An Emulsion Technique For Quantitating High Affinity Uptake of Postprandial Lipoproteins In Vivo: Foundations for a Diagnostic Assay Hanni Christine Gennat (BSc) This thesis is submitted for the Degree of Doctor of Philosophy in the Department of Physiology, The University of Western Australia (2002).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A n Emulsion Technique For
Quantitating High Affinity Uptake
of Postprandial Lipoproteins
In Vivo:
Foundations for a Diagnostic Assay
Hanni Christine Gennat (BSc)
This thesis is submitted for the Degree of Doctor of Philosophy in the Department of Physiology,
The University of Western Australia (2002).

2
SlcfqiozvUdgments
For me, this thesis has been a personal achievement of endurance, pushing my limits
and discovering patience I never thought I possessed. However, it would never have
been conceived if not for A/Prof. John M a m o , and would not have been realised if not
for his encouragement and faith in m y ability. I greatly admire his enthusiasm,
scientific mind and warmth. To Prof. Trevor Redgrave, who has always been
available to give advice, feedback and guidance, for which I am very grateful. I would
not have made it through without you both. I would also like to acknowledge the
Raine Foundation for funding the project.
Without Dr Jane den Hollander who supported me with overwhelming encouragement
and employment, and without w h o m I could not have finished this project. Thank you
so much for being a pillar of strength and a mentor who I greatly admire. This
motivation has also been ongoing and invaluable from m y friends and colleagues in
the laboratory, in particular Donna Vine, Spencer Proctor, Caryn Elsegood, Kenny
Yu, Darrin Smith, Sebely Pal, E m m a Allister, Cheryl Dane-Stewart, Melanie
Voevodin and Tony James. Thank you to you all for your support over the years.
I am also extremely grateful to those who gave me advice with HPLC apparatus,
histology, confocal microscopy and emulsion technology, including Dr Kevin Croft,
Dr Trevor Mori, Alan Light, John Murphy and Dr Ian Martins. Thank you to Dr Phil
Oates, Mark Edwards, Ross Oxwell, and Prof. Don Robertson who were always
available to lend an ear and give their support. Thanks is also extended to the staff at
B S A U Animal Facility and the Animal Research Centre (Royal Perth Hospital) for
their continued assistance, in particular Sandy Goodin and Geoff Billiewicz, Terry
York and Shane Meakins, and Ann Storrie for her surgical assistance.
To my good friends Justina, Nahla, Karin, Vanessa, Vivi, Andrea, Jane, Mari, Corina,
Monica, Chas, Martin, Jude, Alex, Jacque, Vendy, Leonie, Sean, Sato and Adrian and
there many others I have not mentioned because the list is too long. Your friendship
has provided m e with so much strength over the years and I will always remember the
help through hard times, the constant inquiries about "the" PhD, the faith in m y ability

3
to carry this project through and all those yummy dinners. To Robyn and Sandra, a
heartfelt and very special thanks for your continued interest, feedback, and
overwhelming support - I love you both dearly. To m y family who I have missed
incredibly, I hope you understand why I endeavoured to achieve such an incredible
feat and look forward to spending lots of good times with you in the future.
Finally, I would like to thank Janice Halliwell, who has been my companion and pillar
of strength, and who has been through the good times and not-so-good times with m e
through this epic. Your constant love and support is undeniable and very precious and
I look forward to a life with you, and without a thesis to write.

4
abstract of thesis
The metabolism of chylomicrons and their remnants is delayed in certain disease
states. Several studies have shown that the LDL-receptor is the primary mechanism
for the removal of chylomicron remnants from plasma; down regulation of this
receptor may therefore result in a delay in the hepatic clearance of these particles.
However, at present there is no simple method by which high-affinity (receptor)
uptake mechanisms can be detected in humans. Therefore, the principal aim of this
thesis was to develop an emulsion technique for quantitating high affinity uptake of
chylomicron remnants in vivo. The assay is based on the clearance patterns of two
chylomicron-like lipid emulsions. The clearance of normal emulsion represents total
uptake from plasma via high and low affinity mechanisms; in contrast, modified
emulsions do not interact with receptor mechanisms and are cleared via low-affinity
mechanisms only. The difference between the clearances of these two emulsions gives
a measure of high-affinity uptake of postprandial lipoproteins.
The specificity of the proposed technique in quantitating receptor-mediated
uptake of remnants was investigated using in vivo clearance studies (Chapter 3) and
fluorescent probes to examine the uptake of emulsions on a cellular level (Chapter 5).
The findings verified hepatic uptake of normal emulsions via high and low affinity
pathways, as demonstrated in control and LDL-receptor-deficient animal models. In
contrast, modified emulsions did not interact with receptor mechanisms, specifically
the LDL-receptor, and were cleared via non-specific pathways. Hepatic uptake of
emulsion remnant particles in apo E-deficient mice was significantly delayed,
suggesting that apo E is an essential ligand for remnant metabolism via high and low
affinity pathways.
Current monitoring of chylomicron remnant kinetics in vivo involves the
utilisation of radioisotopes as markers of particle clearance, however these are not
suitable for use in humans. The use of vitamin A esters as an alternative to radiolabels
was investigated using animal models. Retinyl esters were incorporated into
emulsions and injected as a bolus dose, directly into the bloodstream. The
incorporation of retinyl esters did not alter the in vivo clearance kinetics of
chylomicron-like emulsions (Chapter 6). Furthermore, the clearance of retinyl
myristate and retinyl palmitate in emulsions closely followed that of radiolabeled

5
cholesteryl oleate in normal and modified emulsions, respectively, suggesting that
retinyl esters are suitable as markers of remnant particles in vivo. The detection of
retinyl esters in plasma by H P L C enabled the calculation of plasma clearance of
retinyl esters over time and the quantitation of chylomicron remnant uptake via high
affinity pathways.
The use of remnant-like emulsions to monitor chylomicron remnant
metabolism is an attractive alternative, as the relative increase in cholesteryl esters
offers properties of stability and the reduced triglyceride mass allows the process of
lipolysis to be bypassed, thereby reducing confounding factors. Chylomicron remnant
composition and size was characterised extensively (Chapter 4), with the objective of
using the data to synthesise remnant-like emulsions representative of nascent
chylomicron remnants. However, the synthesis of remnant-like emulsions based on
these results proved inconclusive and further refinement is required.
Collectively, the data permits the conclusion that normal chylomicron-like
emulsions are taken up via receptor and non-receptor pathways. Modified emulsions
do not interact with receptor mechanisms and therefore their clearance is
representative of low-affinity uptake. Furthermore, retinyl myristate and retinyl
palmitate may be incorporated into chylomicron-like emulsions without altering the
clearance kinetics in vivo, and may be utilised as tracees for normal and modified
remnant particles, respectively. A s a result, high affinity uptake of postprandial
lipoproteins can be quantitated from the clearance of chylomicron-like emulsion
retinyl esters from plasma. The study provides a foundation for further development
of the two emulsion technique for future use in human subjects.

6
Ta/?/e of Contents
Acknowledgements 2 Abstract 4 List of Tables 13 List of Figures 14 List of Non-Standard Abbreviations 16 List of Published Manuscripts 18
Chapter 1: Review of Literature 19
1.1 Introduction 19 1.2 Lipoproteins 21 1.2.1 Lipoprotein Synthesis 21 1.2.2 Chylomicrons 25 1.2.2.1 Synthesis 25 1.2.2.2 Structure and Composition 26 1.2.2.3 Metabolism 27
1.2.3 Chylomicron Remnants 28 1.2.3.1 Synthesis 28 1.2.3.2 Structure and Composition 29
1.2.3.3 Metabolism 30 1.2.4 Very Low-Density Lipoprotein (VLDL) 31
1.2.4.1 Synthesis 31 1.2.4.2 Structure and Composition 32
1.2.4.3 Metabolism 33 1.2.5 L o w Density Lipoproteins (LDL) 33
1.2.5.1 Synthesis 33 1.2.5.2 Structure and Composition 34 1.2.5.3 Metabolism 34
1.2.6 High Density Lipoproteins (HDL) 35
1.3 Lipases 37 1.3.1 Lipoprotein Lipase (LPL) 37 1.3.2 Hepatic Lipase (HL) 39 1.3.3 Cholesterol Ester Hydrolase (CEH) 40 1.3.4 Cholesterol Ester Transfer Protein (CETP) 41
1.4 Apolipoproteins 42 1.4.1 Apolipoprotein B 43 1.4.2 Apolipoprotein E 45 1.4.3 Apolipoprotein C 47
1.5 Lipoproteins, Receptors and Atherosclerosis 48
1.5.1 Atherosclerosis 48 1.5.2 Chylomicron Remnants and Atherosclerosis 52
1.5.3 Receptors 53 1.5.3.1 ApoB-100/E(LDL)-receptor 54

7
1.5.3.2 LDL-receptor-related protein (LRP) 65 1.5.3.3 Lipolysis-stimulated receptor 69
1.5.3.4 VLDL-receptor 70 1.5.3.5 Scavenger receptors 70
1.5.4 Lipoproteins, Cholesterol and Atherosclerosis 71 1.5.4.1 Familial Hypercholesterolemia 81 1.5.4.2 Coronary Artery Disease (CAD) 85 1.5.4.3 Type III Dysbetalipoproteinemia 87
1.5.4.4 Diabetes 87 1.5.4.5 Hypothyroidism 88 1.5.4.6 Obesity 89
1.6 Animal Models for Atherosclerosis 89 1.7 Current Screening for Cardiovascular Risk 92 1.7.1 Vitamin A Fat Load Test 94 1.7.2 Apolipoprotein B Assays 104 1.7.3 LDL-Receptor Function 107 1.7.4 Chylomicron-Like Lipid Emulsions 108 1.7.5 Breath Test 109 1.7.6 Other Methods of Assessing Cardiovascular Disease Risk 110
1.8 The Aims of the Project 112
Chapter 2: General Methods and Materials 119
2.1 Animals 119 2.2 Operative Procedures 120 2.2.1 Lymph Duct Cannulation 120 2.2.2 Duodenal Cannulation 120 2.2.3 Collection of Rat Lymph Chylomicrons 121 2.2.4 Chylomicron Separation 121 2.2.5 Preparation of Chylomicron Remnants In Vivo 122 2.2.6 Chylomicron Remnant Separation 123
2.3 Chylomicron-Like Emulsion Preparation 123 2.3.1 Normal Chylomicron-Like Emulsions 123 2.3.2 Modified Chylomicron-Like Emulsions 124
2.4 Thin Layer Chromatography 124 2.5 Triglyceride and Cholesterol Determination 125 2.5.1 Cholesterol Determination 125 2.5.2 Triglyceride Determination 125
2.6 Phosphorus Determination 126 2.7 Clearance Studies 126 2.7.1 Arterial and Venous Cannulation 126 2.7.2 Clearance Studies in Rats 127 2.7.3 Clearance Studies in Rabbits 128 2.7.4 Calculation of Emulsion Clearance and High Affinity (Receptor)
Uptake In Vivo 128 2.8 Organ Removal and Lipid Extraction 129 2.9 Determination of Radioactivity 130 2.10 Laser Light Scattering 130

2.11 Statistical Analysis 130 2.12 Materials 131 2.12.1 Preparation of Aqueous Solutions 131
2.12.2 Radiochemicals 131 2.12.3 Solvents 131 2.12.4 Chemicals 132 2.12.5 Lipids 133 2.12.6 Thin Layer Chromatography (TLC) 133 2.12.7 Cannulae and Tubing 133 2.12.7.1 Arterial cannulae for carotid artery cannulation 133 2.12.7.2 Venous cannulae for jugular vein cannulation 134
2.12.8 Emulsion Preparation Apparatus 134 2.12.9 Fluorescent Emulsion Preparation Apparatus 134
Chapter 3: Clearance Kinetics of Chylomicron-Like
Emulsions In Vivo 135
3.1 Introduction 135 3.2 Special Methods 142 3.2.1 Animals 142 3.2.2 Experimental Procedure for Chylomicron-Like Emulsion Clearance in
Rats 142 3.2.3 Experimental Procedure for Chylomicron-Like Emulsion Clearance in
Rabbits 143 3.2.4 Emulsion Clearance from Plasma 144
3.2.5 Calculations 144 3.2.6 Statistical Analysis 144 3.2.7 Analysis of Emulsion Composition 145
3.3 Results 146 3.3.1 Composition of Chylomicron-Like Emulsions 146 3.3.2 Removal From Plasma of Injected Emulsion Lipids Following
Separate Injection of the T w o Emulsion Types in Rats 147 3.3.3 Organ Uptake of Injected Emulsion Lipids Following Separate
Injection of the T w o Emulsion Types in Rats 151 3.3.4 Removal from Plasma of Injected Emulsion Lipids Following
Simultaneous Injection of the T w o Emulsion Types in Rats 153 3.3.5 Comparison of Emulsion Cholesteryl Ester Uptake Following Separate
versus Simultaneous Injection of the T w o Emulsion Types Emulsion in
Rats 155 3.3.6 Removal from Plasma of Injected Emulsion Lipids Following Separate
Injection of the T w o Emulsion Types in Rabbits 157 3.3.7 Plasma Clearance of Emulsion Particles Following Simultaneous
Injection of the T w o Emulsion Types in Control and W H H L Rabbits
160 3.3.7.1 Plasma lipid concentrations for control and W H H L rabbits 160

9
3.3.7.2 Removal from plasma of injected emulsion cholesteryl oleate following simultaneous injection of the two emulsion types in rabbits
162 3.3.8 Comparison of Emulsion Removal from Plasma Following Separate
and Simultaneous Injection of the T w o Emulsion Types in Rabbits. 165 3.4 Discussion 168
Chapter 4: Characterisation and Analysis of
Chylomicron Remnants 174
4.1 Introduction 174 4.2 Special Methods 180 4.2.1 Preparation of Chylomicron Remnants 180 4.2.2 Extraction of Lipids Using Thin Layer Chromatography 180
4.2.3 Lipid Assays 181 4.2.4 Determination of Particle Size of Chylomicron Remnants 181 4.2.5 T w o Methods of Preparation of Remnant-Like Emulsions 181
4.3 Results 183 4.3.1 Lipid Analysis of Chylomicron Remnants 183 4.3.2 Synthesis of Remnant-Like Emulsion 188
4.4 Discussion 190 4.4.1 Chylomicron Remnant Composition 190 4.4.2 Comparison and Synthesis of Remnant-Like Emulsions 195
Chapter 5: Hepatic Uptake of Chylomicron and
Remnant-Like Emulsions in Mice 197
5.1 Introduction 197 5.2 Special Methods 200 5.2.1 Animals 200 5.2.2 Materials 200 5.2.3 Chylomicron-Like Emulsion Preparation 200 5.2.4 Remnant-Like Emulsion Preparation 201
5.2.5 Operative Procedures 201 5.2.6 Experimental Procedure for the Preparation of Liver Samples 202 5.2.7 Confocal Laser Scanning Microscopy 202
5.3 Results203 5.3.1 Lipid Composition of Fluorescently-Labelled Chylomicron-Like
Emulsions 203 5.3.2 Hepatic Uptake of Normal Chylomicron-Like Emulsion in LDL-
Receptor-Deficient Mice 205 5.3.3 Hepatic Uptake of Modified Chylomicron-Like Emulsion in LDL-
Receptor-Deficient Mice 208

10
5.3.4 Hepatic Uptake of Normal Chylomicron-Like Emulsion in Apo E-
Deficient Mice 210 5.3.5 Hepatic Uptake of Modified Chylomicron-Like Emulsion in Apo E-
Deficient Mice 213 5.3.6 NaCl 213 5.3.7 Lipid Composition of Fluorescently Labelled Remnant-Like Emulsions
216 5.3.8 Hepatic Uptake Following Simultaneous Injection of Normal and
Modified Remnant-Like Emulsions in Mice 217 5.3.9 Hepatic Uptake Following Simultaneous Injection of Modified and
Normal Remnant-Like Emulsions in Mice 220
5.3.10 NaCl 220 5.4 Discussion 223 5.4.1 Patterns of Emulsion Uptake in LDL-Receptor Deficient Mouse Liver
224 5.4.2 Patterns of Emulsion Uptake in Apo E-knockout Mouse Liver 228 5.4.3 Comparison of Patterns of Uptake of Remnant-Like Emulsions
Injected Following Simultaneous versus Separate Injection 232
5.4.4 Conclusion 233
Chapter 6: The Effect of Retinyl Esters on Clearance
Kinetics of Chylomicron-Like Emulsions In
Vivo 235
6.1 Introduction 235 6.2 Special Methods 241 6.2.1 Animals 241 6.2.2 Preparation of Retinyl Esters 241 6.2.3 Preparation of Normal Chylomicron-Like Emulsions 242 6.2.4 Preparation of Modified Chylomicron-Like Emulsions 242 6.2.5 Emulsion Clearance Studies in Rats 243 6.2.6 Clearance Studies in Rabbits 243 6.2.7 Determination of Radioactivity 243 6.2.8 Organ Extraction in Rats 244 6.2.9 Calculations 244 ,6.2.10 Statistical Analysis 244
6.3 Results245 6.3.1 Particle Size of Chylomicron-Like Emulsions 245 6.3.2 Effect of Retinyl Palmitate Incorporation on Plasma Clearance of
Chylomicron-Like Emulsions in Rats 246 6.3.2.1 Clearance of lipids in normal chylomicron-like emulsions 246 6.3.2.2 Clearance of lipids in modified chylomicron-like emulsions 246
6.3.2.3 High affinity uptake 247 6.3.3 Organ Uptake 250 6.3.4 Effect of Retinyl Palmitate Incorporation on Plasma Clearance of
Chylomicron-Like Emulsions in Rabbits 254

11
6.3.4.1 Clearance of lipids in normal chylomicron-like emulsions 254 6.3.4.2 Clearance of lipids in modified chylomicron-like emulsions 254
6.3.4.3 High affinity uptake 255 6.3.5 Effect of Retinyl Stearate Incorporation on Plasma Clearance of
Modified Chylomicron-Like Emulsions in Rabbits 258 6.3.5.1 Clearance of lipids in modified emulsions 258
6.3.6 Effect of Retinyl Myristate incorporation on Plasma Clearance of Normal Chylomicron-Like Emulsions in Rabbits 260
6.3.6.1 Clearance of lipids in normal emulsions 260 6.4 Discussion 262
Chapter 7: Quantitation of Retinyl Esters in
Chylomicron-Like Emulsions 264
7.1 Introduction 264 7.2 Special Methods 267 7.2.1 Animals 267 7.2.2 Preparation of Retinyl Esters 267 7.2.3 Preparation of Normal Chylomicron-Like Emulsions 268 7.2.4 Preparation of Modified Chylomicron-Like Emulsions 268 7.2.5 Clearance Studies in Rabbits 269 7.2.5.1 1 x lipid mass 269 7.2.5.2 3 x lipid mass 269
7.2.6 Emulsion Clearance from Plasma 270 7.2.7 Organ Extraction 270 7.2.8 High Performance Liquid Chromatography (HPLC) 270 7.2.8.1 Properties of retinyl esters 270 7.2.8.2 Materials 271 7.2.8.3 Standard curves and calculations 271 7.2.8.4 Extraction of blood samples 272 7.2.8.5 H P L C instrumentation 272 7.2.8.6 Organ extraction 273 7.2.8.7 Retinyl Ester Calculations 273
7.2.9 Calculations 274 7.2.10 Statistical Analysis 274
7.3 Results 279 7.3.1 Lipid Composition of Chylomicron-Like Emulsions 279 7.3.2 Clearance of Normal Emulsion Lipids and Retinyl Palmitate (1 x lipid
mass) 280 7.3.3 Plasma Kinetics and Emulsion Clearance of Control Normal and
Modified Emulsions (comparison of 1 and 3 x lipid mass) 281 7.3.3.1 Clearance of lipids in normal chylomicron-like emulsions 281 7.3.3.2 Clearance of lipids in modified chylomicron-like emulsions 281
7.3.3.3 High affinity uptake 7.3.4 Organ Uptake of Lipids (comparison of 1 and 3 x lipid mass)

12
7.3.5 Clearance of Emulsion Lipids for Normal and Modified Control Emulsions and Emulsions Containing 2 5 % Retinyl Palmitate (3 x lipid
mass) 287 7.3.5.1 Clearance of lipids in normal chylomicron-like emulsions 287 7.3.5.2 Clearance of lipids in modified chylomicron-like emulsions 288
7.3.5.3 High affinity uptake 288 7.3.6 Efficiency of Retinyl Ester Extraction 291 7.3.7 Detection of Retinyl Palmitate in Plasma Samples Using Retinyl
Acetate as an Internal Standard (3 x lipid mass) 294 7.3.7.1 Clearance of lipids in normal chylomicron-like emulsions 294 7.3.7.2 Clearance of lipids in modified chylomicron-like emulsions 294
7.3.7.3 High affinity uptake 295 7.3.8 Investigation of Alternate Retinyl Esters as Tracees for Chylomicron-
Like Emulsions 298 7.3.8.1 Retinyl stearate as a marker for chylomicron-like emulsions 298 7.3.8.2 Retinyl oleate as a marker for chylomicron-like emulsions 298 7.3.8.3 Retinyl myristate as a marker for modified chylomicron-like emulsions
299 7.3.9 Purity Check and Analysis of Emulsion Fractions for Radiolabeled
Lipids and Retinyl Myristate 300 7.3.9.1 Characterisation of chylomicron-like emulsion 300
7.3.10 Efficiency of Retinyl Ester Extraction 301 7.3.11 Analysis of Emulsion Fractions During Synthesis 301 7.3.12 Clearance of Normal Emulsion Lipids and Retinyl Myristate in Rabbits
304 7.3.12.1 Clearance of lipids in normal chylomicron-like emulsions 304
7.4 Discussion 306
Chapter 8: General Discussion 311
8.1 Introduction 311 8.2 The Use of Chylomicron-Like Emulsions to Monitor Chylomicron
Remnant Metabolism In Vivo 313 8.3 Development of an Alternate Labelling Technique for Chylomicron-
Like Emulsions 315 8.4 Conclusion 317 8.5 Future Directions 318
References 321
Copies of Published Manuscripts 367

13
List of^TaSCes
Table 1.1 Table 3.1 Table 3.2
Table 3.3
Table 3.4
Table 3.5
Table 3.6
Table 3.7 Table 3.8
Table 3.9
Table 4.1 Table 4.2 Table 4.3 Table 4.4
Table 5.1 Table 5.2 Table 5.3 Table 5.4 Table 5.5
Table 6.1 Table 6.2
Table 6.3
Table 6.4
Table 6.5 Table 6.6 Table 7.1 Table 7.2
Table 7.3
Table 7.4 Table 7.5
Table 7.6 Table 7.7
Table 7.8
Composition and Characteristics of Human Plasma Lipoproteins 24 Lipid Composition of Injected Chylomicron-Like Emulsions 146 Chylomicron-Like Emulsion Cholesteryl Oleate and Triolein Removal in Rats Following Separate Injection of T w o Emulsion Types 150 Organ Uptake of Normal and Modified Emulsions Following Separate Injection of the T w o Emulsion Types in Rats 152 Mean Values for Uptake of Chylomicron-Like Emulsion Cholesteryl Ester Following Simultaneous Injection of the T w o Emulsion Types in Rats 154 Comparison of Emulsion Cholesteryl Oleate Uptake Following Separate and Simultaneous Injection of the T w o Emulsion Types in Rats 156 Plasma Clearance of Emulsion Lipids Following Separate Injection of the T w o Emulsion Types into Control Rabbits 159 Plasma Lipid Profile for Control and Homozygous W H H L Rabbits 161 Plasma Removal of Emulsion Cholesteryl Oleate Following Simultaneous Injection of the T w o Emulsion Types in Rabbits 164 Comparison of Plasma Clearance of Emulsion Cholesteryl Oleate Following Separate versus Simultaneous Injection of the T w o Emulsion Types in
Control Rabbits 167 Lipid Composition and Size of Lymph Chylomicron Remnants 184 Lipid Composition of Lymph Chylomicrons and their Remnants 186 Summary of Remnant-Like Emulsion Synthesis 189 Lipid Composition of Lymph Chylomicrons, Chylomicron Remnants (CM-R M ) , and Remnant-Like Emulsions 196 Comparison of Chylomicron-Like Emulsion Lipid Compositions 204 Qualitative Rating of Fluorescent Intensity in Liver Sections 206 Qualitative Rating of Fluorescent Intensity in Liver Sections 211 Comparison of Remnant-Like Emulsion Lipid Compositions 216 Qualitative Rating of Fluorescent Intensity in Control Mouse Liver Sections
218
Particle Size of Chylomicron-Like Emulsions 245 Mean Area Above Curve Data for Normal and Modified Emulsions in Rats
249
Mean Organ Uptake of Normal and Modified Chylomicron-Like Emulsions
in Rats 253 Mean Area Above Curve Data for Clearance of Normal and Modified
Emulsions in Rabbits 257 Mean Area Above Curve Data for Modified Emulsions in Rabbits 259 Mean Area Above Curve Data for Normal Emulsions in Rabbits 261 Lipid Composition of Injected Normal Chylomicron-Like Emulsions 279 Mean Area Above Curve Clearance Data for Normal and Modified
Emulsions in Rabbits 284 Mean Area Above Curve Data for Normal and Modified Emulsions in
Rabbits 290 Extraction Efficiency of Retinyl Esters at Varying Concentrations 293 Mean Area Above Curve Data for Normal and Modified Emulsions in
Rabbits 297 Lipid Composition of Normal Chylomicron-Like Emulsions 300 Distribution of Radiolabeled Cholesteryl Oleate and Retinyl Myristate in Normal Chylomicron-Like Emulsion Fractions 302 Clearance of Normal Emulsion Lipids and Retinyl Myristate in Rabbits... 305

14
List of figures
Figure 1.1 Major pathways for retinoid transport in the body 97 Figure 3.1 Plasma clearance of triglyceride in non-fasted rats injected separately with a
normal emulsion and a modified emulsion 149 Figure 3.2 Plasma clearance of cholesteryl ester in non-fasted rats injected separately
with a normal emulsion and a modified emulsion 149 Figure 3.3 Plasma clearance of cholesteryl ester in non-fasted rats injected
simultaneously with a normal emulsion and a modified emulsion 154 Figure 3.4 Plasma clearance of cholesteryl ester in rats injected separately with normal
and modified emulsions, and in rats injected simultaneously with normal and
modified emulsions 156 Figure 3.5 Plasma clearance of triglyceride in non-fasted control rabbits injected
separately with a normal emulsion and a modified emulsion 158 Figure 3.6 Plasma clearance of cholesteryl ester in non-fasted control rabbits injected
separately with a normal emulsion and a modified emulsion 158 Figure 3.7 Plasma clearance of cholesteryl ester in non-fasted control and W H H L
rabbits following simultaneous injection with a normal and modified
emulsion 163 Figure 3.8 Plasma clearance of cholesteryl ester in control rabbits injected separately
and simultaneously with normal and modified emulsions 166 Figure 4.1 The lipid composition of lymph chylomicron remnants (total lipid mass).. 185 Figure 4.2 The lipid composition of lymph chylomicron remnants (total molar units). 185 Figure 4.3 Size and chemical composition of chylomicrons and their remnant products.
187
Figure 4.4 A comparison of lipid composition of chylomicron remnants 194 Figure 5.1 Laser scanning confocal micrographs of liver sections from mice 207 Figure 5.2 Laser scanning confocal micrographs of liver sections from mice 209 Figure 5.3 Laser scanning confocal micrographs of liver sections from mice 212 Figure 5.4 Laser scanning confocal micrographs of liver sections from mice 214 Figure 5.5 Laser scanning confocal micrographs of liver sections from control mice. 215 Figure 5.6 Laser scanning confocal micrographs of liver sections from control mice. 219 Figure 5.7 Laser scanning confocal micrographs of liver sections from control mice. 221 Figure 5.8 Laser scanning confocal micrographs of liver sections from control mice. 222 Figure 6.1 Plasma clearance of cholesteryl ester in non-fasted rats injected with a normal
emulsion 248 Figure 6.2 Plasma clearance of cholesteryl ester in non-fasted rats injected with a
modified emulsion 248 Figure 6.3 Hepatic uptake following injection of normal emulsions in rats 251 Figure 6.4 Hepatic uptake following injection of modified emulsions in rats 251 Figure 6.5 Splanchnic uptake following injection of normal emulsions in rats 252 Figure 6.6 Splanchnic uptake following injection of modified emulsions in rats 252 Figure 6.7 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a
normal emulsion 256 Figure 6.8 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a
modified emulsion 256 Figure 6.9 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a
modified emulsion 259 Figure 6.10 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a
normal emulsion 261 Figure 7.1 Calibration curves for retinyl acetate, myristate and oleate (area ratio versus
amount ratio) 275

15
Figure 7.2 HPLC chromatogram of standard mixtures of retinyl esters 276 Figure 7.3 H P L C chromatogram of standard mixtures of retinyl esters 277 Figure 7.4 A typical chromatogram of a plasma extract obtained from a rabbit 3.5 min
post-injection of an emulsion containing retinyl palmitate 278 Figure 7.5 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a
normal emulsion 283 Figure 7.6 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a
modified emulsion 283 Figure 7.7 Organ uptake of remnant particles following injection of normal emulsions in
rabbits 286 Figure 7.8 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a
normal emulsion (3 x lipid mass) 289 Figure 7.9 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a
modified emulsion (3 x lipid mass) 289 Figure 7.10 Plasma clearance of normal emulsion lipids in non-fasted rabbits (3 x lipid
mass) 296 Figure 7.11 Plasma clearance of modified emulsion lipids in non-fasted rabbits (3 x lipid
mass) 296 Figure 7.12 A typical H P L C chromatogram of a normal emulsion containing retinyl
myristate 303 Figure 7.13 Plasma clearance of normal emulsion lipids in non-fasted rabbits (3 x lipid
mass) 305

List of9{pn-StandarcCM6reviations
AAC Abbs ANSA Apo Approx. AU BHT BW CETP CHC13 Ci Co. CO CE CAD CHD CM CMRM d DDW DPM EDTA EM ER Etoh FC FCH FFA FH G gm gm/ml hr HDL HEPES HMGCoA HSPG LD. IDDM IDL KBr kDa kg KOH 1 LCAT
Area above curve Absorbance l-amino-2-nahthol-4 sulphonic acid Apolipoprotein Approximately Arbitrary units Butylated hydroxy toluene (2,6-ditert-butyl-p-cresol) Body weight Cholesteryl ester transfer protein Chloroform Curie Company Cholesteryl oleate Cholesteryl ester Coronary artery disease Coronary heart disease Chylomicrons Chylomicron remnants Density Double distilled water Disintegration's per minute Ethylene diamine tetraacetic acid, di-sodium salt Emulsion Endoplasmic reticulum Ethanol Free (unesterified) cholesterol Familial combined hypercholesterolemia Free fatty acids Familial hypercholesterolemia Gauge Grams Grams per millilitre Hour/s High density lipoprotein N-2-Hydroxyethylpiperzine-N-2-ethanesulfonicacid 3-hydroxy-3-methylglutary coenzyme A Heparan sulphate proteoglycans Internal diameter Insulin dependent diabetes mellitus Intermediate density lipoprotein Potassium bromide Kilo Dalton Kilograms Potassium hydroxide Litre Lecithin cholesterol acyl transferase

17
L D L Low density lipoprotein LDL-receptor Low density lipoprotein receptor LPL LRP MeOH m mg ml M Mm min mwt n NaCl ng nm NEFA NIDDM O.D. PC pH PL RM RO RP RS rpm sec S.E.M. TG TO pCi Ug "1 UV UWA VLDL v/v WA WHHL w/v
Lipoprotein lipase LDL-receptor-related protein Methanol Milli Milligrams Millilitre Molar (moles per litre) Millimoles per litre Minute Molecular weight Number Sodium chloride Nanogram Nanometre Non-esterified fatty acid Non-insulin dependent diabetes mellitus Outer diameter Phosphatidylcholine -log [LT] Phospholipid Retinyl myristate Retinyl oleate Retinyl palmitate Retinyl stearate Revolutions per minute Second Standard error of mean Triacylglycerol/triglyceride Triolein Micro curie Micrograms Micro litre Ultraviolet light The University of Western Australia Very low density lipoprotein Volume per volume Western Australia Watanabe heritable hyperlipidemic Weight per volume

18
List of PublishedManuscripts
1. Gennat, H.C., Redgrave, T.G., Croft, K.D. and Mamo, J.C.L. (1997). An emulsion
technique for monitoring high affinity clearance of chylomicron remnants.
Atherosclerosis. 134 (1,2): 339.
2. Mamo, J.C.L., Elsegood, C.L., Gennat, H.C. and Yu, K. (1996). Degradation of Chylomicron Remnants by Macrophages Occurs via Phagocytosis. Biochemistry. 35:
10210-10214.
3. Mamo, J.C.L., Yu, K.C.W., Elsegood, C.L., Smith, D., Vine, D., Gennat, H.C., Voevodin,
M. and Proctor, S.D. (1997). Is atherosclerosis exclusively a postprandial phenomenon?
Clinical and Experimental Pharmacology and Physiology. 24: 288-293.
4. Gennat, H.C, Redgrave, T.G. and Mamo, J.C.L. (1996). Development of an emulsion technique for monitoring chylomicron remnant clearance for application in man. Clinical
and Experimental Pharmacology and Physiology. 24: A45 (Suppl.).
5. Jelinek, G.A., Gennat, H.C., Celenza, A., O'Brien, D., Jacobs, I. and Lynch, D.M. (2001). Community attitudes towards performing cardiopulmonary resuscitation in Western
Australia. Resuscitation. 51:239-246.
6. Lynch, D.M., Gennat, H.C., Celenza, A., O'Brien, D., Jacobs, I. and Jelinek, G.A. (2001). Snakebite and first aid training in Western Australia. Medical Journal of Australia, (in
print).
7. James, A.P., Pal, S. Gennat, H.C., Vine, D.F. and Mamo, J.C.L. (2001). The
incorporation and metabolism of {J-amyloid into chylomicron-like lipid emulsions. htto://www.biomedcentral.com/inteimedia/wa/mediaget/BMC_D,rrEPvMEDIA.SUBMIS
SION/1164318699285396_ARTICLE.PDF
8. Celenza, A., Gennat, H.C., O'Brien, D.L., Jacobs, I.G., Lynch, D.M. and Jelinek, G.A. (2002). Community competence in cardiopulmonary resuscitation. Medical Journal of
Australia, (in print).
9. Darr, J., Gennat, H., Elston, J., Geia, L., Miller, A., Saunders, V. (Jan-Mar 2002). James Cook University: maternal health education program for health workers. Australian
Indigenous HealthBulletin, Vol 2(1). http://www.heallMnfonet.ecu.edu.au/html/html_buUetin/bmletin_home.hto
Conference (Presentations
Gennat, H.C, Redgrave, T.G. and Mamo, J.C.L. Development of an Emulsion Technique for Monitoring Chylomicron Remnant Clearance for Application in Man. Australian
Atherosclerosis Society Annual Conference, 1996
Gennat, HC, Redgrave, TGR, Croft, K. and Mamo, JCL. An emulsion technique for monitoring high-affinity clearance of chylomicron remnants. International Atherosclerosis
Society Conference, 1997
Gennat, H.C, Redgrave, T.G., Light, A. and Mamo, J.C.L. The metabolism of chylomicron-like emulsions in LDL-receptor-deficient and C57BL/6J mice. Australian Atherosclerosis
Society Annual Conference, 1997

19
Chapter 1: Review of Literature
1.1 Introduction
Cardiovascular disease accounts for 41% of deaths in Australia, while CHD alone is
responsible for 2 3 % of all deaths (National Heart Foundation, 1999) and remains the
most prevalent cause of death and disability in the western world (1988).
Atherosclerosis is a degenerative disease of arterial blood vessels and progressively
develops during the entire life of an individual. It begins with the deposition of
cholesterol within the arterial wall (Gown et al, 1986), (Watanabe et al, 1985), and
causes cholesterol deposition and proliferation of arterial smooth muscle cells.
Moreover, atherosclerosis in coronary arteries may predispose them to spasm.
Because developing coronary artery disease (CAD) is clinically silent for many years
and often presents as sudden death, epidemiologists have spent a considerable effort
defining risk factors, traits, or habits that are associated with increased risk in
individuals during the long asymptomatic phase of C A D (Schaefer, 1990).
Elevated low density lipoprotein (LDL) concentrations in fasting blood plasma
have been established as a risk factor for human coronary heart disease (CHD) by
several epidemiological studies (Gordon et al, 1981b), (Castelli et al, 1986), and
clinical and animal studies have suggested that L D L is the source of the cholesterol
found in atherosclerotic lesions. Zilversmit (Zilversmit, 1979) made the initial
assertion that postprandial lipoproteins may also contribute to the pathogenesis of
atherosclerosis. Our laboratory has supported this hypothesis, by demonstrating that
chylomicron remnants can induce cholesterol loading in macrophages in vitro (Mamo
et al, 1996) penetrate the arterial wall and accumulate within the subendothelial space
( M a m o and Wheeler, 1994), (Proctor and M a m o , 1996), (Proctor and M a m o , 1998).

20
Chylomicron and very low-density lipoprotein (VLDL) remnant lipoproteins
are normally cleared from the circulation by the LDL-receptor (Brown and Goldstein,
1979), (Bowler et al, 1991), (Cooper et al, 1982a). Chylomicron remnants utilise apo
E as a ligand (Arbeeny and Rifici, 1984), (Yu et al, 2000), (Plump et al, 1992),
whereas L D L clearance is mediated by apo B-100. Negative feedback via receptor-
mediated uptake of remnant cholesterol allows the concentration of intracellular
cholesterol to be homeostatically controlled; therefore reduced LDL-receptor activity
may result in increased circulating levels of L D L and chylomicron remnants.
However, the LDL-receptor has a higher affinity for apo E compared with apo B-100
(Mahley et al, 1984), subsequently reduced LDL-receptor expression may have a
more deleterious effect on the clearance of chylomicron remnants compared with
LDL.
The clearance of chylomicron remnants was delayed in subjects with familial
hypercholesterolemia (FH), where LDL-receptor activity is impaired or absent (Mamo
et al, 1998a), (Dane-Stewart et al, 2001), (Yu et al, 1997). Individuals with apo E2
homozygosity have severely retarded elimination of chylomicron remnants (Hazzard
and Bierman, 1976), due to reduced interaction of apo E with the LDL-receptor.
Plasma levels of chylomicron remnants have also been shown to be elevated in
subjects with or at risk of, C A D (Groot et al, 1991), (Karpe et al, 1994), (Weintraub
et al, 1996). The risk of early death due to cardiovascular disease in these subjects is
high, as they often display normal plasma concentrations of L D L and triglyceride and
defective remnant metabolism remains undiagnosed.
Early diagnosis of impaired lipoprotein metabolism, accompanied by effective
medical intervention, can reduce the risks of events that cause development of early
atherosclerosis and death. To date, clinical identification of subjects at risk of
developing atherosclerosis has relied on identifying raised plasma concentrations of
L D L , and the contribution of chylomicron remnants has largely been ignored. Ideally,
a diagnostic assay for LDL-receptor function should consider the clearance of
lipoproteins from endogenous and postprandial sources. Several studies have used apo
B-48 and/or retinyl palmitate quantitation to demonstrate delayed clearance of
chylomicron remnants in human subjects (Dane-Stewart et al, 2001), (Smith et al,
1999), (Curtin et al, 1994), (Isherwood et al, 1997), (Meyer et al, 1996). Apo B-48
is an obligatory, specific marker of chylomicron remnants, however it cannot be used
to quantitate receptor uptake of chylomicron remnants in vivo or minor changes in

21
receptor activity, due to inadequate methods of detection and quantitation. Similarly,
the vitamin A fat load test has the disadvantage that it does not provide a measure of
receptor-mediated uptake of remnants via the LDL-receptor. Chylomicron-like
emulsions readily acquire apo E and apo C, necessary for normal uptake via the L D L -
receptor and have been utilised to monitor chylomicron remnant clearance in humans
(Maranhao et al, 1996), (Martins et al, 1995). However, the use of radiolabels as
markers for remnant clearance is not an acceptable marker for use in vivo.
Consequently, the primary aim of this thesis was to develop a diagnostic assay
suitable for quantifying LDL-receptor activity in vivo, by assessing receptor-mediated
uptake of chylomicron remnants. The significance of the project is that it will assist in
the identification of individuals with reduced LDL-receptor expression, therefore
increased chylomicron remnants and attenuated chylomicron remnant clearance,
hence increased risk of C H D . The proposed technique will utilise chylomicron-like
emulsions labelled with retinyl esters.
1.2 Lipoproteins
1.2.1 Lipoprotein Synthesis
Lipids are insoluble in the aqueous environment, so in order to circulate within the
blood compartment, they are packaged as large aggregates called lipoproteins (Gotto,
1983). Lipoproteins have the function of transporting lipids to tissues for energy,
steroidogenesis, storage, and to maintain cell membrane integrity, and are dynamic
particles that are constantly synthesised, degraded, and removed from the plasma
compartment. Lipoproteins are synthesised by the intestine and liver of all mammals
and serve to transport lipids such as cholesterol and triglyceride in ordered-lipid
protein complexes within the plasma, to other sites such as adipose tissue, muscle and
heart.
The structure of the lipoprotein allows the transport of non-polar substances in
the aqueous plasma environment. Compositional analysis and assumptions that
structure is determined by chemical composition leads to the theory that they are
based upon the same biphasic principle as micelles and lipid bilayers (Miller and
Small, 1983b), (Shen et al, 1977). Lipoproteins generally take the form of spherical
microemulsion particles, comprised of an inner core region containing the

22
hydrophobic cholesteryl esters and triglycerides in variable proportions (Miller and
Small, 1987). Surrounding the non-polar cholesteryl esters and triglycerides is an
outer monolayer of the amphipathic (hydrophilic) lipids, cholesterol and
phospholipid. The separation of lipids is not distinct, with core lipids possessing some
solubility at the surface and cholesterol dissolving in the core (Redgrave, 1983). The
charged head groups of the various phospholipid species and the free hydroxyl group
of cholesterol associate with water dipoles in the surrounding aqueous medium while
the hydrophobic fatty acid chains of phospholipids and the sterol ring structure are in
contact with each other and with the hydrophobic core lipids. The principal core
constituent of the largest species is triglyceride, whereas smaller lipoproteins contain
a progressively higher mole fraction of cholesteryl esters (Kane, 1996). Both of these
lipids are exchanged between different lipoprotein classes.
Lipoproteins are highly ordered lipid-protein complexes. A number of specific
proteins (apolipoproteins) have evolved to interact with lipid microemulsion particles
and associate primarily with the surface phospholipid monolayers (Kane, 1996). Most
apolipoproteins bind weakly to the surface, permit lipid binding, direct the catabolism
of the particle (Dolphin, 1985), and function as cofactors for enzymes, structural
components or ligands for receptor removal (Redgrave, 1983).
Chylomicrons are the major class of lipoprotein formed by the intestine, and
are often referred to as postprandial lipoproteins because they transport dietary lipids.
Chylomicrons float at densities less than 1.006 g/ml, the density of plasma. Dietary
lipids, absorbed by the intestine, enter the plasma via the lymph in chylomicrons.
After feeding, lymph becomes opaque and 'milky' from the presence of large
chylomicrons. During times of fasting, the intestine still remains active in producing
chylomicrons, but they are now smaller in size (Redgrave, 1983). Chylomicrons are
hydrolysed in the plasma compartment by endothelial lipases to form chylomicron
remnants.
The liver also produces triglyceride-rich particles from endogenous lipid. Fatty
acyl chains delivered to the liver in the form of lipoprotein triglyceride and non-
esterified fatty acids or fatty acids newly synthesised in the liver can be incorporated
into triglyceride for delivery to peripheral tissues as V L D L . Analogous to
chylomicrons, V L D L are hydrolysed by endothelial lipases to form denser remnant
particles known as intermediate and low-density lipoproteins (IDL and L D L ,
respectively).

