© 2012 Zoological Society of Japan ZOOLOGICAL SCIENCE 29: 000–000 (2012) Ancient Colonization and Within-island Vicariance Revealed by Mitochondrial DNA Phylogeography of the Mountain Hare (Lepus timidus) in Hokkaido, Japan Gohta Kinoshita 1 * , Mitsuo Nunome 2 , Sang-Hoon Han 3 , Hirofumi Hirakawa 4 , and Hitoshi Suzuki 5 1 Laboratory of Ecology and Genetics, Graduate School of Environmental Science, Hokkaido University, Kita-ku, Sapporo 060-0810, Japan 2 Laboratory of Animal Genetics, Graduate School of Bioagricultural Siences, Nagoya University, Nagoya 464-8601, Japan 3 National Institute of Biological Resources, Environmental Research Complex, Incheon 404-708, Korea 4 Forestry and Forest Products Research Institute, Hitsujigaoka, Toyohira-ku, Sapporo 062-8516, Japan 5 Laboratory of Ecology and Genetics, Faculty of Environmental Earth Science, Hokkaido University, Kita-ku, Sapporo 060-0810, Japan We examined the phylogenetic status and history of the mountain hare Lepus timidus in and around Hokkaido using mitochondrial cytochrome b (cyt b) sequences from 158 samples from Hokkaido and 14 from Sakhalin, as well as four samples from the Korean hare, L. coreanus. The phylogenetic analysis of the cyt b sequences generated in this study and obtained from DNA databases showed the clear genetic specificity of the Hokkaido lineage as a clade. The Hokkaido lineage was esti- mated to have diverged from the other conspecific and L. coreanus lineages 0.46 and 0.30 million years ago (Mya), respectively. These results suggest that the common ancestor of the mitochon- drial lineage in Hokkaido and Korea inhabited Far East Asia before colonization by the present con- tinental lineages of L. timidus, including the Sakhalin population. We estimated the time of the most recent common ancestor of the Hokkaido population to be 0.17 Mya, and found two distinct hap- logroups within the island. One group had greater genetic diversity (mean number of pairwise dif- ferences: π = 0.0188 ± 0.0108) and appears to have expanded from the west to the entire island of Hokkaido. The other had lower genetic diversity (π = 0.0038 ± 0.0037) and its distribution was con- centrated in the east. These contrasting west/east trends indicate that the Hokkaido population was fragmented in the past, and then subsequently expanded. Our study suggests that Hokkaido was an important refugium for boreal species in the far eastern region, and allowed the formation of various population genetic structures within the island. Key words: island refugium, mitochondrial DNA, mountain hare, phylogeography, vicariance INTRODUCTION Glacial cycles during the Pleistocene caused dramatic changes in the geographic distribution and genetic variation of organisms (Hewitt, 2004; Bennett and Provan, 2008; Stewart et al., 2010). During the glacial age, large areas of North America and northern Europe were covered by ice sheets, forcing resident organisms to shift their distribution towards the south (Taberlet et al., 1998; Soltis et al., 2006; Bhagwat and Willis, 2008). Meanwhile, the area from east- ern Siberia to northwestern America remained largely ungla- ciated (Grosswald, 1980, 1998; Abbott and Brochmann, 2003; Svendsen et al., 2004). These areas played an impor- tant role in the phylogeographic history of boreal species. Fedorov et al. (2008) proposed that southeastern Siberia was important for the wood lemming (Myopus schisticolor) and other species as a refugial area. In addition, Hope et al. (2010) found a phylogeographic break for the Eurasian least shrew, Sorex minutissimus, in northeastern Asia. The area, comprised of Primorsky Krai, Sakhalin, and the Hokkaido Islands, is located at the southeastern edge of the distribu- tion for many boreal species (Dobson, 1994; Kawamura, 2007), and is considered to have served as a refugium dur- ing the Quaternary climate changes (Oshida et al., 2005; Sakka et al., 2010). However, the roles of these areas as glacial refugia and the details of the demographic dynamics of the boreal species are not sufficiently understood. Hokkaido Island is located off the eastern coast of north- * Corresponding author. Tel. : +81-11-706-2225; Fax : +81-11-706-2225; E-mail : [email protected] Supplemental material for this article is available online. doi:10.2108/zsj.29.000 0260.R1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

© 2012 Zoological Society of JapanZOOLOGICAL SCIENCE 29: 000–000 (2012)
Ancient Colonization and Within-island Vicariance Revealed by Mitochondrial DNA Phylogeography of the Mountain Hare
(Lepus timidus) in Hokkaido, Japan
Gohta Kinoshita1*, Mitsuo Nunome2, Sang-Hoon Han3,Hirofumi Hirakawa4, and Hitoshi Suzuki5
1Laboratory of Ecology and Genetics, Graduate School of Environmental Science, Hokkaido University,Kita-ku, Sapporo 060-0810, Japan
2Laboratory of Animal Genetics, Graduate School of Bioagricultural Siences, Nagoya University,Nagoya 464-8601, Japan
3National Institute of Biological Resources, Environmental Research Complex,Incheon 404-708, Korea
4Forestry and Forest Products Research Institute, Hitsujigaoka, Toyohira-ku,Sapporo 062-8516, Japan
5Laboratory of Ecology and Genetics, Faculty of Environmental Earth Science,Hokkaido University, Kita-ku, Sapporo 060-0810, Japan
We examined the phylogenetic status and history of the mountain hare Lepus timidus in and around Hokkaido using mitochondrial cytochrome b (cyt b) sequences from 158 samples from Hokkaido and 14 from Sakhalin, as well as four samples from the Korean hare, L. coreanus. The phylogenetic analysis of the cyt b sequences generated in this study and obtained from DNA databases showed the clear genetic specificity of the Hokkaido lineage as a clade. The Hokkaido lineage was esti-mated to have diverged from the other conspecific and L. coreanus lineages 0.46 and 0.30 million years ago (Mya), respectively. These results suggest that the common ancestor of the mitochon-drial lineage in Hokkaido and Korea inhabited Far East Asia before colonization by the present con-tinental lineages of L. timidus, including the Sakhalin population. We estimated the time of the most recent common ancestor of the Hokkaido population to be 0.17 Mya, and found two distinct hap-logroups within the island. One group had greater genetic diversity (mean number of pairwise dif-ferences: ππ = 0.0188 ± 0.0108) and appears to have expanded from the west to the entire island of Hokkaido. The other had lower genetic diversity (ππ = 0.0038 ± 0.0037) and its distribution was con-centrated in the east. These contrasting west/east trends indicate that the Hokkaido population was fragmented in the past, and then subsequently expanded. Our study suggests that Hokkaido was an important refugium for boreal species in the far eastern region, and allowed the formation of various population genetic structures within the island.
