Summary We investigated diurnal and seasonal changes in carbon acquisition and partitioning of recently assimilated car- bon in fast- and slow-growing families of loblolly pine (Pinus taeda L.) to determine whether fast-growing families exhibited greater carbon gain at the leaf level. Since planting on a xeric infertile site in Scotland County, NC, USA in 1993, five Atlan- tic Coastal Plain (ACP) and five “Lost Pines” Texas (TX) fam- ilies have been grown with either optimal nutrition or without fertilization (control). In 1998 and 1999, gas exchange param- eters were monitored bimonthly in four families and needles were analyzed bimonthly for starch and soluble sugar concen- trations. Although diurnal and seasonal effects on net photo- synthesis (A net ) and maximum rate of light-saturated photosyn- thesis (A max ) were significant, few family or treatment differ- ences in gas exchange characteristics were observed. The A net peaked at different times during the day over the season, and A max was generally highest in May. Instantaneous water-use ef- ficiency (WUE i ), derived from gas exchange parameters, did not differ among families, whereas foliage stable isotope com- position (δ 13 C) values suggested that TX families exhibited lower WUE than more mesic ACP families. Although there were no diurnal effects on foliar starch concentrations, needles exhibited pronounced seasonal changes in absolute concentra- tions of total nonstructural carbohydrates (TNC), starch and soluble sugars, and in partitioning of TNC to starch and sugars, mirroring seasonal changes in photosynthesis and shoot and root growth. In all families, foliar starch concentrations peaked in May and decreased to a minimum in winter, whereas reduc- ing sugar concentrations were highest in winter. Some family and treatment differences in partitioning of recently assimi- lated carbon in needles were observed, with the two TX fami- lies exhibiting higher concentrations of TNC and starch and enhanced starch partitioning compared with the ACP families. We conclude that growth differences among the four families are not a function of differences in carbon acquisition or parti- tioning at the leaf level. Keywords: δ 13 C, drought, gas exchange, nitrogen, water-use efficiency. Introduction Net photosynthesis (A net ) of a forest canopy is one of the prin- cipal processes determining net forest carbon gain, storage and productivity. Despite considerable efforts, the degree of genetic control over A net and the associated variation in growth or productivity are poorly understood in field-grown trees (Bongarten and Teskey 1987, Major and Johnsen 1996, Svensson 1996, Johnsen et al. 1999). Recent field studies un- dertaken to determine the physiological basis for observed ge- netic differences in growth of trees and genetic × environment interactions using photosynthetic gas exchange, water rela- tions and leaf carbon isotope discrimination have met with limited success (Major and Johnsen 1996, 1999, Svensson 1996, Johnsen and Major 1999). When photosynthesis has been related to growth, it is usually because estimates of leaf area, or other aspects of light interception (e.g., leaf orienta- tion and morphology), have been incorporated for scaling pur- poses (Bongarten and Teskey 1987, Michael et al. 1990, Wang et al. 1995, Major and Johnsen 1996). However, there is evi- dence of strong correlations between genetic variation in A net and growth under environmental stress, e.g., drought (Major and Johnsen 1996), in seedlings, and when seasonal (Boltz et al. 1986) and diurnal patterns in carbon acquisition and parti- tioning (at the leaf level) are closely examined. Because loblolly pine (Pinus taeda L.) has a wide geo- graphic distribution, it is common to find significant geno- typic variation in growth, particularly between ecotypes adapted to xeric and mesic soils (Bongarten and Teskey 1987). Loblolly pine seedling studies have associated yield variation with photosynthesis, water- and nutrient-use efficiency, and aspects of aboveground biomass partitioning (e.g., Bongarten and Teskey 1987, Seiler and Johnson 1988, Li et al. 1991, Greenwood and Volkaert 1992). However, similar field evi- dence associating genetic variation in productivity of larger trees with A net is lacking. The “Lost Pines” Texas provenance from Bastrop, Fayette and Caldwell Counties (van Buijtenen et al. 1976) is drought hardy and is generally described as slower growing than its more mesic Atlantic Coastal Plain counterparts. Seedling studies suggest that its drought hardi- Tree Physiology 22, 489–498 © 2002 Heron Publishing—Victoria, Canada Diurnal changes in gas exchange and carbon partitioning in needles of fast- and slow-growing families of loblolly pine (Pinus taeda) W. Q. YANG, 1–3 R. MURTHY, 4 P. KING 1 and M. A. TOPA 1 1 Boyce Thompson Institute for Plant Research, Tower Road, Ithaca, NY 14853-1801, USA 2 Present address: North Williamette Research and Extension Center, 15210 NE Miley Road, Aurora, OR 97002, USA 3 Author to whom correspondence should be addressed ([email protected]) 4 Lamont-Doherty Earth Observatory, Columbia University, Biosphere 2 Laboratory, 32540 S. Biosphere Road, Oracle, AZ 85623, USA Received January 30, 2001; accepted August 17, 2001; published online April 1, 2002 Downloaded from https://academic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Summary We investigated diurnal and seasonal changes incarbon acquisition and partitioning of recently assimilated car-bon in fast- and slow-growing families of loblolly pine (Pinustaeda L.) to determine whether fast-growing families exhibitedgreater carbon gain at the leaf level. Since planting on a xericinfertile site in Scotland County, NC, USA in 1993, five Atlan-tic Coastal Plain (ACP) and five “Lost Pines” Texas (TX) fam-ilies have been grown with either optimal nutrition or withoutfertilization (control). In 1998 and 1999, gas exchange param-eters were monitored bimonthly in four families and needleswere analyzed bimonthly for starch and soluble sugar concen-trations. Although diurnal and seasonal effects on net photo-synthesis (Anet) and maximum rate of light-saturated photosyn-thesis (Amax) were significant, few family or treatment differ-ences in gas exchange characteristics were observed. The Anet
peaked at different times during the day over the season, andAmax was generally highest in May. Instantaneous water-use ef-ficiency (WUEi), derived from gas exchange parameters, didnot differ among families, whereas foliage stable isotope com-position (δ13C) values suggested that TX families exhibitedlower WUE than more mesic ACP families. Although therewere no diurnal effects on foliar starch concentrations, needlesexhibited pronounced seasonal changes in absolute concentra-tions of total nonstructural carbohydrates (TNC), starch andsoluble sugars, and in partitioning of TNC to starch and sugars,mirroring seasonal changes in photosynthesis and shoot androot growth. In all families, foliar starch concentrations peakedin May and decreased to a minimum in winter, whereas reduc-ing sugar concentrations were highest in winter. Some familyand treatment differences in partitioning of recently assimi-lated carbon in needles were observed, with the two TX fami-lies exhibiting higher concentrations of TNC and starch andenhanced starch partitioning compared with the ACP families.We conclude that growth differences among the four familiesare not a function of differences in carbon acquisition or parti-tioning at the leaf level.
Keywords: δ13C, drought, gas exchange, nitrogen, water-useefficiency.
Introduction
Net photosynthesis (Anet) of a forest canopy is one of the prin-cipal processes determining net forest carbon gain, storageand productivity. Despite considerable efforts, the degree ofgenetic control over Anet and the associated variation in growthor productivity are poorly understood in field-grown trees(Bongarten and Teskey 1987, Major and Johnsen 1996,Svensson 1996, Johnsen et al. 1999). Recent field studies un-dertaken to determine the physiological basis for observed ge-netic differences in growth of trees and genetic × environmentinteractions using photosynthetic gas exchange, water rela-tions and leaf carbon isotope discrimination have met withlimited success (Major and Johnsen 1996, 1999, Svensson1996, Johnsen and Major 1999). When photosynthesis hasbeen related to growth, it is usually because estimates of leafarea, or other aspects of light interception (e.g., leaf orienta-tion and morphology), have been incorporated for scaling pur-poses (Bongarten and Teskey 1987, Michael et al. 1990, Wanget al. 1995, Major and Johnsen 1996). However, there is evi-dence of strong correlations between genetic variation in Anet
and growth under environmental stress, e.g., drought (Majorand Johnsen 1996), in seedlings, and when seasonal (Boltz etal. 1986) and diurnal patterns in carbon acquisition and parti-tioning (at the leaf level) are closely examined.