23
A n obligatory component of triglyceride-rich lipoproteins are either apo B-100
or apo B-48. The sequence of events in the process of lipoprotein assembly involves
the translocation of apo B into the lumen of the endoplasmic reticulum (ER) where it
associates with the inner leaflet of the E R membrane (Olofsson et al, 1987),
(Alexander et al, 1976). Triglyceride synthesis occurs from cytoplasmic substrates in
association with the membrane. The non-polar triglyceride is deposited within the
hydrophobic environment of the membrane leaflets. A p o B, in association with
triglyceride, buds off into the lumen of the ER. The particle travels through the
secretory network of the Golgi before secretion by a process of reverse pinocytosis
(Borchardt and Davis, 1987), (Sabesin and Frase, 1977). Pulse-chase studies using
human hepatoma cells have shown that V L D L is assembled in the lumen of the E R
via a process in which as the lipid core of the nascent lipoprotein particle was
assembled, apo B was released from the cell membrane (Bostrom et al, 1988).
Lipoproteins are divided into several major classes based on their lipid and
apolipoprotein composition. There are four general thematic roles for plasma
lipoproteins in lipid transport. The first two involve the transport of exogenous and
endogenous triglycerides, respectively, to body tissue. The third is the delivery of
cholesterol to tissues via L D L , and the fourth is the retrieval of lipids from peripheral
tissue via H D L . The plasma lipoproteins vary greatly in biophysical properties.
Chylomicrons are the large triglyceride-rich lipoproteins of enteric origin that carry
exogenous triglycerides into plasma (Kane and Havel, 1994), (Havel and Kane,
1994), whereas V L D L originate in the liver and carry endogenous triglycerides to
similar fates. Lipoproteins consist of a variety of different heterogenous groups of
particles, which are divided into 4 classes due to differences in density, size, chemical
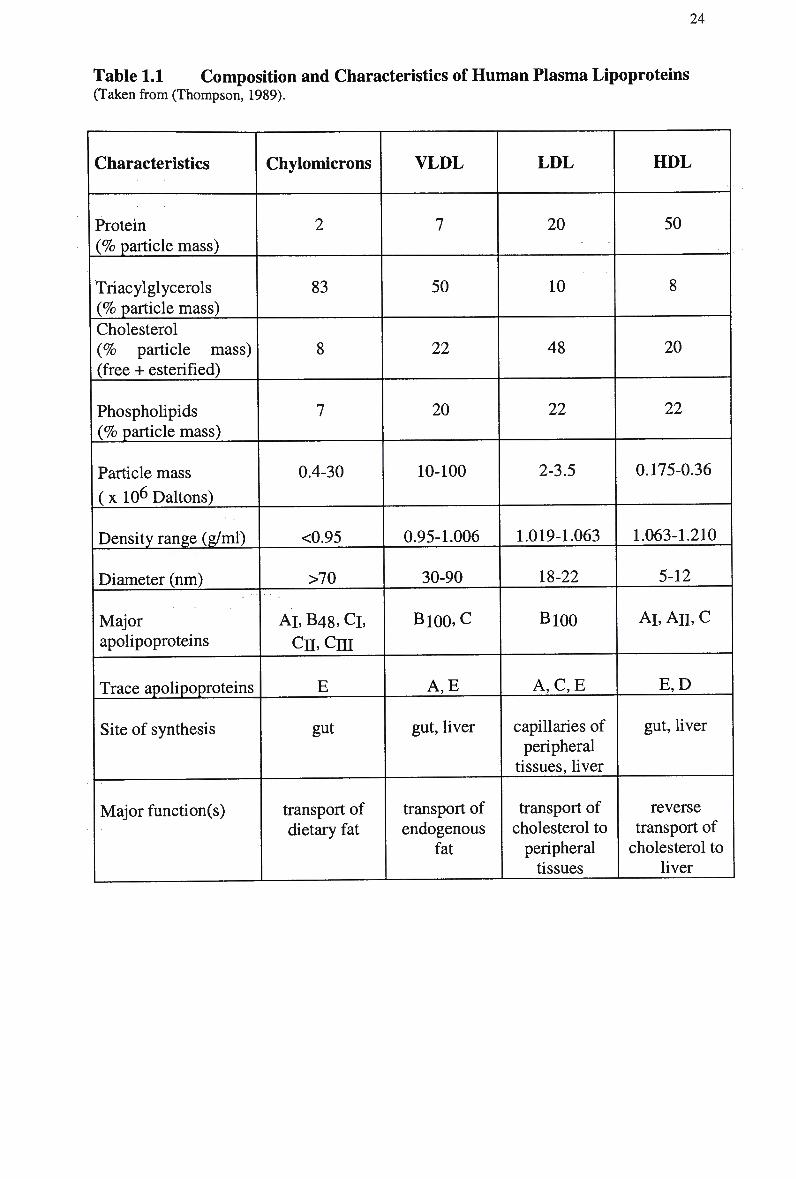
composition and apolipoprotein content (Table 1.1).
24
Table 1.1 Composition and Characteristics of H u m a n Plasma Lipoproteins (Taken from (Thompson, 1989).
Characteristics
Protein (% particle mass)
Triacylglycerols (% particle mass)
Cholesterol (% particle mass) (free + esterified)
Phospholipids (% particle mass)
Particle mass
(x 10 6 Daltons)
Density range (g/ml)
Diameter (nm)
Major apolipoproteins
Trace apolipoproteins
Site of synthesis
Major function(s)
Chylomicrons
2
83
8
7
0.4-30
<0.95
>70
AT, B48, Cl,
cn,cni
E
gut
transport of dietary fat
VLDL
7
50
22
20
10-100
0.95-1.006
30-90
Bl00,C
A,E
gut, liver
transport of endogenous
fat
LDL
20
10
48
22
2-3.5
1.019-1.063
18-22
B100
A , C , E
capillaries of peripheral
tissues, liver
transport of cholesterol to peripheral tissues
HDL
50
8
20
22
0.175-0.36
1.063-1.210
5-12
Al, An, c
E,D
gut, liver
reverse transport of cholesterol to
liver

25
1.2.2 Chylomicrons
1.2.2.1 Synthesis
The major function of chylomicrons is to transport dietary triglyceride and cholesterol
from the intestinal epithelium into the bloodstream, and finally to various cells of the
body (Redgrave, 1999), (Mahley et al, 1981), (Miller and Small, 1987).
Chylomicrons are synthesised and secreted in response to the presence of dietary
intake of fat by adsorptive cells lining the small intestine (Gage, 1920), (Gage and
Fish, 1924.).
The intake of dietary fat in Western civilisation ranges from 50-100g/day and
most of it is in the form of triglyceride and a small fraction consists of cholesteryl
esters and phospholipids. The triglycerides contain fatty acids which can be saturated
or unsaturated. After ingesting a fatty meal, emulsification of dietary lipid begins in
the stomach, but it is in the intestinal lumen where the triglycerides and phospholipids
are hydrolysed to free fatty acid and monoglycerols (Patsch, 1987), (Patsch et al,
1984). In the lumen of the small intestine, the fatty acids liberated by enzymatic
hydrolysis exist as soaps, and in the presence of bile salts form micelles. In this form,
they are presented to the brush border of the epithelial cell membrane and absorbed by
the intestinal mucosa. Within the mucosal cells the fatty acids are reassembled to form
triglycerides, glycerophosphatides and cholesteryl esters and packaged into
triglyceride-rich chylomicron particles by the enterocyte (Patsch, 1987), (Redgrave,
1983), (Imaizumi et al, 1978b).
The fat-soluble vitamins, in particular vitamin A, are incorporated with
cholesteryl esters into the non-polar core of neutral lipid that is surrounded by a coat
of phospholipid, apolipoproteins, and unesterified cholesterol (Patsch, 1987). Once in
the enterocyte, the chylomicrons are thought to be assembled within the Golgi
apparatus of the intestinal mucosa (Assmann, 1982), (Black, 1995), (Herbert et al,
1982). The chylomicron particles are released from the mucosal cells into the
extracellular space when membranes of the trans-cisternae of the Golgi fuse with the
basolateral membranes. Newly synthesised golgi-derived chylomicron particles, with
the associated apolipoproteins are now ready for secretion into the bloodstream
(Dolphin, 1985), (Kane, 1996). Chylomicrons are transported by way of the intestinal
lacteals, the cisterna chyli and transit the thoracic lymph duct to spill ultimately into
blood in the subclavian vein (Kane, 1996), (Assmann, 1982).

26
1.2.2.2 Structure and Composition
Chylomicrons are the largest and least dense of the lipoproteins because they contain
a high proportion of lipid relative to protein. They are heterogenous in size and range
from about 50 to 600 n m (Fraser, 1970), (Redgrave et al, 1993). Their size depends
on factors such as the rate of lipid absorption, the flux of triacylglycerol through the
intestinal cell and the type of dietary fatty acids that predominates in the diet. Thus,
larger chylomicrons are produced after the consumption of large amounts of fat at the
peak of absorption (Redgrave and Dunne, 1975) or when apolipoprotein synthesis is
limiting. W h e n fatty acids are largely unsaturated, the chylomicrons tend to be larger
than when saturated fatty acids are the major fat in the diet. The principal mechanism
by which intestine accommodates large fluxes of triglyceride transport is by
increasing the particle volumes of the chylomicrons secreted, while at lower rates the
smaller sized chylomicrons are the major population of particles (Redgrave and
Dunne, 1975). Thus, the transport of dietary lipid is accomplished principally by
increases in the size of the particles rather than by increases in particle numbers
(Martins et al, 1994), (Fraser et al, 1968).
The lipid composition of the chylomicron is a reflection of the composition of
the diet. For example, a high cholesterol meal will result in relatively small,
cholesteryl ester rich chylomicrons, while a triglyceride-rich meal will result in large,
triglyceride-rich chylomicron particles. The composition of chylomicrons from
animals and man are similar. Chylomicrons are composed largely of triglyceride (75-
90%) and cholesteryl esters (0.5-2%), making them the least dense of the lipoproteins.
The outer surface is composed of phospholipid (6-20%), unesterified cholesterol (0.5-
2 % ) , and proteins (2%). The composition of these lipoprotein particles has been
shown to change with size (Fraser, 1970), (Miller and Small, 1987). Chylomicrons
consist of a hydrophobic oily core, which contains mainly cholesteryl ester and
triacylglycerol with some free cholesterol (Zilversmit, 1965). However, cholesteryl
esters have been shown to be located on the surface of the lipoprotein (Bhattacharya
and Redgrave, 1981), (Janiak et al, 191 A), (Janiak et al, 1979). Studies have shown
that the surface may contain up to 3 % triacylglycerol (Hamilton and Small, 1981).
The chylomicron particles are stabilised by a surface of phospholipids and the
apolipoproteins. The lymph chylomicron contains a complex mixture of proteins,
which are secreted with nascent chylomicrons from the intestine and represent only a

27
small proportion of the total mass of the particle. The major protein of chylomicrons
is apo Al that comprises 38-50% of the chylomicron protein (Imaizumi et al, 1978a),
(Glickman and Green, 1977), (Holt et al, 1979). Lymph chylomicrons also contain
apo C's representing approximately 4 0 % of chylomicron protein. Other middle
molecular weight chylomicron apolipoproteins include apo A-IV and apo E, each
comprising about 10-20% of the total protein (Imaizumi et al, 1978a). The largest
apolipoprotein of chylomicrons is apo B-48, which is an insoluble apolipoprotein with
a molecular weight of 264,000 Daltons and is synthesised in the intestine. There is
one apo B-48 molecule per particle, but comprises approximately 4-5% of the total
chylomicron protein (Kane, 1983). The apolipoproteins A-I, A-II, A-IV, B48 and
small amounts of CII are synthesised on the rough endoplasmic reticulum of the
enterocyte prior to their association with the nascent chylomicron particles.
1.2.2.3 Metabolism
Chylomicrons are metabolised in the circulation by a two-stage process (Mahley and
Hussain, 1991). The first step of chylomicron metabolism involves the hydrolysis of
chylomicron triglyceride and some choline phosphatides within approximately five
minutes by endothelial-bound lipases, forming chylomicron remnant particles.
Second, the liver takes up chylomicron remnants via the interaction of hepatic
receptors with apolipoprotein E, a receptor recognition protein.
Newly secreted chylomicrons are deficient in the C apolipoproteins, however
apolipoproteins E, CI, CII and CDI are obtained either in the lymph or the
bloodstream via transfer from high-density lipoprotein (HDL) particles (Hamilton and
Small, 1981), (Glickman and Sabesin, 1988). Apo Al and ALV are lost from the
surface of these particles (Havel et al, 1973a), (Jeffery and Redgrave, 1982). There is
also an exchange and net loss of surface phospholipids from chylomicrons (Minari
and Zilversmit, 1963), (Redgrave and Small, 1979). Studies have also shown that the
content of free (unesterified) cholesterol in chylomicron particles rises when it enters
the circulation (Miller and Small, 1983b). A rapid redistribution of free cholesterol
has been shown from liposomes with a high free cholesterol/phospholipid ratio to
liposomes with a low free cholesterol/phospholipid ratio (McLean and Phillips, 1981),
(Poznansky and Czekanski, 1979), (Backer and Dawidowicz, 1981). Since
chylomicrons have been shown to have a low free cholesterol/phospholipid ratio, it

28
seems likely that transfer of free cholesterol to these particles occurs in plasma.
Cholesteryl ester transfer protein (CETP) has been shown to be involved in mediating
these exchanges between lipoprotein fractions (Hesler et al, 1987).
O n entry into peripheral capillary beds, chylomicrons come into contact with
an enzyme lipoprotein lipase (LPL); this enzyme is bound to the surface of capillary
endothelium cells of adipose tissue, skeletal and cardiac muscle, and other site (Black,
1995), (Kane, 1996). L P L hydrolyses the triacylglycerols of chylomicrons, producing
fatty acids and glycerol; in this process the apolipoproteins and phospholipids of
chylomicrons are released into the circulation (Mjos et al, 1975), (Redgrave, 1970),
(Redgrave and Small, 1979), (Tall et al, 1979). The adjacent tissue takes up fatty
acids released by hydrolysis of triglyceride for storage or oxidation (Jeffery and
Redgrave, 1982).
As the core of the particle shrinks with the removal of triglyceride, surface
phospholipids are lost by conversion of lysophospholipids or by transfer to other
lipoproteins, particularly the H D L fraction (Chajek and Eisenberg, 1978), (Eisenberg
and Schurr, 1976). Free cholesterol dissolved in the core moves to the surface,
enriching the surface in cholesterol relative to phospholipid (Jeffery and Redgrave,
1982), (Redgrave and Small, 1979). Exchange of apolipoproteins, principally the loss
of apo Al, ATV, CLT and CHI and an enrichment of apo E allows hydrolysis of most of
the remaining triglycerides (Kane, 1996), (Grundy, 1986), (Redgrave, 1970), (Havel
et al, 1973b), (Vigne and Havel, 1981), (McLean and Phillips, 1981), (Jeffery and
Redgrave, 1982). Unesterified cholesterol is also transferred to the H D L fraction of
plasma (Quinn et al, 1982). At least 7 7 % of phospholipid and 3 9 % of protein have
been found to be lost from chylomicrons during lipolysis and transferred to the H D L
fraction of plasma (Redgrave and Small, 1979).
1.2.3 Chylomicron Remnants
1.2.3.1 Synthesis
As hydrolysis of the chylomicron continues and free fatty acids are released, the
triglyceride-rich particle decreases in size and becomes more dense and is termed a
chylomicron remnant (Redgrave, 1983). The kinetics for the removal of chylomicron
triglyceride has been shown to be rapid; the half-life in human subjects is 4.5 min

29
(Grundy and Mok, 1976) and in rats has been calculated to be between 1 and 5
minutes (Harris and Felts, 1970), (Redgrave, 1970). W h e n lipolysis is almost
complete, approximately 70-90% of triglyceride is removed, and the cholesterol-rich
residual chylomicron remnant is released back into the circulation (Redgrave, 1970).
The first evidence of the chylomicron "remnant" was reported by Redgrave
(Redgrave, 1970), who allowed chylomicrons to circulate in functionally
hepatectomised rats and discovered that, as the triglyceride disappeared, a new
particle appeared. W h e n they were injected into intact rats, the chylomicron remnants
were rapidly taken up by the liver. Thus the net result of lipase action is the
production of a lipoprotein core 'remnant' (Redgrave, 1983).
1.2.3.2 Structure and Composition
During the degradation of chylomicrons by LPL, marked changes in surface
chemistry occur. Despite the loss of triglyceride, chylomicron remnants retain
essentially all of the cholesteryl esters (Redgrave, 1988) and acquire cholesteryl esters
in exchange for triglycerides via the action of CETP. The remnant particles are
relatively enriched in cholesteryl esters when compared with the nascent chylomicron
and the ratio of cholesterol to phospholipid contained in the surface area is increased
(Quinn et al, 1982). Chylomicron remnants have been shown to contain
approximately 7 0 % triacylglycerol, 6-7% cholesteryl ester and 5-7% protein (Jeffery
and Redgrave, 1982), however findings from our laboratory suggest that the amount
of triglyceride is approximately 3 5 % (Mamo et al, 1996). During lipolysis,
chylomicrons decrease in size from approximately 500nm to 50nm and increase in
density due to the loss of the loosely packaged core lipids (Redgrave, 1988).
The apolipoproteins present on remnants are apo B-48, E and C (Jeffery and
Redgrave, 1982), (McLean and Phillips, 1981). Chylomicrons and their remnants
contain one molecule of apo B-48 per particle (Martins et al, 1994), (Phillips et al,
1997). Apo B is important in the assembly of the chylomicron, since defective
synthesis of apo B in individual's results in the absence of chylomicrons from their
plasma (Herbert et al, 1982). Investigators have clarified the mechanism of action of
lipoprotein lipases, including the particular role of apo CII as a co-factor (Havel et al,
1973b), (Connelly et al, 1996) and the inhibitory action of all C apolipoproteins on
hepatic uptake of triglyceride-rich lipoproteins (Havel, 1989). Apo E has been shown

30
to be necessary for interaction of chylomicron remnants with hepatic receptors, and
for the transfer of cholesteryl esters from H D L to chylomicrons (Imaizumi et al.,
1978b), (Vigne and Havel, 1981), but remains with the particle as remnants are
formed (Mjos et al, 1975).
1.2.3.3 Metabolism
The chylomicron remnant is the vehicle for the transport of dietary cholesterol and the
metabolism of cholesteryl esters differs dramatically from that of triglycerides. In the
final phase of their metabolism, remnant particles are removed from the circulation by
receptor-dependent endocytosis, almost entirely by hepatic parenchymal cells (Jones
etal, 1984), (Jackie etal, 1991).
Bowler et al. (Bowler et al, 1991) demonstrated that almost all chylomicron
remnants were cleared in vivo by the liver (>90%) via the apo B-100/E (LDL)
receptor. This has also been demonstrated in various systems including perfused liver,
isolated liver membranes, and hepatocytes (Yu et al, 1999), (Barter and Lally, 1979),
(Ha and Barter, 1982), (Mjos et al, 1975), (Nagata et al, 1988), (Tall et al, 1979),
(Tan et al, 1977), (Harris and Felts, 1970), (Linder et al, 1976), (Windier et al,
1980a), (Sherrill and Dietschy, 1978), (Nestel et al, 1963a), (Carrella and Cooper,
1979), (Cooper et al, 1982a). Chylomicrons may also interact with the L D L receptor-
related protein (LRP), heparan sulfate proteoglycans (HSPG) (Mahley et al, 1994),
(Havel, 1998), (Krapp et al, 1996) or other high-affinity uptake processes.
Several studies have indicated that the removal of chylomicron remnants from
the circulation is mediated by the presence of the apo E molecule (Yu et al, 2000),
(Fujioka et al, 1998), (Arbeeny and Rifici, 1984), (Havel et al, 1980), (Sherrill et al,
1980), (Shelbourne et al, 1980), which is recognised by the apo B-100/E receptor
(Brown and Goldstein, 1983). Important to the binding of apo E to the remnant is the
increased ratio of cholesterol to phospholipid in the surface layer of the remnant,
which alters the affinity of the particle for different apolipoproteins. Several studies
have reported that when the remnant has an increased ratio of E to C apolipoproteins,
when compared with the original chylomicron, remnant uptake is facilitated (Havel,
1980), (Redgrave, 1988), (Windier et al, 1980b). In contrast, one study reported that
when reconstituted chylomicrons and chylomicron remnants were decreased in their

31
apo E/apo C ratio from 1.5 to 0.3 for remnants and 0.8 to 0.2 for chylomicrons, uptake
of reconstituted particles was similar to controls (Borensztajn and Kotlar, 1984).
The removal of chylomicron remnant particles appears to be heterogeneous,
with a rapid half-life of less than 5 minutes (Patsch, 1987). Certain pools of remnants
have a very long residence time, while a major proportion leave the plasma
compartment quite rapidly when they are still quite large, i.e. < 75 n m in diameter
(Karpe et al, 1997b). The chylomicron remnants are removed almost as quickly as
they are produced, so there are typically only low concentrations in plasma
(Redgrave, 1988). Studies by Cooper and Y u (Cooper and Yu, 1978) and Sherrill and
Dietschy (Sherrill and Dietschy, 1978) found that the hepatic uptake process for
chylomicron remnants was energy-dependent, saturable, and have a high affinity for
the particles. Following removal from the perfusate, the constituents are degraded
relatively slowly and free cholesterol and amino acids released.
The liver utilises the newly attained cholesterol and cholesteryl esters from the
remnants for conversion to bile acids, which are secreted in the bile as neutral sterol
or incorporated into lipoproteins and released into the plasma as V L D L (Assmann,
1982).
1.2.4 Very Low-Density Lipoprotein (VLDL)
1.2.4.1 Synthesis
The liver, like the gut, synthesises triglyceride-rich lipoproteins and when secreted
into plasma these become V L D L . The main differences between chylomicrons and
V L D L are their site of synthesis and the source of the triglyceride being transported.
The triglyceride and the apolipoproteins of these lipoproteins are synthesised in the
E R and they emerge together with phospholipids and unesterified cholesterol to form
a V L D L particle. Fully lipidated V L D L normally are transported to the Golgi
vesicles, where glycosylation of apolipoproteins proceeds, before the V L D L are
transported to the plasma membrane and released into the space of Disse. The
secretion of triglyceride-rich V L D L provides a pathway by which the liver can export
energy-dense substrate in the form of triglyceride fatty acids and also export
cholesterol, certain exchangeable apolipoproteins, and tocopherol (Kane, 1996) to
extrahepatic cells.

32
The signal for producing V L D L is triggered by the availability of
triacylglycerol for secretion in the lumen of the endoplasmic reticulum. There are two
processes necessary for the assembly of V L D L localised to the ER: translocation of
apo B from the cytoplasmic surface into the lumen, and the addition of lipid to the apo
B nascent chain during its movement into the lumen (Davis, 1997). The production
rate varies greatly, reflecting the supply of disposable energy substrate (Kane and
Havel, 1994) and the rate of apo B-100 secretion (Grundy, 1986). Increases of
triglyceride secretion in V L D L are largely accommodated by increases in particle
volume, though a modest increase in secretion of apo B-100 can take place when
triglyceride secretion is maximal (Pullinger et al, 1989). Genetic alterations in the
apo B gene impair the assembly of V L D L , suggesting that apo B is required for
V L D L assembly and secretion (Young, 1990).
1.2.4.2 Structure and Composition
Apo B-100 is the sole B protein produced in the human liver, and each nascent VLDL
particle contains a single copy of apo B-100 (Kane and Havel, 1994), (Elovson et al,
1988). The protein contains about 40 lipophilic sequences distributed rather evenly
through the molecule that allows it to associate with the particle moieties of V L D L ,
EDL, and L D L with such high affinity that it remains with a single lipoprotein particle
from secretion to endocytosis. Most of the apo B-100 formed in the liver is destroyed
in the endoplasmic reticulum, but its secretion into V L D L is influenced by lipid
metabolism in the liver and it is essential for V L D L assembly and secretion (Lusis et
al, 1987), (Adeli, 1994). In addition to apo B-100, nascent V L D L particles contain
newly synthesised apolipoprotein E and the C apolipoproteins (Grundy, 1986), (Kane,
1996).
V L D L are smaller than chylomicrons, ranging in size from 25 to 100 nm, and
contain less triglyceride but more cholesterol, phospholipid and protein. Smaller
V L D L particles have a lower ratio of apo C: apo B than larger ones. These particles
can undergo lipolysis and the subsequent smaller particles are referred to as V L D L
remnants or IDL.

33
1.2.4.3 Metabolism
As nascent VLDL enters the plasma it acquires apo C as well as cholesteryl esters
from H D L in exchange for triglyceride as catalysed by the CETP. The V L D L particle
undergoes lipolysis by L P L at the surface of endothelial cells and hydrolysis of the
triglyceride results in the formation of a V L D L remnant (IDL). During the course of
lipolysis, the C apolipoproteins are lost from the particle, including apo C-II, the
activator of the enzyme. The loss of other C apolipoproteins removes inhibition of
LPL, allowing hydrolysis of most of the remaining triglycerides (Kane, 1996),
(Grundy, 1986). The circulating half-life of V L D L particles is about 30 to 60 min in
normal humans.
The resultant IDL particles are enriched in cholesteryl esters, and retain a
portion of the original complement of apo E in addition to one copy of apo B-100.
Approximately half of the IDL particles are endocytosed in liver; mediated by the
LDL-receptor (Havel, 1992), which specifically interacts with two protein ligands,
apo B-100 and apo E (Mamo, 1995), (Schneider, 1989). The LDL-receptors have a
higher affinity (20 to 25-fold) for apo E than apo B-100 (Mahley et al, 1984), (Brown
et al, 1981), (Hui et al, 1984a). Although V L D L can interact with the receptor via
apo B-100 to some extent, this tends to be inhibited by the C apolipoproteins. LDL-
receptors can also bind chylomicron remnants, utilising apo E as a ligand (Plump et
al, 1992), and competition studies in perfused liver have confirmed that V L D L
remnants and chylomicron remnants compete for, and share the same hepatic removal
mechanism (Cooper et al, 1982b).
The IDL particles that are not endocytosed may be further processed by
hydrolysis of the surface and core lipids by CETP, whereby hepatic lipase (HL)
progressively removes the rest of the triglycerides and are converted to cholesteryl-
ester-rich daughter particles, LDL.
1.2.5 Low Density Lipoproteins (LDL)
1.2.5.1 Synthesis
The role of LDL is to transport cholesterol to tissues where the cholesterol may be
required for membrane structure or conversion into various metabolites such as

34
steroid hormones. L D L is the major carrier of plasma cholesterol in man, although
this is not so in most other mammals. In animals such as ruminants and some rodents,
most of the cholesterol is transported in high-density lipoprotein (Davis, 1991). The
synthesis of L D L begins when the liver secretes V L D L into the bloodstream. L D L is
then derived from the breakdown of V L D L by LPL. Lipolysis of V L D L produces a
cascade of V L D L intermediates, which contain a progressively lower proportion of
triglycerides and correspondingly richer proportion of cholesterol and phospholipids.
These particles are collectively termed IDL and specifically refer to the intermediate
particles formed during the conversion of V L D L to LDL. Alternately, L D L can be
synthesised directly from the liver.
1.2.5.2 Structure and Composition
LDL differs from its precursor VLDL as it retains only one of the VLDL
apolipoproteins proteins, apo B-100. During the transformation, the apo C and apo E
components are progressively lost. Apolipoprotein B-100 is a large (512 kDa) protein
molecule embedded in the amphipathic coat of the lipoprotein (Brown and Goldstein,
1986), (Cabezas et al, 1994). The apo B-100 appears to be disposed in a
circumferential distribution around the spherical microemulsion particle representing
the lipid core of L D L (Chatterton et al, 1991), (Phillips and Schumaker, 1989),
(Schumaker et al, 1994). Each L D L particle contains one copy of apo B-100, but
each can differ with respect to the amount of bound lipid. Apo B-100 is recognised
and bound by the LDL-receptor on the surface of the cell (Schumaker et al, 1994)
and unlike apo B-48 found on chylomicron remnants, is necessary for hepatic
clearance.
1.2.5.3 Metabolism
The circulating half-life of LDL is approximately 2.5 days in normal humans. The
principal mechanism by which these lipoproteins are removed from blood is
endocytosis into nucleated cells via the LDL-receptor, for which apo B-100 is the
ligand. Endocytosis by hepatocytes accounts for the uptake and degradation of 6 0 % of
L D L that is turned over in the humans (Dietschy et al, 1993). The ligand domain
remains latent in V L D L and IDL of larger particle diameters but then conforms, or is

35
exposed in L D L . Receptor-mediated endocytosis of L D L provides a major source of
cholesterol to cells for the maintenance of cell membranes and plays an important role
in delivery of cholesterol to steroidogenic tissues. Much of the cholesterol supplied to
steroidogenic tissues is thought to be acquired from H D L via SRB1. Receptor number
is increased during cell division and under other circumstances when an increased
supply of cholesterol is required. Once endocytosed, decreasing p H in endosomes
causes dissociation of ligand and receptor. The receptors are returned to the cell
surface, whereas the protein component of L D L is degraded and hydrolysed to amino
acids. Cholesteryl esters are hydrolysed to unesterified cholesterol and free fatty acids
by acid esterase activity, and the cholesterol enters cellular pools. Cholesterol can
then be esterified by A C A T , providing for storage of cholesterol in the ester form
(Kane, 1996), (Lestavel and Fruchart, 1994).
In addition to endocytosis via the LDL-receptor, L D L can be taken up in all
nucleated cells by non-receptor-mediated processes that are of low efficiency but that
become significant as the L D L concentration in extracellular fluid increases greatly,
as in familial hypercholesterolemia. Furthermore, macrophages and transformed
smooth muscle cells can endocytose chemically or physically modified L D L via a pair
of structurally interrelated scavenger receptors (Kodama et al, 1990), (Rohrer et al,
1990). Other receptors residing on macrophages are also capable of removing
oxidised L D L by rapid endocytosis (Stanton et al, 1992), (Endemann et al, 1993).
The synthesis and catabolism of L D L is influenced by both environmental and
genetic factors, such as the type of fat eaten, the mutations in apo B and the mutations
of the LDL-receptor gene.
1.2.6 High Density Lipoproteins (HDL)
Most extrahepatic tissues can synthesise cholesterol de novo; however, the capacity of
extrahepatic tissues to catabolise cholesterol is limited or absent. There is a
mechanism for removing cholesterol from these cells, in order to avoid excessive
accumulation of cholesterol in the extrahepatic tissues. The H D L fraction of the
plasma seem to accomplish this by transporting cholesterol away from the peripheral
tissues and back to the liver for metabolism (Assmann, 1982) and modulating the
surface composition of lipoproteins (Marcel, 1982). The net efflux of cholesterol from

36
tissue or 'reverse cholesterol transport' in effect is thought to inhibit the development
of atherosclerosis (Glomset and Norum, 1973), (Glickman and Sabesin, 1988).
Most H D L particles are found in the density region of 1.063 to 1.21 g/ml
(Mahley et al, 1984) and are typically of 8 to 12 n m in diameter. H D L in the density
region 1.019 to 1.063 g/ml has been classified as H D L 1 . H D L 2 is found in the density
region 1.063 to 1.125 g/ml and the smaller H D L 3 is found in the region 1.125 to 1.21
g/ml (Thompson, 1989), (Oschry and Eisenberg, 1982), (Ha et al, 1984).
Proteins and lipid constituents that form H D L originate in both hepatocytes
and absorptive enterocytes in the intestine, and are initially synthesised as lipid-poor
discs (Hamilton et al, 1986), (Glickman and Magun, 1986) but H D L may also be
formed in plasma during lipolysis. H D L consists of a bilayer composed mainly of
phosphatidylcholine with apo A and apo E at the margins of the disc. O n entering
plasma, they are rapidly converted to spherical H D L by acquiring cholesterol. The
nascent discoidal H D L particle acts as a locus for the action of lecithin: cholesterol
acyltransferase (LCAT), which is the single enzyme responsible for cholesteryl
esterification in the plasma, and the formation of H D L is attributable to L C A T , which
resides on the H D L particles (Hunninghake, 1988). L C A T is plasma-borne and is
secreted by the liver. It is activated primarily by apo A-I, and to a lesser extent by apo
CI, apo E and apo A-IV (Dolphin, 1985), (Czarnecka and Yokoyama, 1995). H D L
can then receive cholesterol from peripheral cells, esterify and transport it to the liver
either directly or by transferring cholesterol to other lipoproteins via C E T P (Barter
and Rye, 1994), (Rajaram and Sawyer, 1996).
Apo A-I also removes cholesterol and phospholipids from cells by an active
transport pathway (Oram and Yokoyama, 1996). As the core of the triglyceride-rich
particle contracts during hydrolysis, the surface phospholipid forms finger-like
projections that bud off from the surface to form disc-like structures (Redgrave,
1983). These structures enter the H D L fraction of plasma and may be hydrolysed by
L C A T and transferred to an adjacent cholesterol molecule, resulting in the formation
of cholesteryl ester and lysolecithin from free cholesterol (Marcel, 1982), (Hamilton
et al, 1986). The hydrophobic cholesteryl esters move from the surface of the H D L to
the core of the particle, which also allows L C A T to receive more free cholesterol
(Mahley, 1990), increasing the size and decreasing the density of the particle. If
L C A T is inhibited (e.g., familial L C A T deficiency), there are only discoidal H D L
particles (HDL3) present in the circulation (Hamilton et al, 1986), (Dolphin, 1985).

37
The H D L 3 particle continues to acquire esterified cholesterol within the core
and consequently becomes larger, with a reduced density. The H D L 3 particle at this
point loses apo A-I and acquires apo E to resemble H D L 2 . Excess cholesterol is
returned to the liver via H D L 2 . The presence of apo E allows the H D L 2 to be
removed by the hepatic apo B-100/E receptor, with the internalised cholesteryl esters
contributing to bile acid synthesis (Dolphin, 1985). One fate of H D L 2 is thought to
involve an exchange protein such as CETP. The cholesteryl esters of the H D L 2 are
postulated to transfer to other lipoproteins and their remnants, a process termed
'forward transport'. This movement of cholesteryl esters is thought to compromise
cholesterol regulation (Fielding, 1992).
Recently, it was shown by Oram et al. (Oram et al, 2000) that lipid-poor H D L
apolipoproteins remove cholesterol and phospholipids from cells by an active
secretory pathway controlled by an A B C transporter called A B C A 1 . The authors
found that c A M P caused increases in apo A-I-mediated cholesterol efflux, A B C A 1
m R N A and protein levels, incorporation of A B C A 1 into the plasma membrane, and
binding of apo A-I to cell surface A B C A 1 . Based on these results, it was concluded
that A B C A 1 is likely to be the cAMP-inducible apolipoprotein receptor that promotes
removal of cholesterol and phospholipids from macrophages. Additionally, mutations
in A B C A 1 impair apo A-I-mediated removal of cellular lipids and cause Tangiers
disease (Lawn et al, 1999), (McNeish et al, 2000), a severe H D L deficiency
syndrome characterised by sterol deposition in tissue macrophages (Assman et al,
1995) and atherosclerosis (Serfaty-Lacrosniere et al, 1994). Therefore this lipid
secretory pathway may play an important role in macrophage lipid trafficking,
lipoprotein metabolism, and protection against atherosclerosis. However, provided
LDL-receptor clearance is adequate, it should promote cholesterol efflux. It is only
when receptor mediated clearance of apo B-containing lipoproteins is compromised
that this process is likely to be short-circuited.
1.3 Lipases and Enzymes
1.3.1 Lipoprotein Lipase (LPL)
A variable portion of plasma free fatty acids comes from the intravascular hydrolysis
of the triglycerides of V L D L and chylomicrons. A high rate of peripheral lipolysis

38
tends to increase F F A uptake by liver, providing fatty acids for oxidation, export of
triglycerides, or ketogenesis, depending on the state of the hepatic substrate economy.
Once chylomicrons and V L D L are secreted into plasma, both marginate in capillaries
where they are subject to lipolysis by lipoprotein lipase, liberating fatty acids that are
chiefly taken up locally. As a result of lipolysis, the particles decrease in size, and the
cores become relatively enriched in cholesteryl esters. The resultant particles are
termed remnants (Kane, 1996).
Lipoprotein lipase is an enzyme necessary for the hydrolysis of chylomicrons
and V L D L triglycerides, with chylomicrons the preferred substrate for L P L in vivo
(Potts et al, 1991). It is a member of a family a lipases that includes hepatic lipase
(HL) and pancreatic lipase. The active sites of all are highly homologous and are each
in the N-terminal domain. The C-terminal domain appears to be required for
appropriate interaction with lipid substrates. Appropriate glycosylation of the enzyme
is also required for activity (Braun and Severson, 1992). Apolipoprotein C-U is an
obligate activator of the enzyme, binding by its C-terminal region to the lipase. The
apolipoprotein CU/lipoprotein lipase system plays a central role in the formation of
remnant lipoproteins, and individuals homozygous for mutations causing deficiency
of apo C-U have severe impairment of removal of chylomicrons and V L D L (Kane,
1996), (Connelly et al, 1987). Premature atherosclerosis can also occur in patients
with familial chylomicronemia as a result of mutations in the L P L gene (Benlian et
al, 1996).
The L P L is synthesised by underlying parenchymal cells in non-hepatic tissues
and is transferred to vascular endothelial cells such as the heart, adipose tissue,
striated muscle, and mammary gland. It moves across endothelium to the luminal
surface in each of these tissues. L P L is bound to the capillary endothelial cell surface
via interaction with cell surface glycosaminoglycans (Dolphin, 1985), (Kane, 1996),
(Mahley, 1990). The suspension of L P L molecules into the vascular lumen by
proteoheparan sulphate molecules attached to the endothelium facilitates the
interaction with chylomicrons and V L D L has also been reported (Scow and
Blanchette-Mackie, 1992).

39
1.3.2 Hepatic Lipase (HL)
Another lipase that participates in the catabolism of triglyceride-rich lipoproteins is
hepatic lipase (HL). H L has been localised primarily to the liver, although it can be
found on other steroidogenic tissues. H L differs from L P L in that it does not require
apolipoproteins for activation, although some do promote activity (Kinnunen et al,
1983). H L is synthesised and secreted by the liver parenchymal cells, and is located
exclusively on the sinusoidal surfaces of the liver endothelial cells, which are thought
to possess receptors which bind H L (Greten, 1982), (Kinnunen et al, 1983). Newly
secreted V L D L and chylomicrons that have not been previously acted upon by LPL,
are poor substrates for H L , which supports the view that H L acts primarily after the
actions of L P L (Dolphin, 1985). H L acts on remnant phospholipids, possibly
changing the conformation or exposure of apo E (Brasaemle et al, 1993).
More recently, Amar et al. (Amar et al, 1998) suggested a role for H L in
mediating the selective uptake of cholesterol from remnant lipoproteins in apo E-
deficient mice, independent of lipolysis. They found that overexpression of inactive
H L in apo E-deficient mice led to reductions in plasma concentrations of cholesterol,
indicating that even in the absence of lipolysis, H L can partially compensate for the
absence of apo E in this animal model. Hepatic uptake of radiolabeled cholesteryl
ester from V L D L was greater in mice expressing either native H L or inactive H L
compared with luciferase controls. The authors concluded that H L serves as a ligand
that mediates the interaction between remnant lipoproteins and cell surface receptors
and/or proteoglycans. One of these pathways may involve the interaction of H L with
L R P or cell surface receptors, such as scavenger receptor, that mediates the selective
uptake of cholesteryl esters.
Hegele et al. (Hegele et al, 1993) showed a delay in the clearance of retinyl
palmitate-labelled chylomicron remnants in HL-deficient subjects, after a fat-load
challenge. Qui et al. (Qui et al, 1998) found that the clearance of chylomicron
cholesteryl ester from the blood was unimpaired in HL-deficient mice, however
chylomicron remnant cholesteryl ester was reduced. Furthermore, endocytosis of
chylomicron cholesteryl esters into liver cells occurred more rapidly in HL-deficient,
than in wild-type mice. The unimpaired hepatic clearance of injected chylomicron
particles in HL-deficient mice demonstrates a critical role for mouse H L in the normal
clearance and processing of chylomicron remnants. Sultan et al. (Sultan et al, 1990)

40
also reported the inhibition of uptake of chylomicron remnant-radiolabelled
cholesteryl ester using an antibody against H L . Reduction or depletion of H L in
isolated perfused rat liver also showed a reduction in the uptake of chylomicron
remnants and triglyceride hydrolysis, however the rate of endocytosis was
significantly increased for those remnants taken up (Shafi et al, 1994). The authors
concluded that H L facilitates the initial uptake of chylomicron remnants in rat liver,
and endocytosis of remnants does not require binding to H L or hydrolysis of remnant
lipids.
1.3.3 Cholesterol Ester Hydrolase (CEH)
Free and esterified cholesterol in liver are both subject to constant turnover. ACAT
esterifies excess intracellular cholesterol with long chain fatty acids (Chang and
Doolitle, 1983), and the resulting cholesteryl esters are utilised for bile and lipoprotein
synthesis or stored in cytosolic lipid droplets and in the membranes of the
endoplasmic reticulum (Hashimoto and Fogelman, 1980). W h e n required,
mobilisation of these esters is brought about by the catalytic action of the neutral
cholesteryl ester hydrolases (CEH) located in the cytosol and in microsomes (Deykin
and Goodman, 1962), (Gandarias etal, 1984).
Soluble cholesteryl esterase has been purified and rather well characterised
(Ghosh et al, 1990), (Ghosh and Grogan, 1991); however, few studies have been
reported on the regulation and function of the microsomal enzyme. The enzyme
displays a circadian rhythm unrelated to the feeding status (Martinez et al, 1991)).
Contradictory reports have appeared with respect to the existence of compensatory
changes in microsomal C E H activity in rat (Stone et al, 1989) and hamster (Ochoa et
al, 1990) in response to altered cholesterol flux to the liver.
It is thought that microsomal C E H may be involved in controlling the
partitioning of cholesteryl esters toward the bile canaliculi or the blood stream and in
maintaining the balance between free and esterified cholesterol in ER. Moreover,
C E H may hydrolyse esterified cholesterol stored in the cytosol (Stokke, 1972),
although the cytosolic C E H is believed to be mainly responsible for this process
(Steinberg, 1976), (Boyd and Gorban, 1980).