Key words: island refugium, mitochondrial DNA, mountain hare, phylogeography, vicariance
INTRODUCTION
Glacial cycles during the Pleistocene caused dramatic changes in the geographic distribution and genetic variation of organisms (Hewitt, 2004; Bennett and Provan, 2008; Stewart et al., 2010). During the glacial age, large areas of North America and northern Europe were covered by ice sheets, forcing resident organisms to shift their distribution towards the south (Taberlet et al., 1998; Soltis et al., 2006; Bhagwat and Willis, 2008). Meanwhile, the area from east-ern Siberia to northwestern America remained largely ungla-
ciated (Grosswald, 1980, 1998; Abbott and Brochmann, 2003; Svendsen et al., 2004). These areas played an impor-tant role in the phylogeographic history of boreal species. Fedorov et al. (2008) proposed that southeastern Siberia was important for the wood lemming (Myopus schisticolor) and other species as a refugial area. In addition, Hope et al. (2010) found a phylogeographic break for the Eurasian least shrew, Sorex minutissimus, in northeastern Asia. The area, comprised of Primorsky Krai, Sakhalin, and the Hokkaido Islands, is located at the southeastern edge of the distribu-tion for many boreal species (Dobson, 1994; Kawamura, 2007), and is considered to have served as a refugium dur-ing the Quaternary climate changes (Oshida et al., 2005; Sakka et al., 2010). However, the roles of these areas as glacial refugia and the details of the demographic dynamics of the boreal species are not sufficiently understood.
Hokkaido Island is located off the eastern coast of north-
* Corresponding author. Tel. : +81-11-706-2225;Fax : +81-11-706-2225;E-mail : [email protected]
Supplemental material for this article is available online.doi:10.2108/zsj.29.000
0260.R1
Editorial Office
ノート注釈
レタープレス様: Supplementary Fileが複数でもここの文言は単数形で問題無いでしょうか?

G. Kinoshita et al.2
eastern Asia, around which the Kuril Islands, Sakhalin, and other parts of the Japanese Archipelago are situated. Between Hokkaido and Honshu, the island located to the south of Hokkaido, lies the Tsugaru Strait, which is deep enough to have separated the islands during the last glaci-ation and is a biogeographic boundary, the Blakiston line (Ohshima, 1991; Masuda, 1999). In contrast, the straits between Hokkaido and Sakhalin or the southern Kuril Islands were comparatively shallow and were probably con-nected by land bridges during the major glacial periods (Ono, 1990; Ohshima, 1991). This allowed boreal species to migrate from the Eurasian continent to Hokkaido and the southern Kuril Islands via Sakhalin (Kawamura, 2007); con-sequently, the present fauna of Hokkaido is similar to that of northeastern Asia (Millien-Parra and Jaeger, 1999). Organ-isms inhabiting Hokkaido could have interacted intermit-tently with conspecific populations on the continent and adjacent islands. Hokkaido Island may therefore have been important for the historical demography and intra-specific evolution of mammals in northeastern Asia during the Pleistocene. Recent genetic studies have provided insights into the phylogeographic history of Hokkaido mammals, demonstrating species-specific patterns of population genetic structures with respect to historical migration from the continent, possibly correlated with behavioral and envi-ronmental factors such as dispersal ability and the existence of suitable habitat (brown bear, Ursus arctos, Matsuhashi et al., 2001; flying squirrel, Pteromys volans, Oshida et al., 2005; red fox, Vulpes vulpes, Inoue et al., 2007; sable, Martes zibellina, Sato et al., 2011). Based on ecological and morphological characteristics, many organisms are consid-ered endemic subspecies in Hokkaido, but the phylogenetic status and population genetic structure of these populations have not yet been surveyed sufficiently.
Lepus timidus is adapted to circumpolar climates and is fairly widespread on the tundra and in boreal forests in the Eurasian continent, with some isolated populations inhabit-ing Scotland, Ireland, the Alps, Sakhalin, Hokkaido, and the southern Kuril Islands. Phylogeographic studies using mito-chondrial DNA (mtDNA) or microsatellites have revealed a degree of genetic endemism for populations in Scotland and Ireland, while there were not significant correlations between geographic and genetic distances for continental popula-tions ranging from Europe to eastern Russia (Waltari and Cook, 2005; Hamill et al., 2006; Melo-Ferreira et al., 2007). By contrast, while mtDNA analyses by Yamada et al. (2002) showed that L. timidus in Hokkaido has unique haplotypes, others have reported phylogenetic affinity in mtDNA analy-ses between L. timidus, especially the lineage from Hokkaido, and the Korean L. coreanus distributed in the Korean Peninsula (Waltari and Cook, 2005; Wu et al., 2005). To better understand the population genetic structure and history of L. timidus in Hokkaido, further phylogeo-graphic analyses of eastern areas with widespread sampling from Hokkaido are required.
In this study, we conducted phylogeographic analyses for L. timidus in Hokkaido using cytochrome b (cyt b) gene sequences, together with sequences from other populations and other Lepus species from the databases. We collected samples throughout Hokkaido and from Sakhalin, and also used samples of Lepus coreanus. Secondly, we surveyed
the population genetic structure within Hokkaido to infer the demographic history after migration from the continent. We discuss how Hokkaido Island contributed to the phylogeo-graphic history of boreal species in northeastern Asia.
MATERIALS AND METHODS
Sampling and DNA extractionOne tissue sample and 157 fecal samples were collected at 158
locations throughout Hokkaido between March 2009 and January 2011. To reduce the risk of multiple samplings of any given individ-ual, only one fecal sample derived from one individual was collected at each location, and sampling locations were separated by at least 2 km. In addition, 11 fecal samples from northern Sakhalin and three tissue samples from central Sakhalin, as well as four tissue samples of L. coreanus from South Korea, were analyzed. DNA was extracted by the standard phenol-chloroform extraction method (Sambrook et al., 2001).
Sequencing analysisThe entire cyt b gene region (1,140 bp) was sequenced in all
samples. The polymerase chain reaction (PCR) was performed for a final volume of 20 μl, and consisted of 1 μl template DNA, 2.5 mM MgCl2, 0.2 mM dNTPs, 0.05 μM of each primer (L14724 and H15915; Irwin et al., 1991), 1 × Taq polymerase buffer, and 0.5 units of AmpliTaq Gold DNA polymerase (Applied Biosystems, Foster City, CA). The first round of PCR was performed to amplify the entire cyt b gene with the following thermal cycling profile: initial denaturation at 96°C for 10 min followed by 35 cycles at 95°C for 30 s, 51°C for 30 s, and 72°C for 30 s. The first PCR products were used as a template in the second PCR to amplify half of the upper and lower segments with two primer sets, L14724 and Lbcytb644R, and Lbcytb514F and H15915, respectively (Lbcytb644R and Lbcytb514F; Nunome et al., 2010). The conditions of the second round of PCR were the same as those of the first PCR. The prod-ucts of the second PCR were sequenced using an ABI 3130 auto-mated sequencer (Applied Biosystems) with a BigDye Terminator cycle sequencing kit (Applied Biosystems), following the manufac-turer’s instructions and using the same primers for the second PCR.The sequences were aligned by eye using ProSeq ver. 2.91 (Filatov, 2001).