Because loblolly pine (Pinus taeda L.) has a wide geo-graphic distribution, it is common to find significant geno-typic variation in growth, particularly between ecotypesadapted to xeric and mesic soils (Bongarten and Teskey 1987).Loblolly pine seedling studies have associated yield variationwith photosynthesis, water- and nutrient-use efficiency, andaspects of aboveground biomass partitioning (e.g., Bongartenand Teskey 1987, Seiler and Johnson 1988, Li et al. 1991,Greenwood and Volkaert 1992). However, similar field evi-dence associating genetic variation in productivity of largertrees with Anet is lacking. The “Lost Pines” Texas provenancefrom Bastrop, Fayette and Caldwell Counties (van Buijtenenet al. 1976) is drought hardy and is generally described asslower growing than its more mesic Atlantic Coastal Plaincounterparts. Seedling studies suggest that its drought hardi-
Tree Physiology 22, 489–498© 2002 Heron Publishing—Victoria, Canada
Diurnal changes in gas exchange and carbon partitioning in needles offast- and slow-growing families of loblolly pine (Pinus taeda)
W. Q. YANG,1–3 R. MURTHY,4 P. KING1 and M. A. TOPA1
1 Boyce Thompson Institute for Plant Research, Tower Road, Ithaca, NY 14853-1801, USA2 Present address: North Williamette Research and Extension Center, 15210 NE Miley Road, Aurora, OR 97002, USA3 Author to whom correspondence should be addressed ([email protected])4 Lamont-Doherty Earth Observatory, Columbia University, Biosphere 2 Laboratory, 32540 S. Biosphere Road, Oracle, AZ 85623, USA
Received January 30, 2001; accepted August 17, 2001; published online April 1, 2002
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022

ness is more a function of drought avoidance mechanisms thandrought tolerance mechanisms because (1) rapid transpirationoccurs when water is available, but water is conserved duringstress, (2) root systems are deep with wide-ranging lateralsand (3) needles are small with deep stomatal pits and fewstomata per unit leaf surface (van Buijtenen et al. 1976, Bilanet al. 1977). It is not known whether the slow growth of field-grown trees is associated with these drought avoidance mech-anisms, in particular, with gas exchange characteristics.
We investigated genetic, seasonal, diurnal and fertilizer ef-fects on carbon acquisition and partitioning of recently assimi-lated carbon in fast- and slow-growing families of loblollypine from a 5-year-old plantation in Scotland County, NC,USA. Specifically, we determined whether (1) fast-growingfamilies exhibited greater carbon gain at the leaf level thanslow-growing families, (2) fast- and slow-growing familiespartitioned recently assimilated carbon differently in needlesand (3) Texas families exhibited higher water-use efficienciesthan Atlantic Coastal Plain families. Gas exchange character-istics were monitored during both a wet and a dry year.
Materials and methods
Study site
The 11-ha study site, neighboring the Southeast Tree Researchand Education Site (SETRES, see Albaugh et al. 1998), is lo-cated in the Georgia–Carolina Sandhills in Scotland County,NC, USA. The soil is sandy, excessively drained, infertile andacidic (pH = 4.6). Mean annual precipitation is 1200 mm andyearly temperature is 17 °C (26 °C in summer and 9 °C in win-ter). Precipitation differed significantly during 1998 and 1999.Annual precipitation for 1998 and 1999 through Septemberwas 1115 and 772 mm, respectively.
From November 1993 to March 1994, loblolly pine seed-lings were planted in 10 replicated blocks with a split-plot de-sign. Nutritional treatment was the main plot (optimal fertil-ization and unfertilized or control) and provenance was thesub-plot (North and South Carolina Atlantic Coastal Plain(ACP) provenance, and a drought-hardy Texas (TX) prove-nance from the “Lost-Pines” area, McKeand et al. 1999). Eachprovenance consisted of five open-pollinated loblolly pinefamilies, with 10 rows of 10 trees planted in each family plot at1.5 × 2 m spacing. Optimal fertilization has been maintainedyearly on one-half of each block by determining annual foliarnutrient ratios (Hockman and Allen 1990). In the presentstudy, only the fastest- (ACP-1 and TX-1) and slowest-grow-ing (ACP-2 and TX-2) families from each provenance wereselected based on evaluation of aboveground height growth of4-year-old control trees (McKeand et al. 1999). We also fo-cused our analysis on three blocks (Retzlaff et al. 2001). Dur-ing each sampling period, the orders of sample collection fromthe three blocks, treatment within each block, and row withineach family plot were randomly selected. The same row wassampled in all three blocks.
Gas exchange and water-use efficiency
During the 1998 and 1999 growing seasons, diurnal changesin gas exchange parameters were monitored bimonthly (LI-6400, Li-Cor, Lincoln, NE); however, in May and Septemberof 1999, only light-saturated maximum photosynthesis (Amax)was measured. Gas exchange data for each block were col-lected on three consecutive days from needles on south-facingbranches exposed to full sunlight in the upper third of the treecanopy. With the exception of the May 1999 sampling, allneedles were at least 80% of their final length. Because of thelarge sample size and inaccessibility of needles, all gas ex-change measurements were made on detached needles within4 min of needle collection. Once removed, needle fascicleswere immediately placed upright in a glass vial containing dis-tilled water, and gas-exchange rates were determined. Gas-ex-change rates were unaffected by using detached needleswithin 4 min of collection. Preliminary tests at various timesthroughout the day indicated that there were no changes in wa-ter relations within the 4-min measuring period with the reso-lution of our equipment. Other studies have found a similarinsensitivity of loblolly pine to short-term changes in VPD,and have used excised needles to measure gas exchange rates(e.g., Samuelson et al. 1992, Porte and Loustau 1998, Samuel-son 2000). All gas exchange measurements are expressed on atotal surface area basis. Needle width was measured with anoptical micrometer and needle area was calculated from lengthand radius of all needles in the cuvette (Murthy and Dougherty1997). Needles from the current flush (usually Flush 1 in thecurrent year) were removed, and needles from the previousyear (Flush 1 or the healthiest flush if multiple flushes existed)were also removed if present.
During measurements, the reference CO2 concentration(400 ppm for diurnal and 385 ppm for Amax) was maintained inthe cuvette with an external CO2 source; photosyntheticallyactive radiation (PAR) and relative humidity were adjusted toapproximate mean light and humidity values for that intervalperiod. For diurnal measurements, two trees from each familyplot were selected and four to six measurement intervals wereconducted depending on day length. Light-saturated maxi-mum photosynthesis was determined on four to six trees fromeach family plot using 1800 µmol m–2 s–1 of PAR between0900 and 1300 h in May and September of 1999. Light-satu-rated maximum photosynthesis data for months other thanMay and September 1999 were the composite of diurnal Anet
data from all intervals before 1300 h exhibiting maximumdaily PAR flux densities. Instantaneous water-use efficiency(WUEi) was calculated as Anet divided by stomatal conduc-tance, gs.
Long-term WUE was assessed by stable carbon isotopecomposition (δ13C) measurements. Needles were removedfrom an upper-canopy branch adjacent to the branch used forgas exchange measurements. Needles were placed immedi-ately on dry ice and stored at –70 °C until freeze-dried. Tissuewas ground with a SPEX® CertiPrep 6750 freezer mill. Foliartissue δ13C and %N in September 1998 and 1999 was obtainedwith a Europa Scientific ANCA-SL Stable Isotope Analysis
490 YANG, MURTHY, KING AND TOPA
TREE PHYSIOLOGY VOLUME 22, 2002
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022
System (Europa Scientific, Crewe, U.K.) at the Cornell BoyceThompson Institute Stable Isotope Laboratory. The stable car-bon isotope ratio was expressed as the 13C/12C ratio in foliagerelative to the Pee Dee Belemnite standard (δ13C, ‰). Preci-sion of the spectrometric analysis of a laboratory standard was± 0.2‰.