41
1.3.4 Cholesterol Ester Transfer Protein (CETP)
CETP is a hydrophobic glycoprotein (Mr 53 000), which acts in plasma to promote
the transfer of cholesteryl esters and triglyceride among different lipoprotein classes.
It is synthesised in the liver, intestine, spleen and adrenal glands. C E T P facilitates the
transfer of cholesteryl ester from chylomicrons, H D L to V L D L , IDL and L D L in
exchange for triglycerides (Barter and Rye, 1994), (Hesler et al, 1987). C E T P is also
responsible for about half of the plasma phospholipid transfer activity (Yen et al,
1989). C E T P therefore plays a central role in the vectorial transfer of cholesteryl
esters from the plasma and peripheral tissues back to the liver for catabolism (Tall,
1993). C E T P has been shown to decrease the core lipid content and size of H D L via
the net mass transfer of cholesteryl esters to V L D L , an effect that may be atherogenic
(Barter et al, 1990), (Agellon et al, 1991).
Patients with a genetic deficiency of C E T P have markedly elevated H D L
cholesterol levels (Koizumi et al, 1991), (Inazu et al, 1990), suggesting a block in
the transfer of cholesteryl ester from H D L to other lipoproteins. Activity of this
protein has been shown to be present in humans and rabbits, but low activity has been
found in rats and mice (Ha and Barter, 1982), (Barter and Lally, 1979). These animals
transport most of their plasma cholesterol in H D L rather than L D L and are relatively
resistant to the development of atherosclerosis. Injection of human C E T P into rats is
associated with a redistribution of cholesterol from H D L to V L D L and L D L (Ha et
al, 1985).
1.3.5 Acyl Coenzyme A: Cholesterol Acyltransferase (ACAT)
Esterification of cholesterol by intestinal A C A T accounts for the cholesteryl ester in
chylomicrons (Martins et al, 1997) and is a catalyst in the synthesis of intracellular
cholesteryl ester (Chang et al, 1997), (Goodman et al, 1965). A C A T also participates
in the secretion of cholesteryl esters assembled in the neutral core of lipoproteins, and
reduced activity is associated with decreased cholesteryl ester in chylomicrons
(Bennett Clark, 1979). More recently, Buhman et al. (Bunman et al, 2000) have
found that mice deficient in A C A T 2 are resistant to diet-induced
hypercholesterolemia and cholesterol gallstone formation, suggesting A C A T 2 has an
important role in the response to dietary cholesterol. Intestinal absorption and hepatic

42
esterification of cholesterol and plasma cholesterol levels were normal in A C A T 1 -
deficient mice, indicating that this enzyme has a limited role in cholesteryl ester
synthesis in the liver and intestine.
1.4 Apolipoproteins
The apolipoproteins of the various lipoproteins regulate lipoprotein metabolism and
therefore determine the unique roles of these lipoproteins. The specific
apolipoproteins may control lipoprotein metabolism by acting either as cofactors or
enzymes, or as ligands for receptors or by maintaining the structure of lipoprotein
particles.
Apolipoproteins associated with human lipoproteins represent several
evolutionary classes. A group of seven with relatively small molecular weights (apo
A-I, A-LT, A-IV, C-I, C-U, C-LTJ, and E) are members of a gene family characterised
by tandem repeats of 11 codons (Luo et al, 1986). Each contains a number of
amphipathic helices than can associate with zwitterionic head groups of phospholipids
of the surface monolayer (Segrest et al, 1992), (Segrest et al, 1994-a). Each protein
binds with relatively high affinity to the lipoprotein particle but has a measurable
solubility in the molecularly dispersed state and is readily exchangeable among
lipoprotein particles (Kane, 1996). Apolipoproteins contain an outer polar face and an
inner hydrophobic core. The outer polar face of the apolipoprotein contains numerous
hydrophobic domains formed by 6-sheets and oc-helices that between the plasma
membrane phospholipid molecules of the outer lipoprotein (Segrest et al, 1994-b),
(Segrest et al, 1994-a), (Young, 1990), (Dolphin, 1985). This is now known to be a
required step in the mechanism of apolipoprotein-mediated cellular lipid efflux, and
the binding ability of the apolipoprotein is critical for efficient membrane micro
solubilization (Gillotte etal, 1999).
Apolipoproteins B and E have been shown to mediate the interaction of
lipoproteins with receptors of the liver and extrahepatic tissues (Mahley and
Innerarity, 1983). The apolipoproteins also function as cofactors for enzymes in lipid
metabolism (Mahley et al, 1984). L P L is activated by apo C-U. The enzyme L C A T ,
which catalyses the esterification of unesterified cholesterol is activated by apo A-I. It
is also suggested that the apolipoproteins may be involved in maintaining the structure

43
of the lipoproteins. In association with phospholipids on the surface of particles, apo
B, Al and E appear to stabilise the micellar structure of the lipoproteins.
The transfer of apolipoproteins C's and apo E to and from triglyceride-rich
lipoproteins exerts profound effects on their capacity to interact with cell receptors
and has a major regulating effect on plasma triglyceride transport and on the remnant
removal processes (Saito et al, 1996). Nascent V L D L and chylomicrons contain
apolipoproteins that are destined to emerge in the protein moieties of H D L . These
proteins, with surface phospholipids and unesterified cholesterol, migrate away from
the triglyceride-rich lipoproteins during lipolysis to become part of the H D L mass.
1.4.1 Apolipoprotein B
The B apolipoprotein group is comprised of two very large proteins, apo B-100 and
apo B-48. Unlike other apolipoproteins, they have extremely high affinity for the
lipoprotein particle and are unexchangeable among lipoprotein particles (Schumaker
et al, 1994), (Kane and Havel, 1994), (Davis, 1999). In man and rat apo B-100 is
formed exclusively in the liver. Apo B-48 is formed only in the intestine in humans,
primates, rabbits and other higher mammalian species but is also synthesised
primarily in the liver in the rat (Krishnaiah et al, 1980), (Lusis et al, 1987),
(Tennyson et al, 1989), (Davidson et al, 1988). Apo B-48 is found in chylomicrons
and is a transcription product of the same gene that produces apo B-100, a protein of
4536 amino acids. Apo B-48, so named because it is completely homologous with the
N-terminal 4 8 % of B-100, results from a tissue-specific editing process that
introduces a stop codon in the transcript. This produces a truncated version of the
protein, which consists of the amino-terminal 2152 residues of apo B-100 (Driscoll
and Cassanova, 1990), (Powell et al, 1987), (Stalenhoef et al, 1984), (Hodges and
Scott, 1992). The activity of the editing factor is high in intestine so apo B-48
accounts for close to 1 0 0 % of apo B produced by this tissue (Young, 1990).
Apo B is important in the assembly of lipoproteins, hepatic apo B-100 in
V L D L biosynthesis and secretion, and intestinal apo B-48 in chylomicron
biosynthesis and secretion (Janiak et al, 191 A). Regulation of apo B synthesis and
secretion by insulin does not occur at the m R N A level (Dashti et al, 1989) and the
m R N A is believed to be constitutively expressed. Regulation of apo B-48 synthesis in
the gut may be under the control of bile salt flux, rather than acute triglyceride flux

44
(Davidson et al, 1986). The protein M T P facilitates the transfer of lipids between
membranes and liposomes, suggesting a role in the assembly of lipoproteins in the
lumen of the E R (Wetterau and Zilversmit, 1986). M T P gene knockout mice have a
marked impairment in the production and secretion of apo B-containing lipoproteins
(Chang et al, 1999), indicating that M T P plays an essential and rate-limiting role in
the secretion of apo B, and may facilitate the translocation of apo B across the E R (Du
et al, 1996). In the absence of sufficient lipid or M T P lipid transfer activity, apo B is
diverted into the ubiquitin-dependent proteasome degradation pathway (Davis, 1999),
(Davis et al, 1989). Specifically, the inability of hypobetalipoproteinemic patients to
absorb and transport triglycerides predicts that a large apo B molecule is essential in
order to assemble large triglyceride-rich lipoproteins (Young et al, 1987).
Findings in H e p G 2 cells that blocked oleic acid stimulated apo B secretion by
inhibiting triglyceride synthesis without affecting the stimulation of cholesterol ester
suggests that cholesterol ester is not rate-limiting for apo B secretion ( W u et al,
1994). More recently, a study by Zhang et al. (Zhang et al, 1999) examined this issue
in primary cultures of hamster hepatocytes. Addition of oleate to medium increased
the mass of triglyceride and cholesterol ester within the hepatocyte and also increased
the secretion of triglycerides, cholesterol ester, and apo B-100 into the medium. The
data indicate a close correlation between the mass of cholesterol ester within the
hepatocyte and apo B100 secretion from it.
N o more than one apo B-48 resides on the chylomicron remnant. Phillips et al.
(Phillips et al, 1997) concluded from studies on E2/E2 individuals, where a remnant
removal defect causes accumulation of intestinally-derived lipoproteins, that apo B
appears unable to transfer among lipoprotein particles containing apo B-48. It seems
as if apo B-48 containing particles are continuously secreted from the enterocyte and
at times of excessive triglyceride availability, lipid droplets fuse with nascent
lipoprotein particles resulting in secretion of enormous chylomicrons (Martins et al,
1994), (Hayashi et al, 1990). Thus the mass of apo B-48 is a measure of the number
of chylomicron particles synthesised by the intestine and entering the lymph.
The second purpose of apo B is to act as a ligand to the LDL-receptor for
delivery of cholesterol to tissues. A p o B-100 has been shown to mediate the
interaction of lipoproteins with apo B-100/E (LDL) receptors of the liver and
extrahepatic tissues (Mahley and Innerarity, 1983). Amino acids 2980 to 3780 of apo
B-100 have been identified as being important for receptor binding which involves

45
clusters of positively charged amino acids. Apo B-48 does not possess the ligand
binding domain so it cannot bind to the LDL-receptor.
The importance of apo B is evident in the hereditary condition of
abetalipoproteinemia, a disease in which patients lack the ability to synthesise and
secrete apo B and consequently have virtually no plasma V L D L or L D L . Apo B in
individuals with this condition is often truncated due to the presence of stop codons.
Other mutations to the apo B gene can result in defective receptor binding regions.
One mutation identified in apo B-100 is at amino acid 3500 where a substitution of
glutamine for arginine can occur resulting in hypercholesterolemia due to raised L D L
levels (Young, 1990).
1.4.2 Apolipoprotein E
Most tissues of rat and man synthesise apo E (Zannis et al, 1985). The intestine
produces small amounts of apo E, but the contribution from this organ is not
considered significant. Nevertheless the liver is the major contributor to plasma apo E
with about 7 0 % of plasma apo E coming from this source (Newman et al, 1985). Apo
E is also synthesised and secreted by several other cell types (Mahley, 1988)). Apo E
is found on chylomicrons, V L D L , remnants and some H D L subclasses. Whether
nascent apo E is secreted on these particles, or becomes associated with them after
secretion, is still controversial (Hamilton et al, 1990), (Hamilton et al, 1991).
Apo E is about 34 000 daltons in molecular weight with several isoforms
possessing carbohydrate and sialic acid. There are three common apo E structural
alleles, namely E-4, E-3 and E-2 (Rail and Mahley, 1992). The phenotypes E4/E4,
E3/E3, E2/E2 are homozygotic whereas E-2/E-3, E-2/E-4 and E-3/E-4 are
heterozygotic. Apo E3/E3 is the most common with around 6 0 % of the population
possessing this phenotype (Mahley et al, 1984). Each isoform is the result of
cysteine-arginine substitutions.
The principle function of apo E is believed to be as a ligand for recognition of
chylomicron remnants by the LDL-receptor, to act as a receptor-mediated uptake
mechanism in the liver and extrahepatic tissues (Mahley et al, 1984), (Plump et al,
1992). Recognition of apo E via the LDL-receptor involves a positively charged
amino acid sequence on the apo E protein made up by a cluster of arginine residues

46
(Yamamoto et al, 1984). Apo E has also been shown to mediate removal of H D L
with apo E (HDL1 and HDLc), V L D L and 0-VLDL (Mahley et al, 1984).
Redgrave (Redgrave, 1999) estimated that large remnants carry five apo E
molecules, whereas small remnants carry only two apo E molecules. Binding to LDL-
receptors requires four apo E molecules, accounting for the slower clearance of small
remnants (Funahashi et al, 1989). Important amino acid residues are at positions 142,
145, 146 and 158. The positively charged lysine and arginine residues at these
positions interact with the negatively charged cysteines on the LDL-receptor (Mahley
et al, 1984) Amino acid substitutions have been shown to affect the receptor binding
capacity of apo E from man. Apo E 2 binds less avidly to the LDL-receptor because of
the substitution with a cysteine at arginine 158. Apo E 4 has cysteine 112 substituted
with an arginine. Apo E also has been shown to modulate the LPL-mediated
processing of triglyceride-rich lipoproteins and can specifically inhibit LPL-mediated
hydrolysis of triglyceride-rich emulsion triglycerides in vivo and in vitro. The arginine
residues in apo E are essential for this effect (Rensen and van Berkel, 1996).
Apo E 2 homozygosity is sometimes associated with type III
hyperlipoproteinemia, which is due to defective interaction of apo E with receptors.
Type in individuals are characterised by hypertriglyceridemia and
hypercholesterolemia (Mahley et al, 1984), and have a severely retarded elimination
of chylomicron remnants (Hazzard and Bierman, 1976). Despite showing normal
binding to LDL-receptors apo E4 is associated with higher than normal plasma
cholesterol (Utermann et al, 1984). Apo E3 has a tendency to associate with H D L in
contrast to apo E4, which preferentially associates with V L D L . The redistribution of
apo E 4 to the V L D L fractions may contribute to the hypercholesterolemia (Steinmetz
et al, 1989). The frequency Of apo E4 in hypercholesterolemic individuals has been
found to be slightly higher than in normolipidemic individuals (Eto et al, 1987).
Cholesterol feeding can sometimes raise the concentration of apo E in plasma
depending on the species (Mahley and Holcombe, 1977) and loading macrophages
with cholesteryl ester increases secretion of apo E up to 10-fold (Basu et al, 1981a).
Whether increasing apo E is beneficial in maintaining homeostasis of lipoprotein
metabolism in unclear. The positive role of apo E in reverse cholesterol transport may
be in delivering cholesterol from peripheral tissues to the liver for excretion. In
abetalipoproteinemia where plasma apo B is lacking, H D L containing apo E may also

47
help deliver cholesterol to peripheral cells of the body (Innerarity et al, 1984). Apo E
may also function to stabilise cholesteryl ester-rich H D L formed by the L C A T
reaction (Mahley et al, 1984).
Yamada et al (Yamada et al, 1992) found that the administration of apo E to
W H H L rabbits prevents the progression of atherosclerosis in this LDL-receptor-
deficient animal model of homozygous FH. Mice generated by gene targeting that
lack apo E develop premature atherosclerosis (Zhang et al, 1992), (Plump et al,
1992), (Breslow, 1993), and Shimano et al (Shimano et al, 1992a), (Shimano et al,
1992b) demonstrated that transgenic mice overexpressing apo E were resistant to
cholesterol-rich diets. These data suggest the antiatherogenic properties of apo E.
1.4.3 Apolipoprotein C
The transfer of apolipoproteins C's to and from triglyceride-rich lipoproteins exerts
profound effects on their capacity to interact with cell receptors and has a major
regulating effect on plasma triglyceride transport and on the remnant removal
processes (Saito et al, 1996).
The most important factors in triglyceride lipolysis and clearance are L P L and
its activator C U (Santamarina-Fojo and Brewer, 1991). Apo CII is necessary as a
cofactor for the action of L P L and apo Cin inhibits activation (Ekman and Nilsson-
Ehle, 1975). Sufficient apo C U is gained by chylomicrons from other lipoproteins by
exchange in plasma or lymph (Redgrave, 1999). Furthermore, apo CII and CHI may
regulate remnant removal by inhibiting uptake by the liver (Windier et al, 1980b),
(Havel, 1980). Apo C U and CUI are mainly associated with H D L during fasting
however during post-prandial lipemia they are transferred to the triglyceride rich
lipoproteins. As the surface phospholipid is lost from chylomicrons and V L D L during
lipolysis, the apo C's are transferred back to the H D L fraction (Weinberg and Spector,
1985), (Schaefer et al, 1982).
Apo CII and CIII are likely to be produced exclusively by the liver (Mahley et
al, 1984), although small amounts of apo CLT and CUI have been shown to be
synthesised by the intestine (Wu and Windmueller, 1979). In hepatocytes, golgi
V L D L were found to be deficient in apo C's compared with serum V L D L , however if
secretion was delayed, more apo C's associate with the nascent V L D L . Most apo C's
were acquired during or after secretion (Nestruck and Rubinstein, 1976).

48
Lipoprotein concentrations of apo CII and apo CIII in V L D L and H D L are
independently regulated. A p o CII and apo CIII levels increased in V L D L as apo B
and triglycerides increased, but the rate of increase of apo CIII was faster than apo
CU. In subjects with hypertriglyceridemia, VLDL-apo CIII was enriched on average
two-fold per V L D L particles compare with apo CII (Le et al, 1988). It was suggested
that denser V L D L type particles bind more apo CIII and that denser particles are more
prevalent in mild hypertriglyceridemia.
Increased ratios of apo CIII-2 to apo CIII-1 have been found in some patients
with chronic renal failure or severe hypertriglyceridemia (Holdsworth et al, 1982).
The triglyceride-rich lipoproteins in these patients were not lipolysed as efficiently as
normal lipoproteins. In another study (Kashyap et al, 1981), some type V
hyperlipoproteinemic individuals contained less apo CHI-0 and more apo CIII-1 in
their triglyceride-rich lipoproteins compared with type IV and normolipaemic
individuals. A p o CJJJ-2 was similar in all three groups.
1.5 Lipoproteins, Receptors and Atherosclerosis
1.5.1 Atherosclerosis
Atherosclerosis is characterised by the progressive formation of plaques on the
arterial wall, which inhibit the flow of blood. The arterial plaques consist of a local
accumulation of cholesterol and fibrous tissue in the intima of the artery, and are also
accompanied by structural changes in the media and thickening of the arterial wall.
The deposition of cholesterol in the subendothelial space of the arterial wall initiates
the formation of an atherosclerotic lesion (Simionescu et al, 1986). This lipid is now
known to be nearly pure cholesteryl ester (Goldstein and Brown, 1977), (Brown and
Goldstein, 1986).
Central to the pathogenesis of atherosclerosis are an abnormally functioning
endothelium and a consequent loss of vascular integrity. The cellular constituents of
"proliferative lesions", the early stage of atherosclerosis, have been identified as
functionally altered endothelial cells, differentiating macrophages, activated T-
lymphocytes, and proliferating smooth muscle cells (Ross et al, 1990). Each of these
cell types is able to form distinct sets of biologically active molecules upon
appropriate stimulation conditions in vitro (Habenicht et al, 1994). The first

49
discernible changes in the arterial wall in experimental atherogenesis are the
adherence of mononuclear cells to the intact endothelium and their subsequent
transendothelial migration into the intima of the arterial wall (Gerrity et al, 1985),
(Jonasson et al, 1986), (Ross et al, 1990). Mononuclear cell become activated before
or concomitant with their migration into the subendothelial space, and begin to form
and secrete molecules with potent biological activities.
T w o schools of thought have emerged about the biochemical and
physiological basis underlying the early stages in atherogenesis. These two
hypotheses have generally been termed the "endothelial injury hypothesis" and the
"lipid infiltration hypothesis" (Steinberg, 1988). It is not clear which process occurs
first, or the relationship between the two. According to the classical "lipid infiltration
hypothesis", lipoproteins, especially L D L , infiltrate the arterial intima prior to
endothelial cell damage (Small, 1977) (Steinberg et al, 1989), (Ross, 1993a),
(Ginsberg, 1994). The major initiator of atherogenesis in this case is the uptake of
lipid, which does not involve cell damage of the arterial wall. Endothelial cells,
smooth muscle cells and monocyte macrophages in the intima appear to accumulate
lipids, particularly cholesterol, from L D L particles and possibly from other
lipoproteins such as chylomicron remnants, V L D L and 6-VLDL. This implies that
circulating cholesterol, principally carried in L D L , is central to the atherogenic
process, and hence atherogenesis does not progress at lower plasma levels (Steinberg,
1988).
The lipid infiltration hypothesis has been further extended as recent evidence
has emerged which states that the oxidative modification of L D L (or other
lipoproteins) is important and possibly obligatory in the pathogenesis of the
atherosclerotic lesion. It appears that macrophages can only accumulate L D L that has
been modified by oxidation (Ox-LDL). L D L is first oxidatively modified by
macrophages or cells such as smooth muscle or endothelial cells in the arterial wall
(Ross, 1993b). The oxidation process is thought to start with the polyunsaturated fatty
acids of phospholipids on the surface of L D L , and propagate to the cholesterol moiety
and apolipoprotein B. This process stimulates the expression of leukocyte adhesion
molecules such as vascular cell adhesion molecule, and the release of a variety of
substances by the endothelial cells. As a result, monocytes adhere to the endothelial
surface and are further stimulated to migrate into the subintima. Once the monocytes
enter the subintima, they ingest the O x - L D L and become activated macrophages that

50
further oxidise the L D L molecules. The resulting cells, called foam cells, are the basis
of fatty streaks. The activated macrophages also release cytokines, which cause a
further proliferation of macrophages and stimulates smooth muscle cells to release
growth factors, which induce both migration and proliferation of the smooth muscle
cells. T lymphocytes also accumulate in the fatty lesion and contribute to the chronic
inflammatory process of atherosclerosis (Dzau, 1994).
Because cholesterol feeding in animal models of atherogenesis leads to rapid
alterations in arterial wall morphology that are similar to atherosclerotic lesions
observed in humans, it has long been suspected that the direct interaction of
lipoproteins with the arterial wall causes injury to the monolayer of endothelial cells,
and may initiate plaque formation (Habenicht et al, 199A). It was on the basis of
cholesterol-feeding studies in monkeys that Ross and Glomset (Ross and Glomset,
1976) proposed a detailed hypothesis of the pathogenesis of atherosclerosis, or
"response to injury" hypothesis. The initial damage could stem from various causes,
such as smoking or elevated blood pressure, or cholesterol. Loss of the structural
integrity of the endothelium may cause the release of platelet-derived growth factors
that could stimulate smooth muscle cell proliferation and secretion of other growth
factors. Thus, repeated episodes of endothelial damage and smooth muscle cell
proliferation can lead to the development of lesions (Ross and Glomset, 1976), (Ross,
1981a).
Injury to the endothelium is thought to allow increased penetration of
lipoproteins into the arterial wall and to promote the adherence of monocytes to the
endothelial areas of damage. Monocytes are attracted to chemokines in the arterial
wall to scavenge cholesterol and subsequently transform into macrophages (Tsukada
et al, 1986), (Filip et al, 1987). Macrophages are then able to accumulate cholesteryl
esters and transform into 'foam cells', which are the most distinct histological feature
of early atherosclerotic regions. Cholesterol accumulation is the result of excess
uptake of lipoprotein-derived cholesteryl ester, which has localised in subendothelial
spaces. This 'fatty streak' formation leads to advanced proliferative fibrous plaque
and atherosclerosis.
Macrophages are thought to play an important role in the formation of the
atherosclerotic lesion, particularly the fatty streak. The process which accounts for the
generation of macrophage-derived foam cells in not known, however there is much
support and evidence for the oxidation of L D L and the subsequent removal of this by

51
macrophages (Steinberg et al, 1989). Prolonged elevation of chylomicron remnants
in the circulation due to a delay in their removal could promote their entry into the
artery wall and contribute (directly) to lesion formation by modulating cellular
activities or by serving directly as a source of cholesterol.
In general, macrophages appear to be a tissue site of uptake of chylomicron
remnants and, with delayed removal by liver, increased macrophage uptake can be
expected (Fujioka et al, 1998). Native chylomicron remnants have been shown to
induce cholesterol loading in macrophages ( M a m o et al, 1997), (Van Lenten et al,
1985), (Yu and M a m o , 2000), suggesting that macrophage foam cells in
atherosclerotic plaques might be derived from the cellular uptake of chylomicron
remnants. Monocytes and macrophages have several pathways that could mediate
chylomicron remnant uptake, including the LDL-receptor, V L D L receptors, LDL-
receptor-related protein (LRP), scavenger receptors and by means of phagocytosis
(Ellsworth et al, 1990) (Van Lenten et al, 1985), (Sakai et al, 1994), (Nakazato et
al, 1996), (Hiltunen et al, 1998), (Floren and Chait, 1981), ( M a m o et al, 1996).
Atherosclerosis is now believed to be a complicated interaction of endothelial
cell damage, lipoproteins and their interactions with the cells and matrix proteins of
the arterial wall, thus atherogenesis is a multifactorial disease (Witzum, 1994). The
result is the unification of both hypotheses, to account for transition from fatty streak
to an advanced lesion (Steinberg 1988).
Major risk factors for atherosclerosis include high levels of L D L cholesterol
(> 4.1 mmoLl"1), H D L cholesterol level < 0.9 mmoLL 1 , family history of C A D in a
first degree relative prior to age 55 years, diabetes mellitus, hypertension, cigarette
smoking; obesity (> 3 0 % of ideal body weight), and presence of peripheral vascular
disease (Panel, 1988). The reason for the accumulation of cholesterol within these
plaques is no doubt multifactorial, however there is strong evidence to suggest that
abnormal lipoprotein metabolism is involved in this process.
The steady state concentration of LDL-cholesterol in the plasma is thought to
greatly increase risk of developing atherosclerotic disease. The organ most closely
linked to the development of atherosclerosis is the liver. The liver is the major site of
production and removal of the lipoproteins involved in atherogenesis and, therefore, it
plays a key role in regulating whole body cholesterol balance and cholesterol
trafficking. The liver is the key organ in control of this balance not only because it
harbours the majority of the LDL-receptors in the body, but also because it is

52
responsible for a large part of the cholesterol synthesised within the body (Havel,
1988). The concentration of cholesterol in hepatocytes seems to be a key factor in
influencing the production of LDL-receptors, and these receptors are tightly regulated
to maintain an optimum cellular content of cholesterol (Grundy, 1991). Consequently,
synthesis of LDL-receptor can be upregulated or downregulated (Brown and
Goldstein, 1986) exclusively at the level of transcription.
1.5.2 Chylomicron Remnants and Atherosclerosis
In recent years, an increasing body of evidence has accumulated that supports a causal
role of chylomicrons and their remnants in the development of atherosclerosis and
C A D (Mamo et al, 1997), (Yu and Cooper, 2001), (Proctor and M a m o , 1996),
(Dallongeville and Fruchart, 1998), (Meyer et al, 1996), (Slyper, 1992), (Steiner,
1993). Despite the focus on L D L as the major vehicle for cholesterol transport in
circulation, chylomicron remnants can carry up to 15 grams of cholesterol per day
(Mamo, 1995). Once chylomicrons undergo hydrolysis in circulation, they become
cholesterol-enriched remnants (Redgrave, 1983). It is the triglyceride-depleted
remnants that are considered to be atherogenic, because they are able to penetrate
arterial tissue and are preferentially retained within the subendothelial space (Proctor
and M a m o , 1998), ( Mamo and Wheeler, 1994). Additionally, chylomicron remnants
can induce substantial macrophage lipid loading (Mamo et al, 1997). O'Brien et al.
(O'Brien et al, 199A) have also suggested that chylomicron remnants are retained in
arterial lesions, after finding apo E localised in atherosclerotic lesions. The uptake,
penetration and retention of cholesterol-rich chylomicron remnants in the
subendothelial space of the arterial wall may play a significant role in initiating the
formation of an atherosclerotic lesion (Williams and Tabas, 1995).
It has been suggested that chylomicron remnants are direct initiators of
atherosclerosis (Zilversmit, 1979), (Karpe and Hamsten, 1995), (Davignon and Cohn,
1996). This hypothesis is supported by a number of elegant studies that have found a
correlation between the delayed clearance of chylomicron remnants and an increase in
the risk of developing atherosclerosis (Groot et al, 1991), (Patsch et al, 1992),
(Karpe et al, 1994), (Weintraub et al, 1996), (Redgrave, 1999). Chylomicron
remnant dyslipidemia has been identified in a number of primary and secondary lipid
disorders ( M a m o et al, 1998a), (De M a n et al, 1996), (Tomono et al, 1994). It is

53
therefore likely that chylomicron remnants do contribute at least, to the formation of
'foam cells' and atherosclerotic plaque.
In order for massive accumulation of cholesteryl esters to occur in arterial
lesions, there are a number of precursors. First, the level of circulating lipoproteins
must be raised. This could be a result of overproduction of endogenous lipoproteins,
or increased plasma concentration of circulating exogenous lipoproteins. Second, the
macrophages in the arterial lesions must have in place mechanisms by which the
uptake of lipoprotein is mediated and are probably not regulated by intracellular
cholesterol concentration, including the LRP, scavenger receptors, and the V L D L -
receptor. Alternately, receptors such as the LDL-receptor that are regulated by
intracellular cholesterol concentration may be downregulated, defective or their
binding capacity reduced, resulting in an attenuation of normal uptake mechanisms,
and greater emphasis placed on alternative pathways.
1.5.3 Receptors
Normal mammalian cells obtain cholesterol either by producing it intracellularly or by
taking it in from the environment as lipoproteins. Cellular cholesterol requirements
are met by endogenous cholesterol synthesis from acetate. The rate-limiting enzyme
in this pathway is 3-hydroxy-3-methylglutaryl coenzyme A ( H M G - C o A ) reductase,
and the activity of this enzyme is increased when cells are actively synthesising
cholesterol (Bilheimer, 1987). The liver removes a large majority of lipoprotein
remnants where the accessibility of the liver to macromolecules owing to the large
pores of the hepatic sinusoids (Redgrave, 1970) and the fenestrated vascular
endothelium (Greeve et al, 1988) permit their passage. The parenchymal cells of the
liver account for the majority of remnant uptake (Jones et al, 1984).
Cholesterol transport is thought to involve two major pathways (Brown and
Goldstein, 1983). The endogenous pathway starts with the production of V L D L by the
liver. V L D L enters plasma and is metabolised, generating progressively smaller
particles, which are usually called IDL (Bierman and Glomset, 1985). IDL may either
be taken up in the liver via the LDL-receptor or converted to L D L , which contain a
major portion of the cholesterol in plasma. L D L is then taken up either by L D L -
receptors in tissues throughout the body or by non-receptor-mediated processes
(Brown and Goldstein, 1983). Early studies have shown that chylomicron remnants

54
are removed by a saturable, high affinity process (Sherrill and Dietschy, 1978), and
the initial event could be binding of the particle to a receptor on hepatic membranes
(Carrella and Cooper, 1979). However there is still much debate regarding the specific
mechanism responsible.
Bierman and Glomset (Bierman and Glomset, 1985) proposed that
chylomicron remnants were taken up in the liver by the chylomicron remnant
receptor, which is thought to be identical to the apo E receptor. This exogenous
pathway is responsible for delivering dietary cholesterol to the liver where it can be
incorporated into other important metabolic pathways, including the one for
endogenous cholesterol transport. However, the clearance of chylomicron remnants in
humans is a complex mechanism still subject to debate. The initial removal of
chylomicron remnants by the liver appears to involve interaction with several
macromolecules on the cells surface, including, heparin-sulfate-bound H L and apo E
(Havel, 1996). Chylomicron remnant clearance is mediated via interaction of apo E
with specific receptors (Windier et al, 1980a) and subsequent non-receptor binding to
the cellular surface in the space of Disse.
There are several candidate receptors proposed to be involved in the clearance
of chylomicron remnants, namely the LDL-receptor, which binds the chylomicron
remnant with high affinity (Yu et al, 2000), (Mamo et al, 1991), (Jackie et al, 1992),
(Bowler et al, 1991), (Beaumont and Assadollahi, 1990). Other proposed
mechanisms include the LRP, or a2-macroglobulin receptor (Hussain et al, 1991),
(Herz et al, 1988), (Beisiegel et al, 1989), heparan sulfate proteoglycans, a hepatic
remnant receptor (Van Dijk et al, 1991), an asialoglycoprotein receptor (Windier et
al, 1991), and a lipolysis-stimulated receptor (Mann et al, 1995).
The receptors suggested to play a role in the uptake of chylomicron remnants are
discussed.
1.5.3.1 Apo B-100/E (LDL)-receptor
The first lipoprotein receptor to be characterised was the LDL-receptor. It was
identified in 1977 as a specific, saturable and high affinity mechanism (Goldstein and
Brown, 1977), using cultured human fibroblasts. LDL-receptors are present within
clathrin-coated pits located on the plasma membrane of a wide variety of tissues, such
as fibroblasts, smooth muscle cells and the liver (Dolphin, 1985). Internalisation of

55
lipoproteins enables the cells to obtain cholesterol, and involves the degradation of the
lipoprotein via several cellular organelles in a sequence that was called the L D L -
receptor pathway (Brown and Goldstein, 1986). This complex lipoprotein-dependent
metabolic pathway is characterised by high-affinity binding of several lipoprotein
classes to the LDL-receptor, endocytotic uptake of the lipoproteins in coated pits, and
the formation of secondary lysosomes. Receptor synthesis and expression is finely
regulated by the intracellular cholesterol concentration, various hormonal factors and
dietary intake of cholesterol and other fats. The discovery of the LDL-receptor has
had a profound impact on cell biology, clinical investigations, therapeutics, and on the
study of atherosclerosis in general (Brown and Goldstein, 1986), (Steinberg, 1988).
The LDL-receptors are synthesised in the rough endoplasmic reticulum and
initially have an apparent molecular weight (Mr) of 120,000 Daltons. They are
transported to the Golgi apparatus where they are converted to a mature form with an
apparent M r of 160,000 Daltons by the addition of carbohydrate (Gianturco and
Bradley, 1987). The LDL-receptor protein contains 839 amino acids, with the gene on
chromosome 19. It contains 8 repeat units of a cysteine rich amino acid sequence; apo
B-100 and apo E proteins have many lysine or arginine residues, respectively. The
structure of the LDL-receptor is a single-chain glycoprotein with five distinct
domains, and is expressed in almost all tissues of the body in mammals (Schneider et
al, 1982), (Van der Westhuyzen et al, 1990).
Following synthesis, the receptor proteins are initially located randomly in the
cell membrane but they migrate to specialised regions that contain clathrin-coated pits
where they cluster by virtue of their cytoplasmic segment. About 8 0 % of the L D L -
receptors are concentrated in these coated pits, which cover about 2 % of the cell
surface. The receptor carries a binding site that protrudes from the cell surface. W h e n
the L D L binds to these receptors, the resulting lipoprotein and receptor complexes are
internalised by invagination of the coated pit to form endocytotic vesicles or
endosomes (Brown and Goldstein, 1975), (Hussain et al, 1999). W h e n the
cytoplasmic segment is absent, the receptor can bind on the surface, but it cannot slide
laterally into the coated pits or take L D L into the cell (Brown and Goldstein, 1975),
(Brown and Goldstein, 1986), (Goldstein and Brown, 1977). The endocytotic vesicles
enter the cytoplasm where their acidification is believed to promote separation of the
L D L from its receptor. The receptor then recycles back to the plasma membrane by

56
clustering with other receptors in a segment of the endosomal membrane, which
pinches off to form a recycling vesicle. Once it reaches the surface the receptor can
bind to another L D L particle and initiate another round of internalisation. The L D L -
receptors on the cell surface internalise once every 10 minutes, whether or not it is
occupied by a lipoprotein. LDL-receptors can support up to 150 passages in acidic
endosomes without losing its functionality, and are degraded with a half-life of 12
hours (Lestavel and Fruchart, 1994), (Brown and Goldstein, 1975), (Goldstein et al,
1976), (Basu et al, 1981b). After its release from its receptor, the lipoprotein is
segregated into vesicles, which fuse with lysosomes. In the lysosomes, acid lipases
hydrolyse the L D L and its components, liberating unesterified cholesterol, which
enters the cellular cholesterol pool. In vivo, the main task of LDL-receptors is to
supply the cells with cholesterol, thereby mediating the removal of cholesterol-rich
lipoprotein particles from the bloodstream.
W h e n adequate amounts of cholesterol are available to the cell, a series of
intracellular adjustments occurs to avoid excess accumulation of intracellular
cholesterol. The presence of cholesterol within the lysosomes mediates negative
feedback control on H M G - C o A reductase and other enzymes of the cholesterol
biosynthetic pathway, which turns off cholesterol synthesis by the cell. Additionally,
synthesis of LDL-receptors is inhibited which suppresses intracellular cholesterol
synthesis and prevents overaccumulation of cholesterol via the receptor pathway. As
the L D L receptors are responsible for L D L removal, an excess of cellular cholesterol
results in the suppression of receptor synthesis. The enzyme A C A T is activated by
cholesterol, which re-esterifies excess cholesterol to enable the cell to store
cholesterol in the cytoplasm as cholesteryl ester droplets (Brown et al, 1980),
(Habenicht etal, 1994), (Brown and Goldstein, 1975), (Lestavel and Fruchart, 1994).
Because the concentration of L D L in plasma and lymph is much greater than
its dissociation constant at the receptor, modification of L D L cholesterol clearance is
achieved by regulating the number of cell surface L D L receptors (Fielding, 1992).
Having such precise feedback suppression allows individuals to maintain a relatively
constant plasma cholesterol level, as changes in cholesterol demand are reflected in
the liver by changes in receptor expression. Therefore the intracellular cholesterol
level maintains cellular cholesterol balance by regulating LDL-receptor activity which
in turn plays a protective role by ensuring that the concentration of cholesterol in
blood is sufficiently low to avoid the build up of atherosclerotic plaques (Habenicht et

57
al, 1994), (Brown et al, 1981), (Brown and Goldstein, 1986). Via the LDL-receptor-
mediated regulatory system, the cell is able to coordinate the utilisation of intra- and
extracellular sources of cholesterol, and for cholesterol to be delivered to peripheral
cells for cell metabolism. Human fibroblasts and other mammalian cells in culture are
able to survive in the absence of a large amount of lipoproteins because they can
synthesise cholesterol from acetyl-CoA (Brown and Goldstein, 1979).
Due to close regulation of the LDL-receptor by the intracellular cholesterol
levels (Brown and Goldstein, 1975), it is unlikely to be involved in the accumulation
of cholesteryl esters in macrophages found in arterial plaque. Angelin et al. (Angelin
et al, 1983) has reported that uptake of chylomicron remnant-derived cholesterol in
the liver reduces both hepatic synthesis of cholesterol, and LDL-receptor activity. In
addition, it is expressed in arterial lesions in very low numbers (Hiltunen et al, 1998).
Brown and Goldstein (Brown and Goldstein, 1984) have previously reported that
when LDL-receptor function is inappropriately diminished, the protective mechanism
is lost, cholesterol builds up in plasma, and atherosclerosis ensues. Alternate receptors
directly involved in the excessive accumulation of cholesterol in macrophages may
assume a greater role to compensate for LDL-receptor function, and potentially
increase the quantity of chylomicron remnants entering the arterial tissue.
A n unresolved question in lipoprotein metabolism is whether the LDL-
receptor is a mechanism by which chylomicron remnants are removed from the
circulation (Mahley et al, 1989). If hepatic LDL-receptor expression or binding were
compromised, it may result in elevated plasma levels of chylomicron remnants, which
utilise this pathway for plasma clearance. In mammals the LDL-receptor has high
affinity for, and specifically interacts with two protein ligands, apo B-100 and apo E
(Mamo, 1995), (Schneider, 1989). The apo B-48 molecule present on chylomicron
remnants lacks the binding domain required for the LDL-receptor. Subsequently,
there is an absolute requirement for apo E to mediate the receptor-mediated uptake of
chylomicron remnants from plasma (Plump et al, 1992), (Innerarity and Mahley,
1978), (Sherrill et al, 1980), (Windier et al, 1980a), (Windier et al, 1988), (Arbeeny
and Rifici, 1984), (Cooper et al, 1982a), (Hui et al, 1984b), (Mortimer et al, 1995a),
(Havel, 1998). This process is mediated by LDL-receptors at the surfaces of
parenchymal liver cells (Francke et al, 1984), (Mamo et al, 1991), (Bowler et al,
1991). These ligands bind to distinct pockets within the general binding domain of the
LDL-receptor (Brown et al, 1991).

