Journal of Biotechnology 91 (2001) 257 – 268 An interdisciplinary research strategy to improve symbiotic nitrogen fixation and yield of common bean (Phaseolus ulgaris ) in salinised areas of the Mediterranean basin J.J. Drevon a, *, C. Abdelly b , N. Amarger c , E.A. Aouani b , J. Aurag d , H. Gherbi a , M. Jebara b , C. Lluch e , H. Payre a , O. Schump a , M. Soussi c,e , B. Sifi f , M. Trabelsi g a INRA, Place Viala, 34000 Montpellier, France b INRST, BP 95, 2050 Hammam-Lif, Tunisia c INRA, BV 1540, 21 034 Dijon, France d Uniersity of Rabat, Rabat, Morocco e Uniersity of Granada, Granada, Spain f INRAT, Ariana, Tunisia g ESA, Mateur 7030, Tunisia Received 6 November 2000; received in revised form 6 April 2001; accepted 3 May 2001 Abstract The main findings of a cooperative research group of agronomists, plant breeders, microbiologists, physiologists and molecularists to improve the symbiotic nitrogen fixation (SNF) and N 2 -dependent yield of common bean under moderate salinity in the Mediterranean basin are summarised. Agronomic surveys in reference production areas show large spatial and temporal variations in plant nodulation and growth, and in efficiency of utilisation of the rhizobial symbiosis. The latter was associated with a large rhizobial diversity, including new bean nodulating species. Macrosymbiont diversity in SNF and adaptation to NaCl was found. However, contrasts between plant genotypes could be altered by specific interactions with some native rhizobia. Therefore, variations in soil rhizobial population, in addition to agronomic practices and environmental constraints, may have contributed to erratic results observed in field inoculations. At the mechanistic level, nodule C and N metabolisms, and abcissic acid content, were related to SNF potential and tolerance to NaCl. Their relation with nodule conductance to O 2 diffusion was addressed by in situ hybridisation of candidate carbonic anhydrase and aquaporin genes in nodule cortex. The limits and prospects of the cooperative strategy are discussed. © 2001 Published by Elsevier Science B.V. www.elsevier.com/locate/jbiotec Abbreiations: ABA, abcissic acid; AI, alcaline invertase; ARA, acetylene reducing activity; DAS, days after sowing; GS, glutamine synthetase; GOGAT, glutamate synthase; ISH, in situ hybridisation; MDH, malate dehydrogenase; PCR, polymerase chain reaction; PEPC, phosphoenolpyruvate carboxylase; SNF, symbiotic nitrogen fixation; SS, sucrose synthase; YEM, yeast extract mannitol. * Corresponding author. Tel.: +33-4-67797440. E-mail address: [email protected] (J.J. Drevon). 0168-1656/01/$ - see front matter © 2001 Published by Elsevier Science B.V. PII:S0168-1656(01)00339-X

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Biotechnology 91 (2001) 257–268
An interdisciplinary research strategy to improve symbioticnitrogen fixation and yield of common bean (Phaseolus�ulgaris) in salinised areas of the Mediterranean basin
J.J. Drevon a,*, C. Abdelly b, N. Amarger c, E.A. Aouani b, J. Aurag d,H. Gherbi a, M. Jebara b, C. Lluch e, H. Payre a, O. Schump a, M. Soussi c,e,
B. Sifi f, M. Trabelsi g
a INRA, Place Viala, 34000 Montpellier, Franceb INRST, BP 95, 2050 Hammam-Lif, Tunisia
c INRA, BV 1540, 21 034 Dijon, Franced Uni�ersity of Rabat, Rabat, Morocco
e Uni�ersity of Granada, Granada, Spainf INRAT, Ariana, Tunisia
g ESA, Mateur 7030, Tunisia
Received 6 November 2000; received in revised form 6 April 2001; accepted 3 May 2001
Abstract
The main findings of a cooperative research group of agronomists, plant breeders, microbiologists, physiologistsand molecularists to improve the symbiotic nitrogen fixation (SNF) and N2-dependent yield of common bean undermoderate salinity in the Mediterranean basin are summarised. Agronomic surveys in reference production areas showlarge spatial and temporal variations in plant nodulation and growth, and in efficiency of utilisation of the rhizobialsymbiosis. The latter was associated with a large rhizobial diversity, including new bean nodulating species.Macrosymbiont diversity in SNF and adaptation to NaCl was found. However, contrasts between plant genotypescould be altered by specific interactions with some native rhizobia. Therefore, variations in soil rhizobial population,in addition to agronomic practices and environmental constraints, may have contributed to erratic results observedin field inoculations. At the mechanistic level, nodule C and N metabolisms, and abcissic acid content, were relatedto SNF potential and tolerance to NaCl. Their relation with nodule conductance to O2 diffusion was addressed byin situ hybridisation of candidate carbonic anhydrase and aquaporin genes in nodule cortex. The limits and prospectsof the cooperative strategy are discussed. © 2001 Published by Elsevier Science B.V.
www.elsevier.com/locate/jbiotec
Abbre�iations: ABA, abcissic acid; AI, alcaline invertase; ARA, acetylene reducing activity; DAS, days after sowing; GS,glutamine synthetase; GOGAT, glutamate synthase; ISH, in situ hybridisation; MDH, malate dehydrogenase; PCR, polymerasechain reaction; PEPC, phosphoenolpyruvate carboxylase; SNF, symbiotic nitrogen fixation; SS, sucrose synthase; YEM, yeastextract mannitol.
* Corresponding author. Tel.: +33-4-67797440.E-mail address: [email protected] (J.J. Drevon).
0168-1656/01/$ - see front matter © 2001 Published by Elsevier Science B.V.
PII: S 0168 -1656 (01 )00339 -X

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268258
Keywords: Nitrogenase; Metabolism; Osmoregulation; Rhizobium; Root-nodule; Symbiosis
1. Introduction
Salinity affects more than 40% of soils in theMediterranean basin. Draining salinised soils orirrigating with high quality water from remotesources are extremely costly. Therefore,selecting plants tolerant to salinity is an alterna-tive strategy for a sustainable agriculture in thosemarginal lands. Legumes, in symbioses with rhi-zobia, are a priority area for research policies ofdeveloping countries because of their ability tosupport production of protein-rich seedsand fodder with fixed nitrogen, in addition totheir beneficial effect on productivity of cerealsand other crops in agricultural rotations. There-fore, a cooperative research project was designedwith the general objective of applying advances inmethodologies and basic knowledge on sym-biotic nitrogen fixation (SNF), which have beenspectacular in the last decade, to (i) select com-mon bean (Phaseolus �ulgaris), and native specificrhizobia for SNF tolerance to NaCl salinity, (ii)assess their yields in soils irrigated with watervarying in salinity in North Africa and SouthEurope, and (iii) progress in understanding bio-chemical mechanisms involved in SNF toleranceto salinity.
