Order Number 8824588 Anatomically preserved Glossopteris and Dicroidium from the Transantarctic mountains Pigg, Kathleen Belle, Ph.D. The Ohio State University, 1988 U MI 300 N. Zeeb Rd. Ann Arbor, MI 48106

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Order Num ber 8824588
A natom ically preserved Glossopteris and Dicroidium from the Transantarctic m ountains
Pigg, Kathleen Belle, Ph.D.
The Ohio State University, 1988
U MI300 N. Zeeb Rd.Ann Arbor, MI 48106

PLEASE NOTE:
In all cases this material has been filmed in the best possible way from the available copy. Problems encountered with this document have been identified here with a check mark V .
1. Glossy photographs or pages >/
2. Colored illustrations, paper or print_______
3. Photographs with dark background > /
4. Illustrations are poor copy_______
5. Pages with black marks, not original copy >/
6. Print shows through as there is text on both sides of p a g e_______
7. Indistinct, broken or small print on several pages \ /
8. Print exceeds margin requirements______
9. Tightly bound copy with print lost in sp ine_______
10. Computer printout pages with indistinct print______
11. Page(s)____________lacking when material received, and not available from school orauthor.
12. Page(s)____________seem to be missing in numbering only as text follows.
13. Two pages num bered . Text follows.
14. Curling and wrinkled pages______
15. Dissertation contains pages with print at a slant, filmed as received_________
16. Other _________________________ __________________________________________
UMI

ANATOMICALLY PRESERVED GLOSSOPTERIS AND DICROIDIUM FROM THE TRANSANTARCTIC MOUNTAINS
DISSERTATION
Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate
School of The Ohio State University
by
Kathleen Belle Pigg, B.S., M.Sc.
* * *
The Ohio State University
1988
Dissertation Committee:Thomas N. Taylor Daniel J. Crawford V. Raghavan Fred D. Sack
Approved By:
Advxser Department of Botany

ACKNOWLEDGMENTS
Support from the following grants during the writing of this dissertation is gratefully acknowledged: NationalScience Foundation Doctoral Dissertation Improvement Grant, Geological Society of America Research Grant, The Ohio State University Graduate School Alumni Research Award, and The Ohio State University Presidential Fellowship.
I would like to thank my reading committee, Drs. D.J. Crawford, V. Raghavan, F. D. Sack, and T. N. Taylor, for their careful editing and helpful comments on the manuscript. Drs. R. E. G. Boerner and D. L. Wrensch generously contributed advice and assistance on statistical procedures. Dr. R. Burry is gratefully acknowledged for the use of his computer facilities, and J. Osborn for assistance with graphics. Valuable insight into the problems of Gondwana paleobotany and gymnosperm phylogeny resulted from conversations with Drs. S. Archangelsky, G.W. Rothwell, R. A. Stockey, and E. L. Taylor. F. A. Bryan, D. F. Chappell, M. L. De Vore, B. L. Gibbins, L. K. Hussey, L. J. Thomas, J. A. Pigg, and E. von Bakonyi provided help and encouragement in countless ways.

I would especially like to thank Dr. T. N. Taylor for his advice, direction, and personal and professional encouragement throughout the course of this study.
iii

VITA
January 27,1958 ............ Born, Urbana, Ohio1978-1980 .................... Research assistant, Ohio
University, Athens, Ohio1980 ......................... B. Sc., Ohio University1980-1982 .................. Graduate teaching assistant
Graduate research assistant, Ohio University
1982 ........................ M. Sc., Ohio University1982-1987 ....................Graduate teaching assistant,
Graduate research assistant, The Ohio State University, Columbus, Ohio
1987-1988 ....................Presidential Fellow, TheOhio State University
1988 - ...................... Assistant Professor, ArizonaState University, Tempe, Arizona
1989 NATO Postdoctoral Fellow, University of Alberta, Edmonton, Alberta, Canada

PUBLICATIONSPigg, K. B., and G. W. Rothwell, 1979. Stem— root
transition of an Upper Pennsylvanian woody lycopsid. Amer. J. Bot. 66: 914-924.
Pigg, K. B., 1983. The morphology and reproductivebiology of the sigillarian cone Mazocarpon. Bot. Gaz. 144: 600-613.
Pigg, K. B., and G. W. Rothwell, 1983a. Chaloneria, gen. nov., heterosporous lycophytes from the Pennsylvanian of North America. Bot. Gaz. 144: 132-147.
Pigg, K. B., and G. W. Rothwell, 1983b. Megagametophyte development in the Chaloneriaceae fam. nov., permineralized Paleozoic Isoetales (Lycopsida). Bot. Gaz. 144: 295-302.
Pigg, K. B., and G. W. Rothwell, 1985. Corticaldevelopment in Chaloneria cormosa (Isoetales), and the biological derivation of compressed lycophyte decortication taxa. Palaeontology 28: 545-553.
Pigg, K. B., and T. N. Taylor, 1985a. Cormophyton gen. nov., a cormose lycopod from the Mazon Creek flora.Rev. Palaeobot. Palynol. 44: 165-181.
Pigg, K. B., and T. N. Taylor, 1985b. Anatomicallypreserved Glossopteris from the Beardmore Glacier area of Antarctica. Ant. J. U. S. 19: 8-10.
Pigg, K. B., R. A. Stockey, and T. N. Taylor, 1986.Studies of Paleozoic seed ferns: Additional studies ofMicrospermopteris aphyllum Baxter. Bot. Gaz. 147: 126-136.
Pigg, K. B., and T. N. Taylor. 1987a. Anatomicallypreserved Glossopteris from Antarctica. VII Simposio Argentino de Paleobotanica y Palinologia Actas, Buenos Aires: 177-180.
Pigg, K. B., and T. N. Taylor. 1987b. Anatomicallypreserved Dicroidium from the Transantarctic Mountains. Ant. J. U. S. (in press).
v

Pigg, K. B.f T. N. Taylor, and R. A. Stockey, 1987.Studies of Paleozoic seed ferns: Heterangiumkentuckyensis sp. nov., from the Upper Carboniferous of North America. Amer. J. Bot. 74: 1184-1204.
FIELDS OF STUDY Major field: Paleobotany

LIST OP TABLES
TABLE PAGE
1. Parameters measured from Glossopteris leavesfor quantitative analysis ..................... 181
2. Species of Indian Glossopteris used in quantitative analysis. Data from Chandraand Surange (1979).................. 182
3. Summary of major cuticular features reported for glossopterid leaves. H=hypostomatic, A=amphistomatic, N=narrow mesh, B=broad mesh s.c. =subsidiary cell, e.c.= epidermal cell, unspec.=unspecialized, gcl=guard cell length, irreg.=irregular orientation, conting. = contiguous stomata, long.=longitudinal orientation, *=fibers present. Data from indicated references.............................. 185
4. Occurrence of features in association withglossopterid cuticular studies (Table 3).1. Mesh shape [narrow (N) vs. broad (B)] . . . . 186
5. Occurrence of features in association withglossopterid cuticular studies (Table 3). 2.Hypostomatic (H) vs. amphistomatic (A), vs. unistomatic ( U ) ................................. 187
6. Occurrence of features in association with glosssopterid cuticular studies (Table 3).3. Epidermal cell wall shape (St= straight anticlinal margins; Si=sinuous, Rt=rectangular; Cv=curved, u=upper epidermis, l=lower epidermis). .188
vii

7. Occurrence of features in association withglossopterid cuticular studies (Table 3). 4.Relationship of mesh shape [narrow (N) vs. broad (B)] to stomatal position [hypo- (H)vs. amphi- (A) vs. unistomatic (U)]............... 189
8. Occurrence of features in association withglossopterid cuticular studies (Table 3). 5.Relationship between stomatal position [hypo- (H) vs. amphi- (A) vs. unistomatic (U)] andmesh shape [narrow (N) vs. broad B) ]............... 190
9. Results of multiple discriminate analysis of Antarctic leaves, based on continuous variables (i.e., mesh length, width, angle of lateral veins with midrib, and mesh area)....................... 191
10. Mean values + standard error for continuous variables for G. schopfii and G. skaarensis . . . .192
viii

LIST OF FIGURES
FIGURES PAGE1. Glosspteris schopfii. Composite of line
diagrams to illustrate morphological variability.......................... 193
2-5. Glossopteris schopfii. External morphology. . .1956-10. Glossopteris schopfii. Anatomical features . .19711-14. Glossopteris schopfii. Anatomical features . .19915-20. Glossopteris schopfii. Anatomical features . . 20121. Glossopteris schopfii. Distribution and
orientation of stomata........................ 20322. Glossopteris schopfii. Reconstruction of
stoma showing organization of stomatal apparatus (Fig. 19)............................ 205
23. Glossopteris schopfii. Reconstruction ofstoma showing single median papilla on subsidiary cells (Fig. 20).................... 205
24-27. Glossopteris schopfii. Morphology of twigs. . .20728. Glossopteris skaarensis. Composite of line
diagrams to illustrate morphological variability.................................... 209
29-32. Glossopteris skaarensis. External morphology. .21133-37. Glossopteris skaarensis. Anatomical features. .21338-41. Glossopteris skaarensis. Anatomical features. .21542-46. Glossopteris skaarensis. Anatomical features. .217
ix

47.
48.
49.
50.
51-55.56.
57.
58.
59-64.65-69.
70-75.
76-80.
81-82.
Glossopteris skaarensis. Line diagramshowing distribution and orientation ofstomata........................................ 219Glossopteris skaarensis. Reconstruction of stomata showing innner region. . ............ 221Glossopteris skaarensis. Reconstruction of stomata from outside. 7 ....................... 221Glossopteris skaarensis. Reconstruction of stomata showing subsidary cells ............. 221Glossopteris skaarensis. Stems................ 223Glossopteris skaarensis. Series of transverse sections thorugh twig to demonstrate phyllotaxy ........... . . . . .225Cluster diagram of Glossopteris leaves, including data from G. schopfii and G.\ skaarensis and Indian leaves................... 227Dicroidium fremouwensis. Composite of line diagrams that illustrate morphological variability in the taxon....................... 229Dicroidium fremouwensis. External morphology. .231 Dicroidium fremouwensis. Anatomical features. .233
Dicroidium fremouwensis. Anatomical features. .235
Dicroidium fremouwensis. Anatomical features.
Dicroidium fremouwensis. Anatomical features.
x

TABLE OF CONTENTSACKNOWLEDGMENTS ................................... iiV I T A ................................................. ivLIST OF TABLES . .................................... viiLIST OF FIGURES...................................... ix
CHAPTER PAGEI. INTRODUCTION .................................... 1
Southern Hemisphere (Gondwana) Floras . . . . 1 The Glossopteris and Dicroidium floras . . . 6Significance of silicified peat ........... 13
II. MATERIALS AND TECHNIQUES........................ 18DIRECTORY OF SPECIMENS .................. 18MATERIALS AND LOCALITIES ................ 22TECHNIQUES................................ 25
III. PART I GLOSSOPTERIS............................ 30INTRODUCTION.............................. 30
IV. DESCRIPTION.................................... 36Glossopteris schopfii Pigg sp. nov. . . . . .36Glossopteris skaarensis Pigg sp. nov........ 45
V. DISCUSSION...................................... 53Generic concept of Glossopteris............ 53Mixed floras............................... 62Species of Glossopteris.................... 69Affinities of the Antarctic l e a v e s ........ 75

Quantitative analysis of Glossopteris . . . .83The Glossopteris leaf f o r m ..................94
VI. PART II DICROIDIUM............................ 109INTRODUCTION...............................109
VII. DESCRIPTION.................................... 114Dicroidium fremouwensis Pigg sp. nov.
VIII. DISCUSSION.................................... 120Generic concept of Dicroidium ............ 120Species of Dicroidium .................... 124Frond architecture........................131
IK. SYSTEMATICS.................................... 138Glossopteris schopfii Pigg sp. nov.
; Diagnosis...........................13 8Glossopteris skaarensis Pigg sp. nov.
Diagnosis..........................140Dicroidium fremouwensis Pigg sp. nov.
Diagnosis ...................142LITERATURE CITED.....................................146TABLES ...............................................181F I G U R E S .............................................193
• •Xll

ANATOMICALLY PRESERVED GLOSSOPTERIS AND DICROIDIUM FOLIAGE FROM THE CENTRAL TRANSANTARCTIC MOUNTAINS
CHAPTER I INTRODUCTION
Southern hemisphere (Gondwana) floras.—Historically, Paleozoic and Mesozoic plants of the southern hemisphere (Gondwana) have been less extensively studied and less understood than their northern hemisphere counterparts. The difficulties of studying this material include the logistics of collecting in remote areas, communication among researchers on several continents, and the typically poor preservation of fossils. However, despite these limitations, paleobotanists have maintained a keen interest in Gondwana floras since the beginning of southern hemisphere exploration. This interest, sparked initially by the discovery of the broad-leafed fossil Glossopteris in India, Australia, and Antarctica (e.g., Seward, 1910, 1914), has continued through the years as the widespread and diverse nature of Gondwana plants became known, and especially as their potential relationship to angiosperm origins has been explored (e.g., Plumstead,
1

1952, 1956, 1958a; Melville, 1969, 1983a, Stebbins, 1974; Retallack and Dilcher, 1981; Doyle and Donoghue, 1986).
The Permian Glossopteris and Triassic Dicroidium floras are of considerable significance to a variety of important, broad-based biological and geological questions. First, their occurrence is indicative of past climatic conditions (Barghoorn, 1961; Plumstead, 1973). The repeated appearance of broad-leafed fossil floras in Antarctica during the Paleozoic and Mesozoic (e.g., Plumstead, 1962, 1975; Schopf, 1973), and the Cretaceous and Tertiary (e.g., Thompson and Burn, 1977; Jefferson, 1980, 1982a, b; Francis, 1986) demonstrates that climatic regimes on a worldwide basis have varied greatly through time (Chaloner and Lacey, 1973; Donn, 1982; Axelrod, 1984). Although Antarctica has remained at a relatively high latitude during much of the earth's history, the South Pole has not always been covered with ice (e.g., Cranwell, 1968; Smith and Hallam, 1970; Axelrod, 1984; Francis, 1986). The present-day polar ice cap is a relatively recent phenomenon thought to have been in existence only since the Miocene (Kemp and Barrett, 1975; Mercer, 1983). The documentation of polar floras at various times in earth's history thus reinforces interest in understanding the major forces in climatic conditions through time (Chaloner and Lacey, 1973; Jefferson, 1982a; Axelrod, 1984; Francis, 1986).

These fossil floras have also been of value first in the establishment of the concept of Gondwana, and subsequently in biostratigraphic correlations of southern hemisphere continents. The widespread distribution of the Gondwana flora throughout the southern hemisphere, together with the discovery of similar occurrences of the therapsid reptile Lvstrosaurus (Elliot, Colbert, Breed, Jenson and Powell, 1970; Colbert, 1973) was historically important as primary evidence for the theory of continental drift (e.g., Sahni, 1926? du Toit, 1957; Wegener, 1966; Schopf, 1970a; Plumstead, 1973; Lele, 1974). As the concepts of continental drift and plate tectonics have become more generally accepted through the years, emphasis has shifted to the value of the plant megafossil record in biostratigraphic and biogeographic correlation (e.g., Rigby and Schopf, 1969? Chaloner and Lacey, 1973? Schopf and Askin, 1980? Archangelsky, 1984; Rigby, 1983, 1985;Francis, 1986). Both megafossils and palynomorphs are particularly significant as index fossils in Gondwana since the marine fossil record is sporadic and difficult to correlate with Permian strata on a worldwide basis, while the nonmarine record is much more extensive (Schopf and Askin, 1980).
Although the stratigraphic and geographic distributions of plant megafossils have contributed a substantial amount of information toward the interpretation of Gondwana

paleogeography and paleoclimatology, Gondwana floras are known only in a very generalized way. At present, biological and structural attributes of the plants themselves remain poorly known. A major milestone in Gondwana paleobotany was reached when Gould and Delevoryas (1977) recognized anatomically preserved reproductive structures of Glossopteris that were leaf-borne, suggesting a relationship with the pteridosperms. However, at present, a clear view of either within-group diversity or the phylogenetic affinities of the dominant Permian and Triassic seed plants (i.e., glossopterids and corystosperms) has not yet emerged (e.g., Surange and Chandra, 1975; Gould and Delevoryas, 1977; Pant, 1977,1982; Petriella, 1981). While the phylogeny of the gymnosperms as a whole remains difficult to elucidate (e.g., Taylor, 1981; Meyen, 1984; Crane, 1985; Doyle and Donoghue, 1986), the position of the Gondwana gymnosperms during the period of radiation and diversity that characterized the late Paleozoic and early Mesozoic ascribes to them an important, but currently enigmatic, place in seed plant phylogeny. Moreover, the potential role of Gondwana plants in the origin of angiosperms (e.g., Thomas, 1933; Plumstead, 1958a; Retallack and Dilcher,1981; Taylor and Taylor, 1987a) remains equally difficult to determine, despite exhaustive comparisons of some of these complex vegetative and fertile structures to those of

extant flowering plants (e.g., Thomas, 1955; Melville, 1983a; Meyen, 1984; Crane, 1985). A more complete elucidation of the structure, biology and putative phylogenetic relationships of southern hemisphere plants will contribute significantly both to questions of vascular plant evolution and to increasing the accuracy of broader scale, paleoenvironmental interpretations.
Perhaps the greatest limitation to the study of southern hemisphere floras has been the type of fossil material available. Gondwana plants are preserved primarily as compression/impression fossils, forms which typically reveal little biological detail. For example, much of the difficulty in interpreting the structure of glossopterid reproductive organs lies in determining which features are anatomical in nature and which are the result of the preservation process (e.g., Plumstead, 1952, 1956; Rigby, 1978; Rex, 1986; Taylor, 1987). It is, therefore, the scarcity of informative material rather than a lack of significant biological and evolutionary questions that has limited the recognition and characterization of Gondwana plants. While, in the northern hemisphere, the diverse assemblage of Carboniferous plants has been extensively characterized from the information-rich permineralized plant remains of the coal swamps (i.e., coal balls; Phillips, 1980), until recently, comparable material has simply not been available from the southern hemisphere.

The Glossopteris and Dicroidium compression/impression floras: — Perhaps the most valuable Gondwana plant fossils relative to occurrence, botanical interest, and utility as biostratigraphic tools have been the compression/impression floras of Permian and Triassic age (e.g., Schopf, 1970a, 1973; Plumstead, 1973; Pant,1982). The reticulate-veined leaves Glossopteris and Ganaamopteris dominate the Permian flora throughout Gondwana (Maheshwari, 1974; Lele, 1976), while the related foliage types Palaeovittaria. Rubidgea, Eurvphvllum. Rhabdotaenia. Belemnopteris. and Pteronilssonia occur less frequently in the same strata (Maheshwari, 1965; Pant,1982? Rigby, 1983). Often associated with these leaves are unusual axes consisting of segmented wood known as Vertebraria (Gould, 1975a), fragments of wood assignable to such form genera as Araucarioxvlon. Dadoxvlon. and Antarctioxvlon (e.g., Krausel, Maithy, and Maheshwari,1961; Krausel, 1962; Maheshwari, 1972; Prasad, 1982, 1986; Pant and Singh, 1987).
Together with vegetative remains, over thirty genera of varied reproductive structures bearing either seeds (e.g, Dictvopteridium. Lidaettonia) or striate bisaccate pollen of the Protohaploxypinus type (e.g., Arberiella. Eretmonia. Glossotheca) have been discovered (Schopf,1970a, 1976; Surange and Chandra, 1975, 1976; Pant, 1977, 1982). The majority of both ovulate and pollen-bearing

reproductive structures consist of a dorsiventral leaf-like unit, usually on a short stalk, either axillary, borne upon, or adnate to, the central portion of a subtending leaf (e.g., Schopf, 1976? Retallack and Dilcher, 1981).Some genera have been interpreted as having several ovulate units per leaf, each bearing an individual ovule (e.g., Denkania. Surange and Chandra, 1971, 1975; Rigby, 1978) while others have multiple cupule-like structures per subtending leaf, bearing numerous ovules (e. g., Partha, Surange and Chandra, 1971, 1975). Still others (e.g., Dictvopteridium. Ottokaria. Pant, 1977; Rigby, 1978) have been interpreted as radial rather than dorsiventral in organization. Pollen-bearing structures demonstrate a similar range of morphological diversity from dorsiventral structures (e.g., Eretmonia. Lacey, Van Dijk and Gordon- Gray, 1975? Surange and Chandra, 1975) to strobilar forms (e.g., Kendrostrobus. Surange and Chandra, 1975).
For many years, the structure of glossopterid fructifications has been the topic of continued debate. As a consequence, various authors have interpreted the phylogenetic relationships among Glossooteris-bearincr plants in differing ways. Plumstead (1952, 1956, 1958a) initially interpreted glossopterid fertile structures as a diverse assemblage, some of which were ovulate-, others pollen-bearing, and still others, presumably bisexual. She classified these structures in the Glossopteridae, as a

class in a position intermediate between the angiosperms and other gymnosperms (Plumstead, 1956, 1958a, b). Surange and Chandra (1975) classified the glossopterids into two orders. They classified the cupulate forms (e.g., Lidaettonia. Denkania and Partha) into thePteridospermales, while other multiovulate forms which they believed cannot be easily homologized with other gymnosperms (e.g., (i.e., Scutum. Dictvopteridium. Ottokaria), they placed in the Glossopteridales (Surange and Chandra, 1975). Rigby (1978) also recognized this diversity, and placed glossopterid reproductive structures into three families within the Pteridospermales (i.e., the Dictyopteridiumaceae, Scutumaceae, and Ottokariaceae).Many authors (i.e., Meyen, 1984; Anderson and Anderson, 1985; Crane, 1985) have emphasized the importance of fertile structures over the vegetative glossopterid leaf for phylogenetic reconstruction. For this reason they have designated the glossopterids as the Arberiales (Meyen,1984), or by other names that are based on reproductive structures rather than the vegetative leaf.
Although glossopterid plants are poorly known, the diversity among reproductive structures has led to the suggestion that leaves with glossopterid venation represent a diverse assemblage of plants that are not necessarily closely related to one another (e.g., Delevoryas, 1973). Whether the glossopterid venation syndrome occurred in a

large plexus of related plants, or represents convergence of unrelated forms, remains, at present, conjectural.
The Permian can be further characterized by the occurrence of a small number of pteridophytic forms. Among the lycopods these include both presumably arborescent forms such as Cvclodendron and Lvcopodiopsis (Edwards,1952; Krausel, 1961; Rayner, 1985), Brasiliodendron (Chaloner, Leistokow, and Hill, 1979), the South African genus Azaniadendron (Rayner, 1986), and smaller, herbaceous species such as Selaqinella harrisiana (Townrow, 1968). Herbaceous sphenopsids include plants referable to the calamite/equisetoid lineage [i. e., Schizoneura. Phvllotheca. Paracalamites and Umbellanhvllites (e.g., Pant and Kidwai, 1968; Rigby, 1969; Pant, Misra and Nautiyal, 1982), Ranioaniia. Stellotheca. (e.g., Pant and Nautiyal, 1967; Maheshwari, 1967; Gould, 1975b), and putative members of the Sphenophyllales (e.g., Trizygia), (e.g., Maithy, 1976; Srivastava and Rigby, 1983; and references cited therein).
Marattialean ferns are represented in the Gondwana flora by vegetative and fertile foliage, in particular of the Asterotheca type (e.g., Damudosorus. Pant and Misra, 1971; Maheshwari, 1974; Anderson and Anderson, 1985), and, rarely, by stems of Psaronius (Brongniart, 1872; White,1908). In addition, osmundaceous ferns are represented by one species of permineralized Palaeosmunda from the Bowen

1 0
Basin of Queensland (Gould, 1970), one from Antarctica (Schopf, 1978) and two species of Osmundacaulis from South America (Herbst, 1975). The enigmatic form Botrvchioosis represents the oldest fern-like plant in the Permian, although some have suggested it may have a gymnospermous affinity (Maheshwari, 1974).
Other types of fern-like foliage found in the Permian of Gondwana include forms assigned to Euramerican taxa (e.g., Alethopteris. Sphenopteris. Pecopteris. and Ptvchocarpus). Although some northern hemisphere forms occur (e.g., Asterotheca. Sphenophvllum. and Corvnepterisl. particularly in western Gondwana as components of "mixed" floras (e.g., Archangelsky and Arrondo, 1969; Anderson and Anderson, 1985; Archangelsky and Cuneo, 1986), a number of additional genera have been recognized for foliage unique to the southern hemisphere. These forms, which occur predominantly in India, include Neomariopteris and Damudopteris (=Sphenopteris), Dichotomopteris (=Pecopteris and Pthvcocarpus) and Dizeuqotheca (=Alethopteris) (Maithy, 1972a, 1975; Pant and Khare, 1974; Maheshwari, 1974).
Apart from the glossopterids, seed plants are not particularly diverse or common in Permian Gondwana strata (Delevoryas, 1975; Pant, 1982; Archangelsky, 1985).Gondwana gymnosperms of Permian age include coniferophytes, putative ginkgophyte foliage (e.g., Psvgmophvllum. Ginkgophvllum. Maheshwari, 1974; Pant, 1982; Anderson and

Anderson, 1985), the enigmatic form Noeqqerathiopsis. and several forms of uncertain affinities. Conifers which are rare in the Gondwana flora (e.g., Archangelsky, 1985) are represented by such genera as Buriadia (Pant and Nautiyal, 1967), Walkomiella (Surange and Singh, 1953), and, in South America, Paranocladus (Surange and Lele, 1956). The oldest taxon, Buriadia. is considered a primitive conifer because it lacks an ovulate cone (Pant, 1982). Archangelsky and Cuneo (1987) have recently proposed the family Ferugliocladaceae for a putative coniferous group which contains ovulate cones with unusual structure. Based on leaf form, some have suggested that Noeqqerathiopsis may represent a southern hemisphere relative of the cordaites (Schopf, 1973; Gould, 1975b), but fertile structures of this plant have not been reported (Pant, 1982).
As in the Permian, the Triassic compression flora is dominated by a single foliage taxon, the bifurcate, pinnate frond Dicroidium (Anderson and Anderson, 1983). Dicroidium is generally regarded as the foliage of the Corystospermales, an endemic group of southern hemisphere pteridosperms (Thomas, 1933, 1955; Townrow, 1957). Other disarticulated plant organs included in the corystosperm seed fern complex are the foliage forms sometimes delimited as Xvlopteris. Johnstonia. and Pachvpteris (Baldoni, 1980; Petriella, 1979, 1981), [but synonomized by other authors with Dicroidium (Townrow, 1957; Archangelsky, 1968a)], the

pollen organs Pteruchus and Pteroma (Thomas, 1933? Petriella, 1980), the ovulate structures Umkomasia. Pilophorosperma and Spermatocodon. (Thomas, 1933;Petriella, 1980), genera recently synonomlzed as Umkomasia by Holmes (1987); and possibly the unusual stem taxon Rhexoxvlon (Archangelsky and Brett, 1961; Archangelsky, 1968a; Petriella, 1978, 1981). Although disarticulated parts of the corystosperms have been described in detail, and plants of this type have been tentatively reconstructed as small trees (Petriella, 1978), very little is known concerning the interrelationship of organs and the variability of taxa within this complex.
Other floristic elements of the Triassic compression flora include southern hemisphere members of the widespread lycopod Pleuromeia/Cvclostrobus complex (Helby and Martin, 1965; Retallack, 1977a; White, 1981), as well as the lycophyte cone Skilliostrobus (Ash, 1979) and the unusual lycophyte axis Cidarophvton (Chaloner and Turner, 1987). Relationships between northern and southern hemisphere members of this group remain poorly understood (e.g., Retallack, 1977a; White, 1981). Sphenopsids are represented by the genera Equisetites and Neocalamites (Lele, 1974), while ferns include the taxa Asterotheca. Cladophlebis. Todites and Dichtvophvllum (Lele, 1974).
Seed plants characteristic of the Triassic Gondwana floras include the conifers Rissikia and Voltziopsis

13(Townrow, 1967), cycadophyte foliage of the Pseudoctenis type (Lele, 1974), the cycad stem Leotocvcas (Delevoryas and Hope, 1971; Delevoryas, 1975), and the putative ginkgophytes Baiera and Ginkaoites (Lele, 1974). Other problematic gymnosperm taxa include the pteridospermous peltasperms (e.g., Lepidopteris. Antevsia. Townrow, 1956; Anderson and Anderson, 1985). In the compression floras of the Triassic Gondwana, these taxa tend to play a minor role, while the foliage form Dicroidium is often dominant (Anderson and Anderson, 1983, 1985).
Although most of the Gondwana pteridophytes can be recognized as members of the same major plant groups that occur in the northern hemisphere (i.e., at the class or ordinal level), southern hemisphere forms are typically distinct at the specific, if not the generic, rank. It is interesting that major seed plant groups at the ordinal and family levels (e.g., glossopterids, corystosperms) appear more likely to be endemic to Gondwana than are major pteridophyte groups (e.g., Archangelsky and Arrondo, 1969; Rigby, 1972). Furthermore, in both the Permian and Triassic these gymnosperms appear to represent the dominant floristic components (Schopf, 1973).
Significance of silicified peat:— Because Glossopteris and Dicroidium represent such significant components of the Permian and Triassic floras, respectively, there has been a continuing desire to

understand more of their anatomical structure, phylogenetic relationships, and the biology of the plants which bore these leaf types. Unfortunately, the paucity of anatomically preserved plant material has limited opportunities to investigate these details. Among the most exciting events in recent years in Gondwana paleobotany were the discoveries of three permineralized plant localities in the southern hemisphere (Gould, 1975b; Gould and Delevoryas, 1977; Schopf, 1970a, 1978; Smoot, Taylor, Collinson and Elliot, 1986; Smoot, Taylor and Collinson, 1987). One of these localities occurs in the Bowen Basin of Queensland, Australia, and is Permian in age (Gould, 1975b, 1980; Gould and Delevoryas, 1977). Two additional localities represent outcrops in the Central Transantarctic Mountains (Schopf, 1970b, 1978; Smoot, et al., 1986; Smoot, et al., 1987). One of these localities (i.e. Skaar Ridge =Mount Augusta sensu Schopf) is Permian in age while the other (Fremouw Peak) is of Triassic age. This material provides the first opportunity to detail the cellular organization of these important but poorly known leaf forms.
In the past several years, plants from these significant localities have been described in several research papers. The Permian-aged Bowen Basin locality yields an assemblage of plant fragments including glossopterid megasporophylls with attached seeds, putative

15vegetative leaves of Glossooteris. Vertebraria roots, Arberiella-like pollen organs bearing Protohaoloxvoinus- type pollen, osmundaceous fern axes, spores, and fungi (Gould, 1970, 1975a; Gould and Delevoryas, 1977).
Of the two Antarctic localities, Skaar Ridge of Permian age contains a fairly low diversity flora dominated by Vertebraria axes, glossopterid leaves (Pigg and Taylor, 1985, Pigg and Taylor, 1987a), and fertile structures (Taylor, 1987), ovules of several types (e.g.,Plectilospermum. Taylor and Taylor, 1987b), some demonstrating polyembryony (Smoot and Taylor, 1986a), a moss (Merceria, Smoot and Taylor, 1986a), putative ferns, and fungi (Schopf, 1970b; Stubblefield and Taylor, 1986).In contrast, the second Antarctic locality at Fremouw Peak contains a rich Triassic flora characterized by many elements (Smoot, et al., 1987) including corystospermaceous foliage (Pigg and Taylor, 1987b; Pigg, 1988); pollen organs (De Vore and Taylor, 1988), cycads (Antarcticvcas. Smoot, Taylor, and Delevoryas, 1985), several ovules (Taylor and Taylor, 1987b; Perovich and Taylor, 1988) and stems, leaves and fertile structures of several undescribed seed plants (Smoot, et al., 1987), several types of filicalean fern (Schopf, 1978; Millay, 1987; Millay, Taylor and Taylor, 1987), sphenophytes (Osborn and Taylor, 1988), putative lycopods, and fungi (Stubblefield and Taylor, 1986; Taylor and Stubblefield, 1987; White and Taylor, 1988).

In contrast to the relatively recent availability of permineralized Gondwana plants, anatomically preserved Carboniferous floras of Euramerica have been studied for over a century. During that time Carboniferous plants have been characterized with a fine degree of resolution, provided by the permineralized coal ball floras. This type of information has greatly influenced the course of northern hemisphere paleobotany. Because of this extensive and detailed data base, studies of Carboniferous plants have become increasingly sophisticated and synthetic in recent years (e.g., Knoll and Rothwell, 1981? Smoot and Taylor, 1985). Researchers using coal ball data have considered such diverse and intricate aspects of their plants as ontogeny (e.g., Eggert, 1961, 1962; Rothwell, 1971), pollen wall ultrastructure (Taylor, 1978; Taylor and Rothwell, 1982), reproductive biology (Millay and Eggert, 1974? Rothwell, 1977), the development of tissue systems such as phloem (Smoot and Vande Wege, 1986) and the vascular cambium (Cichan, 1985, 1986), paleoecology (e.g., DiMichele, Phillips and Olmstead, 1987, and references cited therein), the evolution of fungi (Stubblefield and Taylor, 1987), and evidence of plant-animal interactions (Cichan and Taylor, 1982? Taylor and Scott, 1983). In contrast, the initial descriptive studies of anatomically preserved Permian and Triassic Gondwana plants are only beginning to be published (e.g., Gould, 1970? Gould and

Delevoryas, 1977? Schopf, 1978; Smoot and Taylor, 1986b? Millay, et al., 1987? Smoot, et al., 1985? Pigg and Taylor, 1987a; Taylor and Taylor, 1987a, b, c). With the availability of permineralized plant material in Gondwana it is now possible to both provide initial descriptive data and, more importantly, to address many of the same types of biological questions that have intrigued paleobotanists studying Carboniferous coal floras.
The present study concentrates on the description of anatomical structure of permineralized Glossonteris and Dicroidium leaves from the Central Transantarcitio Mountains. This study has centered on the description and correlation of internal anatomy and external morphological form, the comparison of permineralized leaves to previously recognized compression/impression fossils, and addresses some biological questions concerning these prominent leaf forms that were so successful during the late Paleozoic and early Mesozoic. Through a combination of descriptive and synthetic approaches it is now possible to contribute to the formation of a significantly more realistic picture of plants of the Gondwana landscape than has previously been possible.

