cation of explants was as
given in Fig. 1. Detailed results are given in Table 1. Calculated elongation coef®cients are also indicated above bar graphs.

presence of radioactively labeled ATP with commercially
available CamKII. These experiments revealed that XLEF-1
but not XTCF-3 is phosphorylated by CamKII (Fig. 6B).
In summary, our data suggest that the Wnt/Ca21 pathway
interacts with the Wnt/b-catenin signaling pathway at least
at two different levels. Whereas activated PKC inhibits the
Wnt/b-catenin pathway upstream of GSK3, activated
CamKII does so downstream of b-catenin at the level of
the transcription factor complex.
3. Discussion
Gastrulation is one of the most important steps in verte-
brate development leading to the formation of three germ
layers. Here we analyzed the role of two different Wnt
signaling pathways for proper cell migration during conver-
gent extension movements during Xenopus gastrulation.
Our data indicate that the Wnt/b-catenin pathway is
required for convergent extension by activating the TGF-b
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±7670
Fig. 6. Putative molecular mechanisms of Wnt pathway interactions. (A) Phosphorylation of Dsh by PKC in vitro. Embryos were single injected with either
XPKCa-myc or Dsh-HA. After mixing immunoprecipitates, a kinase reaction was performed in the presence or absence of PKC inhibitor as indicated. A signal
for phosphorylated Dsh can be observed only when PKC and Dsh are mixed in the absence of PKC inhibitor. The control panels show equal input of Dsh-HA
for each reaction as indicated. (B) Phosphorylation of XLEF-1 but not XTCF-3 by CamKII. Coomassie stain of puri®ed fusion proteins of XTCF-3 and XLEF-1
(Pukrop et al., 2001) (left panel). Equal amounts of fusion proteins as judged by SDS-PAGE and Bradford assay were incubated with CamKII and radioactively
labeled 32P-g-ATP. Only XLEF-1 was phosphorylated by CamKII but not XTCF-3 (right panel).

homolog Xnr-3. In contrast, activation of the Wnt/Ca21
pathway has a negative in¯uence on convergent extension
due to its inhibitory function on the Wnt/b-catenin pathway.
Thus, two different Wnt signaling pathways regulate
convergent extension movements in an opposing manner.
3.1. A role for the Wnt/b -catenin pathway for cell migration
Recent publications have suggested a role for Wnts in
regulating convergent extension. Overexpression of a
secreted form of the Wnt receptor Xfz-8, or the endogenous
Wnt inhibitor, FrzA, results in a block of convergent exten-
sion in vivo (Deardorff et al., 1998; Xu et al., 1998).
However, these publications did not investigate the down-
stream signaling pathways affected by these reagents. Here
we show unequivocally that a block of Wnt/b-catenin signal-
ing leads to inhibition of convergent extension movements.
This statement is based on several independent molecular
approaches. Injection of constructs that either directly tether
cytoplasmic b-catenin or lead to a degradation of cytoplas-
mic b-catenin results in disturbed morphogenetic move-
ments. We were able to show the speci®city of these effects
by coinjecting the more downstream component of the Wnt/
b-catenin pathway LEF-1. Although one might argue that the
inhibition of convergent extension in response to blocking
the Wnt/b-catenin pathway merely re¯ects ventralization of
explants we were able to show by rescue experiments that
Wnt/b-catenin signaling is required for both dorsal meso-
derm speci®cation and for convergent extension movements.
Speci®cally, we demonstrated that while siamois is able to
rescue dorsal marker gene expression in Wnt/b-catenin
signaling de®cient embryos, it does not restore proper cell
migration. On the other hand, Xnr-3 fails to rescue conductin
ventralized embryos but it fully rescues convergent exten-
sion movements. Thus, Wnt/b-catenin signaling is required
for convergent extension to occur due to its effect on Xnr-3
expression. This requirement of Wnt/b-catenin signaling for
proper gastrulation has recently also been shown in Droso-
phila (McEwen et al., 2000), and the C. elegans TCF homo-
log POP-1 has been linked to migration processes (Herman,
2001). In summary, our data and those of others indicate that
the Wnt/b-catenin pathway has to be active for proper cell
movements to occur.
Xnr-3 has been shown to be a direct target of Wnt signal-
ing (McKendry et al., 1997). Xnr-3 has also been shown to
alter cell behavior (Smith et al., 1995). Overexpressing Xnr-
3 in embryos results in tube-like extrusions and it has been
speculated that these protrusions might re¯ect an activity of
Xnr-3 in morphogenetic movements (Smith et al., 1995).
