MAGNETIC RESONANCE IN MEDICINE 18, 15-27 ( I99 I ) Analysis of 23Na NMR Spectra from Isolated Perfused Hearts SUSAN J. KOHLER, STANTON B. PERRY, LAURA C. STEWART, DAVID E. ATKINSON, KIERAN CLARKE, AND JOANNE s. INGWALL NMR Laboratory for. Physiological Chemistry, Department $Medicine, Brigham & Women's Hospital and Harvard Medical School, Boston, Massachusetts 021 15 Received December 15, 1989; revised April 2, 1990 The 23Na NMR spectra obtained from isolated hearts perfused with buffer contain- ing the paramagnetic shift reagent dysprosium triethylenetetraminehexaacetic acid, Dy( TTHA)3-, are complex and contain a number of overlapping peaks of different in- tensities. Spectra from rat, rabbit, guinea pig, and ferret hearts obtained during periods of control perfusion are similar and undergo similar changes when the hearts are subjected to periods of ischemia and reflow. The contributions from the intracellular, interstitial, vascular, and bath compartments to the multiple peaks in the spectra of rat hearts have been assigned. The significant contributions to these spectra of bulk magnetic susceptibility effects and incomplete mixing have been demonstrated through a series of modeling ex- periments. Since the spectra from hearts of different species are so similar, the peak as- signments made for the rat are applicable to spectra from rabbit, guinea pig, and ferret hearts as well. This work provides a framework for quantitative analysis of the spectral changes which occur during conditions such as ischemia and reflow. o 1991 Academic Press, Inc. INTRODUCTION The paramagnetic cationic shift reagent dysprosium triethylenetetraminehexaacetic acid, Dy( TTHA)3-, has been shown to be an effective tool for 23Na NMR spectroscopy in biological applications. Since the presence of a high negative charge and its relatively large size prevent the complex from crossing the cell membrane, its use enables the distinction between intracellular and extracellular sodium pools in a variety of cells and tissues (1-6). Of particular interest are studies in which DY(TTHA)~- has been used in isolated, perfused hearts (5,6). Spectra from perfused hearts would be expected to have resonances from four distinct physiological compartments, namely the bath, the vasculature, the interstitium, and the intracellular space. However, the spectra from isolated hearts typically have at least seven or eight poorly resolved peaks. With the exception of the relatively unshifted intracellular sodium signal, definitive peak assignments have not previously been made, although the potential importance of bulk magnetic susceptibility effects has been noted (7, 8). The present study was undertaken to identify the contributions from the four physiological compartments to the 23Na spectra from isolated, perfused hearts. Here we present 23Na spectra of hearts from rats, guinea pigs, rabbits, and ferrets. Although in all cases the spectra are complicated, we show that there is a striking similarity in the appearance of the spectra of hearts from the different species during 15 0740-3 194/9 I $3.00 Copyright 0 IVY1 by Academic Press, Inc. All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MAGNETIC RESONANCE IN MEDICINE 18, 15-27 ( I99 I )
Analysis of 23Na NMR Spectra from Isolated Perfused Hearts
SUSAN J. KOHLER, STANTON B. PERRY, LAURA C. STEWART, DAVID E. ATKINSON, KIERAN CLARKE, AND JOANNE s. INGWALL
NMR Laboratory for. Physiological Chemistry, Department $Medicine, Brigham & Women's Hospital and Harvard Medical School, Boston, Massachusetts 021 15
Received December 15, 1989; revised April 2, 1990
The 23Na NMR spectra obtained from isolated hearts perfused with buffer contain- ing the paramagnetic shift reagent dysprosium triethylenetetraminehexaacetic acid, Dy( TTHA)3-, are complex and contain a number of overlapping peaks of different in- tensities. Spectra from rat, rabbit, guinea pig, and ferret hearts obtained during periods of control perfusion are similar and undergo similar changes when the hearts are subjected to periods of ischemia and reflow. The contributions from the intracellular, interstitial, vascular, and bath compartments to the multiple peaks in the spectra of rat hearts have been assigned. The significant contributions to these spectra of bulk magnetic susceptibility effects and incomplete mixing have been demonstrated through a series of modeling ex- periments. Since the spectra from hearts of different species are so similar, the peak as- signments made for the rat are applicable to spectra from rabbit, guinea pig, and ferret hearts as well. This work provides a framework for quantitative analysis of the spectral changes which occur during conditions such as ischemia and reflow. o 1991 Academic
Press, Inc.
INTRODUCTION
The paramagnetic cationic shift reagent dysprosium triethylenetetraminehexaacetic acid, Dy( TTHA)3-, has been shown to be an effective tool for 23Na NMR spectroscopy in biological applications. Since the presence of a high negative charge and its relatively large size prevent the complex from crossing the cell membrane, its use enables the distinction between intracellular and extracellular sodium pools in a variety of cells and tissues (1-6). Of particular interest are studies in which DY(TTHA)~- has been used in isolated, perfused hearts ( 5 , 6 ) . Spectra from perfused hearts would be expected to have resonances from four distinct physiological compartments, namely the bath, the vasculature, the interstitium, and the intracellular space. However, the spectra from isolated hearts typically have at least seven or eight poorly resolved peaks. With the exception of the relatively unshifted intracellular sodium signal, definitive peak assignments have not previously been made, although the potential importance of bulk magnetic susceptibility effects has been noted (7 , 8). The present study was undertaken to identify the contributions from the four physiological compartments to the 23Na spectra from isolated, perfused hearts.
Here we present 23Na spectra of hearts from rats, guinea pigs, rabbits, and ferrets. Although in all cases the spectra are complicated, we show that there is a striking similarity in the appearance of the spectra of hearts from the different species during
15 0740-3 194/9 I $3.00 Copyright 0 I V Y 1 by Academic Press, Inc. All rights of reproduction in any form reserved.

