Tumor Biology and Immunology An Orally Active Galectin-3 Antagonist Inhibits Lung Adenocarcinoma Growth and Augments Response to PD-L1 Blockade Lynda Vuong 2 , Eleni Kouverianou 1 , Claire M. Rooney 2 , Brian J. McHugh 1 , Sarah E.M. Howie 1 , Christopher D. Gregory 1 , Stuart J. Forbes 3 , Neil C. Henderson 1 , Fredrik R. Zetterberg 4 , Ulf J. Nilsson 5 , Hakon Leffler 6 , Paul Ford 4 , Anders Pedersen 4 , Lise Gravelle 4 , Susan Tantawi 4 , Hans Schambye 4 , Tariq Sethi 2 , and Alison C. MacKinnon 1 Abstract A combination therapy approach is required to improve tumor immune infiltration and patient response to immune checkpoint inhibitors that target negative regulatory receptors. Galectin-3 is a b-galactoside-binding lectin that is highly expressed within the tumor microenvironment of aggressive cancers and whose expression cor- relates with poor survival particularly in patients with non–small cell lung cancer (NSCLC). To examine the role of galectin-3 inhibition in NSCLC, we tested the effects of galectin-3 deple- tion using genetic and pharmacologic approaches on syngeneic mouse lung adenocarcinoma and human lung adenocarcinoma xenografts. Galec- tin-3 / mice developed significantly smaller and fewer tumors and metastases than syngeneic C57/ Bl6 wild-type mice. Macrophage ablation retard- ed tumor growth, whereas reconstitution with galectin-3-positive bone marrow restored tumor growth in galectin-3 / mice, indicating that macrophages were a major driver of the antitu- mor response. Oral administration of a novel small molecule galectin-3 inhibitor GB1107 reduced human and mouse lung adenocarcinoma growth and blocked metastasis in the syngeneic model. Treatment with GB1107 increased tumor M1 macrophage polarization and CD8 þ T-cell infiltration. Moreover, GB1107 potentiated the effects of a PD-L1 immune checkpoint inhibitor to increase expression of cytotoxic (IFNg , granzyme B, perforin-1, Fas ligand) and apoptotic (cleaved caspase-3) effector molecules. In summary, galectin-3 is an important regulator of lung adenocarcinoma progression. The novel galectin-3 inhibitor presented could provide an effective, nontoxic monotherapy or be used in combination with immune checkpoint inhibitors to boost immune infiltration and responses in lung adenocarcinoma and potentially other aggressive cancers. Significance: A novel and orally active galectin-3 antagonist inhibits lung adenocarcinoma growth and metastasis and augments response to PD-L1 blockade. Graphical Abstract: http://cancerres.aacrjournals.org/content/canres/79/7/1480/F1.large.jpg. 1 Centre for Inflammation Research, Queen's Medical Research Institute, University of Edinburgh, Edinburgh Bioquarter, Edinburgh, United Kingdom. 2 Department of Asthma, Allergy and Respiratory Science, King's College London, Guy's Hospital, London, United Kingdom. 3 MRC Centre for Regenerative Medicine, University of Edinburgh, Edinburgh Bioquarter, Edinburgh, United Kingdom. 4 Galecto Biotech, Copenhagen, Denmark. 5 Department of Chemistry, Centre for Analysis and Synthesis, Lund University, Lund, Sweden. 6 Department of Laboratory Medicine, Section MIG, Lund University, Lund, Sweden. Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). T. Sethi and A.C. MacKinnon contributed equally to this article. Corresponding Author: Alison C. MacKinnon, University of Edinburgh, Edin- burgh Bioquarter, 47 Little France Crescent, Edinburgh EH16 4TJ, UK. Phone: 44- 0-1312426855; E-mail: [email protected] doi: 10.1158/0008-5472.CAN-18-2244 Ó2019 American Association for Cancer Research. Cancer Research Cancer Res; 79(7) April 1, 2019 1480 on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Tumor Biology and Immunology
An Orally Active Galectin-3 Antagonist InhibitsLung Adenocarcinoma Growth and AugmentsResponse to PD-L1 BlockadeLyndaVuong2, Eleni Kouverianou1,ClaireM. Rooney2, Brian J.McHugh1, Sarah E.M. Howie1,Christopher D. Gregory1, Stuart J. Forbes3, Neil C. Henderson1, Fredrik R. Zetterberg4,Ulf J. Nilsson5, Hakon Leffler6, Paul Ford4, Anders Pedersen4, Lise Gravelle4,Susan Tantawi4, Hans Schambye4, Tariq Sethi2, and Alison C. MacKinnon1
Abstract
A combination therapy approach is required toimprove tumor immune infiltration and patientresponse to immune checkpoint inhibitors thattarget negative regulatory receptors. Galectin-3 isa b-galactoside-binding lectin that is highlyexpressed within the tumor microenvironmentof aggressive cancers and whose expression cor-relates with poor survival particularly in patientswith non–small cell lung cancer (NSCLC). Toexamine the role of galectin-3 inhibition inNSCLC, we tested the effects of galectin-3 deple-tionusing genetic andpharmacologic approacheson syngeneic mouse lung adenocarcinoma andhuman lung adenocarcinoma xenografts. Galec-tin-3�/�mice developed significantly smaller andfewer tumors andmetastases than syngeneicC57/Bl6 wild-type mice. Macrophage ablation retard-ed tumor growth, whereas reconstitution withgalectin-3-positive bone marrow restored tumorgrowth in galectin-3�/� mice, indicating thatmacrophages were a major driver of the antitu-mor response. Oral administration of a novelsmall molecule galectin-3 inhibitor GB1107 reduced human andmouse lung adenocarcinoma growth and blocked metastasisin the syngeneic model. Treatment with GB1107 increased tumor M1 macrophage polarization and CD8þ T-cell infiltration.Moreover, GB1107 potentiated the effects of a PD-L1 immune checkpoint inhibitor to increase expression of cytotoxic (IFNg ,granzyme B, perforin-1, Fas ligand) and apoptotic (cleaved caspase-3) effector molecules. In summary, galectin-3 is animportant regulator of lung adenocarcinoma progression. The novel galectin-3 inhibitor presented could provide an effective,nontoxic monotherapy or be used in combination with immune checkpoint inhibitors to boost immune infiltration andresponses in lung adenocarcinoma and potentially other aggressive cancers.
Significance: A novel and orally active galectin-3 antagonist inhibits lung adenocarcinoma growth and metastasis andaugments response to PD-L1 blockade.
Graphical Abstract: http://cancerres.aacrjournals.org/content/canres/79/7/1480/F1.large.jpg.
1Centre for Inflammation Research, Queen's Medical ResearchInstitute, University of Edinburgh, Edinburgh Bioquarter, Edinburgh,United Kingdom. 2Department of Asthma, Allergy and RespiratoryScience, King's College London, Guy's Hospital, London, UnitedKingdom. 3MRC Centre for Regenerative Medicine, University ofEdinburgh, Edinburgh Bioquarter, Edinburgh, United Kingdom.4Galecto Biotech, Copenhagen, Denmark. 5Department of Chemistry,Centre for Analysis and Synthesis, Lund University, Lund, Sweden.6Department of Laboratory Medicine, Section MIG, Lund University,Lund, Sweden.
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
T. Sethi and A.C. MacKinnon contributed equally to this article.
Corresponding Author: Alison C. MacKinnon, University of Edinburgh, Edin-burgh Bioquarter, 47 Little France Crescent, EdinburghEH16 4TJ, UK. Phone: 44-0-1312426855; E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-18-2244
�2019 American Association for Cancer Research.
CancerResearch
Cancer Res; 79(7) April 1, 20191480
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

IntroductionGlobally, lung cancer is the leading cause of cancer-related
mortality (1). Non–small cell lung carcinoma (NSCLC) com-prises 80% of total lung cancer cases, with lung adenocarcino-ma being the major subtype (1). In recent years immunecheckpoint therapies targeting various negative regulatoryreceptors on tumor infiltrating cytotoxic T lymphocytes (CTL)such as programmed death-1 (PD-1), programmed death-ligand 1 (PD-L1), cytotoxic T-lymphocyte associated protein4 (CTLA-4), and others, have shown unprecedented efficacy inNSCLC patients even against late-stage disease (2). However,patient response is limited, thus driving intensive researchtoward combining immune checkpoint inhibition with othertargeted agents to overcome resistance (2).
