CANADA IDRC - 145e POLYPH ENOLS IN CEREALS AND LEGUMES Proceedings of a symposium held during the 36th annual meeting of the Institute of Food Technologists, St. Los, Missouri, 10-13 June 1979 Editor: Joseph H. Hulse ARCHIV HULSE 38091

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CANADA
IDRC - 145e
POLYPH ENOLSIN CEREALSAND LEGUMESProceedings of a symposium held during the 36th annual meeting of theInstitute of Food Technologists, St. Los, Missouri, 10-13 June 1979
Editor: Joseph H. Hulse
ARCHIVHULSE38091

The International Development Research Centre is a public corporationcreated by the Parliament of Canada in 1970 to support research designed toadapt science and technology to the needs of developing countries. TheCentre's activity is concentrated in five sectors: agriculture, food and nutritionsciences; health sciences; information sciences; social sciences; andcommunications. IDRC is financed solely by the Government of Canada; itspolicies, however, are set by an international Board of Governors. TheCentre's headquarters are in Ottawa, Canada. Regional offices are located inAfrica, Asia, Latin America, and the Middle East
© 1980 International Development Research CentrePostal Address: Box 8500, Ottawa, Canada Ki G 3H9Head Office: 60 Queen Street, Ottawa
Hulse, J.H.IDRC-1 45 e
Polyphenols in cereals and legumes: proceedings of a symposium heldduring the 36th annual meeting of the Institute of Food Technologists, St.Louis, Missouri, 10-13 June 1979. Ottawa, Ont., IDRC, 1980. 72 p.: ill.
/IDRC publication!, /conference paper!, /nutrition research!, /chemicalanalysis!, !sorghunil, /millet! - /nutritive value!, /acids!, ,'toxins!.
UDC: 612.392.73 ISBN: 0-88936-234-3
Microfiche edition available

Polyphenols in Cereals and LegumesProceedings of a symposium held during the36th annual meeting of the Institute of FoodTechnologists, St. Louis, Missouri, 10-13 June 1979
Editor: Joseph H. Hulse
Cosponsored by
The International Union of Food Science and TechnologyThe Institute of Food TechnologistsThe International Development Research Centre
IDRC- 145e
mILSE

Contents
3 Introduction and Recommendations
9 Introduction et recommandations
15 Vegetable Tannins Structure and Biosynthesis R.K. Gupta and E. Haslam
25 Factors Affecting the Polyphenols of Sorghum and Their Development and Loca-tion in the Sorghum Kernel L.W. Rooney, M.E. Blakely, F.R. Miller, and D.T.
Rosenow
36 Biochemical Effects of Sorghum Polyphenols Larry G. Butler, Ann E. Hagerman,and Martin L. Price
39 Treatments of Sorghum Grain That Reduce the Assayable Tannin Content andTheir Effect on the Nutritional Value of the Grain Martin L. Price and LarryC. Butler
43 Sorghum Polyphenols and Bird Resista nce Roger W. Bullard and Donald J. Elias
50 Polyphenols in Pennisetum Millet R.D. Reichert, C.G. Ynungs, and D.A. Chris-tensen
61 The Nutritional Role of Polyphenols in Beans Ricardo Bressani and Luiz G. Elias
69 Bibliography
Factors Affecting the Polyphenols of Sorghum andTheir Development and Location in the Sorghum Kernel
LW. Rooney', M.E. Blakely', F.R. Miller2, and D.T. Rosenow3
In sorghum grain, polyphenols are located primarily in the pericarp and the testa. Genesregulating pericarp colour and presence of the testa exist. Bird-resistant sorghums have a pigmentedtesta containing condensed tannins that remain in the kernel at maturity. In nonbird-resistantsorghums, the testa is resorbed during maturation. The endosperm of some white, colourlesssorghums contains colour precursors that turn greenish-black when exposed to alkali. Variation incolour, changes in structure during maturation, polyphenol content, and nutritive value of sorghumgenotypes are reviewed.
The major intent of this paper is to review thekernel structure of sorghum and discuss thefactors that affect the polyphenols present in thegrain. No attempt is made to cover all of theliterature on the polyphenols of sorghum as otherpapers cover other specific areas. Certain ideasrecorded here are based on observations gainedover the years and are presented to stimulatediscussion and further elucidation of their vali-dity.
Structure of Sorghum Kernel
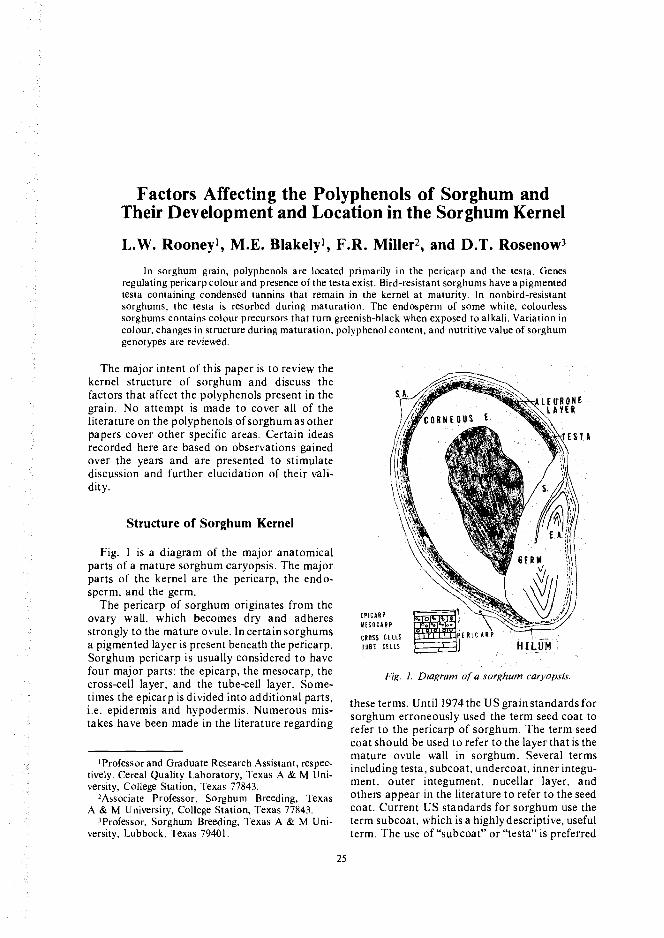
Fig. 1 is a diagram of the major anatomicalparts of a mature sorghum caryopsis. The majorparts of the kernel are the pericarp, the endo-sperm, and the germ.