16 KOHLER ET AL.
periods of normal perfusion, no-flow ischemia, and reperfusion. The work presented here takes advantage of this commonality to design modeling experiments to aid in understanding the factors responsible for the spectral complexity. Thus we present a series of experiments with phantoms and rat hearts designed to identify the spectral contributions of each of the physically distinct compartments. Since the samples are heterogeneous with respect to the distribution of the paramagnetic shift reagent, bulk magnetic susceptibility effects may be an important factor influencing the appearance of the spectra (7 , 8). Similarly, heterogeneity in the ratio of shift reagent to sodium within the sample may lead to spectral complexity. Results are presented from ex- periments which evaluate the contributions of bulk magnetic susceptibility and shift reagent and sodium concentration gradients to the observed spectra. A preliminary report of this work has been made ( 9 ) .
METHODS
Heart Preparation
The hearts used in these experiments were from male Sprague-Dawley rats weighing 350-400 g, male Hartley guinea pigs weighing 350-400 g, male and female New Zealand white rabbits weighing 230-270 g, and castrated male ferrets weighing 1500-
Animals were anesthetized intrapentoneally with sodium pentobarbital or, in the case of the ferret, chloroform. The hearts were quickly removed and placed in ice- cold buffer. The aortic root was secured on a cannula attached to the perfusion ap- paratus and the hearts were perfused via the coronary arteries at a constant temperature of 37°C and at a constant pressure. The perfusion pressure was optimized for each species as measured by the product of heart rate and developed pressure, coronary flow, and high-energy phosphate levels. Perfusion pressure was 60 Torr for guinea pigs and 100 Torr for rats, ferrets, and rabbits. Temperature was maintained at 37°C throughout the protocols by blowing warmed air past the sample. The perfusion and bathing solutions were delivered to the sample chamber via water-jacketed lines main- tained at 37°C.
Langendorff heart preparations were made isovolumic by inserting a water-filled latex balloon into the left ventricle. A drain was also inserted in the left ventricle to prevent fluid accumulation. The heart rate and left ventricular developed pressure were monitored continuously using a Stanton P23dB pressure transducer and Hewlett Packard 8805D pressure amplifier and 7754B recording system.
2000 g.
Perfusion
The perfusion apparatus was fitted with two or three separate reservoirs so that the perfusing medium could be switched rapidly from one buffered solution to another. A second pump maintained an additional reservoir containing the mannitol bathing solution at 37°C; the flow rate of this mannitol bath was one and one-half to two times coronary flow. Each perfusion reservoir was bubbled with O2 :C02 (95:5), main- taining a pH of 7.4. The mixture of mannitol bathing solution and perfusate was aspirated from the sample chamber above the heart. The total volume displaced by

23Na SPECTRA OF HEARTS 17
the buffer and heart was 5-10 ml, depending on the protocol. Monitoring cardiac performance confirmed that all hearts used for ischemia studies exhibited the function typical for Langendorff perfusion.
Solutions
Perfusion buflers. The normal perfusion medium contained 118 m M NaC1, 4.7 mMKC1,1.75 mMCaC12, 1.2 mMMgS04, 0.5 mMNa2H2EDTA, 25 mMNaH2C03, and 11 m M glucose at pH 7.4. For 23Na NMR studies, the perfusion medium was made 10 mMin Na3Dy(TTHA) 6 3NaCI ( 5 , IO), and the NaCl concentration adjusted so that the total sodium concentration remained at 145 mM. To maintain a free [Ca2'] of 1.1 mM, calcium concentrations were increased to 3 m M to compensate for the buffering effects of the shift reagent.
Isotonic low-sodium perfusion bufers. The normal shift reagent perfusion buffer was modified by the replacement of 1 18 m M NaCl with 1 18 m M choline chloride or 118 m M lithium chloride. The Tris form of the shift reagent, Tris3Dy- (TTHA) - 3TrisHC1, was used in place of the Na3Dy( TTHA) - 3NaCI.
Mannitol solution. A sodium-free isotonic mannitol solution containing 10 m M Tris3Dy( TTHA) * 3TrisHC1, 0.25 M mannitol, and 4 m M Tris at pH 7.4 was used either to bathe the hearts or as a perfusion buffer during washout experiments.
NMR Spectroscopy
A Nicolet wide-bore NT 360-MHz NMR spectrometer operating at 95.24 MHz for 23Na and 360 MHz for 'H was used for all experiments. 23Na spectra were acquired with 90" pulses and an interpulse delay of 0.25 s using either a 20-mm probe for rat, guinea pig, and rabbit hearts weighing approximately 1.25 g or a 30-mm probe for ferret hearts weighing approximately 6 g. The sweep width was +2000 Hz and 1024 or 2048 data points were collected. Typically the free induction decays were processed by a baseline correction, exponential multiplication with 8-Hz line broadening, and Fourier transformation; no resolution-enhancing functions were used in these exper- iments. 'H spectra were acquired using a probe tuned to the sodium resonance which was sensitive enough to detect the large 'H signal at 360 MHz. A 2000-Hz sweep width and an interpulse delay of 10 s were used for the 'H spectra. A 5-Hz line broadening was introduced by the exponential multiplication used during signal pro- cessing.
Experimental Protocols
Hearts were perfused with normal perfusion buffer for a control perfusion period and then perfused with buffer containing shift reagent. After equilibration with shift reagent, the perfusate and mannitol bath flows were both stopped to provide a period of no-flow ischemia sufficient to produce moderate to severe injury. This required 28 min for rat, guinea pig, and rabbit, and 120 min for ferret. Subsequent to no-flow ischemia, the ferret heart also experienced a 60-min period of ischemia with only the mannitol bath flowing. Finally the flows of perfusate and mannitol bath were restored for a period of reflow. In some instances the shift reagent was omitted from the perfusate

18 KOHLER ET AL.
and included only in the mannitol bath to provide a situation where all the heart sodium signal remained unshifted.