Tumor-associated macrophages (TAM) are present in the stro-ma of many tumors including NSCLC (3). TAMs acquire analternative (M2)-like macrophage phenotype and secrete angio-genic and anti-inflammatory cytokines, which contribute to theimmunosuppressive milieu of the tumor microenvironment (4).TAMs can also be important direct targets of PD-1/PD-L1inhibition and can also promote drug resistance by removinganti-PD-1 antibodies from T cells (5, 6). Indeed, macrophagedepletion via colony-stimulating factor-1 receptor (CSF-1R)blockade improved T-cell infiltration and antitumor activity ofPD-1 antagonists in preclinical models of melanoma and breastcancer (7, 8), suggesting that strategies aimed at inhibiting mac-rophage responses are necessary to permit effective immunecheckpoint therapy.
One possible target for such combination treatment is galectin-3, a member of a protein family defined by affinity for b-galac-toside-containing glycoconjugates and a conserved carbohydrate-recognition-binding domain (9). Galectin-3 is widely expressedin several cell types such as macrophages, fibroblasts, activatedT-lymphocytes and epithelial cells (10–12) and is highlyexpressed in high fatality cancers such as NSCLC (13). In NSCLCparticularly in adenocarcinoma, increased galectin-3 expressionin tumors, lymphnodes and serum correlates withmetastases andis a negative prognostic indicator (13–18). The galectin-3 geneticpolymorphism rs4652 associated with impaired galectin-3 secre-tion, has been linked to increased survival and response tochemotherapy in NSCLC (18). Galectin-3 can directly enhancecell proliferation (19), apoptosis resistance (20), metastaticpotential (19, 21), as well as lung cancer stemness (22). It is alsoan important constituent of the tumor microenvironment actingon endothelial cells to promote angiogenesis (23). Furthermoremany studies have revealed the inhibitory effects of galectin-3 onactivated cytotoxic T lymphocytes CTLs (24–27) and we haveshown it to be essential for M2 macrophage differentiation (28,29). Hence, galectin-3 forms an ideal candidate target for com-bining with checkpoint blockade.
We examined the role of galectin-3 in NSCLC by utilizing thesyngeneic mouse Lewis Lung Carcinoma (LLC1) model, compar-ing tumor growth inwild-type (WT) andgalectin-3–deficientmiceshowing an essential non-redundant tumor-promoting role forgalectin-3. Bone marrow (BM) transfer and macrophage deple-tion experiments show that macrophages are a major source oftumor-promoting galectin-3. A newly developed, selective smallmolecule galectin-3 inhibitor inhibited mouse and humanNSCLC tumor growth and metastasis and significantly potenti-ated response to an immune checkpoint inhibitor.
Materials and MethodsCell lines, culture, and transfections
LLC1 cells and A549 cells were purchased from the EuropeanCell Culture Collection (ECACC 90020104) and were culturedat 37�C in 5% CO2 (95% air) in DMEM (Sigma D5671)supplemented with 10% FCS, 1% L-glutamine, and 1% peni-cillin/streptomycin. Human cell line A549 was authenticatedby short tandem repeat (STR) DNA profiling by ECACC. Cellswere routinely tested for Mycoplasma every 6 months. VectorpCMV-KDEL-Gluc-1, expressing G.princeps luciferase (LuxBiotechnologies) was transfected by electroporation (Lonzaelectroporation kit VCO-1001). Stably transfected cells wereselected with G418.
AnimalsAll animal experimental work was carried out under a project
license approved by the local Animal Welfare and EthicalReview body (AWERB) and issued in accordance with theAnimals (Scientific Procedures) Act 1986. C57Bl/6 mice andfemale CD1 nude mice were purchased from Harlan Labora-tories. Generation of galectin-3�/� mice by gene-targeting tech-nology has been described previously (30). CD11b-DTR (Diph-theria Toxin Receptor) mice were derived from FVB mice asdescribed (31) and backcrossed over 10 generations onto theC57Bl/6 background. Human galectin-3 knockin mice weregenerated by Cyagen Biosciences using the TurboKnockout(conditional Knockin) approach by inserting the entire humanLGALS3 sequence into exon 1 of mouse Lgals3 so that theexpression of human galectin-3 is under control of the mousegene regulatory element.
Orthotopic LLC1 modelMice were anaesthetized with isofluorane. A 1 mm skin
incision was made below the right shoulder blade. A total of103 LLC1-luciferase cells were injected through the intercostalmuscles into the lung parenchyma prior to the incision beingstapled.
Subcutaneous LLC1 modelLLC1 (2.5 � 105) cells were injected subcutaneously into the
flanks of age-matched male WT and galectin-3�/� C57Bl/6 mice.Each animal received an injection of 2.5� 105 cells suspended in100 mL PBS in both flanks. Tumor volumes were measured withcalipers every 1 to 3 days [tumor volume ¼ p/6 � (L � W)3/2].
LLC1 metastasis modelLLC1 cells were administered via the tail vein (1 � 106 cells)
and lung lobes were harvested at 7 days. RNA was extracted fromlung lobes using a Qiagen RNeasy Kit, converted into cDNA(Quantitect cDNA Synthesis Kit; Qiagen), and luciferase expres-sion was measured by qPCR using primers against Gaussiaprinceps luciferase (50-TCTGCCTGTCCCACATCAAG-30 forwardand 30-CCCTGTGCGGACTCTTTGT-50 reverse; Primer Design)and SYBR Green (ThermoFisher Scientific).
Human adenocarcinoma xenograft modelCD-1 nude female mice received 3 � 106 human lung adeno-
carcinoma cells (A549) in 100 mL 1:1 Matrigel:serum-free DMEMin both flanks. Tumor volumes were measured every 2 to 3 daysusing digital calipers.
Novel Galectin-3 Antagonist Inhibits Lung Cancer Progression
www.aacrjournals.org Cancer Res; 79(7) April 1, 2019 1481
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

Macrophage ablationMacrophages were ablated in C57Bl/6 CD11b-DTR mice
(or WT littermates) by administration of 10 ng/g diphtheriatoxin (DT) intraperitoneally (i.p.) prior to subcutaneous tumorcell injections.
BM transplantMice were injected with 400 mL liposomal clodronate
(Liposoma). After 36 hours, mice were irradiated with 10.5 Gydelivered from an IBL637 gamma irradiator (Gamma ServicesLtd.) at a dose rate of 0.64 Gy/min. Following irradiationmice received a single tail-vein infusion of 107 BM cellsobtained by flushing the femurs of WT and galectin-3�/�
donor mice. Transplanted mice were used 8 weeksposttransplant.
Drug preparationGalectin-3 inhibitor GB1107 (3,4-dichlorophenyl 3-deoxy-
3-[4(3,4,5-trifluorophenyl)-1H-1,2,3-triazol-1-yl]-1-thio-a-D-galactopyranoside; Galecto Biotech; ref. 32) was prepared at aconcentration of 1 mg/mL in 1% polyethylene glycol, 0.5%hydroxypropyl methyl cellulose (HPMC), and stored in ali-quots at �20�C. The anti-PD-L1 monoclonal antibody (clone10F.9G2) used for in vivo blockade experiments was purchasedfrom BioXCell and 200 mg in PBS was administered twiceweekly by interperitoneal injection.
Luciferase assaysLLC1 cells stably transfected with pCMV-KDEL-Gluc-1 were
assessed for luciferase expressionupon addition of n-colenterazine(n-CTZ; Lux Biotechnologies 20001) substrate to a final con-centration of 10 mmol/L to live cells in 96-well plates. Lymphnodes were disaggregated by passing through 40 mm cellstrainers and suspended in PBS. N-CTZ was added at a finalconcentration of 10 mmol/L. Luciferase activity was assessedwith a BioTek SynergyTM HT Luminometer.
ImmunohistochemistryFormalin-fixed paraffin embedded sections were deparaffi-
nized in xylene and rehydrated in graded ethanol. Epitopeswere retrieved by microwaving in 0.01M sodium citrate (pH 6)for Ym1, and cleaved caspase-3 staining and by proteinase Kdigest (1.25 mg/mL) for 5 minutes for F4/80 staining. Sectionswere blocked with serum-free protein block (DAKO) andincubated overnight at 4�C with primary antibodies, rabbitanti-mouse Ym1 (Stem Cell Technologies, 1:200), rat anti-mouse F4/80 (Abcam, 1:100), rabbit anti-mouse ki-67(Abcam, 1:200), and rabbit anti-mouse cleaved caspase-3(clone 5A1; Abcam, 1:1,000). Sections were incubated withspecies-specific biotinylated IgG (Vector), and visualized with3,30-diaminobenzidine (DAB) substrate.