The project, namely FYSAME, was based onmultidisciplinary research including agronomists,microbiologists, plant breeders, physiologists andmolecular biologists from Algeria, France, Mo-rocco, Spain and Tunisia, with the followingtasks, respectively: to survey salinised zones forcollecting local cultivars and Rhizobium strains,and to assess selected symbioses through multilo-cal–pluriannual field trials; to screen macrosym-bionts for SNF tolerance to salinity, and toperform cross-inoculation trials for microsym-biont infraspecificity with and without salinity; todefine optimal procedures for the above screen-ings, and to investigate cellular and molecularSNF mechanisms associated with salt tolerance,through comparative studies of sensitive versustolerant symbioses; to characterise the genetic di-
versity of microsymbionts, and tag the geneticdeterminants of macrosymbiont tolerance for fur-ther investigation of genes involved in SNF toler-ance to salinity.
The objective of this paper is to revise themajor achievements of FYSAME, and addresssubsequent prospects and limits, with an attemptto emphasise what is new from interactions be-tween disciplines in this project.
2. Materials and methods
2.1. Nodulation sur�ey and field trials
The agronomic survey of legume nodulationwithin a production area addresses (i) whetherSNF can satisfy the N demand of a grain legumecrop in production fields, and (ii) what factorslimit the SNF in those fields where the yield of thegrain legume appears to be limited by a deficiencyin N nutrition.
The methodology consists in measuring nodula-tion and growth of the legume in survey stations,i.e. 10×20 m2 homogeneous plot, within a net-work of fields inside the production area. Sam-pling is performed at (i) early vegetative growth,with two to four fully expanded leaves, and (ii)during late flowering or early pod-filling, whenmaximal nodulation had been expressed. At thevegetative stage, a visual scoring of nodulationwas performed. At flowering stage, plants werecollected in four random sites of the station, i.e.two to five neighbouring plants per site dependingon variability in samples, by excavating 20 cmaround the root system. The plant was separatedimmediately from surrounding rhizospheric soil.Soil and plant samples were transferred in plasticbags and stored in a cold room until furtheranalysis. For each individual plant, the shoot wasseparated from the root at the cotyledonary node;the number of nodules was recorded; and thenodule and shoot dry weights were measured afterdrying at 80 °C. At maturity, yield components,and weight and humidity of grains per plant, were

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268 259
measured for two treatments, namely control andnitrogen fertilisation, i.e. 50 kg N as urea at vege-tative, flowering and pod-filling stages in a part ofthe survey station. Other cultural practices weresimilar to those in the field. Thus, a nodulationsurvey required the cooperation of producers ofthe area. It was conducted with a participativeapproach, including extensionists and agron-omists who have gathered knowledge of this areanot only for the expression of the grain legumeyield potential, but also for social relations in-volved in grain legume production.
Within the production area where the abovesurvey was performed, inoculation trials wereconducted with the following treatments:control without N fertilisation or inoculum; inoc-ulation without N fertilisation; inoculation with50 N fertilisation at sowing; inoculation with50 kg N at flowering; inoculation with 50 kg N atsowing and flowering; inoculation with 50 kg N atsowing, flowering and pod-filling. Inoculant waspoured as a suspension of 106 bacteria seed−1
before closing the sowing row. N fertilisation wasapplied as urea. For the other culturalpractises, the cultivation was conducted with localtechniques. The latter differed extensively amongcountries of the project, including variation insalinity of irrigation water. Elementary plots were3.6×6 m2, with six rows at a density of20 plants m−1, distributed in four blocksseparated by a 1 m alley to avoid rhizobial con-tamination. Simultaneously, tests were performedin a sample of 10 production fields chosen fromthe agronomic survey. The trial consisted of aband of six sowing rows in which one treatmentof the above trial was applied, for comparisonwith the local N treatment in the rest of his field.Sampling during growth and harvest were per-formed as described previously for the nodulationsurvey.
These agronomic tasks were performed in theLoukkos perimeter in Morocco, the Mejerda val-ley in Tunisia and the Lauragais corridor inFrance. The reference plants and rhizobia in thesetrials were local cultivars, namely Coco nain blanc(Graphopast, Tunis) in North Africa, Linex (Vil-morin) in France and Rhizobium tropici BCIAT899 (CIAT, Cali).
2.2. Isolation and identification of rhizobia
Rhizobial strains used in this study were iso-lated from root nodules from the above produc-tion areas or from plants grown on soil samplescollected from arid and semi-arid regions of Mo-rocco and Tunisia. Pure strains were usually culti-vated on YEM media (Vincent, 1970). They werestored at −20 °C in 50% YEM glycerol. Theinoculation of new culture media was made fromprecultures at the end of the exponential phase ofgrowth on the same media. The tolerance tosalinity of free-living rhizobia was determined onYEM agar plates containing from 0 to 5% (w/v)NaCl or CaCl2 concentrations or on YEM liquidmedium incubated in a gyratory shaker at200 rpm. Optical density was measured at 600 nmevery 3 h. The generation time was calculatedfrom the logarithmic phase of growth.
The genotypic characterisation was performedby restriction fragment length polymorphism ofthe 16S-rRNA genes amplified by polymerasechain reaction (PCR) according to Laguerre et al.(1994) with the restriction enzymes TaqI, HinfIand MspI.
2.3. Culture of symbiotic beans in glasshouse
Seeds of bean were sterilised in Ca hypochlor-ite, washed with sterile distilled water and germi-nated on 0.8% agar. Plant seedlings weretransferred to 0.5 l plastic pots filled with sandpreviously sterilised during 48 h at 180 °C. Threedays after sowing (DAS), the cotyledons weregently passed through holes fitted in lids of pots,and 1 ml bacterial suspension containing approxi-mately 109 cells was applied on the soil surfacenext to the plant hypocotyle.
For hydroaeroponic culture, seedlings werepassed carefully through the pierced rubber stop-per of a 1 l serum bottle and fixed with cottonfitted around the hypocotyle. The nutritive solu-tion (Vadez et al., 1996) was aerated by com-pressed air at a flow of 400 ml min−1 l−1
solution. As salt treatment, 25 mM NaCl wasadded at the first and subsequent changes ofsolution.

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268260
Plants were harvested at the flowering stage.The weight of plant parts was measured afterdesiccation at 75 °C during 48 h.
2.4. Measurement of nodulated roots O2 uptakeand C2H2 reduction
The nodule O2 uptake was measured by regis-tering with an oxymeter (Abiss, F-91170 ViryChatillon) the changes in O2 concentration duringsuccessive confinement phases of intact nodulatedroot with varying initial pO2 (Ribet and Drevon,1995). After 20 min observation of a stationarynodulated root respiration, as shown by a rectilin-ear slope on the register chart, the nodulated rootenvironment was flushed with a new N2–O2
gaseous mixture, whose pO2 was established withmass flow meters (Tylan, La Verpillere, France).Successive confinements were routinely performedwith initial pO2 of 21, 25, 30, 35, 40, 45, 50 kPa inthe frame of this work.
For each nodulated bean plant, respiration val-ues for successive pO2 were plotted on a graph.The model of regression of nodulated root respi-ration as a function of external O2 concentrationwas established. A linear response in a constantinterval of pO2 close to 21 kPa O2 attested thatnodular conductance was constant. The conduc-tance was therefore calculated by dividing theslope of the above response by the nodule surface.