Vasculature washout. Rat hearts were perfused with normal perfusion buffer for 20 min and then with shift reagent buffer for 20 min. The perfusion medium was then switched to one of the mannitol, choline, or lithium low-sodium buffers for vasculature washout. These experiments were done both with and without the mannitol bath solution flowing.
Atrial mixing. Spectra were obtained for hearts perfused with normal perfusion buffer without shift reagent. The bath line delivering mannitol with shift reagent was either placed in its normal position at the bottom of the sample chamber or introduced into the right atrium of the heart. The aspirating drain remained at the top of the chamber in both cases.
Glass heart experiments. The sample consisted of a heart-shaped bulb placed inside a 20-mm NMR tube containing a 145 m M sodium chloride solution. A series of spectra was taken with the “heart” containing 145 m M NaCl, air, or solutions of NaCl and Dy( TTHA) 3- in varying concentrations.
RESULTS
Species Comparison
Representative spectra from experiments with rat, guinea pig, ferret, and rabbit hearts are shown in Fig. 1. All of the hearts were subjected to similar protocols consisting of a 20-min period of control normoxic perfusion followed by a variable period of 28 or 120 min no-flow ischemia and a period of reperfusion. In all cases, the spectra exhibit a very small resonance at approximately 0-0.8 ppm which increases in intensity during ischemia and then decreases during reperfusion. There is also a large, complex peak centered at approximately 3.5 ppm and spanning a 2-3 ppm range at half-height. This peak has at least one prominent upfield shoulder and one prominent downfield shoulder. The peak sharpens and shifts downfield during ischemia and returns to approximately its original shape upon reperfusion. The increase in area of this peak upon reperfusion is attributed to edema. Finally, all spectra have small downfield shoulders in the 5-7 ppm range. The most downfield of these shifts upfield, sharpens, and greatly increases in size during ischemia.
Spectral Analysis
Analysis of these heart spectra is complicated by the fact that there are more iden- tifiable resonances than there are physically distinct compartments. Sodium may be found in both the heart and its surrounding bath. Sodium in the heart is further partitioned into the intracellular, interstitial, and vascular regions, for a total of four distinguishable compartments. For the purposes of these experiments the veins, arteries, capillaries, and atrial and ventricular spaces are all considered as part of the vascular compartment since all are exposed to the same concentrations of sodium and shift reagent. The interstitium and intracellular spaces are distinct both from each other and from the vascular region.
In addition to the physical compartmentation, other factors may contribute to the observed spectra. Since the shift in the 23Na resonance frequency induced by the
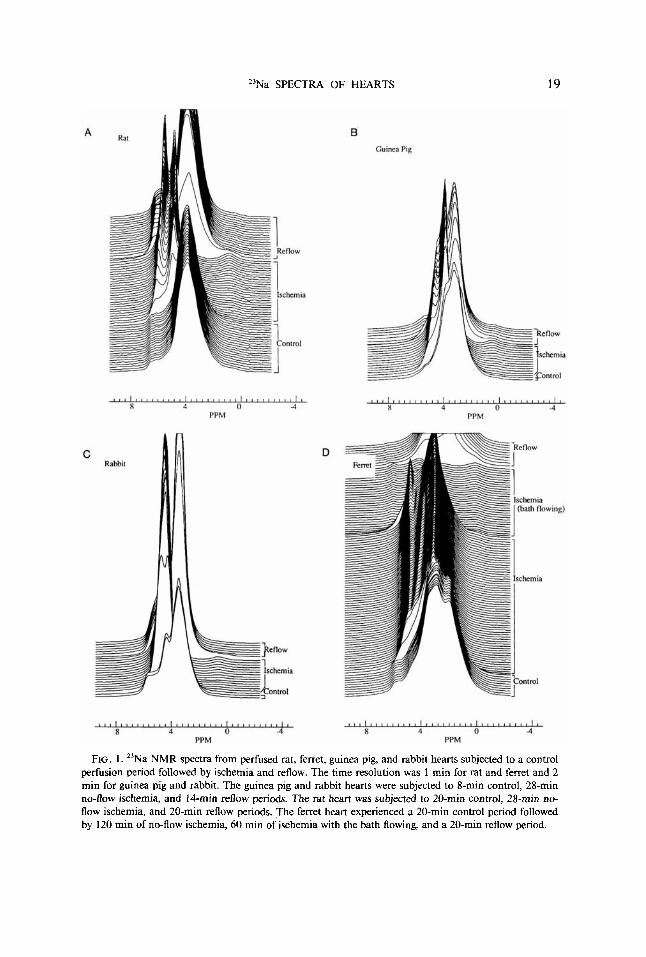
23Na SPECTRA OF HEARTS 19
FIG. 1. "Na NMR spectra from perfused rat, ferret, guinea pig, and rabbit hearts subjected to a control perfusion period followed by ischemia and reflow. The time resolution was I min for rat and ferret and 2 min for guinea pig and rabbit. The guinea pig and rabbit hearts were subjected to 8-min control, 28-min no-flow ischemia, and 14-min reflow periods. The rat heart was subjected to 20-min control, 28-min no- flow ischemia, and 20-min reflow periods. The ferret heart experienced a 20-min control period followed by 120 min of no-flow ischemia, 60 min of ischemia with the bath flowing, and a 20-min reflow period.

20 KOHLER ET AL.
paramagnetic shift reagent Dy( TTHA) 3- is dependent upon both bulk magnetic sus- ceptibility effects and the concentration ratio of shift reagent to sodium (8, l o ) , it was important to define the effects of bulk magnetic susceptibility variations and con- centration gradients within the sample. A series of experiments was undertaken to identify the contributions to the spectra from each of the four physical compartments and to evaluate the possible contributions of bulk magnetic susceptibility and con- centration gradient effects.