Phylogenetic analysisIn addition to the cyt b dataset, sequences of L. timidus from
across a wide distribution (DQ882870–DQ882898, DQ882900–DQ222959, AY745103–AY745109), five other Lepus species (L.arcticus: AF010153; L. othus: AF010154; L. townsendii; AY292729; L. americanus: AY292733; L. brachyurus: AB514337, AB514338, AB514242) and Oryctolagus cuniculus (AY292717) were obtained from international databases (DDBJ/ EMBL/GenBank). Partial cyt bsequences (587 bp) were subjected to phylogenetic analysis using the neighbor-joining method (NJ) and Bayesian inference (BI), and pairwise distances were estimated among populations grouped as described in Wu et al. (2005) and Melo-Ferreira et al. (2007); see Fig. 1. NJ tree construction and the calculation of pairwise distances were conducted with 1,000 bootstrap replications using the program MEGA version 4.0 (Tamura et al., 2007). BI analysis was imple-mented for phylogenetic tree construction and divergence time esti-mation by using BEAST version 1.4.8 (Drummond and Rambaut, 2007). The best-fit model of evolution was selected based on Akaike’s Information Criterion (AIC; Akaike, 1974) by using the program Modeltest version 3.7 (Posada and Crandall, 1998). The dataset was analyzed in a Bayesian uncorrelated lognormal relaxed-clock model and sequences were partitioned according to 1st + 2nd and 3rd codon positions, using a constant population size coalescent as the tree prior. To estimate divergence time, we used three calibration points reported in a previous study (Matthee et al.,

Phylogeography of Lepus timidus in Hokkaido 3
2004); however, these need to be interpreted with caution. The divergence times inferred on the basis of time constraints derived from molecular dating would be over- or underestimates. The fossil records of the genus Lepus are limited and it is difficult to distin-guish Lepus species based on the paleontological record (Kurtén, 1968). The first calibration point was the divergence between Oryctolagus and Lepus, 11.8 Mya. The second was the time of the most recent common ancestors (TMR-CAs) of Lepus, 5.16 Mya. The last was the split between L. timidus and L.townsendii, 1.13 Mya. Time constraints were assigned a normal prior distribution with the 95% confidence interval of the estimates (Matthee et al., 2004). Bayesian searches were conducted using the Markov Chain Monte Carlo (MCMC) method for 10,000,000 gen-erations. Tracer V1.4.1. (Rambaut and Drummond, 2007) was used to assess convergence of MCMC chains and the effective sample size (ESS) values exceeding 200 for all parameters.
Population genetic struc-ture analysis
To study the demo-graphic history, the popula-tion genetic structure in Hokkaido was investigated using complete cyt bsequences (1,140 bp). We constructed median-join-ing networks for all of the Hokkaido samples using
Fig. 1. (A) Population grouping of L.timidus samples from the databases. The species’ ranges are indicated in gray, fol-lowing Flux and Angermann (1990). (B)Sampling localities in Sakhalin and Hokkaido are shown as dots.
Fig. 2. Phylogenetic tree of L. timidus and other species constructed using the neighbor-joining (NJ) method with partial cyt b sequences (587 bp). Haplotypes 1–15 and those of L. coreanus were original samples (see Supplementary Table S1 online), and haplotypes 16–62 and those of other Lepus species were obtained from the databases. Statistical support is indicated by NJ bootstrap/Bayesian posterior probabilities. Only bootstrap values higher than 50 are shown. TMRCAs estimated by Bayesian analysis are shown for each major calibra-tion point.
Editorial Office
取り消し線
Editorial Office
挿入テキスト
hares
Editorial Office
ノート注釈
40字を超えているので、システムに登録されていたRunning headを採用してよろしいでしょうか?
GKinoshita
ノート注釈
了解しました。Running headでよろしくお願いいたします。
GKinoshita
ノート注釈
Fig.1. の中で文字が小さい(A)の“Korean Peninsula”と“Japanese Archipelago”、(B)の“Kuril Island”と“Tsugaru Strait”を一回りフォントを大きくしてもらうことはできますでしょうか? もしくは、本体が10ページになってしまうかと思いますが、Fig.1と2をもう少し拡大してもらうことは可能でしょうか?

G. Kinoshita et al.4
the program Network v 4.6 (Bandelt et al., 1999).Estimations of genetic diversity (mean number of pairwise differ-
ences: π), the mismatch distribution analysis (Li, 1977; Harpending, 1994; Rogers, 1995), and Tajima’s neutrality test (Tajima’s D, Tajima, 1989) were conducted using the ARLEQUIN software. The mismatch distribution for populations at demographic equilibrium has a multi-modal pattern with a smooth wave shape, whereas populations following a recent demographic expansion form a unimodal pattern with a single peak. The good-ness-of-fit of the observed distribution to the expected distribution under the sudden-expansion model (Rogers, 1995) was tested by computing the sum of squares deviation and the raggedness index. The significance of values in the neutrality test and the mismatch analy-sis was tested using a parametric bootstrap approach (1,000 replicates). Moreover, spatial patterns of genetic diversity across each distribution area of two haplo-groups were visualized using the genetic landscape shape interpolation procedure in Alleles in Space (AIS, Miller, 2005). This procedure was initiated by generat-ing a Delaunay triangulation-based connectivity net-work of sampling locations (mean individuals), and then genetic distances calculated to decrease inversely with increasing geographic distances were assigned on grids of the network. Parts of the distribution area with higher genetic diversity between individuals were repre-sented in a darker color in this analysis. We also esti-mated pairwise fixation index (FST) values among sub-populations for a particular haplogroup in ARLEQUIN and constructed an NJ tree using these values in MEGA.
RESULTS
Sequence analysis and variation in the moun-tain hare in Hokkaido and Sakhalin
The complete 1,140-bp cyt b sequences were obtained from 158 samples from Hokkaido and 10 from Sakhalin. No deletions or insertions were detected. Sequences obtained from feces were not divergent from those obtained from tis-sue, and did not show any multiple amplifications of different copies or inconsistent positions in agarose gel electrophoresis. In addition, the base composition of the sequences (A 35.3%, C 36.3%, G 3.2%, T 25.3%) was similar to that of previously reported cyt b sequences of L. timidus(A 38.5%, C 32.3%, G 2.7% and T 26.5%; Melo-Ferreira et al., 2007), proving that the sequences were of mitochondrial origin rather than a nuclear copy. Sampling localities and population group-ings are shown in Fig. 1 and Supplementary Table S1 online.