Diurnal carbohydrate analysis
Current- and previous-year needles were removed for carbo-hydrate analysis from the same trees used for gas exchangemeasurements. Needles were selected from an upper-canopybranch of two trees per family plot. Four fascicles were re-moved over a 24-h period at dawn, dusk and the followingdawn. Preliminary experiments indicated that between-branchvariability in total needle nonstructural carbohydrates and par-titioning was less than 10%, and that maximum starch concen-tration in needles generally occurred by 1300 h, remainingconstant until sunset (data not shown). Foliage tissue wasplaced on dry ice immediately after removal, stored at –70 °Cuntil freeze-dried, ground and extracted with 80% ethanol at80 °C (Topa and Cheeseman 1992). The supernatant was ana-lyzed enzymatically for reducing sugars (glucose + fructose)and sucrose (Jones et al. 1977). The same enzymatic analysiswas used to determine tissue starch concentration after incu-bating the tissue pellet with amyloglucosidase for 24 h at55 °C. Total nonstructural carbohydrate (TNC) concentrationsare the sum of reducing sugars (glucose and fructose), sucroseand starch concentrations. Starch, sugar and TNC concentra-tions are reported as glucose equivalents per gram dry mass oftissue.
Data analysis
Data were analyzed by a repeated measures analysis of vari-ance with a split-plot design: fertilizer treatment was the mainplot, provenance was the subplot, and families were nestedwithin provenance. Between- and within-subject effects weretested at the 0.05 probability level. Trees in each family/blockwere treated as subsamples for both photosynthesis and carbo-hydrate data analyses. Fisher’s protected least significant dif-ference was used in mean comparisons when treatment andprovenance interactions were not significant; or least squaremeans were presented with Bonferroni adjustment where ap-propriate. Linear regression techniques were used to deter-mine relationships between Amax and gs, and between WUEand 13C isotope discrimination. All analyses were performedwith GLM SAS procedures (SAS Institute, Gary, NC) and dif-ferences were considered significant at P ≤ 0.05.
Results
Diurnal Anet
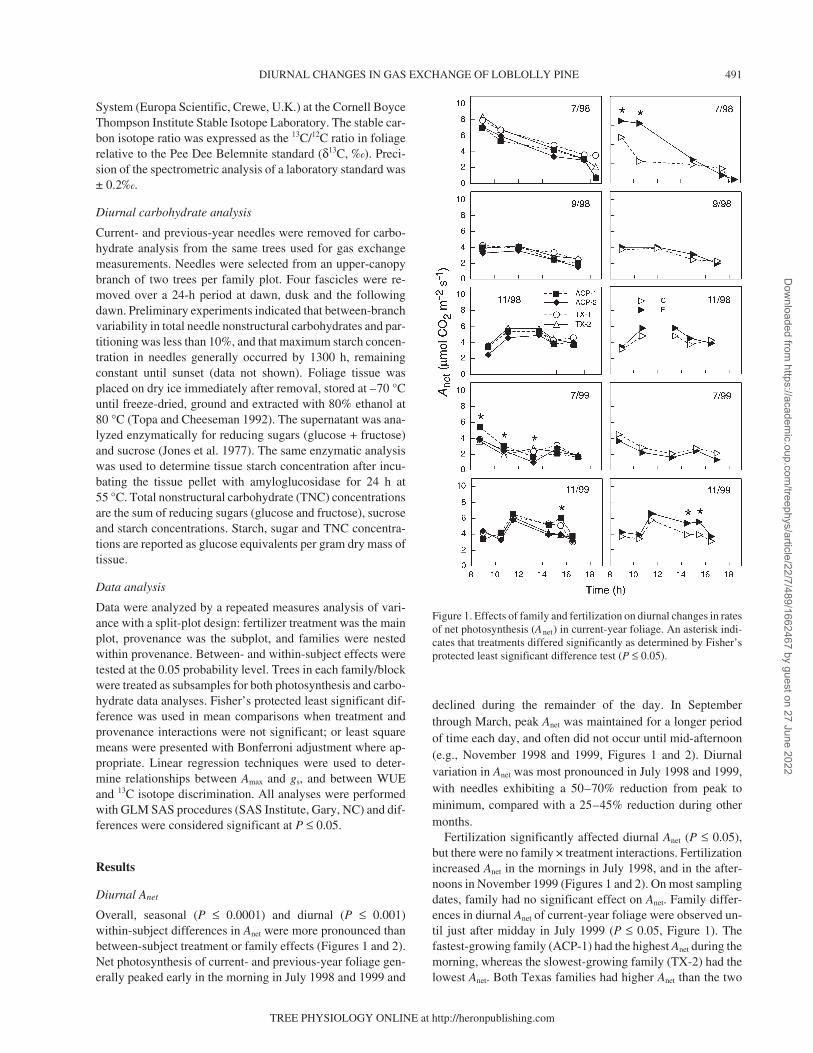
Overall, seasonal (P ≤ 0.0001) and diurnal (P ≤ 0.001)within-subject differences in Anet were more pronounced thanbetween-subject treatment or family effects (Figures 1 and 2).Net photosynthesis of current- and previous-year foliage gen-erally peaked early in the morning in July 1998 and 1999 and
declined during the remainder of the day. In Septemberthrough March, peak Anet was maintained for a longer periodof time each day, and often did not occur until mid-afternoon(e.g., November 1998 and 1999, Figures 1 and 2). Diurnalvariation in Anet was most pronounced in July 1998 and 1999,with needles exhibiting a 50–70% reduction from peak tominimum, compared with a 25–45% reduction during othermonths.
Fertilization significantly affected diurnal Anet (P ≤ 0.05),but there were no family × treatment interactions. Fertilizationincreased Anet in the mornings in July 1998, and in the after-noons in November 1999 (Figures 1 and 2). On most samplingdates, family had no significant effect on Anet. Family differ-ences in diurnal Anet of current-year foliage were observed un-til just after midday in July 1999 (P ≤ 0.05, Figure 1). Thefastest-growing family (ACP-1) had the highest Anet during themorning, whereas the slowest-growing family (TX-2) had thelowest Anet. Both Texas families had higher Anet than the two
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
DIURNAL CHANGES IN GAS EXCHANGE OF LOBLOLLY PINE 491
Figure 1. Effects of family and fertilization on diurnal changes in ratesof net photosynthesis (Anet) in current-year foliage. An asterisk indi-cates that treatments differed significantly as determined by Fisher’sprotected least significant difference test (P ≤ 0.05).
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022
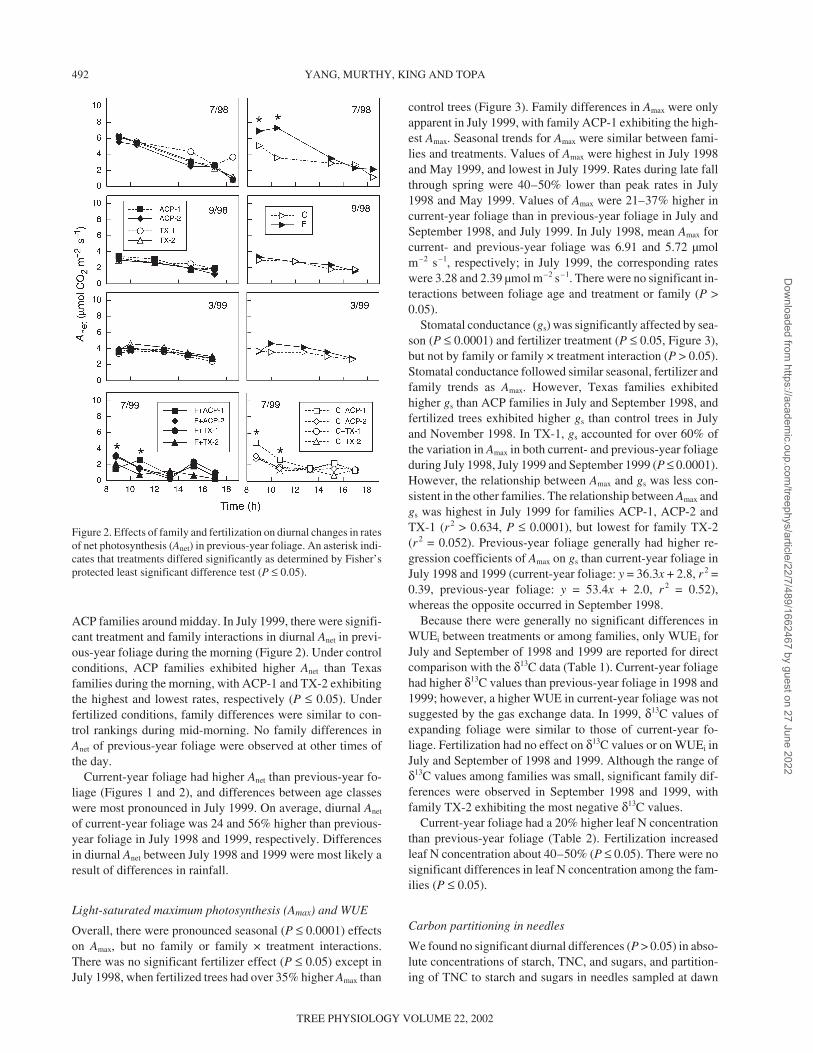
ACP families around midday. In July 1999, there were signifi-cant treatment and family interactions in diurnal Anet in previ-ous-year foliage during the morning (Figure 2). Under controlconditions, ACP families exhibited higher Anet than Texasfamilies during the morning, with ACP-1 and TX-2 exhibitingthe highest and lowest rates, respectively (P ≤ 0.05). Underfertilized conditions, family differences were similar to con-trol rankings during mid-morning. No family differences inAnet of previous-year foliage were observed at other times ofthe day.