Here we showed that overexpressing Wnt-5A on the dorsal
side of the embryo results in a down-regulation of Xnr-3
expression whereas expression of siamois remained unaf-
fected in our hands. Thus, two target genes of the Wnt/b-
catenin pathway are differentially affected by the Wnt/Ca21
pathway. On ®rst sight this seems to be contradictory;
however, this ®nding clearly indicates that at least one
branch of the Wnt/Ca21 pathway exerts its function at the
level of transcription factor complexes. As these complexes
might differ between different target genes of a common
signaling pathway (Nishita et al., 2000) the effect of an
activated Wnt/Ca21 pathway on gene regulation might be
variable as here shown for Xnr-3 and siamois, respectively.
Finally, this observation also explains why Wnt-5A does not
ventralize Xenopus embryos since siamois expression
remains unaffected whereas gastrulation movements are
impaired as Xnr-3 expression is reduced. Our data that
Xnr-3 is involved in regulating convergent extension are
further supported by the observation that embryos that
have been injected with a dominant negative Xfz-7
construct also display problems with convergent extension
(Medina et al., 2000; Djiane et al., 2000; Sumanas et al.,
2000) and these embryos show reduced levels of Xnr-3
expression (Sumanas et al., 2000).
Finally, it is important to note here that our data are also
consistent with recent publications that made the statement
that Wnt/b-catenin signaling might not be required for
convergent extension movements (Wallingford et al.,
2000; Heisenberg et al., 2000). However, those experiments
were done in embryos injected with either a dishevelled
mutant or in silberblick (Wnt-11) mutant zebra®sh embryos
and thus in a situation in which a putative planar cell polar-
ity pathway (Wnt/JNK pathway) was blocked. Thus, the
Wnt/b-catenin pathway is not suf®cient to rescue conver-
gent extension movements if the Wnt/JNK pathway is inac-
tivated.
3.2. Interaction between Wnt/b -catenin and Wnt/Ca21
pathways
With respect to Wnt/Ca21 signaling we found that the
inhibitory effects of Wnt-5A on convergent extension
movements can be phenocopied by either activation of
PKC or overexpressing a constitutively active CamKII
mutant. These effects could be rescued by coinjecting single
components of the Wnt/b-catenin pathway. Consistent with
this antagonism of both the Wnt pathways is the ®nding that
expression of the b-catenin target gene Xnr-3 is reduced in
Xwnt-5A overexpressing embryos. Again, these data argue
for an important role for the Wnt/b-catenin pathway in
regulating convergent extension movements.
Furthermore, our data suggest that the two known cyto-
plasmic mediators of the Wnt/Ca21 signaling pathway, PKC
and CamKII, interact with the Wnt/b-catenin pathway at
two different levels. One candidate for phosphorylation by
PKC is Dsh, since the convergent extension effect of the
PKC activator PMA is reversed by b-catenin and by
dnGSK3 and both function downstream of Dsh in Wnt/b-
catenin signaling. Furthermore Dsh contains a DEP domain,
which recently has been suggested to interact with PKC
(Ponting and Bork, 1996). This hypothesis is also in agree-
ment with the observation that Xwnt-5A can antagonize the
axis-inducing ability of Xwnt-8 but not that of dnGSK
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±76 71

(Torres et al., 1996). Furthermore, Dsh is a phosphoprotein
that gets dephosphorylated in case of overexpressing
dnWnt-11 (Tada and Smith, 2000), which makes it a good
candidate for a PKC target. Finally, we here show that
PKCa is able to directly phosphorylate Dsh in vitro. Further
experiments will have to evaluate whether this also occurs
in vivo. If so, it is tempting to speculate that PKC also
in¯uences the signaling behavior of Dsh with respect to
Wnt/b-catenin and Wnt/JNK signaling.