58
LDL-receptors have been detected in liver cells from several species (Brown
and Goldstein, 1983) and have a higher affinity (20 to 25-fold) for apo E than apo B-
100 (Mahley et al, 1984), (Innerarity and Mahley, 1978), (Brown et al, 1981), (Hui
et al, 1984a). Nagata et al. (Nagata et al, 1988) found that an antibody to the LDL-
receptor markedly decreased the endocytosis of chylomicron remnants by isolated rat
hepatocytes, thus supporting the theory that the presence of apo E on chylomicron
remnants makes them the preferred substrate. Four apo E molecules are needed for
binding of a lipid particle to the LDL-receptor (Funahashi et al, 1989). This also
suggests that the LDL-receptor binds chylomicron remnants and IDL with much
greater affinity than L D L , which has no apo E. Although V L D L can interact with the
receptor via apo B-100 to some extent, this tends to be inhibited by other V L D L
proteins, particularly the C apolipoproteins.
Apo E has been shown to be the ligand responsible for facilitating the
clearance of chylomicron remnants to the LDL-receptor. Plump et al. (Plump et al,
1992) have shown elevated plasma apo B-48 levels in apo E-knockout mice and
extensive complex arterial lesions have also been demonstrated (Zhang et al, 1992),
(Zhang et al, 1994), (Reddick et al, 1994). Using gene knockout mice deficient in
apo E or deficient in LDL-receptor, Mortimer et al. (Mortimer et al, 1995a)
demonstrated that apo E and LDL-receptors are both essential for the normal, fast
clearance of chylomicron remnants by the liver. Martins and Redgrave (Martins and
Redgrave, 1998) repeated these experiments, using chylomicron remnant-like
emulsions labelled with cholesterol[l-13C]oleate, and found no enrichment of 1 3 C 0 2
in the breath of apo E-deficient mice, and the appearance of 13C02 in the breath of
LDL-receptor-deficient mice was markedly decreased. Combined, the results suggest
that in the absence of LDL-receptors, an alternative apo E-dependent pathway
operates to clear the chylomicrons from the plasma, with significantly delayed
catabolism.
A cogent reason for questioning the involvement of L D L (B-100/E)-receptors
in chylomicron remnant clearance is provided by observations in genetic LDL-
receptor deficiency. F H is a genetic disorder in which the LDL-receptor gene is
mutant and exists clinically in two forms: homozygotes and heterozygotes
(Frederickson and Levy, 1972). Characterisation of familial hypercholesterolemic
(FH) individuals and Watanabe heritable hyperlipidemic ( W H H L ) rabbits has shown
a defective expression of LDL-receptors in both groups. The role of the LDL-receptor

59
for removal of chylomicron remnants was assumed to be minimal, based on studies
showing normal plasma clearance of chylomicron remnants in subjects with
homozygous familial hypercholesterolemia (Rubinsztein et al, 1990) and W H H L
rabbits (Kita et al, 1982-b), both of which lack functional LDL-receptors. Since
chylomicron remnants or hypertriglyceridemia have not been observed in the fasting
plasma of patients with homozygous FH, it has been suggested that humans express a
specific chylomicron remnant receptor (Goldstein and Brown, 1983), (Kita et al,
1982-b), or an alternate route(s) of uptake. A possible candidate was the LRP, which
was shown to bind apo E-enriched remnants and was later shown to be the oc-2
macroglobulin receptor (Beisiegel et al, 1989), (Strickland etal, 1990).
The binding of L D L to the LDL-receptor is a univalent interaction (Redgrave,
1999), (Havel, 1986b), whereas it has been postulated that four or five LDL-receptors
are required to bind one chylomicron particle via apo E (Pitas et al, 1979). As a
consequence, small remnants carrying fewer than four molecules of apo E will bind
with reduced efficiency. A decreased concentration of LDL-receptors in clusters on
the cell surface will also reduce binding efficiency. This may be important in diabetic
individuals who have moderate downregulation of the LDL-receptor, resulting in
normal L D L metabolism, but impaired multivalent remnant clearance.
Rubinzstein et al. (Rubinsztein et al, 1990) utilised the retinyl palmitate fat
tolerance test to measure chylomicron remnant clearance in 10 normal subjects (apo E
isotypes 3 or 4 only), 6 normolipidemic apo E2/3 homozygotes and 5 F H
homozygotes. Experiments in vivo revealed no significant differences between
chylomicron remnant clearance in normal compared with homozygous F H subjects,
assessed on the basis of retinyl palmitate levels in postprandial serum, chylomicrons
or chylomicron remnants. However at 7 hour time points, there was a delay in the
plasma clearance of retinyl palmitate. Remnant clearance was greatly decreased at all
times in the apo E2/3 homozygotes, indicative of an important degree of control
exerted by a receptor-mediated step involving apo E as ligand. The absence of any
excess remnant accumulation in F H subjects with varying "impairment' of L D L -
receptor-mediated degradation of apo E-containing lipoproteins, led the authors to the
conclusion that chylomicron remnants are initially cleared from the plasma by apo E-
recognising receptors which are genetically distinct from LDL-receptors (Rubinsztein
et al, 1990). The authors did not preclude the suggestion that low numbers of L D L -
receptors may be sufficient to endocytose remnant particles, which have been

60
'precleared' from the circulation by high capacity binding sites in hepatic sinusoids,
possibly of a glycosaminoglycan nature (Mahley et al, 1989).
In lieu of their finding that chylomicron remnants were removed from
circulation and accumulated in the liver of W H H L rabbits at a normal rate, Kita et al.
(Kita et al, 1982-b) postulated that the plasma clearance of chylomicron remnants
occurred via a receptor-mediated pathway genetically distinct from the apo B100/E
receptor. These rabbits suffer from homozygous LDL-receptor gene mutations
associated with a severe LDL-receptor deficit. Although the results from Kita et al.
(Kita et al, 1982-b) are referred to frequently when discussing the role of the LDL-
receptor (or lack of), the fact that binding of remnants to W H H L hepatocytes was
severely impaired compared with that of hepatocytes from control rabbits is not
discussed. The clearance of chylomicron remnants in W H H L rabbits may be regulated
via an alternate pathway, which has become more efficient, given the LDL-receptor
deficiency.
Demacker et al. (Demacker et al, 1992) also found that the clearance of apo
B-48 in W H H L rabbits was normal, indicating the presence of an uptake mechanism
independent to the LDL-receptor. Since chylomicron remnants have not been
observed in the fasting plasma of patients with homozygous FH, it has been suggested
that humans also express a specific chylomicron remnant receptor (Goldstein and
Brown, 1983). However, chylomicron remnants clear rapidly from plasma, with a
half-life of approximately 20 minutes, therefore are generally not found in fasting
plasma. Weintraub et al. (Weintraub et al, 1987b) support this by suggesting that the
enhanced risk of C H D in F H patients with mild hypertriglyceridemia may be related
to delayed clearance of remnants via the LDL-receptor.
Van Lenten et al. (Van Lenten et al, 1985) have shown that W H H L rabbits
still take up chylomicron remnants, providing evidence of another means of uptake.
Macrophages were loaded with cholesterol and rates of uptake and degradation for
p V L D L , L D L and remnants were monitored. Receptor activities for (3VLDL and L D L
were downregulated in cholesterol-loaded cells, but the rate of uptake and degradation
for remnants was unchanged in cholesterol-loaded cells, suggesting an unregulated
path of uptake for remnants. Similar findings were made in W H H L rabbit alveolar
macrophages. Additionally, (3VLDL and chylomicron remnants were equally effective
in competing for uptake in cholesterol-loaded macrophages and cholesteryl ester

61
accumulation was observed in monocyte-derived macrophages incubated with
remnants, supporting the existence of a remnant (apo E) receptor. F H macrophages
also degraded remnants, providing evidence for another means of uptake.
These observations cannot be considered unequivocal evidence for an LDL-
receptor- independent pathway for chylomicron remnant clearance, as LDL-receptor-
mediated endocytosis of apo E-containing lipoproteins is catalysed at significant rates
in cultured fibroblasts from some F H mutants (Hobbs et al, 1986), including W H H L
rabbits (Wernette-Hammond et al, 1989).
Bowler et al. (Bowler et al, 1991) found the in vivo clearance of chylomicron-
like emulsions in homozygous W H H L rabbits to be severely retarded compared with
control rabbits. The authors demonstrated that the LDL-receptor accounts for more
than 9 0 % of the chylomicron-like remnant removal, indicating that the LDL-receptor
pathway is the primary route of clearance. Based on further studies showing that
chylomicron metabolism in heterozgyous W H H L rabbits was significantly delayed
and L D L was cleared normally (Mamo et al, 1991), it appears that LDL-receptor
remnant clearance is more closely regulated than L D L clearance. Chylomicron
remnant clearance and uptake has also been found to be impaired in W H H L rabbits by
Hussain and colleagues (Hussain et al, 1995), and Beaumont and Assadollahi
(Beaumont and Assadollahi, 1990) observed an increase in plasma retinyl ester in
W H H L rabbits after vitamin A feeding, indicating impaired remnant catabolism.
Redgrave et al. (Redgrave et al, 1995) have utilised chylomicron-like and
chylomicron remnant-like emulsions labelled with [l-14C]oleate to monitor remnant
catabolism via a breath test in intact animals. In homozygous W H H L rabbits, the
appearance of 14C02 in the breath was much slower than in normal control rabbits,
while in heterozygous W H H L rabbits an intermediate level of appearance was found,
consistent with studies discussed above.
More recently, macrophages from W H H L rabbits were found to degrade
remnants and L D L , though to a lesser extent (Yu and M a m o , 1997b). Choi et al.
(Choi et al, 1991) also demonstrated a slower remnant clearance in mice injected
with antibodies to the LDL-receptor. The proposed role of the LDL-receptor in
chylomicron remnant metabolism is in agreement with other in vivo experiments in
rats (Jackie et al, 1992). Together, these clearance studies demonstrate a clear defect
in remnant removal kinetics when the LDL-receptor pathway is compromised, and
have raised doubts on the existence of a remnant receptor separate to the LDL-

62
receptor. In vivo and in vitro studies have confirmed that the LDL-receptor binds
chylomicron remnants, and is responsible for the removal of a substantial portion of
chylomicron remnants from the circulation (Mortimer et al, 1995a), (Ishibashi et al,
1996), (Zhang et al, 1992), (Windier et al, 1988). Thus, considerable evidence
supports the role of hepatic LDL-receptors in both L D L and chylomicron remnant
removal under normal circumstances, and to a lesser extent in peripheral tissue such
as bone marrow and spleen (Bowler et al, 1991).
To elucidate the role of the LDL-receptor in the clearance of chylomicron
remnants in humans, Cabezas et al. (Cabezas et al, 1998) studied remnant clearance
in untreated heterozygous F H subjects using the oral-retinyl palmitate fat-loading test.
Chylomicron remnant clearance was two-fold delayed in F H subjects compared with
controls, as assessed by area under the retinyl palmitate curve. The authors concluded
that the clearance of chylomicrons and chylomicron remnants depends primarily on
the LDL-receptor.
In support of these findings, M a m o et al. ( M a m o et al, 1998a) also found the
metabolism of chylomicron remnants in Japanese subjects with homozygous F H to be
significantly delayed in vivo, utilising apo B-48 and an oral fat load test as markers for
chylomicron remnant clearance. Furthermore, the fasting concentration of apo B-48 in
F H subjects were three-fold greater that normolipidemic controls, suggestive of
massive accumulation of small dense cholesterol-enriched remnants. Assessment of
the metabolism of chylomicron remnants in fibroblasts from subjects with
homozygous F H by Y u et al (Yu et al, 1997) showed that binding and degradation of
chylomicron remnants was reduced by about two-thirds. Interestingly, they found that
degradation of remnants in fibroblasts persisted in the absence of functioning LDL-
receptors, and that their binding to these cells was saturable. Competition and use of
blocking agents ruled out the involvement of the scavenger receptor, and confirmed
the involvement of the oc-2 macroglobulin receptor and a phagocytic uptake pathway.
This could provide an additional mechanism for accumulation of lipids in individuals
with F H and indicates a multi-factorial contribution to the development of
atherosclerosis in patients with FH, whereby L D L clearance is delayed and
chylomicron remnant clearance is significantly reduced.
Studies by Ishibashi et al (Ishibashi et al, 1996) also suggest that remnant
clearance is significantly impaired in LDL-receptor-knockout mice. Studies by Herz

63
et al (Herz et al, 1995) and Mortimer et al (Mortimer et al, 1995a) in LDL-
receptor-deficient mice support the hypothesis that this receptor is not essential for
efficient initial removal of remnants by the liver. They also found post-hepatic uptake,
that endosomal accumulation of cholesteryl esters of chylomicron remnants was
reduced despite the unimpaired rate of initial removal by the liver.
In a study aimed at identifying the primary binding site for chylomicron
remnants, Y u and M a m o (Yu and M a m o , 1997a) showed that smooth muscle cells
displayed saturable high affinity binding of remnants. L D L inhibited binding of
remnants by almost 6 0 % in receptor competition studies, suggesting the LDL-receptor
is the primary route of uptake. Furthermore, chylomicron remnant uptake by smooth
muscle cells was three times greater than L D L , consistent with greater affinity of the
LDL-receptor for lipoproteins containing apo E. In support of these findings, the
binding and degradation of chylomicron remnants by smooth muscle cells from
homozygous and heterozygous W H H L rabbits was impaired to a severe and
intermediate degree, respectively. These findings were confirmed using a polyclonal
antibody specific for the L D L - receptor, which was found to inhibit remnant binding
and degradation by 9 0 % . Koo et al (Koo et al, 1986), (Koo et al, 1988) showed that
chylomicron remnants bound exclusively to the LDL-receptor of human macrophages
and that binding of (3-VLDL (which shares the same binding characteristics as
chylomicron remnants) was significantly depressed in cells from subjects lacking the
LDL-receptor.
Fujioka et al. (Fujioka et al, 1998) undertook two complementary approaches
to identify the potential receptor processes involved in the uptake of chylomicron
remnants by macrophages, using specific inhibitors of the LDL-receptor and the LRP,
and macrophages harvested from normal, LDL-receptor knockout, and apo E
knockout mice. The authors deduced that at least 4 0 % of chylomicron remnant uptake
is due to the LDL-receptor, at least 2 0 % due to another member of the LDL-receptor
family, most likely the LRP, and perhaps the remaining 20-30% due to currently
unidentified mechanisms. The study provided an insight into how chylomicron
remnants can contribute directly to the transport of dietary lipid to artery wall
macrophages and foam cell formation under conditions where atherosclerosis occurs
at an accelerated rate. The results suggest that even in the absence of the LDL-
receptor, macrophages can still bind and metabolise chylomicron remnants, albeit at a

64
slower rate. This assertion predicts that chylomicron remnants could transport a
significant amount of lipid to macrophages, as L R P levels are not affected by lipid
loading.
A description of the interaction of chylomicron remnants and isolated rat
hepatocytes, and the characteristics of receptor-mediated endocytosis, was put
forward by Floren and Nilsson (Floren and Nilsson, 1977a), (Floren and Nilsson,
1977b). The authors suggested that it was specific, saturable, and temperature-
dependent process. Pronase treatment of the cells inhibited the process. Following
uptake, a chloroquine-inhibitable process degraded particles. Both of these
observations are characteristic of LDL-receptor-mediated uptake, and have been
confirmed by Sultan et al (Sultan et al, 1989) in hepatocytes. Similar binding and
uptake has been shown with fibroblasts (Floren et al, 1981) and macrophages (Floren
and Chait, 1981), (Ellsworth et al, 1987). Furthermore, remnants competed with L D L
for binding, and the affinity of remnants was higher than that of LDL. This finding
was consistent with remnants being rich in apo E, and apo E having a higher affinity
for the LDL-receptor than apo B. This was confirmed by W a d e et al. (Wade et al,
1986), who documented remnant binding to the LDL-receptor on ligand blots of liver
membranes prepared from dogs, rats, and rabbits. Based on studies of rat chylomicron
remnant binding to HepG2 cells, Chen et al. (Chen et al, 1991) concluded that most
remnant binding in these cells was regulated and could be accounted for by the LDL-
receptor. Earlier experiments by Greeve et al (Greeve et al, 1988) showed that the
membrane-binding assay represents mainly specific interaction of chylomicron
remnants with the LDL-receptor. In studies conducted by Nagata et al. (Nagata et al,
1988) in rat hepatocytes, the monospecific anti-LDL-receptor antibody blocked at
least 8 0 % of remnant binding and uptake. Jackie et al (Jackie et al, 1992) and Choi
et al. (Choi et al, 1991) have both found that the LDL-receptor may mediate
approximately 5 0 % of the initial removal of chylomicron remnants in normal rats and
mice. Thus it seems that a proportion of remnant uptake could be mediated by the
LDL-receptor.
Cooper et al. (Cooper et al, 1982b) concluded that V L D L remnants and
chylomicron remnants share the same hepatic removal mechanism. Excess
chylomicron remnants or V L D L remnants inhibited removal of 125I-VLDL by the
perfused liver and remnants were removed by the liver at a rate independent of

65
whether the particle was formed in the liver or intestine. Differences in clearance from
plasma may reflect differences in the rate of remnant formation.
Thus the primary mechanism by which arterial smooth muscle cells bind and
degrade chylomicron remnants is via the LDL-receptor. This provides support to the
atherogenic potential of chylomicron remnants, which have been shown to be
delivered to the subendothelial space of arterial vessels (Proctor and M a m o , 1998),
and confirms their ability to act as a substrate for arterial cells. Any delay in clearance
equates to longer circulation time in plasma and blood vessels. Remnants are taken up
via alternate pathways, which are unregulated and phagocytic in nature, and these
pathways target remnants to the artery and not the normal hepatic uptake route. This
allows for penetration of the arterial wall by these particles, both of which have been
demonstrated to penetrate and remain within the endothelium (Proctor and M a m o ,
1998), ( M a m o and Wheeler, 1994), (Nordestgaard et al, 1995). This could result in
increased accumulation in the artery wall and provide the necessary environment for
plaque formation.
There remains widespread speculation about the receptors responsible for the
uptake and clearance of postprandial lipoproteins, however it is generally agreed that
the LDL-receptor plays a role in this process. The contribution of the LDL-receptor
will also depend on the amount of receptor expression, and the health of the
individual in terms of diet, genetic abnormalities and hormonal state. However, if
LDL-receptor expression is absent or defective, the uptake of chylomicron remnants
via alternative mechanisms is not as efficient.
A recent review by Havel (Havel, 1998) suggests that the uptake of chylomicron
remnants by rodent liver is mediated by proteins residing on the microvillus surface of
hepatocytes and occurs in two steps. The initial removal of remnants from the blood
occurs through binding to the LDL-receptor via apo E and to H L via polar lipids and
proteins on the remnant surface. Second, remnants are taken up into the cell mainly by
the LDL-receptor and follow the classical receptor-mediated pathway of endocytosis.
1.5.3.2 LDL-receptor-related protein (LRP)
Recent research has also shown that the clearance of chylomicron remnants from the
blood involves multiple components on the surface of parenchymal liver cells. These
include H L and the recognition of apo E on remnant particles by endocytotic

66
receptors, primarily the LDL-receptor, with other receptors such as the LRP,
participating in the backup mode (Havel, 1996).
The absence of significant accumulation of chylomicron remnants in
homozygous LDL-receptor-deficient animals and humans (Kita et al, 1982b),
(Rubinsztein et al, 1990) initially raised the possibility of the existence of an
additional receptor. The L R P may be the responsible receptor (Brown et al, 1991). In
1988, Herz and co-workers described a candidate protein for the role of the "remnant
receptor". This protein was isolated from a c D N A clone encoding a lipoprotein cell
surface receptor distinct from but closely related to the classical LDL-receptor. The
L R P contains 4,525 amino acids and has a number of structurally distinct regions
(Herz et al, 1988), (Herz et al, 1990). Four of these contain cysteine-rich repeats
homologous to those on the LDL-receptor and other sequences homologous to EGF, a
membrane-spanning region, and a cytoplasmic domain that includes sequence
targeting to coated pits. The protein recycles through endosomal compartments in a
fashion analogous to the LDL-receptor, and is found in hepatic endosomes (Lund et
al, 1989), (Herz et al, 1988). Unlike the LDL-receptor, L R P expression cannot be
downregulated by exogenous cholesterol or upregulated by estrogens (Brown et al,
1991), (Kurt et al, 1989), (Kowal et al, 1989), (De Villiers et al, 1994), (Krieger and
Herz, 1994).
The first demonstration that L R P was identical to the a2-macroglobulin
receptor was made independently by Kristensen et al. (Kristensen et al, 1990) and
Strickland et al. (Strickland et al, 1990), and has since been confirmed (Kowal et al,
1990). Since 0C2-macroglobulin is known to bind plasma lysosomal enzymes and
growth factors, L R P may conceivably function as a "remnant receptor" and as a
"scavenger receptor". The L R P has been termed the "remnant receptor", based on
evidence showing that it binds P-migrating V L D L (which shares similar binding
properties to chylomicron remnants), and apo E-containing liposomes in a Ca +
sensitive way (Herz et al, 1988), (Kowal et al, 1990).
L R P also functions in the cellular uptake of protease-inhibitor complexes
(Breslow, 1993). Although this protein binds chylomicron remnants of plasma poorly,
the apo C and apo E content of the remnant particles can modulate binding of
chylomicrons to the LRP-oc2-M-receptor (Kowal et al, 1990), (Mokuno et al, 1994).
The observation that the 39-kDa inhibitor of L R P (RAP) reduces endocytosis of

67
chylomicron remnants (Mokuno et al, 1994), (Willnow et al, 1994) indicates that the
L R P or a related receptor is involved. However, the role of L R P in the uptake of
chylomicron remnants has been of some debate, and is confounded by contradictory
findings suggesting different contributions to their removal.
It is possible that L P L bound to lipoproteins may also be a ligand for L R P
(Gown et al, 1986) and may facilitate binding to remnant particles by interacting with
heparan sulphate proteoglycans (Brown et al, 1991), (Beisegel et al, 1991).
Similarly, H L bound to heparan sulphate proteoglycans (HSPG) on hepatocytes
interacts with the L R P with high affinity, resulting in the uptake of lipoproteins in
these cells (Krapp et al, 1996), (Havel, 1998). Beisiegel et al (Beisegel et al, 1991)
have shown that L P L can greatly enhance the binding of "L -like" lipoproteins and
apo E-containing liposomes to LRP. The secreted lipases, namely L P L and H L , can
further process remnants and maybe potentiate the cellular uptake of cholesteryl ester
from chylomicron remnants or other lipoproteins. The mechanisms involved in this
may assist in anchoring the lipoprotein to the cell surface and/or hydrolysis of the
esters themselves (Ji et al, 1994b), (Sultan et al, 1990).
In addition to the liver, the L R P is expressed in the brain, placenta and skin
(Kowal et al. 1989, Strickland et al. 1990, (Brown et al, 1991), (Krieger and Herz,
1994), (Herz and Willnow, 1995). The L R P is also a receptor for the milk protein
lactoferrin, proteinase inhibitor ct2-macroglobulin, and tissue-type plasminogen
activator (Chappell et al, 1992), (Willnow et al, 1992), (Huettinger et al, 1992),
(Herz et al, 1992), (Warshawsky et al, 1994), (Krieger and Herz, 1994). This
suggests a broad range of physiological roles in addition to chylomicron remnant
uptake.
Rohlmann et al. (Rohlmann et al, 1998) found massive accumulation of apo
B-48-containing lipoproteins in LDL-receptor knockout mice in which high levels of
R A P in the liver were induced. This did not occur with similar induction of R A P in
wild type mice, indicating that a RAP-sensitive process is responsible for endocytosis
of remnants in LDL-receptor knockout mice. Results from another study showed the
same accumulation in LDL-receptor knockout mice with L R P selectively knocked
out, but not in wild type mice. Combined, the authors concluded that despite
inefficient endocytosis of remnants via LRP, L R P is an effective substitute for the
LDL-receptor in animals given limited amounts of dietary fat. Havel (Havel, 1998)

68
has interpreted these findings as L R P contributing to the initial binding and
endocytosis of chylomicron remnants with large dietary fat loads or under conditions
in which the hepatic LDL-receptor is downregulated. H e concluded that L R P mainly
serves as a backup receptor when an adequate number of LDL-receptors are available.
Willnow et al. (Willnow et al, 199A) used gene transfer technology in mice to
attempt to elucidate the role of L R P in chylomicron remnant uptake. Inhibition of
L R P by the overexpression of R A P was associated with a marked accumulation of
chylomicron remnants in LDL-receptor-negative mice and with some accumulation
on normal mice. This suggests that LDL-receptor and L R P are both involved in
remnant clearance.
A knockout of the L R P gene has been achieved, but homozygous deficiency is
nonviable and heterozygous deficient mice are normal (Herz et al, 1992). Therefore,
this model has not been useful in confirming the role of L R P in chylomicron-remnant
metabolism in vivo, particularly in LDL-receptor normal individuals. Despite this,
Martins et al. (Martins et al, 2000b) utilised remnant-like lipid emulsions and found
the enrichment of 1 3 C 0 2 in the breath of heterozygous LRP-negative mice was
delayed, and in LRP/LDL-receptor-deficient mice was negligible, and concluded that
LDL-receptors are important for the physiological metabolism of chylomicron
remnants under normal circumstances. However, their findings support the hypothesis
that when the LDL-receptors are absent, the L R P may assume a role in the uptake of
chylomicron remnants in liver cells. Alternately, Choi and Cooper (Choi and Cooper,
1993) used an antibody to the LDL-receptor, and demonstrated in mice that the LDL-
receptor accounts for ~ 6 0 % of chylomicron remnant uptake by the liver. In vitro,
they used Chinese hamster ovary cells to demonstrate - 8 0 % inhibition of remnant
uptake and degradation by the LDL-receptor antibody. Additionally, they successfully
used the 39-kDa receptor-associated protein R A P , a high-affinity inhibitor of ligand
binding to the LRP, to block 2 5 % of remnant removal by the liver. More recently, Y u
et al (Yu et al, 1999) found that R A P reduced the removal of remnants by 30-40% in
normal mouse livers. This led the authors to suggest that the LDL-receptor has a more
definitive role in the uptake of chylomicron remnants, and that while the L R P
contributes to this process, it may assume a more 'back-up' or complimentary role.
Havel (Havel, 1998) infers that the LRP, which binds weakly to chylomicron
remnants via apo E, does not appear to have a significant role in the initial removal
process. The remnant particles can be enriched with HSPG-bound apo E present on

69
hepatocyte microvilli, which increases their affinity for L R P to the extent that they are
subject to endocytosis by this receptor, particularly when the LDL-receptor is
deficient or downregulated. This backup mechanism is sufficient to prevent an
appreciable accumulation of chylomicron remnants in humans and other mammals
lacking functional LDL-receptors.
1.5.3.3 Lipolysis-stimulated receptor
Another proposed receptor for the uptake of lipoproteins from plasma is the lipolysis-
stimulated receptor, described by Ji et al. (Ji et al, 1993), (Ji et al, 1994a). In the
presence of free fatty acids, the lipolysis-stimulated receptor recognises either apo B
or apo E, and as a consequence, leads to the internalisation and degradation of the
lipoprotein particles, primarily by the liver. Its affinity is highest for those lipoproteins
most susceptible to lipolysis, the triglyceride-rich lipoproteins.
Ji et al. (Ji et al, 1993), (Ji et al, 1994a) have provided evidence that apo E-
transfected rat hepatoma cells triggers a greatly enhanced remnant uptake involving
HSPGs. H S P G s are polyanionic macromolecules that contain a protein core with
multiple heparan sulfate chains covalently attached. The biological roles of H S P G s
depend on the core protein, which assumes anchoring function and provides a scaffold
for appropriate immobilisation and spacing of heparan sulfate chains (Andres et al,
1992). In hepatocytes, H S P G s are thought to bridge the extracellular matrix and
intracellular cytoskeleton whereas H S P G s bound to endothelial cell plasma
membranes facilitate the binding of circulating macromolecules including growth
factors, LPL, and probably lipoproteins and remnants.
Mahley and Ji (Mahley and Ji, 1999) and others (Mahley and Hussain, 1991),
(Mahley et al, 1994), (Ji et al, 1993), (Ji et al, 1994a) have postulated an initial
sequestration of remnant particles within the space of Disse through interaction with
H S P G s on the surface of hepatocytes. The space of Disse has a high local
concentration of apo E, which is produced by hepatocytes together with an abundance
of H S P G s (Stow et al, 1985), (Hamilton et al, 1990). Mahley and Ji (Mahley and Ji,
1999) suggest that cell-surface H S P G play a critical role in remnant uptake as an
essential or integral component of the H S P G - L R P pathway. The H S P G may serve as
a reservoir for apo E, allowing the particles to be enriched in apo E, which facilitates
their interaction with the LRP. In addition, the authors hypothesise that H S P G appear

70
to function alone as receptor and display unique handling properties for specific
isoform of apolipoprotein E. Zeng et al (Zeng et al, 1998) explored the interaction
between H S P G and endocytosis of remnants, using HepG2 cells with expression of
H S P G inhibited. They showed that remnant binding to liver cells depends on the
LDL-receptor, on the expression of H S P G core proteins, and on the functionality of
heparan sulfate in HSPG. These data support the role of secreted apo E in capturing
chylomicron remnants and P-migrating V L D L .
1.5.3.4 VLDL-receptor
The VLDL-receptor appears to bind only apo E-containing lipoproteins, and has been
found in rabbit and human tissues (Takahashi et al, 1992), (Sakai et al, 1994). The
VLDL-receptor c D N A has been isolated and encodes a protein with a molecular
weight of ~125kDa, and shares considerable structural and sequence homology with
the LDL-receptor (Takahashi et al, 1992). It has been shown to be present in high
numbers in heart, skeletal muscle and adipose tissues, but is absent in the liver.
The VLDL-receptor has been isolated from atherosclerotic lesions in both
humans and rabbits (Hiltunen et al, 1998), (Nakazato et al, 1996), and the V L D L -
receptor expression was primarily in macrophages. The role of the VLDL-receptor in
the uptake of the chylomicron remnants in vivo is not clear, and it appears that apo E
supplementation is necessary for the normal internalisation and degradation of
chylomicron remnants in macrophages (Fujioka et al, 1998).
Niemeier et al (Niemeier et al, 1996) have shown internalisation of chylomicron
remnants in LDL-receptor-negative Chinese hamster ovary cells overexpressing the
VLDL-receptor to be ~3-fold greater than in control cells. A further increase in
chylomicron remnant uptake was observed when a surplus of apo E was added to the
remnants, supporting the role of apo E in chylomicron remnant metabolism and as a
necessary ligand for the VLDL-receptor.
1.5.3.5 Scavenger receptors
Receptors that recognise modified LDL are known as scavenger receptors. When
L D L is chemically modified by acetylation it is taken up by macrophages via a
receptor-mediated process that results in massive accumulation of cholesterol. These

71
receptors are not down regulated by the accumulation of intracellular cholesterol
(Goldstein et al, 1979). The scavenger receptor is not specific for acetylated L D L but
recognises highly negative residues (Goldstein et al, 1979), (Brown et al, 1980) and
so is able to recognise malondialdehyde-modified L D L . The ligand in vivo is
unknown but L D L could be modified by malondialdehyde, which is a by-product of
oxidation of arachidonic acid (Kraemer, 1987). Freeman et al. (Freeman et al, 1991)
have also used Chinese hamster ovary cells expressing scavenger receptors to
demonstrate the role of the scavenger receptor in the accumulation of cholesteryl
esters in response to modified LDL.
To date, there is no evidence that the scavenger receptor is responsible for the
unregulated uptake of chylomicron remnants by macrophages, despite it being a
multi-ligand receptor binding to a range of modified lipoproteins (Floren and Chait,
1981), (Ellsworth et al, 1987), (Mamo et al, 1996).
While there is support for the multiple pathways for chylomicron remnant
removal, the role of the LDL-receptor appears to predominate. Other receptors may
also play a specific role, which becomes more significant when the LDL-receptor is
downregulated or absent. Uptake of remnants via the L R P may be restricted to sites
that specifically secrete apo E and this secretion may increase in circumstances such
as cholesterol loading or LDL-receptor deficiency. The most probable situation is that
the LDL-receptor and the L R P work in concert, with one contributing more in
conditions where the other is down-regulated or absent (Ishibashi et al, 1994b),
(Willnow et al, 1994), (Mortimer et al, 1995a). Alternatively, receptors other than
the LDL-receptor may only assume physiological significance when the LDL-
receptor is downregulated, defective or absent.
1.5.4 Lipoproteins, Cholesterol and Atherosclerosis
The initial hypothesis that cholesterol is involved in the pathogenesis of
atherosclerosis stemmed from early findings of cholesterol in atherosclerotic lesions
(Windhaus, 1910). In 1913, Anitschow and Chalatow showed that feeding cholesterol
to rabbits induced atherosclerosis, supporting this hypothesis (Anitschow and
Chalatow, 1913). Numerous studies have since revealed that a high intake of dietary
cholesterol causes severe hypercholesterolemia and atherosclerosis in many animal
species. In 1916, a Dutch medical practitioner suggested that the Javanese people had

72
minimal atherosclerosis because their traditional diet contained little fat and
cholesterol (De Langen, 1916), (De Langen, 1922). W h e n these people were
subsequently fed European diets rich in animal fat, their blood cholesterol level
increased substantially. Since then, a series of international epidemiological studies
have shown significant correlations between the daily intake of saturated fats and
cholesterol and mortality from C H D (Menotti et al, 1993), (Connor and Conner,
1986), (Keys, 1970), (Kannel and Castelli, 1979). Specifically, the 15-year death rate
in the Seven Countries Study showed that the mortality from C H D and all causes
were related positively to average percentage of dietary energy from saturated fatty
acids. However, death rates were negatively related to the energy percent from dietary
monounsaturated fatty acids, and were low in cohorts with olive oil as the main fat
(Keys et al, 1986). It is now known that there is a direct relationship between the
intake of dietary fatty acids and cholesterol, hyperlipidemia and vascular
atherosclerotic disease. High quantities of cholesterol and saturated fats in the diet
may induce hypercholesterolemia, which is a significant risk factor for atherosclerosis
(Nordoy and Goodnight, 1990), (McNamara etal, 1987).
In primates, the major effect of dietary cholesterol is thought to be an increase
in LDL-cholesterol concentrations. Not only does a high intake of cholesterol increase
the number of circulating L D L particles but it can also change the size and
composition of these particles (Rudel et al, 1985). The rise in serum cholesterol in
animals fed high cholesterol diets may also contribute to an enrichment of cholesteryl
ester in newly secreted lipoproteins at the expense of triglyceride. Initial studies in
tissue culture have clearly demonstrated that increasing the cholesterol content of
cells downregulated the synthesis of LDL-receptors (Goldstein and Brown, 1975). A
down regulation of LDL-receptors is certainly seen in rabbits and hamsters fed
cholesterol (Spady et al, 1985), (Kovanen et al, 1981). Therefore, this may be the
principal mechanism whereby plasma L D L increases with dietary cholesterol.
It has been speculated that a high dietary intake of cholesterol
enhances cholesterol transport of chylomicrons from the intestine to the liver
(Miettinen and Kasaiemei, 1986). The resultant overloading of hepatocytes with
cholesterol leads to several alterations in cholesterol metabolism, including increases
in bile acid synthesis, cholesterol secretion into bile, and a reduction in cholesterol
synthesis. As a result, A C A T activation enhances the esterification and storage of

73
cholesteryl esters in the liver, LDL-receptor is downregulated, and the release of
cholesterol in V L D L increases.
The downregulation of the LDL-receptor only occurs if the suppression of
synthesis is not adequate enough to compensate for the increase in dietary cholesterol.
In the rabbit (Kovanen et al, 1981) and hamsters (Spady et al, 1985), dietary
cholesterol causes marked hypercholesterolemia. Plasma cholesterol in these animals
can increase by up to 10-fold and it appears to be mainly a result of downregulation of
LDL-receptor expression by 60-70%. Hence, the decrease in LDL-receptor expression
in cholesterol-fed rabbits is the causative factor in their hypercholesterolemia
(Kovanen et al, 1981). Rats, however, are remarkably resistant to
hypercholesterolemia when fed cholesterol as LDL-receptor expression is not
downregulated as in rabbits (Roach et al, 1993a). Rats have an efficient capacity to
reduce cholesterol synthesis and increase cholesterol secretion as bile acids to
compensate for the incoming cholesterol in order to maintain cellular cholesterol
homeostasis. There is also a marked increase in the secretion of cholesterol-enriched
VLDL.
The response to dietary cholesterol in humans is more similar to the response
observed in rats than rabbits, as plasma cholesterol levels are not increased to a large
extent with cholesterol administration. There appear to be adequate compensatory
changes in various metabolic pathways, which allow the maintenance of cholesterol
homeostasis in man. The primary response is the suppression of cholesterol synthesis,
but there is also a decrease in fractional absorption of cholesterol from the intestine
and an increase in biliary excretion (McNamara et al, 1987). In addition, the
discovery that A B C A 1 is an apolipoprotein receptor implicates the involvement of
receptor-ligand interactions in regulating cholesterol trafficking (Oram et al, 2000).
Studies of human disease have revealed that A B C A 1 plays a critical role in clearing
excess cholesterol from macrophages and has a major impact on lipoprotein
metabolism (Assman et al, 1995), (Serfaty-Lacrosniere etal, 1994).
Nevertheless, there is still an effect when dietary cholesterol is consumed.
There appears to be a variable effect on serum cholesterol concentrations among
individuals with the same intake of dietary cholesterol (Grundy and Denke, 1990).
This may be related to the individual's ability to process dietary cholesterol (Miettinen
and Kasaiemei, 1986), (Clifton et al, 1990) resulting in some people being responsive
and others non-responsive to dietary cholesterol. The recent discovery of a mouse

74
strain (C57BL/6ByJ) that is resistant to diet-induced hypercholesterolemia and aortic
lesions compared to the C57BL/6J strain, led to the investigation of the gene(s)
underlying the resistant B6By phenotype (Mouzeyan et al, 2000). Genetic crosses
with an unrelated mouse strain revealed a locus, which showed highly significant
linkage between the two mouse types. The authors suggest that this locus represents a
novel gene affecting plasma lipids and atherogenesis in response to diet.
Abnormal lipoprotein levels are associated with several human diseases, most
commonly atherosclerosis. Individuals with atherosclerotic C H D almost invariably
have one or more of four lipoprotein abnormalities: increased LDL-cholesterol;
decreased HDL-cholesterol, usually associated with increased levels of triglyceride-
rich lipoproteins (VLDL); increased levels of IDL-cholesterol and chylomicron
remnants; and high levels of an abnormal lipoprotein; lipoprotein(a) (Breslow, 1993).
M a n y studies have demonstrated that an elevated plasma cholesterol level,
primarily within the L D L fraction, is a risk factor for C H D when their clearance from
plasma is impaired and concentration increased (Levine et al, 1995), (Sniderman et
al, 1997), (Brown and Goldstein, 1986). L D L is a cholesteryl ester-rich lipoprotein
and is the principal carrier of cholesterol in the plasma of persons with plasma
cholesterol concentrations higher than 150 mg/dl (Zilversmit, 1979). In addition,
apolipoprotein B-100 is found in the arterial lesion (Hoff et al, 1975a), (Hoff et al,
1975b), (Hoff et al, 1978). Familial hyperbetalipoproteinemia, in which high levels
of L D L are present prematurely, is associated with early arteriosclerosis and C H D .
L D L is usually elevated in experimental animals that eat atherogenic diets
(Zilversmit, 1979). Humans and animal models with LDL-receptor deficiencies have
slower L D L plasma clearance and develop atherosclerosis and C H D prematurely; and
the smaller size of L D L is thought to enhance passage across arterial endothelium.
A n important consideration is the relationship between high concentrations of
plasma L D L and atherogenesis, and how fatty streaks and foam cell are formed.
Steinberg and co-workers (Steinberg et al, 1989), (Steinberg, 1991) have proposed
that cells forming proliferative lesions of atherosclerosis, in particular macrophages
and endothelial cell, modify native plasma L D L locally within the arterial wall in such
a way that they acquire atherogenic potential. During the process of modification
native L D L loses its ability to bind to the classical LDL-receptor. Moreover, the
modification of L D L involves an alteration of apo B-100 so that the modified versions
of L D L bind to one or more classes of "scavenger receptors". Modified L D L adopts

75
biological properties that are consistent with an atherogenic role, including its role as
a potent promoter of inflammatory tissue reactions. It induces the formation of
cytokines and chemotactic peptides and the synthesis of proinflammatory arachidonic
acid metabolites (Habenicht et al, 1994).
Many researchers believe that chylomicrons and their remnant particles may
be atherogenic, but only in an indirect sense. This hypothesis is based on the fact that
a significant rise in chylomicron concentration is associated with increased
competition for L P L with V L D L particles. In turn, there is an increased concentration
of large V L D L , suggesting that V L D L accounts for much of the postprandial increase
of triglyceride-rich lipoprotein particle number, whereas the majority of postprandial
triglycerides are carried by chylomicrons. Bjorkegren et al. (Bjorkegren et al, 1996)
found that an infusion of chylomicron-like triglyceride emulsion (Intralipid) over a 60
min period elevated plasma triglycerides three-fold over the infusion, concomitant
with a linear increase of large V L D L . The emulsion caused a 75-90% block of the
conversion of large V L D L apo B to small V L D L apo B, estimated from simultaneous
stable isotope studies. The authors concluded that chylomicrons and their remnants
impede the normal lipolytic degradation of V L D L and could thereby be indirectly
implicated in the generation of atherogenic remnant lipoproteins.
Currently L D L is considered the most atherogenic of the lipoproteins and
hence the majority of research has been focussed on this lipoprotein and the
'endogenous' lipoprotein pathway. However there is an increasing body of evidence
to suggest that chylomicrons and their remnants may also contribute to lesion
formation. While L D L carries 7 0 % of plasma cholesterol in circulation (Lees and
Wilson, 1971), chylomicrons and their remnants carry only 2-5% of circulating
cholesterol at any one time (Redgrave, 1988). However, the half-life of chylomicrons
is less than 5 minutes and the total number of pools turned over per day is
approximately 100 (Redgrave, 1983). Despite the low proportion of cholesterol and
cholesteryl esters in the total chylomicron mass, approximately 3 g m of cholesterol
passes daily through the plasma in the form of chylomicrons and chylomicron
remnants. Therefore the flux of cholesterol through plasma is similar for
chylomicrons and L D L (Redgrave, 1999). Furthermore, the slow rate of progression
of vascular disease is consistent with a lipoprotein particle that generally clears
rapidly from plasma (5 min for chylomicron remnants, compared with 6-10 hr for
L D L ) . A n important point is that although remnants transport less than 5 % of total

76
plasma cholesterol at any time, individuals on a western diet will likely have a greater
flux of cholesterol through the chylomicron pathway than through L D L over a 24-hr
period (Mamo, 1995).
Although triglyceride-rich in composition, the large size and rapid turnover
rate of chylomicrons and chylomicron remnants imply that these particles are the
major contributors to cholesterol transport in plasma (Mamo, 1995), (Redgrave,
1983). In a postprandial dyslipidemic state, there is a prolonged elevation and
magnitude of plasma triglycerides, reflecting accumulation of all lipoproteins. This
may be the result of overproduction of triglyceride-rich lipoproteins by the intestine
and/or liver, as well as decrease in their clearance (Watts et al, 1998).
Both L D L and chylomicron remnants share the same primary route of
clearance in vivo namely the LDL-receptor under normal circumstances, and in
certain disease states, the clearance of both of these particles is impaired ( M a m o et
al, 1997). Little research has been dedicated to remnant clearance patterns and
metabolism in LDL-receptor deficient individuals, consequently the contribution of
chylomicrons to elevated plasma cholesterol or the role of the LDL-receptor in the
clearance of remnants it is still not clear. In addition, investigations into the role of
different classes of lipoproteins in producing cholesterol accumulation in
macrophages and in lesion formation of atherosclerotic lesions have been
inconclusive. Defects in chylomicron metabolism can occur at a number of stages in
metabolism. Maranhao et al. (Maranhao et al, 1995) summarised the factors affecting
plasma clearance of chylomicrons and chylomicron-like emulsions as variations in
L P L activity, hepatic receptor activity, and apo E isoforms. The production and
assembly of chylomicrons is also an important factor. Delays in lipolysis are related to
defects in L P L or apo CII, whereas defects in plasma clearance can generally be
attributed to defects in chylomicron remnant removal.
If chylomicron remnant clearance mechanisms such as the LDL-receptor are
defective, chylomicron remnant catabolism will be delayed. Given that chylomicron
remnants are able to penetrate and be retained by the arterial wall, and are degraded
by macrophages, they have an independent atherogenic potential. As Barrett
explained in a recent review (Barrett, 1998), changes induced in the postprandial state
complicate the application and development of models that describe lipoprotein
particle kinetics, and our understanding of the contribution of chylomicrons and their
remnants to the development of atherosclerosis. In addition, increased hepatic lipid

77
substrate supply following a high saturated fat meal also increases secretion of V L D L ,
which competes with chylomicrons for clearance and exacerbates postprandial
dyslipidemia (Brunzell et al, 1973). The hepatic receptor-mediated uptake of
chylomicron remnants is normally saturated for up to seven hours after a fatty meal
and its perturbation is closely related to the development of atherosclerosis (Mamo,
1995).
The putative role of chylomicron remnants in the pathogenesis of
atherosclerosis has generated an interest in the uptake of remnants by extrahepatic
tissues, which may provide an alternative route for chylomicron remnant uptake if the
normal liver pathways are absent or defective. There is evidence to suggest that the
predominant pathway by which macrophages take up chylomicron remnants is
phagocytosis. The evidence that chylomicron remnants are avidly degraded by and
induce lipid loading in macrophages is consistent with the inflammatory aspects of
atherosclerosis ( M a m o et al, 1997). Floren et al (Floren et al, 1981) found that
chylomicron remnants could be taken up by macrophages to form foam cells and can
gain access to human smooth muscle cells following endothelial injury, indicative of
their atherosclerotic capacity. Both Koo et al. (Koo et al, 1988) and Van Lenten (Van
Lenten et al, 1983) have found that chylomicron remnants can bind and are avidly
metabolised by macrophages, a process that is poorly regulated.
Ellsworth et al. (Ellsworth et al, 1986), (Ellsworth et al, 1987), (Ellsworth et
al, 1990) have reported that the uptake of chylomicron remnants by normal
macrophages is mediated primarily by the LDL-receptor. This raises the question of
how these particles contribute to the formation of atherosclerotic lesions. Cholesterol-
rich diets increase the cholesterol content of chylomicrons and their remnants. It is
thus possible that they can contribute to lesion formation by serving as a lipoprotein
source of cholesteryl ester possibly by facilitating the excessive accumulation of lipid
in macrophages in the artery wall and thus the generation of foam cells (Fujioka et al,
1998). This is likely in the case of cholesterol-enriched diet, resulting in release of
cholesterol-rich chylomicron remnants into the plasma (which down-regulates L D L -
receptor) and hence delayed clearance of these particles.
Arterial smooth muscle cells have been shown to rapidly take up certain
lipoproteins from normal or hypercholesterolemic serum. Cholesterol accumulation in
cultured arterial smooth muscle cells occurs after incubation with chylomicron
remnants (Floren et al, 1981). In addition, Zilversmit (Zilversmit, 1979) presented

78
evidence that chylomicron remnants as well as L D L are taken up by arterial cells. The
fact that chylomicrons are the principal carriers of dietary cholesterol in the
bloodstream and that the first stage of their degradation takes place in contact with the
vascular endothelium led Zilversmit to examine the possibility that chylomicrons
might be atherogenic, rather that simply modifying serum cholesterol concentrations.
If chylomicrons are loaded with cholesteryl ester derived from dietary cholesterol, the
remnants are similarly enriched in this lipid fraction (Mjos et al, 1975), (Ross and
Zilversmit, 1977). Liver cells rapidly ingest chylomicron remnants (Noel et al, 1975),
(Cooper, 1977). Cholesterol uptake from chylomicron remnants is much faster than
that from L D L (Andersen et al, 1977) and appears to be a saturable process (Sherrill
and Dietschy, 1978), (Floren and Nilsson, 1977b). It has been shown that fibroblasts
can bind and uptake chylomicron remnants in a saturable and specific manner
(Redgrave et al, 1982), utilising apo E as a ligand (Arnon et al, 1991), (Innerarity
and Mahley, 1978).
Proctor and M a m o (Proctor and M a m o , 1996), (Proctor and M a m o , 1998)
provided evidence for direct uptake of remnants into the arterial wall and fatty lesions.
The authors found that chylomicron remnants penetrate arterial tissue as efficiently as
smaller macromolecules including L D L , H D L and albumin and that there is an
exclusive increase in chylomicron remnant uptake in arterial tissue, whereas L D L
uptake was similar to non-lesioned arterial tissue. The accumulation of pre-formed
chylomicron remnants, chylomicrons and L D L in thoracic aorta of control, W H H L
and cholesterol-fed rabbits, in lesioned and non-lesioned aortic tissues was compared
(Proctor and M a m o , 1996). Aortic accumulation of chylomicron remnants was
substantially greater than for chylomicrons, and both were significantly greater than
LDL . In W H H L rabbits there was significantly greater aortic uptake of chylomicrons
and L D L compared with the carotid wall and, in cholesterol-fed rabbits, significantly
greater aortic uptake of chylomicrons. These findings suggested that in
hypercholesterolemia the lipoprotein retention properties of some arterial beds
change. In arterial fatty lesions from W H H L and cholesterol-fed rabbits there was an
exclusive increase in chylomicron remnant uptake, whereas L D L uptake was similar
to non-lesioned tissue. Thus, chylomicron remnants penetrate arterial tissue efficiently
and are retained in sites of lesion formation, suggesting they are potentially an
atherogenic lipoprotein. Proctor and M a m o (Proctor and M a m o , 1998) monitored the
delivery and retention of lipoproteins by confocal microscopy and found that

79
fluorescent chylomicron remnants widely penetrate arterial tissue via the luminal
surface within minutes of exposure, including the tunica media. Additionally, it was
shown that the efflux of remnants was often not complete after perfusion with buffer,
within the endothelial space and that significant sporadic focal accumulation
occurred. This retention is consistent with the putative atherogenicity of postprandial
lipoproteins.
Fielding (Fielding, 1978b) has studied the quantitative aspects of chylomicron
cholesterol and triglyceride uptake by the perfused rat heart. While 8 0 % of the
triglyceride was removed by the perfusate, 5 0 % of the chylomicron cholesteryl ester
was acquired by the myocardium. The uptake of cholesteryl ester appeared to be non
saturable and was not inhibited by L D L . The uptake of cholesteryl ester was increased
in proportion to the cholesteryl ester content of the chylomicrons. These findings
indicate that uptake of chylomicron cholesteryl ester by the myocardium would
increase when dietary cholesterol intake is high.
Apolipoproteins characteristic of chylomicrons (apo B-48) have been
identified in lesions of human arteries (Hoff et al, 1975a), (Hoff et al, 1975b) and
Scow et al (Scow et al, 1976) presented evidence of the adsorption of chylomicrons
to vascular endothelium, followed by degradation of most of their triglyceride. Rapp
et al (Rapp et al, 1994) and Mora et al. (Mora et al, 1990) have also investigated the
lipoproteins present in human atherosclerotic lesions and found primarily
endogenously derived lipoproteins in lesions. In addition, they examined the L D L
lesion lipid fraction of hypertriglyceridemic patients, and found increased triglyceride
levels, indicating the presence of chylomicron remnants. The presence of a band at the
position of apo B-48 on the L D L fraction immunoblot supported this finding. Yla-
Herttuala et al. (Yla-Herttuala et al, 1988) found apoB-48 present in lesion-free
aortas, consistent with the uptake of chylomicron remnants.
Taken together, these studies support the possibility that chylomicron
remnants possess the potential to penetrate the endothelial space and cause deposition
of cholesterol in the arterial wall. Specifically, there appears to be a role for remnants
in the aetiology of atherogenesis when these particles are elevated due to defective
removal mechanisms. The resultant state of dyslipidemia provides an environment
whereby these events can occur. This indicates a multi-factorial contribution to the
development of atherosclerosis in subjects who demonstrate delayed L D L clearance
and significantly reduced chylomicron remnant clearance.