Immunohistochemistry quantificationsFive to ten fields were scored for each tumor representing
both tumor and stroma. Absolute cell counts were recorded forF4/80 and Ym1 positive cells. Cleaved caspase-3 staining intumors was quantified by inverting 8-bit TIFF files so that DAB-positive areas give the highest pixel intensities. Mean pixelintensities (MPI) were then measured in up to 160 fields ofview covering the entire tumor parenchyma and averaged togive a single value per tumor.
ImmunofluorescenceTumor sections were incubated with rat anti-mouse F4/80
followed by horseradish peroxidase (HRP)-labeled goat anti-ratIgG (DAKO) and tyramide green (Invitrogen). Sections weremicrowaved in 0.01M sodium citrate (pH 6) for 5 minutes,reblocked, and probedwith rabbit anti galectin-3 (R&D) or rabbitanti Ym1 followed by HRP-labeled goat anti-rabbit IgG (DAKO)and tyramide red (Invitrogen) and mounted in fluoromount-Gwith DAPI (eBioscience). Images were captured on a NikonEclipse E600 microscope.
Tumor RNA extraction and RT-PCRTotal RNA from LLC1 tumors and lung tissue was prepared
using RNeasy kits (Qiagen) and reverse transcribed into cDNAusing Quantitect RT kits (Qiagen). cDNA was analyzed usingeither a SYBR green-based quantitative fluorescence method(Invitrogen) and Kiqstart primers (Sigma Aldrich) or Taqmanprimer probe sets (Life Technologies).
SDS PAGE and Western blottingCells were lysed inNP-40 (Invitrogen) and separated by 10% to
15% SDS-PAGE. Proteins were transferred to nitrocellulosemem-brane and probed using antibodies against galectin-3, 1:500(eBioM3/38; eBioscience) and GAPDH, 1:3,000 (14C10; CellSignaling Technology), followed by species specific HRP-conju-gated secondary antibodies (Dako). Bound antibodies weredetected using the Enhanced Chemiluminescence 2 DetectionKit (Pierce).
Tumor dissociation and flow cytometryTumors were minced in serum-free DMEM and digested with
Liberase (2 mg/mL; Sigma-Aldrich) and DNase I (Sigma-Aldrich)at 37�C for 30minutes.Disaggregated tissuewasfiltered through a35-mm nylon mesh, washed, and resuspended in FACS buffer(PBS with 0.1% BSA). Fc receptors were blocked with anti-mouseCD16/32 (Biolegend). Antibody cocktails (anti-mouse Ly6G-pacific blue, CD11b-BV605, Galectin-3-FITC, CD45-PerCP andCD45-APCcy7, MHC-II-PE, CD206-PEcy7, PD-1-APC, F4/80-AF700, CD4-pacific blue, PD-L1 BV605, CD3-PerCPcy5.5,IFNg-PE, CD8-AF700, all from Biolegend) were added to cellsand incubated for 20minutes at room temperature. Samples werefixed and RBCs were simultaneously lysed in RBC Lysis/Fixationsolution (Biolegend). For intracellular staining, cells were per-meabilized with intracellular staining permeabilization washbuffer (Biolegend) and incubated with anti-CD206 or anti-IFNg(Biolegend). Cells were analyzed using an LSR-Fortessa cell ana-lyzer (Beckton Dickenson).
Statistical analysisStatistical analyses were performed using Graphpad Prism 7.0
software. Results are represented as mean � SEM and statisticaltests are described in the figure legends.
ResultsGalectin-3�/�mice donot support the growth andmetastasis ofLLC1 tumors
To examine lung cancer growth within the correct tissue com-partment, LLC1 cells stably expressing G. princeps-luciferase wereinjected (1� 103 cells) through the intercostal space directly intothe lung parenchyma of control and galectin-3�/� mice.
Vuong et al.
Cancer Res; 79(7) April 1, 2019 Cancer Research1482
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

Hematoxylin and eosin staining of lung tissue confirmed thepresence of tumors in control but not galectin-3�/� mouse lungs(Fig. 1A). At 20 days post-injection, 4/10 of control mice hadtumors, whereas none of the galectin-3�/� mice developedtumors (Table 1). In addition, 7/10 control animals displayedgross swelling of the mediastinal lymph nodes (MLN; Table 1),which were positive for metastatic cells as assessed by luciferaseassay on homogenized MLNs. Only 1/11 of galectin-3�/� micehad luciferase positive MLNs (Table 1).
LLC1 cells expressing luciferase were injected subcutaneouslyin both flanks of WT and galectin-3�/� mice. After day 10,subcutaneous tumors from control animals were much largerthan those of galectin-3�/� mice. This difference became statis-tically significant at day 12 (P¼ 0.0004). By the end of the study,tumors of controls had an average volume of 286mm3 comparedwith a volume of 9 mm3 in galectin-3�/� animals (96.9% reduc-tion, P < 0.0001; Fig. 1B and C). The weight of tumors fromcontrol mice was 98% heavier than that of galectin-3�/� mice,
153� 31mg and 3� 2mg, respectively (P < 0.0001; Fig. 1D). Ofall the tumor cell injections receivedby each group, only 11/40 ledto tumors in galectin-3�/�mice compared with 36/40 in controls(Table 1). A total of 5/12 control mice had luciferase positivemetastases in their MLNs whereas galectin-3�/� mice had nometastases (Table 1). These results indicate that galectin-3�/�
mice do not support tumor establishment and spread in a sub-cutaneous LLC1model. Although LLC1 inoculation increased theserum concentrations of anti-galectin-3 IgG antibodies in galec-tin-3�/� mice, no correlation, either negative or positive, wasestablished between antibody production and tumor volume(Supplementary Fig. S1).
M2 macrophages are reduced in tumors from galectin-3�/�
miceTumor stroma F4/80þ macrophages were significantly higher
in galectin-3�/� animals compared with control (P ¼ 0.0217).However, the ratio of Ym1þ/F480þ macrophages was signifi-cantly higher in controls (P¼ 0.0484, Fig. 2A and B), indicatingthat higher galectin-3 levels around the tumor environmentcan drive expansion of M2 macrophages (28). Transcript anal-ysis from whole tumor RNA showed that control tumorshad 2.5-, 3.3-, and 16.7-fold higher levels of IL4, IL10,and IL13 transcripts, respectively (P ¼ 0.04, 0.024, 0.119,respectively), and displayed a 28.8-fold reduction inIFNg mRNA when compared with galectin-3�/� tumors (P ¼0.0066, Fig. 2C). These results indicate a cytokine environmentthat favors M2 macrophage activation in tumors of control butnot galectin-3�/� hosts and suggests an important role forgalectin-3 in the regulation of TAM phenotype.
Figure 1.
Galectin-3�/�mice do not supporttumor growth. LLC1 cells wereinjected into the lung parenchymaof age and sex-matched C57Bl/6WT and galectin-3�/�mice. Tumornodules were enumerated bymacroscopic counting and draininglymph nodes were checked formetastases by luciferase assay.A, Representative hematoxylin andeosin–stained lung sections fromWTand galectin-3�/�mice are shown.B–D, LLC1 cells were inoculatedsubcutaneously into both flanks ofWT or galectin-3�/�mice.Representative images (B), tumorvolume (C), and tumor weight on day19 post-dissection (D) are shown.Arrows, tumor sites. Data inA isderived from a single experiment,n¼ 10–11. Data from B–D arerepresentative of two independentexperiments, n¼ 8. Differences werecompared using an unpairedtwo-tailed t test; ��� , P < 0.001;���� , P < 0.0001 (compared withWT on the same day).
Table 1. Galectin-3�/� mice do not support tumor growth
Orthotopic tumorsGenotype No. ofmicewith primary tumors No. of mice with LNmetastases
WT 4/10 (40%) 7/10 (70%)Gal-3�/� 0/11 (0%) 1/11 (9%)Subcutaneous TumorsGenotype No. ofmicewith primary tumors No. of mice with LNmetastasesWT 36/40 (90%) 5/12 (41.7%)Gal-3�/� 11/40 (27.5%) 0/12 (0%)
NOTE: Prevalence of mice with established primary tumors and inguinal lymphnodemetastases are summarized. Orthotopic and subcutaneous tumor data arerepresentative of one or two independent experiments, respectively.
Novel Galectin-3 Antagonist Inhibits Lung Cancer Progression
www.aacrjournals.org Cancer Res; 79(7) April 1, 2019 1483
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

Figure 2.