CHAPTER II - MATERIALS AND TECHNIQUES DIRECTORY OF SPECIMENS
All specimens of Glossopteris and Dicroidium foliage, and Glossopteris stems and twigs are housed in the Antarctic Paleobotanical Collections, Orton Museum of Geology, The Ohio State University. The material represents a combination of weathered and ddgaged specimens that reveal surface features, and microscope slides of sectioned material that were prepared by serial cellulose acetate peels.Glossopteris schopfii COLLECTION NUMBER FIGUREIndividual leaf specimens from degaged surfaces:Gl.l CB 452 Surface la, 2G1.2 " G1 Surface lb, 3G1.3 " Surface lcG1.4 " " ljG1 . 5 i i i i 5
G1.6 " " liG1.7 " "G1.8 " Fragment IdG1.9 " "
18

1961.10 i i i i
Gl.ll " Gl Surface61.12 " F Base61.13 i i i i
Gl. 14 i i
G1.15 n
Gl. 16 " Fragment61.17Gl. 18 452 Fragment61.19 i i n
61.20 i i n
Gl. 21 435 Leaf 1 seriesGl. 22 Ant 70-1-4361.23 i i
Gl. 24 i i
Gl. 25 i i
G1.26 463 E2 EdgeGl. 27 Ant 70-1-40Gl. 28 i i
Gl. 29 Ant 70-1-222Gl. 30 i i
Gl. 31 Ant 70-1-243Gl. 32 i i
Gl. 33 465 Edge
lgle
If
6, 9, 11-23 4
Slides with specimens from sectioned material: 451, 483, 484, 501, 503, 533, 535

2 0
Glossopteris skaarensisIndividual leaf specimens from degaged surfaces:G2.1 CB 463 E Bot Fragment 28a, 29G2.2 452 Surface 28b, 31G2.3 " FragmentG2.4 " " 28c, 32G2.5 " " 32G2.6 " "G2.7 '• " 28e, 30G2.8 " " 28dG2.9 " " 28fG2.10 " " 28gG2.11 " »G2.12 " "G2.13G2.14 Ant 70-1-43G2.15G2.16 463 E2 EdgeG2.17 Ant 70-1-2G2.18 Ant 70-1-222G2.19G2.20 465 peel series 33, 38-45
Slides with specimens from sectioned material:451, 465, 533

2 1
Dicroidium fremouwensisIndividual leaf specimens from ddgaged surfaces:D1 10137 A Top Surface 58a, 59D2 " •• " 58 fD3 " B Top SurfaceD4 " " "D5 » " "D6 " F Surfacejyj i i H i i
D8 10109 B2 Surface; B2 aside b, and B2c side b 58c, 61, 72
D9 10109 B2 SurfaceDIO " " "Dll " a2 Bot (part)D12 " B Top (counterpart)D13 10128 BD14 10235 B2 58b, 62D15 " •' » 58e, 63D16 " " "D17 " D EdgeD18 10112 58g, 64D19 " 58h

2 2
D20 »D21 10200 C3 58d, 60D22 CB 567Slides with specimens from sectioned material: 567, 602, 10003, 10109, 10137, 10200, 10235.
MATERIALS AND LOCALITIES Permineralized peat of Permian age occurs in the
Beardmore Glacier region of the Central Transantarctic Mountains at Skaar Ridge in Antarctica and contains numerous specimens of petrified glossopterid leaves of two types (Smoot, et al., 1986; Pigg and Taylor, 1985, 1987a). The material source occurs at 84 47' S°, 15' E, Buckley Island Quadrangle (Barrett and Elliot, 1973) at an elevation of approximately 2300 meters. The site occurs approximately 4.8 km from Mt. Augusta at the southeastern end of Skaar Ridge (Smoot, et al., 1986). Stratigraphically, the plant material occurs within the upper Buckley Formation and is considered late Permian in age (Barrett, 1969).
The locality was initially discovered by members of The Ohio State University Byrd Polar Research Center (formerly the Institute for Polar Studies) in the late 1960s (Schopf, 1970b, 1978). Further collections and preparation of the material were undertaken by Schopf

23(1970b, 1971, 1976, 1978) who also contributed a number of papers on the nature of the peat deposits (Schopf, 1970b, 1971). He also incorporated some glossopterid fertile material from the Skaar Ridge locality in a review of glossopterid fructification types (Schopf, 1976). The collecting site was recently recollected by Taylor and colleagues (Smoot, et al., 1986).
The petrified material occurs in a portion of the Buckley Formation and is predominated by carbonaceous siltstone and mudstone interbedded with tabular fining-upward sandstone bodies with erosional bases (Smoot, et al., 1986). Based on sandstone body geometry the depositional environment was initially interpreted by Barrett (1969) as a broad, shallow, low sinuosity stream occurring on a floodplain. More recent observations suggest channel sandstones more readily indicate anastomosing, braided stream deposits (Collinson and Isbell, 1986).
Plant material occurs as permineralized peat in jet black or light brown, siliceous blocks. Schopf (1971) suggested that silicification occurred as a result of infiltration of plant debris by waters highly charged with silica from nearby volcanic activity. He interpreted preservation of this peat to have occurred through several generations of silica, resulting in a matrix containing two generations of chalcedonic mineralization and a final

generation of prismatic quartz (Schopf, 1971). It has been suggested that the poor preservation of pollen and spores, especially in several nearby Triassic deposits may reflect the influence of thermal metamorphism from volcanics of Early- Middle Jurassic age (Kyle and Fasola, 1978). Cuticular material of glossopterid leaves in the present study is frequently poorly preserved and may have also been thus altered.
Triassic age permineralized peat occurs in the Beardmore Glacier region of the Central Transantarctic Mountains at Fremouw Peak (Smoot, et al., 1987). The locality occurs in the Queen Alexandra Range (84° 16' S, 164° 21' E, Buckley Island Quadrangle, Barrett and Elliot,1973) in a col north of Fremouw Peak (Fremouw Formation). Stratigraphically, the locality occurs in the upper Fremouw Formation, and is considered Early/Middle Triassic in age (Smoot, et al., 1987; Taylor, Smoot, and Collinson, 1985). Palynological investigations compare this matrix favorably with the Alisoorites zone, subzones A, B, and possibly C (Kyle, 1977; Kyle and Schopf, 1982), which correspond in. part to the Falcisporites Superzone of Helby, Morgan and Partridge (1987), suggesting an age of Scythian/Ladinian, or lower-middle Triassic (Farabee, Taylor and Taylor,1988a, b). Megafossil data (i.e., Dicroidium. this paper) places this deposit within the D. zuberi zone of Retallack

25(1977b) (=early-middle Scythian) for the Sydney Basin and eastern Australia.
Fossil plant material occurs within several allochthonous clasts that are at approximately the same stratigraphic level within a trough-crossbedded, medium-grained, greenish-grey volcaniclastic sandstone.The clasts are block-shaped and up to 1.0 m thick and 2.5 x 2.5 m wide. Below the peat layer is a zone 0.4 m thick of light grey mudstone. Based on sedimentological evidence, the depositional environment is interpreted as a braided stream containing forested islands that were undercut during flooding. Trough axes in the surrounding sandstone deposit tend to be deflected around the peat blocks, suggesting that the plant material was rafted in and deposited in shallow water. Fossilization is thought to have occurred rapidly, before substantial microbial degradation took place, since the plant material is well preserved (Schopf, 1971). Abundant sources of silica may have been available from both surrounding sands and extensive volcanic activity.
TECHNIQUESPreparation of the Glossopteris and Dicroidium leaf
specimens included a combination of techniques.Glossopterid leaves and fronds of Dicroidium occurring on weathered surfaces were degaged to uncover a maximum area

2 6
from the matrix by a modification of the "micro jackhammer11 technique as described in Pigg, Taylor and Stockey (1987). Specimens were photographed with the use of low-angle lighting to enhance poorly visible surface detail. Line diagrams were prepared by tracing photographs and magnifications were standardized at lOx by enlarging images.
Anatomical detail was determined by closely-spaced serial sections by the cellulose acetate peel technique (Joy, Willis and Lacey, 1956), as modified for silicified peat (Basinger and Rothwell, 1977), with 40% hydrofluoric acid. Paradermal, transverse and various oblique sections were prepared by the peel method in order to study anatomical features and to correlate histology with external morphological form. Pertinent peels were mounted on microscope slides for transmitted light microscopy.Line diagrams of critical sections from peels and/or slides were drawn with the aid of a drawing tube (camera lucida attachment) on a Wild 5D dissecting microscope.
Cuticular remains were bulk macerated from the matrix with 40% hydrofluoric acid and gradually neutralized by 8-10 changes of distilled water. These were then mounted on SEM stubs for microscopy or in glycerine on slides for light microscopy. Additional cuticular specimens were studied directly from acetate peels. Stomatal density was calculated from camera lucida tracings of paradermal peel

27sections showing distribution and orientation of stomata. Since tissue is poorly preserved, selected areas with the best preservation were used for calculations. Preservation did not allow for reliable counts of epidermal cells/area, so stomatal index (=ratio of stomatal number to total epidermal cell number/area) was not calculated.
For a quantitative study of Glossopteris leaf morphology, a total of 17 morphological characters were scored for 10 individual specimens of G. schopfii and 7 of G. skaarensis (Table 1). These include midrib thickness, leaf width, as measured directly or calculated from fragments in which at least 1/2 of the leaf width was present, vein density in central part of leaf/cm2, and marginal vein density/cm2. Also scored (/cm2) were the following types of reticulations: Y (=simple dichotomy),lambda (=simple reticulation), H, N, X, W, and complex reticulations, and total reticulations/cm2. In addition, four parameters of the intercostal or mesh areas were scored: mesh length, width, area, and angle of mesh with the midrib.
Measurements were taken from a standardized central area of each leaf fragment since venation tends to vary in more apical and basal regions. Sample size was established as the area in which 30 meshes of the largest mesh size could be measured (=15.5 cm2). The first 13 characters were scored manually from line diagrams of the leaves ca.

lOx magnification, resulting in one value per specimen; the last 4 (features 14-17 of Table 1) were scored with Zeiss Videoplan image analyzer in the following ways: mesh length (=DMAX parameter), width (=LENGTH parameter), area (=AREA parameter), angle of meshes with the midrib (=ANGLE X parameter), aligning all measurements in the same orientation. DMAX, AREA, and ANGLE X parameters were taken simultaneously, while LENGTH measurements were taken separately. Measurements of the last four parameters resulted in multiple (10-50) measurements per specimen, depending on the number of meshes preserved within the designated grid.
A comparative data set of the same 17 morphological characters was generated from line diagrams of 58 Indian glossopterid leaves, as presented in Chandra and Surange (1979) (Table 2). Representative specimens selected from this monograph included all species where a central region of the leaf was illustrated.
Stepwise and multivariate discriminate analysis and cluster analysis were performed with the glossopterid leaf data (Statistical Analysis Systems, 1985) using three modifications of the data set. For the first analysis, all readings for mesh length, width, area and angle with midrib were included along with the single readings (i.e., one per leaf specimen) of the other 13 parameters. In the second analysis, only the four continuous variables (i.e., mesh

length, width, area and angle with midrib) were included.In the third analysis, a mean value for the four parameters (mesh length, width, area and angle with midrib) was generated and all other single values for other parameters were included. These differing analyses were undertaken to assess and minimize the weight given to parameters with single values.

CHAPTER III - PART I. Glossopteris INTRODUCTION
The genus Glossopteris represents a sphathulate leaf form with entire margins, a persistent midrib, and reticulate venation (Brongniart, 1828; Arber, 1905; Kovdcs-Endrddy, 1981). Glossopteris is the dominant foliage form in the Permian compression/impression floras of Gondwana, and occurs on all the major continents of the southern hemisphere (Pant, 1977, 1982). A widely diverse array of leaves conforms to this genus, and over the years upwards of 200 species have been described (e.g., Pant and Singh, 1971; Chandra and Surange, 1979; Anderson and Anderson, 1985; Boersma and Broekmeyer, 1987, and references cited therein). This large number of species results from several factors. First, the considerable morphological variation within the group provides a basis for the delimitation of many species. Secondly,. since closely circumscribed species have been valuable as biostratigraphic tools, new species often have been created along with new records of stratigraphic occurrence. Specimens of Glossopteris have served as valuable biostratigraphic indicators, particularly in Australia
3 0

(e.g., Rigby, 1983, 1985) and India (Banerjee, 1978; Banerjee and Ghosh, 1970; Chandra and Surange, 1979). Thirdly, since cuticular variability does not often correlate with morphological features, many new species have been created to encompass those forms with cuticle (e.g., Pant and Gupta, 1968, 1971; Pant and Singh, 1971,1974).
Historically, Glossopteris leaves were initially described by Brongniart (1828) and a number of other Europeans interested in the geology and fossil floras of the colonial areas of the southern hemisphere including India, Australia, New Zealand, southern Africa, and areas of South America. By the end of the 19th century, many species, especially from India and Australia, had been established by Feistmantel (Feistmantel 1878-79, 1880a, b 1881, 1882, 1886), and others, (e.g., Bunbury, 1861, Oldham, 1897, Dana, 1849; Seward, 1910) (deJersey, 1968). Extensive collections of Gondwana plants during this time led to several monographic works and reviews (e.g., Tenison-Woods, 1883; Feistmantel, 1878-9; White, 1908). During this period Arber (1905) catalogued the Permian Gondwana holdings in the British Museum (Natural History) In the ensuing years a large number of collections were made from Permian strata of the Gondwana continents, resulting in the description of numerous species of Glossopteris.

Among the most useful summaries of regional Glossopteris floras to date are the following: India(Feistmantel, 1880a, b, 1881, 1882, 1886; Maheshwari, 1965, 1974; Surange, 1973; Lele, 1974, 1976; Chandra and Surange, 1979; and references cited therein); Australia (Dana, 1845; Feistmantel, 1890; Rigby, 1966, 1983, 1985; Rigby, Maheshwari and Schopf, 1980; Gould, 1975b; White, 1978; Retallack, 1980); Antarctica (Seward, 1914; Darrah, 1936; Plumstead, 1962, 1975; Cridland, 1963; Rigby and Schopf, 1969); New Zealand (Mildenhall, 1976; South Africa: Feistmantel, 1889; Kovacs-Endrody, 1976, 1977, 1979, 1983, 1987; Lacey, van Dijk and Gordon-Gray, 1975; Lacey, 1976; LeRoux and Anderson, 1977; Anderson and Anderson, 1983; Rayner and Coventry, 1985); Madagascar (Appert, 1977 and references cited therein); South America (Archangelsky, 1968b; Graham, 1979, 1982, and references cited therein); Argentina (Archangelsky, 1957; Archangelsky and Arrondo, 1969, 1975; Cuneo, 1987; Brazil: White, 1908; Dolianiti, 1946, 1954; Rosier and Fittipaldi, 1981; Perinotto and Rosier, 1984; Uruguay (Herbst, Ferrando, and Jalfin, 1987); the Falkland Islands (Halle, 1911; Seward and Walton,1923).
The widespread Permian occurrence of leaves with the glossopterid venation form, in association with Vertebraria rooting structures, Antarctioxvlon or Araucarioxvlon wood, and glossopterid fertile structures has led some authors to

present a generalized reconstruction of the Glossopteris plant (e.g., Plumstead, 1958b; Pant, 1977? Gould and Delevoryas, 1977). The plant Glossopteris is usually reconstructed as a large arborescent form bearing deciduous leaves, in whorls or a tight helix (Pant and Singh, 1974; Pant, 1977). Fertile structures of glossopterids have been interpreted as variously borne upright, ab-, or adaxially on the plant (e.g., Rigby, 1978? Retallack and Dilcher, 1981; Rex, 1986? Taylor, 1987). Although this has been the general concept of Glossopteris. little evidence supports it as the only, or even the primary type of plant that bore leaves with the glossopterid syndrome.
To date, the only reports of permineralized vegetative leaf remains ascribed to Glossopteris are those characterized by Gould and Delevoryas (1977) in conjunction with glossopterid fertile material from the Bowen Basin of Queensland, Australia, and preliminary reports by Schopf (1970b) of leaves of the Antarctic. Petrified leaves from the Australian locality (Gould and Delevoryas, 1977) demonstrated a combination of significant anatomical features, including radially aligned tracheids in the vascular strands, prominent hypodermis, well-differentiated palisade and spongy mesophyll, and sunken stomata (Fig.2A-E of Gould and Delevoryas, 1977). These leaves resembled the megasporophylls that were the major topic of the paper, however, they were not studied as to

34morphological (surface) features, compared with compression taxa, or even conclusively demonstrated to be Glossopteris.
The anatomically preserved leaves from the Central Transantarctic Mountains described by Schopf (1970b) demonstrate internal anatomical detail that provides substantial new information to our current data base for Glossopteris (Pigg and Taylor, 1985, 1987a). This anatomic information is valuable in addressing several questions at both a systematic and a biological level. The study of internal anatomy coupled with external morphological detail provides, for the first time, a basis for recognizing the underlying cellular structure of plant material that determines external morphological pattern. Anatomical information of this type thus allows for a reassessment of certain morphological features (e.g., midrib, venation structure) that have traditionally been used as taxonomic characters in studies of Glossopteris. To a certain extent, information of this type can also be extrapolated to interpret the structural features of similar leaves preserved only as compression/impression fossils (Pigg and Rothwell, 1985).
The availability of numerous permineralized specimens with similar anatomy provides an opportunity to assess variability within a "species" of Glossopteris based on anatomical features. By quantifying morphological variability present in a group of leaves that all possess

35the same anatomy, an independent measure can be used to reconsider the boundaries of species based on morphology alone.
Apart from systematic considerations, this data provides new information on the biological structure of the leaf form. Whatever their phylogenetic affinities, the glossopterid leaf form was the prevalent leaf type during the Permian of Gondwana. Certainly the internal structure of this leaf form might reflect physiological and structural/functional parameters that were important to the success of plants in the Permian environment. This information is of potential value in understanding more about both basic leaf form and function, and the climactic and ecological factors present during the late Paleozoic of the southern hemisphere.

CHAPTER IV - DESCRIPTION
Glossopteris schopfii: The description of G. schopfiiis based on the study of 30 specimens of leaves occurring on weathered surfaces that show morphological detail, and an additional 200 specimens of fragmentary leaves cut in various planes of section. Studies of the external morphology, including patterns of venation, are based on the first specimens (Fig. 1-6), while sectioned material provides internal anatomical detail (Fig. 7-20). Several specimens (e.g., Fig. 6, 9, 11-20) were peeled in both transverse and paradermal views in order to correlate structural features in a three-dimensional context.
Morphological features typically used as taxonomic characters for Glossopteris include leaf size and shape, length/width ratio, shape of apex and base, and details of venation such as persistence of the midrib, vein density, angle of lateral veins with the midrib and the leaf margin, and types of reticulations (="cross connections") formed between veins (Maheshwari, 1965; Melville, 1969; Chandra and Surange, 1979). In addition, details of the meshes delineated by veins include mesh length, width, area, and shape have been used as descriptive characters (e.g.,
3 6

Banerjee, 1978; Rigby, 1983). These features are the most inclusive since they can be compared with similar features in leaves preserved as impressions (e.g., Cridland, 1963; Chandra and Surange, 1979), (which are limited to these characters), with compressions which exhibit both these features and cuticular detail (e.g., Pant and Gupta, 1968, 1971; Pant and Pant, 1987), and with petrified leaves (Schopf, 1970b; Gould and Delevoryas, 1977; Pigg and Taylor, 1985, 1987a), which can potentially exhibit morphology, cuticle, and, most importantly, internal anatomy.
Leaves of G. schopfii are 1.3-1.8 cm wide based on direct measurements where the total width is preserved and are estimated from partial leaf fragments (where at least 1/2 of the width is preserved) to be up to 3.1 cm (Fig.1-6). No complete leaves were found, but the longest specimen is 6.8 cm and includes the apex (Fig. lb, 3). The width of this specimen remains relatively constant throughout its length, tapering slightly toward the base, suggesting that leaf shape was oblanceolate. Since the base is not preserved, and since glossopterid leaf bases are known to exhibit a wide range of morphologies, including an apetiolate condition (e.g., Chandra and Surange, 1979), the total length cannot be determined with certainty. However, it is unlikely that leaves of this sort exceeded 10-12 cm. Assuming that leaves are around 3

38cm in maximum width and up to 10 - 12 cm long, they would represent either large "small-sized" leaves or small "medium-sized" leaves according to the ranges suggested by Chandra and Surange (1979). Based on these dimensions, leaves have a length/width ratio in the range of 4:1 - 3:1. Six specimens have been discovered with complete apices: they are obtuse to retuse in outline (Fig. la, g, 4, 5).
Stalk-like structures 100 x 50 pm in diameter with a single vascular bundle and a similar histology are found throughout the Skaar Ridge matrix (Fig. 25), but none have been found in attachment to vegetative leaves. It is unclear whether these axes represent the petioles of vegetative leaves, of fertile leaves, or of either ovulate- or pollen-bearing structures attached to fertile leaves. However, other stalk-like structures (Fig. 27) which extend to vascularize the base of ovule-bearing structures have the characteristic anatomy of G. schopfii leaves (Taylor, 1987).
Typical leaves of G. schopfii exhibit a prominent midrib ca. 1.6 mm wide composed of 4-5 parallel-oriented strands which remains distinct for most of the length of the leaf (Fig. 3, 6). In some specimens, the midrib is fractured along the same plane as the remainder of the leaf (Fig. le, f, g, i, 5, 6); in others the midrib may be fractured at a more external plane (Fig. Id, h, j, 4). In the first case, the individual vascular strands of the

midrib are more distinct and the midrib appears to be even with the rest of the leaf, while in the second situation the midrib appears more prominent and individual strands are indistinct (Fig. 3, 4) This disparity is obvious in specimens where portions of the midrib are fractured at both levels (Fig. la, b, 2 and 3 at arrows). Since the external appearance of the midrib may not necessarily reflect its internal structure, the more general term midrib, rather than midvein, is used in the present study. In the upper few millimeters, near the apex, strands tend to anastomose (Fig. la, 5).
Throughout the leaf, 1-3 veins adjacent to the strands of the midrib are vertically aligned and frequently anastomose, creating a region of 1-2 meshes aligned in parallel with the midrib. Meshes of this type are elongate and ca. 3.7 x 0.3 mm (Fig. Ii, 6). Laterally, veins arch out toward the margin, at an angle of 53.30° ± 1.85 (Fig. 16, j, 6). In this region, meshes become longer (3.80 mm ± 0.10 x 0.49 mm ± 0.15) with an area of 1.48 mm ± 0.60, and curve toward the leaf margin. This area is characterized by numerous connections between veins, resulting in a highly reticulate configuration (Fig. 1, 6). Vein interconnections include simple Y-shaped dichotomies (= vein junction type "gamma" of Melville, 1969), Y-shaped reticulations (= "lambda" sensu Melville), X-shaped configurations resulting from a Y-shaped reticulation

4 0
immediately followed by a second dichotomy (= "chi" sensu Melville), H-shaped interconnections (= "eta" of Melville), N-shaped interconnections (="zeta" of Melville), and combinations of dichotomies and reticulations that result in W-shaped and more complex configurations (Fig. 6, 9,12) .
In following a single, lateral vascular bundle from its origin in the midrib, a sequence of 3 - 4 dichotomies and reticulations results in the formation of 2 - 3 meshes. The number of dichotomies and subsequent reticulations in the central region of leaves is approximately equal, with the result that the number of veins/area remains relatively constant. Total vein concentration is ca. 27.3/cm2 with veins in the central region measured at ca. 22.2/cm2 and marginal veins at 21.0/cm2. Veins arch toward the leaf margin, when they meet at an angle of ca. 22°. There is little difference in veins adjacent to the margin compared to those of the central part of the leaf.
In a transverse section through the central region, leaves of G. schopfii are characterized by a central area that is thicker than the lateral laminae (Fig. 7). The area of the midrib ranges in thickness from 0.3 - 0.7 mm; laterally, laminae are 0.2 - 0.3 mm thick. Leaves also vary in thickness and shape along their length: thelaminae of individual leaves are thicker and somewhat rounder in outline in more proximal regions, or areas near

41the petiole. Although it is unclear whether non-laminar petioles were present or if leaves were broadly attached to the stem, the central vascular strand becomes more prominent proximally.
Characteristically, vascular bundles of G. schopfii exhibit vascular strands with conspicuous bundle sheaths (Fig. 7-10). Numerous fibers occur in the vascular bundle sheath, causing the sheath to appear dark and thickened (Fig. 7-10, 12). The anatomical organization of the lateral bundles is the same as that of the midrib, with the only appreciable difference being one of size. The more prominent midrib of some specimens is the result of the combination of the larger size of median vascular bundles and the increase in thickness of the mesophyll itself.
Vascular bundles contain 1 or 2 protoxylem strands surrounded by a group of 20 - 30 metaxylem tracheids in a mesarch to almost exarch arrangement (Fig. 10). Protoxylem tracheids are characterized by annular or helical wall thickening patterns. The associated metaxylem tracheids occasionally exhibit helical wall thickenings, or circular bordered pits, but most possess scalariform/reticulate wall thickening patterns (Fig. 16). In most leaves, the primary xylem is bordered on the abaxial side by a fringe of up to 3 - 8 rows of smaller, radially aligned tracheids that are rectangular in transverse section (Fig. 10). These cells, which presumably represent secondary xylem, are up to 80

42/un in diameter and have scalariform wall thickenings. Individual tracheids within dichotomies and reticulations exhibit a variety of morphologies. Along with regular elongate tracheids are ones that diagonally traverse more than one arm of a W-shaped reticulation (Fig. 9). Still others within dichotomies may bifurcate, with either branch extending into each arm of the dichotomy (Fig. 17). Within some dichotomies as small, transfusion-like tracheids that extend around the inside of the bundle.
Within the vascular bundles, a prominent lacuna occurs on the abaxial side of the strand (0.2 X 0.08 mm), representing the position of the phloem (Fig. 8, 10). No evidence of phloem has been found, but the size of the lacunae suggests that leaves contained a large region of phloem and adjacent tissues. In some specimens the vascular strand is separated, with the radially aligned tracheids displaced in a position within or to the opposite side of the lacuna (Fig. 8, 10, at right). This feature appears to be the result of preservational distortion and does not reflect anatomical differences between vascular bundles.
The bundle sheath is composed of 2-3 layers of elongate cells. The bundle sheath contains numerous elongate fiber-like cells (Fig. 15) that appear rounded in transverse section (Fig. 10). Cells of this type measure about 17 /im wide x 110 /ixm long with thickened walls, and represent bundle sheath fibers. These cells are

43characterized by simple pits in their lateral walls. Pits are most obvious in cells containing amorphous dark material that extends into the wall, delineating their position in lateral and face views (Fig. 15).
Hesophyll is parenchymatous and exhibits little to no differentiation: palisade and spongy layers are absent. Because of the poor preservation of tissues, it is difficult to determine the original nature of the mesophyll, however, in many instances the tissue appears aerenchymatous (Fig. 12, 13). Whether this appearance is due to the original organization and arrangement of this tissue or is primarily the result of substantial shrinkage and distortion of cells during fossilization is unclear.On both ab- and adaxial laminar surfaces is a 1 - 3 celled hypodermis (Fig. 7, 14). Cells of the hypodermis are 25 - 30 fim wide and cuboidal: they frequently contain, dark,amorphous, possibly ergastic substances (Fig. 14).
Cuticle of G. schopfii is only rarely preserved and details are difficult to distinguish. Individual cell margins may be indistinct, and the overall appearance of fragmentary cuticle suggests that it may have been altered during fossilization. Cuticle occurs only on small portions of leaves that have adhered directly to other plant fragments in the matrix, that presumably provided some protection or structural integrity that allowed for its preservation (Fig. 11). Epidermal cells are usually

4 4
absent or torn apart, so that epidermal cell patterns are interpreted primarily from those cuticular fragments that remain.
Epidermal cells on both surfaces are around 53 x 22 Mm, and are elongate with straight-margined anticlinal walls (Fig. 18-20, 22, 23). Papillae 7.5 in diameter are present in the central region of epidermal cells on both surfaces (Fig. 20, 23). Glossopteris schopfii leaves are hypostomatic with randomly distributed stomata that tend to be aligned in parallel with the long axis of the leaf (Fig. 21). Stomata occur in a density of ca.39.2/mm2. Stomatal complexes are slightly sunken, and composed of 4-5 subsidiary cells 65 x 35 fm that are loosely organized around the stomatal apparatus. Guard cells are 43 x 10 pm and have thickened margins. They surround an elongate stoma 25 x 2.5 /m (Fig. 18-20, 22,23).
Five twigs sectioned distal to the level of the stele have been found in transverse and oblique sections. They bear up to 12 leaves (Fig. 24, 26). The identification of these twigs with G. schopfii is based on several prominent anatomical features, including the structure of leaf laminae and the presence on some of the leaves of prominent lacunae, which represent the position of vascular bundles (Fig. 24, at right). The innermost, smallest leaves are 250 x 31 pm and have a prominent, keeled central region but

45only slightly developed lateral laminae (Fig. 24, 26). Leaves to the outside of these are comparable to mature leaves in width. These leaves tend to overlap one another, enclosing the axis (Fig. 24). Mesophyll is not well preserved.
Glossooteris skaarensis — Glossopteris skaarensis Pigg sp. nov. occurs approximately half as frequently in the Skaar Ridge matrix as G. schopfii. Leaves of this type are difficult to recognize in the matrix since they frequently lack internal tissues and are crushed. The description of G. skaarensis is based on the study of 20 fragmentary specimens from weathered surfaces which provide morphological detail, and a total of 50 additional leaves cut in transverse, oblique and paradermal section (Fig. 28-46). Several specimens (e.g., Fig. 33, 38-45) have been cut in both paradermal and transverse section to correlate features as observed in multiple sections.
Leaves of G. skaarensis in which the total width is preserved are up to 2.2 cm wide (Fig. 28-32). The maximum width of leaves is based on from fragmentary specimens in which at least half of the leaf is preserved is 2.7 cm.The most complete leaf fragment is 3.5 cm long (Fig. 28b, 31), and represents only the central portion of the lamina. The total length of leaves cannot be determined with certainty, but in comparison with leaves of similar width preserved as compressions, leaves are estimated to be up to

10 - 12 cm long. As in G. schopfii. no specimens of basal or petiolar areas have been found. However, petiole bases found in attachment to stems are oval-triangular in transverse section, suggesting that petioles of some type were present (Fig. 53, lower right). Because of the fragmentary nature of specimens, length/width ratio and shape are difficult to determine, but several specimens tend to taper proximally (Fig. 28b, e, f), suggesting that leaves were probably oblanceolate. Three leaves with complete apices were discovered: apices are retuse andsometimes asymmetric in outline (Fig. 32).
Externally, G. skaarensis leaves appear to possess a prominent midrib up to 1.1 mm wide which protrudes out from the lateral part of the leaf (Fig. 29-32)..To some extent, as in G. schopfii. this appearance is the result of the midrib and the lateral laminae being fractured at different planes, however, in general the midrib is prominent in this specimen. The midrib persists to the apex, with lateral veins extending from it near the tip (Fig. 32). Lateral veins anastomose to form broad, polygonal (usually pentagonal) meshes 2.86 mm ± 0.77 x 0.60 ± 0.16 mm with an area of 1.44 ± .613 (Fig. 29, 33, 47). At the point of divergence from the midrib, meshes create an angle of 57.82° ± 1.48. Following an individual strand laterally, 3-5 meshes are crossed to reach the margin. Veins are more closely spaced at the margin, which they meet at

10 - 12 cm long. As in G. schopfii. no specimens of basal or petiolar areas have been found. However, petiole bases found in attachment to stems are oval-triangular in transverse section, suggesting that petioles of some type were present (Fig. 53, lower right). Because of the fragmentary nature of specimens, length/width ratio and shape are difficult to determine, but several specimens tend to taper proximally (Fig. 28b, e, f), suggesting that leaves were probably oblanceolate. Three leaves with complete apices were discovered: apices are retuse andsometimes asymmetric in outline (Fig. 32).
Externally, G. skaarensis leaves appear to possess a prominent midrib up to 1.1 mm wide which protrudes out from the lateral part of the leaf (Fig. 29-32). To some extent, as in G. schopfii. this appearance is the result of the midrib and the lateral laminae being fractured at different planes, however, in general the midrib is prominent in this specimen. The midrib persists to the apex, with lateral veins extending from it near the tip (Fig. 32). Lateral veins anastomose to form broad, polygonal (usually pentagonal) meshes 2.86 mm ± 0.77 x 0.60 ± 0.16 mm with an area of 1.44 ± .613 (Fig. 29, 33, 47). At the point of divergence from the midrib, meshes create an angle of 57.82° ± 1.48. Following an individual strand laterally, 3-5 meshes are crossed to reach the margin. Veins are more closely spaced at the margin, which they meet at

47approximately right angles (Fig. 28b, c, e, g, 31). Total vein concentration is ca. 21.1/cm2, with veins in the central region measured at ca. 13.7/cm2 and marginal veins at 19/cm2.
In transverse section, leaves of 6. skaarensis typically are preserved with a thick, inflated midrib area approximately 0.5 mm thick that narrows abruptly to 0.13 - 0.14 mm in the region of the lateral laminae (Fig. 35). Since the internal mesophyll tissues are usually poorly preserved and seldom show tissue continuity, it is difficult to assess whether this configuration represents the original tissue state or a post-taphonomic alteration. However, the increased thickness in the central region of G. skaarensis leaves is apparently the result of a combination of the larger central vascular strands and a thicker mesophyll region in the region of the midrib (Fig. 35). Leaves possess a prominent hypodermis on both surfaces (Fig. 34, 35, 37), and when preserved, thin-walled parenchymatous central mesophyll (Fig. 38). As in 6. schopfii. there is no indication of differentiation of the mesophyll into palisade and spongy layers. Cells of the hypodermis are approximately 48 (36 - 57) /urn in diameter and appear cuboidal in transverse section (Fig. 34). When well preserved they may have thickened walls or dark inclusions up to 16 frn wide. Vascular strands are up to 0.2 mm in diameter in the central region, in lateral

laminae they are less conspicuous. Unlike the strands of G. schopfii. those of fi. skaarensis lack a prominent fibrous bundle sheath. Rather, they are delimited by a bundle sheath of 1-2 elongate thin-walled cells (Fig. 41). The vascular strand contains an area of primary xylem 0.3 mm in diameter in which a small group of protoxylem strands can be distinguished. Wall thickenings on protoxylem and metaxylem elements are typically helical. Abaxial in position to the primary xylem is a zone of 15-20 rows of small, radially aligned tracheids (Fig. 34, 37). These tracheids typically possess helical and scalariform wall thickenings (Fig. 40).
Cuticular and epidermal features of G. skaarensis are better preserved than those of G. schopfii. and allow for a more complete description of their complex structure. A prominent cuticle is present on both surfaces. In transverse section the cuticle appears irregular and jagged, particularly on the abaxial surface (Fig. 34, 39). Epidermal cells possess prominent sinuous anticlinal margins (Fig. 43, 46), and an overall irregular, surface that may be covered with 2-3 small papillae ca. 12 /xm in diameter per cell (Fig. 45, 50). Epidermal cells are ca.45 fim wide x 23 /xm high and display the sinuous margin except for cells overlying the vascular strands, here the anticlinal walls are less sinuous (Fig. 41).

As in fi. schopfii. £. skaarensis leaves are hypostomatic. Stomata are randomly distributed on the abaxial leaf surface in a density of 90.6/mm]2 ] (Fig. 47). Stomata are surrounded by a ring of 5-6 subsidiary cells that range in shape from blunt, rectangular cells 40 x 28 /xm, to more elongate cells with sinuous walls which are 59 x 30 /xm (Fig. 44-46, 48-50). Subsidiary cells have beak-like papillae 12 x 7 /xm that surround the sunken opening of the stoma (Fig. 44, 49). Stomatal complexes may be contiguous such that a given subsidiary cell may be shared by more than one stoma. Guard cells are 34 x 9 /xm, and have thickened margins (Fig. 42, 48). They surround a stomatal opening ca. 9 /xm long.
In spite of the poor preservation of specimens, a considerable amount of information is available from twigs bearing small, scale-like leaves and twelve specimens of larger, woody, branching stems, some with leaf bases and extraxylary tissues (Fig. 51-56). The identity of these specimens with G. skaarensis is established on the basis of anatomical features of the small leaves on twigs, and of leaf traces and cortical histology of larger stems. The most extensive twig is sectioned at the level of the stem axis and bears 24 helically arranged leaves (Fig. 54, 56). The axis is 1.2 mm in diameter at the most proximal level and has a stele ca. 54 /xm in diameter with a broad, hollow pith 34 /xm across, surrounded by a ring of poorly defined

vascular bundles. The stele, in turn, is surrounded by a partially preserved, parenchymatous cortex that extends into lobe-like leaf bases (Fig. 54, 56). The twig exhibits a phyllotaxy that approaches 2/5, as determined by the relative positions of individual diverging leaves (Fig. 56 a-d). The stele is too poorly preserved to allow for the interpretation of primary vascular architecture, but at most levels 5-9 poorly defined, individual bundles are present.
In this species the cortex is composed entirely of primary tissues. Typically the outer portion of the cortex is separated from the stele and no central cortical tissue is preserved. However, in the most complete sections, the cortex has two zones, an inner zone composed of larger, radially elongate parenchymatous cells, and an outer zone of smaller cells adhering to the epidermis.
Twigs of fi. skaarensis are characterized by small, scale-like leaves up to 540 pm long that appear first as lobe-like leaf bases mm extending from the stem margin (Fig. 52, 54, 55, 56). Distal to the point of attachment they are oval-triangular in transverse section (Fig. 54,55, 56). Leaves overlap slightly, but do not encircle one another to the extent of the larger G. schopfii leaves.They are characterized by a prominent midrib area up to 300 pm across that contains a lacuna, representing the position of the midrib (Fig. 55). Lateral laminae are much

51thinner. Histologically, mesophyll is poorly preserved but appears simple and parenchymatous, lacking palisade and spongy layers. In all of these respects, the small, scale-like leaves of this type bear a resemblance to mature leaves of £. skaarensis (Fig. 34-37, 52-56).
Larger stems of 6. skaarensis are up to 2.3 cm in diameter and typically possess a broad pith up to 1.1 mm across (Fig. 51, 53). The pith may be hollow, parenchymatous, or irregularly septate, and may contain cells and/or an area of tissue with dark, amorphous, apparently ergastic substances (Fig. 51, 53). Typically, steles are ca. 1 cm in diameter. Primary vascular strands are difficult to distinguish in transverse section (Fig.51, 53), on the basis of oblique longitudinal sections a marked change in tracheid size and pitting pattern from inner to outer stelar margin can be recognized, presumably reflecting the position of the primary bundles. Protoxylem strands are characterized by helical wall thickening patterns, while metaxylem tracheids have helical or scalariform pitting patterns.
Outside the primary body is a zone of secondary xylem up to ca. 6 mm thick. The wood may appear as a continuous cylinder (Fig. 53) or, at other levels, be dissected radially by the divergence of 5-8 leaf traces (Fig. 51). Woody cylinders usually are either entire or dissected by at least 5-8 leaf traces, rather than only one or two

52traces. Secondary xylem is of the Araucarloxvlon type (Maheshwari, 1972; Stubblefield and Taylor, 1986) and contains numerous uniseriate, parenchymatous rays, from one to a few cells high. Tracheids possess circular bordered pits on their radial, but not tangential walls.
Outside of the secondary xylem, some stems possess a region of cortical tissue up to 5 mm thick (Fig. 53). Cortical tissue is typically parenchymatous, sometimes almost aerenchymatous and may have a similar zonation as that of the small twigs, with an inner zone of larger-celled parenchymatous cells, and a zone of smaller, more compact cells to the periphery (Fig. 53, at right).
Leaf bases attached to the stem extend as lobes of cortex. Vascular strands are initially double with vascular strands from adjacent sympodia joining to form the double leaf trace. These subsequently bifurcate, providing the base of the petiole with four strands (Fig. 53).Distal to its level of attachment the petiole base is 1.5 x .58 mm and oval to triangular in shape (Fig. 37, 53).