In Xenopus, the effect of CamKII on Wnt/b-catenin
signaling might be mediated by direct phosphorylation of
XLEF-1 as shown here. However, we cannot exclude the
possibility that other components of the LEF-mediated tran-
scription factor complex are substrates for CamKII phos-
phorylation. There also remains the possibility that the
effect of CamKII on TCF/LEF factors might be indirectly
mediated by other factors regulated by CamKII. Further
biochemical analyses will have to analyze phosphorylation
of distinct proteins of this complex and the effect of these
phosphorylations on transcription factor assembly, target
gene activation, and, ®nally, convergent extension move-
ments. These experiments will also have to analyze whether
XLEF-1 is also phosphorylated in vivo and whether this
phosphorylation is linked to spatial differences in CamKII
signaling. As there is a ventral to dorsal gradient of CamKII
activity in the Xenopus embryo (KuÈhl et al., 2000a,b) this
implies that CamKII might be able to inactivate XLEF-1 on
the ventral side, while XLEF-1 is more active on the dorsal
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±7672
Fig. 7. Speculative model of how different Wnt signaling pathways ensure proper convergent extension movements. The Wnt/b-catenin pathway is activated
maternally and requires LEF-1 to activate Xnr-3. Activation of this pathway might be through the maternal Xwnt-8b (Cui et al., 1995) or directly through dorsal
accumulation of Dsh (Miller et al., 1999). Activation of the Wnt/Ca21 pathway by maternal Xwnt-11 or Xwnt-5A is also required for convergent extension
movements. The Wnt/Ca21 pathway activates PKC and CamKII that interact with the Wnt/b-catenin pathway at two different levels. PKC inhibits the Wnt/b-
catenin pathway upstream of b-catenin and GSK-3b probably by phosphorylation of Dsh. As Dsh acts in two different Wnt pathways, phosphorylation of Dsh
might function as a molecular switch (S) in this context. CamKII regulates the Wnt/b-catenin pathway downstream of b-catenin at the level of the LEF-1-
mediated transcription factor complex.

side. With respect to the target gene Xnr-3, the phosphor-
ylation of XLEF-1 together with the dorsal enrichment of b-
catenin thus might contribute to the dorsoventral gradient of
Xnr-3 expression. This dorsoventral gradient of Xnr-3
expression due to dorsal b-catenin and ventral CamKII
signaling may also explain why dorsal cells show a more
active migration behavior in convergent extension move-
ments than ventral cells.
3.3. Is there a correlation between Wnt/Ca21 and Wnt/JNK
signaling?
Recent publications suggest that Xwnt-11 is able to acti-
vate JNK in a Dsh-dependent manner (Wallingford et al.,
2000; Heisenberg et al., 2000; Tada and Smith, 2000). As
Wnt-11 is also able to activate PKC and CamKII (Sheldahl et
al., 1999; KuÈhl et al., 2000a) this raises the question about the
relationship between these two pathways. Activation of JNK
by Frizzled has been shown to depend on Dsh and small
GTPases of the rho family (Strutt et al., 1997; Boutros et
al., 1998) although activation of JNK by Dsh seems to be
independent of the small GTPases (Li et al., 1999). It seems
noteworthy that in cell culture systems both Wnt-1 and Wnt-
5A have been able to activate JNK (Li et al., 1999), and JNK
has also been demonstrated to be Ca-sensitive in calmodulin-
dependent and -independent manners (Enslen et al., 1996;
Zohn et al., 1995). Our observations that Dsh can be modi®ed
by PKCa in vitro are consistent with the possibility that PKC
may be involved in regulation of Dsh with respect to JNK
signaling. Thus, several results in different systems are
certainly consistent with the possibility that Wnt/Ca21 and
Wnt/JNK signaling pathways show overlapping properties.
3.4. A model of how Wnts in¯uence convergent extension
One contradiction to be answered is the observation that
the gain of function of Wnt-11/Wnt-5A (Torres et al., 1996)
as well as the prospective loss of function of Wnt-11/Wnt-
5A by expression of dnWnt-11 leads to an inhibition of
convergent extension movements in Xenopus embryos. As
both Wnt/b-catenin signaling (this study) and Wnt/JNK
signaling (Wallingford et al., 2000; Heisenberg et al.,
2000; Tada and Smith, 2000) are required for convergent
extension movements, this contradiction might be explained
by the hypothesis that Wnt/Ca21 signaling serves as a
balancer between these pathways (Fig. 7).