The pericarp of sorghum originates from theovary wall, which becomes dry and adheresstrongly to the mature ovule. In certain sorghumsa pigmented layer is present beneath the pericarp.Sorghum pericarp is usually considered to havefour major parts: the epicarp, the mesocarp, thecross-cell layer, and the tube-cell layer. Some-times the epicarp is divided into additional parts,i.e. epidermis and hypodermis. Numerous mis-takes have been made in the literature regarding
'Professor and Graduate Research Assistant, respec-tively. Cereal Quality Laboratory, Texas A & M Uni-versity, College Station, Texas 77843.
2Associate Professor, Sorghum Breeding, TexasA & M University, College Station, Texas 77843.
3professor, Sorghum Breeding, Texas A & M Uni-versity, Lubbock, Texas 79401.
25
(ICARP
MOSt CA AS
CROSS CttlS
JURO COOLS HILUM
S. A
%.fl'lIn.JL..SI
LEURONELAYER
Fig. 1. Diagram of a sorghum caryopsis.
these terms. Until 1974 the US grain standards forsorghum erroneously used the term seed coat torefer to the pericarp of sorghum. The term seedcoat should be used to refer to the layer that is themature ovule wall in sorghum. Several termsincluding testa, subcoat, undercoat, inner integu-ment, outer integument, nucellar layer, andothers appear in the literature to refer to the seedcoat. Current US standards for sorghum use theterm subcoat, which is a highly descriptive, usefulterm. The use of "subcoat" or "testa" is preferred
UI A
Tab
le I
. The
gen
otyp
e, p
heno
type
, and
mar
ket c
lass
of
som
e co
mm
on s
orgh
um li
nes.
Con
tain
s w
axy
endo
sper
m (
wxw
x) a
ll ot
hers
are
non
wax
y (W
xWs)
.'T
he c
lass
cou
ld p
roba
bly
be y
ello
w, c
urre
ntly
but
, red
efin
inon
of
the
whi
te c
lass
to p
erm
it st
aine
d ke
rnel
s w
ould
let i
t be
clas
sifi
ed a
s w
hite
.
Var
iety
Tes
taSp
read
erC
olou
rM
eso-
End
o-Pl
ant
carp
spe
rm c
olou
rT
esta
BT
x319
7b1
b1B
,B,
SSR
Ryy
iizz
yeye
PPQ
Qno
BT
x378
b1b1
8B,
SSR
RY
YII
7.1
yeye
PPQ
Qno
BT
x399
b1b1
8,B
,SS
RR
YY
iizz
yeye
PPQ
Qno
BT
x398
b1b1
B,B
SSR
RY
Yii
zzye
yePP
noPi
nk K
afir
b1b1
B,B
,SS
RR
YY
IZ
Zye
yePP
noR
ed K
afir
bb1B
,B,
SSR
RY
YII
ZZ
yeye
PPQ
Qno
RT
x700
0bb
B,B
,SS
RR
YY
Iizz
yeye
PPQ
Qno
RT
x7O
78b1
b1B
,B,
SSR
RY
Yii
zzye
yePP
rio
RT
x4l4
b1b1
B,B
,SS
RR
YY
iizz
yeye
PPQ
Qno
RT
xO9
b1b1
B,B
,SS
RR
yytt
zzye
yePP
noD
oubl
e D
war
fB
B1B
,B,
ssR
Ryy
iizz
yeye
PPQ
Qye
sFe
teri
taR
Tx4
28b1
b1B
,B,
SSrr
yyJl
ZZ
yeye
P-q'
qno
RT
x43O
b1b,
B2B
,SS
rryy
llZ
ZY
eYe
PPQ
Qno
RT
x253
6b1
b1B
,B,
SSrr
yyii
ZZ
YeY
e PP
noR
Tx2
566
BB
18,B
2ss
RR
YY
IIZ
Zye
yePP
yes
RT
x256
7b1
bB,B
,SS
rryy
iiZ
ZY
eYe
PPQ
Qno
77C
S2b1
bB,B
,SS
RR
yyll
ZZ
yeye
P-qq
no(S
CO
I7O
-6)
Heg
art
BB
1B,B
,ss
RR
yyii
zzye
yePP
yes
(P13
4911
)
Dob
bsB
1B1B
,B,
SSR
Ryy
ticc
yeye
PPQ
Qye
sSh
allu
B1B
1b,b
,SS
RR
yyii
ZZ
yeye
ppqq
noSC
0748
-5b1
b1B
,B,
SSrr
YY
lIZ
Zye
yePP
noB
Tx6
I5bb
B,B
,SS
RR
yyit
zaye
yepp
qqno
NSA
74O
b1b1
B,B
,SS
RR
yyii
zzye
yePP
no
F-10
4 sd
b1b1
B,B
,SS
RR
yyti
ccye
yePP
no
SCO
IO4
sdB
1BJB
,B,
ssR
RY
YII
ccye
yePP
yes
End
o-Pl
ant
App
eara
nce
Mar
ket
sper
mco
lour
of k
erne
lcl
ass
nony
ello
wpu
rple
chal
ky w
hite
whi
te2
nony
ello
wpu
rple
red
yello
wno
nyel
low
purp
lere
dye
llow
nony
ello
wpu
rple
red
yello
wno
nyel
low
purp
lelig
ht p
inki
sh-r
edye
llow
nony
ello
wpu
rple
brig
ht r
edye
llow
nony
ello
wpu
rple
red
yello
wno
nyel
low
purp
lere
dye
llow
nony
ello
wpu
rple
red
yello
wno
nyel
low
purp
lech
alky
whi
tew
hite
'no
nyel
low
purp
lech
alky
whi
tesp
lotc
hes
brow
n
nony
ello
wre
dw
hite
whi
te'
yello
wpu
rple
yello
wye
llow
yello
wpu
rple
yello
wye
llow
nony
ello
wpu
rple
redd
ish-
brow
nbr
own
yello
wpu
rple
yello
wye
llow
nony
ello
wre
dw
hite
, pea
rly
whi
te'
nony
ello
wpu
rple
chal
ky w
hite
splo
tche
sbr
own
nony
ello
wpu
rple
brow
nbr
own
nony
ello
wta
nw
hite
, pea
rly
whi
te'
nony
ello
wpu
rple
brow
nish
-yel
low
yello
wno
nyel
low
'(w
axy)
nony
ello
w(f
lour
y)no
nyel
low
(sug
ary)
nony
ello
w
tan
purp
le
purp
le
purp
le
chal
ky w
hite
chal
ky w
hite
whi
te
dark
red
dish
-bro
wn
whi
te'
whi
te'
whi
te'
brow
n
Peri
carp
whi
te, t
hick
red,
thic
kre
d, th
ick
red,
thic
kre
d, th
inre
d, th
in w
/inte
n.re
d, th
ick
red,
thic
kre
d, th
ick
whi
te, t
hick
whi
te, t
hick
whi
te, t
hin
w/in
ten.
whi
te, t
hin
wjin
ten.
whi
te, t
hin
w/in
ten.
whi
te, t
hin
whi
te, t
hin
whi
te, t
hin
w/in
ten.
whi
te, t
hick
whi
te, t
hick
whi
te, t
hin
yello
w, t
hin
w/in
ten.
whi
te, t
hick
whi
te, t
hick
whi
te, t
hick
red,
thic
k, w
/inte
n.