Bone marrow–derived macrophages support LLC1 tumor growth. A, Subcutaneous tumors fromWT or galectin-3�/�mice were stained for F4/80 or Ym-1.B,Quantitation of Ym1 and F480 staining in tumor sections. C, RNAwas extracted from these tumors and gene expression of various cytokines was assessed byqPCR. D, LLC1 cells were injected subcutaneously in both flanks of CD11b DTRmice (n¼ 12) and their WT siblings (n¼ 11). All mice were administered DT (10 ng/g) prior to cell injection. D and E, Tumor volume (D) and tumor weights (E) following DT administration. F,WT and galectin-3�/�mice were treated withclodronate, irradiated, and subsequently transplanted with BM cells fromWT or galectin-3�/�mice. LLC1 cells were injected subcutaneously into both flanks oftransplanted mice and tumor volumes (F) and weights (G) were measured. H,Double immunoflourescence staining for F4/80 (green) and galectin-3 (red) wasperformed in tumors from galectin-3�/�mice receiving eitherWT or galectin-3�/� BM transplants. Dotted line represents the boundary between tumor cells (T)and stroma (S). Data are representative of eight mice per irradiation/transplant control group and 12 mice per experimental group. Two-tailed t tests were usedto assess statistical differences. � , P < 0.05; ��� , P < 0.001.
Vuong et al.
Cancer Res; 79(7) April 1, 2019 Cancer Research1484
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

Macrophage depletion impairs tumor initiationWe hypothesized that tumor macrophages may contribute to
tumor growth in the LLC1 model. C57Bl/6 CD11b-DTR trans-genic mice were used as a model of macrophage ablation (33).CD11b-DTR transgenic mice and WT siblings received a singlediphtheria toxin (DT) injection immediately prior to cell implant.At day 12, 15/24 tumors developed in CD11b-DTR micecompared with 20/22 in controls. CD11b-DTR animals hadsignificantly smaller tumor volumes compared with controls(29.4 � 4.1 mm3 and 89.4 � 0.9 mm3, respectively; P ¼0.0005, Fig. 2D) and significantly reduced tumor weights(9.1 � 1.0 and 23.4 � 4.0 mg, respectively; P ¼ 0.0011,Fig. 2E). To assess the efficiency of macrophage ablation in thismodel, DTwas administered tomice with established tumors andF4/80 staining carried out 24 hours after DT administration. An88% reduction in TAMs was observed in the tumors of DTRtransgenic animals (P < 0.0001; Supplementary Fig. S2A).
Galectin-3 phenotype of BM-derived cells in the tumormicroenvironment determines LLC1 tumor growth
To determine which galectin-3–expressing cells are necessaryto support tumor growth, we first altered galectin-3 expressionin recruited cells. Control and galectin-3�/� mice were irradi-ated and transplanted with 107 control or galectin-3�/� BMcells. Eight weeks post-BM transplant, LLC1 cells were injectedsubcutaneously. Transplantation of control BM cells into galec-tin-3�/� mice resulted in significantly increased average tumorvolume and final tumor weight compared with mice trans-planted with galectin-3�/� BM cells (final tumor volume 336mm3 and final weight of 297.6 mg compared with 163.9 and124.4 mg, P < 0.0001 and P ¼ 0.0007, respectively; Fig. 2F andG). Dual immunofluorescence staining showed that the stromaof tumors harvested from galectin-3�/� animals transplantedwith control BM had F4/80 and galectin-3 dual positive cells(Fig. 2H), although the total number of infiltrating macro-phages was not different between control or galectin-3�/� BMtransplanted mice (Supplementary Fig. S2B), suggesting thatgalectin-3 positive macrophages are recruited to the tumorstroma and contribute to tumor growth.
LLC1 cells in vitrodisplay cell surface and cytoplasmic galectin-3staining and release galectin-3 into the culture medium (Supple-mentary Fig. S3A). To determine whether tumor-derived galectin-3 also contributes to tumor growth, galectin-3 was stably knockeddown (KD) in LLC1 cells prior to subcutaneous injection (Sup-plementary Fig. S3B). Although LLC1 proliferation was reducedby galectn-3 KD in vitro (Supplementary Fig. S3C and S3D), tumorgrowth and final tumor weights of LLC1-galectin-3-KD cells wassimilar to WT cells (Supplementary Fig. S3E–S3G).
High-affinity galectin-3 inhibitor prevents human lungadenocarcinoma growth in vivo
Recently, a series of monosaccharide galectin-3 inhibitors withlow nmol/L affinities and good selectivity over other galectinshave been described (32). From this series GB1107 has highaffinity in man at 37 nmol/L but due to species differences inthe galectin-3 carbohydrate binding domain (CBD), the mousegalectin-3 affinity is 38-fold lower. GB1107 has low clearance(1.2ml/min/kg, t1/2¼ 4.5 hours, i.v.) and good uptake upon oraladministration, resulting in high oral availability (F ¼ 75%,orally). As a consequence, dosing GB1107 at 10 mg/kg orallyonce daily results in a plasma concentration abovemouse Kd over
24 hours (Supplementary Fig. S4). CD-1 nude mice bearinghuman lung A549 adenocarcinoma xenografts were treated fromday 18 postimplantation once daily with 10mg/kg GB1107. Thisresulted in significantly reduced tumor growth and final tumorweights (46.2% smaller compared with vehicle control tumorswith final average weights of 117 � 16 and 63 � 11 mg,respectively; P ¼ 0.0132; Fig. 3A). Treatment with GB1107 alsoinhibited LLC1 growth (tumor volumes decreased 48%comparedwith controls on day 18, P < 0.001) and reduced final tumorweights (47� 14mg vs. 120� 29mg controls, P¼ 0.0524) whenadministered daily from theoutset (Fig. 3B). Transcript analysis oftumor RNA from the LLC1 tumors revealed reduced galectin-3(48% less than vehicle, P ¼ 0.018) and mesenchymal markersTGFb (45% less than vehicle, P¼ 0.015) and trends for reductionsof VEGF and aSMA expression (Fig. 3C). Although there was nochange in the M1 marker Nos2, there was a trend towards areduction in expression of the M2 marker Ym1 (50% less thanvehicle) andCD98 (49% less than vehicle), which drives galectin-3–mediated M2 macrophage activation (28), suggesting adecrease in M2 skewed TAM accumulation in the tumor. Toexamine effects on metastasis, LLC1 cells were injected intrave-nously and lung colonization was determined at 7 days postin-jection (Fig. 3D). The presence of metastasis was examined byexpression of Gaussia luciferase transcript in whole lung RNAextracts. GB1107 administered daily from day 1 significantlyreduced tumor burden by 79.2%. These data suggest that inhibi-tion of galectin-3 with an orally active selective galectin-3 inhib-itor can significantly reduce lung adenocarcinoma growth andmetastasis in vivo.
Mice were generated that express the human LGALS3 gene inplace of the mouse gene (Hu-Gal-3-KI). Western blot analysisconfirmed expression of only human galectin-3 in mouse liverlysates from Hu-Gal-3-KI mice (Fig. 3E). LLC1 tumor growth wasinhibited by GB1107 in Hu-Gal-3-KI mice when administrationwas delayed until day 5 after inoculation (Fig. 3F).
Galectin-3 inhibitor blocks LLC1-induced alternativemacrophage activation
Given the altered M1:M2 TAM ratio in LLC1 tumors fromgalectin-3�/� mice and inhibitor-treated mice, we next deter-mined the role of LLC1-derived galectin-3 on macrophagepolarization. Conditioned media from LLC1 cells in vitroincreased IL4-stimulated arginase activity in BM-derivedmacrophages (BMDM) and increased gene expression of argi-nase-1 and fizz1 (Supplementary Fig. S5A and S5B). Thisincrease was inhibited by GB1107, suggesting galectin-3 secret-ed by LLC1 cells induces macrophages to adopt an alternativeM2-like phenotype (Supplementary Fig. S5A and S5B). GB1107did not affect LPS-induced Nos2 expression or nitric oxide(NO) production by BMDMs (Supplementary Fig. S5A andS5C). Although our data show TAMs to be a vital determinantof tumor growth in vivo, treatment of LLC1 cells with inhibitorin vitro also impacted on cell migration and proliferation albeitat higher concentrations (Supplementary Fig. S5D–S5F), sug-gesting some direct effect on galectin-3-mediated tumor cellexpansion and migration.
Galectin-3 depletion reduces M2-like macrophages andenhances infiltration of activated CD8 T cells
TAMs can contribute to T-cell immunosuppression (4). Inparticular, M2-like macrophages secrete more galectin-3 (34) and
Novel Galectin-3 Antagonist Inhibits Lung Cancer Progression
www.aacrjournals.org Cancer Res; 79(7) April 1, 2019 1485
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

Figure 3.