Current-year foliage had higher Anet than previous-year fo-liage (Figures 1 and 2), and differences between age classeswere most pronounced in July 1999. On average, diurnal Anet
of current-year foliage was 24 and 56% higher than previous-year foliage in July 1998 and 1999, respectively. Differencesin diurnal Anet between July 1998 and 1999 were most likely aresult of differences in rainfall.
Light-saturated maximum photosynthesis (Amax) and WUE
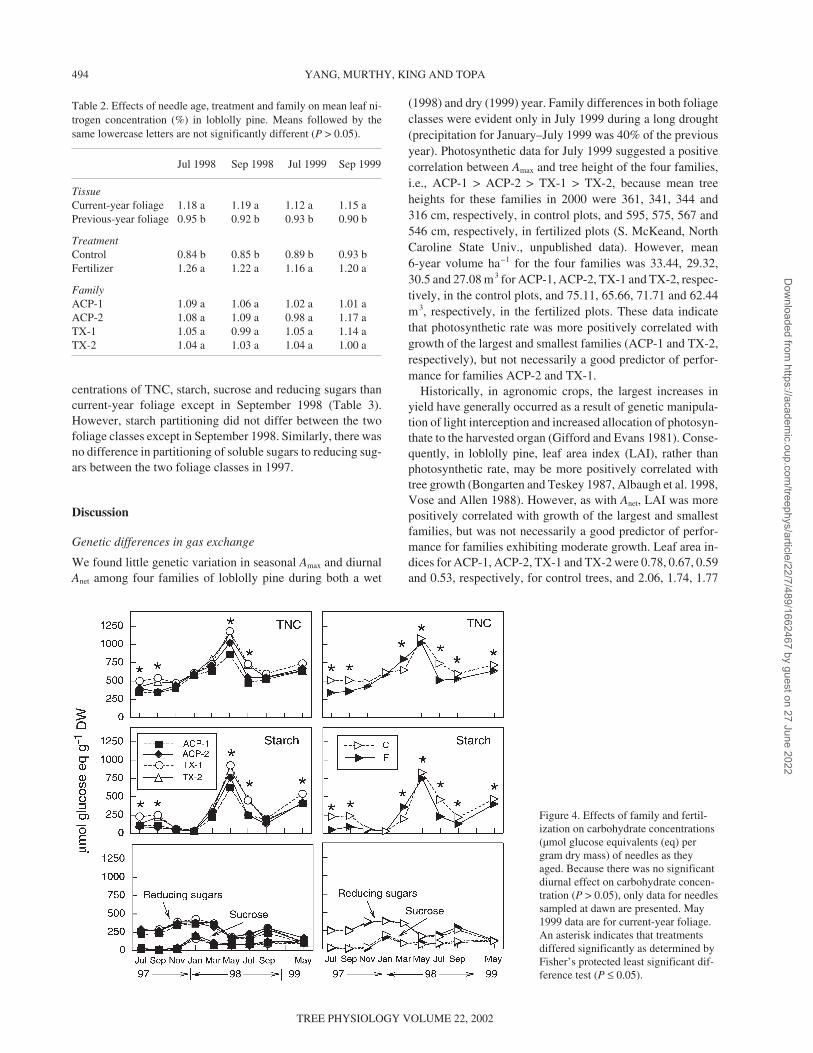
Overall, there were pronounced seasonal (P ≤ 0.0001) effectson Amax, but no family or family × treatment interactions.There was no significant fertilizer effect (P ≤ 0.05) except inJuly 1998, when fertilized trees had over 35% higher Amax than
control trees (Figure 3). Family differences in Amax were onlyapparent in July 1999, with family ACP-1 exhibiting the high-est Amax. Seasonal trends for Amax were similar between fami-lies and treatments. Values of Amax were highest in July 1998and May 1999, and lowest in July 1999. Rates during late fallthrough spring were 40–50% lower than peak rates in July1998 and May 1999. Values of Amax were 21–37% higher incurrent-year foliage than in previous-year foliage in July andSeptember 1998, and July 1999. In July 1998, mean Amax forcurrent- and previous-year foliage was 6.91 and 5.72 µmolm–2 s–1, respectively; in July 1999, the corresponding rateswere 3.28 and 2.39 µmol m–2 s–1. There were no significant in-teractions between foliage age and treatment or family (P >0.05).
Stomatal conductance (gs) was significantly affected by sea-son (P ≤ 0.0001) and fertilizer treatment (P ≤ 0.05, Figure 3),but not by family or family × treatment interaction (P > 0.05).Stomatal conductance followed similar seasonal, fertilizer andfamily trends as Amax. However, Texas families exhibitedhigher gs than ACP families in July and September 1998, andfertilized trees exhibited higher gs than control trees in Julyand November 1998. In TX-1, gs accounted for over 60% ofthe variation in Amax in both current- and previous-year foliageduring July 1998, July 1999 and September 1999 (P ≤ 0.0001).However, the relationship between Amax and gs was less con-sistent in the other families. The relationship between Amax andgs was highest in July 1999 for families ACP-1, ACP-2 andTX-1 (r 2 > 0.634, P ≤ 0.0001), but lowest for family TX-2(r 2 = 0.052). Previous-year foliage generally had higher re-gression coefficients of Amax on gs than current-year foliage inJuly 1998 and 1999 (current-year foliage: y = 36.3x + 2.8, r 2 =0.39, previous-year foliage: y = 53.4x + 2.0, r 2 = 0.52),whereas the opposite occurred in September 1998.
Because there were generally no significant differences inWUEi between treatments or among families, only WUEi forJuly and September of 1998 and 1999 are reported for directcomparison with the δ13C data (Table 1). Current-year foliagehad higher δ13C values than previous-year foliage in 1998 and1999; however, a higher WUE in current-year foliage was notsuggested by the gas exchange data. In 1999, δ13C values ofexpanding foliage were similar to those of current-year fo-liage. Fertilization had no effect on δ13C values or on WUEi inJuly and September of 1998 and 1999. Although the range ofδ13C values among families was small, significant family dif-ferences were observed in September 1998 and 1999, withfamily TX-2 exhibiting the most negative δ13C values.
Current-year foliage had a 20% higher leaf N concentrationthan previous-year foliage (Table 2). Fertilization increasedleaf N concentration about 40–50% (P ≤ 0.05). There were nosignificant differences in leaf N concentration among the fam-ilies (P ≤ 0.05).
Carbon partitioning in needles
We found no significant diurnal differences (P > 0.05) in abso-lute concentrations of starch, TNC, and sugars, and partition-ing of TNC to starch and sugars in needles sampled at dawn
492 YANG, MURTHY, KING AND TOPA
TREE PHYSIOLOGY VOLUME 22, 2002
Figure 2. Effects of family and fertilization on diurnal changes in ratesof net photosynthesis (Anet) in previous-year foliage. An asterisk indi-cates that treatments differed significantly as determined by Fisher’sprotected least significant difference test (P ≤ 0.05).