ROONEY ET AL.: DEVELOPMENT AND LOCATION IN SORGHUM 27
to avoid the confusion that exists among theseterms. The presence of a testa is controlled by B1and B2 genes. Most commercial sorghums grownin the United States do not have a testa.
Other structural features of the sorghum kernelaffect polyphenol content. For instance, the hylarand stylar areas of the kernel are coloured evenwhen the sorghum pericarp is colourless. Theglumes contain polyphenols that sometimesaffect the kernel especially under humid condi-tions or when rainfall occurs during kernelmaturation.
The Appearance of Sorghum (Colour)
The visual appearance of sorghum is basedupon several interacting factors that influence thecolour of the kernel as viewed by the eye. Thepericarp colour and thickness; the presence,colour, and thickness of the testa; and the colourof the endosperm can affect the visual appearanceof sorghum colour. The descriptive terms used torefer to grain colour are confusing unless accom-panied by a description of the genetics of thefactors affecting grain colour.
Attempts to measure and classify the colour ofsorghum have varied from visual rating systemsto use of colour measurements with instruments(Khan et al. 1979). We have used the colour dif-ference meter to measure colour of grains in termsof tristimulus readings. The use of tristimulustechniques is promising and may prove useful topredict the colour of products produced from thegrain. Objective methods of classifying graincolour are needed to help avoid confusion aboutsorghum colour.
Genetics of Sorghum Colour
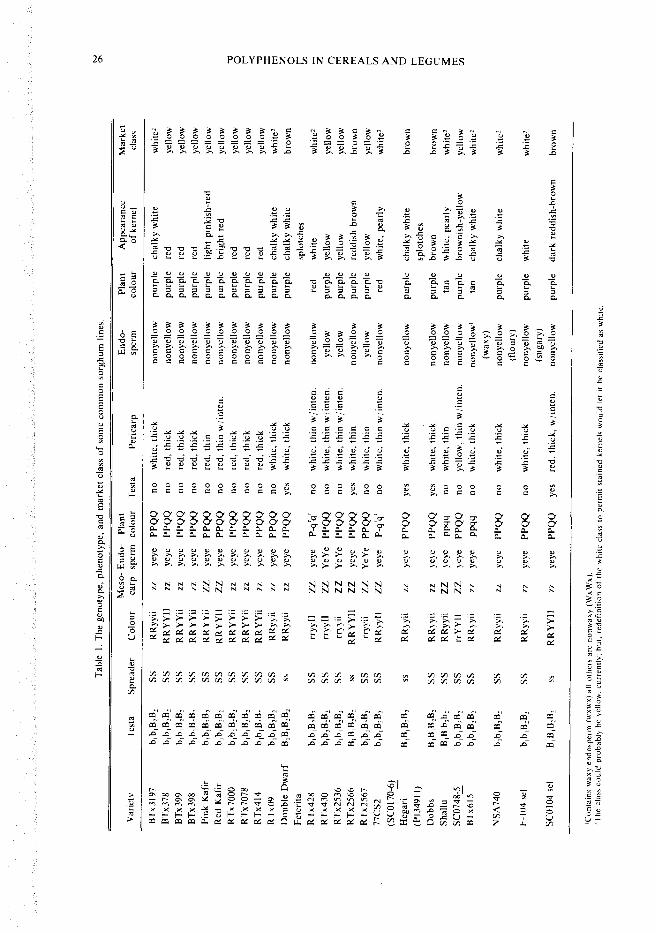
The genetics of grain colour are complex, butconsiderable information is available (Quinbyand Martin 1954; Schertz and Stephens 1966;York 1976). A summary of the genetics of pen-carp colour and thickness, plant colour, andendosperm colour for some sorghum varietiesand lines is presented in Table 1. The genetics isbased on experience with numerous crosses in-volving the sorghum lines and varieties listed inTable 1. It is presented to serve as a reference forfuture work. It is desirable that all other workerspresent the genetics of the material they areworking with in theirstudies. Such information isuseful in the interpretation of results.
Pericarp colour is determined by the following:B1-B2-S-R-Y-1-Z. The presence of the testa is con-trolled by the B1-B2- genes. If they are both domi-nant, the sorghum kernel has a pigmented testa. Ifone set of B1B2 genes is homozygous recessive(i.e. b1bB2B2) then the pigmented testa is absent.The B1B2 genes affect pericarp colour becausewhen they are dominant and a dominant spreadergene (S-) is present, brown pigments appear in thepericarp. The colour is "spread" from the testainto the epicarp of the penicarp. The shade ofbrown in the epicarp is conditioned by the R-Y-I-genes. Thus, it is important to know the geneticsof the pericarp.
The R-Y- genes determine whether the pericarpis red, white, or lemon yellow. The pericarp is redwhen the genes are R-Y-. It is white when thegenes are RRyy or rryy. It is lemon yellow whenthe genes are rrY-. The lemon yellow colour isbleached by the sun and usually turns a dullbrownish-yellow colour by the time the grain hasdried to harvest moisture. Often the lemon yellowcolour is retained in the pericarp under theglumes. Yellow pericarp colour is not associatedwith yellow endosperm and should not beconfused with it. The intensifier (I-) affects the in-tensity of the colour. For instance, a kernel withdominant R-Y- and I- genes will appear brightred when the genotype is R-Y-ii. The testa colouris purple for tptp and brown when Tp- (Casady1975). Hegari and Feterita have the purple testa.