In the wild-type situation, all the different Wnt pathways
are active. The caveat is that the Wnt/Ca21 pathway and the
Wnt/JNK pathway may be the same, or overlapping. The
Wnt/b-catenin pathway is required to ensure expression of
Xnr-3, the Wnt/JNK pathway may be necessary to confer
proper cell polarity or behavior. In a situation characterized
by a Wnt-5A/Wnt-11 gain of function, the Wnt/Ca21 path-
way is strongly activated (Slusarski et al., 1997; Sheldahl et
al., 1999; KuÈhl et al., 2000a,b) and we speculate that PKC
phosphorylates Dsh, leading to preferential signaling
through JNK. In addition, activated CamKII inhibits the
function of XLEF-1. This leads to a loss of Wnt/b-catenin
signaling activity, resulting in an inhibition of convergent
extension movement (this paper). The Wnt/JNK pathway is
still active when Wnt-5A/Wnt-11 is active, thereby provid-
ing polarity information. Thus, elevating the Wnt/Ca21
signaling activity shifts the balance of Wnt/b-catenin and
Wnt/JNK signaling toward the Wnt/JNK side. On the other
hand, overexpressing the dnXwnt-11 construct inhibits acti-
vation of PKC and CamKII (KuÈhl et al., 2000a,b). Thus Dsh
might not be phosporylated by PKC and therefore might
preferentially signal through b-catenin, and in addition
CamKII is not able to inhibit the function of the LEF-1-
mediated transcription factor complex. In other words, over-
expressing the dnWnt-11 construct shifts the balance of the
Wnt/b-catenin and Wnt/JNK pathways toward the Wnt/b-
catenin pathway. The loss of JNK activity then leads to a
perturbation of gastrulation movements (Heisenberg et al.,
2000; Wallingford et al., 2000) and this cannot be rescued
by an overactivation of the Wnt/b-catenin pathway. In the
future it will be important to test whether Dsh is a substrate
for PKC in vivo as suggested by our in vitro data. If it is, this
will allow further testing of the model in which Dsh is a key
control point determining which response to Wnt signaling
will predominate. Indeed, the dishevelled-associated kinase
PAR-1 is an activator of the b-catenin pathway and an
inhibitor of JNK activation (Sun et al., 2001) whereas the
dishevelled interactor naked functions in the opposite direc-
tion (Yan et al., 2001). These ®ndings support the notion
that multiple Wnt signaling pathways are linked via Dishev-
elled in vertebrates.
4. Materials and methods
4.1. Microinjections of embryos
Mature eggs were obtained by injecting females with 500
units of human chorion gonadotropin (HCG, Sigma). One
hour after in vitro fertilization, the jelly coat was removed
by treating the embryos with 2% cysteine hydrochloride in 1/
10 MBS, pH 8.2 (1£ MBSH: 10 mM HEPES, 88 mM NaCl,
1 mM KCl, 0.33 mM Ca(NO3)2, 0.41 mM CaCl2, 0.82 mM
MgSO4, 2.4 mM NaHCO3, pH 7.4). Injection of embryos was
performed in 3% ®coll in 1/10 MBS. At the four-cell stage
both dorsal blastomeres were injected with 5 nl RNA solu-
tion. After injection the embryos were kept for a further 1±2 h
in 3% ®coll in 1/10 MBS to prevent leakage of the embryos
until they were transferred to 1/10 MBS.
4.2. In vitro transcription and RNA constructs
All constructs used in this study were either in pSPT64 or
pCS21 in order to generate in vitro transcribed RNA by use
of mMESSAGE mMACHINE Kit (Ambion). Amounts of
RNA injected were: Xwnt-8, 0.1 ng; XBDe348, 0.5 ng; XB/
U-cadherin, 1.0 ng; Xcadherin-11, 0.5 ng; b-catenin,
0.1 ng; LEF-1, 0.5 ng; XTCF-3, 0.5 ng; siamois, 20±
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±76 73

50 pg; Xnr-3, 0.5 ng; LEFDbBD, 0.5 ng; LEFDHMG,
0.5 ng; conductin, 0.5 ng; Xwnt-5A, 0.5 ng; CamKII
T286D, 1.0 ng; CamKII K42M, 1 ng; dnGSK-3b, 0.5 ng.
4.3. Keller explants
Explants of the dorsal upper blastopore lip were cut at
stage 10.5 (Nieuwkoop and Faber, 1975). Squares of
approximately 0.25 mm2 were cut from control and injected
embryos and cultivated at 188C for 20 h under a piece of
cover glass in modi®ed Barth solution (1 £ MBS). Petri
dishes and cover glasses were coated with BSA. Explants
were scored as described in Section 2. For PMA treatment,
explants were kept in MBS supplemented with 100 nM
PMA.