In total, 26 haplotypes of L. timidus from Hokkaido and four from Sakhalin, as well as haplotypes from four South Korean L. coreanus, were detected in the complete cyt b region (Sup-plementary Table S1 online). These haplotype sequences were deposited in the database (Sup-plementary Table S2 online). In the analysis to examine the phylogenetic status of L. timidus in Hokkaido, the following numbers of haplotypes were used: 13 from Hokkaido, 2 from Sakhalin, and 3 from L. coreanus, detected in the partial cyt
b 587-bp region. The complete cyt b sequences of L. timidusin Hokkaido were used to examine the population genetic structure on the island.
Table 1. Pairwise genetic distances among mountain hare populations and Korean hare using partial cyt b sequences (587 bp).
Hk Sk ES SES EE Al Ir Sc NE L. coreanus
Hk 0.0013
Sk 0.0139 0.0002
ES 0.0146 0.0050 0.0046
SES 0.0195 0.0106 0.0110 0.0164
EE 0.0145 0.0045 0.0055 0.0106 0.0046
Al 0.0163 0.0062 0.0064 0.0121 0.0069 0.0060
Ir 0.0154 0.0054 0.0065 0.0121 0.0060 0.0072 0.0000
Sc 0.0140 0.0040 0.0051 0.0107 0.0046 0.0063 0.0046 0.0003
NE 0.0148 0.0047 0.0055 0.0112 0.0051 0.0068 0.0062 0.0049 0.0052
L. coreanus 0.0097 0.0101 0.0113 0.0161 0.0108 0.0125 0.0116 0.0102 0.0110 0.0020
Fig. 3. (A) Median joining network using complete cyt b sequences (1,140 bp) of all Hokkaido samples. The size of the circles is proportional to the number of sam-ples. The compositions of sample localities after subpopulation grouping (see Fig. 3B) are also reflected in each haplotype. Haplotype names are shown beside the circles. (B) Sample localities of two haplogroups and a subpopulation grouping of them. Darker colors indicate areas of higher altitude.
Phylogeography of Lepus timidus in Hokkaido 5
Phylogenetic relationships and divergence timesThe best-fit model of nucleotide substitution was the
HKY + G model, and the general topology of the NJ tree was identical to that of the BI analysis. In both analyses, the hap62 (AY745108) haplotype from southeastern Siberia was the earliest offshoot among the L. timidus haplotypes (Fig. 2). All of the Hokkaido haplotypes were resolved as monophyletic and distinct from others with high nodal sup-port (94% for NJ and 1.00 for BI). In addition, the haplotypes of L. coreanus formed a clade with the Hokkaido lineage. However, there were obscurities of substructures in the major group, namely, a paraphyletic group that did not include the hap62, Hokkaido, or Korean lineages. This group showed almost no geographic specificity, with low support values in both analyses, although it contained almost all of the L. timidus haplotypes from northern Europe to eastern Russia (as shown by Melo-Ferreira et al., 2007), as well as the other Lepus species, L. arcticus and L. othus. Two haplotypes detected from Sakhalin were very close to each other, and were included in the major group. Pairwise distances among populations were estimated (Table 1). Except for the southeastern population that included a genetically remote haplotype (hap62, Fig. 2), the Hokkaido population was genetically distant from the others (0.0139–0.0163), whereas continental and other insular populations showed relatively close relationships to each other (0.0040–0.0072).
The TMRCA, analyzed using BEAST for each node of the tree, is shown in Fig. 2. The TMRCA esti-mated for all mountain hare species was 0.86 Mya (95% HPD 0.53–1.18 Mya), which is consistent with the earliest fossil record of this spe-cies in the early Middle Pleistocene (Kurtén, 1968). However, Liu et al. (2011) pointed out that the earliest offshoot hap62 might be an intro-gressed haplotype from L. capensisto L. timidus, and if so, the TMRCA obtained for L. timidus could be over-estimated. The split of the Hokkaido/Korean lineage from other L. timiduslineages was estimated to be 0.46 Mya (95% HPD 0.29–0.65 Mya), nearly simultaneous with the diversifi-cation of the major group 0.53 Mya (95% HPD 0.33–0.75 Mya). The Hokkaido lineage diverged from the L. coreanus lineage 0.30 Mya (95% HPD 0.16–0.46 Mya).
Population genetic structure in Hokkaido
The TMRCA for the Hokkaido lin-eage estimated by analysis using partial cyt b sequences (587 bp) was estimated to be 0.17 Mya (95% HPD 0.08–0.27 Mya: Fig. 2). The NJ net-
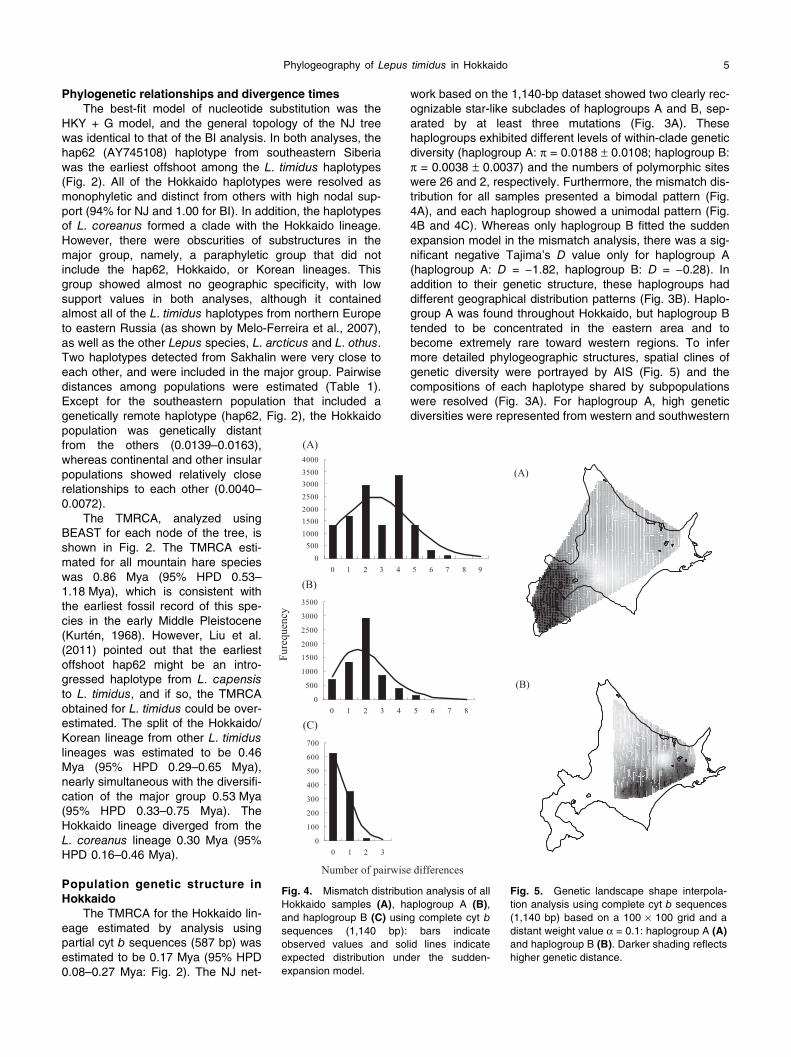