Galectin-3 inhibitor GB1107 inhibits lung adenocarcinoma growth andmetastasis in vivo.A, Female CD-1 nude mice received two subcutaneous injections of3� 106 A549 cells in a 1:1 ratio of Matrigel and serum-free media. Tumors grew to an average of 166 mm3 before commencement of single daily dosing of vehicle(n¼ 6) or 10mg/kg GB1107 (n¼ 6) from day 18. A, Tumor volumes and weights, along with representative images are shown. B, C57Bl/6 mice were injectedsubcutaneously with LLC1 cells and orally dosed once daily with vehicle or 10 mg/kg GB1107 from day 1. Tumor volumes and weights are shown. C, RNAwasextracted from tumors from B and expression of various genes was evaluated by qPCR. D, To test the effect of GB1107 onmetastasis, mice were injected with1� 106 LLC1-luciferase cells via the tail vein, followed by daily oral gavage of vehicle (n¼ 10) or GB1107 (10mg/kg; n¼ 10) from day 1. On day 7, tumor burden inwhole lungs was determined by qPCR using luciferase-specific primers. E,Western blot analysis confirming expression of human galectin-3 in liver lysates fromhu-Gal-3-KI mice. Two-tailed t tests were used to determine statistical significance. � , P < 0.05; �� , P < 0.01; ��� , P < 0.001 (compared with vehicle controls).F, Hu-Gal-3-KI mice bearing bilateral subcutaneous LLC1 tumors received vehicle or 10mg/kg GB1107 once daily orally from day 5. Tumor volumes and weightson day 14 are shown. Results from E represent the mean� SEM of two independent experiments of n¼ 6.
Vuong et al.
Cancer Res; 79(7) April 1, 2019 Cancer Research1486
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

galectin-3 directly impedes T-cell infiltration and activation(24–27, 35). We therefore investigated whether galectin-3–dependent M2 polarization is also associated with changes inT-cell infiltration and activation in vivo. Flow cytometric anal-ysis of tumor digests from hu-Gal-3-KI mice treated withGB1107 showed no increase in macrophage infiltration, butshowed a decrease in macrophage CD206 expression indicativeof reduced M2 TAMs (Supplementary Fig. S6A; Fig. 4A). Sim-ilarly, although there was no significant change in the totalnumber of CD3þ T cells, GB1107 caused an increase in CD8þ
but not CD4þ T cells within tumors (Fig. 4A). This pattern ofimmune infiltration was also observed in tumor digests fromgalectin-3�/� mice compared with WT C57Bl/6 mice (Fig. 4B).Moreover, in galectin-3�/� tumors, infiltrating CD8 T cells butnot CD4þ T cells displayed trends toward increased PD-1 andIFNg expression, together suggesting that galectin-3 depletionmay reprogram the tumor microenvironment to favor proin-flammatory M1-like macrophages and enhance cytotoxic CD8T-cell infiltration and activation. Galectin-3�/� mouse tumorsdisplayed no overall changes in total CD45þ cells, neutrophils,monocytes, or dendritic cells (DC) compared with WT tumors(Supplementary Fig. S6B).
To assess whether other systemic changes in galectin-3–deplet-ed mice may also influence CD8 T-cell activation, we assessedmyeloid populations within the BM of WT and galectin-3�/�
mice. We observed no differences in total CD11bþ cells, neutro-phils, or monocytes including inflammatory Ly6Chi monocytes(Supplementary Fig. S7). There were however increased DCs ingalectin-3�/� compared with WT mice, suggestive of anotherindirect mechanism by which galectin-3 may enhance antitumorT-cell priming (26).
Galectin-3 inhibitor potentiates the antitumor effects of PD-L1blockade
We next examined the effect of galectin-3 inhibition incombination with immune checkpoint inhibition. In thisstudy, GB1107 treatment was delayed until day 6 postimplan-tation. Delayed administration of GB1107 alone did not reducetumor burden and administration of an anti-PD-L1 antibodyadministered twice weekly intraperitoneally from day 6 alsohad no impact on tumor growth. However, a combination ofGB1107 and anti-PD-L1 antibody treatment significantlypotentiated the effect of the single agents (49.5% and 51.4%reduced tumor volumes and weights respectively comparedwith untreated controls; Fig. 5A and B). The reduced tumorgrowth in the combination group was associated with anincrease in PD-1þ CD8þ T cells (Fig. 5C; SupplementaryFig. S8A–S8B) and reduced CD206 expression in macrophages(Fig. 5C). This was combined with a significant decrease intumoral aSMA (Supplementary Fig. S8C) and an increasein expression of T-cell cytotoxic mediators (IFNg perforin-1,granzyme B, and fas ligand; Fig. 5D) and a 10.3% increase(P ¼ 0.005) in the apoptosis marker cleaved caspase-3(Fig. 5E and F) but was not associated with changes in tumorcell proliferation as determined by Ki-67 IHC staining (Sup-plementary Fig. S8D and S8E). There was a correspondingreduction in galectin-3 protein levels within GB1107-treatedtumors (Supplementary Fig. S8F). Together our data suggestthat combination therapy with galectin-3 inhibitor GB1107and PD-L1 blocking antibody promotes tumor cell apoptosisand cytotoxic CD8 T-cell activation.
DiscussionIn this study we show that galectin-3�/� mice do not support
LLC1 tumor growth and deleting galectin-3 in BM-derived cellsrecruited to engrafted mouse lung adenocarcinomas inhibitstumor growth and spread despite high expression of galectin-3in the tumor cells. Furthermore, macrophage depletion reducesmonocyte recruitment and LLC1 tumor growth, confirming thatdepletion of macrophages with liposomal clodronate inhibitsLLC1 tumor growth (36). Although the spleen can be an impor-tant source of TAMs in a KRAS and P53-driven model of lungadenocarcinoma (37), the BM compartment has been shown tobe the major source of TAMs in the LLC1 model (38). Therefore,we sought to restore galectin-3 in tumormacrophages in galectin-3�/�mice by BM transplant with WT galectin-3-positive BM cells.This results in an increase in tumor growth similar to that observedin WT mice. Our previous work has shown that galectin-3 is animportant regulator ofmacrophage function, promoting an "M2"phenotype (28).Our data show thatmacrophages in tumors fromgalectin-3�/� mice or mice treated with GB1107 have reducedCD206þ M2-like macrophages and we observe reduced M2-promoting cytokine transcripts and elevated IFNg expressionwithin galectin-3�/� tumors. In addition, conditioned mediafrom LLC1 cells increases alternative activation of macrophagesin vitro and this can be blocked by coculture with GB1107. Thisdemonstrates that galectin-3 contributes to the M2 immunosup-pressive function of TAMs.
TAMs promote many important features of tumor progressionincluding angiogenesis, tumor cell invasion, motility, and metas-tasis and can also suppress T-cell responses (4). These data showthat galectin-3–expressingmacrophages are recruited to the tumorsite, develop an M2 phenotype and induce downregulation ofCD8þ CTL functions. Galectin-3 has been shown to induce T-celltolerance resulting in T-cell anergy, through various mechanismsincluding inhibiting CD8 and TCR clustering (39), destabilizingthe immune synapse and promoting internalization of TCR andCD3z chains (40). It can also restrict membrane movement andTCR-associated signaling functions of CD45 (41) and inhibit LFA-1 recruitment thus disrupting proper secretory synapse formationand secretion of IFNg (27).
Galectin-3 may also suppress CTL effector function by bindingto LAG-3, a negative regulatory checkpoint, on CD8þ T cells (26)and by inducing apoptosis of CTLs (25) and impairs the antitu-mor functions of natural killer (NK) cells (42). CTLs activated invitro show an alteration in the N-glycome with longer and morebranched N-glycans resulting in the expression of surface glyco-proteins that exhibit high galectin-3 binding (43). The highconcentration of galectin-3 found in tumor microenvironmentscould potentially explain the loss of CTL functions throughreduced motility and signaling functions of surface molecules.
Galectin-3�/�mice have also been shown to have an increase inlymph node plasmacytoid DCs (pDC) compared with WT mice,which are superior in activating CD8þ CTLs compared withconventional DC (26). In addition, galectin-3 knockdown inmonocyte-derived DCs increases the proliferation and IFNg pro-duction from antigen-stimulated CD4þ T cells (44). Our profilingof BM fromWT and gal-3 KOmice showed an increase in CD45þ/MHC-IIþ/CD11b- DCs in BM of gal-3 KO mice compared withWT. Although our study did not distinguish DC subsets, togetherthe data suggest that galectin-3 may indirectly regulate CD8function by promoting DC functions. This requires further study.