Mesocarp Thickness
The Z- genes control the thickness of the pen-carp. A dominant Z- gene gives a sorghum with athin penicarp. Homozygous recessive (zz) gives athick pericarp. The thickness relates to the num-ber of layers in the mesocarp and the presence ofstarch granules. The presence of a thick, starchymesocarp causes the pericarp to mask the colourpigments present in the testa and endosperm. Thethin pericarp (Z-) is translucent and permitscolour of the testa and / or endosperm to affect theappearance of the kernel. The pearly sorghumshave dominant Z genes and usually a high pro-portion of corneous endosperm. Chalky sorghumkernels are those with a thick, starchy mesocarp.The thickness of the pericarp (mesocarp) varieswhen recessive (zz) is present, which indicatesthere may be additional modifiers influencingpericarp thickness, i.e. the difference betweenTx09 and BTx3197. This had led York (1977) topropose a new scheme for classifying the genes forpericarp colour and thickness. The material pre-sented in Table I follows the old system (Schertzand Stephens 1966).

28 POLYPHENOLS IN CEREALS AND LEGUMES
Endosperm Colour
Yellow endosperm sorghums that have highlevels of carotenoid pigments exist. In true yellowendosperm sorghum varieties, the pericarp is thinand colourless (R-yy or rryy), the testa is absent,and the colour of the kernel appears yellow be-cause of the carotenoid pigments in the endo-sperm. If the kernel has a thick mesocarp (zz) itwill sometimes appear white because the yellowcolour of the endosperm is masked by the thickmesocarp. Some sorghums are called bronzesorghums because they have a thin red pericarp,no testa, and a yellow endosperm. Thus the ap-pearance of the kernel is bronze because theyellow endosperni shows through the thin redpericarp and modifies the colour. When a thickpericarp is present, the yellow endosperm colouris masked.
The aleurone of sorghum is usually colourless,although under certain circumstances of insect ordisease attack, pigments have been observed inthe aleurone layer (see Fig. 2C). A coloured aleu-rone exists in barley (Hordeum vulgare) and corn(Zea mays); but none has been reported for sor-ghum yet.
Glume and Plant Colour
Another important consideration is plant andglume colour which affects polyphenol content ofthe grain. Usually the plant colour is consideredas purple or tan. The genes controlling plantcolour are: P-Q-. Tan plant colour is recessive.Actually four colour variations are described:P-Q-, reddish-purple; P-qq, blackish-purple;p..qrqr red; and ppqq, tan. Usually, for practicalpurposes, the plant colour is considered to bepurple or tan. No attempt to distinguish amongthe other genotypes is made.
Glume colour is associated with plant colour.The best way to determine glume colour is to exa-mine the colour of the inside of the glume after thekernel is removed. Glume colour is determinedby the plant colour loci P and Q. Black- and red-glumed plants are P-, and mahogany- and sienna-glumed plants are recessive pp. The Q-locus con-ditions the shade of purple pigmentation present.Glumes that have intent red, black, or purplecolour have a tendency to stain the sorghumkernel because the polyphenolic pigments leachinto the pericarp. In addition, injury to the kernelduring development, by insects or moulds, tendsto produce coloured pigments that are related toplant colour.
Other Genes Affecting Pericarp Colour
Other genes have been reported that affect pen-carp colour and many more probably exist in theworld collection of sorghums. A gene (Pb-) thatcauses purple spots in the pericarp has been re-ported which may function in some commercialsorghums. For instance, new white pericarpsorghums released by the Texas AgriculturalExperimental Station appear to have purplespecks in the pericarp. Brown wash genes (Bw1-Bw2-) exist that have an effect on penicarp colourwhen B 1-B2- genes are present with a homozygousrecessive (ss) spreader. The brown colour washesthrough the pericarp, but it is different from thespreader genes. Another gene, sun red, (Rs1-),causes the colour of the pericarp to become redwhen it is exposed to the sun during maturation.Then it disappears and the pericarp colour isdetermined by the R-Y-I- genes.
Market Class of Sorghums
In the United States Standards for sorghums,four classes of sorghum are recognized, yellow,white, brown, and mixed. The major class of sor-ghum in the US is the yellow class. The sorghumof commerce is usually US number 2 yellow sor-ghum. The market class of sorghum is determinedby visual appearance of the grain; the presence ofa pigmented testa means the grain goes into thebrown class (Table 1).
The class, yellow sorghum, can contain grain ofany colour pericarp as long as it does not containa pigmented testa. Therefore yellow sorghum cancontain grain with white, red, and lemon yellowcolour pericarp. It cannot contain more than 10%of kernels with a pigmented testa. In practice, USyellow sorghum will contain almost no kernelswith pigmented testa.
The white sorghum class contains kernels with98% white pericarps without a testa. This classcannot contain more than 2% of nonwhitesorghum kernels. In addition, kernels with awhite pericarp and spots are considered nonwhitekernels. In practice, the white class is not ofimportance in current marketing of sorghums,but it could become of more importance becausenew sorghum hybrids with high yield potentialhave a white pericarp without a testa. The whiteclass will be most useful for maintenance of grainquality for food use where colour and flavour areprime concerns.
The brown sorghum class consists of kernelswith a pigmented testa (B1-B2-) regardless of pen-carp colour. The brown grain class has depressed

ROONEY ET AL.: DEVELOPMENT AND LOCATION IN SORGHUM 29
nutritional value compared to the yellow andwhite classes of sorghum. The bird-resistant high-tannin sorghums belong to the brown class. Themost effective bird-resistant sorghums containB1-B2-S-R-Y- genes. These genes appear to havethe most serious effects on the nutritional value ofsorghum. They consistently inhibit enzymes andcontain high levels of condensed tannins. Otherbrown sorghums with a testa (B1-B2-ss) have vari-able effects on nutritional value. At this point, thesafest, most conservative method is to continueplacing all sorghums with a testa in the brownclass. The overall quality of brown grain for feedand food is definitely lower than the yellow andwhite grains.
The yellow sorghum of US origin contains littleif any condensed tannins and has nutritionalvalue at least 95% equivalent of corn and greaterfor some species. Certain countries produce bird-resistant, mould-resistant, high-tannin sor-ghums. Problems in marketing arise when sor-ghums with high levels of tannins are sold asyellow sorghum. The buyers are not impressedwith the performance of the sorghums and theydevelop a low opinion of all sorghums. Currentefforts to educate the grain trade about sorghumquality variation include use of methods to detectthe presence of a testa or undercoat. Educationalprograms have helped eliminate the productionof bird-resistant sorghum in Australia and arereducing its production in Argentina. Thesecomments are presented here because of their re-levance to the overall question of tannins in sor-ghum.