CHAPTER V - DISCUSSION
Generic concept of Glossopteris.— Brongniart (1828) established the genus Glossopteris as a section of Filicites (Maheshwari, 1965) to encompass impressions of fossil leaves with entire margins, a "persistent" midrib, and reticulate venation (Arber, 1905? Kovdcs-Endrody,1981). Glossopteris is distinguished from the similar Permian leaf genera Ganaamopterisf Rubidgea,Palaeovittaria. Eurvphvllum. Rhabdotaenia. and Belemnopteris based on a combination of morphological features (Maheshwari, 1965; Pant, 1982; Kovdcs-Endrddy, 1987). While Glossopteris and Rhabdotaenia are characterized by a single persistent midrib, and Belemnopteris by one that is tripartate, Ganaamopteris. Rubidaea. and Eurvphvllum are said to lack a midrib, and in Palaeovittaria the midrib is "partial" (Srivastava, 1956; Surange, 1966; Pant, 1982). Glossopteris. Ganaamopteris and Belemnopteris typically possess cross connections or reticulations between veins, while Rhabdotaenia. Palaeovittaria. Rubidaea and Eurvphvllum lack this feature. An additional distinction occurs in the angle of lateral veins with respect to the midrib. In Rhabdotaenia lateral
53

veins extend from the midrib at right angles, while in Palaeovittaria. Rubidaea and Euryphvllum veins produce an acute angle. Additionally, angles are arched in Rubidaea and Eurvphvllum. but straight in Palaeovittaria. Although it is pinnately compound, Pteronilssonia is included in glossopterid discussions by some authors because it has venation like that of Rhabdotaenia and cuticular structure like that of glossopterids (Pant, 1982). In addition to reticulate venation and a prominent midrib, Belemnopteris is characterized by a sagittate leaf base with two distinctive lateral flanges, each of which is vascularized by one strand of the tripartate midrib (Lacey, et al., 1975; Pant and Choudhury, 1977).
Several authors (e.g., Plumstead, 1973; Lele, 1976; Rigby, 1984) have suggested that a general trend exists in the stratigraphic distribution of the glossopterid genera, such that leaves lacking a midrib (i.e., Ganaamopteris. Rubidaea. Eurvphvllum. and Noeaaerathiopsis) are confined to the Lower Permian, while those with a midrib (Glossopteris. Rhabdotaenia. and Palaeovittaria) occur in Upper Permian strata (Chaloner and Lacey, 1973; Lele,1976). This distribution has been interpreted as reflecting both the evolution within the group (e.g., Meyen, 1971; Bernardes-de-Oliveira, 1978; Rigby, 1984) and a major climactic change, presumably from the cool temperate postglacial Lower Permian, to warm, temperate

55 -conditions in the Upper Permian (Surange, 1975; Lele,1976).
The largest and most widespread glossopterid genera are represented by Glossopteris and Ganqamopteris. Through the years, particular attention has centered around the comparison of, and the difficulty in distinguishing between, these two genera (Haithy, 1965; Maheshwari, 1965; Rigby, 1967, 1984; Pant and Gupta, 1968, 1971; Pant and Singh, 1968; Pant and Choudhury, 1977; Lele, 1976; Kovdcs-Endrody, 1977-1978). The two forms were originally separated by the presence (Glossopteris! or absence (Ganqamopteris) of a midrib (McCoy, 1874-76). They also differ in stratigraphic range: Ganqamopteris is limited tothe Lower Permian (=Talchir and Karharbari Stages) of India (Surange, 1975) and equivalent strata of South Africa (Lacey, et al., 1975), Antarctica (Schopf, 1976), and Australia (Rigby, 1967, 1984), and either rare or absent in the Upper Permian (Maithy, 1965). In contrast, leaves assignable to Glossopteris appear later in the fossil record (e.g., Karharbari of India, Surange, 1975) and increase in diversity until a maximum radiation is reached in the Upper Permian (e.g., Raniganj Stage, Surange, 1975). Although Ganqamopteris has been described from younger strata these reports have mostly received little support.For example, Maithy (1965) and Maheshwari (1965) consider

56that specimens described from the Raniganj as Ganqamopteris should be reassigned to Glossopteris (Surange, 1975).
Much of the difficulty in distinguishing Glossopteris and Ganaamopteris. and in determining if they should be recognized as separate genera, is related to the concept of a midrib and its interpretation in glossopterid leaves. Although both Seward (Seward and Woodward, 1905; Seward, 1910) and Arber (1902, 1905) had difficulty in accepting this generic distinction, and considered merging the two taxa, they made no formal taxonomic changes. Years later, because of the ambiguities involved in interpreting the midrib, Kovacs-Endrody (1977-1978) merged the two genera under the name Glossopteris. which has priority. Although many authors readily acknowledge morphological intergradation between Ganqamopteris and Glossopteris. this taxonomic emendation is generally not accepted, and most workers have retained the two taxa (e.g., Pant and Singh, 1968? Rigby, 1978).
In specimens of Glossopteris where sufficient morphological detail of the venation pattern can be resolved, veins of the midrib usually lack interreticulations (e.g., Pant and Gupta, 1968), although rarely they may be present (e.g., G. stricta. G. fibrosa: Arber, 1905; Pant, 1958). In some genera the midrib persists to the apex, while in others, veins in the central area of the leaf tend to anastomose near the apex (e.g.,

57Fig. 5). Some authors have interpreted the evanescence of the midrib in the apical portion of the leaf as a "partial midrib": evidence of an intergradation betweenGlossopteris and Ganaamopteris (e.g., Pant and Gupta,1968). In other cases, this configuration has been accepted as part of the natural variation within Glossopteris (e.g., Archangelsky, 1957).
In contrast, the central part of the leaf in Ganaamopteris is vascularized by veins that either radiate from the base of the leaf in a fanlike arrangement, or, in other species, by veins that produce a pattern of more parallel, longitudinally oriented, meshes with interreticulations (Maithy, 1965; Pant and Singh, 1971; 1974; Pant and Gupta, 1968, 1971). This second arrangement results in a configuration that superficially simulates a midrib (Maithy, 1965; Pant and Gupta, 1968; Pant and Singh, 1971). Leaf meshes in Ganaamopteris tend to be uniform in size and shape in comparison to the more variable meshes of Glossopteris. Lastly, although cuticular features are similar in both genera, the orientation of stomata in parallel rows is said to occur more commonly in Ganaamopteris (Pant and Gupta, 1968).
The distinction between Glossopteris and Ganaamopteris based on one feature, the presence or absence of a midrib, has disturbed a number of researchers, not only because of the limitations of single character taxonomy, but also

because the character in question is so ambiguous. Both Arber (1905), and Seward (Seaward and Woodward, 1905; Seward, 1910) were reluctant to use the midrib as a major character at the generic level. For example, Seward pointed out that in some extant ferns (e.g.,Scolopendriuml, midribs are inconsistent, while Arber (1902) remarked on the absence of well-defined midribs in the scale leaves of Glossopteris. In order to extend beyond this point, a number of authors have attempted to define their criteria for the midrib more precisely (e.g., Pant and Gupta, 1968; Pant and Singh, 1971, 1974).
Botanically, a midrib is defined as "the principal nerve in a leaf" (Jackson, 1928; Pant and Gupta, 1968), or, by another authority as "the main or central rib or vein of a leaf " (Little and Jones, 1980). In most living plants, the midrib consists of an "abruptly thickened median longitudinal ridge" that may have either one or more than one vascular strand (Pant and Gupta, 1968). In this context, a midrib is construed as a broadly defined region of the leaf, the entire configuration of which results from a combination of an increase in laminar thickness and in size of the midvein or midveins. In her study of Glossopteris and Ganqamopteris leaves, Kovdcs-Endrody (1977-1978) emphasized that the midrib of Glossopteris is composed of several parallel veins, rather than a single vein, and suggested that it therefore does not represent a

59true midrib. In this discussion, Kovdcs-Endrddy (1977-1978) thus restricted her concept of a midrib to a structure consisting of a single vein. Since glossopterids have more than one vein in their midribs, this restricted definition precludes the possibility of a bone fide midrib in any of the glossopterids.
In evaluating criteria more closely, Pant and Gupta (1968) listed six features that characterize a midrib.These include: 1) abruptly thicker lamina in the median area of the leaf; 2) a fold or groove in the lamina? 3) more parallel strands in the median area than laterally; 4) larger angles between secondary veins and midrib; 5) lack of cross connections; and 6) cuticular differences in the central region of the leaf. These authors stressed that although one or several of these characters may be absent, taxonomic value should be placed on a suite of features that together define the midrib, rather than on only the presence of absence of the midrib.
With the availability of anatomically preserved glossopterid leaves, evidence concerning the nature of the midrib can now be studied at four levels of resolution: the gross morphological (superficial) appearance, the venation pattern, cuticular detail, and the level of anatomical detail, as illustrated in £. schopfii and G. skaarensis. While features observed from specimens showing only gross morphology may be somewhat obscure and

6 0
ambiguous, anatomical preservation and cuticular features provide information with a higher degree of resolution. It is now apparent from the study of anatomically preserved leaves, which illustrate all four levels of resolution, that the glossopterid midrib may be the result of several, both real (anatomically based) and superficial, factors.
In terms of anatomically based features, in both <£. schopfii and G. skaarensis the larger "midrib" is the result of a combination of larger veins and an increase in leaf thickness. This occurs more markedly in G. skaarensis where there is an inflated appearance to the region of the midrib in transverse section (Fig. 35), but no other particular, specialized anatomical structure exists in the central region of leaves of this type. In contrast, Gould and Delevoryas (1977), suggest that in putative glossopterid leaves from petrified Australian peat, a thick hypodermis is present, particularly in the region of the midrib, and the presence of this structure results in an increased prominence of the midrib. A similar hypodermal thickening below the upper epidermis, has been suggested by Meyen (1971) for leaves of Tartarina. a peltaspermaceous foliage type from Angara previously thought to be related to Glossopteris (Zimina, 1967, 1977). While some of the figured specimens of Gould and Delevoryas (1977, i.e., Fig. 2E, F, G) resemble those of G. skaarensis in features of venation and hypodermis, the hypodermis in G. skaarensis is

61continuous throughout the width of the leaf. Since the intergradation between central and lateral parts of the Australian leaves is not figured (Gould and Delevoryas,1977) critical comparisons between the two taxa cannot yet be made.
In more superficial terms, the external appearance of weathered glossopterid specimens with the same type of anatomy may give the appearance of either a prominent (Fig. 3, 29) or more delicate (Fig. 4, 31) midrib. In part, this variation, which occurs in both Antarctic species, is the direct result of whether the entire leaf is fractured in the same plane or whether the midrib area has been fractured at a different (more external) level. Presumably this same type of contrast might occur among compression/impression forms. This presence, absence, or "robustness" of a midrib, based on external shape alone, without detail of venation, should be viewed with caution. However, within species of Glossopteris. Banerjee (1978) has used the width of the midrib as a specific character (expressed as percentage of leaf width), and, if measured consistently, this may be a more reliable character than qualitative descriptions such as "prominent, robust, massive", or "ephemeral".
For several reasons, both morphological and pragmatic, the present author concludes that the continued recognition of Glossopteris and Ganaamopteris as separate

taxa is warranted. Although the venation patterns are intergradational, a critical evaluation of the midrib based on several parameters does allow for distinguishing the two taxa, at least most of the time. Furthermore, stratigraphic occurrences have on the whole been consistent. Some researchers have argued the question from the other side, indicating that although intergradations occur between the two taxa, Kovdcs-Endrddy's (1977-1978) all-inclusive synonymy creates an unwieldy and meaningless artificial taxon that aids little in biostratigraphic and paleogeographic concepts (Rigby, 1978). From the available evidence, it seems that Ganqamopteris represents a smaller, more cohesive group of related forms that is intergradational with, and perhaps eclipsed by, the greater diversity of Glossopteris. Presumably, if the two leaf types are related, and occur in stratigraphically sequential order, intermediate forms may be present. Continued critical evaluation of the subfeatures that comprise the midrib, as well as the search for other distinctive features of the leaves, may eventually allow for a more complete understanding of the problem.
Mixed floras: Since glossopterid leaves comprise themajor components of the Permian Gondwana flora, this floristic province is usually referred to as the "Glossopteris flora" (e.g., Arber, 1905; Chaloner and Lacey, 1973; Schopf and Askin, 1980). However, additional

occurrences of leaves with a glossopterid venation pattern have been reported in other geologic levels and geographic regions. Leaves found in geographical and/or stratigraphical settings other than Permian Gondwana, that have been compared with Glossopteris include occurrences in Mexico (Delevoryas, 1969; Delevoryas and Person, 1975; Ash, 1981), Honduras (Ash, 1979-1980); Poland (Bochenski, 1957), the Phetchabun flora of Thailand (Kon'no, 1965; Asama,1969), and the Mesozoic of Greenland and Tonkin (Harris, 1932; Chaloner and Lacey, 1973). In some cases, fossils are too poorly preserved to evaluate with certainty, and reports of this type are generally not widely accepted. However, in other situations, morphology clearly resembles that of Glossopteris in great detail, causing considerable taxonomic and biogeographic confusion.
Of these latter cases, perhaps the most striking resemblance to Glossopteris occurs in Mexialossa. a genus of leaves from Oaxaca, Mexico, which occur in an otherwise typical Jurassic flora (Delevoryas, 1969; Delevoryas and Person, 1975). Six distinctive forms, along with several intergradational specimens, have been described, four of which are compared directly with particular species of Glossopteris (i.e., G. indica. G. browniana. G. taeniopteroides. and G. eurvneura: Delevoryas and Person, 1975). The genus Mexiolossa was established for these leaves since it was unclear what affinities the Mexican

6 4
leaves may have with Gondwana forms, and since leaves of this type are found in association with an unusual fructification (Eerezlaria) which may be related (Delevoryas and Gould, 1971).
A second type of unusual occurrence of Glossopteris leaves is in Permian strata of floristic provinces outside Gondwana. Vegetative leaves similar to Glossopteris have been reported from Angara (Meyen, 1969, 1971, 1977) and Cathaysia (Kon'no, 1965; Asama, 1969). Perhaps the most historically important of these was originally reported by Amlitzky (Arber, 1905; Meyen, 1969, 1971, 1977).Discoveries of this sort were further illustrated by Zalessky (1933) who placed Glossopteris-like leaves in his genus Pursonqia (Meyen, 1977). Leaves of this type lacked cuticle, but the discovery in the same beds of similar leaves with well preserved cuticle led Meyen (1971) to propose the genus Tartarina. Cuticular structure of Tartarina allied these forms with the peltasperms, as did subsequent discoveries of peltaspermaceous ovulate fructifications (Meyen, 1977). Although the Tartarina specimens can be linked with peltasperms and not glossopterids, Meyen (1969, 1971, 1977) suggested that certain isolated leaves in Angara could still be assigned to both Ganqamopteris and Glossopteris (see Meyen, 1969). However, he stressed that the claims are based on only poorly preserved vegetative leaves and there are no

65associated fertile remains present that might suggest affinities of these plants to the southern hemisphere glossopterids. Meyen (1977) also remarked that the occurrence of glossopterid-1ike leaves in Angara are a minor component of an otherwise well diversified flora containing characteristic Angara species.
A second floristic province associated with unusual Glossopteris occurrence is the Cathaysian Giaantooteris flora (Asama, 1969, 1976; Li and Yao, 1985) . Gicrantopteris leaves are a highly diverse group of broad leaves, typically of large size, with reticulate venation.Although they occur primarily in the Cathaysian flora/province, gigantopterids have also been reported from North America (Mamay, 1960; Mamay, Miller and Rohr, 1984; Mamay, Miller, Rohr and Stein, 1986). They differ from Glossopteris in having up to four orders of reticulate venation (Asama, 1976). Ovule-bearing leaves, known under the genus Giaantonomia. bear elliptical seed in two rows along the abaxial leaf surface at right angles to the midrib, while pollen-bearing units known as Gigantotheca (Li and Yao, 1985). Within the Cathaysian flora, there have also been reported a few specimens similar to Glossopteris. However, as in the Angara flora, they represent a minor component of an otherwise highly distinctive provincial flora in most of Cathaysia (Asama, 1969; Hsii, 1976, 1979). Recently, however, several species

66of Glossopteris have been reported from a Tibetan locality that suggest the occurrence of a true Gondwana flora in southern Cathaysian provinces (Li and Yao, 1985).
In addition, geographically within Gondwana, specimens of Glossopteris have been found as components of Triassic floras in India (e.g., Feistmantel, 1880a, 1881, 1886; Lele, 1955, 1962; Banerji and Bose, 1975; Bose, Banerji, and Maithy, 1975; Pal, 1984; Pant and Pant, 1987), Tasmania (Retallack, 1980); Antarctica (Rigby and Schopf,1969), and the Molteno Beds of South Africa (Thomas, 1952, 1958; Anderson and Anderson, 1983). Perhaps the most extensive of these occurrences is at Nidhpuri, India, where 25 species of Glossopteris have been reported (Pant and Pant, 1987). Although Pant and Pant (1987) question the Triassic age of some of these beds, the co-occurrence of numerous Glossopteris and Dicroidium specimens in the same strata at this locality and at others (e.g., Allan Nunatak, Antarctica, Rigby and Schopf, 1969; Molteno Beds, South Africa, Anderson and Anderson, 1985) suggest a gradual, transitional nature of the Permian/Triassic boundary in at least some regions of Gondwana.
Lastly, a different type of unusual floristic distribution occurs among the Glossopteris floras: thepresence of "northern elements" or "mixed floras" (Chaloner and Lacey, 1973; Archangelsky and Arrondo, 1967; Lele,1974). Some of these floras occur within Gondwana, yet

contain glossopterids along with certain "northern elements" particularly pteridophytes, which were presumably introduced into Gondwana. Here, northern hemisphere genera such as Pecopteris. Annularia, and Sphenophvllum. occur in southern hemisphere localities (Archangelsky and Arrondo, 1969; Chaloner and Lacey, 1973). It is interesting that South American and South African localities where these mixtures occur are near the limits of glacial boundaries (Chaloner and Lacey, 1973).
Some authors (e.g., Archangelsky and Arrondo, 1969; Chaloner and Lacey, 1973; Rigby, 1984) have pointed out that the "northern elements" present in these localities are generally pteridophytes. The suggestion has been offered that the strong pteridophyte bias of the northern elements reflects differential migrational ability of spores vs. seeds as dispersal units. An alternative view regarding the preponderance of northern hemisphere pteridophytes is that Gondwana seed plants were evolving more rapidly under stronger selective pressures while pteridophytes were undergoing a period of stasis. An equally plausible possibility is that both groups were undergoing a radiation in response to the changing, but morphological changes in pteridophytic forms were more subtle. In some cases, plants previously recognized as "northern elements" have been reinterpreted in other contexts as southern hemisphere forms (e.g., Rigby, 1984).

68In other cases, true northern taxa may be present within the Gondwana province, whether through migration or the continuation of prominent Lower Carboniferous ancestral forms (Chaloner and Lacey, 1973).
Still other localities contain a flora that may represent a true mixture of floristic realms because they occur near geographic boundaries between paleofloristic zones. The Hazro flora of Turkey represents an example of a flora of this type with mixed Gondwana and Angara elements (Wagner, 1962; Archangelsky and Wagner, 1983). Equally enigmatic is the mixed Cathaysian/Gondwana flora of New Guinea (Kon'no, 1965; Chaloner and Lacey, 1973). These two floras are particularly interesting because of their apparent isolation from any intermediate floras that would link them more closely with either floristic province (Chaloner and Lacey, 1973).
These various types of mixed floras have often been confused with one another and, moreover, authors have disagreed about their significance. It is necessary to regard such terms as "mixed floras" and "northern elements" with caution, and to evaluate each situation individually, with attention to paleogeographic constraints (Chaloner and Lacey, 1973), since they may represent different types of occurrences. True mixed floras may be the result of either migration (Just, 1952), the mixture of northern and Gondwana forms along boundaries (Wagner, 1962), parallelism

6 9
(=homoplasy, Chaloner.and Lacey, 1973? Archangelsky and Arrondo, 1969) and/or any combination of these effects.
Species of Glossopteris.— As a whole, glossopterid leaves are found as disarticulated fragments, and their classification has focused on morphological details of the leaves themselves. This has resulted in a highly artificial taxonomic scheme (e. g., Surange, 1966; Chandra and Surange, 1979). In only a few instances have vegetative leaves been found organically attached to stems (Pant and Singh, 1974), and even less frequently are fertile parts known in attachment (e.g., Pant and Singh, 1974; White, 1978). Leaves assignable to Glossopteris exhibit a wide range of diversity, including size (from 2 cm in length, e.g., [G. taeniensis] to over 20 cm, [e.g.,G. amplal). shape, base, apex, width and persistence of midrib, and venation pattern (e.g., Maheshwari, 1965? Surange, 1975).
Unfortunately, the morphological variation needed to distinguish vegetative leaves has been difficult to assess. The continuous intergradation of some features (e.g., size, shape and venation pattern), has been difficult to categorize, while discontinuous (qualitative) characters (e.g., shape of base or apex), are often only rarely present. This difficulty in species delimitation is further compounded by the contrasting approaches and differing motivations of taxonomists. One group of workers

(e.g., Feistmantel, 1880a, b) has taken a narrow approach, circumscribing species based on one to a few specimens from a single locality, and emphasizing subtle differences. In some of these studies, biostratigraphy has been the motivating factor for defining species (e.g., Chandra and Surange, 1979: Rigby, 1983). Approaches that emphasize vegetative or anatomic fine detail and stratigraphic occurrence (e.g., Pant and Singh, 1971) obviously multiply taxa, sometimes increasing their value as index fossils, but may not address questions of biological and intraspecific variation.
In contrast, a second group of researchers (following the approach of Arber, 1905) have been more broad-based in their taxonomy (Seward, 1897, 1904, 1910; Walkom, 1922). These researchers recognized species based on broader morphological criteria, which encompass what they considered to be natural biological variation. Unfortunately, some of these studies (e.g., Arber, 1905) have tended to combine several morphologically distinct forms into groups that have become so inclusive that they are difficult to define.
Additionally, much confusion has arisen since many early species were described without the designation of a holotype (Banerjee and Ghosh, 1970). Revisionary studies (e.g., Banerjee and Ghosh, 1970; Kovacs-Endrody, 1976; Banerjee, 1978; Rigby, et al., 1980) have greatly aided in

71the clarification of certain characters. Here, lectotypes have been designated for major species and some features (e.g., midrib, vein density) have been quantified. This type of data provides the initial basis from which well delimited species can subsequently be recognized.
Systematic approaches that describe, group or quantify the overall diversity within a genus, or possible relationships of individual species to one another, have not been attempted beyond the more generalized concepts (viz., large vs. medium or narrow-meshed vs. broad-meshed). Although some authors (Chandra and Surange, 1979) have provided size-class designations (e.g., "small" = 1-3 cm) and employ a standardized morphological terminology (Dilcher, 1974; Chandra and Surange, 1979), the process of recognizing or identifying specimens is based on primitive picture-matching (Chandra and Surange, 1979; see Lacey, 1981). Although the fidelity of certain characters has been repeatedly discussed (e.g., Surange, 1966; Pant and Singh, 1971), species designations are based vaguely on descriptive characters which are not taxonomically diagnostic, and there is little agreement on what distinguishes a species of Glossopteris.
A second potential source of diagnostic information for Glossopteris has been explored in specimens from India, Tanganyika, Australia, and Brazil (Zeiller, 1896; Sahni, 1923; Srivastava, 1956; Srivastava and Chandra, 1956; Pant,

1958? Pant and Gupta, 1968, 1971; Pant and Singh, 1971, 1974? Rosier and Fittipaldi, 1981? Pant and Pant, 1987).The first cuticular studies of Glossopteris were of G. indica, as described by Zeiller (1896) and G. anaustifolia (Sahni, 1923). Later, Srivastava (1956) noted cuticular features for 14 species of Indian Glossopteris. 6 species of Ganaamopteris and one species of (the monospecific) Palaeovittaria. Based on this study, Surange and Srivastava (1956) grouped cuticular types into six groups delineated by a combination of the following features: thickness of cuticle, shape of epidermal cells, visibility of venation on the cuticle, distribution, density, and type of stomata, number of subsidiary cells, guard cell shape and thickenings, and presence of epidermal papillae These authors found that Feistmantel's narrowly-defined species, based only on morphological features, corresponded more closely to the cuticular types than did the more broadly defined taxa of Arber (1905). Furthermore, the six groups transgressed the generic bounds between Glossopteris. Ganaamopteris. and Palaeovittaria. with only one group being composed solely of Glossopteris. one of Ganaamopteris. three of both Glossopteris and Ganaamopteris, and one of Glossopteris and Palaeovittaria (Surange and Srivastava, 1956).
Pant (1958) described three additional species of Glossopteris from Tanganyika based on a similar array of

73cuticular features. In subsequent papers, Pant and Gupta (1968, 1971) and Pant and Singh (1971, 1974), described a total of 29 new species of Glossopteris and emended the generic diagnosis (Pant and Gupta, 1968) to include cuticular features. Pant and Singh (1971) provided a dichotomous taxonomic key of cuticular features for all species of Glossopteris with cuticle.
Unfortunately, cuticular studies have contributed little support in delineating species of Glossopteris. Similar cuticle have been described from leaves with very dissimilar venation patterns, and vice versa (Surange,1966). Little is known about the natural biological variability of glossopterid cuticle. Furthermore, although many features have been tabulated (e.g., Pant and Singh, 1971), there has been no critical evaluation of the reliability of features. In some instances it is unclear whether cuticle has been altered during preservation. Although numerous line diagrams are provided in these papers to illustrate cuticular features, many of the photographs of original material are poorly presented, precluding the picture-matching approach to taxonomy that is apparently expected.
Rather than successfully resolving the taxonomy of Glossopteris. the descriptions of cuticular features have further confused the issue. Furthermore, cuticular data has been incorporated into the taxonomy of Glossopteris in

several different ways. Several authors have suggested that species based on morphological features (e.g., venation pattern, size and shape of leaf) alone cannot be included in taxa based on additional cuticle features which they did not possess (e.g., Pant and Singh, 1974; Pant, 1977; Chandra and Surange, 1979). Some authors (e.g., Srivastava, 1956) have retained the original names for those species with cuticle. The situation is further complicated because contemporary workers have disagreed with the taxonomic assignments of several of Srivastava's specimens, and designated new taxa for them (e.g., Pant and Gupta, 1968, 1971; Pant and Singh, 1971). These authors(i.e., Pant, 1958; Pant and Gupta, 1968, 1971; Pant andSingh, 1971, 1974) have created many new species based oncuticular structure in conjunction with morphological features. In some instances, even when cuticular features are known, and even in cases where they represent the only means of distinguishing taxa, they have not always been included in descriptions or diagnoses (i.e., Chandra and Surange, 197?; see Lacey, 1981).
Although many compressions of Glossopteris have been described as containing "structure", "anatomy" or "fine structure" (e.g., Pant, 1958; Pant and Singh, 1971) these leaves lack the three-dimensional preservation which would illustrate the anatomical organization of the mesophyll and veins. Reports of this sort (e.g., Pant, 1958) do provide,

however, in addition to morphology and cuticular/epidermal detail, some detail about individual tracheary elements, or other cell types (e.g., "palisade" or "spongy mesophyll" cells and "fibers" of G. fibrosa and G. hispida. Pant,1958). Material of this type provided Pant (1977) with the impetus to offer a reconstruction of a Glossopteris vegetative leaf in transverse section (viz. Pant, 1977,Fig. 5 a&b). Interpretations of this type remain equivocal, since the three-dimensional, organizational detail was not available and the basis for interpreting mesophyll organization remains unclear.
Affinities of the Antarctic leaves.— The two species of glossopterid leaves described above, G. schopfii and G. skaarensis. represent the first species of Glossopteris for which anatomical features have been delimited. As such they provide a valuable data base from which to characterize structural features of leaves demonstrating the glossopterid venation form. Although the two species are of comparable size and have some similarities in their anatomy they are clearly distinct from one another, and, of the hundreds of specimens studied, no intergrading forms have been discovered.
The first form, G. schopfii. possesses the following distinguishing features: narrow, elongate meshes, arelatively constant laminar thickness in transverse section, elongate fibers in the bundle sheath, a prominent

lacuna in the position of the phloem, metaxylem tracheids characterized by scalariform wall thickenings, epidermal cells with straight-margined anticlinal walls, a stomatal complex with loosely-associated subsidiary cells that tend to be aligned parallel with the long axis of the leaf, and papillate epidermal cells. In contrast, G. skaarensis leaves have broad, polygonal meshes, a thick midrib and abruptly narrow lateral lamina in transverse section, distinctive hypodermis, metaxylem tracheids characterized by helical wall thickening patterns, epidermal cells with prominent sinuous anticlinal margins, irregularly distributed, sunken stomata with a well-defined ring of subsidiary cells possessing beak-like papillae, and a thick, irregular cuticle.
Both Antarctic leaf forms are furthermore distinct from the putative glossopterid leaves described from the Bowen Basin of Queensland, Australia (Gould and Delevoryas, 1977). The Australian leaves differ in possessing well-differentiated mesophyll with palisade and spongy layers (e.g., Fig. 2E, F, G, of Gould and Delevoryas,1977). Although the Australian leaves have yet to be fully characterized, they are reported to possess a distinctive hypodermis on both leaf surfaces that is confined to the region of the midrib. Illustrations (Fig. 2F of Gould and Delevoryas, 1977) further indicate that these leaves lack both a fibrous bundle sheath, like that of G. schopfii. or

77any apparent bundle sheath separating individual vascular strands.
From the published illustrations, some, but not all, of the Australian material, closely resembles specimens of G. skaarensis (compare Fig. 35, 37, 39 with Fig. 2A-C of Gould and Delevoryas, 1977). Since the transition or intergradation between the midrib area (Fig. 2 A-C of Gould and Delevoryas, 1977) and lateral areas of leaves (Fig. 2D-G) is not figured, it is difficult to envision the connection between these very different appearing structures. If Fig. 2A-C represents a leaf closely allied to G. skaarensis. Fig. 2D-G may represent another, less closely related form. Cuticular features of the Bowen Basin leaves are not known.
Thus, at the present time, three distinctive anatomical forms can be recognized for Glossopteris leaves. The suites of anatomical features that distinguish them do not appear to intergrade, based on presently available information. A better understanding of the relationship between these forms, as well as the possible intergradation of glossopterid leaves as a whole, awaits the discovery of additional anatomically preserved specimens.
Glossopteris schopfii and G. skaarensis can also be compared favorably to a number of previously described compression/impression taxa. As a small-medium leaf with narrow elongate meshes, Glossopteris schopfii resembles a

number of the "narrow-meshed" taxa, such as G. anoustifolia and related forms (Banerjee, 1978). The angle produced by the lateral veins with the midrib in both species is between 20-25°, and vein density (22-26/cm2) are comparable in the two forms. Since entire leaves of G. schopfii are not known, overall morphology cannot be compared to that typical of G. ancrustifolia, a linear-lanceolate form, but it is reasonable to suggest that G. schopfii could be of similar length. Glossopteris schopfii can also be compared favorably to at least some specimens designated G. browniana, particularly on the basis of venation (compare Fig. lc to Text-Fig. 2B of Banerji and Bose, 1975).However, leaves of G. browniana are generally much larger than the Antarctic leaves.
In contrast, G. skaarensis has a coarse-meshed venation pattern that is similar to G. conspicua. G. retifera. G. shirleyi. (Lacey, et al., 1975; Banerjee, 1978) and several species of the genus Belemnopteris (Lacey, et al., 1975). Coarse-meshed species of Glossopteris are fewer in number than the narrow meshed forms, and recently two of the most commonly reported taxa, G. retifera and G. conspicua. were synonomized by Rigby, et al., (1980), under the name G. conspicua. Among these taxa, the form G. shirlevi (Rigby,1978), differs from G. elonaata sensu Rigby et al. (1980) in having veins that arch laterally and producing more "D-shaped" meshes (Rigby, 1978). Meshes in G. skaarensis

79are more variable in size than the highly uniform meshes of G. retifera and other coarse-meshed forms, with marginal meshes becoming more elongate than central meshes (Fig.28c, f, 30, 33).
Although Belemnopteris is distinguished from Glossopteris on the basis of a tricostate venation and a sagittate leaf base (Lacey, et al., 1975; Pant and Choudhury, 1977? Rigby, 1978), the coarse venation and cuticular features such as sinuous cell margins, thick cuticle, and stomata with overarching papillae are similar to those in G. skaarensis (e.g., compare Fig. 42-45 with B. pellucida. Text-fig. 4B of Pant and Choudhury, 1977).Since the lower portions of leaves have not been found in G. skaarensis. it is conceivable that leaves of this sort could be belemnopterid rather than glossopterid. Belemnopteris has been reported from Antarctica, in the Permian compression floras of the Transantarctic Mountains (Schopf, 1970a).
Cuticular detail of G. schopfii and G. skaarensis is not as completely known as is that of some compression species (e.g., Pant and Singh, 1971? Pant and Pant, 1987). For this reason it is necessary to compare cuticular features in a more generalized way than that of previous authors. The characters listed in Table 3 represent an assemblage of the features most easily recognizable and comparable from study to study. These include: width and

8 0
length (if known) of leaf fragments; angle of lateral veins, vein density, whether leaves are amphi- or hypostomatic, broad vs. narrow-meshed venation, presence of papillae, number and arrangement of subsidiary cells, stomatal size (as indicated by length of stomatal pore or guard cell), shape of epidermal cells, stomatal density and arrangement, presence of overarching papillae, and whether stomata are sunken.
An overview of these cuticular features in glossopterids demonstrates that there are only a few close correlations to the distribution of characters (Tables 3-8). On the whole, suites of cuticular features do not appear to fall into easily delimited, discontinuous groups. Of the 71 species of cuticular remains surveyed (including species of Glossopteris. putative specimens of Ganaamopteris. Belemnopteris. and Palaeovittaria^, where indicated, roughly 2/3 (67%) have a narrow-meshed venation pattern like that of G. schopfii. G. anaustifolia. and G. browniana, while 30% possess the coarse-meshed form characterized by G. skaarensis. G. retifera and G. shirlevi (Table 4). It is unclear, with this and other features, whether these data reflect true diversity or the tendency for researchers to oversplit the narrow-meshed leaves relative to coarse-meshed glossopterids.
Glossopterid leaves have been described as hypostomatic 79% of the time, with ,,unistomatic,, referring

to a few instances where orientation of leaves could not be determined (Table 5). In the amphistomatic leaves described, the upper epidermis characteristically possesses considerably fewer stomata than the lower surface (e.g., Pant and Gupta, 1968). In at least 59% of leaves surveyed, epidermal cell walls had straight, rather than sinuous, margins (Table 6). This feature often intergrades along one leaf surface, with more elongate epidermal cells with straighter margins covering the midrib, than those of the mesh areas (e.g., Fig. 41, 43). However, in most cases, upper and lower epidermal cells were of the same type, with the same general types of intergradations occurring in both surfaces (Table 3, 6). Leaves with straight-margined epidermal cells predominate, both in narrow- and broad-meshed forms (Table 7), represented by 68% of the narrow-meshed forms and 61% of the broad-meshed taxa.Also, in the relationship of mesh shape and stomatal position, narrow-meshed forms predominate in both hypostomatic (65%) and amphistomatic (88%) leaves (Table 8).
Of the species surveyed, a large portion (52%) were characterized as having irregularly oriented stomata, while 18% had cuticles with contiguous stomata (Table 3).Papillae were restricted to the subsidiary cells in 27% of the cuticles, while they occurred commonly on epidermal cells in 28% of cuticles. When present, papillae could be

82represented by either a single median papilla per cell (26%), several small papillae per cell (16%), or both (11%). Overarching papillae occurred as extensions of subsidiary cells in 34% of the cuticles.
One correlation that did appear to be highly consistent was the presence of stomata described as "unspecialized", and stomata occurring in rows, only in cuticles with straight-margined epidermal cells (Table 3). This interrelationship may be in part a function of cell shape, since the presence of highly sinuous margins tends to result in irregularly oriented cells (e.g., Fig. 46,47), while straight-margined cells tend to be aligned to the longitudinal axis of the leaf (Fig. 18, 21). Stomata occurring in rows were described as, variously, irregularly- (6%), obliquely- (1%), and longitudinally oriented (3%). Ten percent of the species were described as having sunken stomata, and fibers occur on 10% of the leaves.
Florin (1933) believed that Glossopteris was a heterogeneous group, based on variation in cuticular structure of G. indica and G. anoustifolia. as described by Zeiller (1896) and Sahni (1923), respectively. In retrospect, these two reports alone did not provide an adequate sample from which to draw substantial conclusions, in particular because it is difficult to interpret the cuticular features of Zeiller's specimen from his figure

83(Fig. 13 of Zeiller, 1896). It is the opinion of the author that the rectangular cells he illustrates may represent the hypodermis rather than the epidermal or cuticular layer.
However, based on the currently available data (Table 3), cuticular features among glossopterid leaves do appear to be varied in an intergradational manner. As has been observed by many previous authors (e.g., Surange and Srivastava, 1956; Pant, 1977) cuticular structures alone cannot predict morphological features, nor, until variability can be better assessed, and characters better evaluated, should cuticle be regarded as the sole means for defining taxa.
Quantitative analysis of Glossopteris leaves.—Anatomically preserved Glossopteris provide a new,
previously unavailable type of data base from which to analyze glossopterid leaf structure. In the past, characters of Glossopteris have been difficult to delimit, due to their continuous, and ambiguous, nature. The two species from Skaar Ridge are clearly distinct based on anatomy, and at present, are believed to represent the only two glossopterid leaf forms in this low-diversity flora.For this reason, they provide the opportunity to determine the range of morphological variation present within an individual species, as defined by anatomy. No data are available that would determine whether leaves were borne by

84the same genetic individual or breeding population (i.e., equivalence with the paleodeme concept of Anderson (1987) is not implied); however, anatomy provides a much finer degree of resolution and considerably more information than was previously available from Glossopteris compressions and impressions.
Since the continuous nature of morphological features has been difficult to assess, an approach using quantitative methods was undertaken. A suite of easily measurable morphological features was scored for a representative group of leaves of the G. schopfii and G. skaarensis types (Table 1). Although in some specimens, cuticular features were preserved, many of the specimens showing the best morphological surface features (e.g., Fig. 2-5, 29, 30) are highly weathered, possessing little organic material and little or no cuticle. While multiple measurements were available for some characters (i.e.,Table 1, features 14-17), a very limited number of measurements were possible for other parameters (i.e.,Table 1, features 1-13). This second group of parameters are based on a single measurement per leaf specimen, might unrealistically weight the data and provide spurious results. For this reason, three subsets of the data were analyzed, in order to determine in a preliminary manner, the way to get the "best data".