Novel Galectin-3 Antagonist Inhibits Lung Cancer Progression
www.aacrjournals.org Cancer Res; 79(7) April 1, 2019 1487
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244
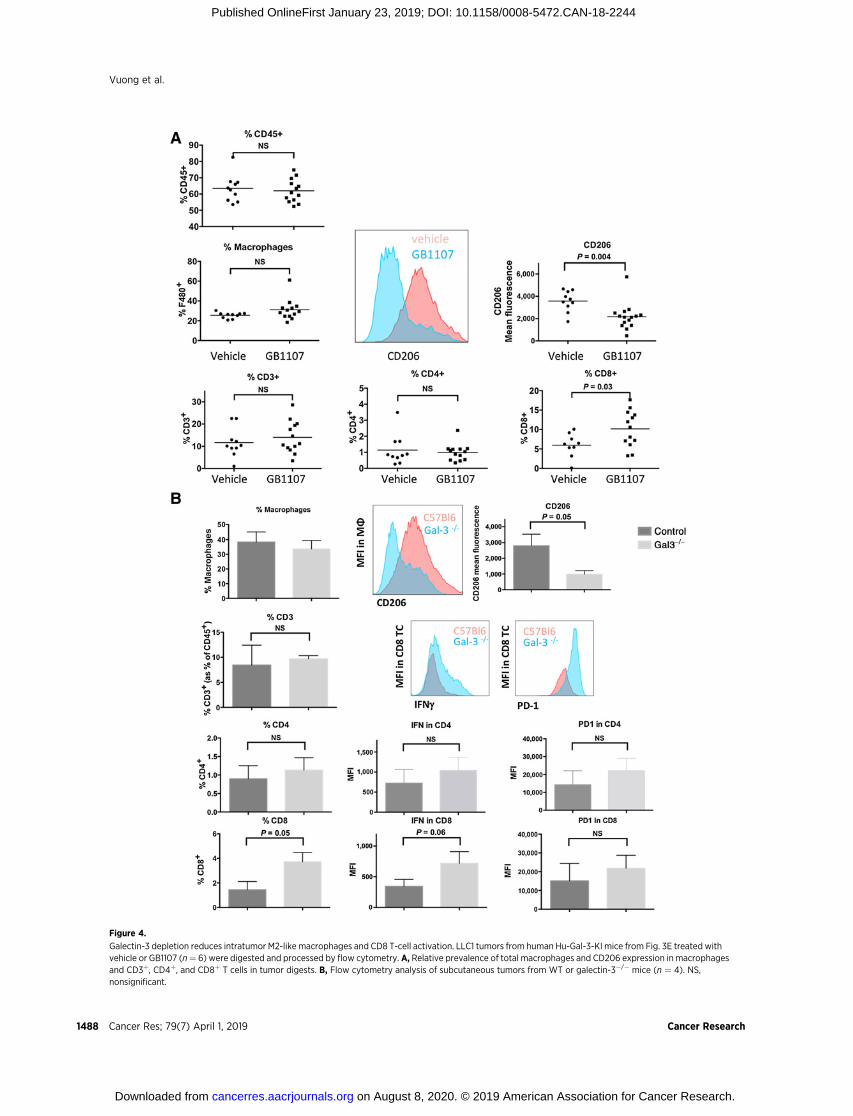
Figure 4.
Galectin-3 depletion reduces intratumor M2-like macrophages and CD8 T-cell activation. LLC1 tumors from human Hu-Gal-3-KI mice from Fig. 3E treated withvehicle or GB1107 (n¼ 6) were digested and processed by flow cytometry. A, Relative prevalence of total macrophages and CD206 expression in macrophagesand CD3þ, CD4þ, and CD8þ T cells in tumor digests. B, Flow cytometry analysis of subcutaneous tumors from WT or galectin-3�/� mice (n ¼ 4). NS,nonsignificant.
Vuong et al.
Cancer Res; 79(7) April 1, 2019 Cancer Research1488
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

Figure 5.
Combination therapy with galectin-3 inhibitor GB1107 and PD-L1 blocking antibody promotes tumor cell apoptosis and cytotoxic CD8 T-cell activation. On day 6,mice bearing subcutaneous LLC1 tumors were randomized into four groups and received either no treatment (n¼ 8), a-PD-L1 neutralizing antibody (200 mgtwice weekly i.p.; n¼ 8), or GB1107 (10 mg/kg once daily orally; n¼ 8) or a-PD-L1 plus GB1107 (n¼ 8). Tumor volumes (A) and weights (B) on day 16 are shown.C, Tumor-infiltrating immune populations were analyzed by flow cytometry (n¼ 4). D, Total RNAwas extracted from tumors in each group and qPCR was usedto assess relative gene expression. E and F, IHC staining of cleaved caspase-3 was quantified as described in Materials and Methods. Scale bars, 100 mm. Datarepresent mean� SE from a single (A, B, and D–F) or two (C) independent experiments. Two-way ANOVAwith Tukey's post hoc test was used to test fordifferences in tumor volume and cleaved caspase-3 IHC scores. One-way ANOVA and Fisher LSD test were used to compare tumor weights, and qPCR and flowcytometry data were compared using two-tailed t tests. � , P < 0.05; �� , P < 0.01; ��� , P < 0.001. ns, nonsignificant.
Novel Galectin-3 Antagonist Inhibits Lung Cancer Progression
www.aacrjournals.org Cancer Res; 79(7) April 1, 2019 1489
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

We show that treatment with GB1107 alone from the outsetinhibits LLC1growth anddelayed treatment inhibits LLC1growthin human galectin-3–expressing mice. This reflects the increasedaffinity this inhibitor has on human versus mouse galectin-3. Inaddition, the galectin-3 inhibitor significantly potentiates theeffect of immune checkpoint blockade with an anti-PD-L1 block-ing antibody. It is believed that the limited patient responses tocheckpoint inhibition is attributable to the lack of T-cell infiltra-tion in so-called "cold" tumors (2). Gordon-Alonso and collea-gues, show that galectin-3 binds to the extracellular matrix and toglycosylated IFNg , preventing release of IFNg-induced CXCL9,which acts as a T-cell chemo-attractant (35). Consistent with this,GB1107 both alone or in combination with anti-PD-L1 increasesthe number of tumor infiltrating CD8 CTLs. Therefore, galectin-3inhibitionmight provide the criticalmeans to turn a "cold" tumor"hot," and thus responsive to immune checkpoint intervention.Furthermore, CD8þ CTLs within the combination drug-treatedtumors are more activated (express more surface PD-1), and thecytokine environment favors tumor rejection with increasedexpression of cytotoxic (IFNg , perforin-1, and granzyme B) andapoptotic (fas ligand) genes with increased caspase activation.
Reduced galectin-3 expression within tumor cells has beenshown to reduce tumor growth in many cancers (reviewed inref. 45), suggested to be due to the anti-apoptotic effect ofcytoplasmic galectin-3 binding to K-RAS and engaging anti-apo-ptotic pathways via its NWGRmotif (20). However, we show thatknockdown of galectin-3 in tumor cells with shRNA had only apartial effect on tumor growth in vitro but had no significant effecton LLC1 growth in vivo. We also show that treatment with thegalectin-3 inhibitor alone could inhibit human adenocarcinomagrowth in CD-1 nudemice, which lack a T-cell response butwhichdisplay innate immunity. These suggest that either tumor derivedormacrophage-derived galectin-3 can impact on tumor growth inthis model, independent of the T-cell–mediated effects.
In conclusion, our results demonstrate that galectin-3 inhibi-tion leads to a reduction in M2-like TAMs and increased infiltra-tion and activity of CD8þ CTLs within LLC1 tumors resulting inreduced tumor growth and metastasis. Several studies have usedother approaches to inhibit galectin-3 in cancer including peptideinhibitors (46), lactulose amines (47), a glycopeptide isolatedfrom cod (48) and large complex plant-derived polysaccharidesincluding modified citrus pectin (49), GCS-100 (39), and galac-tomannans such as GM-CT-01 (50). GCS-100 is currently beingdeveloped for chronic lymphoid leukemia and multiple myelo-ma (51). However recent evidence suggests that these complexcarbohydrates do not act as inhibitors of the canonical carbohy-drate-binding site of galectin-3 and their physiologic effects maybe due to unrelated actions (52). We show using a specific andhigh affinity inhibitor of the galectin-3 carbohydrate site thatpharmacologic inhibition of galectin-3 inhibits lung adenocarci-
noma growth and potentiates the effect of immune checkpointinhibitors. Therefore, galectin-3 has a strong regulatory effect oncancer-related inflammation and could present a key target in themanagement of lung, and potentially other galectin-3–drivencarcinomas, in combination with immune checkpoint blockade.