The move toward production of whitesorghum in the USA can be seen as a positive steptoward utilization of sorghum in foods. The bestway of solving the tannin problem is to growwhite or yellow sorghums and discourage growthof brown where they are not really needed for birdresistance. Nonetheless, there are areas in theUSA and elsewhere in the world where bird-resistant sorghums are required. Thus, there is anurgent need to understand the chemistry ofsorghum-condensed tannins, to develop effectivemethods to eliminate or reduce their detrimentaleffects on nutritional value.
Methods of Tannin Analysis in GrainMarketing
Tannin analyses of sorghum are difficult evenfor research laboratories to perform. Substantialprogress has been made toward better methods ofpolyphenol analysis of sorghum (Daiber 1975;Maxson and Rooney 1972; Price and Butler 1977;
Price et al. 1978; Hagerman and Butler 1978).Application of these analytical methods to grainmarketing situations is extremely difficultbecause of the need for simplicity, speed, reli-ability, and relation to nutritive value. No analy-sis comes close to meeting all these requirements,but the most effective for use in a grain marketingsituation is the chlorox bleach method or pocketknife test to determine the number of kernels witha pigmented testa. The Federal Grain InspectionService of the United States Department of Agri-culture (USDA) detects brown sorghums byvisual examination of the grain sample prior toand after scraping the kernel to look for the testaor subcoat. In certain instances, the chloroxbleach method is used as an unofficial aid to helpdetermine the level of brown grain in sorghumsamples, but it is not an official method.
For inexperienced personnel, the chloroxbleach method is a good method of determiningwhether a sample of sorghum contains grain witha pigmented testa. The percent of brown kernelsin the sample can be used as a semi-quantitativeestimate of the levels of condensed tannins. Thechlorox bleach test was devised to remove thepericarp from cereals to facilitate estimates ofgerm damage (Weak et al. 1972). We have usedthe method for several years to detect brown sor-ghums in yellow sorghums. In 1975 the methodwas demonstrated to the grain industry in nineEuropean countries and has been widely usedsince as an aid to determine the levels of browngrain (kernels with a testa) in market samples.
The chiorox bleach method is based on theremoval of the pericarp which exposes the pig-mented testa. In addition, the testa usuallybecomes an intense dark colour. Therefore thebrown sorghum after bleaching is a dark colour,while nonbrown sorghums have a light colour.Usually it is relatively easy to distinguish betweenthe brown and nonbrown sorghums, butweathering, leaching of pigments into the grain,insect and disease damage can cause nonbrownkernels to bleach darker than usual. Thereforereasonable care must be used in interpreting theresults. Pericarp thickness and ease of removalvaries among sorghum kernels within a varietyand among varieties. In addition, testa thicknessvaries within kernels in a single variety. If thetemperature, heating time, and methods ofstirring are varied, erroneous results can be ob-tained because the testa can be completely orpartially removed. Thus, it is important to set upthe procedure and follow it carefully.
The following procedure had been found mostuseful in our laboratory for market samples.Potassium hydroxide 5.0 g is added to 15.0 g of
30 POLYPHENOLS IN CEREALS AND LEGUMES
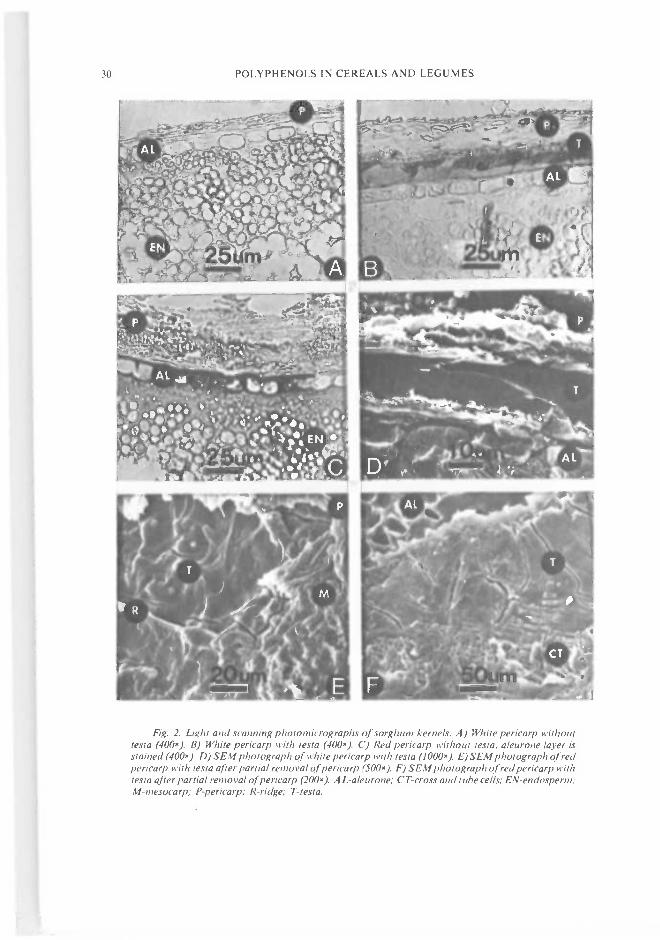
Fig. 2. Light and scanning photomicrographs of sorghum kernels. A) White pericarp withoutlesta (400x). B) White pericarp with testa (4UO). C) Red pericarp without testa, aleurone layer isstained (4OO). D) SEM photograph of white periearp with tesla (IOOO). E) SEM photograph of redpericarp with testa after partial removal oJperwarp (5OO). F) SEM photograph of red pericarp withtesta after partial removal ofpericarp (2OO). A L-aleurone; CT-cross and tube cells; EN-endosperm;tVI-mesocarp; P-pericarp; R-ridge; T-Iesta.

ROONEY ET AL.: DEVELOPMENT AND LOCATION IN SORGHUM 31
sorghum in 70 ml of 5.25% aqueous sodium hypo-.chlorite (NaOC1). The mixture is heated withstirring at 60 °C for 7.0 mm. The sorghum isplaced in a tea strainer and rinsed with tap waterand dried. The procedure can be modified for useunder different circumstances by using knownreference samples of sorghums with and withoutthe testa layer.
The US market class for sorghums with variousgene combinations is presented in Table 1. Thechiorox bleach test is most useful in the market-.ing of grains, but it can also be used to select forbrown sorghums in breeding programs (Kofoid etal. 1978).