4.4. Western blot and ConA sepharose puri®cation
Ten embryos were lysed in 100 ml NOP buffer (PBS
supplemented with 1% Triton X-100, 1% NP40 and 2 mM
CaCl2) and cleared by centrifugation and extraction with
freon. The supernatant was treated with 40 ml ConA-sephar-
ose suspension (Sigma) for 90 min. After centrifugation the
supernatant contains soluble, cytoplasmic proteins, whereas
glyosylated membrane proteins and their interactors were
bound to the sepharose. After washing three times with
RIPA buffer (10 mM Tris, pH 7.5, 2 mM CaCl2, 0.15 M
NaCl, 1% Triton X-100, 1% Na-desoxycholat, 0.1%
SDS), once with high-salt buffer (0.5 M NaCl, 10 mM
Tris, pH 7.5, 2 mM CaCl2) and once with low-salt buffer
(10 mM Tris, pH 7.5, 2 mM CaCl2), proteins were eluted
with protein sample buffer, loaded onto a standard protein
SDS acrylamide gel and analyzed by standard Western blot
procedure. The antibody used to detect b-catenin was from
Transduction Laboratories.
4.5. Expression of fusion proteins and protein
phosphorylation
Cloning, expression, and puri®cation of bacterial fusion
proteins were as described in Pukrop et al. (2001). In short,
puri®cation was as follows: transformed BL21 (DE3)
bacteria were induced by 1 mM IPTG for 2 h at room
temperature. After centrifugation, bacterial pellets were
lysed in lysis buffer (50 mM NaH2PO4, 300±1000 mM
NaCl, 10 mM Imidazole, pH 8.0) including protease inhibi-
tors (complete, Roche) and the cleared supernatent was
loaded onto Ni-NTA resins for 1 h at 48C in a batch proce-
dure. After washing with 50 mM NaH2PO4, 300±1000 mM
NaCl, 10±30 mM Imidazole, pH 8.0, the protein was eluted
10 times with 1 ml of 50 mM NaH2PO4, 300±1000 mM
NaCl, 100±300 mM Imidazole, pH 8.0. Each fraction was
analyzed by SDS-PAGE followed by Coomassie staining
using standard procedures. Phosphorylation of XELF-1 by
CamKII was performed using commercially available
CamKII (NEB) at 308C for 5 min according to the manufac-
turer's instructions.
4.6. RT-PCR analyses
Total RNA was isolated from embryos using Purescript
RNA Isolation Kit (Biozym). First strand cDNAs were
synthesized according to the Gibco protocol using Super-
script II reverse transcriptase. The PCR reaction mixtures
for investigating the expression of marker genes were
prepared using PCR Supermix (Gibco) and PCR was
performed under standard conditions. The sequences of
the used oligonucleotide primers are the following:
Sia F, 5 0-CTCCAGCCACCAGTACCAGATC-3 0
Sia R, 5 0-GGGGAGAGTGGAAAGTGGTTG-3 0
Xnr-3 F, 5 0-CGAGTGCAAGAAGGTGGACA-3 0
Xnr-3 R, 5 0-ATCTTCATGGGGACACAGGA-3 0
Goosecoid F, 5 0-ACAACTGGAAGCACTGGA-3 0
Goosecoid R, 5 0-TCTTATTCCAGAGGAACC-3 0
Chordin F, 5 0-CTGTACCAACCCAATCCGTGCC-3 0
Chordin R, 5 0-CTTGGTGCAACATCTGTCCCGC-3 0
XFD/pintalavis F,
5 0-CCTTAGCCCTACATCTGAACAA-3 0
XFD/pintalavis R,
5 0-TAAAGGGAGCTGAGGATAGGTC-3 0
Xbra F, 5 0-GGATCGTTATCACCTCTG-3 0
Xbra R, 5 0-GTGTAGTCTGTAGCAGCA-3 0
H4 F, 5 0-CGGGATAACATTCAGGGTATCACT-3 0
H4 R, 5 0-ATCCATGGCGGTAACTGTCTTCCT-3 0
Ampli®ed products were analyzed on 1.2% agarose gels.
4.7. PKC phosporylation assay
Xenopus embryos were injected with either XPKCa-myc
or XDsh-HA RNA and lysates were prepared at blastula
stages. PKC and Dsh constructs were immunoprecipitated
separately with antibodies against the tags. Kinase reaction
of mixed immunoprecipitates was for 5 or 10 min as indi-
cated using 32P-g-ATP as cosubstrate and phosphatidyl-l-
serine as a cofactor. The PKC inhibitor used was
GF109203X.
Acknowledgements
We thank Nazan OÈ lmez for technical assistance. The LEF
mutants and the conductin construct were a kind gift of W.