work based on the 1,140-bp dataset showed two clearly rec-ognizable star-like subclades of haplogroups A and B, sep-arated by at least three mutations (Fig. 3A). These haplogroups exhibited different levels of within-clade genetic diversity (haplogroup A: π = 0.0188 ± 0.0108; haplogroup B: π = 0.0038 ± 0.0037) and the numbers of polymorphic sites were 26 and 2, respectively. Furthermore, the mismatch dis-tribution for all samples presented a bimodal pattern (Fig. 4A), and each haplogroup showed a unimodal pattern (Fig. 4B and 4C). Whereas only haplogroup B fitted the sudden expansion model in the mismatch analysis, there was a sig-nificant negative Tajima’s D value only for haplogroup A (haplogroup A: D = −1.82, haplogroup B: D = −0.28). In addition to their genetic structure, these haplogroups had different geographical distribution patterns (Fig. 3B). Haplo-group A was found throughout Hokkaido, but haplogroup B tended to be concentrated in the eastern area and to become extremely rare toward western regions. To infer more detailed phylogeographic structures, spatial clines of genetic diversity were portrayed by AIS (Fig. 5) and the compositions of each haplotype shared by subpopulations were resolved (Fig. 3A). For haplogroup A, high genetic diversities were represented from western and southwestern
Fig. 4. Mismatch distribution analysis of all Hokkaido samples (A), haplogroup A (B), and haplogroup B (C) using complete cyt bsequences (1,140 bp): bars indicate observed values and solid lines indicate expected distribution under the sudden-expansion model.
Fig. 5. Genetic landscape shape interpola-tion analysis using complete cyt b sequences (1,140 bp) based on a 100 × 100 grid and a distant weight value α = 0.1: haplogroup A (A)and haplogroup B (B). Darker shading reflects higher genetic distance.

G. Kinoshita et al.6
Hokkaido, and the central haplotype (1-a, Fig. 3A) was mainly composed of specimens from subpopulations 5 and 6 in the western area. In contrast, haplogroup B showed somewhat high genetic diversity in the southern region of its distribution. Individuals belonging to two tip haplotypes of
the haplogroup B tree were seen only in subpopulations 1 and 2. For haplogroup A, we estimated pairwise FST values among subpopulations and used them to construct the phy-logenetic tree (Table 2 and Fig. 6A). The phylogenetic tree formed three clusters (cluster I-III) containing one or two peripheral subpopulations. Cluster I contained subpopula-tions 3 and 7, and the most frequent haplotype occurring in both subpopulations (haplotype 1-d) was observed only in the area along the Japan Sea coast (Fig. 6B). Cluster II comprised subpopulations 1 and 2, and the most frequent haplotype in each subpopulation was haplotype 1-i and 6, respectively, in haplogroup A. These haplotypes were observed mainly in the west and east, but were rare in the northern and southwestern areas of the island (Fig. 6C). Cluster III was represented by subpopulation 5, and the most frequent haplotype in the subpopulation was haplotype 1-a, which was the most central haplotype in haplogroup A (Fig. 5A) and confined to the west of the island (Fig. 6D).
DISCUSSION
Phylogenetic status of the Hokkaido lineage
Our study suggests that the lineage of L. timidus in Hokkaido is distinct from that of other populations. Melo-Ferreira et al. (2007) found less genetic diversity in L.timidus, even between popu-lations from both ends of the Eurasian continent, and they suggested that this trend may have been caused by a recent expansion from local refugia and subsequent genomic mixing. By contrast, isolated populations in Ireland (hap43) and Scotland (hap20 and 21) demonstrated some-what different genetic spe-cialties (Hamill et al., 2006; Melo-Ferreira et al., 2007), but were much less different from the continental lineage than those of the Hokkaido population (Fig. 2 and Table 1). We added Sakhalin sam-ples and the two haplotypes (hap14 and 15) that were detected were located within the major group (Fig. 2). These results suggest that Hokkaido preserved old lin-eages that diverged earlier than other island lineages.
Phylogenetic relationship between the Hokkaido lin-eage and L. coreanus
We found a sister rela-tionship between L. coreanus
Table 2. Pairwise comparisons of FST using complete cyt bsequences (1,140 bp) between subpopulations of haplogroup A. * P < 0.05.
1 2 3 4 5 6
12 0.010753 0.18607* 0.17982*4 0.04959* 0.02477 0.10954*5 0.9415* 0.08005 0.23821* 0.06271*6 0.24030 0.03628 0.09154* 0.02187 0.06271*7 0.12228* 0.10350 0.01523 0.05420 0.17227* 0.03140
* P < 0.05.
Fig. 6. (A) The subpopulation relationships of haplogroup A are portrayed by constructing a phyloge-netic tree using the NJ method and the Fst values shown in Table 2. The clusters comprised of one or two peripheral subpopulations in the phylogenetic tree are shown with dotted lines. The geographic dis-tributions of the most frequent haplotypes (in haplogroup A) in each peripheral subpopulation are shown with clusters; haplotype 1-d for both subpopulations 3 and 7 in cluster I (B); haplotypes 1-i and 6 in sub-populations 1 and 2, respectively, in cluster II (C); and haplotype 1-a in subpopulation 5 in cluster III (D).