A quantitative analysis was undertaken, using the multivariate statistical procedures of stepwise and multiple discriminate analysis (Statistical Analysis Systems, 1985). In the first step of the analysis, to determine if a quantitative, objective scheme for differentiating G. schopfii and G. skaarensis could be developed, a Stepwise Discriminant Analysis was performed.A total of 426 observations were measured for 10 and 7 specimens of the two taxa, respectively, using the seventeen variables listed in Table 1. This analysis produced a sixteen variable equation for discriminating between these two taxa which accounted for 99.0% of the variation in those variables. Of the seventeen initial variables, only "width" (= width of leaf) was not useful in discriminating between the two species.
Although this sixteen variable model had considerable power, a five variable model based on the variables "innervein, marginal vein, Y-shaped reticulations, W-shaped reticulations, and total reticulations" alone could also account for over 90% of the variation model based on the variables. Moreover, the single variable "innervein" (=density, number of veins/cm2 in inner region of leaf), could account for 46.7% of the variation between G. schopfii and G. skaarensis. Thus, the other 11 variables added relatively little to the power of the discriminant function.

86When the sixteen variable function was then used in a
Multiple Discriminate Analysis to classify the 426 observations into either of the two species (G. schopfii or G. skaarensis), it did so with 100% accuracy. That is, the sixteen variable discriminant function could classify observations into G. schopfii or G. skaarensis without error. The Multiple Discriminate Analysis was then repeated using the five variable model described above.This smaller, more parsimonious discriminant function was also able to classify the 426 observations into the correct species with 100% accuracy. Thus, the five variable model was as effective as a classification tool as was the full sixteen variable model. This indicates that a discriminant functions with as much taxonomic power as a sixteen variable model could be developed for these taxa, at least, with only five measurements per sample. This smaller number of measurements would thus cut the time and effort needed to correctly classify these observations by about 2/3.
With the realization that features 1-13 (Table 1) were weighted heavily in comparison to features 14-17, for which multiple measurements were available, the procedure was repeated, using only features 14-17 in the analysis. (Additionally, the characters "total vein density" and "midrib" (features 1, 2, Table 3), were omitted from all later analyses). Of the four parameters used (i.e., mesh

87length, width, angle of lateral veins with midrib, and mesh area) (Table 9), two (i.e., mesh length and angle of lateral veins with midrib) do not overlap in their ranges. Mesh width overlaps to some extent, while mesh areas are remarkably similar in the two taxa (Table 9).
The results of this analysis were that leaves were still classified into their predicted groups based on anatomy approximately 85% of the time (Table 10).Inspection of the 23 individual results that had a less than 20% chance of predicted identification revealed that the majority of misclassifications could be accounted for by a combination of unusual mesh length and angle in individual meshes.
This analysis sequence was then repeated for the data from 58 Indian taxa, with data taken from line diagrams of Chandra and Surange (1979) (Table 2). This data set presented greater analytical problems as it was much richer in putative taxa but had far fewer observations per taxon. In the first analysis, most of the leaves were classified as predicted, due to the weighting of characters 3-14. In a second analysis of only characters 15-17, there was little correlation of individual measurements with their predicted groups, with the measurements of a given specimen being classified into numerous individual taxa. In the third analysis, using only the four continuous variables, classification became very complex, with numerous

88misclassifications occurring in almost every taxon. In this case, resolution was so poor that the original Indian "species" had little meaning.
Lastly, in order to incorporate the data that were ultimately deleted from the multiple discriminate analyses of both the Antarctic data and the Indian data (i.e., features 3-14), a cluster analysis was performed, using these individual measurements, plus the means and values for features 15-17 (Fig. 57). Although this last method lays no claim to statistical validity, it was done as a preliminary attempt to assess variability present from specimen to specimen.
Results of the cluster analysis were difficult to interpret with confidence (Fig. 57). However, although the nature of the data base limited the amount of useful information, several general trends were apparent (Fig.57). Perhaps most obviously, the Antarctic leaves clustered apart from the Indian forms (Fig. 58, right).This may be in part the result of the differences in sampling techniques, although attempts were made to minimize these affects. This clustering also suggests the hypothesis that characteristic variation in glossopterids may occur in a geographic and/or stratigraphic context.With continued sampling of a large number of glossopterid leaves from throughout Gondwana, along with leaves showing the glossopterid venation structure (e.g., Mexiclossa.

89Delevoryas and Person, 1975), it may be possible to identify certain climatic, ecologic and/or stratigraphic differences among glossopterids in a manner analogous to the variation seen in angiosperms both stratigraphically (e.g., Upchurch and Wolfe, 1987; Wolfe and Upchurch, 1987) and climactically (e.g., Dolph and Dilcher, 1979).
Secondly, even within the constraints of the sample design, the general pattern was one of large groups of putative species clustering (Fig. 57). Considering that many of the species of Glossopteris from India are delimited solely on cuticular, rather than morphological features (Chandra and Surange, 1979), these results are not unexpected. It is apparent that samples in this analysis become either distinct or clustered abruptly, i.e., there is little gradation and/or resolution from which to determine, for instance, a list of valuable taxonomic characters which in combination delimit species easily. It may be that refinements of the sampling technique, in particular, the sampling of many individuals, would create an index by which to better establish salient features of these taxa.
Significant conclusions from these analyses include: 1) it is possible, with a high degree of accuracy (85%) to separate individuals of the species G. schopfii and G. skaarensis on the basis of continuously variable characters of leaf morphology (features 14-17) alone? 2) variability

within the species G. schopfii and G. skaarensis is appreciably less than variability between the two species; 3) if G. schopfii and G. skaarensis were found together as compression/impression taxa, it would be possible, with a high degree of accuracy, to predict to which of these two taxa a specimen belonged; 4) either Glossooteris schopfii or G. skaarensis might serve as hypothesis-generating data bases of the amount of variation one might expect within a species of Glossooteris (However, the degree of intraspecific variation may not be relevant to other taxonomic problems. These two taxa happen to be discontinuous in their suites of morphological features. Other more closely related taxa demonstrating more continuous, intergrading variation could certainly exist); 5) using the data base of variation within a species of G. schopfii and or G. skaarensis. one might establish some type of index for determining whether the degree of variation within another presumed species of Glossooteris based on an assemblage of compressions, should be admissible; 6) with reference to the Indian leaves, here the attempt is to determine whether an assemblage of morphological features might be characteristic for a given taxon, and if so, what features or combinations of characters are most valuable as species "fingerprints". Since the variability among glossopterid leaves as a whole has never been adequately assessed, it is difficult to

determine which features are inherent components of the glossopterid structure and which represent variation within this form. Ultimately, with the addition of representatives from differing stratigraphic and geographic positions, it may be possible to determine whether glossopterid variation reflects environment (e.g., climate, depositional environment, latitude) in Gondwana in a way comparable to that of early angiospermous leaves (e. g., Wolfe and Upchurch, 1987).
Several previous authors have attempted to study Glossooteris through the quantification of a suite of morphological and cuticular features. In many contributions (e.g., Srivastava, 1956? Pant and Gupta,1968; Pant and Singh, 1971), features such as vein density, angle of secondary veins with midrib, stomatal index, and cuticular thickness have often been expressed in a quantified way, but data of this type have never been analyzed or plotted to see what multivariate diversity may exist. Various authors have championed the importance of a particular character or suite of characters, such as Melville (1969) who cataloged types of interconnection (cross-connections) between veins, but no study has attempted to look at an integrated suite of such features. Unfortunately, whether previous studies have taken into account many features or quantified a very few, to my knowledge no study has attempted to synthesize or analyze

92such qualitative and quantitative data to gather an idea of the range of variability present for Glossooteris.
The use of multivariate statistical procedures in the study of fossil leaves is not new. Techniques of this type have been used to aid in the resolution of taxonomic groups (e.g., Scheihing and Pfefferkorn, 1980; Burnham, 1986; Wing and Eckenwalder, 1987; Baghai, 1988); biostratigraphic correlation within a group; identification of morphological trends through time (e.g., Doyle and Hickey, 1976; Hickey and Wolfe, 1975); the use of foliar physiognomy as an indicator of climatic regime (Dolph and Dilcher, 1979;Wolfe and Upchurch, 1987; Upchurch and Wolfe, 1987); and in understanding the interrelationship between taphonomy and paleoecology (e.g., Spicer and Hill, 1979).
Many of the limitations inherent in studying fossil plants, including the necessity of basing a data set on a limited number of specimens, or specimens from only a few localities with little or no intraspecific variability, limit the type of analysis that can be performed, and the confidence with which results are received. Continuous morphological characters of the type difficult to delimit, ironically provide better data for multivariate statistical procedures than do discrete, qualitative data bases, such as the morphological leaf features defined by Hickey (1979) (Dolph, 1976; Hill, 1980; Schwarzenwalder, 1983), since

93different morphological parameters tend to be statistically related (Dolph, 1976).
Approaches of this type have been undertaken most commonly in the study of fossil angiosperms (e.g. Tertiary Ulmoideae, Burnham, 1986; Paleogene Populus. Wing and Eckenwalder, 1987; Miocene Liriodendron. Baghai, 1988), but have also been useful in other groups (e.g., Paleozoic pteridosperm foliage, Scheihing and Pfefferkorn, 1980). In the case of angiosperm leaves, taxonomic characters are evaluated initially from a data base of extant leaves to determine a multivariate set of morphological characters to measure in fossil data (Burnham, 1986). Use of discriminate analysis provides a data set independent of a priori assumptions of which characters will be most valuable.
In the case of many angiosperm leaves whose morphological complexity and cuticular details may be distinctive enough so that variation with confidence, and in comparison to extant data set, a considerable amount of detailed information is understood concerning the morphology and biology of closely related taxa, at least at the level of section, or genus (e.g., Burnham, 1986). This provides a strong initial data base from which to design a study. A contrasting situation occurs in the case of a distinctive, poorly known, heterogenous assemblage of extinct leaf forms such as Glossopteris. In forms that

94cannot be closely tied to living analogues, it is desirable to establish some measure of understanding of the diversity present in the group before the data can be used in an integrative way.
The glossopterid leaf form: The genus Glossopterishas a unusual, if not unique, venation pattern. Although the affinities of Glossopteris-bearina plants remain obscure, this complex anastomosing venation type has been compared to the reticulate-veined leaves of a diverse array of extant and fossil plants. Leaves with reticulate venation are widespread in living vascular plants. They occur among such varied groups as the ophioglossalean, marsilealean, marattialean and filicalean ferns (e.g., Gaudet, 1964; Pray, 1961; Nair and Das, 1978; Wagner,1979), the lycopods (Selaainella. Wagner, Beitel and Wagner, 1982), the cycads (e.g., Stangeria, Brashier, 1968; Foster and Gifford, 1974), Ginkgo (Arnott, 1959), Gnetum (Rodin, 1966, 1967), the monocots (e. g., Inamdar, Shengy, and Rao, 1983), and most dicotyledonous angiosperms (Hickey and Wolfe, 1975). Although the reticulate venation of most dicots is ranked, or hierarchial, there is a great deal of variability within the group. Melville (1969, 1970b,1983a) compared Glossopteris venation patterns in detail with members of the Ranunculaceae, the Lecythidaceae, the Aponcynaceae, and the flower petal vasculature of several forms.

95In addition to the extant plants, reticulate venation
is found widely in fossil gymnospermous leaves (Alvin and Chaloner, 1970), including the foliage of Paleozoic seed ferns (e.g., Linopteris. Lonchopteris. Reticulopteris. Darrah, 1960? Taylor, 1981; Neuropteris reticularis. Oestry-Stidd, 1975), cyadophytes (e.g., Ctenis), and the cycadeoid Dictvozamites. A variety of reticulate leaves also occur among the gigantopterids (Asama, 1976), and the early angiosperms (e.g., Hickey and Wolfe, 1975; Doyle and Hickey, 1976; Wolfe and Upchurch, 1987).
Although leaves with reticulate venation are common to many groups, the superficial resemblances from one group to another do not necessarily provide a basis for recognizing homology. In order to establish more clearly the structural and potential phylogenetic relationships between groups it is instructive to analyze the construction of venation patterns in detail. Few studies of this type have been undertaken, and most of them have been limited to angiosperms. Reports of this type are both descriptive (e.g., Foster and Arnott, 1960; Foster, 1966, 1968) and developmental (Pray, 1955a, b; Slade, 1957) in approach.
Studies of the angiosperms Circaeaster and Kinadonia (Foster and Arnott, 1960; Foster, 1966, 1968) are particularly relevant to the glossopterid venation syndrome. Both leaves possess a dichotomous venation

system that is of a type unusual among the angiosperms. In this venation type, instead of the typical hierarchial venation system of angiosperms, a single order of veins is present. For this reason, this venation type is comparable to that of Glossopteris. Ginkgo (e.g., Arnott, 1959), and some ferns (e.g., Reanellidium; Pray, 1961). Foster described an array of vein morphologies present in these leaves, including "open dichotomous venation", "vein approximations", and a total of five types of reticulations (Foster, 1966, 1968). This typology of reticulations has been used and expanded by other researchers (e.g., Nair and Das, 1978).
The first type of configuration in Circaeaster that departs from the typical open dichotomous venation are the "vein approximations" (Foster, 1966). In this configuration, veins are situated closely alongside one another, but with no anatomical fusion. In a Type I dichotomy, laterally adjacent veins fuse for a short distance, and then diverge. Forms intergradational between vein approximations and true reticulations occur. Foster (1966) remarked upon the difficulty in distinguishing between vein approximations and Type I reticulations in fresh, unprocessed leaves. Only upon clearing the leaves, and seeing the anatomical features, could he determine whether veins were superficially or truly, fused. It may be that in glossopterid leaves where anatomy is unknown, a

97similar difficulty in distinguishing true and apparent fusions would be encountered.
More complex types of reticulations reported for Circaeaster and Kinadonia include Types II-V of Foster (Foster 1966), and the additional Types VI-IX elaborated by Nair and Das (1978). In Type II reticulations, the branches from two laterally dichotomizing veins join, then bifurcate. Type III reticulations are recognized by the joining of two laterally adjacent veins that then continue to the margin as a single vein. In Type IV vein approximations are united by a short "commiseral" vein.Type V reticulations and additional variations described by Nair and Das (1978) among the ferns represent an array of more complex configurations.
Several classifications of vein reticulation types have been proposed and the various typologies are not equivalent. Some previous authors (e.g., Melville, 1969) and the present study use a more descriptive approach of isolated reticulations, Foster's (1963, 1968) typology was based on the relationship of anastomoses to the overall venation pattern, (i.e., whether there are net gains or losses resulting from the reticulation). While this information may eventually be valuable in understanding structural/functional relationships in the leaves, at the present time a more general survey of structural diversity is necessary among the glossopterids. It may be that the

98distribution of particular types of reticulations is valuable in itself both functionally and systematically.
Reticulate venation among the pteridophytes is known in less detail. However, at a gross morphological level, variation is considered to be of taxonomic value (e.g., Wagner, 1979, and references cited therein). Wagner (1979) regards the numerous cases of reticulate fern venation as the result of repeated parallelism, and suggests that reticulate venation has appeared among the ferns independently over fifty times.
Among lycopods, reticulate venation is regarded as an oddity, although it occurs in some specimens of Selaainella (Wagner, et al., 1982). The presence of complex venation in lycopods is at first surprising since it contradicts the concept of the lycopsid microphyll as a simple, univeined leaf (Foster and Gifford, 1974). Selaqinella leaves with complex venation provide an excellent example of plasticity among leaf forms, beyond what might be reasonably expected in this group.
Even less is known regarding reticulate venation patterns of extant gymnosperms. Information from initial studies of Ginkgo (Arnott, 1959), Gnetum (Rodin, 1966,1967), and the cycads (Brashier, 1968) can be contrasted with Glossopteris.
Ginkgo: As a broad-leafed gymnosperm with a densely-packed type of venation, Ginkgo may represent the

closest modern analogue to Glossooteris. Similarities of these features to Glossopteris include the general organization of the venation: Ginkgo. like Glossooteris. has only one order of venation, in contrast to most other reticulate forms, which have a hierarchial organization (e.g., Gnetum. Rodin, 1966, 1967; most angiosperms, Hickey, 1973, 1979). Although Ginkgo is described as a leaf with dichotomizing venation that lacks anastomoses, they do occur sporadically in some specimens. Arnott (1959) documented four different morphological types of reticulations which occurred rarely (1-5 reticulations per individual leaf).
Anatomically, the degree to which the anastomosing veins fuse in Ginkgo is variable (Arnott, 1959). Commonly, the bundle sheath and accompanying transfusion tissue of adjacent bundles are united. In other forms lacking a bundle sheath and transfusion tissues, there is a complete union of the vascular tissue of the two veins. In still other cases, anastomoses merely represent the abutting of individual veins against one another (=vein approximations). These variations contrast with the typical reticulations of Glossooteris schopfii and G. skaarensis, where anastomosing veins are consistently interconnected even at the level of sharing diagonally oriented or bifurcating tracheids (Fig. 9, 17).

Gnetum: In Gnetum. the net venation pattern has thefollowing characteristics (Rodin, 1966, 1967): 1) ahierarchial arrangement of venation (three strongly developed orders and a fourth weakly developed order); 2)the presence of "anomalous veins", i.e., secondary and/or tertiary veins that do not "connect up" completely with higher vein orders; 3) four-thirteen (usually 5-7) separate, parallelly oriented bundles comprising the midvein near the base; 4) fibers immediately underlying the epidermis; matlike, with simple pits in their cell walls;5) laticifers; 6) a mesophyll characterized by well differentiated palisade and spongy layers; 7) frequently scalariform wall thickening patterns on the tracheids, also occasionally circular bordered pits.
Thus, although Gnetum represents a broad-leafed gymnosperm with reticulate venation, it is unlike glossopterids in several ways. Perhaps most significant is the presence of a hierarchial arrangement of venation, but there are also numerous anatomical differences, such as the presence of laticifers and a differentiated mesophyll (Rodin, 1966). Moreover, in Gnetum. the number of vascular bundles decrease distally, as the lateral bundles arch out and dichotomize, while at the apex the single remaining bundle dichotomizes (Fig. 5 of Rodin, 1966). Therefore, in the decrease in the number of veins in the midrib of Gnetum is the result of veins dichotomizing and arching out

101laterally. In contrast, in Glossopteris. veins are fewer distally as a result of both of dichotomizing and anastomosing (Fig. la, 5).
Although the monocots are typically characterized by parallel venation, a character which typically separates them from the dicots, reticulate venation has been reported in twelve species of seven families of monocots including members of the Hydrocharitaceae, Taccaceae, Dioscoreaceae, Smilacaceae, Araceae, Alismataceae, Aponogetonaceae (Inamdar, Shendy, and Rao, 1983).
In contrast to leaves of other vascular plants, the majority of angiosperms are typified by the following features: intercalary growth, stipules, a hierarchy ofprogressively finer levels of venation, several discrete orders of venation, freely ending veinlets, and vein anastomoses between two or more orders of veins (Doyle and Hickey, 1976). It is interesting that although the earliest angiosperm leaves had fewer orders of venation and their higher orders were less organized (Doyle and Hickey, 1976), they nevertheless had a hierarchial organization of venation as early as they are recognized. The presence of this features has suggested to some researchers (e.g.Doyle and Hickey, 1976) that angiosperm leaves are structurally, and possibly developmentally, distinct from other forms. The available evidence suggests that they are unlike Glossopteris as well.

In summary, the venation syndrome of Glossopteris is characterized by a number of features that together comprise a structural organization distinct from that of other leaf forms with reticulate venation. These components include the following: 1. Single order ofvenation rather than hierarchial venation. 2. Midrib composed of several parallel oriented vascular strand rather than a single vein. 3. Apically, veins of the midrib tend to either continue to the midrib or anastomose with one another, rather than arching out laterally and dichotomizing (as in Gnetum) 4. Anatomically, midrib is the result of a combination of increased size of individual vascular bundles and increased mesophyll thickness. There is, in known species, no apparent specialization of ground tissues in region of the midrib that makes it different; also, no large bundle sheath surrounding more than one bundle. 5. If "closed" vs. "open" venation is defined by presence vs. absence of reticulations, then Glossopteris has closed venation. If it is defined by whether there are marginal veins of some type, then glossopterids, which lack marginal veins, are characterized by open venation. 6. In contrast to almost all other forms, Glossopteris represents only form of leaf with one order of venation that consistently shows reticulations as a major part of its venation morphology. In Ginkgo. Circaeaster. and the floral petals of Ranunculus. for instance, the venation is

characteristically dichotomous, with only a few (1-8) reticulations per individual leaf (Arnott, 1959? Arnott and Tucker, 1963; Foster, 1966). In this aspect, perhaps these forms reflect a condition similar to that of putative glossopterid ancestral forms. Certainly within the glossopterids, some forms (e.g., Glossooteris argentina Archangelsky, 1957) have few anastomoses; and they are lacking in the glossopterid foliage genus Palaeovittaria. Other leaves in which this degree of reticulation is an integral part of the structure include several forms of Paleozoic seed fern foliage, (e.g., Linooteris). the Mesozoic seed fern frond Sagenooteris. and Mesozoic and extant cycadophyte foliage (e.g., Ctenis. Stangeria, respectively, Alvin and Chaloner, 1970). These latter forms differ from Glossopteris in representing complex, highly dissected pinnate fronds (Linopteris), or pinnately (Ctenis. Stangeria) or palmately compound leaves (Sagenooteris) rather than entire simple leaves. 7. Anatomically, the tracheids of Glossopteris are more intimately involved in the reticulations than in other forms. For example, individual tracheids dichotomize and extend into both arms of a bifurcating vein, as well as traversing reticulations diagonally.
Although some of these features are characteristic of other groups, none are expressed to the degree of elaboration seen among the glossopterids. Although

104ontogenetic, phylogenetic, physiological, and ecological implications of this leaf form remain yet to be investigated in detail, anatomical information available from the Skaar Ridge leaves provides a significant starting point for assessing the characteristics of Glossopteris in a more biological context.
In recent years, biologists have become increasingly interested in the adaptive significance of leaf form (e.g., Horn, 1971; Parkhurst and Loucks, 1972; Givnish and Vermeik, 1976; Givnish, 1979; Chabot and Chabot, 1982; Parkhurst, 1986, and references cited therein). As the primary organs of photosynthesis, leaves represent dynamic, metabolically active structures which must strike a balance between optimizing carbon dioxide uptake and minimizing water loss (Givnish, 1979). Leaves are intimately tied to their environment and are frequently capable of a great deal of plasticity in response to it. For example, microhabitat stresses commonly lead to the occurrence in the same organism of so-called sun leaves and shade leaves that are morphologically and physiologically distinct (e.g., Chabot and Chabot, 1982 and references cited therein). In a larger sense, such elaborate physiological and morphological adaptations as those associated with C-4 (e.g., Laetsch, 1974) and CAM (e.g., Keeley, 1983) photosynthesis have evolved repeatedly, particularly in groups that inhabit extreme environments.

In studying the adaptive significance of leaf form, some authors (e.g., Parkhurst and Loucks, 1972; Givnish, 1979; Parkhurst, 1986) have invoked arguments of optimal design, that is, that selection acts toward favoring certain "optimal" structural types that survive favorably in their environment (Parkhurst and Loucks, 1972). Some authors (e.g., Parkhurst and Loucks, 1972; Parkhurst, 1986, and references cited therein) have produced mathematical models to account for the physical and physiological components of the leaf structure-function relationship, such as the effect of boundary layers on C02 conductance, the functioning of three-dimensional architecture of the mesophyll, and the physics of light absorbance. In other studies, (e.g., Givnish, 1979) the cost-benefits of leaf production over time have been analyzed, particularly in reference to evergreen leaves. For instance, some authors have suggested that conifer needles, with their secondary compounds, may represent an adaptation to herbivore deterrence, rather than to the positive cost-benefit of leaves that last more than one growing season (Givnish, 1979).
Other studies have focused more on the spatial relationships of leaves in the environment, such as the position of leaves on the tree in relationship to incident light (i.e., adaptive geometry, Horn, 1971), or the occurrence of given leaf forms at different levels in the

106forest canopy (e.g., Givnish, 1982; Givnish and Vermeik, 1976). Together, these approaches lead to several interesting questions. For instance, first, are there optimal leaf types for a given environment? Secondly, can a given leaf form be used to predict the type of environment in which it is found? Although those studying extant ecosystems may focus on the first question (i.e., structural/functional relationships and cost-benefit of particular leaf forms), paleobotanists may find value in data that could address both the first and second question. If leaf forms can be used to predict environment with confidence, they can be considered of great value to paleoenvironmental reconstruction.
Although fossil angiosperm leaves have been valuable in predicting environment on a broad climatic scale (e.g., Dolph, 1976; Hickey, 1979; Upchurch and Wolfe, 1987) in general, disarticulated leaf fossils are limited in the amount of information they can give about the plant which bore them. In cases where the individual organism represented by dispersed plant organs is poorly understood, the microhabitat relationships of such a plant also remain obscure.
Since most of the ecological studies of leaf adaptation have focused on angiosperms, caution must be exercised in the degree of extrapolation made toward Glossopteris, a poorly understood gymnosperm. A better

107 'understanding of glossopterid biology, and the natural variation within this leaf form is needed before such extrapolations can be made with confidence. Modelling of extant gymnosperms such as Ginkgo. which may represent the closest modern analogue to Glossooteris. may be valuable in addressing these questions. Moreover, many of the important parameters studied in extant ecosystems are related to the active physiology of photosynthesis, such as photosynthetic rate, stomatal resistance, transpiration rate, etc. Obviously, these parameters cannot be obtained from fossil data.
Despite these limitations, the Antarctic Glossooteris leaves provide a significant source of data from which to approach leaf structure/function relationships. These specimens provide the first data on the range of variation available for anatomically preserved leaves of this type.The two forms that would both be considered glossopterid based on morphological features have strikingly different anatomical characteristics. These different patterns, i.e., the prominent fibrous bundle sheath and simple stomata of G. schopfii vs. the thick hypodermis and elaborate stomata of G. skaarensis. reflect different structural adaptations to, presumably, the same environment. These anatomically variations in light of morphological similarity present a paradox on interpreting Glossopteris structurally, functionally, and

108phylogenetically. The absence of a well-differentiated mesophyll in both forms may be common in, and significant to glossopterid physiology in general.
Moreover, data from the Antarctic leaves can provide a strong basis for investigating the extensive compression/impression record of Glossooteris in a more informed way than has previously been possible. Reinterpretation of this large plexus of leaves will present a valuable new perspective on the leaf form that dominated the Gondwana landscape during the Permian.

CHAPTER VI - PART II. Dicroidium. INTRODUCTION
The genus Dicroidium Gothan represents a leathery, frond-like foliage form with a bifurcating rachis, entire to several times pinnate laminae, and pinnules demonstrating variable morphologies, (e.g., sphenopterid, odontopterid, alethopterid) (Townrow, 1957; Retallack, 1977b; Petriella, 1981; Anderson and Anderson, 1983). Dicroidium is the dominant foliage form in the Triassic compression floras of Gondwana, and occurs on all major continents of the southern hemisphere.
Because of its widespread distribution throughout Gondwana, Dicroidium has served as one of the most important biostratigraphic megafossils for the Triassic (e.g., Sitholey, 1954; Schopf and Askin, 1980; Anderson and Anderson, 1983; Boersma and Broekmeyer, 1986, and references cited therein). Since the initial illustration of Dicroidium fronds from Tasmania by Morris in 1845 (described under the name Pecopteris odontopteroides) upwards of 30 taxa of Dicroidium have been defined (including varieties [Retallack, 1977b], subspecies and forma [Anderson and Anderson, 1983]). Dicroidium has been
109

reported from Australia, (Carruthers, 1872? Feistmantel, 1878-1879, 1890; Tenison-Woods, 1883; Curran, 1884; Johnston, 1886, 1887, 1888, 1894, 1896? Shirley, 1898? Etheridge, 1890, 1891, 1902; Jack and Etheridge, 1892; Antevs, 1913, 1914; Walkom, 1917, 1922; Jones and deJersey, 1947; Jacob and Jacob, 1950; Townrow, 1962; Douglas, 1969? Retallack, 1977b? Holmes and Ash, 1979; Rowett, 1979; Holmes, 1982); Tasmania (Morris, 1845; Crepin, 1875)? New Zealand (Arber, 1913, 1917); South America (Geinitz, 1876; Szajnocha, 1888; Solms-Laubach and Steinman, 1899; Kurz, 1921? Frenguelli, 1941, 1943, 1950? Gordon and Brown, 1952; Jain and Delevoryas, 1967; Archangelsky, 1968a; Petriella, 1978? Baldoni, 1980; Bortoluzzi, Guerra-Sommer, and Cazzulo-Klepzig, 1985; Mohr and Schoener, 1985); South Africa (Feistmantel, 1889? Seward, 1903, 1908; Seward and Holttum, 1921; du Toit, 1927; Thomas, 1933; Townrow, 1957; Lacey, 1976; Anderson and Anderson, 1983); Antarctica (Plumstead, 1962? Townrow, 1967; Orlando, 1968? Rigby and Schopf, 1969; Lacey and Lucas, 1981; Rigby, 1985; Banerji and Lemoigne, 1987)? Madagascar (Carpentier, 1935), and India (Feistmantel, 1882; Seward, 1932; Lele, 1961a, b; Rao and Lele, 1963? Bose and Srivastava, 1973; Bose and Banerji, 1976? Pal, 1984). Dicroidium compression floras have been used in the stratigraphic correlations from the Late Permian through the early Jurassic of Australia (Retallack, 1977b) where biozones based on fossil plant

Illassemblages have been interpreted as ecological associations. Leaves of this type have also been studied as floristic assemblages in South Africa (Anderson and Anderson, 1983).
Dicroidium is believed to represent the vegetative foliage of a Mesozoic pteridosperm group, the corystosperms (Thomas, 1933, 1955). Thomas (1933) established the Corystospermaceae to encompass an assemblage of South African compressions that contained the helmet-like cupule Umkomasia and the pollen organ Pteruchus. He suggested that Dicroidium represented the vegetative foliage, based on co-occurrence and cuticular similarity (Thomas, 1933,1955). Later authors (e.g., Townrow, 1957? Archangelsky, 1968a; Retallack, 1977b; Petriella, 1979, 1980, 1981; Holmes, 1987) accepted this assignment and expanded the concept of the corystosperms in several ways. Archangelsky (1968a) viewed the constant association of Dicroidium fronds with the unusual gyranospermous stem Rhexoxvlon (Bancroft, 1913? Walton, 1923, 1925, 1956; Archangelsky and Brett, 1961) as evidence that Rhexoxvlon stems bore Dicroidium-tvpe leaves. Accepting this assumption, Petriella (1978) reconstructed the Dicroidium plant as a small tree bearing frond-like leaves. Additional fertile structures assigned to the corystosperms have been described from Australia (Holmes, 1987), Rhodesia (Lacey, 1976)? India (Srivastava, 1971), and South America

112(Frenguelli, 1942; Petriella, 1980) although some authors (e.g., Retallack, 1977b) have expressed doubt as to the corystosperm affinities of some of these reproductive organs.
Although the highly variable frond morphology of Dicroidium has been characterized from numerous specimens, and cuticular detail is known from some species (e.g.,Jacob and Jacob, 1950; Townrow, 1957; Archangelsky, 1968a; Anderson and Anderson, 1983) anatomical features are unknown.
Worldwide, the occurrence of anatomically preserved plants of Triassic age is extremely rare. Although isolated specimens of petrified stems occur at several Triassic localities in South America (e.g., Archangelsky and Brett, 1961, 1963, South Africa and India (Krausel, et al., 1961; Prasad, 1982, 1986), and fragments of petrified wood are widespread throughout the Permian and Triassic of Gondwana (e.g., Krausel, et al., 1961; Krausel, 1962; Maheshwari, 1972), anatomically preserved specimens of other plant organs of Triassic age are poorly represented. With the exception of specimens preserved as lignitic forms in the South Shetland Islands (Lacey and Lucas, 1981) the only permineralized Triassic plant locality with a diverse perraineralized flora occurs in the Central Transantarctic Mountains (Smoot, et al., 1987).

Foliage assignable to Dicroidium represents a very common component of the Fremouw Peak flora (Smoot, et al., 1987; Pigg and Taylor, 1987b; Pigg, 1988). Permineralized Dicroidium specimens from this site provide the first opportunity to document anatomical detail for this leaf type, including details of vascular tissue, frond architecture, and mesophyll organization. From this material the anatomical basis for external morphological features can be established; furthermore, morphology can be compared with the previously described species of Dicroidium known from compression/impression floras (e.g., Retallack, 1977b, Anderson and Anderson, 1983). Anatomical data of this type provides important new information regarding this dominant leaf form of the Triassic, and may provide valuable clues for better understanding both the ecology of corystosperms and their phylogenetic relationships with other gymnosperms.

CHAPTER VII - DESCRIPTION
Specimens of Dicroidium fremouwensis. sp. nov. are represented in the silicified peat by fragments of fronds with attached laminar pinnae (Fig. 58-64). Most of the specimens represent central portions of the frond. Four specimens have been discovered that demonstrate the proximal bifurcation characteristic of Dicroidium (Fig.58e, f, 63, 74), no specimen of the frond apex has been recovered. The following description is based on twenty-two specimens from weathered surfaces, which provide external morphological detail, and forty additional specimens prepared in various planes of section. Five specimens (Fig. 58b, c, d, e, 61-63, 70-75) were peeled in a combination of transverse, oblique and paradermal views to correlate anatomical features in a three-dimensional context.
The most complete fragment of D. fremouwensis is 9 cm long x 4.6 cm wide (Fig. 58a, 59). Since this specimen represents the central portion of a frond, and lacks both the proximal bifurcation and the leaf apex, the entire fronds were probably somewhat larger. Based on correlations with compressions of Dicroidium with similar
114

115structure, it is estimated that fronds could have been to up 15 cm long.
Two types of pinnae occur on the frond, more basal pinnae are bluntly elongate, 1.5 x 0.7 cm, and entire (Fig. 58e, f, g, h, 63, 64). More distal pinnae are lobed, or bipinnatifid with 3-10 rounded lobes (Fig. 58a, b, c, d 59-62). The longest, and most distal pinnae are 1.6 x 0.7 cm and may possess up to 10 lobes (Fig. 58a, b, d, 59-62). All pinnae are attached to the frond rachis at nearly right angles, with laminae broadly attached to the rachis (Fig. 59-64). The pinna rachis itself is broad, 3.6 mm in diameter, and remains fairly constant in size throughout the length of the frond (Fig. 59-64).
Two specimens demonstrate the external morphology of the frond bifurcation diagnostic of Dicroidium (Fig. 58 e, f). Proximal to the bifurcation, the petiole in one specimen (Fig. 58e, 63) is fairly robust (5 mm wide), at a comparable level in the second specimen (Fig. 58f) the rachis is 1.5 mm wide. Distal to this dichotomy, frond axes are somewhat narrower (3.5 and 1.3 mm respectively). Differences in size between the two specimens are thought to reflect the natural biological variability within the taxon.
Venation of laminar pinnae is of the odontopteroid type, with 1-3 veins entering the central part of the pinna (Fig. 58a, b, c, d, 60, 62). Secondary veins diverge at a

116fairly steep angle to the midvein (75-78°)? most of them then dichotomizing about halfway between the midrib and the leaf margin (Fig. 58a, 60, 62). Venation is symmetric in pinnules, with 8 veins/cm2 occurring in the central region of the leaf and 16 veins/cm2 at the margin.
Although specimens of the petiole at a level proximal to the frond dichotomy not have been discovered, three specimens demonstrate the anatomical detail of the bifurcation itself (Fig. 58e, 63, 74). The dichotomy is approximately equal, resulting in rachides about 1.1 cm high and 2 cm wide (Fig. 74). At this level both rachides demonstrate the venation pattern characteristic of the frond as a whole: bundles occur in two groups: an abaxialgroup of six bundles in a ring and an adaxial group of six bundles arranged linearly (Fig. 74). At more distal levels the rachis is 3.5 mm high by 2.5 mm wide in transverse section and is vascularized by at total of 15-20 bundles arranged in two groups (Fig. 75). The abaxial ring consists of 7-8 bundles while on the adaxial surface 5-8 bundles are arranged in a linear group (Fig. 71, 75). Bundles of the abaxial group extend the length of the frond rachis, while those of the adaxial group diverge laterally to vascularize the pinnae. In the most distal sections of the rachis the number of strands decreases to 3-5 (Fig. 72, 73) .