The Prussian blue test (Price and Butler 1977)has excellent potential for use in grain marketing,but it has the disadvantage of requiringuncommon reagents, the glassware is stained, andit works best on ground samples. We have notbeen able to achieve satisfactory results on wholekernels. None of the above disadvantages ismajor, but the chlorox bleach method is muchsimpler for use in the grain marketing system.
The chlorox bleach test can be used in market-ing to determine the percentage of brown grain. Agrain buyer can decide the price reduction for cer-tain levels of brown kernels. In this way thechlorox bleach method can be used as an approx-imate index of the nutritive value of sorghum.Should a user of sorghum wish to determine thenutritive value of the common bird-resistantsorghums grown in the area from which the sor-ghum is received, a value can be arrived at by thismethod. Further work is needed to develop reli-able methods of determining the content ofcondensed tannins in sorghum to be able toaccurately predict its nutritional value.
The Structure of Sorghum Kernels with aTesta
We have examined numerous kinds ofsorghums with light and scanning electron micro-scopy (SEM) to determine where the polyphenolswere located. Fig. 2 is a comparison of thestructure of kernels with and without the testa.The pericarp colour was b1b1B2B2 ssrryyiizzfor the grain in Fig. 2A, while it was B1BB2B2ssrryyiizz for the grain in Fig. 2B. In other wordsboth had a thick white pericarp. A part of thepericarp in Fig. 2A was lost during sectioning.Both appeared chalky white, but the one with atesta would be in the brown market class. Wehave not observed any sorghum kernels that con-tain an unpigmented testa layer. Although, insections of many kernels without a pigmentedtesta, we have seen a remnant tissue which might
be classified as nonpigmented material left fromthe integuments or nucellus.
The testa sometimes appears as distinct blockycells consisting of two overlapping layers (Fig.28). The layers in Fig. 2B are of different colourintensity. One layer was reddish-brown while theother was a light yellowish-brown when viewedwith bright field microscopy without staining.Usually the proportions of the two layers do notremain constant around the kernel. In othersections, the testa appeared as a single stripwithout structure, whereas in yet others blockysubunits were observed. Fig. 2D is a cross sectionviewed with SEM. No evidence of intracellularstructure in the testa was observed even afterattempts were made to extract the tannins withvarious solvents. We have not examined theimmature testa with SEM. A top view of themature testa is presented in Fig. 2E and F. Thissection was prepared by removing the pericarpafter the kernel was softened with ethanol. Insome kernels the testa appeared to have ridgeswhich rose above the main plane of the testa (Fig.2E). However, in others the ridges were notobserved.
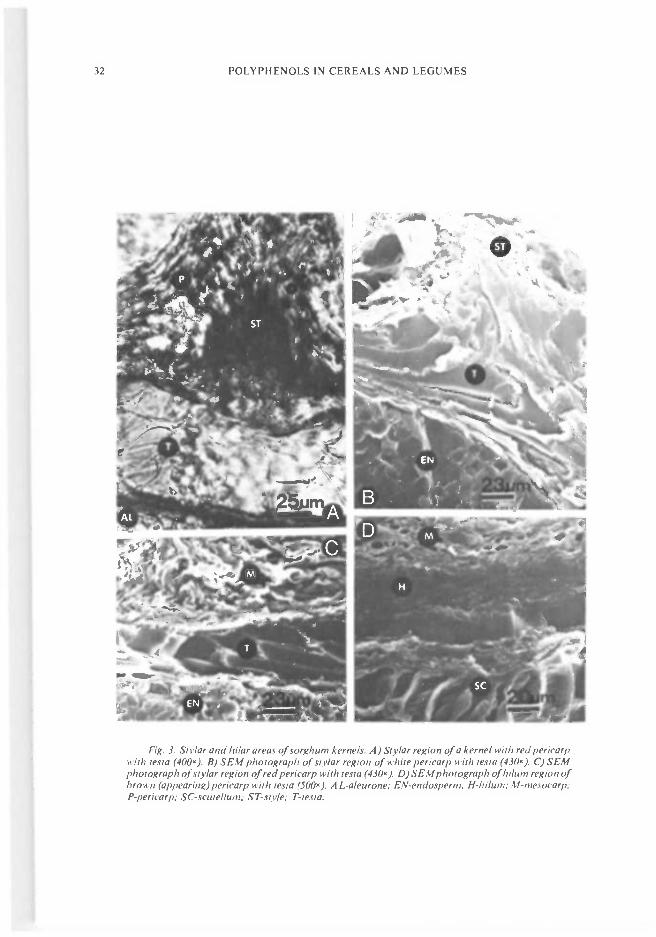
The thickness of the testa varies around thekernel (Blakely et al. 1979). The thickest areas(100-140 Mm) occur just below the stylar region(Fig. 3) whereas the thinnest region (lO-3OMm) islocated on the opposite side of the hilum. Thestructure of the stylar and hilum is presented inFig. 3. The hilum (hilar region) appeared to be acontinuation of the testa, but the pigments aredark black. At the hilum, the testa and aleuronelayers tend to be meshed together. The pigmentsare probably different from those of the testa.Mean and range of testa thickness for five varie-ties measured at each of eight specified locationson the kernel were presented by Blakely et al.(1979). The means ranged from l8Mm to I5OMm.
We have found a sorghum kernel that has apartial testa. The genetics are unknown, but thepartial testa characteristics can be manipulated.Photos of kernels with a partial testa arepresented in Fig. 4. Sorghum kernels with apartial testa have been reported (Quinby andMartin 1954), but they have been difficult to con-firm. However, this partial testa is easily con-firmed because the pericarp is thin. Fig. 4 C and Dwere sections from kernels with the partial testa.The testa disappears (Fig. 4 C and D). Other partsof the kernel such as the aleurone and pericarp areunchanged. The testa, in the kernels with a partialtesta, appeared as small blocky units rather than asmooth stripe. When the testa was absent, thestructure was similar to that of kernels withoutthe testa.
32 POLYPHENOLS IN CEREALS AND LEGUMES
Fig. 3. Stlar and hi/ar areas of sorghum kernels. A) Sivlar region of a kernel with red per/carpwith testa (4OO). B) SEM photograph of sit/ar region of while per/carp with tesla (43O1. C) SEMphotograph of sty/ar region of red pericarp with lesta (43O). D) SEMphotograph of hi/urn region ofbrown (appearing) pericarp with testa (500x). A L-a/eurone; EN-endospertn. H-h/turn; M-tnesocarp;P-per/carp; SC-scute//u,n; S T-szt'/e; T-testa.