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022

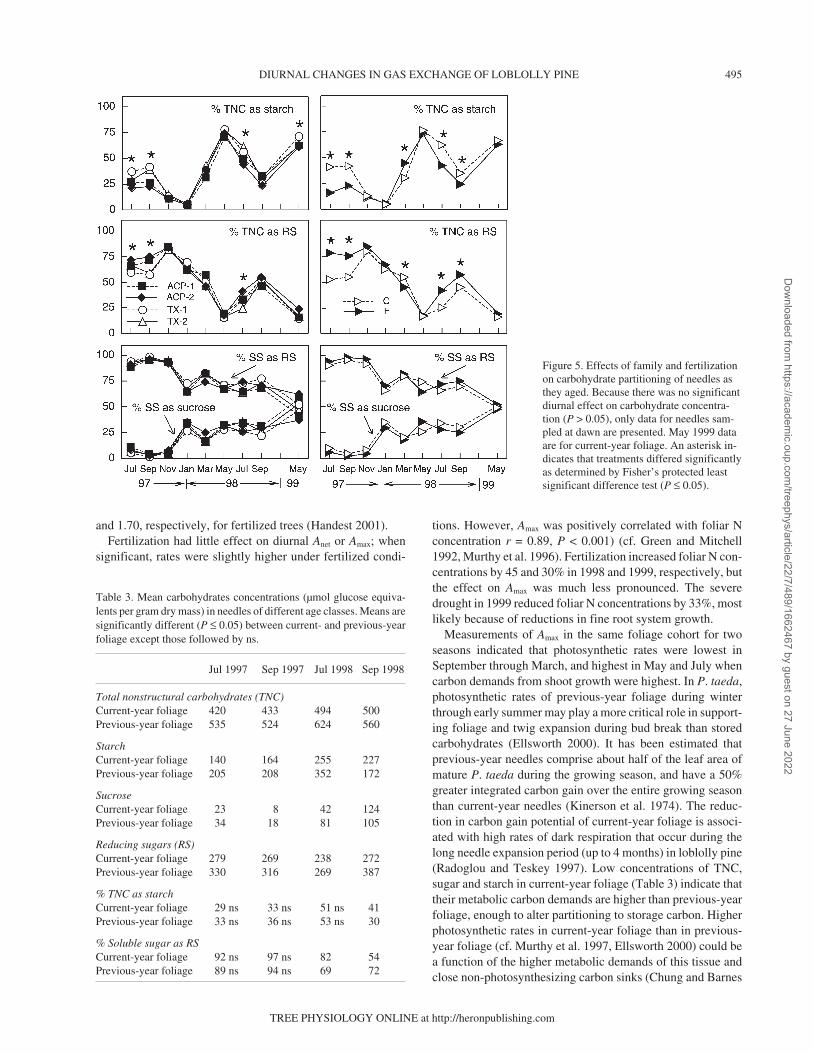
and dusk on all sampling dates. Consequently, only data forthe dawn sampling period are shown. Seasonal differences inneedle concentrations of TNC, starch, reducing sugars and su-crose were highly significant (P ≤ 0.0001, Figure 4), as werethe seasonal differences in partitioning of TNC to starch andsoluble sugars (P ≤ 0.001, Figure 5). Family effects were sig-nificant for all TNC variables (P ≤ 0.029), whereas treatmenteffects were significant for all variables except reducing sugarand sucrose concentrations (P ≤ 0.05).
The highest concentrations of TNC, starch and percent ofTNC partitioned to starch (i.e., starch partitioning) were ob-served in May, whereas reducing sugar concentrations werehighest in winter (November–March). The percent of TNCpartitioned to reducing sugars peaked in late fall, with a mini-mum in May (Figure 5). Reducing sugars were the dominantsoluble sugars in needles, representing over 70% of solublesugars. Consequently, seasonal patterns for soluble sugar con-
centrations mirrored those for reducing sugar (data notshown).
Fertilization generally decreased absolute concentrations ofTNC and starch, and starch partitioning except during the win-ter (November–March), and increased the partitioning of TNCto reducing sugars from summer to early fall. The largest re-ductions in starch concentrations (40–75%) and starch parti-tioning (30–60%) occurred in July and September of bothyears. Family differences in starch and TNC concentrationsand starch partitioning were significant on half of the samplingdates. In general, the two Texas families exhibited higherstarch and TNC concentrations, and starch partitioning thanthe ACP families in May, July and September, which was ac-companied by a decrease in partitioning to reducing sugars. InJuly and September, starch concentrations of the two Texasfamilies were 2–3 times those of the ACP families.
Previous-year foliage had significantly higher needle con-
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
DIURNAL CHANGES IN GAS EXCHANGE OF LOBLOLLY PINE 493
Figure 3. Effects of family and fertil-ization on maximum rates of photo-synthesis (Amax) and stomatal conduc-tance (gs) of needles as they aged. Anasterisk indicates that treatments dif-fered significantly as determined byFisher’s protected least significant dif-ference test (P ≤ 0.05).
Table 1. Effects of foliage age, treatment and family on mean foliage δ13C values and instantaneous water use efficiency (WUEi). Means followedby the same lowercase letters are not significantly different (P > 0.05).
δ13C (‰) WUEi (µmol CO2 mol–1)
Sep 1998 Sep 1999 Jul 1998 Sep 1998 Jul 1999 Sep 1999
TissueCurrent-year foliage –28.36 a –27.93 a 76.9 a 59.4 a 131.3 a 79.3Previous-year foliage –28.97 b –28.72 b 77.6 a 54.4 a 123.4 a –
TreatmentControl –28.87 a –28.25 a 68.6 a 52.2 a 130.4 a 83.9 aFertilizer –28.53 a –28.14 a 85.2 a 55.2 a 124.4 a 75.5 a
FamilyACP-1 –28.59 a –27.90 a 84.5 a 61.7 a 132.6 a 86.7 aACP-2 –28.55 a –27.85 a 84.7 a 52.0 a 128.7 a 82.6 aTX-1 –28.61 a –28.28 ab 70.5 b 54.5 a 110.7 a 77.9 aTX-2 –29.04 b –28.59 b 69.1 b 45.1 a 137.5 a 71.7 a
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022
centrations of TNC, starch, sucrose and reducing sugars thancurrent-year foliage except in September 1998 (Table 3).However, starch partitioning did not differ between the twofoliage classes except in September 1998. Similarly, there wasno difference in partitioning of soluble sugars to reducing sug-ars between the two foliage classes in 1997.
Discussion
Genetic differences in gas exchange
We found little genetic variation in seasonal Amax and diurnalAnet among four families of loblolly pine during both a wet
(1998) and dry (1999) year. Family differences in both foliageclasses were evident only in July 1999 during a long drought(precipitation for January–July 1999 was 40% of the previousyear). Photosynthetic data for July 1999 suggested a positivecorrelation between Amax and tree height of the four families,i.e., ACP-1 > ACP-2 > TX-1 > TX-2, because mean treeheights for these families in 2000 were 361, 341, 344 and316 cm, respectively, in control plots, and 595, 575, 567 and546 cm, respectively, in fertilized plots (S. McKeand, NorthCaroline State Univ., unpublished data). However, mean6-year volume ha–1 for the four families was 33.44, 29.32,30.5 and 27.08 m3 for ACP-1, ACP-2, TX-1 and TX-2, respec-tively, in the control plots, and 75.11, 65.66, 71.71 and 62.44m3, respectively, in the fertilized plots. These data indicatethat photosynthetic rate was more positively correlated withgrowth of the largest and smallest families (ACP-1 and TX-2,respectively), but not necessarily a good predictor of perfor-mance for families ACP-2 and TX-1.
Historically, in agronomic crops, the largest increases inyield have generally occurred as a result of genetic manipula-tion of light interception and increased allocation of photosyn-thate to the harvested organ (Gifford and Evans 1981). Conse-quently, in loblolly pine, leaf area index (LAI), rather thanphotosynthetic rate, may be more positively correlated withtree growth (Bongarten and Teskey 1987, Albaugh et al. 1998,Vose and Allen 1988). However, as with Anet, LAI was morepositively correlated with growth of the largest and smallestfamilies, but was not necessarily a good predictor of perfor-mance for families exhibiting moderate growth. Leaf area in-dices for ACP-1, ACP-2, TX-1 and TX-2 were 0.78, 0.67, 0.59and 0.53, respectively, for control trees, and 2.06, 1.74, 1.77
494 YANG, MURTHY, KING AND TOPA
TREE PHYSIOLOGY VOLUME 22, 2002
Table 2. Effects of needle age, treatment and family on mean leaf ni-trogen concentration (%) in loblolly pine. Means followed by thesame lowercase letters are not significantly different (P > 0.05).