Disclosure of Potential Conflicts of InterestL. Vuong reports receiving other commercial research support from Galecto
Biotech. N.C. Henderson is a consultant/advisory board member of GalectoBiotech. U.J. Nilsson has ownership interest (including stock, patents, etc.) inGalecto Biotech AB and is a consultant/advisory board member of GalectoBiotech AB. H. Leffler reports receiving a commercial research grant fromGalecto Biotech AB, has ownership interest (including stock, patents, etc.) inGalecto Biotech AB, and is a consultant/advisory board member of GalectoBiotechAB.H. Schambye is aCEOatGalecto Biotech andhas ownership interest(including stock, patents, etc.) in Galecto Biotech. T. Sethi is a Chief PhysicianScientist at AstraZeneca and has ownership interest (including stock, patents,etc.) in Galecto Biotech. No potential conflicts of interest were disclosed by theother authors.
Authors' ContributionsConception and design: L. Vuong, E. Kouverianou, S.E.M. Howie,F.R. Zetterberg, U.J. Nilsson, H. Leffler, H, Schambye, T. Sethi, A.C. MacKinnonDevelopment of methodology: L. Vuong, E. Kouverianou, U.J. Nilsson,S. Tantawi, A.C. MacKinnonAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): L. Vuong, E. Kouverianou, C.M. Rooney,B.J. McHugh, C.D. Gregory, A. Pedersen, L. Gravelle, A.C. MacKinnonAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): L. Vuong, E. Kouverianou, B.J. McHugh,S.E.M. Howie, S.J. Forbes, F.R. Zetterberg, U.J. Nilsson, H. Leffler, P. Ford,H, Schambye, A.C. MacKinnonWriting, review, and/or revision of themanuscript: L. Vuong, E. Kouverianou,B.J. McHugh, S.E.M. Howie, C.D. Gregory, N.C. Henderson, F.R. Zetterberg,U.J. Nilsson, H. Leffler, P. Ford, A. Pedersen, H, Schambye, T. Sethi,A.C. MacKinnonAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): S.J. ForbesStudy supervision: T. SethiOther (design and synthesis of GB1107): F.R. Zetterberg
AcknowledgmentsWe thank Frank McCaughan for helpful discussions and kind provision of
reagents, as well as the Biomedical Research Council Flow Cytometry Core(King's College) and the University of Edinburgh Queen's Medical ResearchInstitute's Flow Cytometry facility for flow sorting and flow cytometric analysis.The work presented was funded by Galecto Biotech, a King's Health School'sscholarship to L. Vuong, and a Norman Salvesen Emphysema Research grant toE. Kouverianou.
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received July 22, 2018; revised November 16, 2018; accepted January 17,2019; published first January 23, 2019.
References1. QuaresmaM,ColemanMP, Rachet B. 40-year trends in an index of survival
for all cancers combined and survival adjusted for age and sex for eachcancer in England andWales, 1971-2011: a population-based study. Lancet2015;385:1206–18.
2. Sharma P, Hu-Lieskovan S, Wargo JA, Ribas A. Primary, Adaptive, andAcquired Resistance to Cancer Immunotherapy. Cell 2017;168:707–23.
3. Yuan A, Hsiao YJ, Chen HY, Chen HW, Ho CC, Chen YY, et al. Oppositeeffects of M1 and M2 macrophage subtypes on lung cancer progression.Sci Rep 2015;5:14273. doi: 10.1038/srep14273.
4. Noy R, Pollard JW. Tumor-associated macrophages: from mechanisms totherapy. Immunity 2014;41:49–61.
5. Gordon SR, Maute RL, Dulken BW, Hutter G, George BM, McCracken MN,et al. PD-1 expression by tumour-associated macrophages inhibits phago-cytosis and tumour immunity. Nature 2017;545:495–9.
6. Arlauckas SP, Garris CS, Kohler RH, Kitaoka M, Cuccarese MF,Yang KS, et al. In vivo imaging reveals a tumor-associatedmacrophage-mediated resistance pathway in anti-PD-1 therapy.Sci Transl Med 2017;9. doi: 10.1126/scitranslmed.aal3604.
Vuong et al.
Cancer Res; 79(7) April 1, 2019 Cancer Research1490
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

7. Neubert NJ, Schmittnaegel M, Bordry N, Nassiri S, Wald N, Martignier C,et al. T cell-induced CSF1 promotesmelanoma resistance to PD1 blockade.Sci Transl Med 2018;10.
8. Peranzoni E, Lemoine J, Vimeux L, Feuillet V, Barrin S, Kantari-Mimoun C,et al. Macrophages impede CD8 T cells from reaching tumor cells and limitthe efficacy of anti-PD-1 treatment. Proc Natl Acad Sci 2018;115:E4041–E4050.
9. Hirabayashi J, Hashidate T, Arata Y, Nishi N, Nakamura T, Hirashima M,et al. Oligosaccharide specificity of galectins: a search by frontal affinitychromatography. Biochim Biophys Acta 2002;1572:232–54.
10. Chen HY, Liu FT, Yang RY. Roles of galectin-3 in immune responses.Arch Immunol Ther Exp (Warsz) 2005;53:497–504.
11. Liu FT, Hsu DK, Zuberi RI, Kuwabara I, Chi EY, Henderson WR Jr.Expression and function of galectin-3, a beta-galactoside-binding lectin,in human monocytes and macrophages. Am J Pathol 1995;147:1016–28.
12. Joo HG, Goedegebuure PS, Sadanaga N, Nagoshi M, von Bernstorff W,Eberlein TJ. Expression and function of galectin-3, a beta-galactoside-binding protein in activated T lymphocytes. J Leukoc Biol 2001;69:555–64.
13. Dube-Delarosbil C, St-Pierre Y. The emerging role of galectins in high-fatality cancers. Cell Mol Life Sci 2018;75:1215–26.
14. Mathieu A, Saal I, Vuckovic A, Ransy V, Vereerstraten P, Kaltner H, et al.Nuclear galectin-3 expression is an independent predictive factor of recur-rence for adenocarcinoma and squamous cell carcinoma of the lung.Mod Pathol 2005;18:1264–71.
15. Puglisi F,Minisini AM, Barbone F, IntersimoneD, Aprile G, PuppinC, et al.Galectin-3 expression in non-small cell lung carcinoma. Cancer Lett 2004;212:233–9.
16. Yu LG. Circulating galectin-3 in the bloodstream: an emerging promoter ofcancer metastasis. World J Gastrointest Oncol 2010;2:177–80.
17. Reticker-Flynn NE, Malta DF, Winslow MM, Lamar JM, Xu MJ,Underhill GH, et al. A combinatorial extracellular matrix platformidentifies cell-extracellular matrix interactions that correlate withmetastasis. Nat Commun 2012;3:1122. doi: 10.1038/ncomms2128.
18. Wu F, Hu N, Li Y, Bian B, Xu G, Zheng Y. Galectin-3 genetic variants areassociated with platinum-based chemotherapy response and prognosis inpatients with NSCLC. Cell Oncol 2012;35:175–80.
19. HonjoY,Nangia-Makker P, InoharaH, RazA.Down-regulation of galectin-3 suppresses tumorigenicity of human breast carcinoma cells. Clin CancerRes 2001;7:661–8.
20. Akahani S, Nangia-Makker P, Inohara H, Kim HR, Raz A. Galectin-3: anovel antiapoptotic molecule with a functional BH1 (NWGR) domain ofBcl-2 family. Cancer Res 1997;57:5272–6.
21. Radosavljevic G, Jovanovic I, Majstorovic I, Mitrovic M, Lisnic VJ,Arsenijevic N, et al. Deletion of galectin-3 in the host attenuatesmetastasis of murine melanoma by modulating tumor adhesion andNK cell activity. Clin Exp Metast 2011;28:451–62.
22. Chung LY, Tang SJ, Wu YC, Sun GH, Liu HY, Sun KH. Galectin-3 augmentstumor initiating property and tumorigenicity of lung cancer throughinteraction with beta-catenin. Oncotarget 2015;6:4936–52.