Subsequently, nodule respiration linked to ni-trogen fixation could be calculated for a givenpO2 as the O2 flow diffusing inside the nodule. Itwas equal to nodular conductance multiplied byexternal O2 concentration, according to Fick’slaw.
The acetylene reducing activity (ARA) of nodu-lated root was measured either in situ as previ-ously described by Vadez et al. (1997) forcommon bean, or on detached nodulated rootsfollowing the procedure of Herdina and Silsbury(1991).
2.5. In �itro enzymatic acti�ities
From nodule cytosol extracts, phospho-enolpyruvate carboxylase, malate dehydrogenaseand the NADH-glutamate activities were assayed
by monitoring NADH oxidation, glutamine syn-thetase (GS) and glutamate synthase (NADH-GOGAT) according to Cordovilla et al. (1994),xanthine deshydrogenase and uricase according toSchubert (1981).
Protein was quantified by the Bradford dye-binding assay. Proline, free amino acids and ure-ides were measured according to Irigoyen et al.(1992), Yemm and Cocking (1955) and Vogelsand van der Drift (1970), respectively.
2.6. Differential display and cDNA cloning
Total RNA of P. �ulgaris root nodule cortexwas prepared with RNeasy Plant Mini Kit (Qia-gen, GMBH, Germany) as described by the man-ufacturer. The RNA extract was treated withRNAse-free DNAse (Promega, USA) in order toavoid genomic DNA contamination. For differen-tial display, a reverse transcription was performedon each RNA population with a (dT)11-VNprimer. The resulting cDNA was used in a PCR(Eppendorf, Germany) reaction with the aboveprimer, and an arbitrary 10 bp primer. The ther-mocycling profile was 40 cycles of 94/30–40/120–72/30 °C s−1.
For cloning a candidate gene cDNA from P.�ulgaris, 2 �g of RNA extracted from dissectednodule cortex was reverse transcribed withMMLV (Promega) in the presence of an oligo-d(T) primer and dNTPs in a final volume of 50 �l.Forward and reverse primers were designed fromthe conserved region of several coding sequencesof the candidate gene, for amplifying a 700 bpfragment. The PCR reactions were carried out ina total volume of 25 �l, including 20 �M of eachdNTP, 200 �M of each primer, 0.5 U of TaqDNA polymerase (Promega) and Taq Promegabuffer. The thermocycling profile was 35 cycles at94/60–55/90–72/60 °C s−1. There was no am-plification with control reaction where RNA wastreated as above but without reverse transcriptase.
The amplification products were loaded on asequence gel for profile comparison and selectionof population-specific products. The latter wererecovered by cutting within the gel, and clonedinto a pGEM-T vector (Promega) for sequencingwith the system from Applied Biosystems 373A
J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268 261
(Foster City) and for synthesising 35S-labelledRNA probes. To generate sense or antisenseprobes, plasmids were linearised with ApaI or PstIand transcribed with SP6 or T7 RNA polymerase,respectively. Radioactive labelling was performedas previously described by Gherbi et al. (1997).
2.7. In situ hybridisation and immunolocalisation
For in situ hybridisation (ISH), nodules of ca.3 mm diameter were harvested five weeks afterinoculation, cut into two parts in a transversalsection (Selker, 1988) and immediately transferredinto the fixative of Luther and Bloch (1989).Vacuum was applied at room temperature withseveral interruptions until the nodule halves wereall sunk into the fixative. Thereafter, the nodulehalves were washed three times for 15 min with10 mM EGTA, 10 mM MgCl2, 1 mM DTT and100 mM glycine in 10 mM Pipes buffer at pH 7.5and incubated overnight in the same buffer with-out glycine. The embedding in methacrylate resinwas performed according to Baskin et al. (1992)after addition of 10 mM DTT and bubbling withN2 during 15 min. The exposure to RNA probewas carried out according to Cox et al. (1984)modified by Schump et al. (2000) after removingthe resin according to Kronenberger et al. (1993).Eventually, the slides were coated with autoradio-graphic emulsion (LM1, Amersham) diluted inwater (2/1), processed as recommended by Amer-sham Co. and stored for one month at 4 °C.Development was performed for 1 min with D-19developer (Kodak). Observations were made witha Leica microscope DMRXA.
For immunolocalisation, nodules were fixedduring 30 min in a mixture of 2% (w/v)paraformaldehyde, 0.5% glutaraldehyde in0.1 mM phosphate buffer, 7.2 pH. Abundantwashing in this buffer (+7.5% sucrose) was fol-lowed by a 4 min postfixation in 1% (v/v) OsO4,rapid dehydration in an ethanol series and embed-ding in London white resin. Thin sections werecollected on parlodion-coated gold grids. The im-munogold reaction was performed as previouslydescribed (Serraj et al., 1998). Observations weremade under 80 kV with a 100C JEOL microscope(Tokyo, Japan).
3. Results and discussion
3.1. Spatial and temporal �ariation in beannodulation
Nodulation surveys in Lauragais show largevariation in nodulation from mean field values ofless than five to more than 50 nodules per plant atflowering stage. In addition, large variations innodule number and mass per plant were observedover years at similar developmental stage, in simi-lar soils (Drevon, 2000). In the Medjerda valley,the nodulation of common bean was low, espe-cially in the sites of Beja and Jendouba where nonodules could be detected at both early vegetativeand flowering stage. By contrast, in Bizerte and inCap Bon of NW Tunisia, mean nodulation variedbetween 0.1 and 2.3 nod pl−1. Nodules disap-peared before flowering in many fields where nod-ules were observed at early vegetative stage (Sifi,2000). In Morocco, nodulation was lower inLoukos than in Ain Atiq, with a mean of20 nod pl−1 (Aurag et al., 2000).
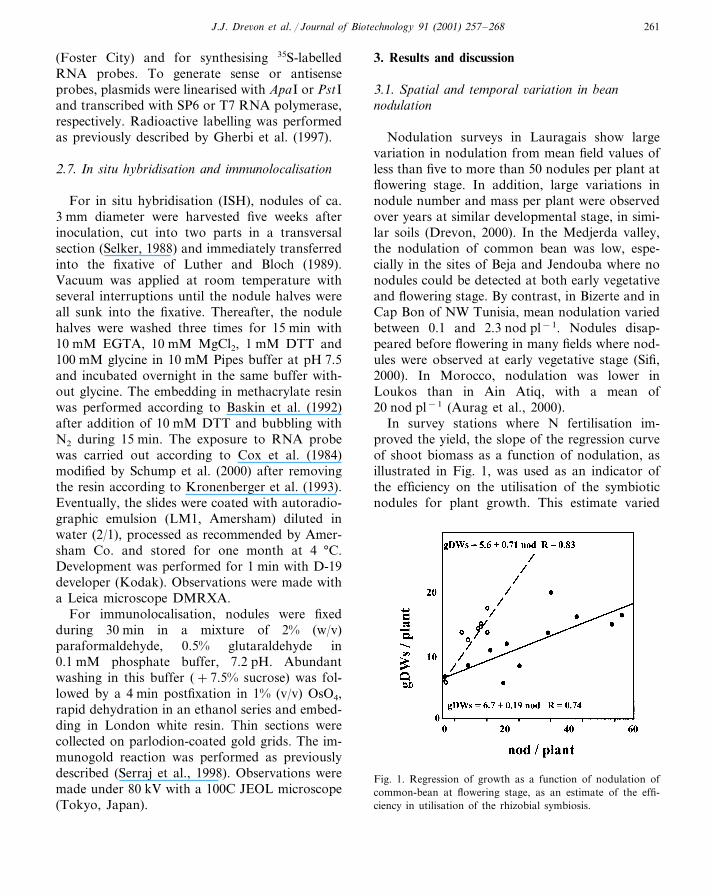
In survey stations where N fertilisation im-proved the yield, the slope of the regression curveof shoot biomass as a function of nodulation, asillustrated in Fig. 1, was used as an indicator ofthe efficiency on the utilisation of the symbioticnodules for plant growth. This estimate varied
Fig. 1. Regression of growth as a function of nodulation ofcommon-bean at flowering stage, as an estimate of the effi-ciency in utilisation of the rhizobial symbiosis.