Jul 1998 Sep 1998 Jul 1999 Sep 1999
TissueCurrent-year foliage 1.18 a 1.19 a 1.12 a 1.15 aPrevious-year foliage 0.95 b 0.92 b 0.93 b 0.90 b
TreatmentControl 0.84 b 0.85 b 0.89 b 0.93 bFertilizer 1.26 a 1.22 a 1.16 a 1.20 a
FamilyACP-1 1.09 a 1.06 a 1.02 a 1.01 aACP-2 1.08 a 1.09 a 0.98 a 1.17 aTX-1 1.05 a 0.99 a 1.05 a 1.14 aTX-2 1.04 a 1.03 a 1.04 a 1.00 a
Figure 4. Effects of family and fertil-ization on carbohydrate concentrations(µmol glucose equivalents (eq) pergram dry mass) of needles as theyaged. Because there was no significantdiurnal effect on carbohydrate concen-tration (P > 0.05), only data for needlessampled at dawn are presented. May1999 data are for current-year foliage.An asterisk indicates that treatmentsdiffered significantly as determined byFisher’s protected least significant dif-ference test (P ≤ 0.05).
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022
and 1.70, respectively, for fertilized trees (Handest 2001).Fertilization had little effect on diurnal Anet or Amax; when
significant, rates were slightly higher under fertilized condi-
tions. However, Amax was positively correlated with foliar Nconcentration r = 0.89, P < 0.001) (cf. Green and Mitchell1992, Murthy et al. 1996). Fertilization increased foliar N con-centrations by 45 and 30% in 1998 and 1999, respectively, butthe effect on Amax was much less pronounced. The severedrought in 1999 reduced foliar N concentrations by 33%, mostlikely because of reductions in fine root system growth.
Measurements of Amax in the same foliage cohort for twoseasons indicated that photosynthetic rates were lowest inSeptember through March, and highest in May and July whencarbon demands from shoot growth were highest. In P. taeda,photosynthetic rates of previous-year foliage during winterthrough early summer may play a more critical role in support-ing foliage and twig expansion during bud break than storedcarbohydrates (Ellsworth 2000). It has been estimated thatprevious-year needles comprise about half of the leaf area ofmature P. taeda during the growing season, and have a 50%greater integrated carbon gain over the entire growing seasonthan current-year needles (Kinerson et al. 1974). The reduc-tion in carbon gain potential of current-year foliage is associ-ated with high rates of dark respiration that occur during thelong needle expansion period (up to 4 months) in loblolly pine(Radoglou and Teskey 1997). Low concentrations of TNC,sugar and starch in current-year foliage (Table 3) indicate thattheir metabolic carbon demands are higher than previous-yearfoliage, enough to alter partitioning to storage carbon. Higherphotosynthetic rates in current-year foliage than in previous-year foliage (cf. Murthy et al. 1997, Ellsworth 2000) could bea function of the higher metabolic demands of this tissue andclose non-photosynthesizing carbon sinks (Chung and Barnes
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
DIURNAL CHANGES IN GAS EXCHANGE OF LOBLOLLY PINE 495
Figure 5. Effects of family and fertilizationon carbohydrate partitioning of needles asthey aged. Because there was no significantdiurnal effect on carbohydrate concentra-tion (P > 0.05), only data for needles sam-pled at dawn are presented. May 1999 dataare for current-year foliage. An asterisk in-dicates that treatments differed significantlyas determined by Fisher’s protected leastsignificant difference test (P ≤ 0.05).
Table 3. Mean carbohydrates concentrations (µmol glucose equiva-lents per gram dry mass) in needles of different age classes. Means aresignificantly different (P ≤ 0.05) between current- and previous-yearfoliage except those followed by ns.
Jul 1997 Sep 1997 Jul 1998 Sep 1998
Total nonstructural carbohydrates (TNC)Current-year foliage 420 433 494 500Previous-year foliage 535 524 624 560
StarchCurrent-year foliage 140 164 255 227Previous-year foliage 205 208 352 172
SucroseCurrent-year foliage 23 8 42 124Previous-year foliage 34 18 81 105
Reducing sugars (RS)Current-year foliage 279 269 238 272Previous-year foliage 330 316 269 387
% TNC as starchCurrent-year foliage 29 ns 33 ns 51 ns 41Previous-year foliage 33 ns 36 ns 53 ns 30
% Soluble sugar as RSCurrent-year foliage 92 ns 97 ns 82 54Previous-year foliage 89 ns 94 ns 69 72
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022

1980, Myers et al. 1999), less potential shading from other fo-liage and higher N concentrations.
Carbon isotopic analysis of foliage is a powerful tool forproviding estimates of long-term WUE (Farquhar and Rich-ards 1984, Farquhar et al. 1989). The δ13C of foliage is an inte-grated measurement of plant physiological processes andenvironmental properties influencing gas exchange during theentire growing season. Differences in δ13C, but not WUEi,among families may partially be a function of strong seasonalchanges in Amax, gs, transpiration rate and WUE. Carbon iso-tope discrimination integrates ci/ca throughout the year; conse-quently, like height or biomass, it reflects the integration ofthese instantaneous measures with environmental conditions.Previous studies with several tree species have observed astronger correlation between WUE (measured by carbon iso-tope discrimination) and growth than between Anet and growth(e.g., Zhang et al. 1994, Major and Johnsen 1996, Johnsen etal. 1999, but see Li 2000). Because changes in ci/ca are a func-tion of photosynthetic capacity or gs, or both, growth of indi-vidual plants may be positively or negatively correlated withleaf δ13C depending on whether the variation in discriminationis associated with changes in photosynthetic capacity orstomatal conductance (Farquhar et al. 1989). In our study, thepositive correlation with δ13C and growth suggests that familydifferences in photosynthetic capacity were the primary causeof variation in δ13C.
Because current-year needles tend to be more sensitive todrought than previous-year needles (Ellsworth 2000), thepresence of older foliage classes during summer may supple-ment carbon demands of younger foliage during drought.However, we found no difference in WUEi between foliageage classes (Table 1). Furthermore, the more negative δ13Cvalues in previous-year foliage indicate a lower WUE than incurrent-year foliage, particularly during the 1999 drought (Ta-ble 3). Although needles were collected from upper-canopybranches exposed to full sunlight, variation in δ13C betweenneedle classes could be a function of variation in solar irrad-iance (Farquhar et al. 1989).
Despite the small range of δ13C values among the four fami-lies, differences were statistically significant and generallymirrored the trend in WUEi, and to some extent, growth of thefour families. The drought-hardy TX-2 was the slowest grow-ing of the four families and generally exhibited the lowestWUEi and δ13C of any family during 1998 and 1999. In con-trast, the faster-growing ACP-1 family had higher WUEi andδ13C during both years. Compared with 1998, all families ex-hibited higher WUEi and δ13C in 1999 because of the drought.Carbon isotope analysis was a good predictor of growth for thefastest- and slowest-growing families, but not for the two fam-ilies exhibiting moderate growth (ACP-2 and TX-1). A stron-ger correlation might exist if all 10 families at the ScotlandCounty site were examined.
Carbon partitioning in needles
Genetic differences in net carbon gain in trees could be a func-tion not only of differences in photosynthetic rate and total leafarea available for carbon fixation, but also of differences in
respiration rate and immediate metabolic demands of leaf tis-sue, including partitioning to storage carbon. Starch accumu-lation in foliage occurs when carbon supply exceeds leafmetabolic demands and carbon export from source leaves. Inloblolly pine, starch accumulation in needles is negatively cor-related with needle respiration (Kuehny and Topa 1998). Lob-lolly pine needles exhibited pronounced seasonal changes inabsolute concentrations of TNC, starch, sugars, and in parti-tioning of TNC to starch and sugars, mirroring seasonalchanges in photosynthesis and shoot and root growth. Concen-trations of TNC and starch in previous-year foliage were high-est in May during spring foliage expansion and peak photo-synthesis, whereas starch concentrations were low in winter.Seasonal peaks in needle starch in the spring near bud breakhave been reported for loblolly and slash (Pinus elliottiiEngelm.) pines (Adams et al. 1986, Cranswick et al. 1987,Gholz and Cropper 1991). High starch concentrations in pre-vious-year foliage are most likely a function of high photo-synthetic rates and reduced carbon demands of expanding fo-liage as their photosynthetic apparatus becomes functionaland respiratory demands decrease (Radoglou and Teskey1997).