Vusculature washout. The vascular volume including capillaries and chambers is estimated to be at least one-half of the total volume of a perfused heart. Under normal perfusion the vascular space contains 145 m M sodium. Thus, the vascular sodium should contribute a major fraction of the total sodium signal. Morphometric analysis has shown ( I 1 ) that the vasculature of the heart collapses in the first few minutes of no-flow ischemia when the perfusion pressure head abruptly changes from 75- 100 Torr to zero. Therefore the sodium which was in the vascular region should be extruded into the surrounding bath and spectra taken during ischemia should show a downfield shift of this signal due to the change in the ratio of sodium to shift reagent. Indeed, examination of the spectra in Fig. 1 reveals that a large fraction of the intensity of the resonance at 3.5 ppm is shifted downfield during ischemia.
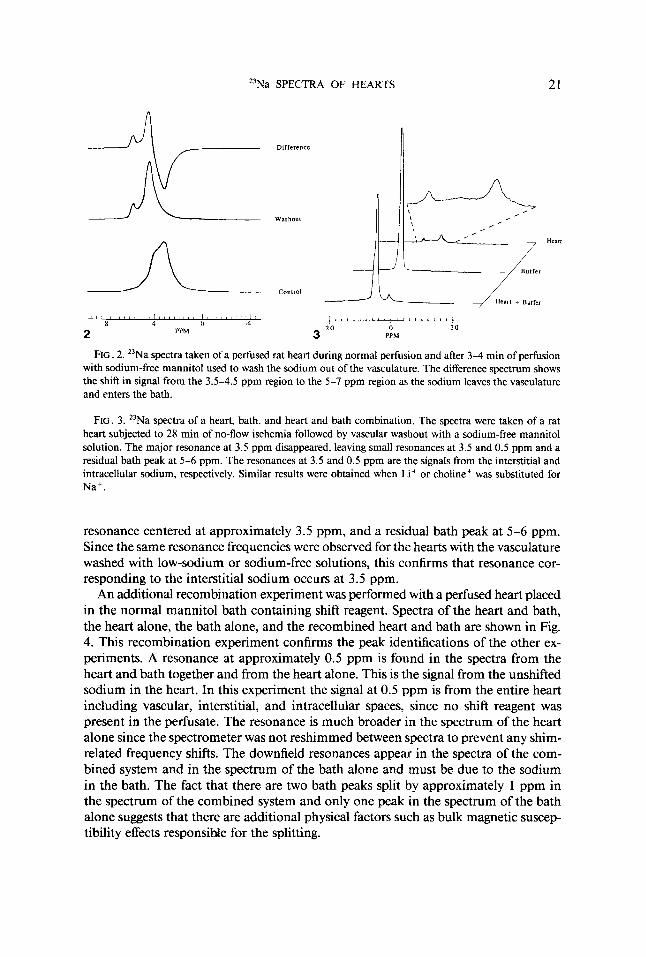
In order to confirm the identification of the vascular component of the spectrum at 3.5 ppm, the vascular region was flushed with a sodium-free mannitol solution or with buffer in which most of the sodium was replaced by lithium or choline. This procedure removes the vascular sodium signal, leaving the intracellular, interstitial, and bath contributions to the spectrum. Figure 2 contains spectra from an experiment with 1-min time resolution for a rat heart in which the vasculature was washed out with a sodium-free mannitol solution. A spectrum from the period of control perfusion is shown along with a spectrum taken in the 3-4 min interval after the washout procedure began. As shown in the difference spectrum, a large amount of signal was shifted from the 3.5-4.5 ppm region to the 5-7 ppm region. This represents a loss of the sodium signal at 3.5-4.5 ppm from the vasculature and a concomitant increase of the sodium signal at 5-7 ppm from the bath.
After the vasculature is washed out and the sodium in the bath is removed by continued mannitol flushing, only the signals from the interstitial and intracellular sodium should remain. In other experiments, spectra recorded at intervals of a few seconds were used to calculate the washout rates. The initial period of washout was fitted to an exponential decay for two mannitol flush experiments, and one experiment each with lithium and choline low-sodium buffers. The mean time constant for the initial vasculature washout in these four experiments was 0.25 min, with a range of 0.22-0.29 min. Thus 95% of the vascular sodium should be washed out within 0.75 min.
Recombination experiments. As shown in Fig. 2, the presence of a large amount of sodium in the bath obscures the interstitial signal in hearts even after vascular collapse in no-flow ischemia. To identify the position of the interstitial sodium resonance, experiments were performed in which the ischemic heart, which should contain sodium only in the intracellular and interstitial spaces, was removed from the bath; spectra were taken of the bath alone and the heart alone, as shown in Fig. 3. As anticipated, the heart spectra had three resonances, one at approximately 0.5 ppm, a very broad
23Na SPECTRA OF HEARTS 21
l , , , , , , , , , f , , , l , / l l l l l l , , l l , l l l I
PPM a 4 0 -4
2
Difference
Washout
Con1rol
Difference
J
i . I
, Heart
Washout
Itear1 + Buffer Con1rol
A
Heart
FIG. 2. "Na spectra taken of a perfused rat heart during normal perfusion and after 3-4 min of perfusion with sodium-free mannitol used to wash the sodium out of the vasculature. The difference spectrum shows the shift in signal from the 3.5-4.5 ppm region to the 5-7 ppm region as the sodium leaves the vasculature and enters the bath.