117Laminar pinnae of both types are vascularized by
usually one, but sometimes up to 3 major veins that divide within the lamina to produce secondary veins, which also dichotomize (Fig. 60, 62). The midrib is composed of a shallow U-shaped vascular region with a prominent region of radially aligned tracheids (Fig. 65, 70, 73). Lateral (secondary) veins are much smaller and more inconspicuous in transverse section (Fig. 66, 67, 70). Secondary veins dichotomize to produce, in turn, the marginal veins.
Vascular strands of the frond rachis and the midvein of pinnae are characterized by a region of 4-8 rows of radially aligned tracheids that occur in a prominent lacuna (Fig. 65, 66, 68). In these veins the primary xylem is usually displaced or separated from the radially aligned tracheids, and is difficult to distinguish (Fig. 65, 69).In lateral (secondary and tertiary) veins of the laminar pinnae, however, primary xylem is better preserved, and is characterized by small terete groups of protoxylem tracheids exarch or marginally mesarch within a group of approximately 5-10 metaxylem tracheids (Fig. 67). Lateral bundles typically lack the radially aligned tracheids (Fig. 67). Tracheids of the primary xylem typically have helical to scalariform-reticulate wall thickening patterns; the radially aligned tracheids are characterized by wall thickenings of the scalariform-reticulate type (Fig.76-77). Adjacent to tracheids within the bundles are

118smaller, barrel-shaped cells 82 x 45 nm with scalariform or reticulate wall thickening patterns (Fig. 76, 78). Cells of this type may represent transfusion tracheids or simply unusually short tracheids. Their abundance within vascular strands suggests that they represent a substantial portion of the xylem present.
Abaxial to the xylem, bundles are frequently characterized by a zone 4-5 cells wide of radially aligned cells that occur in the position of secondary phloem (Fig. 68, 69). Cells of this type are approximately 8 jum in diameter, but are typically crushed and poorly defined. No sieve areas have been found. In some specimens, a region of about 25 elongate fiber-like cells with dark contents subtend this zone (Fig. 68, 69).
The vascular bundle sheath is composed of 1-3 cell layers of narrow, elongate cells 210 x 6 /m with thick walls. Although these cells resemble fibers, they lack any type of pits or wall ornamentation, and have a continuous sheath-like appearance to their surfaces (Fig. 76-78). To the outside of these cells a single layer of large cuboidal cells 30 x 36 fm with large lumens occurs end-to-end along the length of the bundle sheath (Fig. 76).
The ground tissue of large frond members (i.e., petiole and frond rachis) is composed of two zones, an inner, loosely organized parenchymatous zone and an outer zone of smaller, more compact, cuboidal cells (Fig. 71,

1 1 9
75). In some specimens, a palisade layer can also be delimited on the adaxial surface (Fig. 75, top), suggesting that the rachis may have been photosynthetic. Within the ground tissue, many cells have dark contents. It is unclear, however, whether this feature is of anatomical significance or preservational in nature. Elongate cells with amber-colored, presumably resinous contents are infrequently scattered throughout the ground tissue (Fig. 75).
Laminar pinnules possess a well-differentiated mesophyll with palisade and spongy layers. The palisade layer occurs adaxially and is 1-3 cell layers thick. Palisade cells are 110 ;xm high x 40 jum wide, and frequently have dark contents (Fig. 65, 67, 73, 79). Cells of the spongy layer frequently poorly preserved (Fig. 65, 80).
Dicroidium fremouwensis is amphistomatic with stomata infrequently scattered on the upper epidermis. Epidermal cells are polygonal (32 x 42 pm) to elongate (84 x 23 jLtm) , and have straight margins on their anticlinal walls (Fig. 81). In transverse section, epidermal cells appear tabular (Fig. 67). Stomata are dicyclic with 5-7 subsidiary cells which form an incomplete ring (^'pseudosyndetochelic11 of Retallack, 1977b) (Fig. 81, 82). Guard cells are thickened and 23 x 9 /xm, and surround a stomatal opening which is 14 x 5 um in diameter. Subsidiary cells are 26 x 32 /xm.

CHAPTER VIII - DISCUSSION
Generic concept of Dicroidium.— The genus Dicroidium was established in 1912 by Gothan to distinguish southern hemisphere foliage from the similar northern hemisphere genus Thinnfeldia Ettinghausen (Gothan, 1912). According to Gothan, the morphological and cuticular features that separated Dicroidium from Thinnfeldia included bifurcation of the frond, type of venation, thickness of cuticle, epidermal and stomatal cell patterns and stomatal distribution. Gothan (1912) also stressed that Dicroidium was restricted to "the Glossopteris region", or the southern hemisphere (Townrow, 1957).
Although several workers (i.e., Arber, 1917; Walkom, 1917; du Toit, 1927; Medwell, 1952; Jones and deJersey, 1947) continued to include the southern hemisphere forms within Thinnfeldia. other workers accepted Gothan*s (1912) generic designation (e.g., Antevs, 1913, 1914; Frenguelli, 1943; Jacob and Jacob, 1950, Townrow, 1957; Bonetti, 1966; Archangelsky, 1968a; Retallack, 1977b; Holmes and Ash,1979; Petriella, 1981, Holmes, 1982).
Walkom (1925) established the genus Johnstonia for an assemblage of Tasmanian fronds of Triassic age, with or
120

121lobed margins. In 1943, Frenguelli established the additional genera Xvlopteris. Zuberia. Dicroidiopsis. Diplasiophvllum and, in 1950, Tetraptilon for corystosperm foliage from South America on the basis of morphological variation. Townrow (1957) emended the diagnosis of Dicroidium to include the genera Johnstonia and Zuberia. placing emphasis on the similarity of cuticular structure among these taxa. He recognized three major distinctions between Dicroidium and Thinnfeldia: the presence of frond bifurcation, the distribution and morphological details of the stomata, and several features of the epidermal cells (Townrow, 1957). According to Townrow (1957), the most characteristic distinction between the two genera is the presence of a frond bifurcation in Dicroidium and its lack in Thinnfeldia. Secondly, Thinnfeldia was described as hypostomatic with stomata regularly distributed in interveinal bands, rounded stomatal pits and weakly cutinized guard cells (Townrow, 1957). In contrast, Dicroidium leaves were characterized as amphistomatic, with irregularly disposed stomata possessing strongly cutinized guard cells. lastly, epidermal cells were characterized by Townrow (1957) as straight and smooth-surfaced in Thinnfeldia but sinuous and commonly papillate in Dicroidium (Townrow, 1957).
Bonetti (1966) suggested that the genus Dicroidium should probably include the forms recognized as

Diplasiophvllum. Dicroidiopsis. Hoeaia and Xvlopteris. but did not formally amplify Dicroidium to include these taxa. Archangelsky (1968a) made the formal taxonomic revision, synonymizing these four genera with Dicroidium. In including Townrow's (1957) genus Hoeaia in Dicroidium. Archangelsky (1968a) suggested that some cuticular details were quite variable even within individual specimens and not of sufficient value to establish separate taxa. Holmes(1982) also recognized the single genus, Dicroidium. for this plexus of leaf forms, as did Anderson and Anderson(1983), who further synonymized the genus Tetraptilon Frenguelli. While Retallack (1977b) and Petriella (1981) accepted Archangelsky's (1968a) conclusions that these taxa represent closely related forms, they continued to regard Johnstonia and Xvlopteris as generically distinct, based on pinnule morphology. Baldoni (1980) agreed with the latter concept and reviewed forms known as Xvlopteris from Argentine strata.
Dicroidium has also at times been compared or merged with several additional Mesozoic foliage genera possessing dichotomizing fronds (Townrow, 1957; Archangelsky, 1968a; Petriella, 1979, 1981). These taxa include Ptilozamites Nathorst, and Supaia White (Townrow, 1957). Forms with which Dicroidium has been compared which typically lack a frond dichotomy include Pachvpteris Brongniart, Pachvdermophv1lum Thomas and Bose, Cvcadopteris (including

123Lomatopteris) Zingo, Ctenozamites Nathorst, Stenopteris Saporta, Glenopteris Sellards, Protophvllocladus Berry, and Protobelchnum Lesguereux (Townrow, 1957). Although Dicroidium has seldom been formally discussed in relation to the peltaspermaceous foliage form Leoidopteris (Townrow,1956), superficial resemblances between the two taxa have occasionally led to confusion between them (e.g., Bose, pers. comm., 1988).
Although the genus Dicroidium has historically been used by a number of authors, the criteria accepted for its recognition vary (Townrow, 1957; Archangelsky, 1968a; Petriella, 1981). Antevs (1914) accepted the generic name Dicroidium but not the taxonomic criteria as established by Gothan, and included forms that may be more similar to Thinnfeldia (Antevs, 1913, 1914; Townrow, 1957). Jones and deJersey (1947) included Dicroidium sensu Townrow (1957) in the genus Thinnfeldia with reservations. Other authors accepted only part of Gothan's criteria: Frenguelli (1943)distinguished the two forms by the presence of frond bifurcation in Dicroidium but did not accept cuticular studies; while Jacob and Jacob (1950) recognized geographical distribution as the only factor separating Dicroidium and Thinnfeldia.
Although Townrow1s (1957) emendation of Dicroidium has clarified much of the confusion surrounding differences between this taxon and Thinnfeldia. a cursory look at the

124wide variation present in Dicroidium illustrated in more recent publications (e.g., Anderson and Anderson, 1983) suggests that some taxonomic characters are more diverse than previously recognized, and that the genus could now be more broadly defined. In particular, in addition to being "pinnate to bipinnate, with a forked rachis", (Townrow,1957), some fronds of Dicroidium are tripinnate or demonstrate several types of "aberrant" branching, or even lack the characteristic frond bifurcation (Anderson and Anderson, 1983). In addition, many cuticular types illustrated by Anderson and Anderson (1983) have straight-margined epidermal cells (e.g., Fig. 6? Plate 89, 90, of Anderson and Anderson, 1983), in contrast to Townrow's (1957) description of epidermal cells characterized by sinuous walls and lateral extensions.
Tjie present description of the D. fremouwensis. based on permineralized specimens, necessitates a further amplification of the generic concept to include anatomical features that have been found in these leaves conforming to the Dicroidium morphotype. This amplification, however, does not imply that all leaves with the morphological structure known as Dicroidium necessarily possess the anatomical features included.
Species of Dicroidium: Species of Dicroidium havebeen defined on characters of frond morphology and cuticular features and many of the same problems

encountered in generic descriptions have found their way into species delimitation. While some authors have emphasized the morphological variation even to the point to generic rank (e.g., Frenguelli, 1943, in the establishment of the genera Xvlopteris and Johnstonian, others (Jacob and Jacob, 1950; Townrow, 1957) stressed the morphological plasticity within fronds of the same type and suggested that cuticular features were of greater value in delimiting species, still others (e.g., Archangelsky, 1968a) have recognized that the degree of variation in cuticular features previously used to delimit species and genera (e.g., Hoegia) may be found in different parts of the same frond. Anderson and Anderson (1983) have expressed the opinion that although cuticle is known particularly well in some remains (e.g., the Molteno Formation of South Africa) such excellent preservation is not available in many localities and cannot be used in many taxa.
Retallack (1977b), recognizing these limitations, constructed a taxonomic key based on gross morphological features. He recognized three genera (Dicroidium. Johnstonia and Xvlopteris), 23 species, nine of which were split into 25 varieties, of corystosperm foliage from the late Permian to early Jurassic of eastern Australasia. Petriella (1981) succinctly summarized the species of Dicroidium known from Argentina and provided a taxonomic key to these forms that is based on morphological features.

126Anderson and Anderson (1983), in their monograph on South African Dicroidium. recognized 8 polymorphic species and a total of 31 taxa, including species, subspecies and forma. While Retallack (1977b) stressed the utility of his taxa for biostratigraphic purposes, Anderson and Anderson (1983) attempted to include in their classification scheme an element of "population studies" by placing emphasis on the polymorphic nature of species and on co-occurrence of forms at a single locality.
At the present time, all specimens of Dicroidium discovered at Fremouw Peak appear to represent intergradational forms of the same species, based on both anatomical and morphological features (Fig. 59-64; 65-82). If small fragments showing surface morphology were studied in isolation (e.g., Fig. 59, 60, 61, 63), the variation of pinnule type might invite the description of more than one species, but several specimens bear both the more basal, entire, and more distal, bipinnatifid pinnae, demonstrating the intergradation of morphologic form. All specimens studied have the same anatomical features, and, at the present time, no variation is known to indicate that more than one species is present.
Based on morphological features (Fig. 60-66). Dicroidium fremouwensis compares most favorably with D. dubium var. australe (Jacob and Jacob) Retallack (Frame H8 of Retallack, 1977b). Retallack (1977b) described this

127form as a "medium to large irregularly bipinnatifid frond" with relatively obscure and dense secondary venation, and a thick coriaceous lamina. He synonomized his material with Thinnfeldia polvmorpha (Johnston, 1896), T. narrabeensis (Walkom, 1925), Dicroidium australis (Jacob and Jacob,1950), D. walkomi (Jacob and Jacob, 1950), D. sp. cf. feistmanteli (Lele, 1961a), D. antevsiana (Anderson and Anderson, 1970), and D. sp. cf. D. australe (Gould and Retallack, 1977). Of these forms, Walkom's (1925) T. narrabeensis is much larger than D. fremouwensis. and Lele's (1961) D. sp. feistmanteli. is very similar to the present material (see PI. 2, Fig. 9, of Lele, 1961a). Lele (1961a), in his discussion of this taxon, mentions that Seward (1908) commented on the pinnule variability found in certain specimens of D. odontopteroides. that "demonstrate the passage, from linear segments with a midrib giving off clusters of forked veins and with a lamina showing different degrees of lobing to pinnae with short and broad ultimate segments" (Seward, 1908). Although the frond apex of D. fremouwensis is not known, Seward's (1908) description of pinnule variation within an individual frond is appropriate. Dicroidium feistmanteli figured by Jain and Delevoryas (1967, PI. 90, Fig. 6A) is similar to the more basal, entire pinnae of D. fremouwensis (Fig. 58g, h, 64), but pinnules of this type alone (without more information on the entire frond organization), are not

128sufficiently diagnostic to warrant taxonomic assignment.In contrast, a specimen illustrated by Townrow (1967, Fig. 4B) as D. feistmanteli is quite different in pinnule morphology, with blunt, D. zuberi-like pinnules (Archangelsky, 1968). Townrow (1967) compares this specimen to Hoeaia (Townrow, 1957; later D. zuberi; Archangelsky, 1968a), and it is unclear why it is assigned to D. feistmanteli and not Hoeaia initially.
Using Petriella's (1981) taxonomic key for Argentine specimens of Dicroidium. D. fremouwensis falls easily into the species D. dubium (Feistmantel) Gothan, i. e., the larger species D. dubium from which Retallack (1977b) separated D. dubium var. australe discussed above.Anderson and Anderson (1983) list three species as having bipinnatifid fronds, namely D. dubium. D. superbum, and D. elongatum. Of these forms, D. fremouwensis is again most clearly associated with D. dubium types. In particular, D. fremouwensis bears some resemblance to D. dubium subsp. tasmaniense (Johnston) Retallack, et al. 1977.
Lastly, Dicroidium species listed (but not figured) from Antarctica have been limited to only a few taxa, namely, D. odontopteroides. D. feistmanteli and D. sp. from the Transantarctic Mountains and the Sentinel Ranges (Rigby and Schopf, 1967); E>. cf. spinifolia. D. cf. elonaata (Lacey and Lucas, 1981) and forms similar to D. odontopteroides (Banerji and Lemoigne, 1987) from the South

129Shetland Islands and D. dubium. D. odontopteroides. and D. zuberi from South Victoria Land (Rigby, 1985). Townrow (1967) described species of Dicroidium from the Transantarctic Mountains.
Cuticular features of D. fremouwensis are highly variable from specimen to specimen and it is not always possible to determine what features, particularly of cuticular surface, are of biological origin and which are preservational in nature. However, the presence of straight-margined cells, surficial papillae, pseudosyndetochelic stomata described for D. dubium var. australe by Retallack (1977b) are consistent with features seen in D. fremouwensis. Use of cuticular remains in Dicroidium systematics in a meaningful way awaits a critical assessment of variability known in the taxon as a whole and within individual specimens, as well as the discovery of more reliable material.
Since the compression/impression forms with which D. fremouwensis has been compared lack anatomical features themselves, it is not appropriate to formally synonymize D. fremouwensis. a taxon based on a combination of morphological, cuticular, and anatomical features, with these forms. However, D. fremouwensis should be included within this general group of forms and considered a closely related species (Galtier, 1986).

Certain features of frond morphology and cuticular structure have recently led to the suggestion that the present material may be more closely allied with Lepidooteris (Townrow, 1956), than Dicroidium. or perhaps represent an intergradational form between Dicroidium and Lepidooteris (Archangelsky, Bose, pers. comm., 1988). However, based on the currently available evidence, the present author believes the Antarctic material described herein more closely conforms to the genus Dicroidium. Salient features that distinguish the two taxa include the presence in Leoidopteris of interpinnules, or entire pinnules interspersed with lobed pinnae, bump-like tubercles on the surface of the frond rachis, and small extensions on the guard cells (i.e., lappets).Additionally, fronds of Lepidooteris are hypostomatic while those of Dicroidium are characteristically amphistomatic (Townrow, 1956, 1957). Dicroidium is furthermore characterized by the presence of a major frond dichotomy, a character not recognized as a distinguishing feature of Leoidopteris.
Dicroidium fremouwensis is characterized by fronds with two types of pinnules, but there is no evidence of an alternation of the two types, as would be expected in frond fragments of the Leoidopteris type. Secondly, although the rachis has a somewhat irregular surface, it lacks prominent tubercles like those that occur in Lepidooteris.

Dicroidium fremouwensis is an amphistomatic frond, with considerably fewer stomata on the adaxial surface, a situation common in Dicroidium. Lastly, stomata of D. fremouwensis do possess several bumps or thickenings per guard cell that may represent "lappets" (Townrow, 1956) like those of Leoidopteris. but similar characters have been described for many of other species of Dicroidium (e.g., Anderson and Anderson, 1985), and in the literature this character has not been clearly restricted to Lepidooteris. Clearly, at the present time the generic concepts of these foliage forms are in a state of flux. As additional information is available from both compressions and petrifactions, perhaps the relationship of these taxa will be more clearly delimited.
Frond architecture— Anatomically preserved plant material from Fremouw Peak provides the first opportunity to describe the internal structure of Dicroidium leaves (Pigg and Taylor, 1987a, Pigg, 1988). Perhaps the most important anatomical features of D. fremouwensis are the vascular architecture of the frond, the anatomy of the vascular bundle and bundle sheath, and the complexity of mesophyll tissues. In transverse section, the frond rachis of D. fremouwensis is characterized by a radial group of bundles occurring abaxially and a linear group in an adaxial position (Fig. 70-75). This particular pattern of frond vascular architecture most closely resembles that of

132several extant cycad genera. Worsdell (1906) described the petioles of cycad leaves as having bundles with "their xylems slightly directed towards each other" (see Fig. 15 of Worsdell, 1906), and compared them with the petioles of the medullosan frond Mveloxvlon. Bierhorst (1971) illustrates a petiole of C. revoluta showing a vascular arrangement with a nearly complete abaxial ring of 14 bundles that extend on either end to vertical rows of three bundles each, that, in turn, join onto adaxially positioned lateral flanks of four bundles each (Fig. 20-4 D of Bierhorst, 1971). It is not difficult to imagine in C. revoluta. with the loss of the vertical bundles,' an arrangement strikingly like that of D. fremouwensis (Fig. 75) .
The frond vasculature of Dicroidium is also somewhat like the petioles of some filicalean ferns (Keating, 1968). In particular, Dicroidium resembles Pteridium aquilinum and Hemitelia choricaroa. in having adaxial and abaxial (="ventral and dorsal", Keating, 1968) divisions to the vascularization of their petioles (compare Fig. 75 with Fig. 24 and 26 of Keating, 1968). Dicroidium can also be compared to the petiole of Rubus. which has a completely radial organization of its vascular strands (Fig. 199 of Napp-Zinn, 1973).
In comparison to other pteridosperms for which frond anatomy is known, the frond vascular architecture of

Dicroidium appears unique. Lyginopterid, calamopityean, and callistophytalean seed ferns are characterized by a vascular strand that is either single and U-shaped, (e.g., Lvainopteris. Blanc-Louvell, 1966; Microspermopteris. Pigg, Stockey, and Taylor, 1985; Feraxotheca. Millay and Taylor, 1977, 1978), single and band-like (Callistophvton.Rothwell, 1975) double or composed of several individual bundles (Calamopitvs. Galtier, 1974; Heterangium. Shadle and Stidd, 1974; Pigg, Taylor and Stockey, 1987; Schopfiastrum, Rothwell and Taylor, 1972; Stidd and Phillips, 1978; Taylor and Millay, 1977). Neither does Dicroidium show the complex frond architecture known for the medullosans (Basinger, Rothwell and Stewart, 1974), such as fronds of Neuropteris ovata and N. scheuzeri in attachment to Mveloxvlon (Beeler, 1983) and Medullosa endocentrica (Hamer and Rothwell, 1988). In these forms the petioles are characterized by an outer ring of numerous vascular bundles and an inner region of either scattered bundles (Beeler, 1983), or bundles arranged linearly (Beeler, 1983; Hamer and Rothwell, 1988).
Anatomical features of the bundle sheath include the presence of marginally exarch/mesarch primary bundles, radially aligned tracheids, abundant transfusion tissue, and sheath-like elongate fiber-like cells (Fig. 78). Although often used as an important taxonomic character, the type of primary xylem maturation has been

intergradational and/or difficult to assess in many seed ferns, particularly Heterangium (Pigg, et al., 1987).Cycads are typically characterized as either marginally exarch or mesarch (Worsdell, 1906; Foster and Gifford,1974), or "weakly mesarch but near exarch (Bierhorst,1971). Additionally, Foster and Gifford (1974) have remarked that Cvcas has "both endarch and exarch xylem in the midrib bundle of the pinnae as well as in the vascular strands in the leaf rachis". In D. fremouwensis. protoxylem strands are difficult to identify, but reflect this general intergradation of marginally mesarch/near exarch. It may be that that character is of limited systematic use when looking at petioles. Perhaps however, more characteristic of cycadlike anatomy is the presence of abundant transfusion tracheids in the vascular strands of D. fremouwensis that resemble those found in cycad leaves (compare Fig. 69 with Worsdell, 1898, Fig. 16).
Lastly, the mesophyll of D. fremouwensis. like that of many if not most gymnospermous leaves, is well differentiated into palisade and spongy layers. The bundle sheath is composed of unusual fiber-like cells that have a sheath-like appearance but appear to lack simple pits or other ornamentation on their lateral walls. Resinous cells are scattered, and do not appear to have distinct positions in relation to vascular bundles as occurs among a few seed ferns (e.g., Feraxotheca. Millay and Taylor, 1978) and many

135conifers. Taken together, anatomical features of D. fremouwensis strongly suggest its affinities may be more closely with cycads than has previously been considered (e.g., Thomas, 1955; Petriella, 1981).
The anatomical description of D. fremouwensis provides the first basis upon which to compare anatomical details of the Mesozoic seed fern fronds of the corystosperms with leaves of the well known Paleozoic pteridosperms, the cycads, and other gymnospermous forms. Among pteridosperms, certain features are common to calamopiteans, most of the lyginopterids, some of the medullosans, and the corystosperms. These include the occurrence of pinnate fronds with a basal dichotomy, a two-parted ground tissue in larger frond members, sometimes differentiated into sparganum and/or dictyoxylon cortex, and usually multivascularized laminar pinnules (Galtier, 1974; Taylor and Millay, 1977; Pigg et al., 1987).Petriella (1981) summarized a number of features of corystosperms, lyginopterids, and medullosans (Table II of Petriella, 1981) including features of stem structure, anatomy and leaf trace; unfortunately the anatomical evidence he used for stem and leaf trace features of Dicroidium is unclear. However, the association of the unusual stem Rhexoxvlon (Bancroft, 1913; Walton, 1923,1925; Archangelsky, 1968a) led Archangelsky (1968a) to suggest it might represent the stem that bore Dicroidium.

136Archangelsky and Brett (1961) illustrate structures in the cortex of Rhexoxvlon stems which they designate as leaf traces (i.e., Fig. 19, 20 of Archangelsky and Brett, 1961), which may be the evidence used by Petriella for his discussion, as well as his (1978) reconstruction of Dicroidium. It is important to underscore, though, that at the present time, the relationship between Rhexoxvlon and Dicroidium remains unclear, and reconstructions of this type are equivocal.
Although Dicroidium foliage at Fremouw Peak has yet to be found in attachment to its parent stem, vegetative leaves of this type occur together with a fertile frond bearing pollen organs of the Pteruchus type with Alisnorites and Pteruchopol1enites pollen (De Vore and Taylor, 1988). The similarities in frond organization and histological features of vegetative foliage described herein and this fertile material, suggest strongly that they may represent the same plant. Also associated with this material are several types of ovules (Perovich and Taylor, 1988; Taylor and Taylor, 1987a; Taylor, Taylor and Millay, 1987), some of which occur in cupulate structures. Considering the rich floristic diversity at Fremouw Peak, it is difficult to assign many of the various disarticulated plant organs to a particular group, however, as similar histological features may reflect either generalized responses to the environment of unrelated

137groups or conservative tissue types occurring in a larger plexus. Until either organic attachments or distinctive histological features comparable in value to the capitate glands of Lvqinooteris (Oliver and Scott, 1903), are found, reconstructions of the Dicroidium plant remain unfounded.
For some time, the relationships of the Mesozoic seed fern groups both among themselves, to Paleozoic seed ferns, to other gymnosperms and to the angiosperms have been difficult to assess (Thomas, 1955; Petriella, 1981? Taylor, 1981). The paucity of anatomically preserved material has contributed to the lack of resolution with which these groups are presently viewed. Studies such as the present study provide the potential to greatly expand the data base of anatomical information concerning these plants, and will hopefully provide a new dimension to our characterization of these groups.

CHAPTER IX - SYSTEMATIC SECTION
Glossopteris schopfii Pigg sp. nov. Diagnosis:Leaves 1.3-1. 8 cm wide (based on complete widths), estimated from partially preserved fragments up to 3.1 cm in width? largest fragment 6 . 8 cm long, leaves estimated up to 1 0 - 1 2 cm in length; oblanceolate; 1/w ratio estimated 4:1-3:1; apex obtuse to retuse; venation composed of prominent midrib up to 1.62 mm wide (10 leaves) of 4-5 parallel strands that anastomose only near apex; lateral strands anastomose to form narrow, elongate meshes ca. 3.8 ± .10 mm x 0.49 ± .53 mm (10 leaves); area of these meshes is 1.48 ± 0.60 (10 leaves). Veins arch from midrib at angle of 53.30 ± 1.85 (10 leaves); marginal (lateral) veins meet margin at angle of around 228. Vein interconnections include simple Y-shaped dichotomies and reticulations, X,H, and N-shaped connections, and more complex configurations resulting from combinations of these forms. Total vein concentration 27.3/cm2 (9 leaves), veinconcentration in central region 2 2 .2 /cm2 ( 1 0 leaves), marginal vein concentration 2 1 .0/cm2 ( 1 0 leaves).
In transverse section, lamina 0.3-0.7 mm thick in region of midrib, 0.2-0.3 mm thick laterally. Vascular
138

strands of midrib somewhat larger than laterals but of identical anatomy. Bundles mesarch to marginally exarch. Protoxylem strands with annular or helical wall thickenings; metaxylem strands mostly with scalariform wall thickenings but occasionally exhibiting circular bordered pits. Radially aligned tracheids 3-8 cells/row, with scalariform wall thickenings. Prominent lacuna presumably representing position of phloem 0 . 2 x 0.08 mm wide.Vascular strand surrounded by prominent bundle sheath composed of elongate cells; many cells with thickened walls with simple pits and internal dark, amorphous contents, representing fibers, 17 x 110 jum. Mesophyll relatively undifferentiated, lacking palisade and spongy layers, composed of apparently loosely organized, parenchymatous tissue. Hypodermis on both surfaces, of 1-2 cells in thickness, composed of cuboidal cells 25-30 nm wide, frequently containing amorphous, dark material.
Upper epidermis lacking stomata; leaves hypostomatic. Epidermal cells elongate, 53 x 35 f m, straight-margined, frequently with centrally occurring papilla 7.5 nm in diameter. Lower epidermis with simple stomata arranged parallel to long axis of leaf but not in regular rows, ca. 39.2 stomata/mm2. Stomata slightly sunken; surrounded by 4-5 subsidiary cells 65 x 35 fm together comprising loosely organized stomatal complex: subsidiary cells not

140specialized. Guard cells 43 x 10 fmj with thickened margins; stoma elongate, 25 x 2.5 im.
Holotvoe; Specimen G1.21, 435 #4a, Figs: 6 , 9, 11-20. Specimens are deposited in the Antarctic Paleobotanical Collections, Orton Museum of Geology, The Ohio State University.Paratvpes: Specimens in 452; 483, 533. Figs. la, b, 2, 3,8 , 10. Specimens are deposited in the Antarctic Paleobotanical Collections, Orton Museum of Geology, The Ohio State University.Etymology: This species is named in honor of the late Dr. James M. Schopf in recognition of his significant contributions to Antarctic paleobotany.
Glossopteris skaarensis Pigg sp. nov. Diagnosis: Leavesup to 2 . 2 cm wide based on specimens with complete widths, estimated up to 2.7 cm based on fragmentary leaves with at least 1/2 of width; largest specimen 3.5 cm long, leaves estimated up to 1 0 - 1 2 cm in length; leaves probably oblanceolate, some taper proximally; apex retuse, sometimes asymmetric. Venation composed of a prominent midrib up to 1 . 1 mm wide, composed of a single broad strand, persisting to the apex where several strands anastomose; lateral strands anastomose to form broad, polygonal (pentagonal) meshes 2.86 ± .77 x.060 ± 0.16 mm2 (7 leaves); mesh area

1411.44 ± 0.61 mm2 (7 leaves). Veins arch from midrib at angle of 57.8 ± 1.48° (7 leaves); marginal (lateral) veins meet margin at approximately right angles. Total vein concentration ca. 2 1 .1/cm2; vein concentration in central region of leaf ca. 13.7/cm2, marginal vein concentration ca. 19/cm2.
In transverse section, lamina 0.5 mm thick in region of midrib, 0.13 - 0.14 mm thick laterally. Vascular strand of midrib appreciably larger than lateral veins but of similar histology. Bundles oval, terete, with ca. 20 tracheids, with helical or more commonly, scalariform wall thickenings. Vascular strands contained in bundles surrounded by 2-3 layers of bundle sheath composed of thin-walled, elongate cells sometimes containing dark amorphous material; fibers lacking. Mesophyll relatively undifferentiated, lacking palisade and spongy layer differentiation, composed of apparently loosely organized, parenchymatous tissue. Hypodermis prominent on both surfaces; 3-4 cells thick; composed of cuboidal cells 48 (36-57) Mm wide, frequently containing amorphous, dark material 16 Mm across.
Epidermal cells with sinuous anticlinal walls; leaves hypostomatic. Epidermal cells with rough, bumpy surfaces sometimes with 2-3 small papillae ca. 1.4 Mm in diameter. Cells overlying veins elongate. Epidermal cells 45 Mm wide x 24 mm high. Sunken stomata surrounded by 5-6 subsidiary

142cells organized in a ring. Stomata frequently contiguous, sharing 1-3 subsidiary cells with adjacent stoma or stomata. Subsidiary cells of two types: blunt rectangular cells 40 x 28 nm, elongate cells with sinuous margins 59 x 30 jLtm. Subsidiary cells with abaxially elongate beaklike projections 12 x 7 fm that extend over stomata. Stomatal complex up to 118 /Ltm in diameter. Guard cells somewhat thickened, stoma 9 /m wide, guard cells 34 x 9 jum; stomata randomly distributed and irregularly oriented? stomata concentration ca. 90.6/mm2.
Holotype: 465 C3 and C4, Fig. 33, 38-45. Specimens are deposited in the Antarctic Paleobotanical Collections,Orton Museum of Geology, The Ohio State University. Paratypes: Specimens in 452, 533, 484, Figs. 28a, c, 29,32, 34, 37. Slides 0,000-0,000 in the Antarctic Paleobotanical Collections, Orton Museum of Geology, The Ohio State University.Etymology: The species is named skaarensis in reference to its type locality, Skaar Ridge, in the Central Transantarctic Mountains.
Dicroidium fremouwensis. sp. nov. Pigg. Bifurcating frond at least 9 x 4.6 cm, with bluntly elongate, entire pinnae basally, 1.5 x 0.7 cm? longer, bipinnatifid pinnae more distally with 3-10 rounded lobes/pinna? longest pinnae

1431 . 6 x 0.7 cm with 10 lobes; pinnae form right angles with pinna rachis, laminae broadly attached to rachis. Pinna rachis thick, 3.6 mm, and of constant diameter throughout length of frond. Proximal-most petiole 5 mm in diameter; bifurcation results in two branches somewhat narrower than original axis, (3.6 mm); venation of pinnae mostly odontopteroid type with 1-3 veins entering central part of pinnule; secondary veins extending at steep angles of approximately 75-78°, most dichotomizing distally; venation symmetric in pinnules; 8 veins/cm2 in central region of pinnule (adjacent to midvein); 16 veins/cm2 at pinnule margin.
Frond rachis 3.5 mm high by 2.5 mm wide, broadly triangular-ovate in transverse section, vascularized by 15-20 bundles arranged in two groups: abaxial group of 7-8bundles arranged in a ring and adaxial group of 5-8 bundles arranged linearly; bundles of abaxial group extending the length of the rachis, decreasing to 3 distally; adaxial bundles diverging laterally to vascularize pinnae. Primary xylem exarch-marginally mesarch, with 7-10 tracheids/group; tracheids of the primary xylem with helical and scalariform-reticulate wall thickening patterns; primary xylem subtended by 3-8 rows of radially aligned tracheids, tracheids up to 23 fm in diameter with reticulate-scalariform wall thickenings; short, barrel-shaped cells 82 x 45 jLcm with reticulate wall

144thickenings (=transfusion tissue) occurring adjacent to tracheids; xylem subtended abaxially by 4-6 rows of poorly defined, radially aligned cells 8 fm in diameter, representing secondary phloem; vascular bundle sheath 1-3 cells thick, composed of elongate cells 2 1 0 fm long x 6 /m in diameter with thickened walls, lacking any pitting or wall thickening patterns; short, barrel-shaped cells adjacent to bundle sheath.
Ground tissue of frond rachis composed of inner parenchymatous, loosely organized zone and outer zone of smaller, more cuboidal cells, many cells with dark contents; randomly occurring, elongate resinous cells 72 fxm in diameter, sparsely distributed. Laminar pinnae with well-differentiated mesophyll with palisade and spongy layers; palisade layer of 1 - 2 cell layers occurring adaxially, cells 110 fm long x 40 fm across.
Leaves amphistomatic, stomata infrequent on upper epidermis, epidermal cells polygonal (32 x 42 fj.m) to elongate (84 x 23 fm), with straight anticlinal margins; stomata with dicyclic, incomplete ring of subsidiary cells (="pseudosyndetochelic" of Retallack, 1977b); guard cells thickened, 23 x 9 nm; surrounding stomatal opening 14 x 5 fm; subsidiary cells 26 x 32 fm.
Holotype: Specimen D21 on 10200 C3 Surface, and paradermal pull series; Fig. 58d, 76-82. Specimens are deposited in

the Antarctic Paleobotanical Collections, Orton Museum of Geology, The Ohio State University.Paratypes; Specimen D1 on 10137 A Surface, Fig. 58a, 59? Specimen D8 on 10109 B2 Surface, B2 A Side B, B2 C Side B, Fig. 58c, 61, 72. Specimen D14 on 10235 B2, Fig. 58b, 62; Specimen D15 on 10235 B2, Fig. 58e, 63. Specimens are deposited in the Antarctic Paleobotanical Collections, Orton Museum of Geology, The Ohio State University. Etymology: The specific epithet, fremouwensis. refers to Fremouw Peak, the locality in the Central Transantarctic Mountains from which the material was obtained.

146
LITERATURE CITEDAlvin, K., and W. G. Chaloner. 1970. Parallel evolution
in leaf venation: an alternative view of angiospermorigins. Nature 226: 662-663.
Anderson, H. M., and J. M. Anderson. 1970. A preliminary review of the biostratigraphy of the uppermost Permian, Triassic and lowermost Jurassic of Gondwanaland. Palaeont. afr. 13: 1-22.
Anderson, J. M. 1987. Evolutionary patterns inPermo-Triassic plant life. XlVth International Botanical Congress. Berlin. Int. Bot. Congr. Abstr. 17: 285.
, and H. M. Anderson. 1983. Palaeoflora of SouthernAfrica Molteno Formation (Triassic). Volume 1. A. A. Balkema. Rotterdam.
, and _____. 1985. Palaeoflora of Southern Africa.Podromus of South African megafloras. Devonian to Lower Cretaceous. A. A. Balkema. Rotterdam.
Antevs, E. 1913. Results of Dr. E. Mojberg's Swedish scientific expeditions to Australia 1910-1913. V.Some Mesozoic plants. K. svenska Vetensk.-Akad.Handl. 52: 1-6.
. 1914. Die Gattungen Thinnfeldia. Ett. undDicroidium Goth. K. svenska Vetensk.-Akad. Handl. 51: 1-71.
Appert, O. 1977. Die Glossopterisflora der Sakoa inSudwest-Madagascar. Palaeontographica 162B: 1-50.
Arber, E. A. N. 1902. The Clarke collection of fossilplants from New South Wales. Quart. J. geol. Soc. S. Afr. 58: 1-26.
. 1905. Catalogue of the fossil plants of theGlossbpteris flora in the Department of Geology, British Museum (Natural History). Br. Mus. Nat. Hist. London. 255p.

147 . 1913. On the earlier Mesozoic floras of New
Zealand. Proc. Cambridge Phil. Soc. 17: 122-131. . 1917. The earlier Mesozoic floras of New Zealand.
New Zealand Geol. Surv., Palaeont. Bull. 6 .Wellington. 80p.
Archangelsky, S. 1957. Las Glossopterideas del Bajo de la Leona. Rev. Assoc. Geol. Argentina 12: 135-164.
. 1968a. Studies on Triassic fossil plants fromArgentina. IV. The leaf genus Dicroidium and its possible relation to Rhexoxvlon stems. Palaeontology 11: 500-512.
. 1968b. Palaeobotany and palynology in SouthAmerica: a historical review. Rev. Palaeobot.Palynol. 7: 249-266.
. 1984. Floras neopaleozoicas del Gondwana y suzonacion estratigrafica. Aspectos paleogeograficos conexos. Commun. Ser. Geol. Portugal 70: 135-150.
. 1985. Aspectos evolutivos de las coniferasgondwanicas del Paleozoico. Bull. Section des Sciences, 1985, 8 : 115-124.
, and 0. G. Arrondo. 1969. The Permian taphoflorasof Argentina with some considerations about the presence of "northern” elements and their possible significance, in: A. J. Amos. Gondwana stratigraphy (1st IUGS Gondwana Symp., Buenos Aires) 71-89.
, and ____ . 1975. Paleogeografia y plantas fosilesen el Permico inferior Austrosudamericano. Actas I Congreso Argentino de Paleontol. y Biostrat. 1: 479-496.
, and D. W. Brett. 1961. Studies on the Triassicfossil plants from Argentina. 1. Rhexoxvlon from the Ischigualasto Formation. Phil. Trans. Roy. Soc. London. 244B? 1-19.
_, and __________. 1963. Studies on Triassic fossil plantsfrom Argentina. II. Michelilloa waltonii nov. gen. et spec, from the Ischigualasto formation. Ann. Bot. 27: 147-153.
, and R. Cuneo. 1986. Corynepteris australis sp.nov., primer registro de una coenopteridal en el Permico inferior de Chubut, Argentina. Actas. IV Congr. Arg. Pal. Bioestrat. Mendoza 1: 177-185.