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268262
between nodulation stations within an area, andbetween areas and years (Drevon, 2000). A highslope of the regression can be interpreted as highefficiency of the symbiotic N supply to the plantgrowth and adequate complementation betweenboth sources of N, i.e. atmosphere and soil. Dif-ferences in efficiency may be due to variation inthe symbiotic potential of the host legume and/orthe microsymbiont.
3.2. Di�ersity of nati�e rhizobia nodulatingcommon bean
From the 300 isolates of the Tunisian collec-tion, the eight following taxons have been iden-tified so far: Rhizobium gallicum, R. etli, R.leguminosarum bv phaseoli, R. giardinii, S. fredii,S. meliloti, S. medicae and ‘pseudo-Agrobac-terium ’ (Mhamdi et al., 1999). This large diversityhas also been revealed by serological studies. In-deed, 62% of representative isolates of theTunisian collection distributed in 19 serogroups(Fekki et al., 2000). In addition, the structure ofrhizobial populations differed significantly be-tween the Medjerda valley, the Cap Bon and thesemi-arid south of Tunisia. In Morocco, so far,two isolates were identified as R. leguminosarumbv phaseoli, four as ‘pseudo-Agrobacterium ’ andsix as R. tropici B (Boumouch et al., 2000).
Such Rhizobium species as R. gallicum and R.giardinii had been proposed previously as newspecies (Amarger et al., 1997), in addition to R.etli and R. leguminosarum bv phaseoli, which arecommonly found outside Latin America in soilswhere common bean has been grown for years.By contrast, the Sinorhizonium spp. nodulating P.�ulgaris were new in the Mediterranean basinsoils. R. tropici B was previously found in soils oftropical Latin America and Africa. However, itwas not found either in Tunisia, where this ab-sence was confirmed by serological studies, or inFrance. Therefore, it needs further exploration.The isolation of Agrobacterium spp. from com-mon bean nodules agrees with a similar observa-tion previously reported by de Lajudie et al.(1999) who worked with Acacia spp. nodules.
Nineteen and 30 rhizobia from Morocco andTunisia, respectively, were found to be at least as
efficient, or more efficient that R. tropici BCIAT899 in symbiosis with the local cultivarCoco. A large variability was found in the toler-ance of native rhizobia to salinity in free livingculture on YEM (Aouani et al., 1998; Aurag etal., 2000). In addition, mutants were obtainedfrom R. etli and R. tropici with altered sensitivityto salinity (Ben Abdelkhalek et al., 2000).
3.3. Selection of symbiotic P. �ulgaris
Numerous lines of P. �ulgaris showed bettergrowth and yield than the local Coco in theMedjerda valley during the three years analysed(1996 considered as a dry year, 1997 as a regularone, and 1998 a humid one) (Trabelsi, 2000).Although this difference varied with the year, 14lines were selected as superior than Coco. Theirnodulation with local rhizobia varied between 0and 27 nod pl−1 in KID53 and BRB17, respec-tively, whereas Coco harboured a significantlydifferent mean of 11 nod pl−1. In tests of theselines within a production area in 1999, DOR585,SVM29-21, FLAMINGO, KID53, RUDDY,CAN74, BRB17, WAF147 expressed a mean yieldhigher than the 1.3 t ha−1 of Coco (Trabelsi andSifi, 2000). With regard to the farmer’s demand,these lines could be sown for pod production.However, none had the seed characteristics tosubstitute Coco for grain for the local marketdemand of white grains. In Lauragais, variabilityin yield was found among 27 white seed lines,with Diego being superior to the local cultivarLinex, with a mean yield of 2.5 t ha−1. Nodula-tion varied between 5 and 50 nod pl−1 for T815and Diego (Rey-Poiroux et al., 2000).
The sensitivity to moderate salinity of linesselected from field observations, including thosecited above, was tested in a glasshouse with Nnutrition depending on either the rhizobial sym-biosis or the supply of urea or NO3
−. The plantgrowth was less affected by the salt treatmentwith mineral N than with N2, and the amount ofN fixed was the most limited (Saadallah et al.,2000). Salinity inhibited not only the nodulationprocess but also the nodule growth, except forDOR585, and the nodule function. This highersensitivity of symbiotic plants was associated with
J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268 263
an excessive accumulation of Cl− in leaves, andoverloading of nodules with toxic ions Na+ andCl− (Lachaal et al., 2000). In symbiosis withCIAT899, Coco was more sensitive than BAT477,DOR364, DOR585 and Flamingo, whereasBRB17, ABA16 and Dark were the most sensitive(Boughribil et al., 2000; Abdelly et al., 2000).
The higher sensitivity of Coco compared toBAT477 was associated with higher content ofNa+ and mostly Cl− of leaves, though there wasno significant difference in K+ (Abdelly et al.,2000). Content of Na+ in the roots tissue was alsohigher for Coco than BAT477. It was muchhigher than that of leaves, which agrees with theknown exclusive behaviour of common bean.Content of Na+ of nodules was higher in BAT477and Dark than in Coco. However, it was muchlower than nodule content of Cl−, which did notdiffer significantly between contrasting lines. Nod-ule P content was not affected by salinity. Asplit-root experiment confirmed that the sensitiv-ity of symbiotic common bean to NaCl was notdue to toxic effects of Na or Cl accumulation innodules or leaves (Lachaal et al., 2000). Mostprobably the NaCl inhibition was associated withalteration of other nutrients acquisition. In addi-tion, the contrasting responses of common beanlines to salinity was not a consequence of anydifference in initial vigour of the seedlings (Ab-delly et al., 2000).
Whatever the line, the nodulation was muchhigher in the glasshouse than in the fields. Thelow nodulation of common bean in the variousareas of field observation may be due to suchadverse environmental conditions as water defi-ciency or excess of soil nitrate or salinity, or tospecific interactions with the native rhizobia. Toanalyse the latter, cross-inoculation trial betweenselected rhizobial isolates and bean lines was car-ried out.