FIG. 3. "Na spectra of a heart, bath, and heart and bath combination. The spectra were taken of a rat heart subjected to 28 min of no-flow ischemia followed by vascular washout with a sodium-free mannitol solution. The major resonance at 3.5 ppm disappeared, leaving small resonances at 3.5 and 0.5 ppm and a residual bath peak at 5-6 ppm. The resonances at 3.5 and 0.5 ppm are the signals from the interstitial and intracellular sodium, respectively. Similar results were obtained when Li+ or choline+ was substituted for Na+.
resonance centered at approximately 3.5 ppm, and a residual bath peak at 5-6 ppm. Since the same resonance frequencies were observed for the hearts with the vasculature washed with low-sodium or sodium-free solutions, this confirms that resonance cor- responding to the interstitial sodium occurs at 3.5 ppm.
An additional recombination experiment was performed with a perfused heart placed in the normal mannitol bath containing shift reagent. Spectra of the heart and bath, the heart alone, the bath alone, and the recombined heart and bath are shown in Fig. 4. This recombination experiment confirms the peak identifications of the other ex- periments. A resonance at approximately 0.5 ppm is found in the spectra from the heart and bath together and from the heart alone. This is the signal from the unshifted sodium in the heart. In this experiment the signal at 0.5 ppm is from the entire heart including vascular, interstitial, and intracellular spaces, since no shift reagent was present in the perfusate. The resonance is much broader in the spectrum of the heart alone since the spectrometer was not reshimmed between spectra to prevent any shim- related frequency shifts. The downfield resonances appear in the spectra of the com- bined system and in the spectrum of the bath alone and must be due to the sodium in the bath. The fact that there are two bath peaks split by approximately 1 ppm in the spectrum of the combined system and only one peak in the spectrum of the bath alone suggests that there are additional physical factors such as bulk magnetic suscep- tibility effects responsible for the splitting.
22 KOHLER ET AL.
d l 8 1 I 1 1 / 1 I I I 1 8 ’ ’ , 8 > f 1 1 I , , , , ’ ’ , 1 , / , / I I , I I , 8 1 8 I 1 I , # I I , 1 8 , I I I , , I 1 I 8 4 0 4
PPM 5 PPM 4
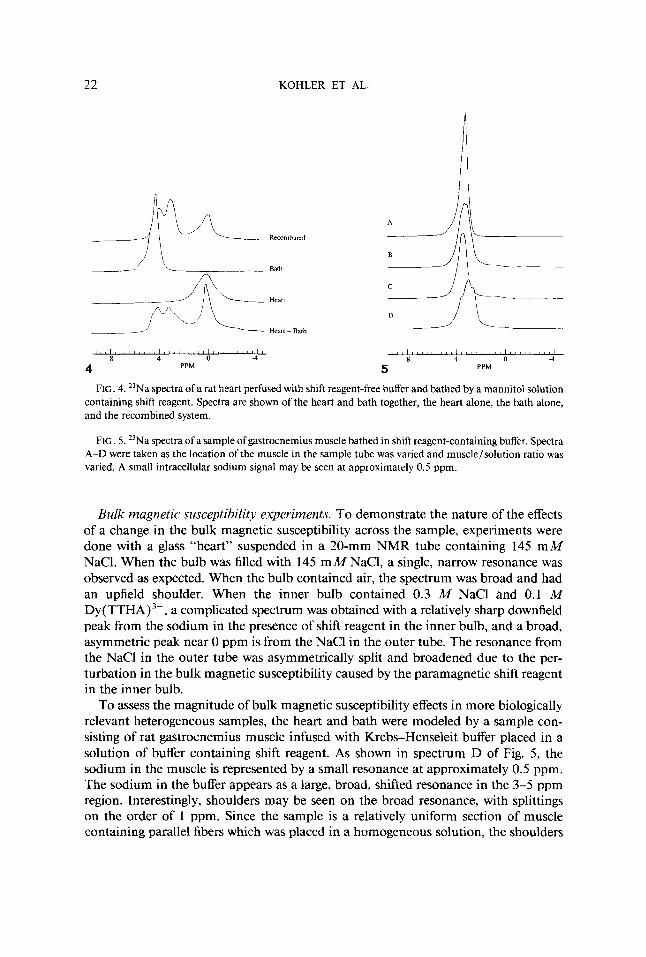
FIG. 4. 23Na spectra of a rat heart perfused with shift reagent-free buffer and bathed by a mannitol solution containing shift reagent. Spectra are shown of the heart and bath together, the heart alone, the bath alone, and the recombined system.
FIG. 5. 23Na spectra of a sample of gastrocnemius muscle bathed in shift reagent-containing buffer. Spectra A-D were taken as the location of the muscle in the sample tube was vaned and muscle/solution ratio was vaned. A small intracellular sodium signal may be seen at approximately 0.5 ppm.
Bulk magnetic susceptibility experiments. To demonstrate the nature of the effects of a change in the bulk magnetic susceptibility across the sample, experiments were done with a glass “heart” suspended in a 20-mm NMR tube containing 145 m M NaC1. When the bulb was filled with 145 m M NaC1, a single, narrow resonance was observed as expected. When the bulb contained air, the spectrum was broad and had an upfield shoulder. When the inner bulb contained 0.3 M NaCl and 0.1 M DY(TTHA)~-, a complicated spectrum was obtained with a relatively sharp downfield peak from the sodium in the presence of shift reagent in the inner bulb, and a broad, asymmetric peak near 0 ppm is from the NaCl in the outer tube. The resonance from the NaCl in the outer tube was asymmetrically split and broadened due to the per- turbation in the bulk magnetic susceptibility caused by the paramagnetic shift reagent in the inner bulb.