148
, and _____. 1987. Ferugliocladaceae, a new coniferfamily from the Permian of Gondwana. Rev. Palaeobot. Palynol. 51: 3-30.
, and R. H. Wagner. 1983. Glossopteris anatolica sp.nov. from uppermost Permian strata in south-east Turkey. Bull. Br. Mus. nat. Hist. (Geol). 37:81-91.
Arnott, H. J. 1959. Anastomoses in the venation of Ginkgo biloba. Amer. J. Bot. 46: 405-411.
, and S. S. Tucker. 1963. Analysis of petal venationin Ranunculus repens v. pleniformis. Amer. J. Bot.50: 821-830.
Asama, K. 1969. Parallelism in Palaeozoic plants between Gondwanaland and Cathaysia land. Gondwana stratigraphy IUGS Symposium Buenos Aires, 1-15 October 1967. 127-136.
. 1976. Gigantopteris flora in southeast Asia and itsphytopalaeogeographic significance. Tokyo natl. sci. mus. bull. 1 2 .
Ash, S. R. 1979. Skilliostrobus gen. et sp. nov. a new lycopsid cone from the early Triassic of Australia. Alcheringa 3: 73-89.
. 1979-1980. Glossopterid leaves from the earlyMesozoic of Northern Mexico and Honduras. Palaeobotanist 28-29: 201-206. (issued 1981).
Axelrod, D. I. 1984. An interpretation of Cretaceous and Tertiary biota in polar regions. Palaeogeogr., Palaeoclimatol., Palaeoecol. 45: 105-148.
Baghai, N. L. 1988. Liriodendron (Magnoliaceae) from the Miocene Clarkia flora of Idaho. Amer. J. Bot. 75: 451-464.
Baldoni, A. M. 1980. Revision de las especies del genero Xvlopteris (Corystospermaceae) en el Triasico de Argentina, Australia y Sudafrica. Ameghiniana 17: 135-155.
Bancroft, N. 1913. Rhexoxvlon africanum. a newmedullosan stem. Trans. Linn. Soc. London (Bot.) 8 : 87-103.

149Banerjee, M. 1978. Genus Glossonteris Brongniart and its
stratigraphic significance in the Palaeozoics of India. Part 1 — a revisional study of some species of the genus Glossooteris. Bull. Botan. Soc. Bengal 32: 81-125.
, and A. K. Ghosh. 1970. Further studies on thegenus Glossopteris Sternb. and its stratigraphic significance. Second Gondwana Symposium Proc. and Papers South Africa, 1970: 561-570.
Banerji, J., and M. N. Bose. 1975. Some Lower Triassic plant remains from Asansol region, India. Palaeobotanist 24: 202-210. (issued 1977).
, H. K. Maheshwari, and M. N. Bose. 1974. Some plantfossils from the Gopad River section near Nidpur, Sidh District, Madhya Pradesh. Palaeobotanist 23: 59-71.(issued 1976).
, and Y. Lemoigne. 1987. Significant additions tothe Upper Triassic flora of Williams Point, Livingston Islands, South Shetlands (Antarctica). Geobios 20 (4): 469-487.
Barghoorn, E. S. 1961. A brief review of fossil plants of Antarctica and their geologic implications. Chapter 1 in Nat. Acad. Sci. Pub. 839: 5-9.
Barrett, P. J. 1969. Stratigraphy and petrology of the mainly fluviatile Permian and Triassic Beacon rocks, Beardmore Glacier area, Antarctica. Ohio State Univ. Inst. Polar Studies Rept. 34: 1-132.
, and D. H. Elliot. 1973. Reconnaissance geologicmap of the Buckley Island Quadrangle, Transantarctic Mountains, Antarctica. U. S. Geol. Survey Map A-3.
Basinger, J. F., and G. W. Rothwell. 1977. Anatomically preserved plants from the Middle Eocene (Allenby Formation) of British Columbia. Can. J. Bot. 55: 1984-1990.
, _____, and W. N. Stewart. 1974. Caulinevasculature and leaf trace production in medullosan pteridosperms. Amer. J. Bot. 61: 1002-1015.
Beeler, H. E. 1983. Anatomy and frond architecture of Neuropteris ovata and N. scheuchzeri from the Upper Pennsylvanian of the Appalachian Basin. Can. J. Bot. 61: 2352-2368.

150Bernardes-de-Oliveira, M. E. C. 1978. Ensalo sobre a
utilizacao de carachetes biometricos das glossopterodofitas em estratigrafla. Bol. Inst. 6 eoci.f Univ. Sao Paulo, 1978 9: 91-95.
Bierhorst, D. W. 1971. Morphology of vascular plants. Macmillan, N. Y. 560 p.
Blanc-Louvel, C. 1966. Etude anatomique comparee des tiges et des petioles d'une pteridospermee du Carbonifere du genre Lvainopteris Potonie. Mem. Mus. Nat. Hist. Nat. Ser C 18: 103 pp.
Bochenski, T. 1957. Preliminary report of the appearance in the Bunter Sandstone in borehole Radoszyce 3 of a plant of the Glossopteris type— Glossopteridium J. Czarnockii, n. sp. Warsaw Inst. Geol. Bull. 124: 189-191.
Boersma, M., and L. M. Broekmeyer. 1986, Index of figured plant megafossils — Triassic 1976-1980. Rev. Palaeobot. Palynol. 49: 235-344.
, and . 1987. Index of figured plantmegafossils — Permian 1976-1980. Rev. Palaeobot. Palynol. 53: 11-140.
Bonetti, M. I. R. 1966. Consideraciones sobre algunos representantes de la familia Corystospermaceae. Ameghiniana 4: 389-395.
Bortoluzzi, C. A., M. Guerra-Sommer, and M.Cazzulo-Klepzig. 1985. A tafoflora Triassica da Formacao Santa Maria, RS, Brasil; III.Pesquisas— Instit. Geoscienc. Univer. Fed. do Rio Grande do Sul 17: 215-232.
Bose, M. N., and J. Banerji. 1974. Some fragmentary plant remains from the Lower Triassic of Auranga Valley, District Palamau. Palaeobotanist 23: 139-144.(issued 1976).
, _____, and P. K. Maithy. 1975. Some fossil plantremains from Ramkola-Tatapani Coalfields, Madhya Pradesh. Palaeobotanist 24: 108-117. (issued 1977).
Brashier, C. K. 1968. Vascularization of cycad leaflets. Phytomorphology 18: 35-43.
Brongniart, A. 1828. Podrome d'une histoire des vegetaux fossiles. Paris.

151 . 1872. Notice sur le Psaronius brasiliensis. Bull.
Soc. Botan. France 19: 3-10.Bunbury, C. J. F. 1861. Notes on a collection of fossil
plants from Nagpur, Central India. Q. J. Geol. Soc. 17: 325-349.
Burnham, R. J. 1986. Foliar morphological analysis of the Ulmoideae (Ulmaceae) from the early Tertiary of western North America. Palaeontographica 20IB: 135-167.
Carpentier, A. 1935. Etudes paleobotaniques sur le groupe de la Sakoa et le groupe de la Sakamena (Madagascar). Annals geol. Serv. Mines. Madagascar 5: 1-32.
Carruthers, W. 1872. Notes on fossil plants fromQueensland, Australia. Q. Jl. geol. Soc. Lond. 28: 350-359.
Chabot, B. F., and J. F. Chabot. 1977. Effects of light and temperature on leaf anatomy and photosynthesis in Fraoaria vesca. Oecologia 26: 363-^377.
, and D. J. Hicks. 1982. The ecology of leaf lifespans. Ann. Rev. Ecol. Syst. 13: 229-259.
Chaloner, w. G., and W. S. Lacey. 1973. The distribution of late Palaeozoic floras. Spec. Pap. Palaeontol. 12: 271-289.
, K. U. Leistokow, and A. Hill. 1979. Brasilodendrongen. nov. and B. pedroanum (Carruthers) comb, nov., a Permian lycopod from Brazil. Rev. Palaeobot. Palynol. 28: 117-136.
Chandra, S, and K. R. Surange. 1979. Revision of the Indian species of Glossopteris. Monograph No. 2. Birbal Sahni Institute of Palaeobotany. Lucknow, India. 291 p.
Cichan, M. A. 1985. Vascular cambium and wood development in Carboniferous plants. I. Lepidodendrales. Amer.J. Bot. 72: 1163-1176.
. 1986. Conductance in the wood of selectedCarboniferous plants. Paleobiology 12: 302-310.
, and T. N. Taylor. 1982. Wood-borings inPremnoxvlon: plant-animal interactions in theCarboniferous. Palaeogeogr. Palaeoclimatol., Palaeoecol. 39: 123-127.

152
Colbert, E. H. 1973. Wandering lands and animals. E. P. Dutton and Co. New York.
Collinson, J. W., and J. Isbell. 1986. Permo-Triassic sedimentology of the Beardmore Glacier region. Ant.J. U. S. (1986 Review): in press.
Crane, P. R. 1985. Phylogenetic analysis of seed plants and the origin of angiosperms. Ann. Missouri Bot. Gard. 72: 716-793.
Cranwell, L. M. 1968. Palynological intimations of some Preoligocene Antarctic climates, in: E. M. vanZinderen Bakkeer, ed. Palaeoecology of Africa and the surrounding islands and Antarctica. 5: 1-19.
Crepin, F. 1875. Note sur le Pecooteris odontopteroides Morris. Bull. Acad. roy. des Sciences de Bruzelles 2e Series 39: 258-263.
Cridland, A. A. 1963. A Glossopteris flora from the Ohio Range, Antarctica. Amer. J. Bot. 50: 186-195.
Cuneo, R. 1987. Phytopaleoecology of the Rio Genoa formation at Lomas Chatas, Permian of Chubut, Argentina. Ameghiniana 24: 3-15.
Curran, J. M. 1884. On some fossil plants from Dubbo, New South Wales. Proc. Linnean Soc. N. S. W. 9: 250-254.
Dana, J. D. 1849. Fossil plants, Appendix 1, in UnitedStates Exploring Expedition during the years 1838-1842 under the Command of Charles Wilkes, U. S. N., C. Sherman, Philadelphia 10: Geology pp. 714-720.
Darrah, W. C. 1936. Discussion: Antarctic fossil plants.Science 83: 390-391.
. 1939. Textbook of paleobotany. D.Appleton-Century, N. Y.
deJersey, N. J. 1968. Palaeobotany and palynology in Australia: a historical review. Rev. Palaeobot.Palynol. 6 : 111-136.
Delevoryas, T. 1969. Glossopterid leaves from the Middle Jurassic of Oaxaca, Mexico. Science: 895-896.

153 . 1973. Postdrifting Mesozoic floral evolution, in:
B. J. Meggers, E. S. Ayensu, and W. D. Duckworth, eds. Tropical forest ecosystems in Africa and South America: a comparative review. Smithsonian Inst.Press, Washington, D. C. pp. 9-19.
. 1975. Mesozoic cycadophytes. in: K. S. W.Campbell, ed. Gondwana geology. Papers from the Third Gondwana Symposium, Canberra, Australia, 1973. Australian National University Press. p. 173-191.
, and R. E. Gould. 1971. An unusual fossilfructification from the Jurassic of Oaxaca, Mexico. Amer. J. Bot. 58: 616-620.
, and A. D. Hope. 1971. A new Triassic cycad and itsphyletic implications. Postilla 150: 1-21.
, and C. P. Person. 1975. Mexialossa varia gen. etsp. nov., a new genus of glossopteroid leaves from the Jurassic of Oaxaca, Mexico. Palaeontographica 154B: 114-120.
De Vore, M. L., and T. N. Taylor. 1988. Mesozoic seed plants: a pollen organ from the Triassic of Antarctica. Amer. J. Bot. 75: 106.
Dilcher, D. L. 1974. Approaches to the identification of angiosperm leaf remains. Bot. Rev. 40: 1-157.
DiMichele, W. A., T. L. Phillips, and R. G. Olmstead.1987. Opportunistic evolution: abiotic environmentalstress and the fossil record of plants. Rev.Palaeobot. Palynol. 50: 151-178.
Dolianiti, E. 1946. Noticia sobre novas formas na "Flora de Glossopteris" do Brasil Meridional. Brazil, Min. Agr. Dept. Nacl. Prod. Mineral., Div. Geol. Mineral., Nota Preliminaria 34: 1-6.
. 1954. A paleobotanica no Brasil. Brazil. Min.Agr., Dept. Nacl. Prod. Minrta;., Div. Geol. Mineral., Bol. 148: 7-56.
Dolph, G. E. 1976. Interrelationships among the gross morphological features of angiosperm leaves. 1976. Bull. Torrey Bot. Club 103: 29-34.
., and D. L. Dilcher. 1979. Variation in leaf sizewith respect to climate in the tropics of the Western Hemisphere. Bull. Torrey Bot. Club. 107: 154-162.

154Donn, W. L. 1982. The enigma of high-latitude
paleoclimate. Palaeogeogr., Palaeoclimatol., Palaeoecol. 40: 199-212.
Douglas, J. S. 1969. The Mesozoic floras of Victoria.Parts 1 and 2. Mem. geol. Surv. Viet. 28: 1-310.
Doyle, J. A., and M. J. Donoghue. 1986. Seed plant phylogeny and the origin of angiosperms: Anexperimental cladistic approach. Bot. Rev. 52: 321-431.
, and L. J. Hickey. 1976. Pollen and leaves from theMid-Cretaceous Potomac Group and their bearing on early angiosperm evolution, in: C. B. Beck, ed.Origin and early evolution of angiosperms. Columbia Univ. Press, pp.39-206.
du Toit, A. 1927. The fossil flora of the Upper Karroo Beds. Ann. South Afr. Mus. 22: 289-420.
. 1957. Our wandering continent. Edinburgh. Oliverand Boyd.
Edwards, W. N. 1952. Lvcopodiopsis: a southern hemispherelepidophyte. Palaeobotanist 1: 159-164.
Eggert, D. A. 1961. The ontogeny of Carboniferous arborescent Lycopsida. Palaeontographica 108B:43-92.
. 1962. The ontogeny of Carboniferous arborescentSphenopsida. Palaeontographica HOB: 99-127.
Elliot, D., E. Colbert, W. Breed, J. Jensen and J. Powell. 1970. Triassic tetrapods from Antarctica: Evidencefor continental drift. Science 169: 1197-1202.
Etheridge, R. 1890. Coal and plant-bearing beds of the Palaeozoic and Mesozoic Age in Eastern Australia and Tasmania with special reference to the fossil flora. Dept. Mine. Mem. Geol. Surv. N. S. W. Palaeontology, No. 3: 1-183.
. 1891. Description of some South Australia Silurianand Mesozoic fossils. Parliamentary papers relating to exploration, mining and geology. (Papers in coal bearing area in neighbourhood of Leigh Creek, etc.) 150: 9-14.

155 . 1902. More complete evidence of Thinnfeldia
odontooteroides Morris in the Leigh's Creek Coal Measures. Contrib. Palaeont. S. Aust. 12: 29.
Farabee, M. J., E. L. Taylor, and T. N. Taylor. 1988a. Palynological assemblages from the Falla Formation (Upper Triassic) of South Victoria Land, Antarctica. Amer.. J. Bot. 75: 108-109.
, _____, and _____ . 1988b. Pollen and sporeassemblages from the Falla Formation (Upper Triassic). Central Transantarctic Mountains, Antarctica. Rev. Palaeobot. Palynol., in press.
Feistmantel, O. 1878-1879. Palaeozoische und mesozoische Flora des ostlische Australien. Palaeontographica Cassel, suppl•, 3: 55-130? (1878), 130-195 (1879).
. 1880a. Fossil flora of the Gondwana System. Theflora of Damuda-Panchet divisions. Palaeont. indica, Mem. geol. Surv. India Ser 12, 3: 1-77.
. 1880b. Palaeontological notes from the Karharbariand South Rewah Coal-Fields. Rec. geol. Surv. India 13: 176-190.
. 1881. Fossil flora of the Gondwana system. Theflora of the Damuda-Panchet divisions. Palaeont. indica, Mem. geol. Surv. India, Ser. 12, 3: 78-149.
. 1882. The fossil flora of the Gondwana System;flora of the South Rewa Gondwana Basin. Mem. geol. Surv. India. Palaeont. indica, ser XII, 4: 1-52.
. 1886. The fossil flora of the Gondwana System. 2.The fossil flora of some of the Coalfields in Western Bengal. Palaeont. indica, Mem. geol. Surv. India, Ser 12, 4: 1-6.
. 1889. ubersichtliche Darstellung dergeologisch-palaeontologischen Verhaltnisse Sud-Afrikas. Th 1: Die Karroo-Formation und diedieselbe unterlagernden Schichten. Abh. K. Bohm. Ges. Wiss. Prague 7: 1-89.
. 1890. Geological and palaeontological relations ofthe coal and plant bearing beds of eastern Australia. Mem. geol. Surv. N. S. W. (palaeont). 3: 1-183.

156Florin, R. 1933. Studien uber die Cycadales des
Mesozoikums nebst erortungen uber die Spaltoffnungsapparate der Bennettiales. Kongl. svensk. Vetnsk. Hand. 12: 4-134.
Foster, A. S. 1966. Morphology of anastomoses in the dichotomous venation of Circaeaster. Amer. J. Bot.53: 588-598.
. 1968. Further morphological studies on anastomosesin the dichotomous venation of Circaeaster. J. Arn. Arb. 49: 52-72.
, and H. J. Arnott. 1960. Morphology and dichotomousvasculature of the leaf of Kinadonia uniflora. Amer. J. Bot. 75: 684-698.
, and E. M. Gifford. 1974. Comparative morphology ofvascular plants. 2d. ed. W. H. Freeman and Co. San Francisco. 751 p.
Francis, J. 1986. Growth rings in Cretaceous and Tertiary wood from Antarctica and their palaeoclimatic implications. Palaeontology 29: 665-684.
Frenguelli, J. 1941. Dicroidium stelznerianum (Gein) n. comb. Notas Mus. La Plata Paleontologia 6 : 393-403.
. 1943. Resena critica de los generos atribuidos ala 'serie de Thinnfeldia'. Revta Mus. La Plata n.s. (Paleont). 2: 225-342.
. 1950. Addenda a la flora del Gondwana superior enla Argentina II. Revta Asoc. geol. argent. 5: 15-30.
Galtier, J. 1974. Sur 1'organisation de la fronde desCalamopitvs. Pteridospermales probables du Carbonifere inferieur. C. R. Acad. Sc. Paris ser. D. 279: 975-978.
. 1986. Taxonomic problems due to preservation:comparing compression and permineralized taxa. in:R. A. Spicer and B. A. Thomas, eds. Systematic and taxonomic approaches in palaeobotany. The Systematics Association sp. vol. 31, pp. 1-16.
Gaudet, J. J. 1964. Morphology of Marsilea vestita. Leaves. Amer. J. Bot. 51: 591-597.

157Geinitz, H. B. 1876. Uber rhatischen Pflanzen und
Thierreste in den Argentinschen Provinzen. La Rioja, San Juan, uhnd Mendoza. Palaeontographica suppl. 3: 1-14.
Givnish, T. 1979. On the adaptive significance of leafform, in 0. T. Solbrig, et al., eds. Topics in plant population biology. Columbia Univ. Press, pp. 376-407.
. 1982. On the adaptive significance of leaf heightin forest herbs. Amer. Nat. 120: 353-381.
., and G. J. Vermeij. 1976. Sizes and shapes ofliane leaves. Amer. Nat. 110: 743-778.
Gordon, M., and R. W. Brown. 1952. Plantas triassicas do sol do Brasil. I: Estudas preliminares. Notasprelim. Estud. Div. geol. Miner. Bras. 54: 1-7.
Gothan, W. 1912. Uber die Gattung ThinnfeldiaEttinghausen. Abh. natur. Gesell. Nurnberg. 19:1-14.
Gould, R. E. 1970. Palaeosmunda. a new genus ofsiphonostelic osmundaceous trunks from the Upper Permian of Queensland. Palaeontology 13: 10-28.
. 1975a. A preliminary report on petrified axes ofVertebraria from the Permian of eastern Australia, in: K. S. W. Campbell, ed. Gondwana geology. Papers from the Third Gondwana Symposium, Canberra,Australia, 1973. Australian National University Press, p. 109-115.
. 1975b. The succession of Australian Pre-Tertiarymegafossil floras. Bot. Rev. 41: 453-483.
, and T. Delevoryas. 1977. The biology ofGlossopteris: evidence from petrified seed-bearingand pollen-bearing organs. Alcheringa 1: 387-399.
Graham, A. 1979. Literature on vegetation history, inLatin America. Supplement I. Rev. Palaeobot. Palynol. 27: 29-52.
. 1982. Literature on vegetation history in LatinAmerica. Supplement II. Rev. Palaeobot. Palynol.37: 185-223.

158Halle, T. G. 1912. On the geological structure and history
of the Falkland Islands. Bull. geol. Instit. Univ. Upsala. 11: 115-226.
Hamer, J., and G. W. Rothwell. 1988. The vegetative structure of Medullosa endocentrica Baxter (Pteridospermopsida). Can. J. Bot., in press.
Harris, T. M. 1932. The fossil flora of Scoresby Sound east Greenland. Part 3. Caytoniales and Bennettitales. Medd. om Gronland 85: 5-133.
Helby, R., and A. R. H. Martin. 1965. Cvclostrobus. gen. nov. cones of lycopsidean plants from the Narrabeen Group (Triassic) of New South Wales. Aust. J. Bot.13: 389-404.
, R. Morgan, A. D. Partridge. 1987. Apalynological zonation of the Australian Mesozoic, in: P. A. Jell, ed. Studies in Australian Mesozoicpalynology, Association of Australian Palaeontologists, Sydney, 1-94.
Herbst, L. A., L. A. Ferrando, and G. A. Jalfin. 1987.Florula Permica de Melo (Depto. Cerro Largo), Uruguay. Bol. Asoc. Latinoamericano Paleobot. Palin. 10: 1-5.
Herbst, R. 1975. On Osmundacaulis carnieri (Schuster) Miller and Osmundacaulis braziliensis (Andrews)Miller, in: K. S. W. Campbell, ed. Gondwana geology. Papers from the Third Gondwana Symposium, Canberra, Australia, 1973. Australian National University Press, p. 117-123.
Hickey, L. J. 1973. Origin of the major features ofangiospermous leaf architecture in the fossil record. Cour. Foursch.-Inst. Senckenberg 30: 27-34.
. 1979. 4. A revised classification of thearchitecture of dicotyledonous leaves, in: C. R.Metcalfe and L. Chalk, eds. Anatomy of the dicotyledons. Vol I. pp. 25-39. 2nd ed. Oxford University Press, Oxford.
, and J. A. Wolfe. 1975. The bases of angiospermphylogeny: vegetative morphology. Ann. Missouri Bot.Gard. 62: 538-589.
Hill, R. S. 1980. A numerical taxonomic approach to the study of angiosperm leaves. Bot. Gaz. 141: 213-229.

159Holmes, W. B. K. 1982. The Middle Triassic flora from
Benolong, near Dubbo, central-western New South Wales. Alcheringa 6 : 1-33,
, 1987. New corystosperm ovulate fructifications fromthe Middle Triassic of eastern Australia. Alcheringa 11: 165-173.
, and S. R. Ash. 1979. An early Triassic megafossilflora from the Lorne Basin, New South Wales. Proc. Linn. Soc. N. S. W. 103: 47-70.
Horn, H. S. 1971. The adaptive geometry of trees. Princeton University Press, Princeton. 144p.
Hsu, J. 1976. On the palaeobotanical evidence forcontinental drift and Himalayan uplift. Palaeobotanist 25: 131-143. (issued 1978).
. 1979. Glossopteris flora found in southern Tibetand continental drift. 9th Int. Cong. Carb. Strat. Geol.
Inamdar, J. A., K. N. Shendy, and N. V. Rao. 1983. Leaf architecture of some monocotyledons with reticulate venation. Ann. Bot. 52: 725-736.
Jack, R. L., and R. Etheridge. 1892. The geology and palaeontology of Queensland and New Guinea.Queensland Dept. Mines, Geol. Surv. Queensland Publ. 92, 768 pp.
Jackson, B. D. 1928. A glossary of botanic terms.London.
Jacob, K., and C. Jacob. 1950. A preliminary account of the structure of the cuticles of Dicroidium (Thinnfeldia) fronds from the Mesozoic of Australia. Proc. Nat. Inst. Sci. India 16: 101-126.
Jain, R. K., and T. Delevoryas. 1967. A Middle Triassic flora from the Cacheuta Formation, Minas de Petroleo, Argentina. Palaeontology 10: 564-589.
Jefferson, T. H. 1980. Angiosperm fossils in supposedJurassic volcanogenic shales, Antarctica. Nature 285: 157-158.
. 1982a. Fossil forests from the Lower Cretaceous ofAlexander Island, Antarctica. Palaeontology 25: 681-708.

160 . 1982b. The preservation of fossil leaves in
Cretaceous volcaniclastic rocks from Alexander Island, Antarctica. Geol. Mag. 119: 291-300.
Johnston, R. M. 1886. General observations regarding the classification of the upper Palaeozoic and Mesozoic rocks of Tasmania, together with a full description of all the known Tasmanian coal plants, including a considerable number of new species. Pap. Proc. R.Soc. Tasm. 1885: 343-387.
. 1887. Fresh contributions to our knowledge of theplants of Mesozoic age in Tasmania. Pap. Proc. R.Soc. Tasm. 1886: 160-182.
. 1888. A systematic account of the geology ofTasmania. Hobart, government printer. 408 pp.
. 1894. Further contributions to the fossil flora ofTasmania. Part 1. Pap. Proc. R. Soc. Tasm. 1893:170-178.
. 1896. Further contributions to the fossil flora ofTasmania. Part 2. Pap. Proc. R. Soc. Tasm. 1896: 57-63.
Jones, o. A., and N. J. DeJersey. 1947. The flora of the Ipswich Coal Measures— morphology and floral succession. Pap. Dep. Geol. Univ. Qd. n. s., 3:1-88.
Joy, K. W., A. J. Willis, and W. S. Lacey. 1956. A rapid cellulose acetate peel method in paleobotany. Ann. Bot. 20: 635-637.
Just, T. 1952. Fossil floras of the southern hemisphere and their phytogeographic significance. Bull. Amer. Mus. Nat. Hist. 99: 189-203.
Keating, R. C. 1968. Trends of specialization in the stipe anatomy of Dennstaedtia and related genera.Amer. Fern J. 58: 126-140.
Keeley, J. E. 1983. Crassulacean acid metabolism in the seasonally submerged aquatic Isoetes howellii. Oecologia 58: 57-62.
Kemp. E. M., and P. J. Barrett. 1975. Antarcticglaciation and early Tertiary vegetation. Nature 258: 507-508.

161Knoll, A. H., and 6. W. Rothwell. 1981. Paleobotany:
Perspectives in 1980. Paleobiology 7: 7-35.Kon1no, E. 1965. Some connections between the Cathaysian
flora and the Glossooteris flora in India during the later Permian Age. Palaeobotanist 14: 26-35.(issued 1966).
Kovdcs-Endrddy, E. 1976. Notes on some Glossopterisspecies from Hammanskraal (Transvaal) Palaeobot. afr. 19: 67-86.
. 1977-1978. The systematic position of the genusGanaamopteris McCoy, in: R. P. Stapleton, E. M.Beer, E. Kovdcs-Endrody, and W. G. Siesser. Papers on biostratigraphic research. Dept. Mines, Geol. Survey. South Africa. Bull 60: 19-71.
. 1977. The taxonomic status of the genus Rubidgea.Bothalia 12: 313-317.
. 1979. A re-evaluation of the venation structure ofGlossopteris. Ann. Geol. Surv. S. Afr. 12 (1977-1979): 107-141.
. 1981. "Broad-sense interpretation" of Glossopterisleaves: a critique. Palaeont. afr. 24: 35-38.
. 1983. Note on Glossopteris conspicua Feistmanteland on its assumed stratigraphic significance. Ann. Geol. Surv. Rep. S. Af. 1983. 17: 69-85.
. 1987. Clarification of the genus PalaeovittariaFeistmantel. Palaeontol. Afr. 26: 59-68.
Krausel, R. 1961. Lvcopodiopsis derbvi Renault und einige andere Lycopodiales aus dem Gondwana Schichte. Palaeontographica 109B: 61-92.
. 1962. Antarctic fossil wood, in E. P. Plumstead.Transantarctic expedition. 1955-1958, Sci. Rept. 9 (Geology) 133-154.
, P. K. Maithy, and H. K. Maheshwari. 1961.Gymnospermous woods with primary structures from Gondwana rocks— a review. Palaeobotanist 10: 97-107.
Kurtz, F. 1921. Atlas de plantas fosiles de la Republica Argentina. Actas Acad. Nac. Cienc. Cordoba 7:129-139.

162Kyle, R. A. 1977. Palynostratigraphy of the Victoria Group
of South Victoria Land, Antarctica. N. Z. J. of Geol. and Geophys. 20: 1081-1102.
, and A. Fasola. 1978. Triassic palynology of theBeardmore glacier area of Antarctica. Palinologia 1: 313-319.
, and J. M. Schopf. 1982. Permian and Triassicpalynostratigraphy of the Victoria Group, Transantarctic Mountains, in: C. Craddock, ed.Antarctic geosciences Univ. Press, Wisconsin.
Lacey, W. S. 1976. Further observations on the Molteno flora of Rhodesia. Arnoldia 7: 1-14.
> 1981. Revision of the Indian species ofGlossopteris. Book review, IOP Newsletter 14: 13-14.
., and R. C. Lucas. 1981. The Triassic flora ofLivingston Island, South Shetland Islands. Br. Antarct. Surv. Bull. 53: 157-173.
, D. E. van Dijk, and K. D. Gordon-Gray. 1975.Fossil plants from the Upper Permian in the Mooi River district of Natal, South Africa. Ann. Natal Mus. 22: 349-420.
Laetsch, W. M. 1974. The C4 syndrome: A structuralanalysis. Ann. Rev. Plant Physiol. 25: 27-52.
Lele, K. M. 1955. Plant fossils from Parsora in the South Rewa Gondwana Basin, India. Palaeobotanist 4: 23-34.
. 1961a. Studies in the Indian Middle Gondwanaflora: 1. On Dicroidium from the South Rewa GondwanaBasin. Palaeobotanist 10: 48-68. (issued 1962).
. 1961b. Studies in the Indian Middle Gondwana Flora-2. Plant fossils from the South Rewa Gondwana Basin. Palaeobotanist 10: 69-83. (issued 1962).
. 1974. Late Paleozoic and Triassic floras of Indiaand their relation to the floras of northern and southern hemisphere. Palaeobotanist 23: 89-115.(issued 1976).
1976. Palaeoclimatic implications of Gondwana floras. Geophytology 6 : 207-229.

163LeRoux, S. F., and H. M. Anderson. 1977. A review of the
localities and flora of the Lower Permian Karoo strata at Vereeniging, South Africa. Palaeontol. Afr. 20: 27-42.
Li, Xing-xue. 1983. Note on three new species ofGlossopteris of Qubu Formation, South Xizang, Tibet and discussion on age of formation. Acta Palaeontol. Sinica 22: 2.
, and Yao, Zhao-qi. 1985. Carboniferous and Permianfloral provinces in east Asia, in: J. T. Dutro andH. W. Pfefferkorn, eds. Neuvieme congres international de stratigraphie et de geologie du carbonifere. IX-ICC Compte rendu 5 (Paleontology, Paleoecology, Paleogeography): 95-101.
Little, R. J., and C. E. Jones. 1980. A dictionary of botany. Van Nostrand Reinhold Co. New York.
Maheshwari, H. K. 1965. Studies of the Glossopteris flora of India. — 31. Some remarks on the genus Glossopteris. Palaeobotanist 14: 36-45.
. 1967. Studies in the Glossopteris flora of India.— 34. On a record of Phvllotheca australis Brongn. from Jhara Coalfield, Bihar. Palaeobotanist 16: 167-169. (issued 1968).
. 1972. Permian wood from Antarctica and revision ofsome Lower Gondwana wood taxa. Palaeontographica 138B: 1-43.
. 1974. Floristics of the Permian and TriassicGondwanas of India. Palaeobotanist 23: 145-160.(issued 1976).
Maithy, P. K. 1965. Studies in the Glossopteris flora of India. — 32. On the genus Gangamopteris McCoy. Palaeobotanist 14: 46-51. (issued 1966).
. 1972a. A revision of the Lower GondwanaSphenopteris from India. Palaeobotanist 21:
70-80. (issued 1974). . 1972b. Dichotomopteris. a new type of fern frond
from the Lower Gondwana of India. Palaeobotanist 21: 365-367 (issued 1974).
. 1976. Further observations on Indian LowerGondwana SphenophyHales. Palaeobotanist 25:266-278. (issued 1978).

164
Mamay, S. H. 1960. Gigantopteridaceae in Permian floras of the southwestern United States. Geol. Surv. Resch. 1960: B380.
, J. M. Miller, and D. M. Rohr. 1984. LateLeonardian plants from West Texas: the youngestPaleozoic plant megafossils in North America. Science 223: 279-281.
, _____, , and W. E. Stein. 1986. Delnorte. anew genus of Permian plants. Phytologia 60: 345-346.
McCoy, F. 1874-1876. Podroraus of the paleontology of Victoria, or figures and descriptions of Victorian organic remains. Victoria Geol. Survey, 1st decade, pp. 1-43, pis 1-10 (1874), 2nd decade pp. 1-37, pis. 9-20 (1875), 4th decade, pp. 1-32, pis 31-40 (1876).
Medwell, L. 1954. A review and revision of the flora of the Victorian Lower Triassic. Proc. Roy. Soc. Viet, n. s. 65: 63-11.
Melville, R. 1969. Leaf venation and the origin of the angiosperms. Nature 224: 121-125.
. 1970. Links between the Glossopteridae and theangiosperms. Proc. Second Gondwana Symposium., C. S.I. R. South Africa 585-588.
. 1983. Glossopteridae, Angiospermidae and theevidence for angiosperm origin. Bot. J. Linn. Soc.8 6 : 279-323.
Mercer, J. H. 1983. Cenozoic glaciation in the southern hemisphere. Ann. Rev. Earth Planet Sci. 1983. 11:99-132.
Meyen, S. V., 1969. New data on relationship between Angara and Gondwana Late Palaeozoic floras in Gondwana Stratigraphy IUGS. Symposium, Buenos Aires. 1967. Paris. Unesco. 139-148.
. 1971. Parallelism and its significance for thesystematics of fossil plants. Geophytology 1: 34-47.
. 1977. Relation of Angara and Gondwana floras: acentury of controversies.
1984. Basic features of gymnosperm systematics and phylogeny as evidenced by the fossil record. Bot.Rev. 50: 1-111.

165
Mildenhall, D. C. 1976. Glossopteris ampla Dana from New Zealand Permian sediments. N. Z. J. Geol. Geophys.19: 130-132.
Millay, M. A. 1987. Triassic pteridophytes from Antarctica. Amer. J. Bot. 74: 710-711.
, and D. A. Eggert. 1974. Microgametophytedevelopment in the Paleozoic seed fern family Callistophytaceae. Amer. J. Bot. 61: 1067-1075.
, and T. N. Taylor. 1977. Feraxotheca gen. n., alyginopterid pollen organ from the Pennsylvanian of North America. Amer. J. Bot. 64: 177-185.
, and _____. 1978. Fertile and sterile frondsegments of the lyginopterid seed fern Feraxotheca. Rev. Palaeobot. Palynol. 25: 151-162.
_____, _____ , and E. L. Taylor. 1987. Studies ofAntarctic fossil plants: an association of ferns fromthe Triassic of Fremouw Peak. Ant. J. U. S. (in press).
Morris, J. 1845. Fossil flora, 245-254. in: P. E. DeStrezelecki, Physical description of New South Wales and van Diemans Land. Longman, Brown, and Green, London.
Mohr, B., and F. Schoener. 1985. Eine obertriassischeDicroidium Flora suedoestlich Alto del Carmen, Reion de Atacama (Chile). Neues Jahrbuch Geol. Palaeontol. Monatshefte 1985: 6 : 368-379.
Nair, N. C., and A. Das. 1978. The venation pattern in ferns. 3. Anastomoses and other features in Pteris vittata. Acta. Bot. Indica.
Napp-Zinn, K. 1973. Anatomie des Blattes, II.Blattanatomie der Angiospermen, A. topographische Anatomia des Angiospermenblattes. I. Lieferung, Handbuch der Pflenzenanatomie. Bd. VIII Teil 2A, Gebruder Borntraeger, Stuttgart.
Oestry-Stidd, L. L. 1979. Anatomically preservedNeuropteris rarinervis from American coal balls. J. Paleontol. 53: 37-43.
Oldham, R. D. 1897. On a plant of Glossopteris with part of the rhisome attached, and on the structure of Vertebraria. Rec. Geol. Surv. India 30: 45-50.