Birchmeier, Berlin. R.T.M. is an Investigator of the Howard
Hughes Medical Institute. This work was supported by DFG
grants to D.W. and M.K.
References
Behrens, J., von Kreis, J.P., KuÈhl, M., Bruhn, L., Wedlich, D., Grosschedl,
R., Birchmeier, W., 1996. Functional interaction of b-catenin with the
transcription factor LEF-1. Nature 382, 638±642.
Behrens, J., Jerchow, B.A., WuÈrtele, M., Grimm, J., Asbrand, C., Wirtz, R.,
KuÈhl, M., Wedlich, D., Birchmeier, W., 1998. Functional interaction of
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±7674

an axin homolog, conductin, with b-catenin, APC, and GSK-3b.
Science 280, 596±599.
Boutros, M., Paricio, N., Strutt, D.I., Mlodzik, M., 1998. Dishevelled acti-
vates JNK and discriminates between JNK pathways in planar polarity
and wingless signalling. Cell 94, 109±118.
Bradley, R.S., Cowin, P., Brown, M.C., 1993. Expression of Wnt-1 in PC-
12 cells results in modulation of plakoglobin and E-cadherin and
increased cell adhesion. J. Cell Biol. 123, 1857±1865.
Brannon, M., Gomperts, M., Sumoy, L., Moon, R.T., Kimelman, D., 1997.
A b-catenin/Xtcf-3 complex binds to the Siamois promoter to regulate
speci®cation of the dorsal axis in Xenopus. Genes Dev. 11, 2359±2370.
Broders, F., Thiery, J.P., 1995. Contributions of cadherins to directional cell
migration and histogenesis in Xenopus embryos. Cell Adhes. Commun.
3, 419±440.
Cui, Y., Brown, J.D., Moon, R.T., Christian, J.L., 1995. Xwnt-8b: a mater-
nally expressed Xenopus Wnt gene with a potential role in establishing
the dorsoventral axis. Development 121, 2177±2186.
Deardorff, M.A., Tan, C.G., Conrad, L.J., Klein, P.S., 1998. Frizzled-8 is
expressed in the Spemann organizer and plays a role in early morpho-
genesis. Development 125, 2687±2700.
Djiane, A., Riou, J.F., Umbhauer, M., Bouocaut, J.C., Shi, D.L., 2000. Role
of frizzled7 in the regulation of convergent extension movements
during gastrulation in Xenopus laevis. EMBO J. 127, 3091±3100.
Du, S.J., Purcell, S.M., Christian, J.L., McGrew, L.L., Moon, R.T., 1995.
Identi®cation of distinct classes and functional domains of Wnts
through expression of wild-type and chimeric protein in Xenopus
embryos. Mol. Cell. Biol. 15, 2625±2634.
Enslen, H., Tokumitsu, H., Stork, P.J., Davis, R.J., Soderling, T.R., 1996.
Regulation of mitogen-activated protein kinases by a calcium/calmo-
dulin-dependent protein kinase cascade. Proc. Natl. Acad. Sci. USA 93,
10803±10808.
Gradl, D., KuÈhl, M., Wedlich, D., 1999. The Wnt/Wg signal transducer b-
catenin controls ®bronectin expression. Mol. Cell. Biol. 19, 5576±5587.
Hadeball, B., Borchers, A., Wedlich, D., 1998. Xenopus cadherin-11
(Xcadherin-11) expression requires the Wnt/Wg signal. Mech. Dev.
72, 101±113.
Heisenberg, C.P., Tada, M., Rauch, G.J., SauÂde, L., Concha, M.L., Geisler,
R., Stemple, D.L., Smith, J.C., Wilson, S.W., 2000. Silberblick/Wnt-11
mediates convergent extension movements during zebra®sh gastrula-
tion. Nature 40, 76±81.
Herman, M.A., 2001. C. elegans POP-1/TCF functions in a canonical Wnt
pathway that controls cell migration and in a noncanonical Wnt path-
way that controls cell polarity. Development 128, 581±590.
Howard, J.E., Smith, J.C., 1993. Analysis of gastrulation: different types of
gastrulation movement are induced by different mesoderm-inducing
factors in Xenopus laevis. Mech. Dev. 43, 37±48.
Keller, R.E., 1986. The cellular basis of amphibian gastrulation. In: Brow-
der, L. (Ed). The Cellular Basis of Morphogenesis, Developmental
Biology: A Comprehensive Synthesis. Plenum Press, New York, pp.