23. Markowska AI, Jefferies KC, Panjwani N. Galectin-3 proteinmodulates cellsurface expression and activation of vascular endothelial growth factorreceptor 2 in human endothelial cells. J Biol Chem 2011;286:29913–21.
24. Ruvolo PP. Galectin 3 as a guardian of the tumor microenvironment.Biochim Biophys Acta 2016;1863:427–37.
25. Peng W, Wang HY, Miyahara Y, Peng G, Wang RF. Tumor-associatedgalectin-3 modulates the function of tumor-reactive T cells. Cancer Res2008;68:7228–36.
26. Kouo T, Huang L, Pucsek AB, Cao M, Solt S, Armstrong T, et al. Galectin-3shapes antitumor immune responses by suppressing cd8þ t cells via lag-3and inhibiting expansion of plasmacytoid dendritic cells. Cancer ImmunolRes 2015;3:412–23.
27. Petit AE,DemotteN, Scheid B,WildmannC, Bigirimana R,Gordon-AlonsoM, et al. A major secretory defect of tumour-infiltrating T lymphocytes dueto galectin impairing LFA-1-mediated synapse completion. Nat Commun2016;7:12242.
28. MacKinnon AC, Farnworth SL, Hodkinson PS, Henderson NC, AtkinsonKM, Leffler H, et al. Regulation of alternative macrophage activation bygalectin-3. J Immunol 2008;180:2650–8.
29. Voss JJ, Ford CA, Petrova S, Melville L, Paterson M, Pound JD, et al.Modulation of macrophage antitumor potential by apoptotic lymphomacells. Cell Death Differ. 2017.
30. HsuDK, Yang RY, Pan Z, Yu L, SalomonDR, Fung-LeungWP, et al. Targeteddisruption of the galectin-3 gene results in attenuated peritoneal inflam-matory responses. Am J Pathol 2000;156:1073–83.
31. Cailhier JF, Partolina M, Vuthoori S, Wu S, Ko K, Watson S, et al.Conditional macrophage ablation demonstrates that resident macro-phages initiate acute peritoneal inflammation. J Immunol 2005;174:2336–42.
32. Zetterberg FR, Peterson K, Johnsson RE, Brimert T, Hakansson M, LoganDT, et al. Monosaccharide derivatives with low-nanomolar lectin affinityand high selectivity based on combined fluorine-amide, phenyl-arginine,sulfur-pi, and halogen bond interactions. ChemMedChem 2018;13:133–7.
33. Duffield JS, Forbes SJ, Constandinou CM, Clay S, Partolina M,Vuthoori S, et al. Selective depletion of macrophages reveals distinct,opposing roles during liver injury and repair. J Clin Invest 2005;115:56–65.
34. Novak R, Dabelic S, Dumic J. Galectin-1 and galectin-3 expression profilesin classically and alternatively activated human macrophages.Biochim Biophys Acta 2012;1820:1383–90.
35. Gordon-Alonso M, Hirsch T, Wildmann C, van der Bruggen P. Galectin-3captures interferon-gamma in the tumor matrix reducing chemokinegradient production and T-cell tumor infiltration. Nat Commun 2017;8:793. doi: 10.1038/s41467-017-00925-6.
36. Schmall A, Al-Tamari HM, Herold S, Kampschulte M, Weigert A, Wietel-mannA, et al.Macrophage and cancer cell cross-talk viaCCR2 andCX3CR1is a fundamental mechanism driving lung cancer. Am J Respir Crit CareMed 2015;191:437–47.
37. Cortez-Retamozo V, Etzrodt M, Newton A, Rauch PJ, Chudnovskiy A,Berger C, et al. Origins of tumor-associated macrophages and neutrophils.PNAS 2012;109:2491–6.
38. Shand FH,Ueha S,OtsujiM,Koid SS, Shichino S, Tsukui T, et al. Tracking ofintertissue migration reveals the origins of tumor-infiltrating monocytes.PNAS 2014;111:7771–6.
39. DemotteN,WieersG, VanDer Smissen P,MoserM, SchmidtC, ThielemansK, et al. A galectin-3 ligand corrects the impaired function of human CD4and CD8 tumor-infiltrating lymphocytes and favors tumor rejection inmice. Cancer Res 2010;70:7476–88.
40. Chen HY, Fermin A, Vardhana S, Weng IC, Lo KF, Chang EY,et al. Galectin-3 negatively regulates TCR-mediated CD4þ T-cellactivation at the immunological synapse. PNAS 2009;106:14496–501.
41. Stillman BN,HsuDK, PangM, BrewerCF, JohnsonP, Liu FT, et al. Galectin-3 and galectin-1 binddistinct cell surface glycoprotein receptors to induce Tcell death. J Immunol 2006;176:778–89.
42. Rabinovich GA, Toscano MA. Turning `sweet' on immunity: galectin-glycan interactions in immune tolerance and inflammation. Nat RevImmunol 2009;9:338–52.
43. Antonopoulos A, Demotte N, Stroobant V, Haslam SM, van der Bruggen P,Dell A. Loss of effector function of human cytolytic T lymphocytes isaccompanied by major alterations in N- and O-glycosylation. J Biol Chem2012;287:11240–51.
44. Mobergslien A, SioudM.Galectin-1 and -3 gene silencing in immature andmature dendritic cells enhances T cell activation and interferon-gammaproduction. J Leukoc Biol 2012;91:461–7.
45. Wang L, Guo XL. Molecular regulation of galectin-3 expression andtherapeutic implication in cancer progression. Biomed Pharmacother2016;78:165–71.
46. Zou J, Glinsky VV, Landon LA, Matthews L, Deutscher SL. Peptidesspecific to the galectin-3 carbohydrate recognition domain inhibitmetastasis-associated cancer cell adhesion. Carcinogenesis 2005;26:309–18.
47. Glinsky VV, KiriakovaG,GlinskiiOV,Mossine VV,Mawhinney TP, Turk JR,et al. Synthetic galectin-3 inhibitor increases metastatic cancer cell sensi-tivity to taxol-induced apoptosis in vitro and in vivo. Neoplasia 2009;11:901–9.
48. Guha P, Kaptan E, Bandyopadhyaya G, Kaczanowska S, Davila E, Thomp-son K, et al. Cod glycopeptide with picomolar affinity to galectin-3
Novel Galectin-3 Antagonist Inhibits Lung Cancer Progression
www.aacrjournals.org Cancer Res; 79(7) April 1, 2019 1491
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

suppresses T-cell apoptosis and prostate cancer metastasis. PNAS 2013;110:5052–7.
49. Pienta KJ, Naik H, Akhtar A, Yamazaki K, Replogle TS, Lehr J, et al.Inhibition of spontaneous metastasis in a rat prostate cancer model byoral administration of modified citrus pectin. J Natl Cancer Inst 1995;87:348–53.
50. Demotte N, Bigirimana R, Wieers G, Stroobant V, Squifflet JL, Carrasco J,et al. A short treatment with galactomannan GM-CT-01 corrects the
functions of freshly isolated human tumor-infiltrating lymphocytes.Clin Cancer Res 2014;20:1823–33.
51. Harazono Y, Nakajima K, Raz A. Why anti-Bcl-2 clinical trials fail: asolution. Cancer Metastasis Rev 2014;33:285–94.
52. Stegmayr J, Lepur A, Kahl-Knutson B, Aguilar-Moncayo M, Klyosov AA,Field RA, et al. Low or no inhibitory potency of the canonical galectincarbohydrate binding site by pectins and galactomannans. J Biol Chem2016;291:13318–34.
Cancer Res; 79(7) April 1, 2019 Cancer Research1492
Vuong et al.
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244

2019;79:1480-1492. Published OnlineFirst January 23, 2019.Cancer Res Lynda Vuong, Eleni Kouverianou, Claire M. Rooney, et al. BlockadeAdenocarcinoma Growth and Augments Response to PD-L1 An Orally Active Galectin-3 Antagonist Inhibits Lung
Updated version
10.1158/0008-5472.CAN-18-2244doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2019/01/23/0008-5472.CAN-18-2244.DC1
Access the most recent supplemental material at:
Overview
Visual
http://cancerres.aacrjournals.org/content/79/7/1480/F1.large.jpgA diagrammatic summary of the major findings and biological implications:
Cited articles
http://cancerres.aacrjournals.org/content/79/7/1480.full#ref-list-1
This article cites 49 articles, 17 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/79/7/1480To request permission to re-use all or part of this article, use this link
on August 8, 2020. © 2019 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 23, 2019; DOI: 10.1158/0008-5472.CAN-18-2244
Related Documents