3.4. Interaction of nati�e rhizobia with selectedbean lines
The initial cross-inoculation trials were per-formed in sand culture, with eight plant lines andfour local rhizobial isolates chosen among thedifferent species, e.g. in Tunisia, 31c3 (R. legumi-
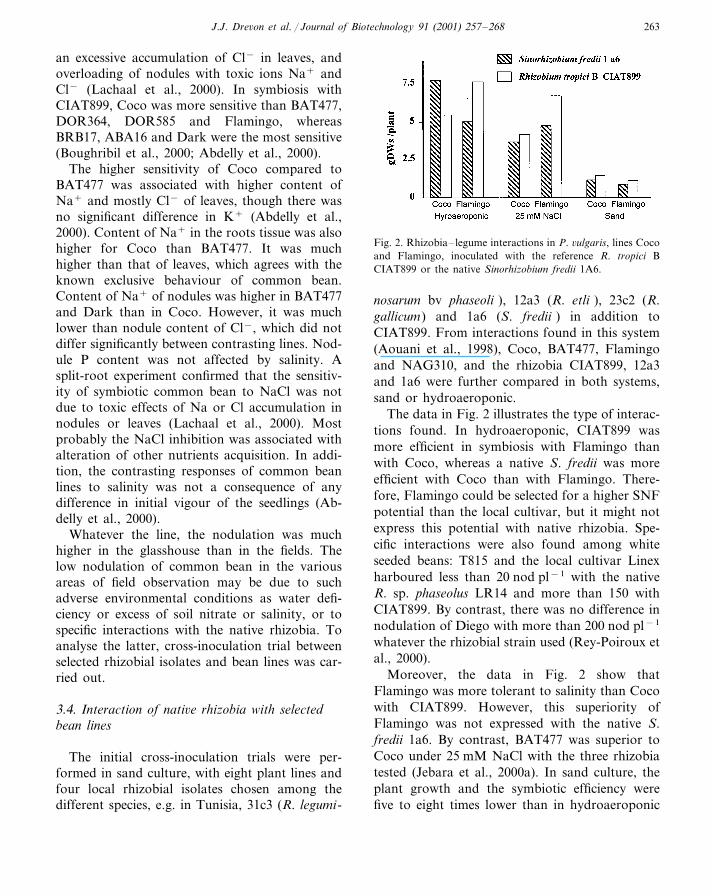
Fig. 2. Rhizobia– legume interactions in P. �ulgaris, lines Cocoand Flamingo, inoculated with the reference R. tropici BCIAT899 or the native Sinorhizobium fredii 1A6.
nosarum bv phaseoli ), 12a3 (R. etli ), 23c2 (R.gallicum) and 1a6 (S. fredii ) in addition toCIAT899. From interactions found in this system(Aouani et al., 1998), Coco, BAT477, Flamingoand NAG310, and the rhizobia CIAT899, 12a3and 1a6 were further compared in both systems,sand or hydroaeroponic.
The data in Fig. 2 illustrates the type of interac-tions found. In hydroaeroponic, CIAT899 wasmore efficient in symbiosis with Flamingo thanwith Coco, whereas a native S. fredii was moreefficient with Coco than with Flamingo. There-fore, Flamingo could be selected for a higher SNFpotential than the local cultivar, but it might notexpress this potential with native rhizobia. Spe-cific interactions were also found among whiteseeded beans: T815 and the local cultivar Linexharboured less than 20 nod pl−1 with the nativeR. sp. phaseolus LR14 and more than 150 withCIAT899. By contrast, there was no difference innodulation of Diego with more than 200 nod pl−1
whatever the rhizobial strain used (Rey-Poiroux etal., 2000).
Moreover, the data in Fig. 2 show thatFlamingo was more tolerant to salinity than Cocowith CIAT899. However, this superiority ofFlamingo was not expressed with the native S.fredii 1a6. By contrast, BAT477 was superior toCoco under 25 mM NaCl with the three rhizobiatested (Jebara et al., 2000a). In sand culture, theplant growth and the symbiotic efficiency werefive to eight times lower than in hydroaeroponic

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268264
(Fig. 2). Thus, sand culture may not be adequateto select the most efficient symbiotic partners.Moreover, the symbioses ranking for SNF wasdifferent from that found in hydroaeroponic,probably because of differences in the sensitivityto water deficiency, presumed to be a major limi-tation of the sand culture.
In this project, no correlation was found be-tween the variation of tolerance of native rhizobiafor growth in free living culture under salinity,and that for symbiotic efficiency under moderatesalinity. This contrasts with the higher nitrogenfixation levels observed by Ben Abdelkhalek et al.(2000) with the Tn5 mutants which displayedhigher tolerance to salinity than the parent R. etliCFN42 in YEM.
3.5. Nodule metabolism and O2 permeability
Since NaCl tolerant symbioses were selectedpartly in controlled environment, physiologicalassays could be performed in order to search formechanisms involved in the tolerance. Nodulephysiology is characterised by an intensecatabolism of sucrose from photosynthesis andrespiration to support the reduction of N2 bynitrogenase. Reduced N2 is incorporated into glu-tamate via glutamine synthetase (GS)–glutamatesynthase (GOGAT), and subsequently transami-nated into glutamine or asparagine, ormetabolised into ureides for xylemic export toother plant organs. A central role is played byPEPC which carboxylates trioses into C4 organicacids for amino acid synthesis, bacteroids supplyand probably participates in the osmotic and pHregulations within nodule.
Higher nodule PEPC and malate dehydroge-nase (MDH) activities were found in NaCl toler-ant symbioses (Pliego et al., 2000). This wasassociated with higher contents of total solublesugars which could contribute to the energeticand/or osmoregulation requirements of the nod-ule metabolism. In the control plants, PEPC andalcaline invertase (AI) were stimulated by NaClduring vegetative growth whereas MDH and su-crose synthase (SS) were inhibited during flower-ing, though the nitrogenase activity (ARA) wasdrastically reduced during all stages (Pliego et al.,
2000). GS, GOGAT, xanthine dehydrogenase anduricase were significantly decreased (Khadri et al.,2000) as previously observed in Vicia faba byCordovilla et al. (1994) and in Cicer arietinum bySoussi et al. (1998).
The contents of ureides and aminoacids weredecreased by NaCl. This contrasts with previousobservations by Soussi et al. (1998) in chickpeaand by Gordon et al. (1997) in soybean, where anaccumulation of aminoacids was observed as adirect effect of salt. These discrepancies may bedue to differences between not only species, butalso timing of salt application. Finally, prolineaccumulation was not correlated with levels andtime of salt application. This confirms that prolineaccumulation was more a symptom of susceptibil-ity to salt than an adaptive response.