80
In contrast, two recent studies in humans show that a major proportion of
chylomicron remnants are removed from plasma long before they attain a size at
which they may penetrate the arterial wall (Karpe et al, 1997b), Le et al (Le et al,
1997). Karpe (Karpe, 1999) has suggested that in addition to these properties of
remnants, the amount of cholesterol carried by apo B-48 remnants, both in fasting and
postprandial plasma, is very small compared with all other potentially atherogenic
lipoprotein species in human plasma. However, these observations may not be
appropriate for humans with defective receptor-mediated mechanisms, where
cholesterol-rich particles are circulating for longer periods of time in plasma, are
hydrolysed to smaller particles, and are able to penetrate the arterial wall.
It is feasible then, that circulating chylomicron (or V L D L ) remnants may bind
and penetrate the arterial surface, to be adsorbed and then degraded to remnants at the
arterial surface. The reactions leading to endocytosis of remnants by smooth muscle
cells may take place at sites where local injury has removed the endothelium, or they
may occur at endothelial surfaces, followed by vesicular transport to subendothelial
layers (Scow et al, 1976).
Cholesterol feeding has been shown to retard the clearance of isotopically labelled
cholesterol from lymph chylomicrons in rabbits, compared with normally fed rabbits
(Redgrave et al, 1976). These results suggested that the hypercholesterolemia
observed in cholesterol-fed rabbits was due to the accumulation of chylomicron
remnants in plasma. Ross et al. (Ross et al, 1978) found that rabbits fed either a
cholesterol-free, low-fat, semi-synthetic diet or rabbit chow plus cholesterol
developed hypercholesterolemia in the range of 300-600 mg/dl. After 4 weeks of
feeding, the semi-synthetic group carried most of its cholesterol as L D L , and the
cholesterol-fed group showed mostly V L D L and IDL. Zilversmit (Zilversmit, 1979)
concluded that the hypercholesterolemia of the cholesterol-fed rabbits is largely due
to chylomicron remnants, whereas the animals fed the semi-synthetic diet show
elevated levels of lipoproteins of endogenous origin. These findings support the
hypothesis that cholesterol of exogenous and endogenous origin is equally
atherogenic. In addition, it has been demonstrated that cholesterol-fed rabbits
accumulate retinyl palmitate-labelled chylomicron remnants in plasma to a much
greater extent than rabbits fed on a chow diet (Ross and Zilversmit, 1977). This was
assumed to be due to down-regulation of the LDL-receptor, based on a study by
Kovanen et al. (Kovanen et al, 1981).

81
1.5.4.1 Familial Hypercholesterolemia
The most convincing evidence for a close relationship between atherogenesis and
disorders of lipid metabolism is the striking incidence of C H D in patients afflicted
with the LDL-receptor negative phenotype of familial hypercholesterolemia
(homozygous F H ) (Scrott et al, 1972). Brown and Goldstein found that subjects with
F H had a substantially increased risk of atherosclerosis (Goldstein and Brown, 1974),
(Goldstein and Brown, 1973). The development of severe atherosclerotic disease in
F H demonstrated the preventive role of the LDL-receptor in the atherosclerotic
process, in addition to the direct correlation between developing atherosclerosis and
plasma cholesterol levels (Bilheimer, 1987), (Soutar et al, 1977).
In 1939, Muller identified F H as an inborn error of metabolism causing high
cholesterol levels and heart attacks in young people (Brown and Goldstein, 1984). He
showed that it was transmitted as a single autosomal dominant trait determined by a
single gene. In the 1970's, Khachadurian demonstrated that there are two forms of
FH, a heterozygous form and a more severe homozygous form (Khachadurian, 1971).
The heterozygous form occurs with an estimated frequency of one in 500 in the
general population, making it one of the most common inherited disorders in humans.
The much more severe homozygous form is rare, occurring with a frequency of about
one in 1,000,000 in the general population (Brown and Goldstein, 1986).
Familial hypercholesterolemia is characterised by elevated L D L cholesterol
levels, the presence of tendon and cutaneous xanthomas, and the development of
premature atherosclerosis (Goldstein and Brown, 1983). The most common
manifestations of atherosclerosis in heterozygotes are angina pectoris, myocardial
infarction and sudden death (Stone et al, 191 A), (Mabuchi et al, 1986). The
hypercholesterolemia also promotes cholesterol deposition in other tissues; e.g.
cholesterol infiltration in macrophages eventually causes nodular swellings of the
tendons in various sites (Goldstein and Brown, 1983). F H is generally associated with
normal V L D L concentrations (type Ua phenotype), but they may have increased
V L D L levels (type Hb phenotype) (Fredrickson et al, 1967), (Levy et al, 1972).
Heterozygous F H produce on average one half of the normal number of LDL-
receptors since they only have one normal gene (Brown and Goldstein, 1986). Their
plasma L D L level can be twice the normal level (< 9 mmol/litre) and they start to

82
experience heart attacks in their early 40's if untreated (Mackness and Durrington,
1992), (Kwiterovich et al, 191 A). Being an entirely genetic syndrome, elevated levels
of cholesterol are found throughout childhood and there are good reasons for
commencing at least dietary therapy and other coronary prevention measures in
childhood (Mackness and Durrington, 1992). F H homozygotes have inherited one
abnormal gene for the LDL-receptor from each parent and therefore have little or no
functioning LDL-receptor activity on their tissues. Consequently, they exhibit L D L
cholesterol levels 6 to 8-fold above normal (< 15 mmol/litre) (Goldstein and Brown,
1983) and they typically develop heart disease before the age of 20, often before age
10 (Brown and Goldstein, 1986). If untreated, these individuals generally do not
survive beyond age 30.
The membrane-spanning domain of the LDL-receptor is important for
anchoring the receptor in the cell membrane, and cytoplasmic domain is critical for
proper lateral migration of the receptors to the coated pits where internalisation of
L D L occurs (Brown and Goldstein, 1986). More than 600 mutations including
insertions, deletions, nonsense and missense mutations, have now been described in
the LDL-receptor gene. These affect synthesis, post-transcriptional processing, ligand
binding activity or internalisation of the LDL-receptor (Lombardi et al, 1995),
(Tolleshaug et al, 1983), (Goldstein and Brown, 1983), (Russell et al, 1989).
Four classes of mutation, each affecting a different region of the gene and
therefore a different function of the molecule, have thus far been defined. Class 1
mutations are those in which no receptors are synthesised (LDL-receptor-negative). In
most individuals with these mutations, the amount of m R N A is markedly reduced,
although in some a defective m R N A is present (Hobbs et al, 1988), (Tolleshaug et
al, 1983). Class 2 mutations are those in which the receptor is synthesised but not
properly transported to or through the Golgi complex (Leitersdorf et al, 1989). Class
3 mutations are those where the receptor molecules are inserted into the membrane
but then fail to bind lipoprotein, presumably because of defects in the ligand-binding
domain of the receptor (Hobbs et al, 1989). Class 4 mutations are those where the
receptors move to the cell surface and bind L D L , but then fail to migrate to the coated
pits for internalisation. The defects for this type of mutation have been found in the
cytoplasmic domain of the LDL-receptor (Bilheimer et al, 1979), (Schaefer, 1990).
The heterogeneity of the mutant alleles at the LDL-receptor locus appears to
explain some variability in the clinical course of F H homozygotes. The worst

83
combination of homozygous F H is the combination of two mutations that result in a
failure of receptor expression rather than a defective receptor (Mackness and
Durrington, 1992) and are associated with the most severe clinical course (Goldstein
and Brown, 1983), (Sprecher et al, 1985). Based in functional LDL-receptor assays
in cultured cells, such patients are described as 'receptor-negative' (Goldstein and
Brown, 1983). However, if the cells of homozygotes exhibit some receptor function
(up to 2 5 % of normal), the onset of atherosclerotic complications may be delayed and
life expectancy is somewhat longer. Such patients are called 'receptor-defective'.
Although these mutations produce different phenotypes at the protein level,
they produce a uniform clinical syndrome. All mutations disrupt the efficiency with
which L D L is catabolised, causing the plasma L D L concentration to increase and
produce significant and sustained hypercholesterolemia. This leads to chronic injury
to the endothelial lining of arterial walls and accelerated atheroma formation (Brown
and Goldstein, 1986), (Ross, 1986), (Bilheimer etal, 1979), (Soutaref a/., 1977).
Studies with radiolabelled L D L show that in F H homozygotes, the L D L
particles remain in the blood stream about two and half times longer than they do in
people with normal functioning LDL-receptors (Langer et al, 1972). In addition,
homozygotes actually produce twice as much L D L per day as normal people. This is
because the precursors of LDL, V L D L and LDL also remain in the plasma longer
since they are not removed from plasma through the LDL-receptor, causing an
increase in the production of LDL. This synergistically acts with the decreased
removal of L D L to increase the level of L D L in plasma. This condition illustrates a
causal relationship between elevated plasma L D L concentration and atherosclerosis.
Bilheimer and colleagues (Bilheimer et al, 1984) demonstrated the importance of the
hepatic LDL-receptor in L D L removal. By transplanting livers of homozygous
hypercholesterolemia patients with livers from normal subjects, there was a marked
reduction in the plasma LDL.
One of the most important animal models in the field of hyperlipidemia and
atherosclerosis has been the W H H L rabbit. The W H H L rabbit is a strain of Japanese
rabbit discovered by Watanabe and is characterised by grossly elevated levels of
serum cholesterol, phospholipids, and triglycerides (Watanabe, 1980), (Watanabe et
al, 1981). This animal is deficient in L D L receptors, the absence of which has been
confirmed using cultured skin fibroblasts (Tanzawa et al, 1980) and hepatocytes
(Attie et al. 1981). The genetic defect arises from an in-frame deletion of 12

84
nucleotides, which results in the loss of four amino acids from the ligand-binding
domain of the LDL-receptor (Kita et al, 1981), (Yamamoto et al, 1986-b). This
animal spontaneously develops hypercholesterolemia and atherosclerosis in a way
that is analogous to the situation in humans with F H (Bilheimer et al, 1982).
As a consequence of LDL-receptor deficiency, W H H L rabbits clear L D L from
plasma with reduced efficiency (Suckling and Jackson, 1993). The defect in L D L
catabolism occurs via a reduced affinity for apo B, allowing it to be transported to the
cell surface at only one-tenth the normal rate (Bilheimer et al, 1982), (Bilheimer et
al, 1979), (Soutar et al, 1982), (Kesaniemi et al, 1983), (James et al, 1989), (Pitman
et al, 1982), (Kita et al, 1982-a). These authors have shown that essentially all of the
L D L was catabolised via receptor-independent process(es) in F H and the W H H L
rabbit versus one-third of L D L catabolism in normal counterparts. Attie et al. (Attie et
al, 1981) found that W H H L hepatocytes degrade L D L , and proposed that these cells
also possess a nonsaturable, receptor-independent pathway for the catabolism of LDL.
In addition to increased levels of L D L , these animals have been found to display
increased concentrations of DDL and V L D L , both of which are enriched in cholesterol
and apo B-100 (Havel et al, 1982). W H H L rabbits are also characterised by low
concentrations of H D L and increased concentrations of apo E in plasma. A decreased
rate of apo E catabolism would be expected in the absence of the LDL-receptor,
which binds apo E-containing lipoproteins as well as these containing apo B (Brown
etal, 1981).
The clearance of L D L in F H patients and W H H L rabbits is severely impaired
but both are said to have normal clearance of chylomicrons. Havel et al. (Havel et al,
1982) found only trace amounts of apo B-48 in plasma of W H H L rabbits, consistent
with efficient hepatic clearance of chylomicron remnants. However, Bowler et al.
(Bowler et al, 1991) clearly demonstrated defective chylomicron remnant clearance
in homozygous W H H L rabbits, and delayed clearance in heterozygous W H H L
rabbits, compared with controls. The presence of defective LDL-receptors and a slow
rate of plasma chylomicron removal were found to be factors involved in
atherosclerosis from studies using the W H H L rabbits (Bowler et al, 1991), (Mamo et
al, 1991). In addition, Dane-Stewart et al. (Dane-Stewart et al, 2001) recently found
that heterozygous F H subjects had significantly elevated plasma concentrations of
apoB-48 and remnant-like particle-cholesterol compared with controls. The findings
suggest that patients with heterozygous F H have elevated plasma concentrations of

85
T R L remnants, including those of intestinal origin, and may be a consequence of
decreased clearance of these particles by the LDL-receptor.
Thus the cause of F H has been accepted as defective receptor-mediated L D L
catabolism. Therefore, L D L defects effect chylomicron remnant metabolism and
clearance and early intervention and diagnosis can be extremely helpful in terms of
therapy approach. Current diagnosis of F H relies primarily on accurate measurement
of plasma L D L cholesterol and triglyceride levels in the fasting state. Other tools for
diagnosing F H include: specific testing of LDL-receptor function on cultured
cutaneous fibroblasts or circulating peripheral lymphocytes (Cuthbert et al, 1986),
and testing of LDL-receptor function on cells cultured from amniotic fluid obtained
by amniocentesis (Brown et al, 1978). A non-radioactive dot blot assay with L D L
conjugated to colloidal gold is also available. This has been used to measure the LDL-
receptor activity of unstimulated human circulating mononuclear cells by Roach et al
(Roach et al, 1993b), w h o showed that the high plasma cholesterol of F H subjects
was related to a low in vivo LDL-receptor activity. Detection of abnormal genes for
the LDL-receptor has also been demonstrated, by analysis of D N A obtained from
leucocytes (Sudhof et al, 1985). A n enormous number of different mutations can
occur in the syndrome making a D N A marker for F H generally applicable outside
individual kindred's an unlikely possibility. Hence there is a continuing reliance on
clinical features for F H diagnosis (Mackness and Durrington, 1992).
1.5.4.2 Coronary Artery Disease (CAD)
Barritt (Barritt, 1956.) first suggested the association between postprandial
hyperlipidemia and C A D . Zilversmit then hypothesised in 1979 that plasma
chylomicrons in persons who ingest a cholesterol-rich diet are atherogenic
(Zilversmit, 1979). Since then, interest in chylomicrons has been increasing, in part
owing to the realisation that current concepts of lipoprotein metabolism are
inadequate to account completely for the development of arteriosclerosis (Redgrave,
1999). Epidemiological studies have repeatedly shown significant degrees of
correlation between intake of cholesterol and other lipids with the prevalence of C H D
in various populations (Grundy et al, 1982), (McGee et al, 1984), (Keys, 1988), and
it is apparent that atherosclerosis is a long-term disease ( M a m o et al, 1998a). Animal
studies have shown that experimentally induced hypercholesterolemia is followed by

86
the accumulation of cholesterol in the large arteries, and high levels of cholesterol-
rich chylomicron remnants. Zilversmit (Zilversmit, 1979) supported the view that
arterial lipid accumulates as the result of not only abnormally high concentrations of
L D L in the blood plasma, but also as a consequence of the normal process of lipid
absorption and transport. This normal process may be innocuous in persons who
ingest a low-fat diet, but is probably pathogenic in those who consume a diet rich in
fat and cholesterol.
Perhaps the most promising evidence for the involvement of chylomicrons and
their remnants in the pathogenesis of atherosclerosis, is a number of studies that have
found delayed clearance of postprandial lipoproteins in normolipidemic subjects with
C A D (Maranhao et al, 1996), (Groot et al, 1991), (Karpe et al, 1994), (Weintraub et
al, 1996). Approximately 3 0 % of patients with C A D do not have elevated levels of
fasting lipoproteins, as measured by total plasma cholesterol or triglyceride
concentrations (Genest et al, 1991). Wieland et al. (Wieland et al, 1990) investigated
the metabolism of postprandial lipoproteins in 4 male patients who suffered from
premature (< 60 yrs) angiographically proven C H D . They had no remnant to low
plasma total and L D L cholesterol levels. Using the vitamin A fat loading test they
were able to show that the clearance of chylomicron remnants was delayed compared
with 3 healthy control subjects. Their data strongly favour the concept that defective
removal of postprandial lipoproteins leads to the development of C H D in these
patients, and that these postprandial lipoproteins may be atherogenic. Additionally, fat
challenge tests in subjects with normotriglyceridemic profiles in the fasting state have
been shown to independently predict coronary disease (Groot et al, 1991), (Patsch et
al, 1992), (Simons etal, 1987).
Data from the M R F I T study of C H D in men show that 5 2 % of myocardial
infarction patients have a plasma cholesterol concentration within the accepted range
(Martin et al, 1986). Kinlay (Kinlay, 1988) also questioned the practice of cholesterol
screening and its effectiveness, when results of an Australian survey into C H D
revealed that 3 0 % of deaths occurred in patients with plasma cholesterol levels within
an acceptable range. These studies also implicate a role for chylomicron remnants in
the development of C H D (Zilversmit, 1979), (Simionescu and Simionescu, 1991),
(Keys, 1988), (Mamo et al, 1997), (Steiner, 1993), (Groot et al, 1991).

87
1.5.4.3 Type III Dysbetalipoproteinemia
Another classical human genetic model of dyslipoproteinemia that supports the
atherogenicity of remnant particles is Type III dysbetalipoproteinemia. Type III is a
defective ligand-receptor interaction due to a genetic abnormality in apo E (E2/E2
homozygosity). Apo E 2 has a lower binding affinity than apo E3, E 4 for the LDL-
receptor (Schneider et al, 1981). Lipoproteins utilising this protein as the ligand are
cleared inefficiently, and subjects who possess the apo E2/E2 phenotype have
significantly delayed chylomicron remnant metabolism (Weintraub et al, 1987a),
(Hazzard and Bierman, 1976), (Kane, 1983), (Mahley and Angelin, 1984), (Conner et
al, 1987). Approximately 1 % of the population is apo E2/E2 homozygous and
approximately 1 in 50 of these individuals develop Type III hyperlipoproteinemia,
which is characterised by fasting hyperlipidemia (Mahley and Angelin, 1984).
Atherosclerosis is prevalent in individuals with Type III because the clearance of
chylomicron remnants and IDL is severely impaired (Thompson, 1989), (Mahley and
Rail, 1989). Further evidence is provided by studies showing only slightly improved
remnant clearance in Type Ul Hyperlipoproteinemic patients after up-regulation of the
LDL-receptor using Lovastatin (Gylling et al, 1995).
1.5.4.4 Diabetes
Diabetes mellitus and its associated hypertriglyceridemia are strongly associated with
the early onset of plaque formation and development of atherosclerotic vascular
disease (Steiner, 1997), (Howard, 1987). The reasons for the high risk of
atherosclerosis associated with diabetes are multi-factorial, however a defect in
lipoprotein metabolism is considered to be primary factor in the aetiology of vascular
disease (Steiner, 1997), (Bar-on et al, 1984), (Nordestgaard and Tybaerg-Hansen,
1992), (Kostner and Karadi, 1988), (Howard, 1987). Indeed, a number of studies have
demonstrated a decrease in the clearance of remnant lipoproteins in
hypertriglyceridemic and normotriglyceridemic type II diabetic patients (Lewis et al,
1991), (Chen et al, 1993), (Cavallero et al, 1995), (Durlach et al, 1996), (Reznik et
al, 1996), (Curtin et al, 1994), 1994). Additionally, it has been demonstrated that the
postprandial levels of triglyceride-rich lipoproteins in diabetic populations are higher
compared with control populations (OMeara et al, 1992).

88
Apo E is the ligand responsible for interaction of remnants with the L D L -
receptor, and chylomicrons and their remnants produced by streptozotocin-induced
diabetic rats have been shown to be depleted of apo E (Levy et al, 1985). Redgrave
and Callow (Redgrave and Callow, 1990) also found a reduction in the amount of apo
E associated with chylomicron-like emulsions in streptozotocin-induced diabetic rats
and a decrease in the clearance of remnant lipoproteins (Redgrave et al, 1991),
(Redgrave and Callow, 1990). Utilising chylomicron remnant-like emulsions labelled
with [l-14Cloleate, Redgrave et al. (Redgrave et al, 1995) found that the appearance
of 1 4C02 label in the breath in diabetic rats and LDL-receptor-deficient mice was
delayed compared with their respective controls. Thus the impaired clearance of
chylomicron remnants in diabetic patients may be a result of reduced/compromised
interaction of chylomicron remnants with the LDL-receptor, as a result of attenuated
apo E concentrations and reduced LDL-receptor expression.
Recently w e postulated that receptor expression might be decreased and high-
affinity uptake of remnant lipoproteins compromised with insulin deficiency, whether
absolute (IDDM) or relative (NI D D M ) . A study in our laboratory assessed particle
uptake of chylomicron-like lipid emulsions in control and alloxan-diabetic rabbits, as
a possible cause of altered lipoprotein metabolism. The data suggests that both
triglyceride secretion rate and L P L activity were unaltered by alloxan-induced
diabetes. Rather, the observed hypertriglyceridemia may be due to a delay in plasma
clearance of triglyceride-rich lipoproteins. W e concluded that the reduction in
emulsion remnant clearance observed in diabetic rabbits was due to decreased
chylomicron remnant particle uptake via uptake mechanisms (Gennat et al,
unpublished observations).
1.5.4.5 Hypothyroidism
Hypothyroid patients, characterised by reduced plasma thyroxine concentrations, are
at elevated risk of C A D . Thyroid status has been shown to regulate LDL-receptor
expression, therefore the metabolism of remnants would be expected to be
compromised in hypothyroid patients. Redgrave and co-workers (Redgrave et al,
1991) found the clearance of chylomicron remnants to be delayed in methimazole-
induced hypothyroid rats. Thyroxine replacement therapy has also been shown to
have a positive effect on the fasting plasma lipoprotein profile, suggesting an increase

89
in LDL-receptor expression (Mishkel and Crowther, 1977), (Arem and Patsch, 1990),
(Packard etal, 1993), (Zerbinatti etal, 1991).
1.5.4.6 Obesity
Obesity is a significant risk factor for atherosclerosis, and is often a precursor to
diabetes. Visceral obesity is also considered an important risk factor and may promote
insulin resistance (Wilson et al, 1987). Smith et al. (Smith et al, 1999) have shown a
delay in chylomicron remnants metabolism, as demonstrated by the vitamin A fat load
test and apo B-48 concentration. The vitamin A fat-load method has been utilised for
the purpose of characterising chylomicron remnant catabolism in human subjects
(Berr, 1992), (Berr and Kern Jnr, 1984), (Borel et al, 1998). This method has been
used to detect abnormalities in postprandial lipid clearance in disease states such as
chronic renal failure (Wilson et al, 1985a) and hypertriglyceridemia (Wilson et al,
1985b), hyperlipidemia (Sprecher et al, 1991), (Weintraub et al, 1987b), and
hyperbetalipoproteinemia (Genest et al, 1986). Delayed clearance of chylomicron
remnants has also been documented in patients with secondary hyperlipidemias and
this has been associated with increased risk of coronary disease.
Chylomicron remnant metabolism may be impaired as a result of defective
clearance via the LDL-receptor or increased synthesis in the intestine as a primary and
secondary hyperlipidemia, in a number of disease states. A delay in clearance would
result in elevated concentrations of circulating postprandial lipoproteins, and increase
the exposure, flux and finally, deposition of chylomicron remnants into the arterial
wall. Add to this that humans are in a postprandial state for most of their lives, hence
chylomicrons are continually being synthesised, even in the absence of dietary fat.
1.6 Animal Models for Atherosclerosis
Animal models have been used extensively for studying lipoprotein metabolism, and
the rabbit was the first animal to be used as a model for atherosclerosis (Saltykow,
1908), (Ignatowski, 1909). The principal advantage of the use of the rabbit as a model
has been the rapidity with which the animal becomes hypercholesterolemic and hence
atherosclerotic when fed a variety of natural or semi-synthetic diets (Clarkson, 1963),
(Clarkson, 1972), (Kritchevsky, 1969), (Carroll, 1971). With suitable manipulation of

90
the rabbit through dietary and hormonal means, lesions resembling those of the
human type can be produced (Constantinides, 1961), (Clarkson et al, 1970). Further
support for the use of the rabbit as a model for the study of atherogenesis appeared
when it was shown that rabbits use the same method for the transport of lipid into
atherosclerotic plaque as occurs in human aortic lesions (Walton, 1973), utilising
C E T P for transfer of lipoprotein cholesterol. In man and rabbits, C E T P plays a central
role in the transfer of cholesteryl esters from H D L to other lipoproteins, whereas the
rat has low or absent levels of C E T P activity (Ha and Barter, 1982). Rabbits treated
with a neutralising monoclonal antibody to C E T P demonstrated increased cholesteryl
ester in H D L , and cholesteryl ester clearance from plasma was delayed (Whitlock et
al, 1989).
However, as in man, there are considerable inter- and intra-strain variations in
the susceptibility of rabbits to both hypercholesterolemia and atherosclerosis.
Differences between sexes of rabbits have also been observed; male rabbits have been
shown to have significantly lower plasma cholesterol concentrations than female
rabbits (Laird et al, 1970), (Fillios and Mann, 1956.). Despite these disadvantages the
rabbit has continued to be used for such purposes. The W H H L rabbit provides a
unique opportunity to determine the consequences of LDL-receptor deficiency for
many aspects of lipoprotein metabolism (Havel et al, 1982), including a model for
quantitating receptor activity.
The availability of the technology to transfer genes associated with lipid
metabolism into mice has resulted in a rapid growth in publications describing
transgenic mice with properties of significance to human lipid metabolism (Suckling
and Jackson, 1993). The mouse is the best mammalian system for the study of genetic
contributions to disease. This is because of the easy breeding, short generation time,
and availability of inbred strains. Unfortunately the mouse is highly resistant to
atherosclerosis. The introduction of genetically engineered mouse models has helped
to increase our knowledge of how certain proteins affect atherosclerosis (Breslow,
1996).
For the purpose of using mouse models for lipoprotein metabolism and
atherosclerosis, lipoprotein transport genes are either been added to the germ line of
mice by transgenic techniques or knocked out by homologous recombination in
embryonic stem cells. The resultant over- or under-expression of these genes has
resulted in new insights about how these genes function in the body and their role in

91
lipoprotein metabolism (Breslow, 1993). Genetically manipulated animals overcome
some of the more c o m m o n problems by isolating the cause of atherosclerotic lesions
and providing a model for specifically targeting the problem, and reduce the
variability inherent in the use of animals. These animal models provide a more
realistic and specific model for human lipoprotein disorders and atherosclerosis and
the more c o m m o n genetic mutations that occur in society (Breslow, 1993).
Thus homozygous apo E knockout mice have provided a new model of
atherosclerosis (Zhang et al, 1992), (Plump et al, 1992). Homozygous apo E
knockout mice have cholesterol levels of 10.3-12.9 mmol/1 on a chow diet. W h e n fed
a western-type diet (0.15% cholesterol and 2 0 % fat), they respond with cholesterol
levels of approximately 46.3 mmol/L O n both diets, triglyceride levels are minimally
elevated (Breslow, 1993). This would suggest a defect in the uptake of lipoprotein
remnants, rather than a deficiency in hydrolysis of lipoprotein particles. Metabolic
studies indicate a severe defect in lipoprotein clearance from plasma, as predicted
from the known function of apo E as a ligand for lipoprotein receptors. Extensive
complex arterial plaques have formed after being placed on a normal laboratory chow
diet (0.01% cholesterol) or a western-type diet (0.15% cholesterol). In addition, both
plasma apo E and apo B-48 levels were markedly elevated but apo B-100 levels
remained unaffected (Plump et al, 1992), (Zhang et al, 1992), (Piedrahita et al,
1992). The results suggested that L D L metabolism was unimpaired, however the
delayed clearance of chylomicron remnants was an important factor involved in the
promotion of atherosclerosis in these mice. The single genetic lesion causing E
absence and severe hypercholesterolemia is sufficient to convert the mouse from a
species that is highly resistant to one that is highly susceptible to atherosclerosis
(Breslow, 1993).
The advantages of animal models for the study of atherosclerosis include
short-term induction of experimental atherosclerosis and control of dietary and
environmental factors. In addition, data from a variety of animal models can be
integrated to allow for the collection of complementary data/findings, hypotheses
relating to the aetiology of atherosclerosis can be tested, and pre-clinical testing of
diagnostic procedures and chemical/drug intervention for prevention of
atherosclerosis can be carried out. One main disadvantage to the use of animal models
is that the results obtained in animals may not be validly applied to man. Despite the
fact that in all animal models with spontaneous atherosclerosis, hypercholesterolemia

92
produced by dietary cholesterol exacerbates the lesion (Clarkson, 1972),
atherosclerotic lesions have been induced by hypercholesterolemia in species that do
not naturally develop the lesions (Clarkson et al, 1970).
1.7 Current Screening for Cardiovascular Risk
Considerable interest in lipoprotein metabolism has been generated in the past few
decades because of the apparent correlation between the levels of serum lipoproteins
and atherosclerosis (Brown and Goldstein, 1984). The cholesterol of atherosclerotic
plaques is thought to be derived from L D L lipoprotein particles that circulate in the
blood stream and there is a strong positive relationship between the level of L D L in
the plasma and the risk of C A D (McNamara et al, 1987), (Nordoy and Goodnight,
1990).
Plasma H D L are cholesterol rich particles, however evidence suggests that
H D L play an important role in the movement of cholesterol from non-hepatic tissues
to the liver for catabolism and excretion. This process is known as "reverse
cholesterol transport" (Glomset, 1968). High levels of H D L appear to be
antiatherogenic since it acts as a scavenger of surplus free cholesterol from other
lipoproteins and also from cells of the arterial wall where atherogenic plaques form
(Gordon et al, 1977), (Nordoy and Goodnight, 1990). Therefore, there is a strong
negative relationship between the level of H D L in the plasma and the risk of C H D .
This relationship is independent of plasma L D L , other lipoproteins such as V L D L and
other risk factors such as obesity, smoking and blood pressure.
It is now known that the hypercholesterolemia can be adequately controlled to
reduce the risk of developing premature atherosclerosis. Due to the fact that C H D is
relatively common, it is important that physicians be alert to its manifestation and
diagnose it early in its course, when appropriate therapy can be instituted to prevent or
retard the progression atherosclerosis and its clinical sequelae. The lipid and
lipoprotein parameters that are predominantly measured and effectively comprise the
traditional lipoprotein profile include total cholesterol, H D L cholesterol, L D L
cholesterol, and triglyceride (Myers et al, 1994). Final classification and potential
intervention is ultimately based on the measurement of L D L cholesterol.
The current approach is to identify individuals with elevated L D L cholesterol
values, by screening fasting serum or plasma cholesterol levels. In screening for all

93
disorders of lipid transport or to document whether L D L cholesterol levels are indeed
elevated, the measurement of plasma or serum cholesterol triglyceride, and H D L
cholesterol on samples obtained after a 12 to 14 hour fast is taken. Sixty to seventy-
five percent of total plasma cholesterol is transported on L D L . Therefore, at all ages,
the total plasma cholesterol usually is a reflection of L D L cholesterol values. If lipid
values are within the normal range, usually no further lipoprotein workup is required
(Schaefer, 1990). The maintenance of cellular cholesterol homeostasis is undoubtedly
the result of two opposing processes: the ability of L D L to deliver cholesterol to
arteries and the ability of H D L to remove excess cholesterol from them. Therefore, it
is the ratio of plasma L D L / H D L and not just plasma L D L levels alone which is
perhaps the best predictor for the development of C A D (Nordoy and Goodnight,
1990).
Plasma lipids and lipoproteins are generally measured in the fasting state, and
treatment strategies for prevention of cardiovascular disease are based on such
measurements despite the fact that most of our lives are spent between the
consumption of regular meals (Karpe, 1999). Fasting levels of these lipoproteins do
not, however, sufficiently discriminate between patients with and without coronary
disease (Mamo, 1995). Given that humans spend most of their time in the
postprandial state, this approach is not always the most effective.
Chylomicron remnant dyslipidemia has now been identified in a number of
primary and secondary lipid disorders (Mamo et al, 1998a), (De M a n et al, 1996),
(Tomono et al, 1994) and clearly it has become important to determine remnant
metabolism. In terms of substantiating that chylomicrons and their remnants do
contribute to the pathogenesis of atherosclerosis, the progress has been slow. This is
in part due to the fact that the measurement of chylomicron particles in vivo is
difficult, as they clear rapidly from plasma (half-life of ~ 5 min) and have the fastest
turnover in plasma of all lipoproteins (Redgrave, 1999). A similar quantity of
cholesterol passes through the plasma in chylomicrons and L D L on a daily basis, yet
chylomicrons contribute a small amount to total plasma cholesterol, at any time. Thus
an increase in the contribution of chylomicron cholesterol to total plasma cholesterol
may not be evident, except in cases where plasma cholesterol is grossly abnormal
(Redgrave, 1999). Furthermore, chylomicrons and their remnants cannot be separated
exclusively from other lipoproteins by routine density gradient ultracentrifugation
because they float in the same density range as V L D L , IDL, L D L and H D L .

94
Several tests are routinely used to assess post-prandial lipoprotein kinetics.
Elevated fasting plasma triglyceride and post-prandial triglyceride response have been
widely used as markers for the metabolism of chylomicrons. However, assessment of
the concentration of chylomicrons and their remnants in a clinical setting requires the
measurement of a marker that is specific for postprandial lipoproteins. Currently two
methods are available for the determination of chylomicrons and their remnants
during the postprandial phase. The first is the use of a fat-load containing vitamin A,
which acts as an indirect marker of chylomicron metabolism. The second is the
quantitation of the protein apo B-48, a marker specific for chylomicrons.
1.7.1 Vitamin A Fat Load Test
The term vitamin A is used as a generic description for all derivatives having the
same (6-ionine) ring structure and the same biological activity as retinol, excluding
provitamin carotenoids. Retinol is a low molecular weight, fat-soluble compound that
can partition into membranes. Vitamin A occurs naturally only in animals. In most
animal tissues, the predominant retinoid is retinyl palmitate, but retinyl oleate and
retinyl stearate are also found. If present in excessive amounts, retinol can disrupt
normal membrane structure and function. Therefore, vitamin A in excess of
immediate tissue requirements is stored as the ester of retinol with long-chain fatty
acids; the primary site of such storage is the liver. To be transported through the
aqueous environment and to limit its level in membranes, retinol is normally either
bound to proteins both extracellularly and intracellularly, is esterified to long chain
fatty acids for transport in lipoproteins, or is stored in cytoplasmic lipid droplets
(Blomhoff et al, 1991), (Blomhoff, 1994).
Several researchers suggested that retinyl esters might be an appropriate
endogenous label for chylomicron remnants. The rationale for this approach is based
on the metabolic fate of ingested vitamin A (Hazzard and Bierman, 1976), (Ross and
Zilversmit, 1977), and the fact that dietary retinol within the recommended safe range
does not increase serum retinol levels. In the intestinal mucosa, absorbed retinyl
palmitate is hydrolysed to retinol by several enzymes and solubilised by bile salts
before being absorbed by the enterocyte by diffusion (Weber, 1981), (Blomhoff,
1994). In the enterocyte, retinol is esterified with long-chain fatty acids by acyl CoA:
retinol acyltransferase and lecithin: retinol acyltransferase (Helgerud et al, 1983),

95
(MacDonald and Ong, 1988). It is then incorporated into the core of chylomicrons,
secreted into intestinal lymph, and remains largely within the remnant particle during
triglyceride lipolysis. The resultant chylomicron remnants still contain the retinyl
esters and are taken up irreversibly via receptor-mediated endocytosis by liver
parenchymal cells (Goodman et al, 1965), (Goodman et al, 1966), (Huang and
Goodman, 1965), (Sherrill and Dietschy, 1978), (Sherrill et al, 1980), (Goodman,
1980), (Nagasaki et al, 1994), (Havel, 1994), (Haghpassand and Moberly, 1995).
In the liver, retinyl esters are hydrolysed to vitamin A alcohol (retinol) at the plasma
membrane or in early endosomes, probably by retinyl ester hydrolase (Harrison and
Gad, 1989). Retinoids serve as the body's reserve of vitamin A and are used to
maintain a steady state concentration of vitamin A in plasma. Retinol is either stored
in fat-storing cells (Blomhoff et al, 1982), (Blomhoff et al, 1984) or transferred to
the E R and resecreted as unesterified retinol bound to a specific carrier protein called
plasma retinol-binding protein (RBP) (Blomhoff, 1994), (Goodman, 1980), (Kanai et
al, 1968), (Smith et al, 1973), (Smith and Goodman, 1979). After removal of
esterified retinol by the liver, it does not recirculate (Berr and Kern Jnr, 1984),
Thompson et al. 1983). Retinyl esters have to be hydrolysed to retinol for their
delivery into the blood. Serum retinol levels are homeostatically maintained within a
narrow range by R B P , which delivers it to target tissues. Surface receptors for R B P
have been found on many cells, including intestinal cells, epithelial testicular cells and
interstitial cells. Specific binding proteins known as cellular R B P have been detected
in these tissues and seem to be responsible for binding retinol once it has entered the
cell (Haghpassand and Moberly, 1995), (Nagasaki et al, 1994). The formation and
hydrolysis of retinyl esters are key processes in the metabolism of the fat-soluble
micronutrient vitamin A. Long-chain acyl esters of retinol are the major chemical
form of vitamin A (retinoid) stored in the body. Although retinyl esters are found in a
variety of tissues and cell types, up to 8 0 % of the body's total retinol is present in the
liver; mainly located in stellate (Ito) cells (Norum and Blomhoff, 1992) in the liver.
Retinyl esters with various fatty acids packed together in cytoplasmic lipid droplets
(Redgrave and Vakakis, 1976) represent the major storage forms. Thus, these esters
represent the major endogenous source of retinoid that can be delivered to peripheral
tissues for conversion to biologically active forms (Harrison, 2000). The major
pathways for retinoid transport and storage in the body are shown in Figure 1.1.