241±327.
KuÈhl, M., Wedlich, D., 1996. Xenopus cadherins: sorting out types and
functions in embryogenesis. Dev. Dyn. 207, 121±134.
KuÈhl, M., Wedlich, D., 1997. Wnt signalling goes nuclear. BioEssays 19,
101±104.
KuÈhl, M., Finnemann, S., Binder, O., Wedlich, D., 1996. Dominant nega-
tive expression of a cytoplasmic deleted mutant of XB/U-cadherin
disturbs mesoderm migration during gastrulation in Xenopus laevis.
Mech. Dev. 54, 71±82.
KuÈhl, M., Sheldahl, L.C., Malbon, C.C., Moon, R.T., 2000a. Ca21/calmo-
dulin-dependent protein kinase II is stimulated by Wnt and Frizzled
homologs and promotes ventral cell fates in Xenopus. J. Biol. Chem.
275, 12701±12711.
KuÈhl, M., Shedahl, L.C., Park, M., Miller, J.R., Moon, R.T., 2000b. The
Wnt/Ca21 pathway: a new vertebrate Wnt signaling pathway takes
shape. Trends Genet. 16, 279±283.
Laurent, M.N., Blitz, I.L., Hashimoto, C., RothbaÈcher, U., Cho, K.W.Y.,
1997. The Xenopus homeobox gene twin mediates Wnt induction of
goosecoid in establishment of Spemann's organizer. Development 124,
4905±4916.
Lee, C.H., Gumbiner, B.M., 1995. Disruption of gastrulation movements in
Xenopus by a dominant-negative mutant for C-cadherin. Dev. Biol.
171, 363±373.
Li, L., Yuan, H., Xie, W., Mao, J., Caruso, A.M., McMahon, A., Sussman,
D.J., Wu, D., 1999. Dishevelled proteins lead to two signalling path-
ways. Regulation of LEF-1 and c-jun N-terminal kinase in mammalian
cells. J. Biol. Chem. 274, 129±134.
McEwen, D.G., Cox, R.T., Peifer, M., 2000. The canonical Wg and JNK
signaling cascades collaborate to promote both dorsal closure and
ventral patterning. Development 127, 3607±3617.
McKendry, R., Hsu, S.C., Harland, R., Grosschedl, R., 1997. LEF-1/TCF
proteins mediate Wnt-inducible transcription from the Xenopus nodal-
related 3 promoter. Dev. Biol. 192, 420±431.
Medina, A., Reintsch, W., Steinbeisser, H., 2000. Xenopus frizzled 7 can
act in cannonical and non-cannonical Wnt signaling pathways: impli-
cations on early patterning and morphogenesis. Mech. Dev. 92, 227±
237.
Miller, J.R., Rowning, B.A., Larabell, C.A., Yang-Snyder, J.A., Bates,
R.L., Moon, R.T., 1999. Establishment of the dorsal±ventral axis in
Xenopus embryos coincides with the dorsal enrichment of dishevelled
that is dependent on cortical rotation. J. Cell Biol. 146, 427±437.
Molenaar, M., van de Wetering, M., Oosterwegel, M., Peterson-Maduro, J.,
Godsave, S., Korinek, V., Roose, J., Destree, O., Clevers, H., 1996.
XTCF-3 transcription factor mediates b-catenin induced axis formation
in Xenopus embryos. Cell 86, 391±399.
Molenaar, M., Roose, J., Peterson, J., Venanzi, S., Clevers, H., Destree, O.,
1998. Differential expression of the HMG-box transcription factors
XTCF-3 and XLEF-1 during early Xenopus development. Mech.
Dev. 75, 151±154.
MuÈller, H.A., KuÈhl, M., Finnemann, S., Schneider, S., van der Poel, S.Z.,
Hausen, P., Wedlich, D., 1994. Xenopus cadherins: the maternal pool
comprises distinguishable members of the family. Mech. Dev. 47, 213±
223.
Nieuwkoop, P.D., Faber, J., 1975. Normal table of Xenopus laevis
(Daudin), North Holland, Amsterdam.
Nishita, M., Hashimoto, M.K., Ogata, S., Laurent, M.N., Ueno, N.,
Shibuya, H., Cho, K.W.Y., 2000. Interaction between Wnt and TGF-
b signalling pathways during formation of Spemann's organizer.
Nature 403, 781±785.