Oxygen plays a critical role in nodule physiol-ogy. Indeed, O2 is toxic for nitrogenase though itis necessary for the regeneration of the ATP sup-porting nitrogenase activity. The response of nod-ule ARA to rhizospheric pO2 suggests that thenodule O2 concentration is regulated at a subopti-mal level in ambient conditions (Jebara et al.,2000b), as previously found with soybean(Drevon et al., 1988). A low and variable perme-ability of the nodule cortex may control the O2
entry within the organ (Minchin, 1997; Drevon etal., 1995). The methodology for measuring nodulepermeability was shown to be valid in commonbean (Jebara et al., 2000b), although changes inpermeability were slightly faster in common beanthan in soybean (Roy, 1993). Nodule permeabilitywas increased under salinity. This was associatedwith a decrease in nodulation, and shared similar-ities with previous observations in soybean underphosphorus deficiency (Ribet and Drevon, 1995).Although nodule individual area was higher underNaCl excess than under P deficiency, this higherpermeability under salinity contrasts with theshort-term decrease in nodule permeability whichwas observed immediately after NaCl application(Serraj et al., 1994) and was associated with acontraction of nodule inner-cortex cells (Serraj etal., 1995).
Since a hypothesis of osmoregulation of thenodule permeability was postulated from previousobservations (Drevon et al., 1995), and since ab-

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268 265
cissic acid (ABA) has been associated with thecontraction of the stomatal guard cell in responseto NaCl application, the ABA content of com-mon bean nodules was measured. The NaCl ap-plication increased nodule ABA content, and theexogenous ABA application decreased nodulerespiration (Irekti and Drevon, 2000). The latterwas compensated by an increase in rhizosphericpO2, thereby indicating that nodule permeabilityhad been decreased (Jebara et al., 2000b). TheABA application decreased leghemoglobin con-tent in pea, but it did not affect nodule SS, noraspartate amino transferase, AI, MDH, GS,uridine diphosphoglucose pyrophosphorylase,isocitrate dehydrogenase and GOGAT (Gonzalezet al., 2000). Interestingly, the pretreatment withABA decreased the inhibitory effects of NaCland the related N2-dependent growth of thenodulated common bean (Lluch, personal com-munication).
3.6. Gene expression in nodule tissue
Since the NaCl-linked variation of nodule per-meability has been localised within the nodulecortex, an in situ hybridisation (ISH) methodol-ogy was developed in order to test whether it wasassociated with any detectable change in geneexpression within nodule cortical cells, and fur-ther understand the nodule function under highNaCl concentrations. Thus, the quantitative tran-scriptional analysis of specific genes at the tissuelevel could be a major link between physiologyand genomics within a multidisciplinary project.
The nodule-specific carbonic anhydrase (CA)was chosen because its expression was restrictedto the nodule cortex in Medicago sati�a (de laPena et al., 1997). Moreover, CA catalyses thehydration of CO2 into bicarbonate, the substrateof PEPC, an enzyme which may play a role innodule response to salinity according to theabove results. The P�ca gene obtained from anmRNA extract of bean nodule cortex shared 90%homology with that of M. sati�a and expressedexclusively in the two to four cell layers betweenthe vascular traces and the most internal non-in-fected cells surrounding the infected zone(Schump et al., 2000). This corresponded to the
nodule inner-cortex parenchyma where cell sizewas shown to decrease when permeability wasdecreased by salt or excess O2 (Serraj et al.,1995). Although the intensity of the ISH signalvaried among nodules and plants of a single ex-periment, a higher mean intensity was associatedwith a higher nodule permeability due to P defi-ciency. These results are consistent with an os-moregulatory regulation of the nodulepermeability: the CA would be involved in thesynthesis of malate subsequently accumulated inthe vacuoles as an osmoticum; malate would alsoact as a counter anion of the putative K+ accu-mulation driving the water accumulation andsubsequent turgidity of the inner-cortex cells(Drevon et al., 1995). However, it does not ex-clude other roles of the CA, such as (i) supplyingorganic acids for increased bacteroidalmetabolism and aminoacid syntheses, or (ii) fa-vouring the nodule evolution of CO2 by highersolubilisation and diffusion of bicarbonatethrough the occluded intercellular spaces of thenodule cortex. Aquaporin was the other candi-date gene chosen in this project since bothtonoplastic (Serraj et al., 1998) and plasmalemic(Fleurat-Lessard et al., 2000) aquaporins werefound by immunolocalisation to be overexpressedin the nodule inner cortex in soybean nodule. Apreliminary result showed that the expression ofthe 680 pb P�aq obtained from the bean nodulecortex localised in the nodule inner cortex.
In order to search for other genes of which thealtered expression in the nodule cortex would beassociated with variation of the nodule perme-ability, a differential display experiment was per-formed by Gherbi et al. (2000) with mRNAsisolated from nodule cortex exhibiting differencesin permeability. Among 55 cDNA cloned, eightconfirmed differential expression by dot–blot ex-periments. In particular, the clone 43 was furthershown by northern to have higher expression innodules with higher permeability. The ISH re-vealed that the clone 43 was expressed in thenodule external cortex. In addition, its sequenceanalysis revealed significant homology with aparietal pathogen-related protein (Gherbi et al.,2000).

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268266
4. Conclusion and prospects
The multidisciplinary work during theFYSAME project benefited mostly from thesharing of controls and the exchanging of mostpromising results between disciplines. The abilityto distinguish native Rhizobiaceae species fromproduction areas made it possible to test theirinteraction with the selected plant material. Sincevariations were observed in these interactions,more work is needed in this field, which mayreinforce the interaction between bacteriologistand plant breeders that this project has promoted.In addition, the screening in controlled environ-ment gave the opportunity for plant breeders andphysiologists to interact. It is now required toexpand this link to the functional genomics withsuch methodologies as immunolocalisation and insitu hybridisation, among others to be furtherexplored.
The most difficult interaction was probably atthe field level. Indeed, it depended not only on theability of the above disciplines to interact withagronomists and producers, but also on the socialorganisation in production areas. The latter wasprobably the major reason why it has been moredifficult to perform the field observations in theless developed countries of Morocco and Tunisiathan in France. Therefore, this level of interactionalso needs more work in order to implement anintegrated improvement of the symbiotic nitrogenfixation in a grain legume like common bean.Such progress might contribute to the full benefitof the disciplines involved in rhizobiology to pro-gress in a cooperative group.
Acknowledgements
This work was supported by Project No. ER-BIC18C960081 (FYSAME) of the INCO DC pro-gramme of the European Union.
References
Abdelly, C., Saadallah, K., Drevon, J.J., 2000. Fixation bi-ologique de l’azote en conditions de salinite et de deficience
en phosphore chez deux varietes de haricot BAT477 (toler-ante) et Coco (sensible). In: Drevon, J.J., Sifi, B. (Eds.),Fixation Symbiotique de l’Azote et DeveloppementDurable dans le Bassin Mediterraneen. INRA Les Collo-ques, Paris (in press).
Amarger, N., Macheret, V., Laguerre, G., 1997. Rhizobiumgallicum sp. nov. and Rhizobium giardinii sp. nov. fromPhaseolus �ulgaris nodules. Int. J. Syst. Bacteriol. 47, 996–1006.
Aouani, M.E., Mhamdi, R., Mars, M., Ghrir, R., 1998. Nodu-lation and growth of common bean under NaCl-stress. SoilBiol. Biochem. 301, 473–1475.