96
Harrison et al. (Harrison et al, 1995) studied chylomicron remnant
[3H]retinyl ester metabolism during and after uptake into the liver of rats. Labelled
retinyl esters were rapidly cleared from plasma (half time - 1 0 min) and appeared in
the liver. The co-localisation of both neutral and acid, bile salt-independent retinyl
ester hydrolase and labelled esters in plasma membrane/endosomal fractions within
the liver suggests a probable role for these enzymes in the initial hepatic metabolism
of chylomicron remnant retinyl esters. This conclusion was supported by the
observation that plasma membrane/endosomal fractions were active in catalysing the
hydrolysis of chylomicron remnant retinyl esters in vitro.
Vitamin A is required for vision, reproduction, foetal development and the
development and maintenance of differentiated tissues. It also plays an important role
in reducing infectious disease morbidity and mortality by enhancing immunity, an
effect that is partly mediated by macrophages. Hence, knowing how these cells take
up vitamin A is important. Hagen et al. (Hagen et al, 1999) have shown that
macrophages efficiently take up chylomicron remnant retinyl esters and RBP-bound
retinol by specific and saturable mechanisms. L P L increased the binding of
chylomicron remnant [3H|retinyl ester by approximately 3 0 % and the uptake of
chylomicron remnant [3H]retinyl ester by more than 300%. The authors concluded
that both LDL-receptor and L D L R P are involved in the uptake of chylomicron
remnant [3H]retinyl ester in macrophages. Van Bennekum et al. (van Bennekum et
al, 1999a) also hypothesised that L P L contributes to this extrahepatic clearance of
chylomicron vitamin A, and investigated the distribution of uptake of [3H]retinyl
ester-containing rat lymph chylomicrons after injection into mice. Based on the sites
of uptake in wild type and LPL-null mice, they concluded that L P L expression does
influence accumulation of chylomicron retinoid in extrahepatic tissues, with greater
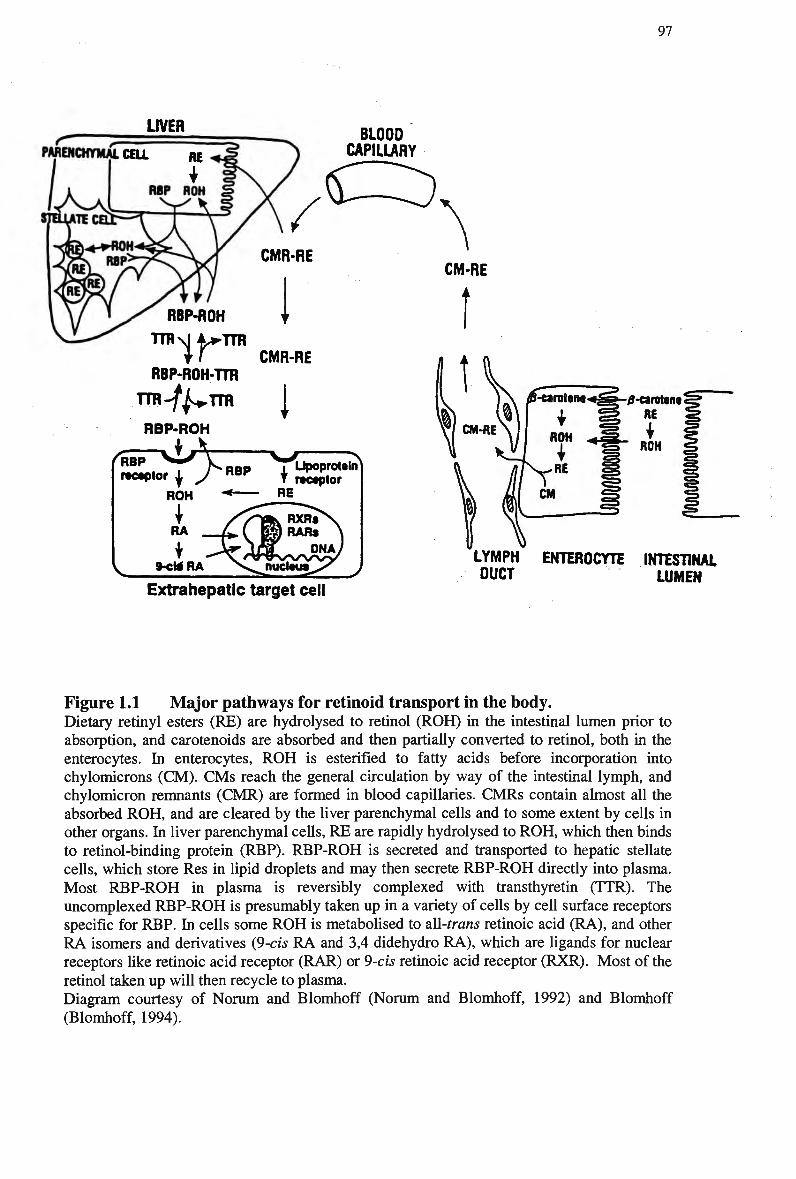
97
Extrahepatic target cell
Figure 1.1 M a j o r pathways for retinoid transport in the body. Dietary retinyl esters (RE) are hydrolysed to retinol (ROH) in the intestinal lumen prior to absorption, and carotenoids are absorbed and then partially converted to retinol, both in the enterocytes. In enterocytes, R O H is esterified to fatty acids before incorporation into chylomicrons (CM). C M s reach the general circulation by way of the intestinal lymph, and chylomicron remnants ( C M R ) are formed in blood capillaries. C M R s contain almost all the absorbed R O H , and are cleared by the liver parenchymal cells and to some extent by cells in other organs. In liver parenchymal cells, R E are rapidly hydrolysed to R O H , which then binds to retinol-binding protein (RBP). R B P - R O H is secreted and transported to hepatic stellate cells, which store Res in lipid droplets and may then secrete R B P - R O H directly into plasma. Most R B P - R O H in plasma is reversibly complexed with transthyretin (TTR). The uncomplexed R B P - R O H is presumably taken up in a variety of cells by cell surface receptors specific for RBP. In cells some R O H is metabolised to all-trans retinoic acid (RA), and other R A isomers and derivatives (9-cis R A and 3,4 didehydro R A ) , which are ligands for nuclear receptors like retinoic acid receptor (RAR) or 9-cis retinoic acid receptor (RXR). Most of the retinol taken up will then recycle to plasma. Diagram courtesy of Norum and Blomhoff (Norum and Blomhoff, 1992) and Blomhoff
(Blomhoff, 1994).

98
L P L activity increasing the amount of retinoid taken up from chylomicrons and/or
their remnants. In addition, it has been found by Myhre et al (Myhre et al, 1998) that
following uptake of chylomicron remnant retinyl esters by the macrophage cell line
J774, the retinyl esters are hydrolysed to retinol before retinol is further metabolised
to retinal and the various retinoic acid isoforms. The authors concluded that this
physiologic plasma transport molecule for vitamin A that might be covalently linked
to proteins.
The capacity to transport retinyl esters through the plasma compartment is
considered a unique attribute of chylomicrons and their remnants. That property, and
the hypothesis that retinyl esters are conserved during chylomicron catabolism,
constitutes the theoretical basis for their use as markers of lipoproteins originating in
the intestine (Sprecher et al, 1991), (Hazzard and Bierman, 1976), (Zilversmit, 1978),
(Ross and Zilversmit, 1977), particularly in disease states (Havel, 1994). The most
commonly used approach to measure chylomicron metabolism in humans involves the
incorporation of retinyl esters into the chylomicron particle via a fat load. This
procedure is based on the observation that dietary retinol is esterified in intestinal
cells, packaged in the chylomicron core (Goodman et al, 1966). The vitamin A fat
load test utilises orally administered retinyl palmitate, which is incorporated within
the core of the chylomicron on packaging in the enterocyte, and secreted into the
intestinal lymph (Lovegrove et al, 1999), (Berr and Kern Jnr, 1984).
As chylomicrons undergo lipolysis to become remnants the retinyl ester
remains within the particles until their removal as remnants by the liver, and are not
resecreted in hepatic-derived lipoproteins in humans. Hence the concentration in
plasma reflects the combination of secretion and clearance (Goodman, 1980), (Smith
et al, 1999). Therefore, by monitoring the concentration of plasma retinyl ester or
retinyl ester associated with the chylomicron fraction, information about the
appearance and clearance of chylomicrons can be determined (Barrett, 1998).
Most laboratories measure retinyl ester in a density range of < 1.006 g/ml
because at greater densities the vitamin is suggested to represent transfer to more
dense lipoproteins and not indicative of chylomicron remnant concentrations ( M a m o
et al, 1998b). However, more recent studies in vitro and in vivo suggest that esterified
vitamin A is not subject to transfer between lipoproteins and that its isolation at
densities greater > 1.006 g/m represents the generation of small dense postprandial
remnants (Peel et al, 1993), ( M a m o et al, 1998a). In fact in situations where high

99
affinity uptake is compromised one would predict increased appearance of remnants
in more dense plasma fractions as a consequence of increased interaction with
lipolytic enzymes. Given that it is the small cholesterol-rich chylomicron remnants
that are considered to be most atherogenic, it is important to ensure their complete
recovery by isolating lipoproteins at sufficiently dense gradients ( M a m o et al,
1998b).
Central to the use of retinyl esters as an appropriate marker is the fact that it
does not exchange or become transferred to other non-intestinally derived lipoprotein
classes. Clearly a significant exchange of chylomicron core components with other
lipoproteins impacts upon the analysis and interpretation of the retinol based data
(Barrett, 1998). However, some studies indicate that it might not be an ideal marker
for chylomicrons since retinyl palmitate transfers to H D L and L D L (Zilversmit et al,
1982). Cohn et al (Cohn et al, 1993) observed that up to 2 5 % of retinyl palmitate
was contained in the apo B-100 triglyceride-rich lipoproteins. This retinyl palmitate
fraction appeared at late time points, which suggests that it derived from the transfer
of core lipids between lipoproteins. However, results from specific quantification of
chylomicron remnants and V L D L of varying particle size are sparse (Zilversmit and
Shea, 1989). Furthermore, Karpe et al. (Karpe et al, 1995) suggested that
chylomicrons and chylomicron remnants might not be uniformly labelled with retinyl
esters after a vitamin A fat load meal, with smaller particles carrying fewer retinyl
ester molecules. This hypothesis was based on in vivo turnover studies in healthy
human subjects, and the authors proposed the number of retinyl ester molecules
incorporated into the core of the particle depended on the very early uptake of large
chylomicron remnants and the intestinal secretion of small chylomicrons. The vitamin
A fat load approach has also been criticised by Krasinski et al. (Krasinski et al,
1990a) because retinyl palmitate is transferred from chylomicrons to other circulating
lipoprotein particles in human subjects. Hence this molecule progressively becomes a
less-selective marker of chylomicronemia as time passes after ingestion of the fat-rich
meal containing retinol.
While many researchers dispute the specificity and reliability of retinyl esters
as a marker for chylomicron remnants, most of these describe transfer of retinyl esters
to other lipoprotein classes at later time points. Krasinski et al. (Krasinski et al,
1990b) investigated postprandial vitamin A metabolism by measuring retinyl ester,
triglyceride and apo B-48 in the plasma lipoproteins of human subjects before and

100
after fat feeding. In triglyceride-rich lipoproteins of d< 1.006 g/ml, retinyl esters
similarly peaked at 6 hrs, whereas triglyceride as well as apo B-48 peaked at 3 hours.
Although retinyl esters were found mainly in triglyceride-rich lipoproteins in the
initial postprandial period (84% 3 hrs, 8 3 % 6 hrs), in fasting and postprandial plasma
a large percentage of plasma retinyl esters were in L D L density fraction (44% fasting,
9 % 3 hrs, 9 % 6 hrs, 1 9 % 9 hrs, 3 2 % 12 hrs). A small percentage of retinyl esters were
also found in postprandial H D L ( 2 % to 7 % ) . The authors concluded that retinyl esters
do not always serve as markers for intestinal apoB-48-containing triglyceride-rich
lipoproteins in fasting or postprandial plasma. The authors expressed the amount of
retinyl ester in L D L as a percentage of retinyl ester in total plasma and found that as
much as a third of total retinyl esters were contained in the L D L fraction in fasting
plasma.
Alternately, Berr and co-workers (Berr and Kern Jnr, 1984), (Berr et al, 1985)
found that retinyl palmitate to be a stable label for the core of chylomicrons and their
remnants, with only 5-7% of retinyl palmitate transferring from chylomicrons and
V L D L to L D L during postprandial lipemia or during in vitro incubation. Wilson and
co-workers also analysed the distribution of retinyl esters in plasma lipoproteins of
humans after a vitamin A fat load meal, and studied the in vitro transfer of rat
chylomicron retinyl esters in plasma (Wilson and Chan, 1983), (Wilson et al, 1983).
These investigators considered the amount of retinyl ester in L D L to be insignificant
when compared with the amount of retinyl ester absorbed, and are limited over an 8 hr
period. Hazzard and Bierman (Hazzard and Bierman, 1976) further proposed that
after uptake of chylomicron remnants by the liver, the retinyl palmitate is not
incorporated into V L D L but enters a storage pool. Ross and Zilversmit (Ross and
Zilversmit, 1977) found that retinyl palmitate was less subject to protein-mediated
transfers than cholesteryl esters. Thus retinyl palmitate detected in plasma represents
intestinally derived lipoproteins only. Blomhoff et al. (Blomhoff et al, 1982)
concluded that retinyl palmitate is transported in the core of chylomicrons and their
remnants in plasma and removed with the remnant particle, after monitoring clearance
of radiolabeled retinyl palmitate. Recently, it was shown that even patients who have
a very protracted residence time for chylomicrons (Type I Hyperlipidemics)
demonstrate very little movement of retinyl ester to other lipoproteins (Sprecher et al,
1991).

101
Despite a number of studies indicating that retinyl palmitate transferred to
H D L and L D L , Martins et al. (Martins et al, 1991) found no transfer of the retinyl
palmitate label from emulsion particles to H D L when incubated with rat or rabbit
plasma, even in the presence of lipid transfer enzymes. Almost all of the cholesteryl
oleate and retinyl palmitate labels remained in fraction 1 and 2, corresponding to
V L D L , emulsion and emulsion remnants. Only 4 % of retinyl palmitate transferred in
human plasma incubated with H D L , and this was found in the L D L subfraction. At
the same time cholesteryl oleate label transferred to the H D L fraction with rabbit and
human plasma, but not with rat plasma.
The consensus is that plasma removal of retinyl palmitate, at least in it major
kinetic component, reflects plasma removal of chylomicron remnants and only 5-10%
is transferred to other lipoprotein classes. The findings support the concept that in the
initial stages of postprandial lipemia, or in the initial period after injection of
chylomicron, retinyl ester-rich plasma, retinyl esters are predominantly associated
with postprandial lipoproteins. Retinyl esters do not appear to be resecreted by the
liver (Thompson et al, 1983), (Lenich and Ross, 1987) and their exchange among
lipoproteins is minimal (Hazzard and Bierman, 1976), (Wilson et al, 1983), (Wilson
and Chan, 1983), (Berr and Kern Jnr, 1984), (Brenninkmeijer et al, 1987), (Cortner et
al, 1987). As a result, it is generally accepted that the measurement of retinyl
palmitate concentration provides a suitable method for monitoring chylomicron and
chylomicron remnant clearance (Berr et al, 1983), (Foger and Patsch, 1993),
(Rubinsztein et al, 1990). However, while Bitzen and colleagues (Bitzen et al, 1994)
found retinyl palmitate clearance to be relatively constant within individuals, they
warn that there may be considerable variation in the number and type of particles
synthesised between individuals.
Another finding is that the postprandial retinyl ester response is delayed
compared with that of apo B-48, suggesting that retinyl palmitate is questionable as a
marker of chylomicrons and their remnants. Karpe et al. (Karpe et al, 1995) studied
the metabolism of chylomicrons and their remnants in the postprandial state in
normolipidemic healthy men by measuring apoB-48 and retinyl palmitate in
lipoprotein fractions after a vitamin A oral fat load mixed meal. Compared with the
peak plasma concentration of apoB-48, the peak plasma concentration of retinyl
palmitate was delayed. Approximately 2000 and 4000 retinyl palmitate molecules
were carried in each chylomicron particle in the 3 and 6 hour samples, respectively, in

102
contrast to the remnant fraction in which 100 to 600 retinyl palmitate molecules were
found for each lipoprotein particle. The author suggested that the limited retinyl
palmitate exchange between lipoprotein particles indicates that the smaller intestinal
lipoproteins do not originate primarily from larger Sf >400 chylomicron particles but
instead are secreted directly into the Sf 20 to 400 fraction and subsequently converted
to smaller chylomicron remnants.
The delay in retinyl ester response relative to apo B-48 has been highlighted
previously (Krasinski et al, 1990b), (Cohn et al, 1989), and has raised the possibility
that retinyl ester m a y transfer to other lipoprotein classes. Another explanation is that
exogenous retinyl ester may not be incorporated within the particle at the same rate as
the dietary triacylglycerol and the endogenous apo B-48 (Lovegrove et al, 1999).
This view is shared with many researchers, but the fat load method remains a popular
technique in clinical studies. Retinyl ester measurements can provide additional data
regarding the incorporation of other constituents of the chylomicron particle, and clear
information on the patterns of lipaemic response to meal ingestion (Lovegrove et al,
1999).
Zilversmit (Zilversmit, 1979) describes the use of a dietary radioactive retinol
label as being incorporated in the chylomicron fraction primarily in the esterified form
remains with the chylomicrons through their degradation to remnants and subsequent
uptake by the liver. H e found that triglyceride disappearance from the bloodstream
differs little in cholesterol-fed and control animals, whereas the disappearance of the
retinyl ester and cholesteryl ester portions of the chylomicron were greatly retarded.
In rabbits fed cholesterol for 4 days, at least two-thirds of the V L D L cholesterol
appeared to be a result of chylomicron remnants (Ross and Zilversmit, 1977).
Zilversmit and others ((Rodriguez et al, 1976), (Kushwaha and Hazzard, 1978)) have
concluded that a partial blockade exists in the removal of cholesteryl ester-laden
remnants from the circulation, and that the predominant cholesterol-containing
lipoprotein in plasma is composed of chylomicron remnants.
The utilisation of retinyl esters as tracees for chylomicron remnant metabolism
is not new. Since the original investigations of Gage and Fish (Gage and Fish, 1924.)
into the dynamics of large chylomicron particles during postprandial lipemia,
measurements of triacylglycerol-rich lipoproteins after ingestion of fat-rich meals
have been utilised to provide information about the metabolism of these intestinal
lipoprotein particles in vivo (Havel, 1997). Hazzard and Bierman used retinyl esters as

103
a marker for circulating intestinally derived lipoproteins in 1976 (Hazzard and
Bierman, 1976), and showed that the clearance of chylomicron remnants in subjects
with type III hyperlipidemia was impaired. This method has been widely used for
estimating postprandial responses in animal (Kita et al, 1982-b), (Ross and
Zilversmit, 1977) and human (Berr and Kern Jnr, 1984), (Berr et al, 1986), (Berr et
al, 1985), (Berr et al, 1983), (Berr, 1992), (Krasinski et al, 1990a), (Borel et al,
1998), (Wilson etal, 1983), (Wilson etal, 1985a), (Weintraub etal, 1987a) studies.
The vitamin A fat-load method has also been utilised successfully for the
purpose of identifying patients with abnormalities in postprandial lipid clearance,
including chronic renal failure (Wilson et al, 1985a), hypertriglyceridemia (Wilson et
al, 1985b), hyperlipidemia (Weintraub et al, 1987b), (Sprecher et al, 1991),
hyperbetalipoproteinemia (Genest et al, 1986), type HI hyperlipoproteinemia
(Cortner et al, 1987), (Brummer, 1998), homozygous F H ( M a m o et al, 1998a),
hypothyroidism (Zerbinatti et al, 1991) and C A D (Groot et al, 1991). It has also
been used to monitor postprandial responses to dietary regimens such as fatty acid
(Fielding et al, 1996), fish oil intake (Brown and Roberts, 1991) and cholesterol
feeding (Ross and Zilversmit, 1977), in addition to contraceptive steroid use (Berr et
al, 1986). Boquist et al. (Boquist et al, 1999) investigated the relationship between
alimentary lipemia and common carotid artery intima media thickness, in an effort to
diagnose chylomicron remnant metabolism in individuals at risk of developing
atherosclerosis. In addition, the vitamin A fat tolerance test has been used to
demonstrate that thyroxine replacement therapy enhances the clearance of
chylomicron remnants in normolipidemic patients with hypothyroidism (Weintraub et
al, 1999).
Despite some uncertainties, the vitamin A technique remains the method of
choice for most laboratories that wish to assess post-prandial lipoprotein kinetics
(Smith et al, 1999). However, Redgrave (Redgrave, 1999) cautions that results from
the fat load challenges are reliable in the detection of severe derangement's of
remnant clearance, but possibly unreliable in detecting minor abnormalities of
remnant physiology. It is also important to consider during interpretation of results,
that plasma data is affected by clearance as well as kinetics of absorption of vitamin A
from the intestinal lumen. Retinyl esters may transfer to other lipoproteins in the
plasma, particularly in the later stages of the fat load test, and the esters may serve as
a substrate for LPL, causing further complications in data interpretation. Furthermore,

104
it has not been established whether chylomicron metabolism is altered by transport of
a large dietary load of vitamin A
A number of considerations have to be made when delivering any fat
challenge meal, including meal fatty acid composition, size of the meal, time of day,
and what environmental factors need to be taken into consideration (Karpe, 1997b). A
key issue is whether or not the meal should contain other nutrients as well as the
lipids. In a number of previous studies, cream has been used as the only component of
the meal. Although this is a fairly simple and reproducible meal, it has clear
disadvantages. Firstly, dairy fats contain a substantial amount of short- and medium-
chain fatty acids, which cannot form chylomicrons. Secondly, the saturated fatty acid
content is high. This affects the physico-chemical behaviour of the chylomicrons
when the temperature of plasma or isolated fractions of triglyceride-rich lipoproteins
is lowered. Oils are more suitable, but they all have to be prepared before ingestion, as
emulsification is important for both palatability and absorption. With regards to meal
size, the total fat mass must constitute a provocation of the metabolic system; on the
other hand, if the meal is too large, interfering with effects on absorption, drastically
delay gastric emptying and problems with palatability will appear. A n appropriate
meal size is probably between 20-120 g fat.
1.7.2 Apolipoprotein B Assays
Another method for monitoring chylomicron metabolism is the measurement of the
unique structural protein of intestinally derived lipoproteins, apo B-48 (Kane et al,
1980), (Phillips et al, 1997). Apo B-48, a protein found exclusively with
chylomicrons and their remnants and is undoubtedly an appropriate marker of
chylomicron kinetics because it is endogenous, is indicative of particle number and is
not transferred to other plasma lipoproteins. Smith et al. (Smith et al, 1999) have
even suggested that fasting plasma concentration of apo B-48 appeared to be a good
surrogate marker for the degree of post-prandial lipidemia and may circumvent the
need for oral fat challenges. This is based on the fact that in the fasting state apo B-48
concentration is a marker of chylomicron remnants. Only one apo B-48 molecule is
associated with each intestinally derived postprandial particle (Phillips et al, 1997),
(Martins et al, 1994), therefore the concentration of apo B-48 is an excellent marker
for particle number. There has been some debate as to whether the number or size of

105
particles increases during a fatty meal, as this could have bearing on the interpretation
of results. Martins et al. (Martins et al, 1994) suggested from animal studies that the
number of particles remains the same. However, human studies indicate that the
number of particles is increased following a fatty meal ( M a m o et al, 1998a).
The mass of apo B-48 in fasting and postprandial plasma is small, which limits
quantification, and therefore apo B-48 kinetics is not commonly used to study
chylomicron remnant metabolism (Smith et al, 1999). Although apo B-48 is an
exclusive marker for chylomicron remnants, the quantitation of apo B-48 at present
remains difficult and non-standardised. The advent of methods to quantify apo B-48
and apo B-100 separately in triglyceride-rich lipoproteins (Bergeron et al, 1996),
(Karpe et al, 1996) evidently can circumvent this problem, because the apo B
components remain exclusively with the chylomicron and V L D L particles until they
are removed from the blood (Havel, 1997). A number of researchers (Schneeman et
al, 1993), (Bergeron and Havel, 1996) are now applying quantitative techniques of
apo B-48 and apo B-100 analysis to measure the contribution of chylomicrons and
V L D L particles to postprandial lipemia.
Smith et al. (Smith et al, 1997) have developed a method for quantification of
fasting serum apo B-48 concentration which requires density gradient
ultracentrifugation of the total lipoprotein fraction and the use of a sensitive but time-
consuming method, involving western blotting and enhanced chemiluminescence
detection of the apo B-48 protein. The mass of apo B-48 is calculated using a purified
apo B-48 standard whose concentration is determined by colorimetric protein assay.
This method is relatively new, and hence there is minimal data available on the
normal range of fasting plasma apo B-48 concentrations or its use in a clinical setting
(Smith et al, 1999). In addition, many researchers utilise a fat load meal to measure
chylomicron metabolism, and measure plasma concentration of apo B-48 and retinyl
esters concurrently. A common finding is that the retinyl ester response is delayed
compared with that of apo B-48, suggesting that both or either of these markers are
questionable as a means of quantifying postprandial lipoproteins.
The contribution of the intestinally derived lipoproteins to total lipemia after
altering diet fatty acid compositions was assessed by Lovegrove et al. (Lovegrove et
al, 1999), using apo B-48 and retinyl ester concentrations. The authors found that
retinyl ester and apo B-48 measurements provided broadly similar information with
respect to lack of effects of dietary or meal fatty acid composition and the presence of

106
single or multiple peak responses. However, the apo B-48 and retinyl ester
measurements differed with respect to the timing of their peak response times, with a
delayed retinyl ester peak, relative to apo B-48, of approximately 2-3 hr for one of the
diets. Lovegrove et al (Lovegrove et al, 1999) concluded that there are limitations of
using retinyl ester in a fat load as a specific chylomicron marker, whereas apo B-48
quantitation was found to be a more appropriate method for chylomicron and
chylomicron remnant quantitation. Similarly, Karpe et al. (Karpe et al, 1995) studied
the metabolism of chylomicrons and their remnants in the postprandial state in
normolipidemic healthy m e n by measuring apoB-48 and retinyl palmitate after a
vitamin A oral fat load. Fasting plasma apoB-48 concentrations were low in most
subjects and increased in response to the test meal in Sf > 20 lipoprotein fractions.
However, compared with the peak plasma concentration of apoB-48, the peak plasma
concentration of retinyl palmitate was delayed. The delayed appearance of the retinyl
ester peak response relative to apo B-48 and even triacylglycerol have been
highlighted previously (Cohn et al, 1989), (Krasinski et al, 1990b). This observation
raises the possibility that retinyl ester may transfer to other lipoprotein classes or that
the two methods are marking different populations of chylomicrons and their
remnants.
As a result of their studies in humans, Krasinski et al. (Krasinski et al, 1990b)
suggest that following a fat-rich meal, the earliest intestinal chylomicrons entering
plasma are rich in triglyceride. As absorption proceeds and lipid is depleted from the
intestine, chylomicrons entering plasma decrease in number (as indicated by the
decrease in apoB-48) and become less rich in triglyceride, but are increasingly rich in
retinyl ester. The net effect is that in postprandial triglyceride and apoB-48 peak
earlier than retinyl ester. A n alternate hypothesis is that a late retinyl ester peak could
result from a late input of retinyl ester in V L D L from the liver. Alternately, the delay
in clearance and peak times may signify that apo B-48 and retinyl ester are marking
different populations of chylomicrons, this indicates that the two markers do not
mirror each other. Another explanation is that exogenous retinyl ester may not be
incorporated within the particle at the same rate as the dietary triacylglycerol and the
endogenous apo B-48 (Lovegrove et al, 1999).
The measurement of apo B-48 is generally agreed to be the most appropriate
measurement against which other methods should be compared (Lovegrove et al,
1999). Apo B-48 is a good indicator of fasting levels of postprandial lipoproteins, and

107
has shown to be an indicator of visceral obesity and C A D as diagnosed by
angiography (Smith et al, 1999). However, the measurement of apo B-48 does not
distinguish defects in lipolysis from defects in remnant clearance. Bergeron and Havel
(Bergeron and Havel, 1995) found a slightly greater maximal response of apo B-48 to
a meal rich in polyunsaturated fat as compared with a meal rich in saturated fat, but
return to baseline values was similar. The authors also found prolonged responses of
both apo B-48 and apo B-100 to a challenge meal in individuals with apo E4/apo E3
phenotype as compared with those of apo E3/apo E3 phenotype (Bergeron and Havel,
1996). It is generally considered that the rate of return to post-absorptive values
reflects clearance mechanisms rather than that the rate of fat absorption (Havel, 1997).
Hence, their results were interpreted to suggest that clearance of remnants of intestinal
chylomicrons and hepatic V L D L is impaired in the presence of an allele specifying
apoE4.
1.7.3 LDL-Receptor Function
Analysis of LDL-receptor function or of the LDL-receptor gene is generally reserved
for the diagnosis of familial hypercholesterolemia. Specific testing of LDL-receptor
function on cultured cutaneous fibroblasts or circulating peripheral lymphocytes can
detect F H heterozygotes and homozygotes (Cuthbert et al, 1986). Similar testing of
L D L receptor function on cells cultured from amniotic fluid obtained by
amniocentesis has led to the diagnosis of homozygous F H in utero (Brown et al,
1978). It is also possible to detect the abnormal genes for the L D L receptor by
analysis of D N A obtained from the leukocytes in a small amount of venous blood
(Sudhof et al, 1985). This technology is likely to play a major role in the future
diagnosis of patients with various L D L receptor defects (Humphries, 1986).
More than 150 mutations including insertions, deletions, nonsense and
missense mutations, have been described in the LDL-receptor gene. These affect the
synthesis, post-transcriptional processing, ligand-binding activity or internalisation of
the LDL-receptor (Lombardi et al, 1995). Different mutations in the LDL-receptor
gene produce different phenotypic effects upon the receptor protein. Approximately
two-thirds of the mutations are single base changes or small structural rearrangements
and about one third of the mutations consist of major structural rearrangements
detectable by Southern blotting. In most individuals with these mutations, the amount

108
of m R N A is markedly reduced, although in some a defective m R N A is present
(Hobbs et al, 1988). Thus, in most individuals with these mutations, the LDL-
receptor is absent. These genes are coordinately regulated by the sterol status of the
cell in parallel with the LDL-receptor (Rudling, 1992). The expression of the LDL-
receptor is primarily regulated at the transcriptional level. Therefore, to define m R N A
translation, gene transcription, and LDL-receptor binding activity, the level of LDL-
receptor protein and m R N A are measured in human cells. The amount of LDL-
receptor protein is quantified using the enhanced chemiluminescence method and the
level of m R N A for the LDL-receptor is quantified by the reverse transcription
polymerase chain reaction.
However, the L D L receptor function test utilises in vitro analysis with
lymphocytes, and is an expensive method of obtaining this information. In addition,
the rate of progression of atherosclerosis in any homozygous F H patient cannot be
predicted from LDL-receptor status alone. The advantage of such an approach is that
it may provide more specific information for aggressive and effective treatment. For
the definitive diagnosis of genetic dyslipidemias, family-screening studies and
additional biochemical assays are essential. Unfortunately, such data often are not
available to many physicians who are treating patients with lipid disorders.
1.7.4 Chylomicron-Like lipid Emulsions
Chylomicron metabolism may also be traced by injecting radiolabeled nascent
chylomicrons (Redgrave et al, 1976) directly into circulation. A simpler approach to
the measurement of chylomicron remnant (particle) clearance is the injection of
chylomicron-like emulsions with labelled cholesteryl ester (Maranhao et al, 1986),
(Oliveira et al, 1988), (Redgrave et al, 1993). Chylomicron-like emulsions are
metabolised similarly to nascent lymph chylomicrons (Redgrave and Maranhao,
1985) and their composition can be manipulated to isolate different aspects of
chylomicron metabolism. These techniques permit the clearance phase of
chylomicron metabolism to be measured, and can give an estimate of chylomicron
remnant residence time (Redgrave et al, 1993). Chylomicron-like emulsions are
primarily used to evaluate the kinetics of chylomicron metabolism in animal studies
(Redgrave et al, 1991), (Bowler et al, 1991), (Mamo et al, 1991), ( Mamo et al,
1994), (Mortimer et al, 1994a), (Mortimer et al, 1994b), (Mortimer et al, 1995a).

109
Radiolabeled chylomicron-like emulsions have been used to investigate defects in
chylomicron removal in human subjects (Redgrave et al, 1993), but the use of
radioactive tracers renders them unsuitable for use in humans. Despite this
disadvantage, emulsions have been utilised to study the postprandial effects of C H D
in humans (Maranhao et al, 1996), (Martins et al, 1995), (Hungria et al, 1999).
Chylomicron-like emulsions have been used extensively to study chylomicron
metabolism in animal models of insulin deficiency (Redgrave and Callow, 1990),
hypothyroidism (Redgrave et al, 1991), hyperphagia (Martins et al, 1994), familial
hypercholesterolemia (Mamo et al, 1991), (Mamo et al, 1994), (Bowler et al, 1991)
and gene-manipulated animal models (Mortimer et al, 1997), (Mortimer et al,
1994b).
1.7.5 Breath Test
One other means of assessing chylomicron and chylomicron remnant metabolism is
via a breath test. Redgrave et al. (Redgrave et al, 1995) have utilised chylomicron-
like and chylomicron remnant-like emulsions labelled with [l-14C]oleate to monitor
remnant catabolism in intact animals. The appearance of label in the breath in diabetic
rats, and apo E-deficient and LDL-receptor-deficient mice was delayed compared
with their respective controls. In homozygous W H H L rabbits, the appearance of
14C02 was much slower than in normal control rabbits, while in heterozygous W H H L
rabbits an intermediate level of appearance was found. Martins and Redgrave
(Martins and Redgrave, 1998) have also utilised chylomicron remnant-like emulsions
labelled with [l-13C]oleate to assess the metabolism of triglyceride-rich lipoprotein
remnants in mice, in accordance with the breath test.
The authors concluded that the breath test provides an integrated and
quantitative information assessment of capacity for clearance and subsequent
metabolism of the remnants of triglyceride-rich lipoproteins in intact experimental
animals. The breath test is simple to perform and non-invasive, and can be used to
define the roles of certain disease states for example insulin deficiency and LDL-
receptor deficiency, on the roles of lipid constituents on remnant clearance (Martins et
al, 2000b). The breath test also lends itself to repeated measurements to assess the
effects of dietary and/or pharmacological manipulations. To date, studies indicate that
the breath test reliably measures the metabolism of chylomicron remnants and that

110
cholesteryl ester fatty acid is metabolised by mitochondrial pathways (Martins et al.,
2000a), (Redgrave et al, 1995), (Martins et al, 2000b). However, the process of fatty
acid metabolism and oxidation to C O 2 by the liver is complex, therefore enrichment
of [1-13C] in the breath is subject to variation caused by [1-13C] in foods, depending
on their origin. The validity of the breath test has been established in animal models
(Martins and Redgrave, 1998). However, clinical trials in humans are still needed to
establish the predictive value of the breath test in the diagnosis of increased risk of
cardiovascular disease and to determine its value in measuring the effects of
therapeutic interventions (Redgrave et al, 2001).
1.7.6 Other Methods of Assessing Cardiovascular Disease Risk
Another common approach to monitoring postprandial lipemia is to use triglyceride as
a marker for chylomicron kinetics after ingestion of a fat load meal. However, this
provides little information as to the clearance and uptake of the remnant particle, as
triglycerides are cleared as a consequence of lipolysis and do not remain with the
particle after this process (Krasinski et al, 1990b). For remnants, it is insufficient to
measure plasma triglycerides as an indicator of metabolism because slow clearance is
not always sufficient to induce chronic hypertriglyceridemia and lipolysis is not
usually the rate-limiting factor.
A n immunoseparation assay has recently been commercially developed for the
purpose of measuring both chylomicron and V L D L remnant cholesterol (remnant-like
particle cholesterol or RLP-C) in human serum. The assay removes lipoproteins using
monoclonal anti-apo B-100 (LDL and V L D L ) and anti-apo A-l (chylomicrons and
H D L ) immunoaffinity mixed gels. Following separation of RLP, the assay
enzymatically measures the cholesterol in the unbound fraction. The measurement of
RLP-C provides a simple and cost-effective marker for remnant lipoproteins, and may
be useful in large-scale clinical or epidemiological studies (Nakajima et al, 1993),
(Nakajima et al, 1996). Devaraj and colleagues (Devaraj et al, 1998) have used this
assay to detect elevated fasting RLP-C in normolipidemic patients with C A D .
However, unless the relative content of apo B-48 and apo B-100 in the RLP-C
fraction are evaluated, the contribution of hepatic versus intestinal particles cannot be
determined (Sakata et al, 1998). In addition, the RLP-C test lacks specificity for

Ill
distinguishing defects in lipolysis from defects in remnant clearance (Redgrave et al,
2001).
Deficiencies in LDL-receptors may result in accumulation of chylomicron
remnants in plasma and subsequent dyslipidemia. Therefore measurement of LDL-
receptor status is one method of identifying possible delays in receptor-mediated
clearance of chylomicron remnants and to monitor the effectiveness of therapeutic
intervention. One approach to studying the role of the LDL-receptor in the plasma
clearance of chylomicron remnants is to measure postprandial chylomicron
metabolism in subjects with homozygous FH, especially if the LDL-receptor defects
are defined with respect to the recognition and metabolism of apo E-containing
ligands (Mahley et al, 1989). Since chylomicron remnant and (3-VLDL clearance are
both known to be mediated by apo E (Mahley and Innerarity, 1983), (3-VLDL
degradation rates in cultured skin fibroblasts are used as an index of the capacity of
hepatic LDL-receptors in F H subjects to mediate chylomicron remnant clearance.
A microemulsion with the size and structure of L D L lipid portion has been
utilised by Maranhao et al (Maranhao et al, 1997), with the assumption that the
emulsion was taken up by LDL-receptors when injected into the bloodstream. In vitro
experiments in mononuclear cells demonstrated that LDL-emulsion strongly
competed with native L D L for cell uptake, via a saturable mechanism. W h e n injected
into hypercholesterolemic human subjects, the plasma disappearance curve was
markedly slower than that from normolipidemic subjects. Additionally, the plasma
clearance rate of the LDL-microemulsion in hypertriglyceridemic patients was not
significantly different from controls. Several experimental observations in patients
with myeloid leukemia, whose LDL-receptor function is upregulated, have conveyed
the concept that LDL-emulsion is picked up by the LDL-receptor (Maranhao et al,
1992), (Maranhao et al, 1993), (Maranhao et al, 1994). They also demonstrated that
the plasma disappearance curve obtained from hypercholesterolemic patients was
markedly lower than that from control normolipidemic subjects, though the degree of
hyperlipidemia did not necessarily determine the clearance rate of the LDL. The
authors concluded that the LDL-emulsion could be a useful tool to study lipoprotein
metabolism, however the emulsion was radiolabeled with [14C]oleate, which is not
acceptable for use in humans. In addition, it may not give a true indication of LDL-

112
receptor activity, as chylomicron remnants have a higher affinity for the LDL-receptor
than does L D L .
LDL-receptor detection using colloidal gold-LDL conjugates also allows for
the detection of the LDL-receptor on nitrocellulose paper (Roach et al, 1987), (Roach
et al, 1993b), (Roach et al, 1993a). The method is inexpensive, safe and sensitive,
and the colloidal gold-LDL conjugates can be used to quantify tissue and cell L D L -
receptors, when combined with silver staining and scanning densitometry. However,
the method utilises solubilised liver membrane proteins and is based on L D L versus
chylomicron remnant uptake via the LDL-receptor. A more recent method described
by Pal et al. (Pal et al, 2000) involves the binding of colloidal gold-labelled
chylomicron remnants to detect LDL-receptor binding activity in fibroblasts. Binding
of gold-labelled remnants was greater than that of gold-labelled L D L , supporting a
greater affinity of the LDL-receptor for lipoproteins containing apo E, and suggesting
that changes in LDL-receptor expression might be more readily identified using gold-
labelled remnants. However, while the method avoids the use of liver biopsy, it
assumes that human skin fibroblasts are suitable surrogate markers for the hepatic
receptor and does not give a clear indication of remnant clearance via the L D L -
receptor in vivo.
Despite the large number of techniques currently available to monitor
chylomicron clearance in vivo and in vitro, the metabolism of chylomicron remnants
is not considered in routine clinical screening or studies for subjects with elevated
cholesterol or who possess coronary risk factors. This dilemma lies partly in the
difficulty in separating intestinal from non-intestinal lipoproteins, complexity of the
procedures, standardisation of any procedure, and data reduction into a manageable
number of concise variables and context for interpretation of these variables.
Therefore, there is no simple, reproducible technique available at present to detect
elevated plasma levels of chylomicron remnants or to estimate LDL-receptor activity.
1.8 The Aims of the Project
The primary aim of this thesis was to develop a diagnostic assay to assess receptor-
mediated uptake of chylomicron remnants by monitoring particle clearance and
quantitating LDL-receptor activity in vivo, and was investigated utilising the
following methods:

113
• T o demonstrate that normal and modified emulsions represented the uptake of
chylomicron remnants by total and low affinity uptake mechanisms, respectively, and
to investigate the validity of these two emulsions as a model for monitoring receptor-
mediated uptake, the clearance of the two emulsions was studied in animal models.
W H H L rabbits were utilised to investigate the possibility that chylomicron remnants
are catabolised via a receptor distinct from the LDL-receptor. The clearance kinetics
of separate and simultaneous injection protocol were also compared.
• T o synthesise a remnant-like emulsion that would physiologically mimic
chylomicron remnants in vivo, the lipid composition of remnants derived from nascent
chylomicrons was investigated, allowing accurate synthesis of replica particles.
• Fluorescent labelling techniques were used in knockout animal models to
confirm the uptake of normal chylomicron-like emulsions types by the liver, and the
delayed uptake of modified chylomicron-like emulsions at a cellular level. The
simultaneous injection of normal and modified emulsions was also investigated, to
eliminate the possibility that this would interfere with the kinetics of particle uptake.
• To establish that retinyl esters had no deleterious effect on the clearance
kinetics of chylomicron-like emulsion triglyceride and cholesteryl oleate clearance,
representing lipolysis and particle uptake, respectively, the clearance of emulsions
with and without retinyl esters incorporated were compared.
• T o ensure that retinyl esters would mimic particle uptake, the clearance of
emulsion retinyl esters and radiolabeled cholesteryl oleate were monitored and
compared in animal models. To assess the suitability of utilising retinyl esters for
quantitation of receptor-mediated uptake, high affinity was calculated as the
difference in clearance of the retinyl esters in the two emulsion types.
There is an increasing body of evidence to suggest that chylomicrons and their
remnants m a y contribute to lesion formation, and the atherogenic potential of
postprandial lipoproteins is now becoming more widely recognised. Karpe suggests
that the reasons for the relative lack of clinical evidence demonstrating an
involvement of postprandial lipid and lipoprotein metabolism in the development of
atherosclerosis is a consequence of biological, statistical and methodological issues
(Karpe, 1999). The perturbations of lipid metabolism in the postprandial state are
complex, with changes in both composition and concentration of potentially
atherogenic lipoproteins. However, with the development of new and more sensitive

114
techniques to quantify chylomicrons and their remnants, a slightly revised concept for
the clinical implications of the metabolism of postprandial lipoproteins is emerging.
Chylomicron remnants are removed from the circulation by LDL-receptors,
using apo E as the ligand. Remnants share the same receptor as L D L under normal
circumstances, and in certain disease states, the clearance of both of these particles is
impaired. The metabolism of L D L is reasonably well understood in LDL-receptor
deficient individuals, however little research has been dedicated to remnant clearance
patterns and metabolism. Consequently, it is still not clear what role the LDL-receptor
performs in the clearance of chylomicron remnants. Information of this nature is
important in furthering our understanding of the metabolism of remnants in L D L -
receptor deficient individuals. Defects in LDL-receptors or in apo E may lead to
chylomicron remnant accumulation in the plasma. A delay in chylomicron remnant
clearance results in increased arterial exposure and uptake followed by extracellular
accumulation of chylomicron remnants which initiates an inflammatory response as
evidenced by phagocytic degradation by macrophages. Therefore, cholesterol in early
atherosclerotic lesions could represent lipid primarily derived from chylomicron
remnants and not L D L .
Plasma cholesterol measured in the fasting state is a poor predictor of C H D
risk, as one half of heart attacks occur in individuals with plasma cholesterol in the
normal range (Martin et al, 1986). Therefore, many individuals with abnormal
postprandial lipoprotein metabolism may remain undetected in routine fasting clinical
testing. Weintraub et al. (Weintraub et al, 1996) and others have shown that
normolipidemic subjects can still display remnant dyslipidemia. There is also
evidence to suggest that, although chylomicron remnants have greater affinity for the
LDL-receptor than L D L , they in fact require greater expression of receptors for
proper removal (Yu et al, 1999), (Bowler et al, 1991), (Choi and Cooper, 1993).
Familial hypercholesterolemia is a common genetic abnormality (1/200), and these
individuals are hypercholesterolemic, with mild hypertriglyceridemia as evident in
Type lib Familial combined hypercholesterolemia. Observations such as these might
represent individuals with sufficient receptor expression for proper removal of L D L ,
but not for chylomicron remnants (Smith et al, 1999).
Despite the substantial clinical evidence suggestive of a causal role of post
prandial dyslipidemia and C A D , studies of chylomicron remnant metabolism in vivo
have been hampered by lengthy procedures that discourage subject participation

115
(Smith et al, 1999). The availability of quantitative methods to measure lipids, apo B-
48 and retinyl palmitate in fat loading has provided a very useful tool in determining
the intestinal contribution to postprandial lipemia, and in identifying individuals with
greater postprandial responses. However, these methods are complex and insensitive
to small defects in chylomicron remnant clearance (Martins and Redgrave, 1998). In
addition, apo B-48 is present at very low concentrations in plasma, even in the
postprandial state, and measurement of apo B-48 does not distinguish between the
processes of lipolysis and uptake. The primary problem with the vitamin A oral fat
test is that the concentration of retinyl palmitate in plasma reflects the combination of
secretion and clearance (Smith et al, 1999).
The metabolism of chylomicron remnants may be a sensitive index of
susceptibility to atherosclerosis, but at the present time the assessment of an
individual's capacity to metabolise chylomicron remnants is unavailable because
direct measurement has not been possible and nascent chylomicrons cannot be
collected for study. Attempts to identify surrogate markers that might predict the rate
of remnant removal have been met with only limited success. This is in part due to the
fact that chylomicron remnants clear rapidly from the plasma (t Vi ~ 15 mins), thereby
making these particles difficult to monitor. This is in keeping with the concept of
exposure in terms of concentration multiplied by time, whereby the concentration of
remnants in plasma is sufficient for accurate measurement for a limited time period
only. There is also a paucity of means to quantify the contribution of receptor
mechanisms to postprandial lipemia. Specific methods that can be applied practically
and rapidly in clinical research and practice are still needed in order to investigate
factors effecting levels of chylomicron and their remnants, and their importance in
C H D risk. Currently, screening for receptor function involves complex and expensive
methods that are restricted to high-risk patients.
A safe, low intrusive procedure for quantifying net receptor activity in vivo
would be valuable in identifying individuals with defective postprandial metabolism,
and improving procedures for screening individuals at risk of developing vascular
disease by relating high-affinity uptake to remnant clearance and L D L cholesterol
concentration. In certain disease states, the clearance of both L D L and chylomicron
remnant particles is impaired and atherosclerosis is often not recognised until a
clinical event occurs. It is therefore likely that many individuals have an undiagnosed
receptor deficiency (and reduced lipoprotein clearance). Quantitation of net receptor

116
activity in vivo may be a better predictor of 'risk' than the plasma concentration of
LDL-cholesterol. In particular, a technique such as this could possibly establishing
additional criteria for selecting individuals for dietary and/or therapeutic intervention.
It may also assist in determining whether LDL-receptor activity is reduced or down
regulated and could potentially be used to screen for the existence of functional
abnormalities of apo E or to distinguish them from defects in LDL-receptor-related
function, or to disclose the effectiveness of drug treatments or dietary regimens.
Consequently it was the aim of this project to develop a reproducible and cost-
effective diagnostic assay whereby receptor-mediated uptake of chylomicron
remnants could be assessed quickly and simply. This procedure could be used to
monitor lipoprotein particle clearance for use in man, and permit quantitation of (net)
LDL-receptor activity in vivo. The project involved the development of two
lipoprotein-like emulsions which when injected simultaneously gives a measure of
receptor activity (based on their difference in clearance). This procedure has
previously been used successfully in animal models (Redgrave et al, 1987), (Mamo
et al, 1991), (Bowler et al, 1991), (Redgrave et al, 1995), but there is a need to
develop non-isotopic tracees for application in man. It was the aim of the project to
utilise vitamin A esters to trace clearance of the lipid emulsions. The procedure
described will enable the assessment of defective chylomicron remnant clearance as a
risk factor for vascular disease.
Lipid emulsions have been utilised extensively for monitoring the metabolism
of chylomicron remnant clearance and have been established as reliable models for
chylomicron metabolism in humans (Redgrave et al, 1993) and animal models
(Redgrave et al, 1992a), (Redgrave et al, 1995), (Martins and Redgrave, 1998),
(Mortimer et al, 1994b), (Mortimer et al, 1997). Emulsions have also been widely
used to monitor the effects of hypothyroidism (Redgrave et al, 1991), diabetes
(Redgrave and Callow, 1990), (Martins et al, 1994), familial hypercholesterolemia
(Mamo et al, 1991), (Mamo et al, 1994), (Bowler et al, 1991) C H D (Maranhao et
al, 1996), (Martins et al, 1995), renal transplant (De Lima et al, 1998), multiple
myeloma (Hungria et al, 1999), hyperlipidemia (Maranhao et al, 1997), and
hypertension (Mackintosh et al, 1996) on the metabolism of chylomicron remnants in
humans and animal models. Lipid emulsions have been utilised as a means of specific
diagnosis and for targeting therapeutic drugs, thus, they have are a potentially strongly
diagnostic tool for screening those at risk of developing a number of disease states.