166
Oliver, F. W., and D. H. Scott. 1903. Laaenostoma lomaxi. the seed of Lvainodendron. Proc. Roy. Soc. London 71B: 477-481.
Orlando, H. A. 1968. A new Triassic flora from Livingston Island, South Shetland Islands. Brit. Antarctic survey Bull. 16: 1-13.
Osborn, J. M., and T. N. Taylor. 1988. Mesozoicsphenophytes from Antarctica. Amer. J. Bot. 75:113.
Pal, P. K. 1984. Triassic plant megafossils from the Tiki Formation, South Rewa Gondwana basin, India. Palaeobotanist 32: 253-309.
Pant, D. D. 1958. The structure of some leaves and fructifications of the Glossopteris flora of Tanganyika. Bull. Brit. Mus (Nat. Hist.) Geology 3: 127-175.
. 1977. The plant of Glossopteris. J. Indian Bot.56: 1-23.
, 1982. The Lower Gondwana gymnosperms and theirrelationships. Rev. Palaeobot. Palynol. 37: 55-70.
, and A. Choudhury. 1977. On the genus BelemnopterisFeistmantel. Palaeontographica 164B: 153-166.
______ and K. L. Gupta. 1968. Cuticular structure of someIndian Lower Gondwana species of Glossopteris Brongniart. Part 1. Palaeontographica 124B: 45-81.
, and _____. 1971. Cuticular structure of someIndian Lower Gondwana species of Glossopteris Brongniart. Part 2. Palaeontographica 132B:130-152.
., and P. F. Kidwai. 1968. On the structure of stemsand leaves of Phvllotheca indica Bunbury, and its affinities. Palaeontographica 121B: 102-121.
, and P. K. Khare. 1974. Damudopteris gen. nov.— anew genus of ferns from the Lower Gondwanas of the Raniganj Coalfield, India. Proc. R. Soc. London. B 186: 121-135.

167 , and L. Misra. 1977. On two genera of pteridophylls
Damudosorus gen. nov. and Trilithecopteris gen. nov. from the Lower Gondwanas of the Raniganj coal-field. Palaeontographica 164B: 76-86.
, _____, and D. D. Nautiyal. 1982. On the structureof stems and leaves of Schizoneura oondwanensis. Palaeontographica 183B: 1-7.
, and D. D. Nautiyal. 1967. On the structure ofBuriadia heterophylla (Feistmantel) Seward and Sahni and its fructification. Phil. Trans. Roy. Soc.London. (Biol. Sci.) 252B: 27-48.
, and _____. 1984. On the morphology and structureof Ottokaria zeilleri sp. nov.: a femalefructification of Glossopteris. Palaeontographica 193B: 127-152.
, and R. Pant. 1987. Some Glossopteris leaves fromIndian Triassic beds. Palaeontographica 205B:165-178.
, and K. B. Singh. 1968. On the genus GanqamopterisMcCoy. Palaeontographica 124B: 83-101.
, and _____. 1971. Cuticular structure of someIndian Lower Gondwana species of Glossopteris Brongniart. Part 3. Palaeontographica 135B:l-40.
, and _____. 1974. On the stem and attachment ofGanaamopteris and Glossopteris leaves, Part II. Structural features. Palaeontographica 147B: 42-73.
i and V. K. Singh. 1987. Xylotomy of some woods fromRaniganj Formation, Permian, Raniganj coalfield,India. Palaeontographica 203B: 1-82.
Parkhurst, D. F. 1986. Internal leaf structure: athree-dimensional perspective, in: T. J. Givnish,ed. On the economy of plant form and function. Cambridge University Press, Cambridge, pp. 215-246., and 0. L Loucks. 1972. Optimal leaf size in relation to environment. J. Ecol. 60: 505-537.

168Perinotto, J. A. J., and 0. Rosier. 1984. Glossopterideas
de Fluviopolis (Penniano superior) bo sul do Estado do Parana. Paleobot e palino na America do Sul: 1983: contr. IV reun. de paleobot e palin. in: J. Alexandre, J. A. J. Perinotto, and T. R. Fairchild, eds. Bol. IG Int. Geocien. Univ. Sao Paulo 15:90-96.
Perovich, N. E., and E. L. Taylor. 1988. Permineralizedovules from the Triassic of Antarctica. Amer. J. Bot. 75: 113-114.
Petriella, B. 1978. La reconstruccion de Dicroidium(Pteridospermopsida, Corystospermaceae). Obra Centen. Mus. La Plata 5: 107-110.
. 1979. Sinopsis de las Corystospermaceae(Corystospermales, Pteridospermophyta) de Argentina.I. Hojas. Ameghiniana 16: 81-102.
. 1980. Sinopsis de las Corystospermaceae(Corystospermales, Pteridospermophyta) de Argentina.II. Estructures fertiles. Ameghiniana 17: 168-180.
. 1981. Sistematica y vinculaciones de lasCorystospermaceae H. Thomas. Ameghiniana 18:221-234.
Phillips, T. L. 1980. Stratigraphic and geographicoccurrences of permineralized coal-swamp plats— Upper Carboniferous of North America and Europe, in: D. L.Dilcher and T. N. Taylor (eds.), Biostratigraphy of fossil plants— successional and paleoecological analysis, pp. 25-92. Dowden, Hutchinson, and Ross, Stroudsburg, PA.
Pigg, K. B. 1988. Corystosperms from the Triassic of Antarctica: Dicroidium. Amer. J. Bot. 75: 114.
, and G. W. Rothwell. 1985. Cortical development inChaloneria cormosa (Isoetales), and the biological derivation of compressed lycophyte decortication taxa. Palaeontology 28: 545-553.
, R. A. Stockey, and T. N. Taylor. 1985. Studies ofPaleozoic seed ferns: additional studies ofMicrospermopteris aphvllum Baxter. Bot. Gaz. 147: 126-136.
, and T. N. Taylor. 1985. Anatomically preservedGlossopteris from the Beardmore Glacier area of Antarctica. Ant. J. U. S. 19: 8-10.

169
, and . 1987a. Anatomically preservedGlossopteris from Antarctica. VII Simposio argentino de Paleobotanica y Palinologia Actas: 177-180.
, and . 1987b. Anatomically preservedDicroidium from the Transantarctic Mountains. Ant. J. U.S. (in press).
, , and R. A. Stockey, 1987. Studies ofPaleozoic seed ferns: Heterancrium kentuckvensis sp.nov., from, the Upper Carboniferous of North America. Amer. J. Bot. 74: 1184-1204.
Plumstead, E. P. 1952. Description of two new genera and six new species of fructifications borne on Glossopteris leaves from South Africa. Trans, geol. soc. S. Afr. 55: 281-328.
. 1956. Bisexual fructifications borne onGlossopteris leaves from South Africa. Palaeontographica 100B: 1-25.
. 1958a. Further fructifications of theGlossopteridiae. Trans, geol. Soc. S. Afr. 61:51-76.
. 1958b. The habit of growth of Glossopteridae.Trans. Proc. Geol. Soc. South Africa 61: 81-96.
. 1962. Fossil floras of Antarctica, in:Transantarctic Expedition Sci. Rep. 9 (geology):1-154.
. 1973. The Late Palaeozoic Glossopteris flora, in:A. Hallam, ed. Atlas of paleobiogeography. Elsevier Scientific Publ. N. Y.: 187-205.
. 1975. A new assemblage of plant fossils fromMilorgfjella, Dronning Maud Land. Brit. Antarctica Survey Sci. Rept. 83: 1-30.
Prasad, M. N. V. 1982. An annotated synopsis of Indian Palaeozoic gymnospermous woods. Rev. Palaeobot. Palynol. 38: 119-156.
. 1986. Xylotaphoflora of the Kamthi Formation,Indian Lower Gondwana, with remarks on the biostratigraphic importance of its taphoflora. Palaeontographica 201B: 111-134.

170Pray, T. R. 1955a. Foliar venation of angiosperms. II.
Histogenesis of the venation of Liriodendron. Amer.J. Bot. 42: 18-27.
. 1955b. Foliar venation of angiosperms. IV.Histogenesis of the venation of Hosta. Amer. J. Bot. 42: 698-706.
. 1960. Ontogeny of the open dichotomous venation inthe pinna of the fern Nephrolepis. Amer. J. Bot. 47: 319-328.
. 1961. Ontogeny of the closed dichotomous venationof Reanellidium. Amer. J. Bot. 49: 464-472.
Rao, A. R., and K. M. Lele. 1962. On the cuticle of Dicroidium (Thinnfeldia) Sahni (Seward) with some observations on the genera Thinnfeldia and Dicroidium. Palaeobotanist 11: 7-12 (issued 1963).
Rayner, R. J. 1985. The Permian lycopod Cyclodendronleslii from South Africa. Palaeontology 28: 111-120.
. 1986. Azaniadendron. a new genus of lycopod fromSouth Africa. Rev. Palaeobot. Palynol. 47: 129-143.
_____, and M. K. Coventry. 1985. A Glossopteris florafrom the Permian of South Africa. S. Afr. J. Sci. 81: 21-32.
Retallack, G. 1977a. The life and times of a Triassic lycopod. Alcheringa 1: 3-29.
. 1977b. Reconstructing Triassic vegetation ofeastern Australasia: a new approach for thebiostratigraphy of Gondwanaland. Alcheringa 1: 247-277, and Alcheringa-fiche 1: Gl-jl7,ISSN0311-5518.
. 1980. Late Carboniferous to Middle Triassicmegafossil floras from the Sydney Basin, in: C.Herbert and R. J. Helby, eds. A guide to the Sydney Basin. Bull. Geol. Surv. New South Wales 26:385-430.
, and D. L. Dilcher. 1981. Arguments for aglossopterid origin of angiosperms. Paleobiology 7: 54-67.

171t
Rex, G. 1986. Experimental modelling as an aid tointerpreting the original three-dimensional structure of compressions, in: R. A. Spicer and B. A. Thomas,eds. Systematic and Taxonomic Approaches in Palaeobotany. The Systematics Association sp. vol.31.
Rigby, J. F. 1966. The Lower Gondwana floras of the Perth and Collie Basins, Western Australia.Palaeontographica 118B: 113-152.
. 1967. On Ganaamonteris walkomii sp. nov. Rec.Australian Mus. 27: 175-182.
. 1969. Permian sphenopsids for Antarctica. Prof.Pap. U. S. geol. Surv. 613F, F1-F13.
. 1972. The Gondwana palaeobotanical province atthe end of the Palaeozoic. 24th IGC, 1972— Section 7: 324-330.
. 1978. Permian glossopterid and other cycadopsidfructifications from Queensland. Geol. Surv. Queensland Publ. 367 (Palaeontol. Paper 41): 1-21.
. 1983. The role of the Glossopteris flora inbiostratigraphy: a preliminary assessment in theReids Dome Beds. Geol. Surv. Queensl., Brisbane, Queensland. Geol. Soc. Aust. 221-229.
. 1984. The origin of the Glossopteris flora— somethoughts based on macrophyte remains. Evolu. Bot. and Biostr. (19-28). A. K. Ghosh Comm. Vol.:19-28.
. 1985. Some Triassic (middle Gondwana) floras fromSouth Victoria Land, Antarctica. Hornibrook Symposium, Dept. Sci. and Ind. Resch., New Zealand, Christchurch, New Zealand: 78-79.
%
, H. K. Maheshwari, and J. M. Schopf. 1980. Revisionof Permian plants collected by J. D. Dana during 1839-1840 in Australia. Queensland Dept, of Mines, Geol. Survey. Qd. Pub. 376, Pal. pap. 471-25.
, and J. M. Schopf. 1969. Stratigraphic implicationsof Antarctic paleobotanical studies. Gondwana stratigraphy earth sci. 2: 91-106.
Rodin, R. J. 1966. Leaf structure and evolution in American species of Gnetum. Phytomorphology 16:56-68.

172 . 1967. Ontogeny of foliage leaves in Gnetum.
Phytomorphology Panchanan Maheshwari Memorial Volume: 118-128.
Rosier, 0., and F. C. Fittipaldi. 1981. 0 paleobotanicono estudo e prospeccao de Carvao. Atasdo III Simposio Regional de Geologia da Socieda de Brasileira de Geologia, Nucleo de Sao Paulo (Curitiba, Parana, Setembro, 1981) 1: 274-283.
Rothwell, G. W. 1971. Ontogeny of the Paleozoic ovule,Callospermarion p u s ilium. Amer. J. Bot. 58: 706-715.
. 1975. The Callistophytaceae (Pteridospermopsida):1. Vegetative structures. Palaeontographica 15IB:171-196.
. 1977. Evidence for a pollination-drop mechanism inPaleozoic pteridosperms. Science 198: 1251-1252.
, and T. N. Taylor. 1972. Carboniferous pteridospermstudies: morphology and anatomy of Schopfiastrumdecussatum. Can. J. Bot. 50: 2649-2658.
Rowett, A. I. 1979. The megafossil flora of the LeighCreek Coal measures with emphasis on the Dicroidium - Xvlopteris - Johnstonia complex. BSc. Hons, thesis, Univ. Adelaide.
Sahni, B. 1923. On the structure of the cuticle inGlossopteris ancmstifolia Brongn. Rec. Geol Surv. India 54: 277-280.
. 1926. The southern fossil floras: a study in theplant geography of the past. (Pres. Ass.) Proc. 13th Ind. Sci. Cong. Bombay: 229-254.
. 1936. Wegener's theory of continental drift in thelight of palaeobotanical evidence. J. Indian Bot.Soc. 15: 319-332.
Scheihing, M. H., and H. W. Pfefferkorn. 1980. Morphologic variation in Alethopteris (Pteridosperms, Carboniferous) from St. Clair, Pennsylvania, USA. Palaeontographica 172B: 1-9.
Schopf, J. M. 1970a. Relation of floras of the southern hemisphere to continental drift. Taxon 19: 657-674.
1970b. Petrified peat from a Permian coalbed in Antarctica. Science 169: 274-277.

173 . 1971. Notes on plant tissue preservation and
mineralization in a Permian deposit of peat from Antarctica. Amer. J. Sci. 271: 522-543.
. 1973. The contrasting plant assemblages fromPermian and Triassic deposits in southern continents. Can. Soc. Petr. Geol. Calgary, Alta., 1973: 379-397.
. 1976. Morphologic interpretation of fertilestructures in glossopterid gymnosperms. Rev. Palaeobot. Palynol. 21: 25-64.
. 1978. An unusual osmundaceous specimen fromAntarctica. Can. J. Bot. 56: 3083-3095.
, and R. A. Askin, 1980. Permian and Triassic floralbiostratigraphic zones of southern land masses, in:D. L. Dilcher and T. N. Taylor (eds.).Biostratigraphy of fossil plants. Dowden, Hutchinson, and Ross, Stroudsburg, Pa.
Schwarzwalder, R. N. 1983. The use of multivariatestatistics in paleosystematics. Amer. J. Bot. 70 (Abstr.): 78.
Seward, A. C. 1897. On the association of Siaillaria and Glossopteris in South Africa. Quart. J. Geol. Soc. Lond. 53: 315-340.
. 1903. Fossil floras of Cape Colony. Ann. S. Afr.Mus. 4: 1-122.
. 1904. On collections of Natal fossil plants. II:From the Drakensberg range in west Natal. Report of the Geological survey of Natal and Zululand 2:101-104.
. 1908. On a collection of fossil plants from SouthAfrica. Q. Jl. geol. Soc. London. 64: 83-108.
. 1910. Fossil plants. Cambridge. 624p. . 1914. Antarctic fossil plants. Brit, antarct.
Terra Nova Exped. (Geol.) 1: 1-49. . 1932. On some fossil plants from the Parsora
Stage, Rewa. Rec. geol. Surv. India 6 6 : 235-243. , and R. E. Holttum. 1921. On a collection of fossil
plants from Southern Rhodesia. Bull. geol. Surv. Zth. Rhod. 8 : 39-45.

174 , and J. Walton. 1923. On a collection of fossil
plants from the Falkland Islands. Quart. J. Geol.Soc. London. 79: 313-333.
, and A. S. Woodward. 1905. Permocarboniferousplants and vertebrates from Kashmir. Palaeont. indica, Mem. Geol. Surv. India 2. Mem. No. 2.
Shadle, G. L., and B. M. Stidd. 1974. The frond of Heteranaium. Amer. J. Bot. 62: 67-75.
Shirley, J. 1898. Additions to the fossil flora of Queensland, mainly from the Ipswich Formation. Trias-Jura system. Bull. geol. Surv. Qd. 7:1-25.
Sitholey, R. V. 1954. Mesozoic and Tertiary floras of India— a review. Palaeobotanist 3: 55-69.
Slade, B. 1957. Leaf development in relation to venation, as shown in Cercis siliouastrum L., Prunus serrulata Lindl., and Acer pseudoplatanus L. New Phytol. 56:281-300.
Smith A. G. and A. Hallam. 1970. The fit of the southern continents. Nature 225: 139-144.
Smoot, E. L., and T. N. Taylor. 1985. Paleobotany:recent developments and future research directions. Palaeogeogr., Palaeoclimatol., Palaeoecol. 50:149-162.
, and . 1986a. Evidence of simple polyembryonyin Permian seeds from Antarctica. Amer. J. Bot. 73: 1079-1081.
, and _____. 1986b. Structurally preserved fossilplants from Antarctica: II. A Permian moss from theTransantarctic Mountains. Amer. J. Bot. 73:1683-1691.
/ _____, and J. W. Collinson. 1987. Lower Triassicplants from Antarctica: Diversity and paleoecology.VII Simposio argentino de Paleobotanica y Palinologia, Actas: 193-196.
/ _____, _____, and D. H. Elliot. 1986. Structurallypreserved Permian plants from Skaar Ridge, Beardmore Glacier region. Ant. J. U. S. (1986 Review): inpress.

175 , , and T. Delevoryas. 1985. Structurally
preserved fossil plants from Antarctica. I. Antarcticvcas. gen. nov., a Triassic cycad stem from the Beardmore Glacier area. Amer. J. Bot. 72: 1410-1423.
, and M. Vande Wege. 1986. Phloem anatomy inStauronteris biseriata from the Pennsylvanian of North America. Amer. J. Bot. 73: 1043-1048.
Solms-Laubach, H., and G. Steinman* 1899. Das Auftretenund die Flora der rhatischen Kohlenschichten von La Ternera (Chile). in: Stenmann, G. Beitrage zurGeologie und Palaeontologie con Sudamerika, VII.Neues JB Miner. Geol. Palaont. 12: 581-609.
Spicer, R. A., and C. R. Hill. 1979. Principal Components and Correspondence Analysis of quantitative data from a Jurassic plant bed. Rev. Palaeobot. Palynol. 28: 273-299.
Srivastava, A. K., and J. F. Rigby. 1983. Sphenophvllum.Trizygia. and Gondwanaphvton from Barakar formation of Raniganj coalfield, India with a revision of Lower Gondwana Sphenophyllales. Geophytology 13: 55-62.
Srivastava, P. N. 1956. Studies in the Glossopteris floraof India— 4. Glossopteris. Ganqamopteris. and Palaeovittaria from the Raniganj coalfield. Palaeobotanist 5: 1-45.
Srivastava, S. C. 1971. Some gymnospermic remains fromthe Triassic of Nidpur, Sidhi district, Madhya Pradesh. Palaeobotanist 18: 280-296.
Statistical Analysis Systems. 1985. SAS user's guide:statistics. Version 5 Edition. SAS Institute, Cary, NC.
Stebbins, G. L. 1974. Flowering plants: Evolution abovethe species level. Harvard University Press, Cambridge, Massachusetts.
Stidd, B. M., and T. L. Phillips. 1973. The vegetative anatomy of Schopfiastrum decussatum from the Middle Pennsylvanian of the Illinois Basin. Amer. J. Bot.60: 463-474.
Stubblefield, S. P., and T. N. Taylor. 1986. Wood decay in silicified gymnosperms from Antarctica. Bot. Gaz. 147: 116-125.

176 , and _____. 1987. Tansley review no. 12. Recent
advances in palaeomycology. New Phytol. 108: 3-25.Surange, K. R. 1966. The present position of the genus
Glossopteris. Proc. Autumn School Bot. Mahabaleshwar 1966: 316-327.
. 1975. Indian Lower Gondwana floras: a review.in: K. S. W. Campbell, ed. Gondwana geology, p. 135-147.
., and S. Chandra. 1971. Denkania indica gen. et sp.nov — A glossopteridean fructification from the Lower Gondwana of India. Palaeobotanist 20: 264-268.(issued 1973).
, and _____ . 1975. Morphology of the gymnospermousfructifications of the Glossopteris flora and their relationship. Palaeontographica 149B: 153-180.
, and _____ . 1976. Morphology and affinities ofGlossopteris. Palaeobotanist 25: 509-524. (issued1978).
., and K. M. Lele. 1956. Studies in the Glossopterisflora of India— 6 . Plant fossils from Talchir Beds of South Rewa Gondwana basin. Palaeobotanist 5:82-90. (issued 1957).
, and P. Singh. 1953. The female dwarf shoot ofWalkomiella indica— A conifer from the Lower Gondwanas of India. Palaeobotanist 2: 5-8.
, and P. N. Srivastava. 1956. Studies in theGlossopteris flora of India— 5. Generic status of Glossopteris. Gancramopterisr and Palaeovittaria. Palaeobotanist 5: 46-49.
Szajnocha, L. 1888. Uber fossile Pflanzenreste aus Cacheuta in der Argentinischen Republik. Sber. Akad. Wiss. Wien. Mathematisch— naturwissenschaftliche klasse 97: 219-245.
Taylor, E. L. 1987. Glossopteris reproductive organs: ananalysis of structure and morphology. XlVth International Botanical Congress, Berlin. Int. Bot. Congr. Abstr. 17: 287.
Taylor, T. N. 1978. The ultrastructure and reproductive significance of Monoletes (Pteridospermales) pollen. Can. J. Bot. 56: 3105-3118.

177 . 1981. Paleobotany. An introduction to fossil
plant biology. McGraw-Hill Book Company, New York. , and M. A. Millay. 1977. Morphologic variability of
Pennsylvanian lyginopterid seed ferns. Rev.Palaeobot. Palynol. 32: 27-62.
, and G. W. Rothwell. 1982. Studies of seed fernpollen: development of the exine in Monoletes(Medullosales). Amer. J. Bot. 69: 570-578.
, and A. C. Scott. 1983. Interactions of plants andanimals during the Carboniferous. BioScience 33: 488-493.
., E. L. Smoot, and J. W. Collinson. 1985.Paleoenvironment of Upper Triassic plants from the Fremouw Formation. Ant. J. U. S. (in press).
, and S. P. Stubblefield. 1987. A fossil mycoflorafrom Antarctica. VII Simposio argentino de Paleobotanica y Palinologia Actas 187-189.
, and E. L. Taylor. 1987a. An unusual gymnospermousreproductive organ of Triassic age. Ant. J. U. S. (in press).
, and _____. 1987b. Structurally preserved fossilplants from Antarctica. III. Permian seeds. Amer. J. Bot. 74: 904-913.
, _____, and M. A. Millay. 1987. Mesozoic seedplants from Antarctica: Multiovulate cupules. Amer.J. Bot. 74 (part 2): 691-692.
Tenison-Woods, J. E. 1883. On the fossil flora of thecoal deposits of Australia. Proc. Linnean Soc. N. S. W. 8 : 37-167.
Thomas, H. H. 1933. On some pteridospermous plants from the Mesozoic rocks of South Africa. Phil. Trans. Roy. Soc. London 222B: 193-265.
. 1952. A Glossopteris with whorled leaves.Palaeobotanist 1: Birbal Sahni Memorial Volume:435-438.
. 1955. Mesozoic pteridosperms. Phytomorphology 5:177-185.

178 . 1958. Lidaettonia. a new type of fertile
Glossopteris. Bull. Brit. Mus. (Nat. Hist.) Geol. London 3: 179-189.
Thompson, M. R. A., and R. W. Burn. 1977. Angiosperm fossils from latitude 70s S. Nature 269: 139-141.
Townrow, J. 1956. The genus Lepidooteris and its southern hemisphere species. Avh. norske Vidensk. Akad. Oslo Mat.-Naturv. Klasse. 2: 1-28.
. 1957. On Dicroidium. probably a pteridospermousleaf, and other leaves now removed from this genus. Trans, geol. Soc. Afr. 60: 21-60.
. 1962. On Pteruchus. a microsporophyll of the (Corystospermaceae. Bull. Br. Mus. (Nat. Hist.) Geol 6 : 280-320.
. 1967. Fossil plants from Allan and.CarapaceNunataks, and from the Upper Mill and shackleton Glaciers, Antarctica. N. Z. Jl. Geol. Geophys. 10: 456-473.
. 1968. A fossil Selaqinella from the Permian of NewSouth Wales. J. Linn. Soc. (Bot.) 61: 13-23.
Upchurch, G. R., and J. A. Wolfe. 1987. Mid-Cretaceous to Early Tertiary vegetation and climate: evidence fromfossil leaves and woods, in: E. M. Friis, W. G.Chaloner, and P. R. Crane, eds. The origin of angiosperms and their biological consequences. Cambridge University Press pp. 75-104.
Wagner, R. H. 1962. On a mixed Cathaysia and Gondwanaflora from S. E. Anatolia (Turkey). C. R. 4eme Congr. Strat. et Geol. Carb., 1 Heerlen (1958) 3: 745-752.
Wagner, W. H. 1979. Reticulate veins in the systematics of modern ferns. Taxon 28: 87-95.
, J. M. Beitel, and F. S. Wagner. 1982. Complexvenation patterns in the leaves of Selaqinella: Megaphyll-like leaves in lycophytes. Science 218: 793-794.
Walkom, A. B. 1917. Mesozoic floras of Queensland. Pt.1: The flora of Ipswich and Wallon Series (c)Filicales, etc.? (d) Ginkgoales; (e) Cycadophyta; (f) Coniferales, Pubis, geol. Surv. Qd. 257: 1-67.

179 . 1922. Palaeozoic floras of Queensland. Part 1.
The flora of the Lower and Upper Bowen series. Publ. geol. Surv. Qd. 270: 1-65.
. 1925. Notes on some Tasmanian Mesozoic plants.Part I. Pap. Proc. Roy. Soc. Tasmania for 1924:73-89.
Walton, J. 1923. On Rhexoxvlon Bancroft, a Triassic genus of plants exhibiting a liane-type of vascular organization. Philos. Trans. R. Soc. London 212B: 79-102.
. 1925. On some South African fossil woods. Ann. S.Afr. Mus. 22: 1-26.
. 1956. Rhexoxvlon and Dadoxvlon from the LowerShire region of Nyasaland and Portuguese East Africa, compared with previously described genera from Africa and other parts of the world. Colon. Geol. Min. Resour. 6 : 159-168.
Wegener, A. 1966. The origin of continents and oceans.J. Biram, tr. Dover Publ., N. Y.
White, D. 1908. Fossil flora of Coal-Measures of Brazil. Com. Est. Mines Carvao Brasil. Rel. Final. III. Rio de Janeiro.
White, J., and T. N. Taylor. 1988. A Triassic fungus from Antarctica with possible ascomycetous affinities.Amer. J. Bot., in press.
White, M. E. 1978. Reproductive structures of theGlossopteridales in the plant fossil collection of the Australian Museum. Rec. Austr. Mus. 31: 473-505.
. 1981. Cvclomeia undulata (Burges) gen. et comb.nov., a lycopod of the early Triassic strata of New South Wales. Rec. Aust. Mus. 33: 723-734.
Wing, S. L., and J. E. Eckenwalder. 1987. Quantitativearchitectural comparisons of fossil and modern leaves. Amer. J. Bot. 74: 694.
Wolfe, J. A., and G. R. Upchurch. 1987. North American nonmarine climates and vegetation during the Late Cretaceous. Palaeogeogr., Palaeoclimatol.,Palaeoecol. 61: 33-77.

180Worsdell, W. C. 1898. The vascular structure of the
sporophylls of the Cycadaceae. Ann. Bot. 12:203-241.
. 1906. The structure and origin of the Cycadaceae.Ann. Bot. 20: 129-159.
Zalessky, M. D. 1933. Sur les vegetaux fossiles nouveaux du Systeme Anthracolithique du Bassin de Kousnetzk. Bull. Acad. Sci. URSS, sci. math, et natur. 8 : 1213-1258.
Zeiller, R. 1896. Etude sur qUelques plantes fossiles, in particulier Vertebraria et Glossopteris des environs de Johannesburg (Transvaal). Bull. Soc. Geol. du France. 3rd ser. 24: 349-378.
Zimina, V. G. 1967. On Glossopteris and Ganqamopteris in Permian deposits of the Southern Premorje (Far East). Palaeont. Zh. 3: 96-107.
. 1977. The flora in the Early and early LatePermian of southern Primor'ye. Nauka, Moscow.

181
Table 1. Parameters measured from Glossopteris leaves for quantitative analysis.
21. Total vein density/cm2. Midrib thickness (mm)3. Leaf width (mm)
24. Vein density in central part of leaf/cm25. Marginal vein density/cm
26 . Concentration of Y-shaped dichotomies/cm
27. 11 11 Lambda-shaped reticulations/cm8 •• >• H n ii
9 11 11 h— " w «10. " " X - " 11 "1 1 ii ii yj_ n n ii
12. " " Complex " 11 "13. " " Total 11
14. Mesh length (mm)15. Mesh width (mm)
216. Mesh area (mm )17. Angle of mesh with midrib

182Table 2. Species of Indian Glossopteris used in
quantitative analysis. Data from Chandra andSurange (1979).
Identification Species and author Counts Text Figs. Number (from
Chandra and Surange 1979)
1 . 3.2 G. angusta Pant & Gupta 14 1 2 b32 . 3.4 G. pandurata Pant & Gupta 5 46 T3. 3.5 G. giridihensis Pant & Gupta 6 45 C4. 3.6 G. karharbariensis Chandra
& Surange 30 49 N5. 3.7 G. longicaulis Feistmantel 8 2 1 al6 . 3.8 G. zeilleri Pant & Gupta 6 45 K7. 3.9 G. communis Feistmantel 1 1 45 E8 . 3.10 G. decipiens Feistmantel 6 46 Q9. 3.11 G. nimishea Chandra & Surange 18 47 K
1 0 . 3.12 G. stenoneura Feistmantel 13 44 I1 1 . 3.13 G. churiensis A. K. Srivastava 29 54 H
•CMiH 3.14 G. syaldiensis Chandra
& Surange 2 0 51 B13. 3.16 G. saksenae Chandra
& Surange 14 54 M14. 3.17 G. intermittens Feistmantel 15 44 L15. 3.18 G. nakkarea Chandra & Surange 8 48 N16. 3.19 G. barakarensis Kulkarni 25 48 E17. 3.21 G. tortuosa Zeiller 25 49 E18. 3.22 G. lanceolatus Pant
& K. B. Singh 17 52 F

183Table 2, <continued.
19. 3.24 G. maculata Pant& R. S. Singh 17 54 B
2 0 . 3.25 G. euryneura Maheshwari 15 50 D2 1 . 3.26 G. mohudaensis Chandra
& Surange 2 0 51 G2 2 . 3.27 G. rhabdotaenioides
Pant & K. B. Singh 13 53 H23. 3.28 G. srivastavae Surange
& Maheshwari 16 50 A24. 3.29 G. divergens Feistmantel 3 51 A25. 3.30 G. sahnii P. N. Srivastava 1 0 52 I
•VOCM 3.31 G. nautiyalii Pant
& K. B. Singh 14 47 0
27. 3.32 G. gondwanensis Pant & Gupta 15 54 G
28. 3.33 G. formosa Feistmantel 2 28 C<29. 3.34 G. conspicua Feistmantel 7 50 H30. 3.35 G. retifera Feistmantel 17 50 B31. 3.36 G. intermedia Feistmantel 8 50 K32. 3.37 G. subtilis Pant & Gupta 16 17 d33. 3.38 G. oldhamii Pant
& R. S. Singh 1 0 52 G34. 3.40 G. rewaensis Chandra & Surange 25 50 C35. 3.41 G. orbicularis Feistmantel 18 49 C36. 3.42 G. feistmanteli Rigby 15 48 B37. 3.43 G. vulgaris Pant & Gupta 8 46 0

184Table 2, continued.
38. 3.44 G. bosei Chandra & Suranqe 3 1 2 g39. 3.45 G. searsolensis Pant
& R.S. Singh 1 0 50 M40. 3.46 G. spatulata Pant
& K. B. Singh 9 45 A41. 3.47 G. emarginata Maheshwari
& Prakash 1 1 18 e]42. 3.48 G. retusa Maheshwari 1 1 48 H43. 3.49 G. pantii Chandra & Surange 29 48 G44. 3.50 G. arberii P. N. Srivastava 18 45 G45. 3.51 G. tenuifolia Pant & Gupta 2 44 M46. 3.52 G. angustifolia Brongniart 23 46 X47. 3.54 G. tenuinervis Pant & Gupta 26 47 L
00 • 3.56 G. raniganjensis Chandra & Surange 13 44 H
49. 3.57 G. leptoneura Bunbury 30 46 H50. 3.58 G. indica Schimper 9 45 J51. 3.59 G. musaefolia Bunbury 5 51 J52. 3.60 G. stricta Bunbury 25 51 J53. 3.61 G. senii Shyam C. Srivastava 16 49 Q54. 3.62 G. gopadensis Banerji,
Maheshwari & Bose 2 0 49 P55. 3.63 G. brongniartii Pant & Gupta 1 0 2 1 g56. 3.65 G. radiata Pant & K. B. Singh 8 49 K57. 3.66 G. browniana Brongniart 25 47 D58. 3.67 G. varia Pant & Gupta 3 25 h