GKinoshita
ノート注釈
FstのstをST(下付き)に直してください

Phylogeography of Lepus timidus in Hokkaido 7
and lineages from Hokkaido (Fig. 2). Previous mtDNA and nuclear DNA analyses suggested that L. coreanus was dis-tinct from L. timidus (Koh et al., 2001; Koh and Zhang, 2010). However, no samples from Hokkaido were included in their studies. Contrastingly, an mtDNA analysis by Wu et al. (2005) demonstrated a close relationship between L.timidus in Hokkaido and L. coreanus using one haplotype from each lineage, and Waltari and Cook (2005) also reported a genetic affinity between L. coreanus and L. timidus. Our data reveal that L. coreanus was embedded within the L. timidus group as a lineage closely related to the Hokkaido lineages. However, introgression from L. timidus to some other hare populations has also been reported in various regions in Europe and northern Asia (Alves et al., 2008; Liu et al., 2011). For example, allopatric hares on the Iberian Peninsula harbor high frequencies of mtDNA (in L.castroviejoi, L. europaeus, and L. granatensis) and the telo-metric region of the X chromosome (in L. granatensis) from L. timidus by ancient introgression as a consequence of dis-tribution replacement after deglaciation (Alves et al., 2003, 2008; Melo-Ferreira et al., 2005, 2011). Considering these findings and taxonomic status based on the morphological characters (Flux and Angermann, 1990), the mitochondrial lin-eage of L. coreanus is likely to be a result of introgression from the ancestor of the Hokkaido lineage. However, a more synthetic analysis with many L. coreanus samples or using nuclear DNA is needed to define the classification of the Hokkaido lineage and L. coreanus.
Origin and migration history of the Hokkaido lineageThere are two possible routes of mammalian migration
from the continent to Hokkaido: the northern route from eastern Siberia via Sakhalin, and the southern route from the Korean Peninsula via Honshu. On the islands south of Honshu, the indigenous Japanese hare, L. brachyurus, is widely distributed; however, there are no records of either extant or extinct L. timidus. Phylogeographic analysis of L.brachyurus using mtDNA also showed no imprint of histori-cal contact with L. timidus (Nunome et al., 2010). Therefore, the Hokkaido lineage must have migrated using the northern route, like other boreal species, such as P. volans and Martes zibellina (Oshida et al., 2005; Kawamura, 2007; Sato et al., 2011). Moreover, our data suggest that the migration of the Hokkaido lineage may have occurred only once before the last glacial, because they formed one monophyl-etic group with the estimated tMRCA of 0.17 Mya in the mtDNA analysis.
The results of our phylogenetic analysis (Fig. 2 and Table 1) indicate that the ancestor of L. timidus in Hokkaido may have inhabited Far East Asia and may have contacted L. coreanus in the middle Pleistocene, and that the offspring remained on Hokkaido Island. While climatic oscillations have diluted the phylogeographic structure among continen-tal populations of L. timidus (Melo-Ferreira et al., 2007) with recent re-entrance to Sakhalin, the Hokkaido population seems not to have been affected by subsequent migration from the continent via Sakhalin.
Population genetic structure and lineage separation in Hokkaido
Our results indicate that there are two distinct mitochon-
drial haplogroups within the Hokkaido lineage that show dif-ferent patterns of distribution and genetic diversity (Fig. 3A). Haplogroup A is found across the entire island and has greater genetic diversity than the other, particularly in the southwest (Figs. 3B and 5A). The most central haplotype (1-a) of this group is normally found in the west (Fig. 3A, B). Conversely, the genetic diversity within haplogroup B is much lower, and its distribution is concentrated in the east, where tip haplotypes are also mainly found (Fig. 3A and 3B). The west/east phylogeographic pattern seems to be a result of past vicariance on Hokkaido Island, given the distinctive-ness of the Hokkaido lineage from other lineages (Fig. 2) and the migration route from the north. The TMRCA of the Hokkaido population was estimated to be 0.17 Mya (Fig. 2); therefore, the vicariance occurred after that.
The geographic patterns of distribution and genetic diversity in Hokkaido suggest that haplogroup A and B expanded from the west and east sides of the island, respectively. In addition, the relationship between subpopu-lations in haplogroup A formed a triple-branched structure for clusters I–III, and the characteristic distribution was shown for the most frequent haplotype in each cluster (Fig. 6). These results suggest that the predicted expansion routes of haplogroup A can be separated into two streams: the Japan Sea coastal line (subpopulations 3, 4, 6, and 7) and the Pacific Ocean coastal line (subpopulations 1, 2, and 4–6) derived from a source area from the southwest to the west of the island, where high genetic diversity and the con-centration of the most central haplotype were observed. For some other mammals in Hokkaido, such as U. arctos(Matsuhashi et al., 2001) and V. vulpes (Inoue et al., 2007), phylogeographic analyses using mtDNA revealed that they were composed of two or three distinctive haplogroups, which were thought to be the result of multiple migrations from the continent. However, there are no previous reports of mammals that suggest ancient population fragmentation on Hokkaido. An alpine belt runs north–south in central Hokkaido (Fig. 3B), and glaciers appeared at some higher elevations during the last glacial age (Ono et al., 2005). At that time, taiga forests and tundra grasslands developed in Hokkaido and on the southern Kuril Islands, which were accessible via land bridges (Ono, 1990; Takahara et al., 2010; Igarashi and Zharoh, 2011). Such topographical struc-tures and climatic changes may have caused the past vicar-iance on the island and the present distribution pattern of the two haplogroups. In addition, the climate in southwest-ern Hokkaido is generally milder than that in the east (Nakamura et al., 2007). This may have allowed haplogroup A in the southwest to retain a large population and high genetic diversity during the glacial age compared to haplo-group B in the east.
The role of Hokkaido for boreal species in the Far EastHokkaido Island and the surrounding area are located at
the southern edge of the distribution of many boreal species in northeastern Asia, and have recently attracted attention as refugia for these species in glacial ages (Fedorov et al., 2008; Hope et al., 2010). The Hokkaido population and the mitochondrial lineage of L. coreanus seem to be remnants of the ancient demographic expansion of L. timidus in this region. Previous studies based on mtDNA analysis reported

G. Kinoshita et al.8
that mammals in Hokkaido showed species-specific phylo-geographic patterns. In addition, the timing of their migra-tions to Hokkaido has been proposed based on molecular data or geographic evidence. Some have close genetic rela-tionships with continental populations, which are probably the result of the last glacial migration via Sakhalin (Martes zibellina, Sato et al., 2011; Sorex unguiculatus, Ohdachi et al., 2001; Apodemus peninsulae, Serizawa et al., 2002). Meanwhile, other species with unique lineages in Hokkaido, such as L. timidus, seem to have migrated to Hokkaido before the last glacial age (S. caecutiens, Ohdachi et al., 2001; P. volans, Oshida et al., 2005). In addition, there are species that maintain multiple lineages, having migrated at various times (U. arctos, Matsuhashi et al., 2001; S.gracillimus, Ohdachi et al., 2001; V. vulpes, Inoue et al., 2007). Consequently, Hokkaido allows boreal species to maintain unique lineages as a result of the geographic his-tory in which the island was connected intermittently to the continent by the northern land bridge, and was not covered by massive ice sheets during the major glacial age (Ono, 1990; Ohshima, 1991).