To assess the magnitude of bulk magnetic susceptibility effects in more biologically relevant heterogeneous samples, the heart and bath were modeled by a sample con- sisting of rat gastrocnemius muscle infused with Krebs-Henseleit buffer placed in a solution of buffer containing shift reagent. As shown in spectrum D of Fig. 5 , the sodium in the muscle is represented by a small resonance at approximately 0.5 ppm. The sodium in the buffer appears as a large, broad, shifted resonance in the 3-5 ppm region. Interestingly, shoulders may be seen on the broad resonance, with splittings on the order of 1 ppm. Since the sample is a relatively uniform section of muscle containing parallel fibers which was placed in a homogeneous solution, the shoulders
"Na SPECTRA OF HEARTS 23
on the solution resonance must be due to variations in the local magnetic field within the sample caused by changes in the bulk magnetic susceptibility between the muscle and the solution containing the paramagnetic Dy ( TTHA)3-. Confirmation of this comes from the observation that the splittings seen in these spectra are highly dependent on the sample geometry; variations either in the ratio of muscle to solution or in the position of the muscle within the solution alter the fine details of the spectrum, as seen in spectra A-C of Fig. 5.
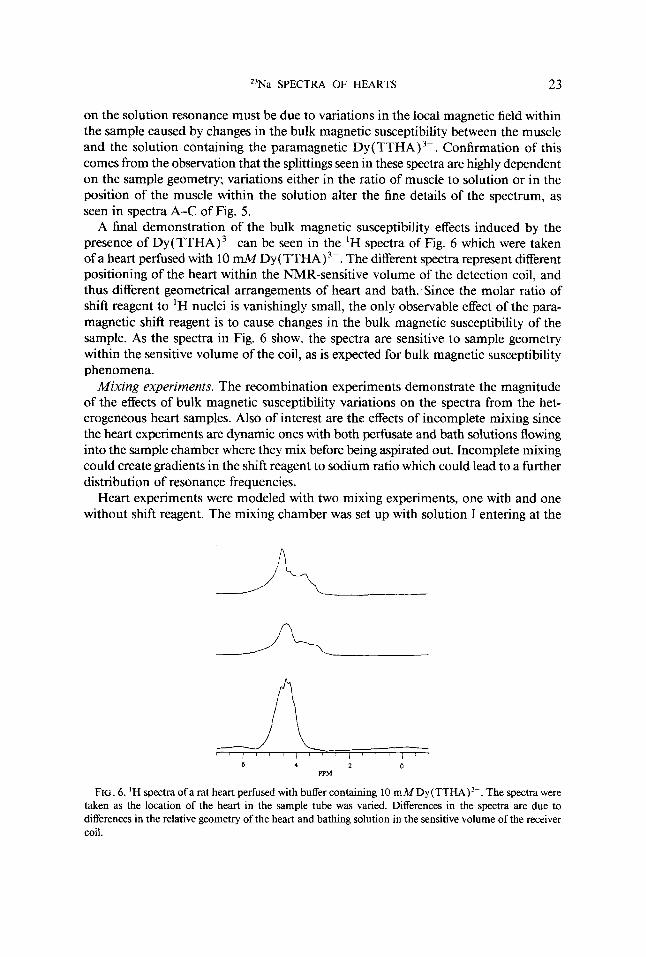
A final demonstration of the bulk magnetic susceptibility effects induced by the presence of DY(TTHA)~- can be seen in the 'H spectra of Fig. 6 which were taken of a heart perfused with 10 ~ M D Y ( T T H A ) ~ - . The different spectra represent different positioning of the heart within the NMR-sensitive volume of the detection coil, and thus different geometrical arrangements of heart and bath. Since the molar ratio of shift reagent to 'H nuclei is vanishingly small, the only observable effect of the para- magnetic shift reagent is to cause changes in the bulk magnetic susceptibility of the sample. As the spectra in Fig. 6 show, the spectra are sensitive to sample geometry within the sensitive volume of the coil, as is expected for bulk magnetic susceptibility phenomena.
Mixing experiments. The recombination experiments demonstrate the magnitude of the effects of bulk magnetic susceptibility variations on the spectra from the het- erogeneous heart samples. Also of interest are the effects of incomplete mixing since the heart experiments are dynamic ones with both perfusate and bath solutions flowing into the sample chamber where they mix before being aspirated out. Incomplete mixing could create gradients in the shift reagent to sodium ratio which could lead to a further distribution of resonance frequencies.
Heart experiments were modeled with two mixing experiments, one with and one without shift reagent. The mixing chamber was set up with solution I entering at the
6 4 2 0 PPM
FIG. 6. 'H spectra o fa rat heart perfused with buffer containing 10 ~ M D Y ( T T H A ) ~ - . The spectra were taken as the location of the heart in the sample tube was vaned. Differences in the spectra are due to differences in the relative geometry of the heart and bathing solution in the sensitive volume of the receiver coil.

24 KOHLER ET AL.
top of the cell, solution I1 entering near the bottom, and the drain near the top. Solution I1 flowed into the cell at twice the rate of solution I in both experiments. This is the physical arrangement of solution entry and exit ports used in the heart experiments in which the solution I1 (mannitol bath) flow rate was either 1.5 or 2 times the solution I (perfusate) flow. Spectra from a simple mixing experiment in which solution I contained 150 m M NaCl and solution I1 contained no sodium have one narrow resonance at 0 ppm. The integrated area under this peak fell by a factor of 2.4 when the flow of solution I1 was turned on. Thus the steady-state concentration of sodium in the sample tube with both solutions flowing was 62 mM, close to the value of 50 m M anticipated in the case of complete mixing.