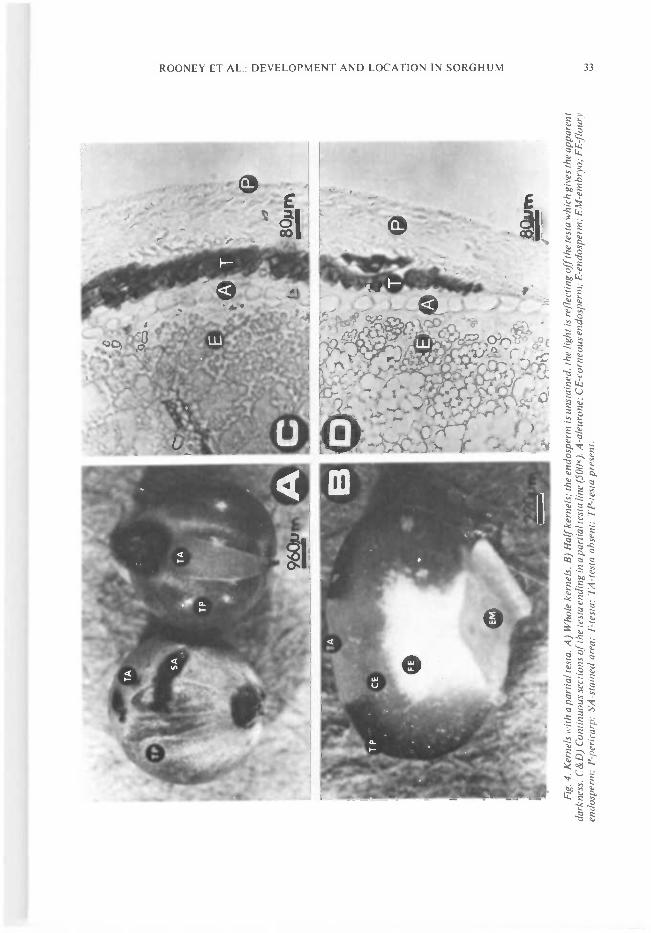
Fig
. 4. K
erne
ls w
ith a
par
tial t
esta
. A)
Who
le k
erne
ls. B
) H
alf k
erne
ls: t
he e
ndos
perm
is u
nsta
ined
, the
ligh
t is
refle
ctin
g of
f the
test
a w
hich
give
s th
e ap
pare
ntda
rkne
ss. C
&D
) C
ontin
uous
sec
tions
of t
he te
sta
endi
ng in
a p
artia
l tes
ta li
ne (
500*
). 4
-ale
uron
e; C
E-c
orne
ous
endo
sper
m; E
-end
ospe
rm; E
M-e
mbr
yo; F
E-f
lour
yen
dosp
erm
; P-p
eric
arp;
SA
-sta
ined
are
a; T
-tes
ta; T
A-t
esta
abs
ent;
TP
-tes
ta p
rese
nt.
34 POLYPHENOLS IN CEREALS AND LEGUMES
1000-
900-
800
700-
600-
400-
300-
200-
100-
0
F
a,I gatiI,
a
a,I \DDF,/
- '-I --15 20 25 30 35 40
DAYS AFTER BLOOM
' DDF\I,,
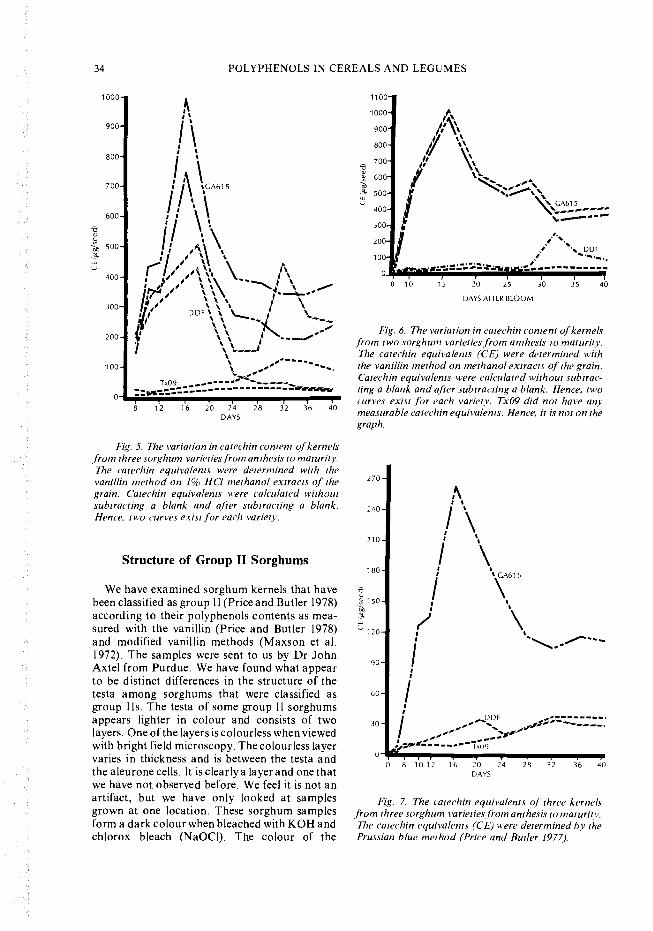
\ \ - Fig. 6. The variation in catechin content of kernels- from two sorghum varieties from anthesis to maturity.
The catechin equivalents (CE) were determined with-- . the vanillin method on methanol extracts of the grain.'
Catechin equivalents were calculated without subtrac-TxDS --- ting a blank and after subtracting a blank. Hence, two
8 12 16 20 24 28 32 36 40 measurable catechin equivalents. Hence, it is not on theDAYS
curves exist for each variety. Tx09 did not have any
graph.
Fig. 5. The variation in catechin content of kernelsfrom three sorghum varieties from anthesis to maturity.The catechin equivalents were determined with thevanillin method on 1% HCI methanol extracts of thegrain. atechin equivalents were calculated withoutsubtracting a blank and after subtracting a blank.Hence, two curves exist for each variety.
Structure of Group II Sorghums
We have examined sorghum kernels that havebeen classified as group II (Price and Butler 1978)according to their polyphenols contents as mea-sured with the vanillin (Price and Butler 1978)and modified vanillin methods (Maxson et al.1972). The samples were sent to us by Dr JohnAxtel from Purdue. We have found what appearto be distinct differences in the structure of thetesta among sorghums that were classified asgroup us. The testa of some group II sorghumsappears lighter in colour and consists of twolayers. One of the layers is colourless when viewedwith bright field microscopy. The colourless layervaries in thickness and is between the testa andthe aleurone cells, It is clearly a layer and one thatwe have not observed before. We feel it is not anartifact, but we have only looked at samplesgrown at one location. These sorghum samplesform a dark colour when bleached with KOH andchlorox bleach (NaOCI). The colour of the
270
1\Is240 I\210
/t\
I S180 \615
150
(1u120
90 I
60 /DDF -
30 /
I\
"
iGA615
'
\\\\\ \ _...