We further revealed that the Hokkaido lineage of L.timidus has a characteristic population genetic structure, which indicates past vicariance on the island. Our study demonstrates the significance of the eastern and western regions of Hokkaido for delineating the population genetic structure of L. timidus. Several previous studies examined the phylogenetic relationships of small mammal populations on Hokkaido and the southern Kuril Islands (Iwasa et al., 2000; Ohdachi et al., 2001). However, they examined only a few samples from the eastern region of Hokkaido and the southern Kuril Islands, and the role of the regions as a sub-population source area was not mentioned. To assess the phylogeographic history of the island’s organisms, a more detailed analysis including more samples from the southern Kuril Islands is required.
In conclusion, our study suggests that Hokkaido played a significant role as an isolated refuge for L. timidus and other boreal species. Furthermore, we stress that the finding of ancient within-island vicariance in L. timidus may offer new insights into the demographic history of many organ-isms in Hokkaido.
ACKNOWLEDGMENTS
We extend sincere gratitude to Alexey P. Kryukov for providing valuable comments on an earlier version of the manuscript. This work was supported by a Grant-in-Aid for Scientific Research (B) to HS (19405010) from Japan Society for the Promotion of Science (JSPS). This study was supported in part by a Grant-in-Aid for Fellows of the Japan Society for the Promotion of Science (JSPS) to NM (21002319) from the Ministry of Education, Culture, Sports, Science and Technology (MEXT). We thank Fadeev NA, Higuchi K, Hirabayashi T, Hemmi H, and Vesta Z for kind help in supplying the valuable samples used in this study.
REFFERENCES
Abbott RJ, Brochmann C (2003) History and evolution of the Arctic flora: in the footsteps of Eric Hultén. Mol Ecol 12: 299–313
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Contr 19: 716–723
Alves PC, Ferrand N, Suchentrunk F, Harris DJ (2003) Ancient intro-gression of Lepus timidus mtDNA into L. granatensis and L.
europaeus in the Iberian Peninsula. Mol Phylogenet Evol 27: 70–80
Alves PC, Melo-Ferreira J, Freitas H, Boursot P (2008) The ubiqui-tous mountain hare mitochondria: multiple introgressive hybrid-ization in hares, genus Lepus. Philos Trans R Soc B-Biol 363: 2831–2839
Bandelt HJ, Forster P, Rohl A (1999) Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol 16: 37–48
Bennett KD, Provan J (2008) What do we mean by ‘refugia’? Qua-ternary Sci Rev 27: 2449–2455
Bhagwat SA, Willis KJ (2008) Species persistence in northerly gla-cial refugia of Europe: a matter of chance or biogeographical traits? J Biogeogr 35: 464–482
Dobson M (1994) Patterns of distribution in Japanese land mam-mals. Phylogenetic Relationships of Crocidura. Mamm Rev 24: 91–111
Drummond AJ, Rambaut A (2007) BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol Biol 7: 1–8
Fedorov VB, Goropashnaya AV, Boeskorov GG, Cook JA (2008) Comparative phylogeography and demographic history of the wood lemming (Myopus schisticolor): implications for late Qua-ternary history of the taiga species in Eurasia. Mol Ecol 17: 598–610
Filatov D (2001) Processor of sequences manual. University of Bir-mingham. http://www.biosciences.bham.ac.uk/filatov/proseq.html
Flux JEC, Angermann R (1990) The hares and jackrabbits. In “Rab-bits, Hares and Pikas: Status Survey and Conservation Action Plan” IUCN, Gland, Switzerland
Grosswald MG (1980) Late weichselian ice sheet of Northern Eurasia. Quaternary Res 13: 1–32
Grosswald MG (1998) Late-Weichselian ice sheets in Arctic and Pacific Siberia. Quat Int 45/46: 3–18
Hamill RM, Doyle D, Duke EJ (2006) Spatial patterns of genetic diversity across European subspecies of the mountain hare, Lepus timidus L. Heredity 97: 355–365
Harpending RC (1994) Signature of ancient population growth in a low-resolution mitochondrial DNA mismatch distribution. Hum Biol 66: 591–600
Hewitt GM (2004) Genetic consequences of climatic oscillations in the Quaternary. Philos Trans R Soc B-Biol Sci 359: 183–195
Hope AG, Waltari E, Dokuchaev NE, Abramov S, Dupal T, Tsvetkova A, et al. (2010) High-latitude diversification within Eurasian least shrews and Alaska tiny shrews (Soricidae). J Mammal 91: 1041–1057
Igarashi Y, Zharoh EA (2011) Climate and vegetation change during the late Pleistocene and early Holocene in Sakhalin and Hokkaido, northeast Asia. Quat Int 237: 24–31
Inoue T, Nonaka N, Mizuno A, Morishima Y, Sato H, Katakura K, Okul Y (2007) Mitochondrial DNA phylogeography of the red fox (Vulpes vulpes) in northern Japan. Zool Sci 24: 1178–1186
Irwin DM, Kocher TD, Wilson AC (1991) Evolution of the cytochrome b gene of mammals. J Mol Evol 32: 128–144
Iwasa MA, Utsumi Y, Nakata K, Kartavtseva IV, Nevedomskaya IA, Kondoh N, Suzuki H (2000) Geographic patterns of cytochrome band Sry gene lineages in the gray red backed vole Clethrionomys rufocanus from Far East Asia including Sakhalin and Hokkaido.Zool Sci 17: 477–484
Kawamura Y (2007) Last glacial and Holocene land mammals of the Japanese islands: their fauna, extinction and immigration. The Quaternary Research 46: 171–177
Koh HS, Zhang KH (2010) Genetic Distinctness of the Korean Hare, Lepus coreanus (Mammalia, Lagomorpha), Revealed by Nuclear Thyroglobulin Gene and mtDNA Control Region Sequences. Biochem Genet 48: 706–710
Koh HS, Chun TY, Yoo HS, Zhang Y-P, Wang J, Zhang M, Wu C-H (2001) Mitochondrial cytochrome b gene sequence diversity in the Korean hare, Lepus coreanus Thomas (Mammalia,

Phylogeography of Lepus timidus in Hokkaido 9
Lagomorpha). Biochem Genet 39: 417–429Kurtén B (1968) Pleistocene Mammals of Europe. Weidenfeld and
Nicholson, LondonLi WH (1977) Distribution of nucleotide differences between two ran-
domly chosen cistrons in a finite population. Genetics 85: 331–337