Gholz and Cropper (1991) found little seasonal variation insugar concentrations in slash pine needles. We found signifi-cant seasonal differences in foliar soluble sugar concentra-tions (sucrose and reducing sugars), but the seasonal differ-ences were not as pronounced as those for starch. Unlikeseedling studies with loblolly and pond pines (Pinus serotinaMichx.) (Topa and Cheeseman 1992, Kuehny and Topa1998), sucrose was not the predominant sugar in needles, andreducing sugars represented over 70% of soluble sugars inloblolly pine needles. Whole-leaf sucrose concentrations gen-erally reflect vacuolar storage because the pool available fortransport is small, mobile and continuously balanced by thevacuolar pool (Wardlaw 1990). Reducing sugars are more re-flective of immediate metabolic demands of tissues and, assuch, exhibit high turnover.
Seasonal peaks in TNC and starch concentrations generallyfollowed those of photosynthesis. The absence of diurnalchanges in leaf starch concomitant with pronounced diurnalpatterns in Anet indicates that diurnal changes in Anet are inde-pendent of chloroplastic starch concentrations, and would ar-gue against feedback inhibition of photosynthesis by chloro-plastic starch. The lack of diurnal effects on foliar starchconcentrations and starch partitioning is surprising given thestrong diurnal pattern in Anet in loblolly pine, and evidence forcoordinated diurnal partitioning of newly fixed carbon tostarch and sucrose in herbaceous species (e.g., Fondy and Gei-ger 1985, Fondy et al. 1989, Servaites et al. 1989a, 1989b, Liet al. 1992) and in seedlings of some woody species (Topa andCheeseman 1992, Wullschleger et al. 1992, Topa et al. 2001).However, diurnal starch accumulation (i.e., dusk–dawn) inloblolly pine may be more pronounced in seedlings because ofa higher dependency on recently assimilated carbon comparedwith older trees (Vogt et al. 1993).
Although fertilization had little effect on Anet, it reducedTNC concentrations and starch partitioning throughout most
496 YANG, MURTHY, KING AND TOPA
TREE PHYSIOLOGY VOLUME 22, 2002
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022

of the growing season. Texas families exhibited higher con-centrations of TNC and starch, and starch partitioning. LowerTNC and starch concentrations may be a dilution effect as a re-sult of fast growth, a reflection of enhanced metabolic de-mands of leaves, or the result of increased export of sucroseout of leaves. If sugar concentrations are not similarly altered,partitioning of recently assimilated carbon to sugars may bemaintained at the expense of starch. Field studies with P. taeda(Adams et al. 1986) and P. elliottii (Gholz and Cropper 1991)indicated minimal effects of fertilization on foliar starch andsugar concentrations, but marked differences attributable toneedle age. Previous-year foliage consistently exhibitedhigher concentrations of TNC, starch and soluble sugars thancurrent-year foliage (Table 3).
Conclusion
In field-grown loblolly pine, diurnal and seasonal effects onphotosynthesis were more pronounced than family or treat-ment effects. However, family differences might be more pro-nounced under the consistent severe drought conditions thatare experienced by Texas loblolly pine trees in their naturalhabitat. Although loblolly pine from the “Lost Pines” region inTexas is known for its drought hardiness, Texas families ex-hibited lower water-use efficiencies than the more mesic ACPfamilies. Carbohydrate analysis of foliage indicated somefamily and treatment differences in partitioning of recently as-similated carbon, with the two Texas families exhibitinghigher concentrations of TNC and starch, and enhanced parti-tioning toward storage carbon than the ACP families. We con-clude that growth differences among the four families ofloblolly pine are not a function of differences in carbon acqui-sition or partitioning at the leaf level. Ongoing research at theScotland County site suggests that the growth differences maybe closely associated with light interception, i.e., LAI (Hand-est 2001), and with genetic differences in whole-tree carbonallocation, in particular, fine root system carbon demands(Retzlaff 2001).
Acknowledgments
This research was conducted by the Boyce Thompson Institute forPlant Research and North Carolina State University as part of cooper-ative agreement by U.S. DOE Agenda 2020 Sustainable Forestry Re-search Initiative DE-FC Ø 7-97ID13527. Additional funds were pro-vided by the endowment of Boyce Thompson Institute for PlantResearch, the North Carolina State University Tree ImprovementProgram, Bowater, Champion International, Georgia-Pacific Corp.Rayonier, Westvaco and the Weyerhauser Company. We thank DavidMcDermitt, Kai Blaisdell and Richard Phillips for technical assis-tance. This manuscript has not been subjected to a U.S. DOE reviewand should not be construed to represent policies of this agency.
References
Adams, M.B., H.L. Allen and C.B. Davey. 1986. Accumulation ofstarch in roots and foliage of loblolly pine (Pinus taeda L.): effectsof season, site and fertilization. Tree Physiol. 2:35–46.
Albaugh, T.J., H.L. Allen, P.M. Dougherty, L.W. Kress and J.S.King. 1998. Leaf area and above- and belowground growth re-sponses of loblolly pine to nutrient and water additions. For. Sci.44:317–328.
Bilan, M.V., C.T. Hogan and H.B. Carter. 1977. Stomatal opening,transpiration and needle moisture in loblolly pine seedlings fromtwo Texas seed sources. For. Sci. 33:457–462.
Boltz, B.A., B.C. Bongarten and R.O. Teskey. 1986. Seasonal pat-terns of net photosynthesis of loblolly pine from diverse origins.Can. J. For. Res. 16:1063–1068.
Bongarten, B.C. and R.O. Teskey. 1987. Dry weight partitioning andits relationship to productivity in loblolly pine seedlings fromseven sources. For. Sci. 33:255–267.
Chung, H.H. and R.L. Barnes. 1980. Photosynthate allocation inPinus taeda. III. Photosynthate economy: its production, consump-tion and balance in shoots during the growing season. Can. J. For.Res. 10:348–356.
Cranswick, A.M., D.A. Rook and J.A. Zabkiewicz. 1987. Seasonalchanges in carbohydrate concentration and composition of differ-ent tissue types of Pinus radiata trees. N.Z. J. For. Sci. 17:121–130.
Ellsworth, D.S. 2000. Seasonal CO2 assimilation and stomatal limita-tions in a Pinus taeda canopy. Tree Physiol. 20:435–445.
Farquhar, G.D. and R.A. Richards. 1984. Isotope composition ofplant carbon correlation with water-use efficiency of wheatcultivars. Aust. J. Plant Physiol. 11:539–552.
Farquhar, G.D., J.R. Ehleringer and K.T. Hubick. 1989. Carbon iso-tope discrimination and photosynthesis. Annu. Rev. Plant Physiol.Mol. Biol. 40:503–537.
Fondy, B.R. and D.R. Geiger. 1985. Diurnal changes in allocation ofnewly fixed carbon in exporting sugar beet leaves. Plant Physiol.78:753–757.
Fondy, B.R., D.R. Geiger and J.C. Servaites. 1989. Photosynthesis,carbohydrate metabolism, and export in Beta vulgaris L. andPhaseolus vulgaris L. during square and sinusoidal light regimes.Plant Physiol. 89:396–402.
Gholz, H.L. and W.P. Cropper, Jr. 1991. Carbohydrate dynamics inmature Pinus elliotti var. elliottii trees. Can. J. For. Res. 21:1742–1747.
Green, T.H. and R.J. Mitchell. 1992. Effects of nitrogen on the re-sponse of loblolly pine to water stress. I. Photosynthesis andstomatal conductance. New Phytol. 122:627–633.
Greenwood, M.S. and H.A. Volkaert. 1992. Morphophysiologicaltraits as markers for the early selection of conifer families. Can. J.For. Res. 22:1001–1008.
Handest, J. 2001. Effect of nutrient amendments and genotype onstand productivity and crown characteristics of loblolly pine (Pinustaeda L.). M.Sc. Thesis, North Carolina State Univ., Dept. For-estry, Raleigh, NC, 40 p.