The spectra became more interesting when 6 m M DY(TTHA)~- shift reagent was added to each of the solutions. In this case the integrated area again dropped by a factor of 2.4 when the flow of solution I1 was turned on, but the shape of the resonance changed as well. The main peak at approximately 4 ppm acquired a prominent shoulder approximately 1 ppm upfield. The location and intensity of the shoulder were quite stable, reflecting the establishment of a new steady state. Similar spectra were seen when the chamber was modified such that flow of solution I was impeded by a small latex balloon with three holes cut in it. Spectra from this experiment are shown in Fig. 7. In this case the upfield shoulder was more prominent, and an even smaller very broad shoulder was seen approximately 1 ppm further upfield. When the flows of both solutions were stopped, simulating no-flow ischemia conditions, the prominent shoulder collapsed, but the smaller shoulder remained. Since the shift reagent con- centration was the same throughout, these experiments demonstrate the spectral com- plexity introduced by a variation in the shift reagent to sodium ratio in different regions of the chamber. When flow to the chamber is stopped, diffusion equalizes the shift reagent to sodium ratio in the main part of the sample tube and the prominent shoulder disappears. The small shoulder remains, perhaps reflecting a difference in the shift reagent to sodium ratio between the small latex balloon and the much larger external solution. Due to the impedance of diffusion by the balloon such a difference would be expected to persist for much longer.
Atrial mixing. The results from the simple mixing experiments and the recombi- nation experiments show that the signal from the bath may be spread over a wide range of chemical shifts due to both mixing effects and bulk magnetic susceptibility effects. In typical heart experiments, the heart almost fills the central portion of the NMR tube. The bathing solution is introduced at the bottom of the tube, and the bath and perfusate are aspirated from the top. Experiments done with the dye methylene blue in the perfusate have shown that the mixing of the bath and perfusate is probably incomplete ( 5 ) . The extent of mixing achieved is likely to be different during the control and reflow periods since the heart shrinks during ischemia and becomes edem- atous during reflow. Further, the extent of mixing during the period of ischemia itself will again be different since all flows are turned off. In an effort to assess the magnitude of mixing effects in experiments with intact hearts, experiments were done in which the site of introduction of the bathing solution was varied. The case in which the bath flow entered at the bottom of the NMR tube was compared to that in which the mannitol bath line was inserted into the right atrium of the heart. In both cases, the aspirating drain remained at the top of the chamber. When the bath was introduced
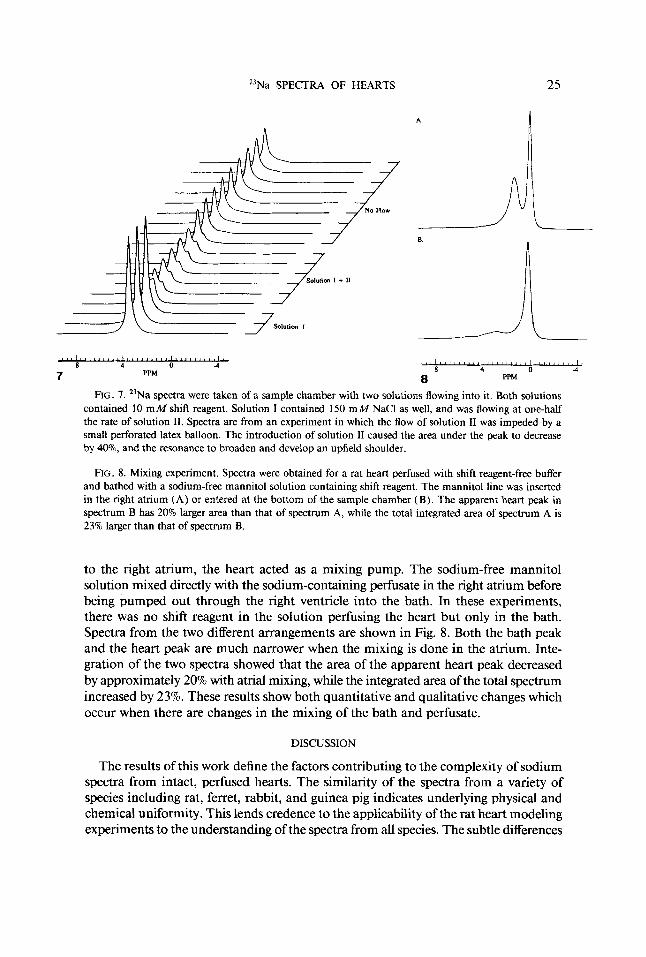
23Na SPECTRA OF HEARTS 25
FIG. 7. 23Na spectra were taken of a sample chamber with two solutions flowing into it. Both solutions contained 10 m M shift reagent. Solution I contained 150 m M NaCl as well, and was flowing at one-half the rate of solution 11. Spectra are from an experiment in which the flow of solution I1 was impeded by a small perforated latex balloon. The introduction of solution I1 caused the area under the peak to decrease by 40'70, and the resonance to broaden and develop an upfield shoulder.
FIG. 8. Mixing experiment. Spectra were obtained for a rat heart perfused with shift reagent-free buffer and bathed with a sodium-free mannitol solution containing shift reagent. The mannitol line was inserted in the right atrium ( A ) or entered at the bottom of the sample chamber (B). The apparent heart peak in spectrum B has 20% larger area than that of spectrum A, while the total integrated area of spectrum A is 23% larger than that of spectrum B.
to the right atrium, the heart acted as a mixing pump. The sodium-free mannitol solution mixed directly with the sodium-containing perfusate in the right atrium before being pumped out through the right ventricle into the bath. In these experiments, there was no shift reagent in the solution perfusing the heart but only in the bath. Spectra from the two different arrangements are shown in Fig. 8. Both the bath peak and the heart peak are much narrower when the mixing is done in the atrium. Inte- gration of the two spectra showed that the area of the apparent heart peak decreased by approximately 20% with atrial mixing, while the integrated area of the total spectrum increased by 23%. These results show both quantitative and qualitative changes which occur when there are changes in the mixing of the bath and perfusate.