Ponting, C.P., Bork, P., 1996. Pleckstrin's repeat performance: a novel
domain in G-protein signaling? Trends Biochem. Sci. 21, 245±246.
Pukrop, T., Gradl, D., Henningfeld, K.A., KnoÈchel, W., Wedlich, D., KuÈhl,
M., 2001. Identi®cation of two regulatory elements within the HMG-
box transcription factor XTCF-4. J. Biol. Chem. 276, 8968±8978.
Shulman, J.M., Perrimon, N., Axelrod, J.D., 1998. Frizzled signaling and
the developmental control of cell polarity. Trends Genet. 14, 452±
458.
Sheldahl, L.C., Park, M., Malbon, C.C., Moon, R.T., 1999. Protein kinase C
is differentially stimulated by Wnt and Frizzled homologs in a G-
protein-dependent manner. Curr. Biol. 9, 695±698.
Slusarski, D.C., Corces, V.G., Moon, R.T., 1997. Interaction of Wnt and
Frizzled homologue triggers G-protein-linked phosphatidylinositol
signalling. Nature 390, 410±413.
Smith, W.C., McKendry, R., Ribisi Jr., S., Harland, R.M., 1995. A nodal-
related gene de®nes a physical and functional domain within the
Spemann organizer. Cell 82, 37±46.
Sokol, S.Y., 1993. Mesoderm formation in Xenopus ectodermal explants
overexpressing Xwnt-8: evidence for a cooperating signal reaching the
animal pole by gastrulation. Development 118, 1335±1342.
Strutt, D.I., Weber, U., Mlodzik, M., 1997. The role of RhoA in tissue
polarity and Frizzled signalling. Nature 387, 292±295.
Sumanas, S., Strege, P., Heasman, J., Ekker, S.C., 2000. The putative Wnt
receptor Xenopus Frizzled ± functions upstream of b-catenin in verte-
brate dorsoventral patterning. Development 127, 1981±1990.
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±76 75

Sun, T.-Q., Lu, B., Feng, J.-J., Reinhard, C., Jan, Y.N., Fantl, W.J.,
Williams, L.T., 2001. PAR-1 is a Dishevelled associated kinase and a
positive regulator of Wnt signaling. Nature Cell Biol. (in press).
Tada, M., Smith, J.C., 2000. Xwnt-11 is a target of Xenopus Brachyury:
regulation of gastrulation movements via dishevelled, but not through
the canonical Wnt pathway. Development 127, 2227±2238.
Torres, M.A., Yang-Snyder, J., Purcell, S.M., Demarais, A.A., McGrew,
L.L., Moon, R.T., 1996. Activities of the wnt-1 class of secreted factors
are antagonized by the wnt-5 A class and by a dominant negative
cadherin in early Xenopus embryo. J. Cell Biol. 133, 1123±1137.
Wallingford, J.B., Rowning, B.A., Vogeli, K.M., RothbaÈcher, U., Fraser,
S.E., Harland, R.M., 2000. Dishevelled controls cell polarity during
Xenopus gastrulation. Nature 405, 81±85.
Winklbauer, R., 1994. Mesoderm migration in the vertebrate gastrula.
Semin. Dev. Biol. 5, 91±99.
Winklbauer, R., SchuÈrfeld, M., 1999. Vegetal rotation, a new gastrulation
movement involved in the internalization of the mesoderm and endo-
derm in Xenopus. Development 126, 3703±3713.
Wodarz, A., Nusse, R., 1998. Mechanisms of Wnt signaling in develop-
ment. Annu. Rev. Cell Dev. Biol. 14, 59±88.
Xu, Q., D'Amore, P.A., Sokol, S., 1998. Functional and biochemical inter-
actions of Wnts with FrzA, a secreted Wnt antagonist. Development
125, 4767±4776.
Yan, D., Wallingford, D., Sun, T.Q., Nelson, M.A., Sakanaka, C., Reinhard,
C., Harland, R.M., Fantl, W.J., Williams, L.T., 2001. Cell autonomous
regulation of multiple Dishevelled-dependent pathways by mammalian
naked. Proc. Natl Acad. Sci. USA 98, 3802±3807.
Zohn, I.E., Yu, H., Li, X., Cox, A.D., Earp, H.S., 1995. Angiotensin II
stimulates calcium-dependent activation of C-Jun N-terminal kinase.
Mol. Cell. Biol. 15, 6160±6168.
M. KuÈhl et al. / Mechanisms of Development 106 (2001) 61±7676
Related Documents