Aurag, J., Boumouch, I., Filali-Maltouf, A., 2000. Ecophysi-ologie des souches de Rhizobium nodulant le haricot(Phaseolus �ulgaris L.) dans les sols salins. In: Drevon, J.J.,Sifi, B. (Eds.), Fixation Symbiotique de l’Azote et Devel-oppement Durable dans le Bassin Mediterraneen. INRALes Colloques, Paris (in press).
Baskin, T.I., Busby, C.H., Fowke, L.C., Sammut, M., Gubler,F., 1992. Improvements in immunostaining samples em-bedded in methacrylate: localization of microtubules andother antigens throughout developing organs in plants ofdiverse taxa. Planta 187, 405–413.
Ben Abdelkhalek, H., Sanjuan, J., Olivares, J., Lluch, C.,2000. Amelioration de la symbiose Rhizobium etli–Phaseo-lus �ulgaris dans des conditions de stress salin par l’utilisa-tion de souches bacteriennes tolerantes a la salinite. In:Drevon, J.J., Sifi, B. (Eds.), Fixation Symbiotiquede l’Azote et Developpement Durable dans le Bassin Med-iterraneen. INRA Les Colloques, Paris (in press).
Boughribil, S., Lachaal, M., Abdelly, C., Soltani, A., Hajji,M., 2000. Implication du statut ionique des nodules dans lafixation de l’azote en milieu sale chez le haricot. In:Drevon, J.J., Sifi, B. (Eds.), Fixation Symbiotiquede l’Azote et Developpement Durable dans le Bassin Med-iterraneen. INRA Les Colloques, Paris (in press).
Boumouch, I., Brhada, F., Filali-Maltouf, A., Aurag, J., 2000.Selection of osmotolerant and effective strains of Rhizo-bium for inoculation of common bean (Phaseolus �ulgarisL.) in Moroccan saline soils. Agronomie, submitted.
Cordovilla, M.P., Ligero, F., Lluch, C., 1994. The effect ofsalinity on N2 fixation and assimilation in Vicia faba. J.Exp. Bot. 45, 1483–1488.
Cox, K.H., DeLeon, D.V., Angerer, L.M., Angerer, R.C.,1984. Detection of mRNAs in sea urchin embryos by insitu hybridization using asymmetric RNA probes. Dev.Biol. 101, 485–502.
de Lajudie, P., Willems, A., Nick, G., Mohamed, S.H., Torck,U., Coopman, R., Filali Maltouf, A., Kersters, K., Drey-fus, B., Lindstrom, K., Gillis, M., 1999. Agrobacterium bv.1 strains isolated from nodules of tropical legumes. Syst.Appl. Microbiol. 22, 119–132.
de la Pena, T.C., Frugier, F., McKhann, H.I., Bauer, P.,Brown, S., Kondorosi, A., Crespi, M., 1997. A carbonicanhydrase gene is induced in the nodule primordium andits cell-specific expression is controlled by the presence ofRhizobium during development. Plant J. 11, 407–420.

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268 267
Drevon, J.J., 2000. Nodular diagnostic: agronomic basis for anintegrated improvement of symbiotic nitrogen fixation andthe N2-dependent yield of grain-legumes. In: Morot-Gaudry, J.F. (Ed.), Nitrogen Assimilation by Plants. Sci-ence Publishers Inc./INRA, Paris, pp. 417–425.
Drevon, J.J., Kalia, V.C., Heckmann, M.O., Pedelahore, P.,1988. In situ open-flow assay of soybean root nodularacetylene reduction activity: influence of acetylene andoxygen. Plant Physiol. Biochem. 26, 73–78.
Drevon, J.J., Deransart, C., Fleurat-Lessard, P., Jaillard, B.,Ndjiondjop, M.N., Payre, H., Ribet, J., Roy, G., Serraj,R., 1995. Is the symbiotic fixation osmoregulated by re-versible contraction of cells in the legume-nodule innercortex? In: Tikhonovitch, I.A., Provorov, N.A., Romanov,V.I., Newton, W.E. (Eds.), Nitrogen Fixation: Fundamen-tals and Applications. Kluwer Academic, Dordrecht, p.598.
Fekki, S., Aouani, M.E., Mhamdi, R., Jebara, M., Mars, M.,Ghrir, R., 2000. Caracterisation serologique d’une collec-tion de Rhizobium nodulant le haricot et application auxetudes de competitivite. In: Drevon, J.J., Sifi, B. (Eds.),Fixation Symbiotique de l’Azote et DeveloppementDurable dans le Bassin Mediterraneen. INRA Les Collo-ques, Paris (in press).
Fleurat-Lessard, P., Michonneau, P., Maeshima, M., Serraj,R., Drevon, J.J., 2000. Expression of PAQ1 and g-TIPaquaporins is tissue dependent soybean (Glycine max) rootnodules. Planta, submitted.
Gherbi, H., Duhoux, E., Franche, C., Katharina, P., Nassar,A., Berry, A.M., Bogusz, D., 1997. Cloning of a full-lengthsymbiotic hemoglobin cDNA and in situ localization of thecorresponding mRNA in Casuarina glauca root nodule.Physiol. Plant 99, 608–616.
Gherbi, H., Schump, O., Payre, H., Drevon, J.J., 2000. Ex-pression differentielle de genes du cortex de nodules deharicot. In: Drevon, J.J., Sifi, B. (Eds.), Fixation Symbio-tique de l’Azote et Developpement Durable dans le BassinMediterraneen. INRA Les Colloques, Paris (in press).
Gonzalez, E.M., Galvez, L., Arrese-Igor, C., 2000. Abcissicacid induces a decline in nitrogen fixation that involvesleghaemoglobin, but is independent of sucrose synthaseactivity. J. Exp. Bot., in press.
Gordon, A.J., Minchin, F.R., Skot, L., James, C.L., 1997.Stress-induced declines in soybean N2 fixation are relatedto nodule sucrose synthase activity. Plant Physiol. 114,937–946.
Herdina, J.A., Silsbury, J.H., 1991. Estimating nitrogenaseactivity of faba bean (Vicia faba L.) by acetylene reduction(AR) assay. Aust. J. Plant Physiol. 17, 489–502.
Irekti, H., Drevon, J.J., 2000. Acide abscissique et reponse dela permeabilte nodulaire a la salinite. In: Drevon, J.J., Sifi,B. (Eds.), Fixation Symbiotique de l’Azote et Developpe-ment Durable dans le Bassin Mediterraneen. INRA LesColloques, Paris (in press).
Irigoyen, J.J., Emerich, D.W., Sanchez-Diaz, M., 1992. Waterstress induced changes in concentrations of proline andtotal soluble sugar in nodulated alfalfa (Medicago sati�a)plants. Physiol. Plant 84, 55–60.
Jebara, M., Aouani, M.E., Drevon, J.J., 2000a. Interactionlignees de haricots-rhizobium-specifiques: effets du systemede culture et de NaCl. In: Drevon, J.J., Sifi, B. (Eds.),Fixation Symbiotique de l’Azote et DeveloppementDurable dans le Bassin Mediterraneen. INRA Les Collo-ques, Paris (in press).