DISCUSSION
The results of this work define the factors contributing to the complexity of sodium spectra from intact, perfused hearts. The similarity of the spectra from a variety of species including rat, ferret, rabbit, and guinea pig indicates underlying physical and chemical uniformity. This lends credence to the applicability of the rat heart modeling experiments to the understanding of the spectra from all species. The subtle differences

26 KOHLER ET AL.
which have been observed in the spectra from different species may be due to effects caused by differences in heart geometry and size as well as by varying degrees of vascularization due to differences in age and species.
Our results with model systems illustrate the effects of variations in bulk magnetic susceptibility and concentration gradients in the biological samples. The bulk magnetic susceptibility effects have been shown to cause splittings on the order of one part per million in both physiological and nonphysiological samples. These effects are pro- nounced because DY(TTHA)~- is paramagnetic and regions of the sample with high Dy (TTHA) 3- concentration have a significantly different bulk magnetic susceptibility than neighboring regions of low Dy( TTHA) 3- concentration. The bulk magnetic susceptibility effects are highly dependent on the geometry and orientation of the shift reagent-containing compartments. The bulk magnetic susceptibility effects are most obvious in the splitting and broadening of the bath resonances, but also cause the other resonances to be inhomogeneous as well. Similar effects have been reported in a study of the field dependence of 23Na spectra of rat gastrocnemius muscle infused with shift reagent (3 ) .
We have identified the contributions to the sodium NMR spectrum from the four physical compartments within our heart samples. The intracellular sodium resonance occurs at approximately 0.5 ppm, and in spectra taken during the period of control perfusion this peak contributes less than 3% of the total signal in a rat heart bathed with a sodium-free mannitol solution. The interstitial sodium signal is a broad reso- nance centered at about 3.5 ppm, as is the sodium in the vascular region. Depending upon the completeness of mixing before aspiration, the resonance from the sodium in the bath may be spread over a wide spectral range due to gradients in the shift reagent to sodium ratio and bulk magnetic susceptibility effects from inhomogeneous distribution of the shift reagent. During ischemia when all flows are stopped, sodium is extruded from the heart into the bath and the bath resonances sharpen. On the basis of our analysis of the effects of mixing, these resonances in the bath would be expected to sharpen as the perfusate and bath mix more completely and the shift reagent to sodium ratio in the bath becomes more uniform. During ischemia when the bath sodium concentration is high and the peaks are sharp, it is often possible to see splittings of up to 1 ppm in the bath resonances due to bulk magnetic susceptibility effects. Thus both mixing and bulk magnetic susceptibility effects contribute to the changes in the spectra which occur during ischemia.
The results from these experiments are important for our interpretation of sodium NMR spectra from perfused hearts and have identified several important factors which need be considered when designing heart experiments. This information will provide a basis for a quantitative analysis of the spectral changes which occur during conditions such as ischemia and reflow.
ACKNOWLEDGMENTS
This work was supported in part by National Institutes of Health Grant PO1 HL38189. S.B.P. was supported by a Clinical Investigator Award HL 02246 from National Institutes of Health, L.C.S. was a Medical Research Council of Canada Fellow, D.E.A. was a Fellow of the Sarnoff Foundation, and K. C. was the Howard B. Sprague Fellow of the Massachusetts Affiliate of the American Heart Association.

23Na SPECTRA OF HEARTS 27
REFERENCES
1. M. M. PIKE, E. T. FOSSEL, T. W. SMITH, AND C. S. SPRINGER, Amer. J. Physiol. 246, C528 ( 1984). 2. H. HOFELER, D. JENSEN, M. M. PIKE, J. L. DELAYRE, v. P. CIRILLO, C. s. SPRINGER, E. T. FOSSEL,
AND J. A. BALSCHI, Biochemistry 26,4953 (1987).
138 (1989). 3. J. A. BALSCHI, S. J. KOHLER, J. A. BITTL, C. S. SPRINGER, AND J. S. INGWALL, J. Magn. Reson. 83,
4. J. A. BALSCHI, J. A. BITTL, C. S. SPRINGER, AND J. S. INGWALL, NMR Biomed. 3, 47 (1990). 5. M. M. PIKE, J. C. FRAZER, D. F. DEDRICK, J. S. INGWALL, P. D. ALLEN, C. S. SPRINGER, AND T. W.
6. J. A. BALSCHI, J. C. FRAZER, J. K. FETTERS, K. CLARKE, C. S. SPRINGER, T. W. SMITH, AND J. S.
7. D. BURSTEIN, Biophys. J. 54, 191 (1988). 8. S. C.-K. CHU, Y. Xu, J. A. BALSCHI, AND C. S. SPRINGER, Magn. Reson. Med. 13,239 (1990). 9. S. J. KOHLER, K. CLARKE, S. B. PERRY, D. E. ATKINSON, L. C. STEWART, T. KONISHI, AND J. S.
INGWALL, “Proceedings of the Eighth Annual Meeting of the Society of Magnetic Resonance in Medicine,” p. 500, 1989.
10. S. C.-K. CHU, M. M. PIKE, E. T. FOSSEL, T. W. SMITH, J. A. BALSCHI, AND C. S. SPRINGER, J. Magn. Reson. 56, 33 (1984).
11. S. HUMPHREY, personal communication.
SMITH, Biophys. J. 48, 159 (1985).
INGWALL, Circulation 72(11), 1420 (1985).
Related Documents


![Roleof carcinoma perfused by31P NMR - PNAS · Proc. Natl. Acad. Sci. USA90 (1993) 2647 erage, 1.4 g); diameter, 1.2-1.8 cm]. Surgery to isolate and perfusethetumorexvivowasperformedbyaprocedure(27](https://static.cupdf.com/doc/110x72/5f0f5f5f7e708231d443d5a1/roleof-carcinoma-perfused-by31p-nmr-pnas-proc-natl-acad-sci-usa90-1993-2647.jpg)