117
The advantages of using chylomicron-like lipid emulsions is that particles are
of defined size range and composition, similar to nascent lymph chylomicrons
(Redgrave and Maranhao, 1985), (Rensen et al, 1997), the lipids are of known purity,
and it is possible to bypass problems associated with kinetics of intestinal absorption.
Emulsions rapidly incorporate the apolipoproteins from plasma, either by the release
of apolipoproteins from plasma lipoproteins or by the association of apolipoproteins
present free in plasma (Erkelens et al, 1981), (Weinberg and Scanu, 1982), (Tajima et
al, 1983). They are metabolised similarly to chylomicrons consistent with rapid LPL-
mediated hydrolysis of triacylglycerols, followed by hepatic uptake of the remnants
derived from the emulsions (Lenzo et al, 1988), (Carlson and Hallberg, 1963). In
addition, (Martins et al, 1989) found that a portion of labelled phospholipids from
emulsions injected into intact rats transferred rapidly to H D L fractions, modelling the
transfer that occurs with lymph chylomicrons (Redgrave and Small, 1979). Following
injection of radiolabeled emulsion particles into rats and human subjects, Redgrave et
al. (Redgrave et al, 1993) concluded that information about chylomicron metabolism
can be obtained via plasma clearance data following injection of suitably labelled
chylomicron-like emulsions in man and rats. Oliveira et al. (Oliveira et al, 1988) and
Redgrave and Maranhao (Redgrave and Maranhao, 1985) have also found that
nascent chylomicron remnants compete with chylomicron-like emulsion lipids.
Another advantage of the chylomicron-like emulsion technique is the ability to
inject a known lipid mass as a bolus injection. This enables the bypassing of intestinal
processing time (digestion and absorption), and would reflect plasma clearance. It will
also reduce circulation time and the likelihood of retinyl ester transfer.
T o achieve a diagnostic assay whereby the receptor-mediated uptake of
chylomicron remnants can be assessed in human subjects in vivo, it was necessary to
find a suitable substitute for radiolabeled cholesteryl oleate to mimic particle uptake.
Based on the documented utilisation of the vitamin A fat load test, retinyl esters were
chosen as the appropriate marker to be incorporated into chylomicron-like emulsion
particles. Retinyl ester concentrations in plasma following intravenous introduction of
the two emulsion types (using a different retinyl fatty acid for each) could be used to
trace particle uptake. Retinyl esters were chosen for the study because, similar to
cholesteryl esters, they are hydrophobic compounds and are incorporated into the core
of the chylomicron particle upon assembly. Retinyl esters are naturally occurring
compounds and bolus injections of retinyl palmitate-labelled plasma have been used

118
to monitor chylomicron remnant metabolism in vivo, however their incorporation into
chylomicron-like emulsions has not been studied previously as an alternative to their
incorporation following a vitamin A fat load meal.
In order to characterise remnant clearance in normal humans, Berr and Kern
(Berr and Kern Jnr, 1984) and Berr (Berr, 1992) isolated chylomicrons from human
plasma after feeding retinol in a fatty meal. After a delay to allow the rest of the fed
retinol to disappear from the patient's plasma, particles were pulse-injected directly
into plasma of subjects over 3-5 min. This avoided the problem of kinetics of
intestinal absorption, however, there was the additional difficulty of collecting
postprandial plasma from subjects before re-injection (Berr and Kern Jnr, 1984), (Berr
et al, 1985), (Berr et al, 1986). The first rapid component (t ¥218.8 min) was shown
to represent clearance of chylomicron remnants. The authors proposed a one-
compartment model for chylomicron remnant removal and further suggested that the
rate of chylomicron remnant formation was not the limiting factor in chylomicron
clearance, but that remnant removal was, and that catabolism by the liver appears to
be saturable by ordinary lipid intake in healthy humans.
Karpe et al. (Karpe et al, 1995) have also injected retinyl palmitate-labelled
postprandial plasma into fasting subjects, with subsequent tracing of retinyl palmitate
in fractions of triglyceride-rich lipoproteins. Based on their findings, the authors
suggested that limited retinyl palmitate exchange between lipoprotein particles
indicates that the smaller intestinal lipoproteins do not originate primarily from larger
Sf >400 chylomicron particles but instead are secreted directly into the Sf 20 to 400
fraction and subsequently converted to smaller chylomicron remnants. Others have
established that retinyl esters do no transfer to other lipoproteins (Martins et al, 1991)
and several studies have established retinyl esters as reliable markers for chylomicron
metabolism in normal and hyperlipidemic subjects (Weintraub et al, 1987b), (Wilson
et al, 1983). Hence, the use of retinyl esters remains a favoured method for
monitoring post-prandial response in vivo. Based on previous chylomicron-like
emulsion studies in humans (Maranhao et al, 1996), (Martins et al, 1995), w e
envisaged the proposed technique proving to be a more rapid process compared with
the fat load test, with reduced circulation time for retinyl ester transfer.

119
Chapter 2: General Methods and Materials
2.1 Animals
New Zealand White (NZW) and semi-lop (NZW/ - cross) rabbits, weighing between
1.8 to 3.5 kg, were obtained from the Animal Resources Centre, Murdoch, Western
Australia. A colony of Watanabe-heritable-hyperlipidemic ( W H H L ) rabbits were
maintained at the Biological Sciences Animal Unit (BSAU), at the University of
Western Australia. All rabbits were housed individually in steel cages at the B S A U .
The air temperature in the room was maintained at 25°C, and the rabbits had free
access to tap water and commercial rabbit chow.
Male albino Wistar rats, weighing between 250 to 400 g m were obtained from
the Animal Resources Centre, Murdoch, Western Australia, and were housed in cages
at a temperature of 25°C. They had free access to tap water and were fed on a
commercial rat pelleted diet containing approx. 5 % fat.
Unless stated, animals were not fasted before use, but were allowed free
access to food and water until experimentation commenced. The animal handling and
surgical procedures were approved by the Animal Welfare Committees of the
University of Western Australia and Royal Perth Hospital, which are both signatory to
the 'Code of Welfare for the Handling and Experimentation on Animals' document
established by the National Health and Medical Research Council of Australia.

120
2.2 Operative Procedures
2.2.1 Lymph Duct Cannulation
The surgical procedure was adapted from Bollman et al. (Bollman, 1948) and
modified (Redgrave and Martin, 1977), (Redgrave et al, 1982), (Umeda et al, 1995).
The temperature of the surgery room was kept at room temperature (approx. 20-25 °C)
at all times. Plastic cannulas (polyvinylchloride, internal diameter of 0.5 m m , outer
diameter of 0.8 m m ) used in the operation were siliconised (when necessary, as
stated) using an aqueous solution of silicone (0.01% v/v; AquaSil™, Pierce Chemical
Co., Illinois, U S A ) to prevent clotting. Rats were anaesthetised using 0.1 ml of
sodium pentobarbitone (60 mg/ml; Nembutal®, Boehringer Ingelheim Pty Ltd.) per
100 g body weight. While under full anaesthesia, the abdomen was opened by mid
line incision. The stomach and intestine were displaced to the right of the animal. A
large hypodermic needle (14G) was inserted under the inferior vena cava to pierce the
right hand side of the dorsal wall. A siliconised cannula was threaded through the
needle from the exterior of the rat. The needle was then removed leaving the cannula
in place. The cannulas were presiliconised using an aqueous silicone solution to
prevent clotting. The cannula was inserted into the superior mesenteric lymph duct,
which emerges from the duodenum, and fused with Superglue™ (Selleys Chemical
Co., Padstow, N S W , Australia) to prevent the cannula detaching.
2.2.2 Duodenal Cannulation
A hypodermic needle (14 G) was used to pierce the left hand side of the dorsal wall,
and a polyvinylchloride cannula (LD. 0.5 m m ; O.D. 0.8 m m ) inserted through the
needle. The needle was then removed, leaving the cannula in place. A triangle was
stitched in place in the stomach (Silk No.l) and a small incision made in the triangle.
The gastric cannula was fed through the stomach into the duodenum with its end
placed approximately 5 c m beyond the pylorus. The silk triangle was pulled tight and
tied with silk to secure the cannula (gastric tube) in place. Following lymph duct and
duodenal cannulation, the abdominal wall and skin was sutured with Silk N o 2.

121
2.2.3 Collection of Rat L y m p h Chylomicrons
Post-operatively, the rats were restrained in individual Bollman cages before
recovering full consciousness. Tap water was freely available. A 0.15 M NaCl
solution was infused through the gastric tube at 2 ml/h using a peristaltic pump
(Gilson, Paris, France). After an hour of saline infusion and full recovery from
anaesthesia, Intralipid™ (2% v/v; Kabi Pharmacia, Sweden.) in 4 % (w/v) glucose was
infused through the same gastric tube at 2 ml/h. The lymph fluid was initially clear
but became 'milky' within an hour of infusion of Intralipid™. The milky lymph was
collected over a period of 24 hr into conical flasks containing 0.4 M E D T A , p H 7.4
(500 pi). Contaminating cells, i.e., leukocytes were removed by pelleting after low
speed centrifugation in a microcentrifuge (BHG Hermle Z 230M) at 3000 rpm (1200
gav) for 10 min.
2.2.4 Chylomicron Separation
The technique of density gradient ultracentrifugation was used to isolate nascent
chylomicrons from lymph (Redgrave et al, 1975). The density of the lymph was
raised to 1.10 gm/ml by adding solid potassium bromide (0.14 gm/ml). A density
gradient was pre-formed by the sequential underlayering of saline solutions of
increasing densities (6 ml of 1.065, 1.040, 1.020, and 1.006 gm/ml, respectively). U p
to 14 ml of lymph at density =1.10 gm/ml was underlayered onto the pre-formed
density gradient. Chylomicrons were isolated from lymph by ultracentrifugation at 27
000 rpm (100 000 gav) for 40 min at 20°C in a Beckman Optima-L ultracentrifuge by
using a Beckman S W 2 8 rotor (Beckman Instruments Inc., Palo Alto, CA, U S A ) . At
the end of the spin, a distinct 'creamy' layer (chylomicrons) appeared at the top of the
solution, which is removed by a combination of lifting with a teflon-coated spatula
and gentle aspiration. The chylomicrons were kept under nitrogen to prevent
oxidation, and used within 2 days after isolation. The size of the particles was
determined by laser light scattering using a BI-90 particle sizer (Brookhaven
Instruments Corp., Ronkonkoma, N Y , USA).

122
2.2.5 Preparation of Chylomicron R e m n a n t s In Vivo
Isolating chylomicron remnants from postprandial plasma pose some difficulties
because of contamination from lipoproteins (particularly V L D L and chylomicrons)
and low yields (Mamo et al, 1996). In the past, chylomicron remnants have been
'made' in vitro by treating human and animal lymph chylomicrons with lipoprotein-
deficient serum (to lipolyse chylomicrons) but this method gave inconsistent sizes of
chylomicron remnants and contamination with excessive free fatty acids (Floren et
al, 1981). Our laboratory has solved these problems by devising a novel technique to
isolate 'pure' chylomicron remnants (Mamo et al, 1996). Purity of chylomicron
remnants was based on the absence of apoB-100 containing lipoproteins following
sodium dodecyl sulfate-polyacrylamide gel electrophoresis (Proctor and M a m o ,
1996). The apolipoprotein profile showed significant quantities of apo E (-52%) and
lesser quantities of apo C (-18%), apo B-48 (-6%) and apo A-l (-7%), and a constant
diameter between 40 and 50 n m compared to greater than 200 n m for nascent lymph
chylomicrons as determined by laser light scattering. In addition, there was a
reduction in the triglyceride to cholesterol ratio from approximately 20 (i.e.,
chylomicrons) to a ratio of 2 (i.e., chylomicron remnants).
Rabbits (2.5-4 kg) were anaesthetised with intramuscular injections of
ketamine (30 mg/kg) and xylazine (5 mg/kg). Following a mid-line incision, the
coeliac and mesenteric arteries were cleared of surrounding fat and connective tissue
and ligated within 0.5 c m beyond their origin from the aorta (to exclude the hepatic
artery). The cardio-oesophageal junction was ligated just above the stomach, followed
by the rectum and associated blood vessels. Finally, the portal vein was ligated to
ensure no blood flow to the liver. The abdominal wall was sutured, and then the
hepatectomised rabbits were given a large dose of chylomicrons (1.5 g m of
chylomicron-triglyceride/3 kg B W ) by injection into a femoral vein catheter over a 5
min period. A large dose of triglyceride was chosen to dilute residual V L D L . To
prevent hypoglycaemia, glucose was infused at 1.5 ml/min by way of the femoral
cannula during the procedure. The chylomicrons were left to circulate and undergo
lipolysis in the anaesthetised animal. Complete anaesthesia was maintained by
intramuscular administration of ketamine (30 mg/kg) and xylazine (5 mg/kg) at 45-90
min intervals. After 2.5-3 hr, the rabbit was exsanguinated by bleeding from the
abdominal aorta using 0.4 M E D T A (500 pi) as the anti-coagulant. During bleeding,

123
25 ml of 0.15 M NaCl solution was infused into the femoral vein to increase recovery
of plasma. Blood was centrifuged at 3500 rpm for 15 min and the supernatant
(plasma) removed with a pipette.
2.2.6 Chylomicron Remnant Separation
Chylomicron remnants were isolated from the plasma by density gradient
ultracentrifugation. The density of the plasma was adjusted to 1.10 gm/ml by the
addition of potassium bromide (0.14 g/ml). A density gradient was pre-formed by the
sequential underlaying of saline solutions of 1.065 (6 ml), 1.020 gm/ml (6 ml), 1.006
gm/ml (8 ml) and plasma (density 1.10 gm/ml; 17 ml). Chylomicron remnants were
separated from plasma by ultracentrifugation at 27,000 rpm (100,000 gav) for 16 h at
20°C in a Beckman Optima™/L8-50M, using a Beckman S W 2 8 rotor. The milky
layer of chylomicron remnants (d < 1.006 gm/ml) was collected from the top of each
tube by gentle aspiration and used within 24 hr. The remnants were purged with
argon to prevent oxidation and were kept in the dark at 20°C. Remnants prepared in
vivo had a diameter of 45 ± 5 nm. Our laboratory has previously provided
electrophoretic evidence to exclude contamination by V L D L .
2.3 Chylomicron-Like Emulsion Preparation
2.3.1 Normal Chylomicron-Like Emulsions
Lipid emulsions resembling chylomicrons in composition and size were prepared as
previously described (Redgrave and Maranhao, 1985), (Lenzo et al, 1988). Triolein
(70 mg), cholesteryl oleate (3 mg), cholesterol (2 mg) (Nu Chek Prep, Elysian, M N )
and egg phosphatidylcholine (25 mg) (Lipid Products, Surrey, U K ) , each greater than
9 9 % pure, were dispensed from stock solutions into vials. The emulsions were
labelled with either radioactive cholesteryl[3H]oleate and [14C]triolein, or with
cholesteryl[14Cloleate and [3H]triolein (Amersham, Surry Hills, N S W , Aust). Solvents
were then evaporated under a stream of nitrogen before overnight desiccation to
eliminate residual solvent traces. The dried lipid mixture was emulsified by sonication
in 150 mM-NaCl in l O m M Hepes buffer (pH 7.4) solution. The lipids were sonicated
in 8.5 ml of the buffer solution at 55-56°C (above the liquid crystalline-to-isotropic

124
liquid melting transition of cholesteryl oleate, monitored by a thermocouple in the
vessel) with the atmosphere above the mixture purged with nitrogen to prevent lipid
oxidation. Sonication was for 20 min using a 1cm probe of the Branson Cell
Disruptor. Sonication was at a continuous output of 90-110 W with a Vibra-Cell high-
intensity ultrasonic processor (Sonics and Materials, Inc, Danbury, C N ) . The density
of the crude emulsion was increased to 1.10 gm/ml by adding KBr (0.14 gm/ml).
After placing 4 ml portions at the bottom of two centrifuge tubes, 2.5 ml of NaCl
solutions of densities 1.065, 1.020 and 1.006 gm/ml were then sequentially layered
above. The tubes were centrifuged in a S W 4 1 swinging bucket rotor of a Beckman
L8-70M ultracentrifuge for 22 min at 14 000 rpm (24191 gav) at 20°C. The large
coarsely emulsified particles were removed and replaced with a 1.006 gm/ml solution.
This was followed by a second centrifugation of 20 min at 24 000 rpm (71092 gav) at
20°C. The emulsion particles that floated to the surface were analysed for size and
lipid composition and injected into animals within 24 hr of preparation. Oxidation
was prevented by the addition of a quenching antioxidant, reduced glutathione (50
pg/ml) and storage under argon.
2.3.2 Modified Chylomicron-Like Emulsions
Modified chylomicron-like emulsion particles were prepared as described in Section
2.3.1, according to the method of Redgrave and Maranhao (Redgrave and Maranhao,
1985). Free (unesterified) cholesterol ( 2 % of total lipid mass) was omitted from the
original lipid mixture to form particles with modified clearance characteristics
(Bowler et al, 1991), (Redgrave et al, 1987). The modified chylomicron-like lipid
emulsions contained a different isotopic form of cholesteryl oleate so that clearance
from plasma of the two emulsion types could be distinguished.
2.4 Thin Layer Chromatography
The lipid composition of chylomicrons, chylomicron remnants and chylomicron-like
emulsions was determined using thin layer chromatography (TLC). The lipoproteins
(0.5 ml) were extracted in glass stoppered tubes or scintillation vials with 2.5 ml of
methanol, 5 ml of chloroform and 2.5 ml of D D W as described (Folch et al, 1957).
The tubes were mixed and allowed to stand overnight for adequate separation of the

125
phases. A volume of 4.5 ml of chloroform was removed from the lower phase and
dried under nitrogen. A n aliquot of 150 pi of chloroform: methanol mixture (2:1, v/v)
was added to the tube and 100 pi of this solution were applied in a narrow band to
thin layer plates precoated with 0.2 m m layers of silica gel. The samples were
developed for one hour in a glass tank in a solvent system petroleum ether (40-
60°C)/ether/acetic acid (90:10:1), v/v). The bands of different lipids were visualised
using iodine vapour and separated in sequence. The triglyceride, cholesteryl ester and
unesterified cholesterol bands were scraped from the plate. Triglyceride was then
extracted into chloroform and assayed, and the unesterified cholesterol and
cholesteryl ester bands were hydrolysed and assayed. The concentrations of
phosphorus in the chylomicrons and remnants were determined prior to extraction,
except for the chylomicron characterisation procedure, where samples were extracted
in methanol and assayed.
2.5 Triglyceride and Cholesterol Determination
2.5.1 Cholesterol Determination
Plasma lipids were determined from blood samples drawn throughout the kinetic
studies. All plasma samples were analysed within 8 hr of collection.
Cholesterol mass in plasma samples and lipoprotein fractions were determined
via the cholesterol oxidase procedure by an enzymatic assay kit purchased from Trace
Scientific (kit catalogue number: TR13015, Australia) in accordance to the
manufacturer's instructions. The assay does not discriminate between free and
esterified cholesterol, and therefore lipids were separated by thin-layer
chromatography and assayed independently. Colour development in samples was read
in a Beckman D U 6 5 0 spectrophotometer at a wavelength of 500 nm.
2.5.2 Triglyceride Determination
Triglyceride mass in plasma samples and lipoprotein fractions were quantified
following correction for free glycerol by subtraction of a constant, by an enzymatic
assay kit purchased from W a k o Pure Chemical Company (catalogue numbers: 430-
11291, 432-11491, 436-11391, 438-11591, Osaka, Japan) in accordance with the

126
manufacturer's instructions. Colour development in samples was read in a Beckman
D U 6 5 0 spectrophotometer at a wavelength of 600 nm. Concentrations were corrected
for free glycerol. The triglyceride mass was calculated using a reference serum
purchased from Trace Scientific Pty. Ltd. (Clayton, Vic, Australia).
2.6 Phosphorus Determination
Plasma lipid and emulsion phosphorus was determined by a modification of the
method described by Bartlett (Bartlett, 1959). Lipid phosphorus was measured
directly on emulsion and lipoprotein suspensions prior to extraction. Duplicate
samples and standards (range 0.4 to 2 pg of inorganic phosphorus KH2PO4) were
aliquoted into glass tubes, 0.3 ml 10 N sulphuric acid was added and the tubes heated
for 60 min at 200°C. One drop of 3 0 % hydrogen peroxide was then added to the
tubes. If the solution did not turn colourless a further drop of hydrogen peroxide was
added. The samples were then heated for a further 60 min at 200°C, cooled, and 2.0
ml of ammonium molybdate solution (0.22% in D D W ) was added and the tubes
mixed. 50 pi A N S A solution (0.25% w/v l-amino-2-naptha-4-sulphonic acid in a
solution of 0.5% sodium sulphite and 1 5 % sodium metabisulphite) was then added to
all tubes and mixed well. The tubes were then heated for 7 min at 100°C before
cooling to room temperature. Optical density was read at 820 n m in a Shimadzu U V -
120-02 spectrophotometer.
2.7 Clearance Studies
2.7.1 Arterial and Venous Cannulation
Non-fasted rats were anaesthetised using 0.1 ml of sodium pentobarbitone (60 mg/ml;
Nembutal®, Boehringer Ingelheim Pty Ltd.) per 100 g m body weight. Plastic
cannulas were placed in the left jugular vein and carotid artery for injections and
blood sampling, respectively. The venous and arterial cannulae were inserted through
a channel made through the skin and subdermal fascia at the back of the neck. A
saline-filled Teflon cannula (0.76 m m O.D. x 0.33 m m I.D., Small Parts, Inc.) was
inserted through the left common carotid artery so that the tip was located in the aortic
arch, and a venous vinyl cannula (0.8 m m O.D. x 0.5 I.D., Dural, N.S.W. Australia)

127
was inserted near the junction of the left jugular and subclavian veins. The tip was
advanced to lie in the superior vena cava. Heparin was not used but clotting was
prevented by treatment of the tubing with Aquasil™ (Pierce, Rockford, LL.) before
use. After surgery, the animals were allowed to recover from the effects of
anaesthesia in individual restraint cages for 2-4 h before the injection study
commenced, and were allowed free access to water during this time. Hydration of the
rats and patency of the cannulas were maintained by injections of small volumes of
0.15 M NaCl solution.
2.7.2 Clearance Studies in Rats
Chylomicron-like emulsions containing < 2.5 mg of triglyceride in a volume of 0.35-
4.0 ml were injected as a bolus into the bloodstream of conscious animals. Blood
samples of 0.35 ml were taken at 0, 1.5, 3, 5, 8, 12, 15, 20, 25, 30 and 40 min after
injection through the cannula in the carotid artery, and placed in microfuge tubes
containing 4 units of heparin. Each withdrawal was replaced with an equivalent
volume of 0.15 M NaCl solution. Plasma was separated immediately from blood cells
by centrifugation (2000 gav) in a microcentrifuge ( B H G Hermle Z 230M), for 15 min
at 4°C. After separation, the plasma collected was measured for radioactivity without
extraction, by taking 150ul and adding 5 ml of Emulsifier-Safe™ (Packard) and
counting in a Beckman LS 3800 Liquid Scintillation counter (Beckman Instruments
Inc, U S A ) . After completion of the clearance study 20 m g of sodium pentobarbitone
(Nembutal®) was injected and the liver and spleen were removed and washed
thoroughly in ice-cold 0.15 M NaCl.
Plasma clearance kinetics were computed as area under the curves during the
entire 30-40 mins after injection. To calculate the percentage of injected radioactivity
remaining at each time point the values obtained from dual-label counting were
multiplied by the total plasma volume, which was calculated as ( B W °'725 x 0.174).
The percent radioactivity remaining in the plasma was plotted on a logarithmic scale
(ordinate) against time on a linear scale (abscissa). Clearance of emulsion was bi-
exponential over 1.5-40 min for rats.

128
2.7.3 Clearance Studies in Rabbits
Chylomicron-like emulsions were injected into conscious rabbits via the lateral ear
vein. The amount of triglyceride injected was < 2.5 m g in a maximum volume of 800
ul. Blood samples of 0.5 ml were subsequently taken from the opposite ear vein at 0,
3, 5, 8, 12, 15, 20, 25, 30 and 40 min after injection for chylomicron emulsion studies.
25 G needles (Terumo Corp.) were used for injection and sampling; neither cannulas
nor heparin were used in the sampling procedure. Coagulation of blood was prevented
by the addition of 20 units of heparin to samples, which were kept on ice until plasma
could be separated by centrifugation. Plasma was separated from blood cells by
centrifugation at 2000 rpm in a microcentrifuge (BHG Hermle Z 230M), for 15 min at
4°C. After separation the plasma collected was measured for radioactivity without
extraction, by adding 5 ml of toluene-based flour (Emulsifier-Safe™, Packard) to 400
ul plasma and counting it in a Beckman LS 3800 Liquid Scintillation counter
(Beckman Instruments Inc, USA).
To determine the percentage of injected activity remaining at each time point,
the radioactivity values were multiplied by the total plasma volume (assumed to be
3.3% of total B W ) (Hussain et al, 1989). The percent radioactivity remaining in the
plasma was plotted on a logarithmic scale (ordinate) against time on a linear scale
(abscissa). Clearance of emulsion was bi-exponential over 3-40 min for rabbits.
2.7.4 Calculation of Emulsion Clearance and High Affinity
(Receptor) Uptake In Vivo
Plasma clearance was expressed as the incremental change in plasma concentration
following injection of a chylomicron-like emulsion, that is, the area under the curve
(amount of emulsion remaining in plasma). Emulsion plasma clearance kinetics were
calculated with the assistance of computer aided software (Prism, Graphpad software,
CA, USA ) . The area of a peak was determined by the trapezoid rule. The area under
the curve was then subtracted from the total value (100% of injected dose) and the
area above the curve (AAC) values were used in all studies as a measure of plasma
clearance of chylomicron remnants.
The A A C values for radiolabeled chylomicron-like emulsion triglyceride and
cholesteryl oleate were calculated for normal and modified chylomicron-like

129
emulsions, to provide a measure of the amount of radiolabeled lipid cleared from
plasma, in rat and rabbit studies. Chylomicron remnants contain some residual triolein
therefore triolein removal is the sum of two processes, hydrolysis by L P L and
removal with remnant particles (Mamo et al, 1994). Cholesteryl oleate is a
constituent of the lipid particle core and remains within the core during hydrolysis of
triolein by LPL, therefore its removal is indicative of remnant clearance.
Quantitation of high-affinity clearance of chylomicrons in vivo was achieved
by using normal and modified chylomicron-like emulsions. Aliquots of the two
emulsions were either injected on consecutive days or mixed immediately prior to
injection and injected separately into the recipient animal after first taking a blood
sample for measurements of plasma lipids. The total amount of lipid injected was <
2.5 mg, in a volume of less than 800pl. The clearance of normal chylomicron-like
emulsion represents total uptake, which includes both high-affinity (receptor) and
low-affinity (non-specific uptake) mechanisms. The modified emulsion gave a
measure of low-affinity uptake. High-affinity uptake was calculated as the difference
between total uptake and low-affinity uptake. Therefore, the A A C attributable to
receptor activity (AACrec) was derived by difference of the A A C for normal
emulsions less the A A C of modified emulsions (AACtotai - AACnsu).
Total Uptake = High Affinity Uptake A n d L o w Affinity Uptake
The A A C calculations represent the mass of lipid cleared from plasma,
therefore A A C was chosen as the appropriate tool for measuring emulsion clearance
as it provides an indication of the amount of lipid cleared from plasma by high and
low affinity uptake mechanisms.
2.8 Organ Removal and Lipid Extraction
After completion of the clearance study, the animals were injected with a lethal dose
of sodium pentobarbitone (Nembutal®) and removed from the restraint cage. A cut
was made from the lower abdomen to the borders of the rib cage and the liver and
spleen were removed. Lipids of the liver and spleen were extracted within 4 hr of
removal from the animals using a modified procedure of Folch et al. (Folch et al,
1957). Adipose tissue and mesentery were removed from the organs before lipid

130
extraction. The organs were weighed, minced and 0.5-1.0 g m portions of the minced
organs extracted with 20 vol. of chloroform/methanol (2:1, v/v).
After the organs had been extracted the chloroform/methanol/organ mixture
was then filtered through Whatman N o 1 filter paper into glass counting vials and the
organs were further washed with 2 ml of chloroform: methanol (2:1, v/v). 10 ml 0.03
M KC1 was added to the vial to allow the phases to separate. The chloroform layer
(infranatant) was removed and dried under a stream of nitrogen. The solvent was then
evaporated under a stream of nitrogen. The radioactivity in the purified lipid extract
was measured in 15 ml of scintillant (Emulsifier-Safe™, Packard).
2.9 Determination of Radioactivity
Radioactivity of plasma and organs after lipid extraction was determined in a
Beckman L S 3800 Liquid Scintillation Counter (Beckman Instruments Inc. U S A )
using a program designed to simultaneously measure the radioactivity of 3 H and 14C
in dpm, over 10 min. Quenching was corrected by the channel ratio method of
Hendler (Hendler, 1964). Samples were counted using Emulsifier-Safe™ (Packard)
scintillant. The radioactivity in each vial of dried lipid extracts was measured after 48
hr, after being kept in the dark to reduce chemiluminescence.
2.10 Laser Light Scattering
Laser light scattering using a BI-90 particle sizer (Brookhaven Instruments Corp.,
Ronkonkoma, N Y ) was used to determine the size of emulsion particles. A n emulsion
volume of 50 pi was suspended in distilled water in plastic cuvettes and placed into
the sample cell compartment of the particle sizer. Measurements of particle size are
made in groups of 100 cycles. After every group of cycles the polydispersity and
effective diameter are determined. These values characterise the mean diameter and
relative width of the size distribution for the sample.
2.11 Statistical Analysis
Statistical comparisons were made on the data of the plasma clearance (calculated
from the individual A A C values) by analysis of variance. The f-test for independent

131
means was used to compare differences of individual points. The clearance of
chylomicron-like lipid emulsions in control and W H H L rabbits were compared by t-
test for independent means. Probability values of < 0.05 were accepted as significant.
Potential relationship between two variables was assessed using the Pearsons
correlation coefficient.
Regression analysis was carried out using the least-squares method. The area
under curve was integrated, and descriptive statistics were calculated with computer
software (Prism, Graphpad software, C A , U S A ) . The 9 5 % confidence interval was
used as a measure of statistical significance, i.e. p < 0.05.
2.12 Materials
2.12.1 Preparation of Aqueous Solutions
All aqueous salt solutions were prepared by dissolving the salts in distilled water,
which had been filtered using the Milli-Q system (Millipore Inc, Milford, M A , U S A )
at a resistivity of 18.2. M£2cm"\
2.12.2 Radiochemicals
[3H]Cholesteryl oleate ether- 30-6- Ci/mmol
Cholesteryl! l-14C]oleate- 50-60 Ci/mmol
[3H] triolein
[l-14C]triolein
2.12.3 Solvents
Solvents used for assaying retinyl esters were HPLC grade and were filtered and
degassed prior to use. The remaining solvents used were of analytical grade and were
used without further purification.
(Amersham International)
(Amersham International)

2.12.4 C h e m i c a l s
All chemicals were of analytical grade unless specified otherwise.
Ammonium molybdate tetrahydrate (A.R.)
Ammonium bicarbonate
A N S A (l-amino-2 napthol-4-sulphonic acid)
Argon
Aquasil
Boric Acid
Butylated Hydroxy-toluene
Chromotropic acid disodium salt dihydrate
D-Glucose (A.R.)
Di-sodium hydrogen orthophosphate
Ethylenediamine tetraacetic acid di-sodium salt
Glucose
Glutathione, reduced crystalline
(-L-glutamyl-L-cysteinylglycine)
Heparin, sodium
HEPES (N-2 hydroxyethylpiperazine
N'-2-ethane sulfonic acid)
Hydrogen peroxide
Intralipid (10% w/v soybean oil;
1.2% w/v fractionated egg-phospholipids
and 2.25% w/v glycerol)
Iodine
Ketapex (Ketamine)
Nembutal®
Paraformaldehyde
Peroxidase
Polyvinylalcohol
Potassium Bromide
Potassium Dihydrogen Phosphate
Retinyl Palmitate
Retinyl Stearate
Retinyl Oleate
David Brown Scientific
Ajax Chemical Co. Ltd
Calbiochem
CIG, Western Australia
Pierce Chemical Co.
B D H Chemicals
Sigma Chemical Co.
B D H Chemicals
Ajax Chemical Co. Ltd
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd
Sigma Chemical Co.
Cmwlth Serum Lab
Sigma Chemical Co.
Ajax Chemical Co. Ltd.
Pharmacia (Sweden)
Mallinkrodt
Apex Lab. Pty
Australia
Abbott Lab., Australia
Sigma
Sigma Chemical Co.
B D H Chemicals
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd.
Sigma Chemical Co.
Dept. Chemistry, U W A
Dept. Chemistry, U W A

133
Retinyl Myristate
Sodium Azide
Sodium Bromide
Sodium Bicarbonate
Sodium Cacodylate
Sodium Chloride
Sodium Citrate
Sodium dihydrogen orthophosphate
Sodium Metabisulphite
Sodium Sulphate
Sodium Sulphite
Sucrose
Superglue™
Trichloracetic acid
Xylazine (Xylazine hydrochloride)
Dept. Chemistry, U W A
Ajax Chemical Co. Ltd.
Mallinkrodt Australia
Ajax Chemical Co. Ltd
Ajax Chemical Co. Ltd
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd.
Ajax Chemical Co. Ltd.
Selleys Chemical Co.
Ajax Chemical Co. Ltd.
Troy Lab. Pty Ltd.
2.12.5 Lipids
Cholesterol (>99% purity)
Cholesteryl oleate (>99% purity)
L-phosphatidylcholine dioleoyl
Triolein (>99% purity)
Nu Chek Prep, Elysian, M N
Nu Chek Prep, Elysian, M N
Sigma Chemical Company
Avanti Polar Lipids
(Birmingham, AL)
Nu Chek Prep, Elysian, M N
2.12.6 Thin Layer Chromatography (TLC)
Thin-Layer Chromatography Aluminium Plates
TLC Standards for neutral lipid
Kieselgel 60, Merck, Darmstadt
Nu-Chek Prep, Elysian, M N
2.12.7 Cannulae and Tubing
2.12.7.1 Arterial cannulae for carotid artery cannulation
Teflon tube (30G lightweight) Small Parts Inc., Miami,
(Wall 0.006", LD. 0.012")
27GX»/2" needles
FLA.
Terumo Corporation

134
2.12.7.2 Venous cannulae for jugular vein cannulation
Clear vinyl tube (medical grade) Dural Plastics and
(O.D. 0.8 mm, LD. 0.5 mm)
25G X 1" needles
Dialysis tubing (width 10 mm)
Engineering, Dural, N S W
Terumo Corporation
Union Carbide Co. (Aust.)
2.12.8 Emulsion Preparation Apparatus
Dynava™freeze-drying unit
Vibra cell high intensity ultrasonic processor
Optima™ ultracentrifuge
BI-90 Particle Sizer
Dynavac, Aust.
Daintree Industries, USA
Beckman Inst. Corp., Palo Alto,
USA
Brookhaven Instruments Corp.,
Ronkonkoma, N Y
2.12.9 Fluorescent Emulsion Preparation Apparatus
[cholesteryl BODIPY FLC12 ]
(Cholesteryl-4,4-difluoro-5,7-dimethyl-4-
bora-3oc4a-diaza-s-indacene-3-dodecanoate)
Sofijet sterile dental needles - 30 G (0.3 x 23am)
Molecular Probes Inc.
(Eugene, OR)
Sofic, BP, Mazamet, France.