Hockman, J.N. and H.L. Allen. 1990. Nutritional diagnosis in youngloblolly pine stands using a DRIS approach. In Sustained Produc-tivity of Forest Soils. Eds. S.P. Gessel, D.S. Lacate, G.F. Weetmanand R.F. Powers. Univ. British Columbia, Faculty of Forestry Pub-lication, Vancouver, B.C., pp 500–514.
Johnsen, K.H. and J.E. Major. 1999. Shoot water relations of matureblack spruce families displaying a genotype × environment interac-tion in growth rate. I. Family and site effects over three growingseasons. Tree Physiol. 19:367–374.
Johnsen, K.H., L.B. Flanagan, D.A. Huber and J.E. Major. 1999. Ge-netic variation in growth, carbon isotope discrimination, and foliarN concentration in Picea mariana: analyses from a half-diallelmating design using field-grown trees. Can. J. For. Res. 29:1727–1735.
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
DIURNAL CHANGES IN GAS EXCHANGE OF LOBLOLLY PINE 497
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022

Jones, M.G.K., W.H. Outlaw, Jr. and O.H. Lowry. 1977. Enzymic as-say of 10–7 to 10–14 moles of sucrose in plant tissues. Plant Physiol.60:379–383.
Kinerson, R.S., K.O. Higginbotham and R.C. Chapman. 1974. Thedynamics of foliage distribution within a forest canopy. J. Appl.Ecol. 11:347–353.
Kuehny, J.S. and M.A. Topa. 1998. Diurnal changes in allocation andpartitioning of recently assimilated carbon in loblolly pine seed-lings. Physiol. Plant. 104:317–328.
Li, B., D.R. Geiger and W.J. Shieh. 1992. Evidence of circadian regu-lation of starch and sucrose synthesis in sugar beet leaves. PlantPhysiol. 99:1393–1399.
Li, B., H.L. Allen and S.E. McKeand. 1991. Nitrogen and family ef-fects on biomass allocation of loblolly pine seedlings. For. Sci. 37:271–283.
Li, C. 2000. Population differences in water-use efficiency of Euca-lyptus microtheca seedlings under different watering regimes.Physiol. Plant. 108:134–139.
Major, J.E. and K.H. Johnsen. 1996. Family variation in photosyn-thesis of 22-year-old black spruce: a test of two models of physio-logical response to water stress. Can. J. For. Res. 26:1922–1933.
Major, J.E. and K.H. Johnsen. 1999. Shoot water relations of matureblack spruce families displaying a genotype × environment interac-tion in growth rate. II. Temporal trends and response to varying soilmoisture conditions. Tree Physiol. 19:375–382.
McKeand, S.E. and J. Svensson. 1997. Sustainable management ofgenetic resources. J. For. 94:4–9.
McKeand, S.E., J.E. Grissom, J.A. Handest, D.M. O’Malley and H.L.Allen. 1999. Responsiveness of diverse provenances of loblollypine to fertilization—age 4 results. J. Sustain. For. 10:87–94.
Michael, D.A., D.I. Dickmann, J.G. Isebrands and N.D. Nelson.1990. Photosynthesis patterns during the establishment year withintwo Populus clones with contrasting morphology and phenology.Tree Physiol. 6:11–27.
Murthy, R. and P.M. Dougherty. 1997. Estimating foliage area ofloblolly pine shoots. For. Sci. 43:299–303.
Murthy, R., P.M. Dougherty, S.J. Zarnoch and H.L. Allen. 1996. Ef-fects of carbon dioxide, fertilization and irrigation on photosyn-thetic capacity of loblolly pine trees. Tree Physiol. 16:537–546.
Murthy, R., S.J. Zarnoch and P.M. Dougherty. 1997. Seasonal trendsof light-saturated net photosynthesis and stomatal conductance ofloblolly pine trees grown in contrasting environments of nutrition,water and carbon dioxide. Plant Cell Environ. 20:558–568.
Myers, D.A., R.B. Thomas and E.H. DeLucia. 1999. Photosyntheticresponse of loblolly pine (Pinus taeda L.) needles to experimentalreduction in sink demand. Tree Physiol. 19:235–242.
Porté, A. and D. Loustau. 1998. Variability of the photosyntheticcharacteristics of mature needles within the crown of a 25-year-oldPinus pinaster. Tree Physiol. 18:223–232.
Radoglou, K. and R.O. Teskey. 1997. Changes in rates of photosyn-thesis and respiration during needle development of loblolly pine.Tree Physiol. 17:485–488.
Retzlaff, W.A., J.A. Handest, D.M. O’Malley, S.E. McKeand andM.A. Topa. 2001. Whole-tree biomass and carbon allocation of ju-venile trees of loblolly pine (Pinus taeda): influence of geneticsand fertilization. Can. J. For. Res. 31:960–970.
Samuelson, L.J. 2000. Effects of nitrogen on leaf physiology andgrowth of different families of loblolly and slash pine. New For.19:95–107.
Samuelson, L.J., J.R. Seiler and P.P. Feret. 1992. Gas exchange andcanopy structure of 9-year-old loblolly pine, pitch pine and pitch ×loblolly hybrids. Trees 6:28–31.
Seiler, J.R. and J.D. Johnson. 1988. Physiological and morphologicalresponses of three half-sib families of loblolly pine to water-stressconditioning. For. Sci. 34:487–495.
Servaites, J.C., D.R. Geiger, M.A. Tucci and B.R. Fondy. 1989a. Leafcarbon metabolism and metabolite levels during a period of sinu-soidal light. Plant Physiol. 89:403–408.
Servaites, J.C., B.R. Fondy, B. Li and D.R. Geiger. 1989b. Sources ofcarbon for export from spinach leaves throughout the day. PlantPhysiol. 90:1168–1174.
Svensson, J.C. 1996. Genetic differences in carbon dioxide assimila-tion, nutrition and biomass production in a 10-year-old loblollypine (Pinus taeda L.) plantation. Ph.D. Thesis, Dept. of Forestry,North Carolina State Univ., Raleigh, NC, 123 p.
Topa, M.A. and J.M. Cheeseman. 1992. Carbon and phosphorus par-titioning in Pinus serotina Michx. seedlings growing underhypoxic and low-P conditions. Tree Physiol. 10:195–207.
Topa, M.A., D.W. Vanderklein and A. Corbin. 2001. Effects of ele-vated ozone and low light on diurnal and seasonal carbon gain insugar maple. Plant Cell Environ. 24:663–677.
van Buijtenen, J.P., M.V. Bilan and R.H. Zimmerman. 1976.Morpho-physiological characteristics related to drought resistancein Pinus taeda. In Tree Physiology and Yield Improvement. Eds.M.G.R. Cannell and F.T. Last. Academic Press, London, pp349–359.
Vogt, K.A., D.A. Publicover, J. Bloomfield, J.M. Perez, D.J. Vogtand W.L. Silver. 1993. Belowground responses as indicators of en-vironmental change. Environ. Exp. Bot. 33:189–205.
Vose, J.M. and H.L. Allen. 1988. Leaf area, stem wood growth, andnutrition relationships in loblolly pine. For. Sci. 34:547–563.
Wang, T., P.M.A. Tigerstedt and A. Vihera-Aarnio. 1995. Photosyn-thesis and canopy characteristics in genetically defined families ofsilver birch (Betula pendula). Tree Physiol. 15:665–671.
Wardlaw, I.F. 1990. The control of carbon partitioning in plants.Tansley Review No. 27. New Phytol. 116:341–381.
Wullschleger, S.D., R.J. Norby and D.L. Hendrix. 1992. Carbon ex-change rates, chlorophyll content, and carbohydrate status of twoforest tree species exposed to carbon dioxide enrichment. TreePhysiol. 10:21–31.
Zhang, J.W., L. Fins and J.D. Marshall. 1994. Stable carbon isotopediscrimination, photosynthetic gas exchange, and growth differ-ences among western larch families. Tree Physiol. 14:531–539.
498 YANG, MURTHY, KING AND TOPA
TREE PHYSIOLOGY VOLUME 22, 2002
Dow
nloaded from https://academ
ic.oup.com/treephys/article/22/7/489/1662467 by guest on 27 June 2022
Related Documents