Jebara, M., Aouani, M.E., Drevon J.J., 2000b. Genotypicvariation in nodule conductance to the oxygen diffusion incommon bean (Phaseolus �ulgaris). Agronomie, submitted.
Khadri, M., Pliego, L., Soussi, M., Ocana, A., Lluch, C., 2000.Assimilation de l’ammonium et metabolisme des ureideschez Phaseolus �ulgaris : effet du stress salin. In: Drevon,J.J., Sifi, B. (Eds.), Fixation Symbiotique de l’Azote etDeveloppement Durable dans le Bassin Mediterraneen.INRA Les Colloques, Paris (in press).
Kronenberger, J., Desprez, T., Hefte, H., Caboche, M., Traas,J.G., 1993. A methacrylate embedding procedure devel-oped for immunolocalization on plant is also compatiblewith in situ hybridization. Cell Biol. Int. 17, 1013–1021.
Lachaal, M., Boughribil, S., Soltani, A., Hajji, M., 2000.Sensibilite au sel du haricot (Phaseolus �ulgaris L.) enrelation avec l’accumulation de Na+ et Cl− dans lesnodosites. In: Drevon, J.J., Sifi, B. (Eds.), Fixation Symbi-otique de l’Azote et Developpement Durable dans leBassin Mediterraneen. INRA Les Colloques, Paris (inpress).
Laguerre, G., Allard, M.R., Revoy, F., Amarger, N., 1994.Rapid identification of rhizobia by RFLP analysis of PCR-amplified 16S rRNA genes. Appl. Environ. Microbiol. 60,56–63.
Luther, P.W., Bloch, R.J., 1989. Formaldehyde-amine fixativesfor immunocytochemistry. J. Histochem. Cytochem. 37,75–82.
Mhamdi, R., Jebara, M., Aouani, M.E., Ghrir, R., Mars, M.,1999. Genotypic diversity and symbiotic effectiveness ofrhizobia isolated from root nodules of Phaseolus �ulgarisL. grown in Tunisian soils. Biol. Fertil. Soils 28, 313–320.
Minchin, F.R., 1997. Regulation of oxygen diffusion in legumenodules. Soil Biol. Biochem. 29, 881–888.
Pliego, L., Khadri, M., Soussi, M., Ocana, A., Lluch, C., 2000.Effet de la salinit sur la fixation de l’azote et lemetabolisme du carbone dans les nodules de haricot(Phaseolus �ulgaris). In: Drevon, J.J., Sifi, B. (Eds.), Fixa-tion Symbiotique de l’Azote et Developpement Durabledans le Bassin Mediterraneen. INRA les Colloques, Paris(in press).
Rey-Poiroux, X., Drevon, J.J., 2000. Variabilite genotypiquede la fixation d’azote chez le haricot de type lingot. In:Drevon, J.J., Sifi, B. (Eds.), Fixation Symbiotiquede l’Azote et Developpement Durable dans le Bassin Med-iterraneen. INRA Les Colloques, Paris (in press).
Ribet, J., Drevon, J.J., 1995. Increase in permeability tooxygen and in oxygen uptake of soybean nodules underlimiting phosphorus nutrition. Physiol. Plant 94, 298–304.
Roy, G., 1993. Respiration et Diffusion de l’Oxygene dans laSymbiose Fixatrice d’Azote: Soja-Bradhyryzobiumjaponicum. Ph.D. thesis, Ecole Nationale SuperieureAgronomique de Montpellier, 98 pp.

J.J. Dre�on et al. / Journal of Biotechnology 91 (2001) 257–268268
Saadallah, K., Abdelly, C., Drevon, J.J., 2000. Nodulation etcroissance nodulaire chez le haricot (Phaseolus �ulgaris L.)sous contrainte saline. Agronomie, in press.
Schubert, K.R., 1981. Products of biological nitrogen fixationin higher plants: synthesis, transport and metabolism. Ann.Rev. Plant Physiol. 37, 539–574.
Schump, O., Gherbi, H., Escoute, J., Gamas, P., De Billy, F.,Payre, H., Drevon, J.J., 2000. Fine expression of nodule-specific carbonic anhydrase by in situ hybridization onmethacrylate embedded root-nodules of common bean(Phaseolus �ulgaris). J. Plant Physiol., submitted.
Selker, J.M.L., 1988. Three-dimensional organization of unin-fected tissues in soybean root nodules and its relation tocell specialization in the central region. Protoplasma 147,178–190.
Serraj, R., Roy, G., Drevon, J.J., 1994. Salt stress induces adecrease in the oxygen uptake of soybean nodules and intheir permeability to oxygen. Physiol. Plant 91, 161–168.
Serraj, R., Fleurat-Lessart, P., Jaillard, B., Drevon, J.J., 1995.Changes in soybean nodule cortical cells under short-termsalt stress and altered oxygen concentration. Plant CellEnviron. 18, 455–462.
Serraj, R., Frangne, N., Maeshima, M., Fleurat-Leussard, P.,Drevon, J.J., 1998. A g-TIP cross reacting protein is abun-dant in the cortex of soybean nitrogen-fixing nodules.Planta 206, 681–684.
Sifi, B., 2000. Diagnostic nodulaire et inoculation des culturesde pois chiche et du haricot en Tunisie. In: Drevon, J.J.,
Sifi, B. (Eds.), Fixation Symbiotique de l’Azote et Devel-oppement Durable dans le Bassin Mediterraneen. INRALes Colloques, Paris (in press).
Soussi, M., Ocana, A., Lluch, C., 1998. Effects of salt stress ongrowth, photosynthesis and nitrogen fixation in chick-pea(Cicer arietinum L.). J. Exp. Bot. 49, 1329–1337.
Trabelsi, M., 2000. Caracterisation biologique et agronomiqued’une collection de genotypes de haricot. In: Drevon, J.J.,Sifi, B. (Eds.), Fixation Symbiotique de l’Azote et Devel-oppement Durable dans le Bassin Mediterraneen. INRALes Colloques, Paris (in press).
Trabelsi, M., Sifi, B., 2000. Investigations on common-beansymbiosis in Tunisia. Agronomie, submitted.
Vadez, V., Rodier, F., Payre, H., Drevon, J.J., 1996. Nodulepermeability to O2 and nitrogenase-linked respiration inbean genotypes varying in the tolerance of N2 fixation to Pdeficiency. Plant Physiol. Biochem. 34, 871–878.
Vadez, V., Lasso, J.H., Beck, D.P., Drevon, J.J., 1997. Utiliza-tion of the acetylene reduction assay to screen for toleranceof N2 fixation to limiting P nutrition in common bean.Physiol. Plant 99, 227–232.
Vincent, M., 1970. A Manual for the Practical Study of RootNodule Bacteria. In: IBP Handbooks, vol. 15. BlackwellScientific Publications, Oxford.
Vogels, G.D., van der Drift, C., 1970. Differential analyses ofglyoxylate derivatives. Anal. Biochem. 33, 143–157.
Yemm, E.W., Cocking, E.C., 1955. The determination ofamino acids with ninhydrin. Analyst 80, 209–213.
Related Documents