Liu J, Yu L, Arnold ML, Wu CH, Wu SF, Lu X, Zhang YP (2011) Reticulate evolution: frequent introgressive hybridization among chinese hares (genus Lepus) revealed by analyses of multiple mitochondrial and nuclear DNA loci. BMC Evol Biol 11: 223
Masuda R (1999) Blakiston’s line and genetic investigation on bio-geography of mammals in Japan. Mammal Sci 39: 323–328 (in Japanese)
Matsuhashi T, Masuda R, Mano T, Murata K, Aiurzaniin A (2001) Phylogenetic relationships among worldwide populations of the brown bear Ursus arctos. Zool Sci 18: 1137–1143
Matthee CA, van Vuuren BJ, Bell D, Robinson TJ (2004) A molecu-lar supermatrix of the rabbits and hares (Leporidae) allows for the identification of five intercontinental exchanges during the Miocene. Syst Biol 53: 433–447
Melo-Ferreira J, Boursot P, Suchentrunk F, Ferrand N, Alves PC (2005) Invasion from the cold past: extensive introgression of mountain hare (Lepus timidus) mitochondrial DNA into three other hare species in northern Iberia. Mol Ecol 14: 2459–2464
Melo-Ferreira J, Boursot P, Randi E, Kryukov A, Suchentrunk F, Ferrand N, Alves PC (2007) The rise and fall of the mountain hare (Lepus timidus) during Pleistocene glaciations: expansion and retreat with hybridization in the Iberian Peninsula. Mol Ecol16: 605–618
Melo-Ferreira J, Alves PC, Rocha J, Ferrand N, Boursot P (2011) Interspecific X-chromosome and mitochondrial DNA introgres-sion in the Iberian hare: selection or allele surfing? Evolution 65-7: 1956–1968
Miller MP (2005) Alleles In Space (AIS): Computer software for the joint analysis of interindividual spatial and genetic information. J Hered 96: 722–724
Millien-Parra V, Jaeger JJ (1999) Island biogeography of the Japanese terrestrial mammal assemblages: an example of rel-ict fauna. J Biogeogr 26: 959–972
Nakamura Y, Krestov PV, Omelko AM (2007) Bioclimate and zonal vegetation in Northeast Asia: First approximation to an inte-grated study. Phytocoenologia 37: 443–470
Nunome M, Torii H, Matsuki R, Kinoshita G, Suzuki H (2010) The influence of Pleistocene refugia on the evolutionary history of the Japanese hare, Lepus brachyurus. Zool Sci 27: 746–754
Ohdachi S, Dokuchaev NE, Hasegawa M, Masuda R (2001) Intraspecific phylogeny and geographical variation of six spe-cies of northeastern Asiatic Sorex shrews based on the mito-chondrial cytochrome b sequences. Mol Ecol 10: 2199–2213
Ohshima (1991) The late-Quaternary sea-level change of the Japanese Islands. Geogr 100: 967–975
Ono Y (1990) The northern landbridge of Japan. Quat Res 29: 183–192
Ono Y, Aoki T, Hasegawa H, Dali L (2005) Mountain glaciation in Japan and Taiwan at the global Last Glacial Maximum. Quat Int 138/139: 79–92
Oshida T, Abramov A, Yanagawa H, Masuda R (2005) Phylogeog-raphy of the Russian flying squirrel (Pteromys volans): implica-tion of refugia theory in arboreal small mammal of Eurasia. Mol
Ecol 101: 219–224Posada D, Crandall KA (1998) MODELTEST: testing the model of
DNA substitution. Bioinformatics 14: 817–818Rambaut A, Drummond AJ (2007) Tracer v1.4. http://
beast.bio.ed.ac.uk/TracerRogers AR (1995) Genetic evidence for a Pleistocene population
explosion. Evolution 49: 608–615Sakka H, Quere JP, Kartavseva I, Pavlenko M, Chelomina G,
Atorkin D, et al. (2010) Comparative phylogeography of four Apodemus Speciens (Mammalia: Rodentia) in the Asian Far East: evidence of Quaternary climatic in thir genetic structure. Biol J Linn Soc 100: 797–821
Sambrook J, Russell DW (2001) Molecular Cloning: A Laboratory Manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York
Sato JJ, Hosoda T, Kryukov AP, Kartavtseva IV, Suzuki H (2011) Genetic diversity of the sable (Martes zibellina, Mustelidae) in Russian Far East and Hokkaido inferred from mitochondrial NADH dehydrogenase subunit 2 gene sequences. Mamm Stud 36: 209–222
Serizawa K, Suzuki H, Iwasa MA, Tsuchiya K, Pavlenko MV, Kartavtseva IV, et al. (2002) A spatial aspect on mitochondrial DNA genealogy in Apodemus peninsulae from East Asia. Biochem Genet 40: 149–161
Soltis DE, Morris AB, McLachlan JS, Manos PS, Soltis PS (2006) Comparative phylogeography of unglaciated eastern North America. Mol Ecol 15: 4261–4293
Stewart JR, Lister AM, Barnes I, Dalén L (2010) Refugia revisited: individualistic responses of species in space and time. Proc Biol Sci 277: 661–671
Svendsen JI, Alexanderson H, Astakhov VI, et al. (2004) Late Qua-ternary ice sheet history of northern Eurasia. Quaternary Sci Rev 23: 1229–1271
Taberlet P, Fumagalli L, Wust-Saucy AG, Cosson JF (1998) Com-parative phylogeography and postglacial colonization routes in Europe. Mol Ecol 7: 453–464
Tajima F (1989) Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123: 585–595
Takahara H, Igarashi Y, Hayashi R, Kumon F, Liew PM, Yamamoto M, et al. (2010) Millennial-scale variability in vegetation records from the East Asian Islands – Taiwan, Japan and Sakhalin. Quaternary Sci Rev 29: 2900–2917
Tamura K, Dudley J, Nei M, Kumar S (2007) Molecular Evolutionary Genetics Analysis (MEGA) Software Version 4.0. Mol Biol Evol24: 1596–1599
Waltari E, Cook JA (2005) Hares on ice: phylogeography and histor-ical demographics of Lepus arcticus, L. othus, and L. timidus(Mammalia: Lagomorpha). Mol Ecol 14: 3005–3016
Wu C, Wu J, Bunch TD, Li Q, Wang Y, Zhang YP (2005) Molecular phylogenetics and biogeography of Lepus in Eastern Asia based on mitochondrial DNA sequences. Mol Phylogenet Evol 37: 4561
Yamada F, Takaki M, Suzuki H (2002) Molecular phylogeny of Japanese Leporidae, the Amami rabbit Pentalagus furnessi, the Japanese hare Lepus brachyurus, and the mountain hare Lepus timidus, inferred from mitochondrial DNA sequences. Genes Genet Syst 77: 107–116
(Received December 27, 2011 / Accepted June 8, 2012)
Editorial Office
取り消し線
GKinoshita
ノート注釈
Svendsen JI, Alexanderson H, Astakhov VI, Demidov I, Dowdeswell JA, Funder S, et al. (2004)
GKinoshita
ノート注釈
issue no.省略でお願いします。
Related Documents