-o
R
1100
1000
700
600
500
400
300
200
100
0
8 10
0
0 8 10 12 16 20 24 28 32 36 40DAYS
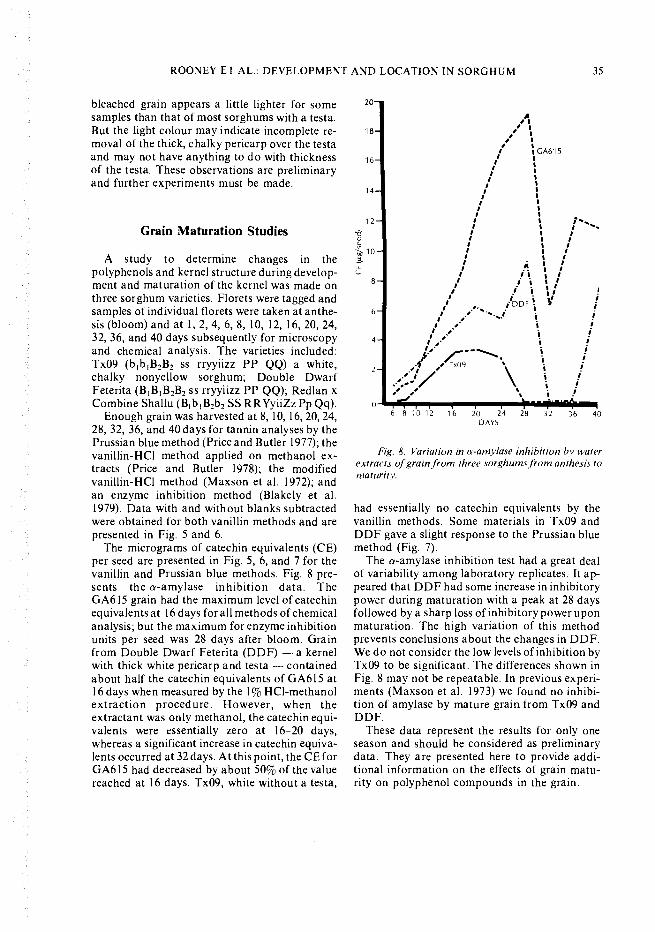
Fig. 7. The catechin equivalents of three kernelsfrom three sorghum varieties from an thesis to maturity.The catechin equivalents (CE) were determined by thePrussian blue method (Price and Butler 1977).
ROONEY ET AL.: DEVELOPMENT AND LOCATION IN SORGHUM 35
bleached grain appears a little lighter for somesamples than that of most sorghums with a testa.But the light colour may indicate incomplete re-moval of the thick, chalky pericarp over the testaand may not have anything to do with thicknessof the testa. These observations are preliminaryand further experiments must be made.
Grain Maturation Studies
A study to determine changes in thepolyphenols and kernel structure during develop-ment and maturation of the kernel was made onthree sorghum varieties. Florets were tagged andsamples of individual florets were taken at anthe-sis (bloom) and at 1, 2, 4, 6, 8, 10, 12, 16, 20, 24,32, 36, and 40 days subsequently for microscopyand chemical analysis. The varieties included:Tx09 (b1b1B2B2 ss rryyiizz PP QQ) a white,chalky nonyellow sorghum; Double DwarfFeterita (B1B1B2B2 ss rryyiizz PP QQ); Redlan xCombine Shallu (B1bB2b2 SS RRYyiiZz Pp Qq).
Enough grain was harvested at 8, 10, 16,20,24,28, 32, 36, and 40 days for tannin analyses by thePrussian blue method (Price and Butler 1977); thevanillin-HCI method applied on methanol ex-tracts (Price and Butler 1978); the modifiedvanillin-HCI method (Maxson et al. 1972); andan enzyme inhibition method (Blakely et al.1979). Data with and without blanks subtractedwere obtained for both vanillin methods and arepresented in Fig. 5 and 6.
The micrograms of catechin equivalents (CE)per seed are presented in Fig. 5, 6, and 7 for thevanillin and Prussian blue methods. Fig. 8 pre-sents the a-amylase inhibition data. TheGA615 grain had the maximum level of catechinequivalents at 16 days for all methods of chemicalanalysis; but the maximum for enzyme inhibitionunits per seed was 28 days after bloom. Grainfrom Double Dwarf Feterita (DDF) a kernelwith thick white pericarp and testa - containedabout half the catechin equivalents of GA6 15 at16 days when measured by the 1% HC1-methanolextraction procedure. However, when theextractant was only methanol, the catechin equi-valents were essentially zero at 16-20 days,whereas a significant increase in catechin equiva-lents occurred at 32 days. At this point, the CE forGA615 had decreased by about 50% of the valuereached at 16 days. Tx09, white without a testa,
.. I..--
STxOS
06 8 10 12 16 20 24 28 32 36 40
DAYS
Fig. 8. Varialion in a-amylase inhibition by waterextracts of grain from three sorghums from anthesis tomaturity.
had essentially no catechin equivalents by thevanillin methods. Some materials in Tx09 andDDF gave a slight response to the Prussian bluemethod (Fig. 7).
The cw-amylase inhibition test had a great dealof variability among laboratory replicates. It ap-peared that DDF had some increase in inhibitorypower during maturation with a peak at 28 daysfollowed by a sharp loss of inhibitory power uponmaturation. The high variation of this methodprevents conclusions about the changes in DDF.We do not consider the low levels of inhibition byTx09 to be significant. The differences shown inFig. 8 may not be repeatable. In previous experi-ments (Maxson et al. 1973) we found no inhibi-tion of amylase by mature grain from Tx09 andDDF.
These data represent the results for only oneseason and should be considered as preliminarydata. They are presented here to provide addi-tional information on the effects of grain matu-rity on polyphenol compounds in the grain.
S 'S
'S
\/ /
20
18
ASI'ISI II II iGA615
16 SS/ I
S14 SI
S
II
S I12
S II1 I
10
IU
8.,/ Il S/t
I I'SI'I I
6 /I ,._ /DDFI.
S .5S s
'
Related Documents