185Table 3. Summary of major cuticular features reported
for glossopterid leaves.' : ' 1
.1'1
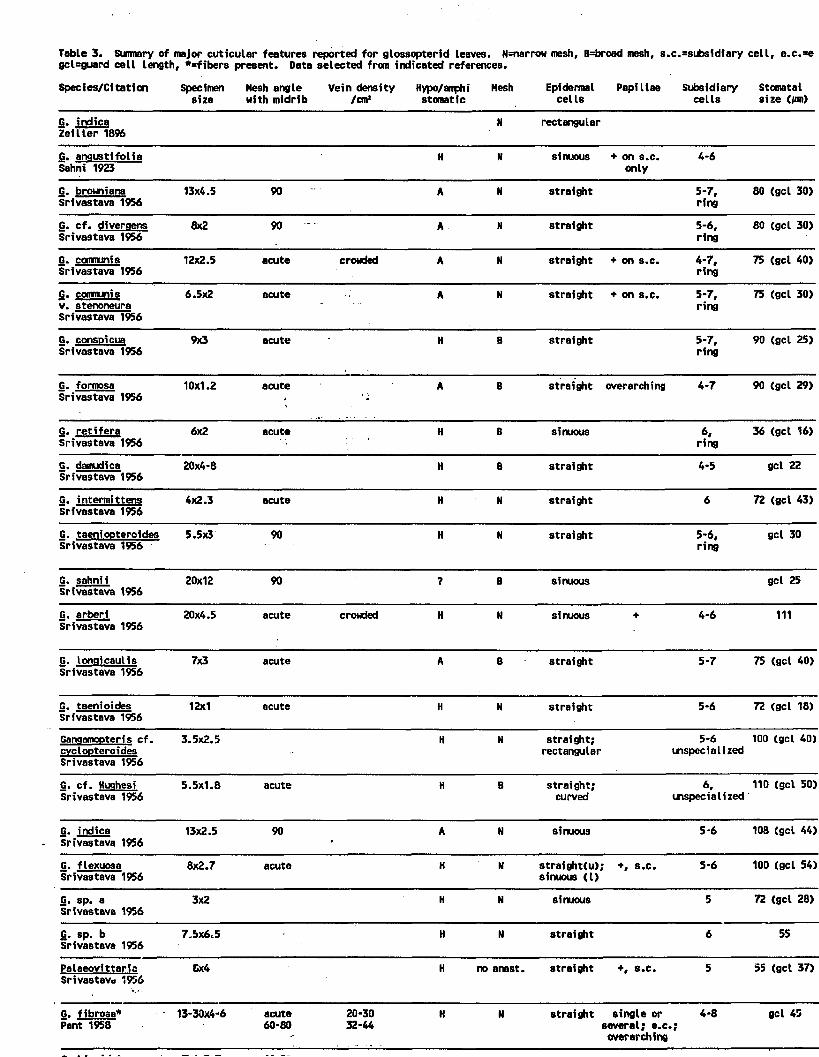
Table 3. Suimary of major cutlcular features reported for glossopterfd leaves. N=narrow mesh, B=broad mesh, 8.c.=sii>sidiary cell, e.c.=^gcl=guard cell length, *=fibers present. Data selected from indicated references.
Species/Citation Specimen Mesh angle Vein density Hypo/amphi Mesh Epidermal Papillae Subsidiary Stomatal size with midrib /cm1 stomatic cells cells size (pm)
G. indica Zeiller 1896
N rectangular
G. anaustifolia Sahni 1923
H N sinuous + on s.c. 4-6 only
G. brouniana Srivastava 1956
13x4.5 90 A N straight 5-7,ring
80 (gel 30)
G. cf. diveraens Srivastava 1956
8x2 90 A N straight 5-6,ring
80 (gel 30)
G. communis Srivastava 1956
12x2.5 acute crowded A N straight + on s.c. 4-7,ring
75 (gel 40)
G. communis v. stenoneura Srivastava 1956
6.5x2 acute A N straight ♦ on s.c. 5-7,ring
75 (gel 30)
G. consoicua Srivastava 1956
9x3 acute H B straight 5-7,ring
90 (gel 25)
G. formosa Srivastava 1956
10x1.2 acute'•
A B straight overarching 4-7 90 (gel 29)
G. retifera Srivastava 1956
6x2 acute H B sinuous 6,ring
36 (gel 16)
G. damudica Srivastava 1956
20x4-8 H B straight 4-5 gel 22
G. intermittens Srivastava 1956
4x2.3 acute H N straight 6 72 (gel 43)
G. taeniooteroides Srivastava 1956
5.5x3 90 H N straight 5-6,ring
gel 30
G. sahnii Srivastava 1956
20x12 90 ? B sinuous gel 25
G. arberi Srivastava 1956
20x4.5 acute crowded H N sinuous + 4-6 111
G. lonnicaulis Srivastava 1956
7x3 acute A B straight 5-7 75 (gel 40)
G. taenioides Srivastava 1956
12x1 acute H N straight 5-6 72 (gel 18)
Ganaamooteris cf. cvclooteroides Srivastava 1956
3.5x2.5 H N straight; 5-6 rectangular unspecialized
100 (gel 40)
G. cf. Hughesi Srivastava 1956
5.5x1.8 acute H B straight; 6, curved mspecialized
110 (gel 50)
G. indica Srivastava 1956
13x2.5 90•
A N sinuous 5-6 108 (gel 44)
G. flexuosa Srivastava 1956
8x2.7 acute H N straight(u); +, s.c. 5-6 sinuous (I)
100 (gel 54)
G. sp. aSrivastava 1956
3x2 H N sinuous 5 72 (gel 28)
G. sp. bSrivastava 1956
7.5x6.5 H N straight 6 55
Palaeovittaria Srivastava *956
8x4 H no anast. straight +, s.c. 5 55 (gel 37)
G. fibrosa* Pant 1958
13-30x4-6 acute60-80
20-3032-44
H N straight single or 4-8 several; e.c.; overarching
gel 45

turrmary of major cuticular features reported for glossopterid leaves. N=narrow mesh, B=broad mesh, s.c.=subsidiary cell, e.c.=epidermal cell,* U length, *«fibers present. Data selected from indicated references.
:ation Specimen Mesh angle Vein density Hypo/amphi Mesh Epidermal Papillae Subsidiary Stomatal Stomata Isize with midrib /cm1 stomatic cells cells size (/on) arrangement
N rectangular
olia H N sinuous + on s.c. only
4-6 i rregular
a1956
13x4.5 90 A N straight 5-7,ring
80 (gel 30) irregularcontiguous
raens1956
8x2 90 A N straight 5-6.ring
80 (gel 30) irregularcontiguous
195612x2.5 acute crowded A N straight + on s.c. 4-7,
ring75 (gel 40) irregular
ira1956
6.5x2 acute A N straight + on s.c. 5-7.ring
75 (gel 30) i rregular
a1956
9x3 acute H B straight 5-7.ring
90 (gel 25) longitudinal orientation in rows; sunken
195610x1.2 acute A B straight overarching 4-7 90 (gel 29) irregular
orientation in rows; contiguous
19566x2 acute H B sinuous 6.
ring36 (gel 16) irregular
195620x4-8 H B straight 4-5 gel 22
tens1956
4x2.3 acute * H N straight 6 72 (gel 43) irregular;contiguous
eroides1956
5.5x3 90 H N straight 5-6.ring
gel 30 irregular orientation in rows
195620x12 90 7 B sinuous gel 25 sparse;
irregular
195620x4.5 acute crowded H N sinuous + 4-6 111 irregular;
contiguous
lis1956
7x3 acute A B straight 5-7 75 (gel 40) irregular orientation in rows; contiguous
es1956
12x1 acute H N straight 5-6 72 (gel 18) scattered
is cf.des1956
3.5x2.5 H N straight;rectangular
5-6unspecialized
100 (gel 40) crowded;irregular
esi1956
5.5x1.8 acute H B straight;curved
6,unspecialized
110 (gel 50) oblique orientation in rows
195613x2.5 90
■A N sinuous 5-6 108 (gel 44) irregular;
contiguous
19568x2.7 acute H N straight(u);
sinuous (I)+, s.c. 5-6 100 (gel 54) irregular
19563x2 H N sinuous 5 72 (gel 28)
19567.5x6.5 H N straight 6 55
ds1956Gx4 H no anast. straight +, s.c. 5 55 (gel 37) crowded;
i rregular; contiguous
13-30x4-6 acute60-80
20-3032-44
H N straight single or several; e.c, overarching
4-8■ *
gel 45 125/mm*; sunken
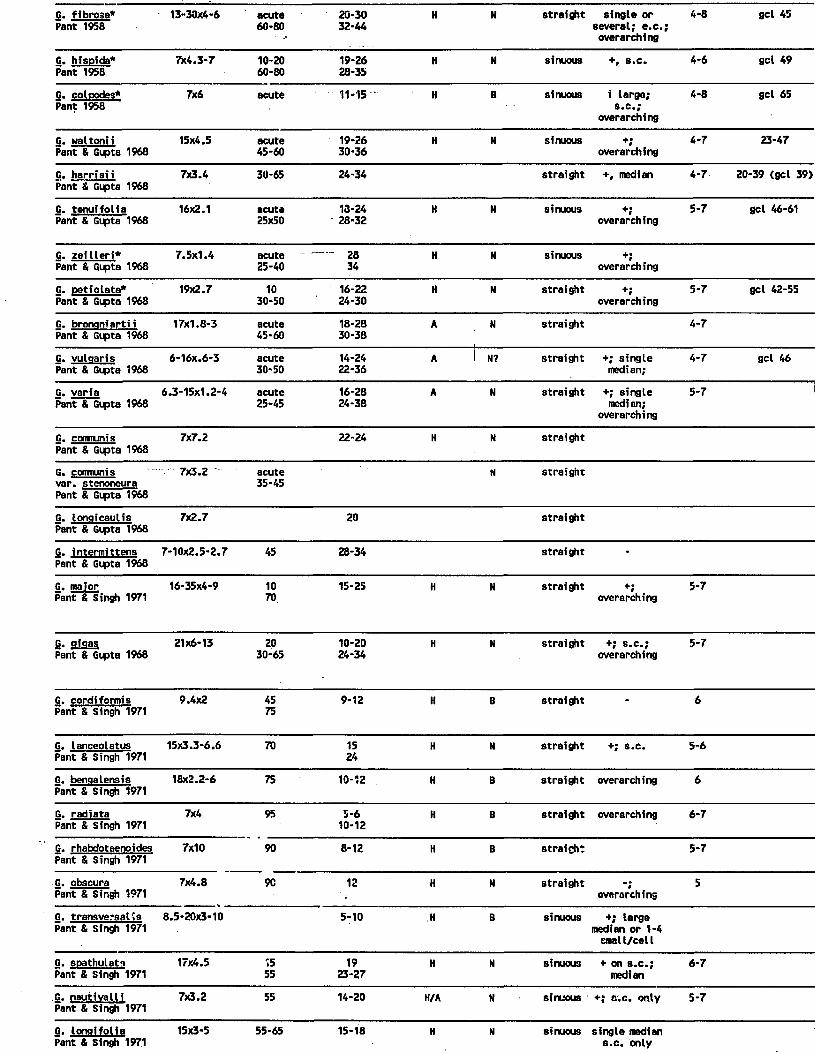
G. fibrosa* Pant 1958
13-30x4-6 acute60-80
20-3032-44
H N straight single or several; e.c.; overarching
4-8 gel 45
G. hism'da* Pant 1958
7x4.3-7 10-2060-80
19-2628-35
H N sinuous +, s.c. 4-6 gel 49
G. colMdes* Pant 1958
7x6 acute 11-15 H B sinuous i large; s.c.;
overarching
4-8 gel 65
G. waltonii Pant & Gupta 1968
15x4.5 acute45-60
19-2630-36
H N sinuous +;overarching
4-7 23-47
G. harrisii Pant & Gupta 1968
7x3.4 30-65 24-34 straight +, median 4-7 20-39 (gel 39)
G. tenuifolia Pant & Gupta 1968
16x2.1 acute25x50
13-2428-32
H N sinuous +;overarching
5-7 gel 46-61
G. zeilleri*Pant & Gupta 1968
7.5x1.4 acute25-40
----- 2834
H N sinuous +;overarching
G. Detiolata*Pant & Gupta 1968
19x2.7 1030-50
16-2224-30
H N straight +;overarching
5-7 gel 42-55
G. bronnniartii Pant & Gupta 1968
17x1.8-3 acute45-60
18-2830-38
A i N straight 4-7
G. vulqaris Pant & Gupta 1968
6-16x.6-3 acute30-50
14-2422-36
A 1 N? straight +; single median;
4-7 gel 46
G. variaPant & Gupta 1968
6.3-15x1.2-4 acute25-45
16-2824-38
A N straight +; single median;
overarching
5-7
G. communis Pant & Gupta 1968
7x7.2 22-24 H N straight
G. communis var. stenoneura Pant & Gupta 1968
7x3.2 acute35-45
N straight
G. lonqicautis Pant & Gupta 1968
7x2.7 20 straight
G. inter-mittens Pant & Gupta 1968
7-10x2.5-2.7 45 28-34 straight -
G. majorPant & Singh 1971
16-35x4-9 1070.
15-25 H N straight +;overarching
5-7
G. gigasPant & Gupta 1968
21x6-13 2030-65
10-2024-34
H N straight +; s.c.; overarching
5-7
G. cordiformis Pant & Singh 1971
9.4x2 4575
9-12 H B straight “ 6
G. tanceolatus Pant & Singh 1971
15x3.3-6.6 70 1524
H N straight +; s.c. 5-6
G. bengalensis Pant & Singh 1971
18x2.2-6 75 10-12 H B straight overarching 6
G. radiata Pant & Singh 1971
7x4 95 5-610-12
H B straight overarching 6-7
G. rhabdotaenoides Pant & Singh 1971
7x10 90 8-12 H B straight 5-7
G. obscura Pant & Singh 1971
7x4.8 9C 12 H N straightoverarching
5
G. transversal is Pant & Singh 1971
8.5-20x3-10 5-10 H B sinuous +; large median or 1-4 email/cell
G. soathuleta Pant & Singh 1971
17x4.5 1555
1923-27
H N sinuous ♦ on s.c.; median
6-7
G. nautivalli Pant & Singh 1971
7x3.2 55 14-20 H/A N sinuous '+; s.c. only 5-7
G. longifolia Pant & Singh 197.1
15x3-5 55-65 15-18 H N sinuous single median s.c. only

ouaiyui. O
6x4 H no anast. straight +, s.c. 5 55 (gel 37) crowded;irregular;contiguous
13-30x4-6 acute60-80
20-3032-44
H N straight single or several; e.c.; overarching -
4-8 gel 45 125/mm*;sunken
7x4.3-7 10-2060-80
19-2628-35
H N sinuous S*Ca 4-6 gel 49 70/mm*;irregular
7x6 acute 11-15 H B sinuous i large; s.c.;
overarching
4-8 gel 65 66/mm*; sunken
15x4.5 acute45-60
19-2630-36
H N sinuous «■;overarching
4-7 23-47
7x3.4 30-65 24-34 straight +, median 4-7 20-39 (gel 39) scattered;irregular
16x2.1 acute25x50
13-2428-32
H N sinuous +;overarching
5-7 gel 46-61 138/nrn’
7.5x1.4 acute25-40
.. - 2834
H N sinuous +;overarching
19x2.7 1030-50
16-2224-30
H N straight +;overarching
5-7 gel 42-55 111/mm*; i rregular
17x1.8-3 acute45-60
18-2830-38
A i “ straight 4-7 121/mn*;scattered
6-16X.6-3 acute30-50
14-2422-36
A 1 N? straight +; single median;
4-7 gel 46 76/mm1; irregular
6.3-15x1.2-4 acute25-45
16-2824-38
A N strai ght +; single median;
overarching5-7 71/mm*;
irregular
7x7.2 22-24 H Ni
straight
7x3.2 ' acute35-45
N straight
7x2.7 20 straight
7-10x2.5-2.7 45 28-34 straight -
16-35x4-9 1070
15-25 H N straight *;overarching
5-7 178/mm*;irregular;contiguous;
sunken
21x6-13 2030-65
10-2024-34
H N straight +; s.c.; overarching
5-7 200/mm*;irregular;contiguous;
sunken
9.4x2 4575
9-12 H B straight • 6 133/mm*;irregular;sunken
15x3.3-6.6 70 1524
H N straight +; s.c. 5-6 102/nsn*
18x2.2-6 75 10-12 H B straight overarching 6 160/mm*;irregular
7x4 95 5-610-12
H B strai ̂it overarching 6-7 i rregular
7x10 90 8-12 H B straight 5-7
7x4.8 9C 12 H N straight Ҥoverarching
5 192/mm*; i rregular
8.5-20x3-10 5-10 H B sinuous ♦; large median or 1-4 cmall/cell
80-120/rnn*; irregular
17x4.5 1555
1923-27
N N sinuous + on s.c.; median
6-7 167/mm*; irregular

G. opscura fx.it.a w -ml h n siraignt -; 9Pant & Singh 1971 overarchingG. transversal is Pant & Singh 1971
8.5-20x3-10 5-10 H B sinuous +; large median or 1-4 cmetl/cetl
G. soathulatn Pant & Singh 1971
17x4.5 1555
1923-27
H N sinuous + on s.c.; median
6-7
G. nautivalli Pant & Singh 1971
7x3.2 55 14-20 H/A N sinuous +; s.c. only 5-7
G. Lonaifolia Pant & Singh 1971
15x3-5 55-65 15-18 H N sinuous single median s.c. only
G. sinqularis Pant & Singh 1971
7x4.8 51 * H B straight +; even; overarching
5-6 129
G. damudica Pant & Singh 1971
6545-60
30 straight
G. aonduanensis Pant & Gupta 1971
7.5-14x0.4-1.2 1010-30
10-16 H B straight-sinuous(mostlystraight)
+; s.c. only;
5-6 23-41
G. subtilis*Pant & Gupta 1971
12x4 10-2050-60
10-1612-20
H B straight-sinuous(mostlystraight)
29-44
G. tenuinervis Pant & Gupta 1971
7x2 45-6075-85
28-34 (mid-If) 36-40 (margin)
H N straight-curved
on s.c. 5-7 gel 17-:
G. ahusikensis Pant & Gupta 1971
10x2.1 35-5050-70
16-2226-32
H N straight gel 45
G. contracts Pant & Gupta 1971
7.2x2.3 10-1545-50
16-2322-32
H N straight-sinuous
+; single median;
overarching
gel 35
G. angusta Pant & Gupta 1971
10x2.6 20-45 16-2222-28
H N sinuous +; numerous small
24-46
G. airidihensis Pant & Gupta 1971
7x3.3 20-4050-65
20-2226-30
H N straight +; single median;
overarching
2-7 32-40
G. Dandurata* Pant & Gupta 1971
3.8x1.5 30 45-50 straight
G. sp. APant & Gupta 1971
9.6x1.7 +
G. searsolensis Pant & Singh 1974
7x3.5 acute25-42
12-2016-24
H N straight overarching unspecialized 39
G. maculata Pant & Singh 1974
9.8x3.8 12-40 5-1313-20
H B curved-sinuous
wspecialized 10-34
G. sastrii Pant & Singh 1974
14x4 17-5061-94
.4-910-26
H B sinuous overarching 6-7/ 16-31.!
G. oldhami Pant & Singh 1974
15.4x6.8 30-7470-88
8-1415-36
H N straight gel 42-<
Belemnooteris saqittifolia Pant & Choudhury 1977
5-16x0.4-7.5 20-80 6-1310-18
H B sinuous overarching 5-6 35-41.!
G. oellucida Pant & Choudhury 19 77
16.5x7.5 48-709-12
8-9 H Ni
sinuous overarching 5-6; ring 31.5
G. nitssonioides Pant & Pant 1987
11.5x3.5 40-5070-80
20-3030-40
U N straight + on s.c. 6-8;unspecialized
G. sidhiensis Pant & Pant 1987
8x4.5 30-4570-80
15-178-15
U N straight + on s.c. 7;unspecialized
G. schoofii Pigg 1988
7x1.8-3.1 22 2221
H N straight +; median 5-6 s.c. and unspecialized s.c.
G. skaarensis Pigg 1988
7x2.7 57 14 H B sinuous +; several small/cell; overarching
5-6; ring

7x4.8 9C 12 H N straight "foverarching
5 192/mm*;irregular
8.5-20x3-10 5-10 H B sinuous +; large median or 1-4 cmall/celt
80-120/nrn*;irregular
17x4.5 1555
1923-27
H N sinuous + on s.c.; median
6-7 167/rnn*; irregular
7x3.2 55 14-20 H/A N sinuous +; s.c. only 5-7 89/mm*
15x3-5 55-65 15-18 H N sinuous single median s.c. only
133/mm*;irregular
7x4.8 51 • N B straight +; even; overarching
5-6 129
6545-60
30 straight i rregular
7.5-14x0.4-1.2 1010-30
10-16 H B straight-sinuous(mostlystraight)
■*•; s.c. only;
5-6 23-41 152/mm*;irregular
12x4 10-2050-60
10-1612-20
H B straight-sinuous(mostly
straight)
29-44 57/itm*;irregular
7x2 45-6075-85
28-34 (mid-If) 36-40 (margin)
H N straight-curved
on s.c. 5-7 gel 17-35 133/mm*;irregular
10x2.1 35-5050-70
16-2226-32
H N straight gel 49 118/rnn*
7.2x2.3 10-1545-50
16-2322-32
N N straight-sinuous
+; single median;
overarching
gel 39 171/irm*
10x2.6 20-45 16-2222-28
H N sinuous +; numerous small
24-46 95/mm*; i rregular
7x3.3 20-4050-65
20-2226-30
N N straight +; single median;
overarching
2-7 32-40 114/mn’
3.8x1.5 30 45-50 straight
9.6x1.7 +
7x3.5 acute25-42
12-2016-24
H N straight overarching wspecialized 39 157/mm*;irregular
9.8x3.8 12-40 5-1313-20
H B curved-sinuous
unspecialized 10-34 39/mm*;irregular
14x4 17-5061-94
.4-910-26
H B sinuous overarching 6-7/ 16-31.5
15.4x6.8 30-7470-88
8-1415-36
H N straight gel 42-63
5-16x0.4-7.5 20-80 6-1310-18
H B sinuous overarching 5-6 35-41.5 9/nin*;irregular;sunken
16.5x7.5 48-709-12
8-9 H H
!
sinuous overarching 5-6; ring 31.5 24/rrm1;irregular
11.5x3.5 40-5070-80
20-3030-40
U N straight + on s.c. 6-8;unspecialized
8x4.5 30-4570-80
15-178-15
U N straight + on s.c. 7;unspecialized
7x1.8-3.1 22 2221
H N straight +; median 5-6 s.c. and wspecialized s.c.
longitudinal orientation in rows
7x2.7 57 14 H B sinuous +; several small/cell; overarching
5-6; ring irregular;contiguous;
sunken

186
Table 4 Occurrence of features in association with glossopterid cuticular studies (Table 3)1. Mesh shape [narrow (N) vs. broad (B)].
?N N B no reticu- TOTALlations
Number: 1Percentage 1.5%
44 2067% 30%
11.5%
67100%

187
Table 5. Occurrence of features in association with glossopterid cuticular studies (Table 3).2. Hypostomatic (H) vs. amphistomatic (A) vs. unistomatic (U).
Number:Percentage:
H U H/A
5079%
10 216% 3%
12%
TOTAL
63100%

188
Table 6. Occurrence of features in association with glosssopterid cuticular studies (Table 3).3. Epidermal cell wall shape (St= straight anticlinal margins; Si= sinuous, Rt= rectangular; Cv= curved; u=upper epidermis; l=lower epidermis).
St Si Rt Rt St St St (u) TOTAL/St /Cv /Si /Si(l)
Number41 18 1 1 2 5 1 69
Percentage:60% 27% 1% 1% 3% 7% 1% 100%

189
Table 7. Occurrence of features in association with glossopterid cuticular studies (Table 3).4. Relationship of mesh shape [narrow (N) vs. broad (B)] to stomatal position [hypo- (H) vs. amphi- (A) vs. unistomatic (U).]
St/N Si/N St/B Si/B Other Total
Number: 26 12 9 6 11 64
Percentage: 40% 19% 14% 10% 17% 100%

1 9 0
Table 8. Occurrence of features in association with glossopterid cuticular studies (Table 3).5. Relationship between stomatal position [hypo- (H) vs. amphi- (A) vs. unistomatic (U)] and mesh shape [narrow (N) vs. broad (B)].
H&N H&B A&N A&B U&N H/A&B
Number:32 17 7 2 2 1
Percentage:59% 31% 13% 4% 4% 2%

191
Table 9. Results of multiple discriminate analysis ofAntarctic leaves, based on continuous variables (i.e., mesh length, width, angle of lateral veins with midrib, and mesh area)
Observations classified into taxon:G. schopfii
From G. schopfii:Number ofobservations: 208Percentage: 84.55%
From G. skaarensis:Number ofobservations: 26Percentage: 14.44%
TOTAL: 234PERCENT: 54.93%
G. skaarensis TOTAL
38 24615.45% 100%
154 18085.56% 100%
192 42645.07 100%

192
Table 10. Mean values + standard error for continuous variables for G. schopfii and G. skaarensis.
Taxon:
Mesh length (mm)
Mesh width (mm)
2Mesh area (mm )
Angle of lateral veins with midrib
G. schopfii
3.8 + 0.1
0.49 + 0.15
1.48 + 0.6
53.3° + 1.85
G. skaarensis
2.86 + 0.77
0.6 + 0.16
1.44 + 0.613
57.82° + 1.48

193
Glossopteris schopfii. Fig. 1. Composite of line diagrams to illustrate morphological variability. These specimens represent part of the data base for the quantitative* analysis (see text for details). a=Gl.l, 452 Surface, (Fig. 2); b= G1.2, 452 G^ Surface, (Fig. 3); c= G1.3, 452 Surface? d= G1.8. 452 Fragment; e= G1.12, 452 F Base; f= G1.15, 452 F Base; g= Gl.ll, 452 Gj Surface; h= G1.10, 452 Surface; i= G1.6, 452 Surface; j= G1.4, 452 Surface. Scale bar = 1 cm.

194

195
Glossopteris schopfii. External morphological features. Pig. 2-5.
Fig. 2. Specimen showing general morphological features. Note vascular strands in midrib (arrow). Gl.l, 452 Surface (=Fig. la) x 2.5.
Fig. 3. Most extensive specimen, showing detail ofvenation. Note 4-5 veins of midrib in region where leaf has split paradermally (arrow). G1.2, 452, G^ Surface (=Fig. lb) x 2.2.
Fig. 4. Specimen showing detail of retuse apex. G1.22, Ant 70-1-43 x 3.2.
Fig. 5. Specimen of small (scale?) leaf showing detail of anastomosing venation near emarginate apex. G1.5, 452 Surface x 4.1.

1 9 6

1 9 7
Glossopteris schopfii. Anatomical features. Fig. 6-10. (f=fiber, h=hypodermis, ph=phloem, px=protoxylem, vb=vascular bundle).
Fig. 6. Paradermal section to show narrow elongate meshesand various types of reticulations. 435 no. 2 a x 8.
Fig. 7. Transverse section through several leaves. Noteprominent vascular bundles (vb), hypodermis (h). 533 A Bot no. 8 a x 32.
Fig. 8. Transverse section through leaf showing several vascular bundles. Mesophyll is greatly distorted.483 C Top l a x 70.
Fig. 9. Paradermal section to show venation in W-shapedreticulation. Note tracheids that crosses diagonally through reticulation (arrow). 435 no. 4 alpha x 112.
Fig. 10. Transverse section through three vascularbundles. Note metaxylem, mesarch-exarch position of protoxylem strands (px), radially aligned tracheids, prominent fibers of bundle sheath (f), and large lacuna representing position of phloem (ph). In bundle on right, xylem has separated. 483 C Top a no.1 X 6 0 .

198
x. A

1 9 9
Glossopteris schopfii. Anatomical features. Fig. 11-14. (cuticle, m=mesophyll).
Fig. 11. Paradermal preparation of leaf with lower cuticle preserved (c). Leaf is sectioned through mesophyll at top (m). Positions of stomata in cuticle illustrated in Fig. 21. 435 no. 4 alpha x 50.
Fig. 12. Paradermal section through mesophyll (m), showing an H-shaped vein reticulation. Cells of the mesophyll are poorly preserved. 435 no. 8 alpha x 70. *
Fig. 13. Paradermal section through mesophyll. 435 no. 8 alpha x 66.
Fig. 14. Paradermal section through hypodermis showing small cuboidal cells with dark contents. 435 no. 2 alpha x 112.

2 0 0

2 0 1
Glossopteris schopfii. Anatomical features. Fig. 15-20. (p=papilla, sc=subsidiary cell).
Fig. 15. Paradermal section of leaf showing detail of bundle sheath fibers. Dark spots represent simple pits of bundle sheath in face view (arrow). 435 no. 9alpha, x 106.
Fig. 16. Tracheids with reticulate/pitted wall thickenings. 435 no. 30 alpha, x 325.
Fig. 17. Y-shaped dichotomy. Note bifurcating tracheid (arrow). 435 no. 22 beta x 275.
Fig. 18. Paradermal section of lower cuticle showing stoma (arrow), straight-margined epidermal cells, and some dark, irregular thickenings on cells. 435 no. 9 alpha x 232.
Fig. 19. Cuticle of lower leaf surface, showing stoma and straight epidermal cell walls. 435 no. 5 x 400.
Fig. 20. Stoma surrounded by five subsidiary cells (see Fig. 23). Note centrally positioned papillae on subsidiary cells (p), 435 no. 4 alpha x 400.

2 0 2
9

203
Glossopteris schopfii. Fig. 21. Distribution andorientation of stomata. Specimen is illustrated in Fig. 6, 11. Dotted lines represent areas for which reliable samples could be counted. Vectors represent lingitudinal axis of guard cells, which tend toward being aligned in rows.

*« ♦

205
Glossopteris schopfii. Stomata. Fig. 22. Reconstruction of stoma showing simple organization of stomatal apparatus.
Fig. 23. Reconstruction of stoma showing single median papilla on subsidiary cells of this species.

206

207
Glossopteris schopfii. Morphology of stems. Fig. 24-27. (1= small leaves, vb=vascular bundle).
Fig. 24. Transverse section through bud ofsmall twig surrounded by G. schopfii leaves. Bud has been sectioned distal to level of stem. Note innermost leaves with thickened midrib and shorter lateral laminae (1), and prominent vascular bundle area on outermost leaves (vb). 484 B Bot no. 4 beta x 32.
Fig. 25. Transverse section through small stalk (arrow), 533 A Bot a no. 4 x 27.
Fig. 26. Transverse section through small bud cut distal to stem level. Note thickened midrib and short lateral laminae of leaves, 484 B Bot no. 1 c x 42.
Fig. 27. Oblique-transverse section through the base of ovule showing double vascular strand (vb) with anatomical features of G. schopfii leaves. 533 B Top no. 16 x 22.

208

209
Glossopteris skaarensis. Fig. 28. Composite of line diagrams to illustrate morphological variability. These specimens represent part of the data base for the quantatitive analysis (see text for details). a= G2.1, 463 E Bot Fragment (=Fig. 29); b= G2.2, 452 Surface, (=Fig. 31); c=G2.4, 452 Fragment, (Fig. 32); d=G2.8, 452 Fragment; e=G2.7, 452 Fragment, (=Fig. 30); f=G2.9, 452 Fragment; g=G2.10, 452 Fragment. Scale bar = 1 cm.
i

2 1 0

2 1 1
Glossopteris skaarensis. External morphological features. Fig. 29-32.
Fig. 29. Specimen showing general morphological features. Note polygonal shaped meshes, and thickened area of midrib. G2.1, 463 E Bot Fragment (=Fig. 28a) x 2.
Fig. 30. Specimen showing general features. 0,2.1, 452 Fragment (=Fig. 28f) x 2.6.
Fig. 31. Specimen showing denser venation pattern. G2.2, 452 Fragment (=Fig. 28b) x 2.7.
Fig. 32. Small leaves. Leaf on right shows slightlyemarginate apex. Note venation. G2.4, G2.5, 452 Fragment (=Fig. 28c) x 4.1.

212

213
Glossopteris skaarensis. Anatomical features. Pig. 33-37. (c=cuticle, h=hypodermis, mr=midrib, vb=vascular bundle).
Fig. 33. Paradermal section showing general features of venation. Note midrib (mr), 465 C4 Side no. 5 x 7 .
Fig. 34. Transverse section of leaf showing prominent hypodermis (h), thickened abaxial cuticle (c) and central vascular bundle (vb), 484 D Top no. 7 f x 70.
Fig. 35. Transverse section of leaf. Note inflatedappearance of midrib in comparison to lateral laminae. 484 D Top no. 4 f x 42.
Fig. 3 6. Transverse section through leaf at proximal level. Note prominent double vascular bundles in midrib, keeled appearance of midrib, and short lateral laminae. Compare with petiole base in Fig. 53 (bottom right), 533 B Top. no. 16 x 22.
Fig. 37. Transverse section of two leaves. Note prominent hypodermis and oval vascular bundles (vb), 484 C Bot. no. 1 g x 40.

2 1 4

215
Glossopteris skaarensis. Anatomical features. Fig. 38-41. (e=epidermal cell).
Fig. 38. Paradermal section of mesh to show mesophll.Darkened material represents cuticle. 465 C4 side no. 2 x 33.
Fig. 39. Transverse section through leaf to show thickened cuticle on upper and lower epidermis, and tabular cells of the epidermis (e). 484 C Bot. no. 1 g X 176.
Fig. 40. Paradermal section through leaf to show tracheid with helical wall thickening (arrow). 465 C4 side 1no. 5 x 216.
Fig. 41. Paradermal section through leaf. Note vascular bundle sheath of elongate cells, lacking fibers. 465 C4 SI no. 5 x 216.

2 1 6

217
Glossopteris skaarensis. Anatomical features. Fig. 42-46. (sc=subsidiary cell).
Fig. 42. Lower cuticle two guard cells (arrows). Notethickened guard cells. See Fig. 48. 465 C4 side no.13 x 363.
Fig. 43. Cuticle showing sinuous anticlinal wall pattern of cells. 465 C4 side alpha no. 12 x 176.
Fig. 44. Stomatal apparatus. Sunken stoma is surrounded by ring of six subsidiary cells (sc). See Fig. 50.465 C4 side 1 no. 5 x 336.
Fig. 45. Two sunken stomata (arrows) surrounded by rings of 5 -6 subsidiary cells. Note prominent beaklike papillae of subsidiary cells. See Fig. 49. 465 C4 side 1 no. 5 x 336.
Fig. 46. Cuticle showing sinuous anticlinal cell walls. Note stomata (s) x 550.

218

2X9
Glossopteris skaarensis. Fig. 47. Distribution andorientation of stomata. Specimen is illustrated in Fig. 33.

220
I

221
Glossopteris skaarensis. Stomata. Fig. 48.Reconstruction of detail of stomata from inside of leaf. See Fig. 42.
Fig. 49. Reconstruction of subsidiary cells with beaklike papillae surrounding two continguous stomata. See Fig. 44.
Fig. 50. Reconstruction of detail of stomatal apparatus with ring of subsidiary cells, see Fig. 45.


223
Glossopteris skaarensis. Stems. Fig. 51-55. (co = cortex, l=small leaves, lb=leaf base, pi=pith, st=stele, vb=vascular bundle, w=wood).
Fig. 51. Transverse section of stem showing septate pith and broad zone of wood. 451 E Top no. 52 alpha x 14.
Fig. 52. Oblique longitudinal section through small budwith attached scale-like leaves. Note stele (st) and leaves (1). 451 D Top no. 6 beta x 31.
Fig. 53. Transverse section of stem with leaf bases. Note broad pith (pi), cylinder of wood (w), parenchymatous cortex (co), and leaf base (lb). 451 D Bot no. 31 taux 24.
Fig. 54. Transverse section through small bud withattached leaves (1), and lobelike leaf bases (lb).See Fig. 56. 451 D Bot. no. 32 theta x 50.
Fig. 55. Oblique section through small bud that does notpass through axis. Note small scale-like leaves withshort lateral laminae (1), and large lacuna (vb) representing position of vascular bundle. 533 B Top no. 5 x 50.

224

225
Glossopteris skaarensis. Fig. 56. Series of transversesections through bud in Fig. 54. Leaves are numbered in order of occurrence from proximal to more distal sections (Section d is most proximal). Phyllotaxy approaches 2/5. Series 451 D top theta. a= no.63, b= no. 55, c=no. 40, d=no. 30 x 50.

226

227
Cluster diagram of Glossopteris leaves, including data from G. schopfii and G. skaarensis, and Indian leaves (Chandra and Surange, 1979), as indicated in Table 2. Fig. 57.

1.15
M1.121.11MO2J2.7
25
228
.1310 20 30 40 50 eo

229
Dicroidium fremouwensis. Composite of line diagrams thatillustrate morphological variability in the taxon.Fig. 58. a= Dl, 10137 A Top Surface (Fig. 59); b=D14, 10235 B2 (Fig. 62); c= D8, 10109 B2 Surface (Fig. 61); d= D21, 10200 C3, (Fig. 60); e= Dl5f 10235 B2 (Fig. 62); f=D2, 10137 A Surface; g= D18, 10112, (Fig. 64); h= D19, 10112.

o , - . P '
HH,____^ It

231
Dicroidiuxn fremouwensis. External morphological features. Fig. 59-64.
Fig. 59. Distal portion of frond showing bipinnatid pinnae (=Fig. 58a). Dl, 10137 A Surface x 2.2.
Fig. 60. Portion of bipinnatifid pinnae showing lobes and venation pattern. (=Fig. 58d), D21, 10200 Surface x 2.6.
Fig. 61. Note bipinnatifid pinnae. Specimen has been recut at arrow (see Fig. 72), =58c), D8, 10109, Surface x 3.
Fig. 62. Bipinnatifid pinna with venation. (=Fig. 58b),D14, 10235 B2 Surface x 2.6.
Fig. 63. Note frond bifurcation, entire basal pinnae, and venation (=Fig. 58e). D15, 10235 B2 Surface x 2.6.
Fig. 64. Note entire basal pinnae (=Fig. 58g). D18, 10112Surface x 2.

232

233
Dicroidium fremouwensis. Fig. 65-69. Anatomical features. (f=fiber-like cells, pa=palisade, ph=phloem, sp=spongy mesophyll, tt=transfusion tracheids).
Fig. 65. Transverse section through laminar pinna to show prominent midvein and palisade (pa) and spongy (sp) mesophyll. 10033 B Bot no. 13 x 22.
t
Fig. 66. Transverse section through laminar pinna at level where lamina is constricted. Note lateral veins (arrow), palisade layer and vascular bundles of midrib. 580 CSR no. 2d x 34.
Fig. 67. Transverse section through laminar pinna showing lateral veins. Note palisade (pa) mesophyll. 580 C Bot. no. la x 110.
Fig. 68. Transverse section of vascular bundle showingradially aligned tracheids and zone of dark, radially aligned cells interpreted as secondary phloem (arrow). 567 B^ Top alpha no. 1 x 256.
Fig. 69. Transverse section of central vascular bundle. Note transfusion tissue (tt), secondary phloem (arrow), and fiber-like cells (f). 567 B Top no. 3e x110.


235
Dicroidium fremouwensis. Anatomical features. Fig. 70-75. (r=radial group of abaxial bundles, re=resinous cells).
Fig. 70. Transverse section through laminar pinna. 580 C Bot no. 1 x 27.
Fig. 71. Oblique transverse section of frond rachis at proximal level showing parenchymatous ground tissue. 602 B Bot no. 1 x 32.
Fig. 72. Transverse section through frond rachis at level indicated in Fig. 61. Vascular bundles are arranged in abaxial ring of five bundles (r) and an adaxial line of three bundles (numbered 1-3). D8, 10109 B2aside b no. 1 alpha x 22.
Fig. 73. Transverse section through either laminar pinna or rachis at distal level, extending into lateral laminae on either side. Note five prominent vascular bundles of midvein, in U-shaped arrangement. 567 B Top no. 3e x 23.
Fig. 74. Oblique section through the two branches of frond rachis distal to bifurcation (arrows). Rachis on right is in transverse section, rachis at left is obliquely cut. On right, note abaxial ring of five bundles, and linear arrangement of adaxial bundles.602 C Top no. 20 x 22.

236
Fig. 75. Transverse/oblique section of frond rachis showing abaxial ring of eight bundles (r) and an adaxial line of four bundles. Note resinous cells (re). 10109 B2aside b no. 1 beta x 22.

237

238
Dicroidium fremouwensis. Anatomical features. Fig. 76-80. (cu=cuboidal cells, f=fiber-like cells, t=tracheid, tt=transfusion tissue)
Fig. 76. Longitudinal section through laminar pinna showing vascular bundle sheath. Note cuboidal cells of bundle sheath (cu, at left), elongate fiber-like cells (f), and transfusion tissue (tt). 10200 C3 Pull alpha no. 8 x 176.
Fig. 77. Longitudinal section through vascular bundle.Note tracheids (t). 10200 C3 Pull alpha no. 8 x 216.
Fig. 78. Longitudinal section through vascular bundle.Note transfusion tissue (tt). 10200 C3 Pull alpha no.5 x 216.
Fig. 79. Paradermal section through palisade layer of lamina. 10200 C3 Pull alpha no. 5 x 70.
Fig. 80. Paradermal section through spongy layer of lamina. 10200 C3 Pull alpha no. 5 x 216.

239

240
Dicroidium fremouwensis. Anatomical features. Fig. 81-82.
Fig. 81. Cuticle of lower epidermis. Note polygonal and elongate epidermal cells (e), stomata. 10200 C3 Pull alpha no. 1 x 214.
Fig. 82. Detail of stoma. 10200 C3 Pull alpha no. 1 x 343.

2 4 1
Related Documents