135
Chapter 3: Clearance Kinetics of
Chylomicron-Like Emulsions In
Vivo
3.1 Introduction
The triacylglycerols present in lipoproteins transporting lipids from the intestine and
from the liver (i.e. chylomicrons and V L D L ) are rapidly hydrolysed by the enzyme
L P L at the endothelial surface of capillaries in adipose tissue, skeletal muscle, cardiac
muscle and other sites (Havel, 1986b). W h e n lipolysis is almost complete, a
cholesterol-residual lipoprotein, called the chylomicron remnant, is released back in
the circulation (Redgrave, 1983). The remnant lipoprotein is then removed by the
liver, via a receptor-mediated process with apolipoprotein E as the ligand (Plump et
al, 1992), (Wilson etal, 1991), (Funahashi etal, 1989).
As discussed in Chapter 1, the mechanisms and pathways responsible for high-
affinity chylomicron remnant removal have received considerable attention, however
a consensus on the events underlying remnant removal has not been reached. At least
two genetically and functionally distinct receptors are proposed in the process of
remnant uptake. The receptor primarily responsible for chylomicron removal is
considered to be the LDL-receptor. A second pathway responsible for chylomicron
remnant uptake other than the LDL-receptor, was suggested by Kita et al. (Kita et al,
1982-b). However, Bowler et al. (Bowler et al, 1991) studied homozygote and
heterozygote W H H L rabbits in an attempt to identify the postulated second receptor.
Their findings did not support the existence of a second high-affinity removal

136
mechanism for chylomicron remnants and they concluded that the LDL-receptor is
the primary route of clearance of chylomicron remnants from the plasma.
Interestingly, heterozygous W H H L rabbits cleared L D L normally, however
chylomicron remnant clearance was ~ 5 0 % of control rabbits. This may indicate that
chylomicron clearance is more closely regulated by LDL-receptor activity than is
L D L clearance (Mamo et al, 1994). Beaumont and Assadollahi (Beaumont and
Assadollahi, 1990) found delayed chylomicron remnant metabolism in W H H L rabbits
and concluded that the hyperlipoproteinemia of the W H H L rabbits is at least partly of
exogenous origin and a defect in the apo B-100/E (LDL)-receptor may explain the
impairment. The LDL-receptor is suggested to account for approx. 8 0 % of total
remnant removal from the plasma (Mamo et al, 1991). There is high affinity binding
of the chylomicron remnant to the LDL-receptor, which occurs via the ligand apo E
(Floren etal, 1981).
Uptake mechanisms other than the LDL-receptor, which may be involved with
the removal of chylomicron remnants from plasma, include the scavenger cell
pathway, fluid endocytosis and the L D L receptor-like protein (LRP, also known as the
a2-macroglobulin receptor). Such uptake mechanisms are suggested to be low-
affinity, with both the scavenger cell pathway and endocytosis characterised as
receptor-independent mechanisms (Simionescu and Simionescu, 1991), (Herz, 1993).
The L R P receptor is expressed in the liver, although unlike the LDL-receptor, L R P
expression is not regulated in response to ligand binding and internalisation (Herz,
1993). However, L R P will only bind to lipoproteins artificially enriched in vitro with
apo E. Native lipoproteins (even those with high endogenous apo E content) do not
bind effectively (Fielding, 1992) to LRP. Therefore, it is not clear whether L R P plays
a significant role in clearance of remnants in the liver in vivo.
Emulsion models of triglyceride-rich lipoproteins containing triolein,
phospholipid, cholesteryl oleate and cholesterol have been shown to be metabolised
like natural chylomicrons when injected into conscious rats (Redgrave and Maranhao,
1985), (Redgrave et al, 1991), (Martins et al, 2000a), rabbits (Bowler et al. 1991,
(Redgrave et al, 1995), and man (Redgrave et al, 1993), (Maranhao et al, 1996),
(Martins et al, 1995). The clearance of chylomicron-like lipid emulsions is indicative
of high-affinity (receptor) plus non-specific uptake (nsu) mechanisms. The clearance
of chylomicron triglycerides reflects the sum of two processes, lipolysis and particle

137
uptake, while clearance of chylomicron cholesteryl ester reflects tissue uptake from
the plasma of remnant particles in various states of delipidation (Redgrave, 1999),
(Redgrave and Maranhao, 1985).
Maranhao et al (Maranhao et al, 1986) reported that the amount of
cholesterol in emulsion particles had a profound effect on their clearance in plasma,
and, in the absence of cholesterol, remnant particles were slowly cleared. The authors
reported that emulsions rich in cholesteryl ester but poor in free cholesterol were
metabolised like nascent chylomicron particles, whereas emulsions poor in cholesteryl
ester but rich in free cholesterol showed remnant-like behaviour. Further work with
emulsion lipid metabolism in vivo by Redgrave et al (Redgrave et al, 1987)
confirmed that free cholesterol was necessary for chylomicron-like emulsions to
mimic high-affinity clearance. The authors found that emulsions lacking cholesterol
were acted on by the enzyme L P L but the resultant triacylglycerol-depleted remnant
particle remained in the plasma instead of being rapidly taken up by the liver.
Heparin-stimulated lipolysis failed to increase the rate of cholesteryl ester removal,
indicating that the enzyme L P L was not the limiting factor to remnant uptake in the
cholesterol free emulsions. The authors concluded that the presence of emulsion
cholesterol is a critical determinant of early metabolic events. Saito et al. (Saito et al,
1996) have also studied the effects of cholesterol, and found that when emulsions
contained no cholesterol, changes in the composition of the core lipids had little effect
on the properties of the surface. W h e n cholesterol was incorporated into the emulsion
particle, changes in the core lipids had pronounced effects on surface rigidity, and on
the binding of apolipoproteins.
The mechanism(s) of the cholesterol effect remains unclear, but it is possible
that cholesterol changes the properties of the surface. The alterations in the metabolic
behaviour are thought to arise from changes in the conformation and binding
properties of apolipoproteins. Comparison of the apolipoprotein profile showed that
emulsions with a high content of free cholesterol bound less A-I, A-IV and C
apolipoproteins with relative increases in the amount of apo E (Maranhao et al,
1986). Such a result suggested that free cholesterol was required for the association of
apolipoprotein E to the particle surface. The knowledge that apo E mediates remnant
uptake indicates that such emulsion particles may lack the ability to bind apo E to the
particle surface. Alternately, the bound apo E may have a decreased affinity to the
receptor, or both (Maranhao et al, 1986), (Redgrave et al, 1987). A close relationship

138
between conformational changes in apolipoproteins and the thermal transition of
surface and core lipids has been suggested (Mims et al, 1990), (Banuelos et al,
1995). It is well established that apo E is a necessary ligand for chylomicron remnant
uptake and it has been shown that the high-affinity LDL-receptor is predominantly
responsible for chylomicron remnant uptake in vivo (Bowler et al, 1991), (Fielding,
1992). Changes to the amount and proportion of relative apolipoproteins, which
surround the lipoprotein particle surface and direct the catabolism of the particle, may
also cause a defect in the removal of the lipoprotein from the plasma.
What is clear is that unesterified cholesterol plays an essential role in the
metabolism of chylomicrons and conferring remnant-like behaviour (Redgrave et al,
1987). Cholesterol significantly affects the structure and physical properties of
bilayers, and the cholesterol in chylomicrons probably alters the physical character of
the particle surface as the cholesterol concentration increases relative to
phospholipids. Cholesterol also influences the packing of the acyl chains of the
membrane bilayer. Cholesterol molecules are partly dissolved in the oily core of the
particle and partly interdigitated between the acyl chains of the phospholipids, with a
partition coefficient in a model system of about 24 hr in favour of the surface (Ekman
et al, 1988). In general, an ordering effect is seen for acyl chains in the liquid-
crystalline state (Levine and Wilkins, 1971). Cholesteryl ester influences the partition
of cholesterol between the core and surface of triglyceride-rich particles (Li and
Sawyer, 1993). Cholesterol, in turn, affects the partition of cholesteryl esters between
the core and surface (Li et al, 1990), (Li and Sawyer, 1992). In model systems,
cholesterol packs with precise stoichiometrics, of one cholesterol for each
phospholipid, or one cholesterol for every two phospholipid molecules (Presti et al,
1982). Natural membranes possibly consist of domains where there is a 1:1
stoichiometry, mingled with domains of phospholipid lacking cholesterol, or of
domains with a 1:2 stoichiometry. Cholesterol may be needed at the surface of
remnant emulsions to promote the binding of apolipoprotein E in a conformation
appropriate for recognition as a ligand by specific hepatic receptors (Redgrave et al,
1987).
Chylomicron-like emulsion particles are similar in size and composition to
nascent chylomicrons. These are made from purified lipids and contain no proteins
(Redgrave and Maranhao, 1985) but acquire apo E once in the circulation. Clearance
of chylomicron-like lipid emulsions is indicative of high affinity (receptor) plus non-

139
specific uptake (nsu) mechanisms. W h e n the usual component of unesterified (free)
cholesterol ( 2 % of total lipid mass) is omitted from the emulsion mixture, the
resultant emulsion particles are consistent with chylomicrons in size, and interact
efficiently with hydrolytic enzymes in vivo, and are thus converted to a remnant at the
same rate. However, the 'modified' chylomicron-like emulsion particles do not
interact with high affinity uptake (receptor) pathways and remain in plasma. Thus
receptor-mediated uptake is delayed or prohibited, and the particles are taken up via
low-affinity mechanisms (Redgrave et al, 1987), (Bowler et al, 1991), ( M amo et al,
1991), (Redgrave et al, 1995). The clearance from plasma of the modified emulsion
particles is therefore representative of non-specific uptake (nsu), or low-affinity
mechanisms. A difference in clearance of normal emulsion particles (receptor + nsu)
versus modified particles (nsu) provides a measure of net receptor uptake in vivo.
Because chylomicron remnants and L D L are primarily cleared via the L D L -
receptor, their concentration in plasma is indicative of receptor activity. The objective
of this set of experiments was to investigate the validity of these two emulsions as a
model for monitoring receptor-mediated uptake. To achieve this, the clearance of
chylomicron-like emulsions with or without the normal component of unesterified
cholesterol was compared in control and homozygous W H H L rabbits. The rabbit is a
suitable model for the study of hyperlipidemia and atherogenesis, since it has been
shown that rabbits use the same method for the transport of lipid into atherosclerotic
plaque as occurs in human aortic lesions (Walton, 1973), utilising C E T P for transfer
of lipoprotein cholesterol. In man and rabbits, C E T P plays a central role in the
transfer of cholesteryl esters from H D L to other lipoproteins and since the diagnostic
assay is based on cholesteryl ester kinetics in vivo, it was necessary to establish that
cholesteryl ester transfer is minimal.
Specifically, W H H L rabbits lack functioning L D L receptors and are a model
for FH. The clearance of L D L in F H patients and W H H L rabbits is severely impaired
but both are said to have normal clearance of chylomicrons (Kita et al, 1982-b).
Hence it has been suggested that chylomicron remnants are taken up via a mechanism
genetically distinct from the LDL-receptor. However, previous studies with W H H L
rabbits found no difference in plasma clearance, when normal or modified emulsions
were injected (Bowler et al, 1991), ( M amo et al, 1991). Consistent with the absence
of functional L D L receptors in W H H L rabbits, the authors demonstrated that there is
no high affinity mechanism for remnants in W H H L rabbits. Utilising a [14C] breath

140
test, Redgrave et al. (Redgrave et al, 1995) found the appearance of C 0 2 in breath
was much slower in homozygous W H H L rabbits than in normal control rabbits, while
in heterozygous W H H L rabbits, an intermediate level of appearance was found.
Our laboratory has postulated that chylomicron-like emulsion clearance may
be impaired in the homozygous W H H L rabbits by competition with V L D L remnants
(DDL) for the LDL-receptor. To investigate this possibility the amount of receptor-
mediated chylomicron remnant clearance was measured in W H H L rabbits. In
addition, W H H L rabbits were utilised to investigate the possibility that chylomicron
remnants are catabolised via a receptor distinct from the LDL-receptor. To determine
the proportion of chylomicron clearance that is receptor-mediated the clearance of
normal and modified chylomicron-like emulsions were compared in individual
rabbits. Emulsions that lack unesterified cholesterol do not bind apo E and are not
recognised by receptors (Redgrave et al, 1987) and are used to determine the amount
of non-receptor mediated (low-affinity) chylomicron clearance. The two emulsion
types were injected simultaneously, and the clearance of emulsion particles compared.
Receptor-mediated clearance was calculated as the difference in clearance of the two
emulsion types. The organ uptake of chylomicron-like emulsions was also compared
in control and W H H L rabbits.
In keeping with the hypothesis that chylomicron remnants are cleared in vivo
via the LDL-receptor, it was expected that there would be no difference in the
clearance of the modified chylomicron-like emulsion, in control or W H H L rabbits.
Alternately, the clearance of the normal chylomicron-like emulsion would be
expected to vary significantly between the rabbit types, with control rabbits utilising
the rapid LDL-receptor pathway for chylomicron clearance, while W H H L rabbits
utilise an alternate, low-affinity pathway. W H H L rabbits lack LDL-receptors so any
receptor-mediated chylomicron-like emulsion removal should indicate the presence of
an alternate receptor and decreased clearance through competition, and that the L D L -
receptor deficiency has a direct effect on chylomicron remnant metabolism.
To determine the most appropriate approach to the two-emulsion method, the
clearance kinetics of separate and simultaneous injection protocol were compared.
The rationale for performing clearance studies simultaneously is that this is the
preferred method for use in human subjects. Previous clearance studies have assessed
normal and modified emulsion kinetics on consecutive days, which is not appropriate
to human subjects due to logistics of attendance and day-to-day variability in

141
lipoprotein metabolism. The single injection method provides an indication of the
metabolism of both emulsion types at a single point in time. M a m o et al. (Mamo et
al, 1994) utilised the simultaneous injection method, however the lipid load was very
small (< 2.5 m g ) and unlikely to effect lipid kinetics in vivo. In this series of
experiments, the lipid mass injected was increased as a result of simultaneous
injection of the two emulsion types. To study the kinetics of the two emulsion types
when injected separately, the clearance of normal and modified chylomicron-like
emulsions was studied in individual control rabbits in vivo. This approach allowed a
period of 24 hr for clearance of radioactivity before injection of the second emulsion,
and avoided any possible contamination of the modified emulsion with esterified
cholesterol. The clearance data was then compared with data obtained from
simultaneous emulsion injection studies.
All experiments were repeated using the rodent (rat) as an animal model, as
blood samples are easily obtained, and previous work has established that the same
mathematical model can accurately describe triglyceride and cholesteryl ester
clearance data from human and rat emulsion studies (Redgrave et al, 1993). Unlike
rabbits, rats do not produce CETP, however the use of rats as an experimental model
for studying the in vivo kinetics of chylomicron-like emulsions is well established in
our laboratory ( M a m o et al, 1993), (Martins et al, 1994), (Redgrave et al, 1992-b),
(Redgrave et al, 1991), (Redgrave and Maranhao, 1985).

142
3.2 Special Methods
3.2.1 Animals
Male New Zealand White (NZW) and semi-lop (NZW/ - cross) rabbits, weighing
between 1.8 to 3.5 kg, were obtained from the Animal Resources Centre, Murdoch,
Western Australia and were used as controls. W H H L rabbits were from a colony
maintained at the Biological Sciences Animal Unit at the University of Western
Australia. Male albino Wistar rats, weighing between 250 to 400 gm, were obtained
from the Animal Resources Centre, Murdoch, Western Australia.
3.2.2 Experimental Procedure for Chylomicron-Like Emulsion
Clearance in Rats
In the first set of clearance studies in rats, chylomicron-like emulsions (normal and
modified) containing radioactive cholesteryl[14C]oleate and [3H]triolein were
prepared (described in Chapter 2). The carotid artery and the left jugular vein of rats
were cannulated and injected with an aliquot of normal or modified emulsion. Rats
were sacrificed following each clearance study; therefore different sets of rats were
used to study the kinetic properties of the two emulsion types. The plasma lipid
concentration profiles of rats were determined before experimentation. The average
plasma concentrations for triglyceride and cholesterol were 0.95 mg/ml and 0.59
mg/ml, respectively, with little variation. Approx. 2 m g emulsion triglyceride in a
volume of 0.35 - 0.4 ml was injected into the jugular venous cannula as a bolus dose.
The mass of emulsion triglyceride injected was equivalent to 2 0 % of the recipient
animals' total plasma triglyceride pool. Blood samples of 0.35-0.4 ml were removed
at 0, 1.5, 3, 5, 8, 12, 20 and 30 min. After completion of the clearance study, 20 m g of
sodium pentobarbitone was injected and organs removed from the rats were processed
as described in Chapter 2. Plasma samples were measured for emulsion cholesteryl
ester and triglyceride radioactivities, and clearance of emulsions was determined from
the decline in plasma radioactivities.
In the second set of emulsion clearance studies in rats, the normal and
modified chylomicron-like lipid emulsions contained different isotopic forms of
cholesteryl oleate so that clearance from plasma of the two emulsion types could be

143
distinguished (Bowler et al, 1991), ( M a m o et al, 1991). Emulsion triolein was not
monitored. The procedures described above were used, with the exception that both
normal and modified emulsions were injected simultaneously. The emulsions were
mixed immediately prior to injection, and approx. 3 m g emulsion triglyceride was
injected. The mass of emulsion triglyceride injected was equivalent to 3 0 % of the
total plasma triglyceride pool, and it is known from previous experiments in our
laboratory that up to 5 0 % of the total triglyceride pool can be injected without
saturating clearance kinetics in the rat.
3.2.3 Experimental Procedure for Chylomicron-Like Emulsion
Clearance in Rabbits
In the first set of emulsion clearance studies in rabbits, the normal and modified
chylomicron-like emulsions were injected on separate days, to allow sufficient time
for the radioactivity in plasma to clear and to avoid contamination of the modified
emulsion with esterified cholesterol. Both emulsions contained radiolabeled triolein
and cholesteryl oleate to monitor chylomicron hydrolysis and particle uptake,
respectively. In the second set of experiments, emulsions were injected
simultaneously into rabbits. The two emulsion types were mixed immediately prior to
injection. The rationale for performing clearance studies simultaneously is that this is
the preferred method for use in human subjects. The same animals were re-used for
all experiments, thus acting as controls for the clearance of two emulsion types.
Injection of tracer was into a lateral ear vein of conscious, restrained rabbits.
The total amount of emulsion triglyceride injected was between 2.5 -3.5 m g per
rabbit in a ma x i m u m volume of 800 „1 for separate injection studies (equivalent to 4-
5 % of the recipient animals' total plasma triglyceride pool). For simultaneous
injection studies, a total of 5-7 m g triglyceride was injected in a maximum volume of
1000 ul, equivalent to 3-10% of the total plasma triglyceride pool of W H H L and
control rabbits, respectively. Blood samples of 0.5-1.0 ml were subsequently taken
from the opposite ear vein at regular time intervals between 3 and 40 min for
chylomicron studies. After completion of the clearance study, the plasma samples
were measured for emulsion cholesteryl ester radioactivities. At the completion of this
set of experiments, a lethal dose of sodium pentobarbitone was injected and organs
removed and processed as described in Chapter 2.

144
3.2.4 E m u l s i o n Clearance from P l a s m a
The clearances of the normal and modified chylomicron-like emulsions, radiolabeled
with cholesterol[14C]oleate and [3H]triolein, and cholesterol [3H] oleate and
[14C]triolein, respectively, were measured. Clearances of emulsion triglyceride and
cholesteryl oleate were determined from the decline in plasma radioactivities.
Clearance of emulsion was bi-exponential over 1.5 to 40 min for rats and 3 to 40 min
for rabbits. The radioactivity in plasma at each time point was expressed as a percent
of the injected dose (as described in Section 2.7).
3.2.5 Calculations
Emulsion plasma clearance kinetics were calculated as described in Section 2.7.4.
Area above curve values were used for analysis of plasma clearance of chylomicron
remnants. The A A C values for radiolabeled chylomicron-like emulsion triglyceride
and cholesteryl oleate were calculated for normal and modified chylomicron-like
emulsions, to provide a measure of the amount of radiolabeled lipid cleared from
plasma, in rat and rabbit studies. The A A C values were utilised in these studies as a
measure of plasma clearance, with high affinity and low affinity uptake represented
by the clearance of normal and modified emulsion cholesteryl oleate, respectively,
and high-affinity uptake calculated as the difference in clearance between the two
emulsion types.
3.2.6 Statistical Analysis
The statistical significance between group means of the plasma clearance was tested
by student's t-test. The clearance of chylomicron-like lipid emulsions in control and
W H H L rabbits were compared by f-test for independent means. Probability values of
< 0.05 were accepted as significant. Descriptive statistics were calculated with
computer software (Instat), with p values of < 0.05 accepted as statistically
significant.

145
3.2.7 Analysis of Emulsion Composition
The lipid composition of chylomicron-like emulsions was determined using TLC. The
emulsion lipids (0.5 ml) were extracted in glass stoppered tubes with 2.5 ml of
methanol, 5 ml of chloroform and 2.5 ml of D D W as described (Folch et al, 1957).
The tubes were mixed and allowed to stand overnight for adequate separation of the
phases. A volume of 4.5 ml of chloroform was removed from the lower phase and
dried under nitrogen. A n aliquot of 150 pi of chloroform: methanol mixture (2:1, v/v)
was added to the tube and 100 pi of this solution was applied in a narrow band to
T L C plates precoated with 0.2 m m layers of silica gel (Merck). The samples were
developed for 1-2 hr in a glass tank in the solvent system: petroleum ether (40-
60°C)/ether/acetic acid (90:10:1, v/v). The bands of different lipids were visualised
using iodine vapour and separated in sequence. The triglyceride, cholesteryl ester and
unesterified cholesterol were scraped from the plate. Triglyceride was then extracted
into chloroform and assayed, and the unesterified cholesterol and cholesteryl ester
bands were hydrolysed and assayed. Phospholipid was measured directly on emulsion
and lipoprotein suspensions. The size of the emulsion particles was determined by
laser light scattering using a BI-90 particle sizer (Brookhaven Instruments Corp.,
Ronkonkoma, N Y ) .
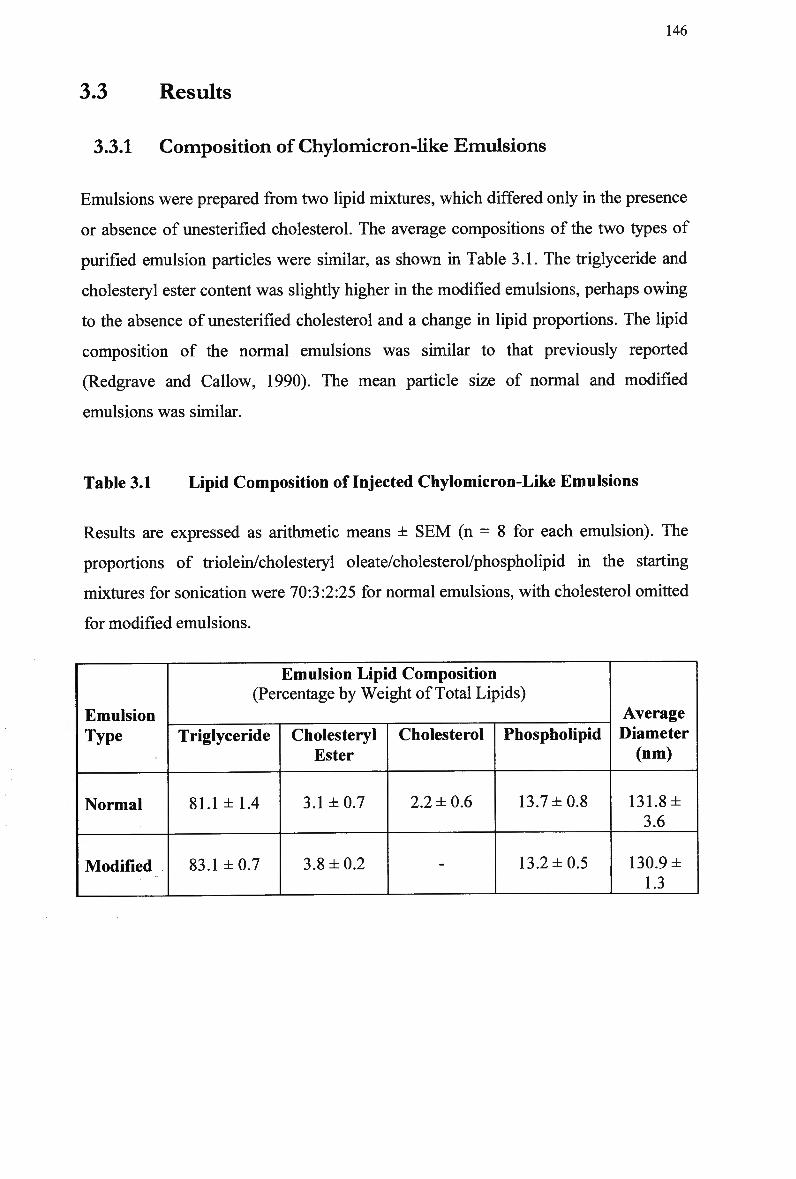
146
3.3 Results
3.3.1 Composition of Chylomicron-like Emulsions
Emulsions were prepared from two lipid mixtures, which differed only in the presence
or absence of unesterified cholesterol. The average compositions of the two types of
purified emulsion particles were similar, as shown in Table 3.1. The triglyceride and
cholesteryl ester content was slightly higher in the modified emulsions, perhaps owing
to the absence of unesterified cholesterol and a change in lipid proportions. The lipid
composition of the normal emulsions was similar to that previously reported
(Redgrave and Callow, 1990). The mean particle size of normal and modified
emulsions was similar.
Table 3.1 Lipid Composition of Injected Chylomicron-Like Emulsions
Results are expressed as arithmetic means ± SEM (n = 8 for each emulsion). The
proportions of triolein/cholesteryl oleate/cholesterol/phospholipid in the starting
mixtures for sonication were 70:3:2:25 for normal emulsions, with cholesterol omitted
for modified emulsions.
Emulsion Type
Normal
Modified
Emulsion Lipid Composition (Percentage by Weight of Total Lipids)
Triglyceride
81.1 ±1.4
83.1 ±0.7
Cholesteryl Ester
3.1 ±0.7
3.8 ±0.2
Cholesterol
2.2 ± 0.6
-
Phospholipid
13.7 ±0.8
13.2 ±0.5
Average Diameter (nm)
131.8± 3.6
130.9 ± 1.3

147
3.3.2 Removal From Plasma of Injected Emulsion Lipids
Following Separate Injection of the Two Emulsion Types in
Rats
Initial studies of emulsion lipid clearance were done using the rat as an animal model,
as blood samples are easily obtained and rats have been utilised extensively as an
experimental model for studying the in vivo kinetics of chylomicron-like emulsions in
our laboratory. The clearance of normal and modified emulsions was compared with
previous observations that the absence of unesterified cholesterol in emulsions
delayed the clearance of remnant particles. Modified emulsion particles were
synthesised by excluding unesterified cholesterol, in order to prevent receptor-
mediated uptake. In this study, cholesteryl oleate and triolein in normal and modified
emulsion particles were differentially radiolabeled and injected separately into
unanaesthetised rats to demonstrate the difference in remnant uptake. The A A C
values for normal and modified emulsions are presented in Table 3.2, and clearance
curves presented in Figures 3.1-3.2.
The general metabolic pattern for the normal emulsion was similar to the
behaviour of natural chylomicrons (Redgrave and Maranhao, 1985), with a lower
mean plasma clearance for cholesteryl ester than triglyceride (Table 3.2). Hydrolysis
of emulsion triglycerides and the removal of remnants occur in the initial rapid phase
(Lenzo et al, 1988). Consistent with previous findings (Redgrave et al, 1987),
normal emulsions hydrolysed quickly, with more than 9 0 % of particle triolein
disappearing from the plasma by 12 min after injection (Figure 3.1), followed by a
rapid disappearance of particle remnants, with over 9 0 % of labelled cholesteryl oleate
removed by 30 min. This behaviour reflects the normal pattern for chylomicron
metabolism, i.e., triglyceride clearance proceeds first because removal is via two
processes, lipolysis within the plasma to form triglyceride depleted remnants,
followed by remnant uptake and removal of residual triglyceride, whilst cholesteryl
ester clearance can only occur during remnant uptake. The modified emulsion was
also hydrolysed rapidly, with more than 9 0 % of particle triolein disappearing from
plasma by 12 min, however the particle removal was delayed compared with removal
of normal emulsion, with 5 0 % cholesteryl oleate still remaining in plasma at 30 min
(Figure 3.2).
In contrast to the emulsion containing cholesterol, the mean A A C for
radiolabeled cholesteryl oleate was less for the modified emulsion (2426.4 ± 124.3

148
and 1217.5 ± 87.9, respectively). This difference was statistically significant. In this
study, as with previous chylomicron clearance studies, the clearance of radiolabeled
cholesteryl oleate was indicative of remnant removal. The slower phase of removal of
modified emulsions represents a delay in the uptake of remnant lipoprotein particles
(Figure 3.2). The removal of emulsion remnants via high affinity mechanisms
accounted for approx. 4 8 % of total uptake, while low affinity mechanisms accounted
for approx. 5 2 % of the total uptake in rats.
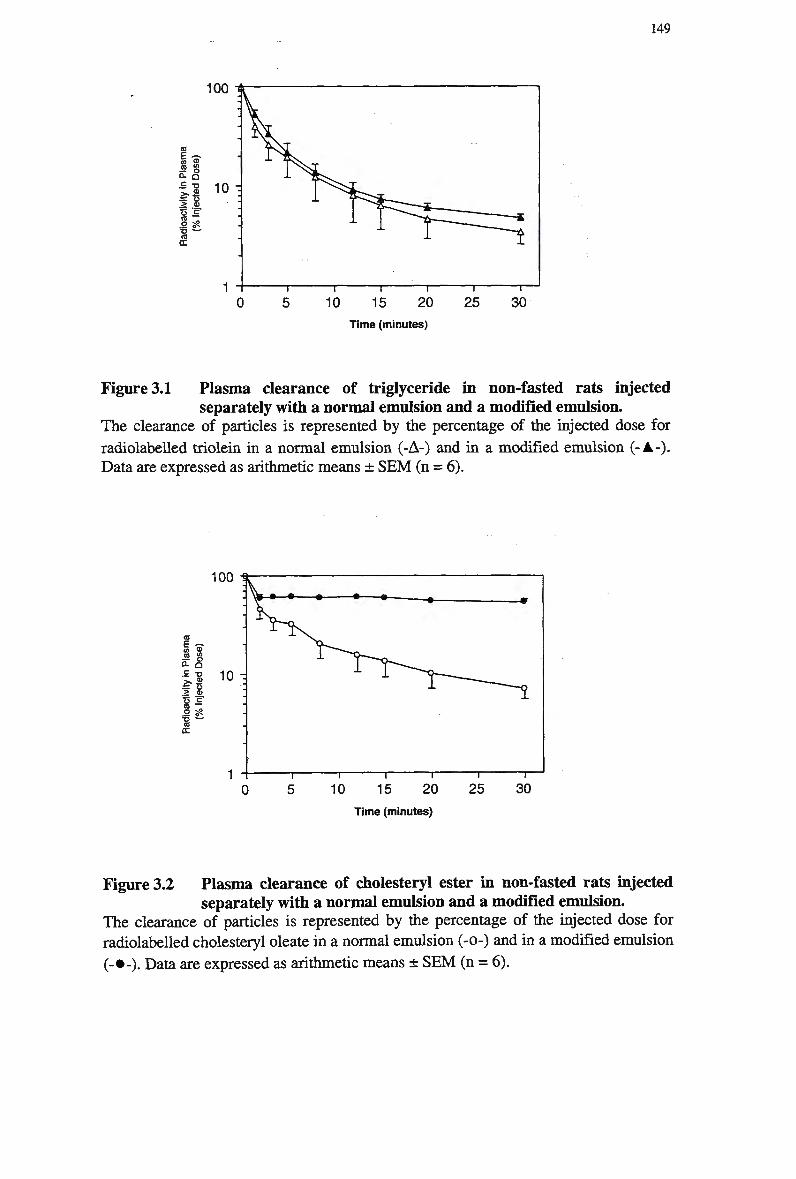
149
100
a
« s 0. _
f 8 .2 S«
DC
10 -
10 15 20 Time (minutes)
Figure 3.1 Plasma clearance of triglyceride in non-fasted rats injected separately with a normal emulsion and a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for
radiolabeled triolein in a normal emulsion (-A-) and in a modified emulsion (-A-). Data are expressed as arithmetic means ± S E M (n = 6).
Time (minutes)
Figure 3.2 Plasma clearance of cholesteryl ester in non-fasted rats injected separately with a normal emulsion and a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate in a normal emulsion (-0-) and in a modified emulsion
(-•-). Data are expressed as arithmetic means ± S E M (n = 6).

150
Table 3.2 Chylomicron-Like Emulsion Cholesteryl Oleate and Triolein Removal in Rats Following Separate Injection of T w o Emulsion Types
Emulsion Type
Normal
Modified
High-Affinity Uptake
Area Above Curve (AU)
CO
2425.01 ± 126.3
1226.3 ±81.5
t
1198.7 ±180.1
TO
2621.5 ±101.7
2553.4 ±55.9
The area above curve values (arbitrary units: A U ) for rats are tabulated for radiolabeled triolein (TO) and cholesteryl oleate (CO), following injection of normal and modified emulsions. Data are expressed as arithmetic means ± S E M (n = 6).
* p < 0.05, ** p<0.01,,f p< 0.001 vs clearance of normal emulsion.

151
3.3.3 Organ Uptake of Injected Emulsion Lipids Following
Separate Injection of the Two Emulsion Types in Rats
Normal and modified chylomicron-like emulsions were injected into unanaesthetised
rats. Liver uptake was measured at 30 min post-injection. Spleen uptake was also
monitored to exclude significant uptake by the reticuloendothelial system. The organ
uptake values for normal emulsions and modified emulsions are presented in Table
3.3, as a percent of injected dose.
Less than 1 % of radioactivities were recovered in the spleen and the
splanchnic uptake of cholesteryl oleate and triolein in modified emulsions was
significantly higher compared with normal emulsions. In contrast, hepatic recoveries
of triolein and cholesteryl oleate of modified emulsion were lower than for normal
emulsion. Recovery of cholesteryl oleate label in the liver was 9.4% for modified
emulsion, which was significantly less than for the normal emulsion (38.1%). The
recovery of triolein in liver of modified emulsion was also significantly lower than
that of normal emulsion (5.52% and 8.27%, respectively).
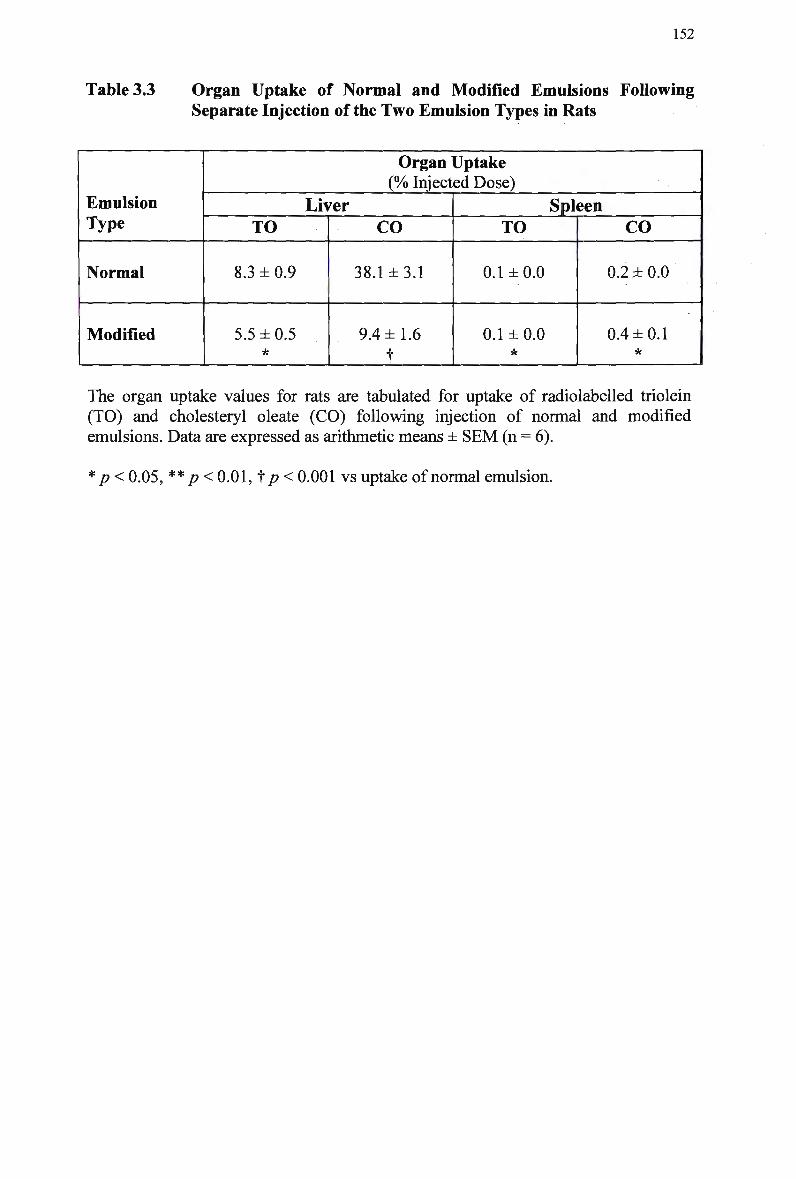
152
Table 3.3 Organ Uptake of Normal and Modified Emulsions Following Separate Injection of the Two Emulsion Types in Rats
Emulsion Type
Normal
Modified
Organ Uptake (% Injected Dose)
Liver TO
8.3 ± 0.9
5.5 ±0.5 *
CO
38.1 ±3.1
9.4 ±1.6
f
Spleen TO
0.1 ±0.0
0.1 ±0.0
CO
0.2 ± 0.0
0.4 ±0.1 *
The organ uptake values for rats are tabulated for uptake of radiolabeled triolein (TO) and cholesteryl oleate (CO) following injection of normal and modified emulsions. Data are expressed as arithmetic means ± SEM (n = 6).
p < 0.05, ** p<0.01,'fp< 0.001 vs uptake of normal emulsion.

153
3.3.4 Removal from Plasma of Injected Emulsion Lipids
Following Simultaneous Injection of the Two Emulsion
Types in Rats
Normal chylomicron-like emulsions were labelled with cholesteryl[14Cjoleate and
modified emulsions were labelled with cholesteryl[ Hjoleate, to allow the emulsion
particles to be monitored simultaneously in vivo. Simultaneous injection of the two
emulsion types was employed to provide an indication of the metabolism of receptor
activity at a single point in time, thereby avoiding the day-to-day variability in
lipoprotein metabolism. In addition, previous studies have not utilised this approach
to quantitating receptor uptake in rats. Normal and modified emulsions were injected
simultaneously into unanaesthetised rats. The A A C data was calculated from the bi-
exponential curves of residual plasma radioactivities between 0 and 30 min after
injection, and expressed as a percentage of the injected dose. The patterns of removal
from plasma of cholesteryl oleate are shown in Figure 3.3. The A A C data for normal
and modified emulsions are presented in Table 3.4.
The mean A A C for radiolabeled cholesteryl oleate (modified emulsion) was
1902.9 ± 101.9, significantly lower than for the normal emulsion (2470.8 ± 66.2). In
this experiment, as with the previous chylomicron clearance study utilising separate
injection of the two emulsion types, the clearance of radiolabeled cholesteryl oleate is
indicative of remnant removal. The slower initial phase of removal of modified
emulsions represents a delay in the uptake of remnant lipoprotein particles (Figure
3.3). The removal of emulsion remnants via high affinity mechanism accounted for
approx. 2 3 % of total uptake, while low affinity mechanisms accounted for approx.
7 7 % of the total uptake in rats.
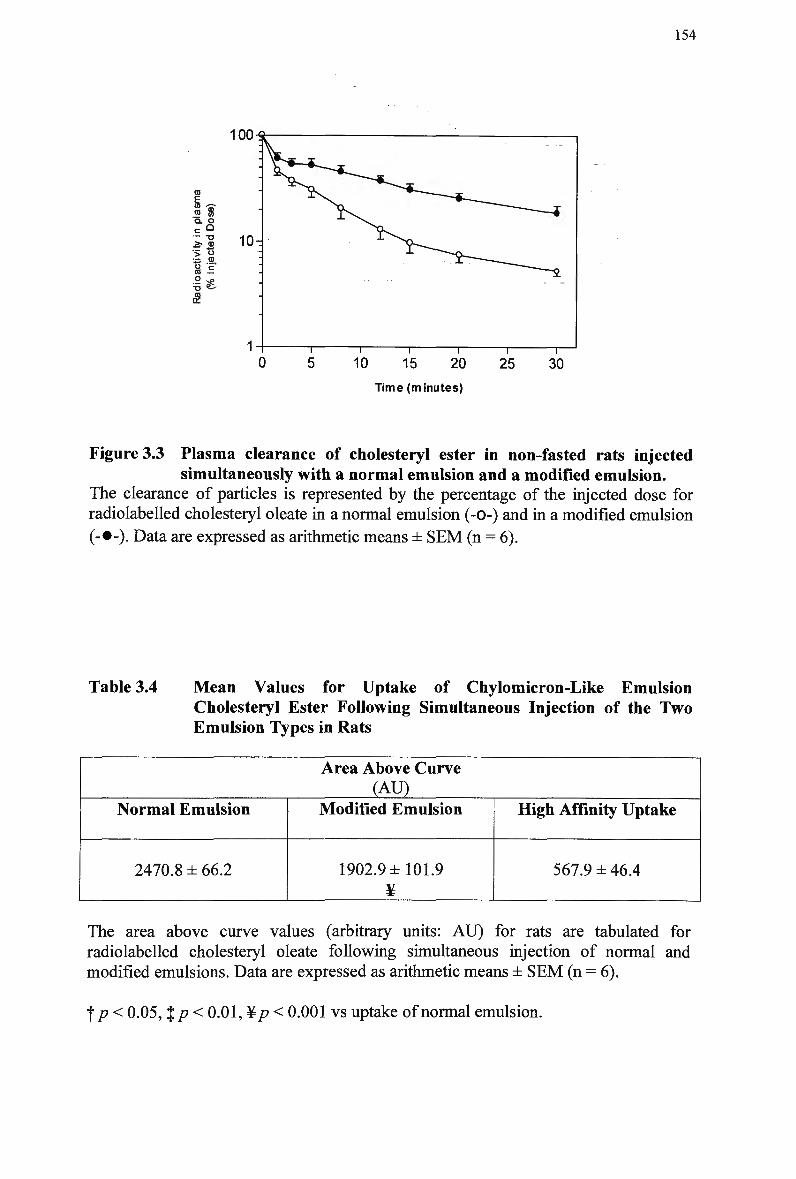
154
100-S
If Q. O — -o .& aj > o = (D
a '= CO —
- &
Time (minutes)
Figure 3.3 Plasma clearance of cholesteryl ester in non-fasted rats injected simultaneously with a normal emulsion and a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate in a normal emulsion (-0-) and in a modified emulsion
(-•-). Data are expressed as arithmetic means ± SEM (n = 6).
Table 3.4 Mean Values for Uptake of Chylomicron-Like Emulsion Cholesteryl Ester Following Simultaneous Injection of the Two Emulsion Types in Rats
Area Above Curve (AU)
Normal Emulsion
2470.8 ± 66.2
Modified Emulsion
1902.9 ±101.9
High Affinity Uptake
567.9 ± 46.4
The area above curve values (arbitrary units: AU) for rats are tabulated for radiolabeled cholesteryl oleate following simultaneous injection of normal and modified emulsions. Data are expressed as arithmetic means ± SEM (n = 6).
f p < 0.05, %p< 0.01, ¥/? < 0.001 vs uptake of normal emulsion.

155
3.3.5 Comparison of Emulsion Cholesteryl Ester Uptake
Following Separate versus Simultaneous Injection of the
Two Emulsion Types in Rats
The results from Sections 3.3.2 and 3.3.4 for control rats are compared. The rationale
for comparing separate and simultaneous injection was to determine if simultaneous
injection of the two emulsion types altered clearance kinetics of emulsion remnants in
vivo, as this has not previously been determined. Figure 3.4 shows the clearance of
emulsion cholesteryl oleate in rats, following separate and simultaneous injection of
the two emulsion types. The remnant uptake of normal emulsions was similar for both
injection modes, as evident by the A A C curve data and clearance curves. However,
the clearance of modified emulsion cholesteryl oleate was significantly greater
following simultaneous injection of the two emulsion types. Specifically, the modified
emulsion remnants cleared to a greater extent following simultaneous injection, with
2 0 % of injected dose remaining in plasma at 30 min compared with 5 5 % for separate
injection of the two emulsion types.
Normal chylomicron-like emulsions are cleared by low and high affinity
mechanisms, while modified emulsions are cleared by low affinity mechanisms only.
The difference between the clearances of the two emulsions can be largely accounted
for by receptor-mediated clearance, therefore the area between the two curves is
indicative of high-affinity clearance. The high affinity uptake as calculated by A A C
data, was significantly greater (2.1 times) following separate injection compared with
simultaneous injection of the two emulsion types (Table 3.5). High affinity
mechanisms accounted for 4 8 % and 2 3 % of the total emulsion uptake for separate and
simultaneous injection, respectively. Consequently, uptake of emulsion remnants via
low affinity pathways accounted for 5 2 % following separate injection and 7 7 % for
simultaneous injection.
The decrease in high affinity uptake values for the simultaneous injection
study was primarily due to an increase in the uptake of emulsion remnants by low
affinity mechanisms, represented by the uptake of modified emulsion cholesteryl
oleate (Figure 3.4). It is noteworthy that the amount of injected emulsion triglyceride
increased from 2 0 % of the total triglyceride pool for separate injections, to 3 0 % for
simultaneous injections. These values are both below the amount expected to saturate
remnant clearance kinetics in the rat (50%).
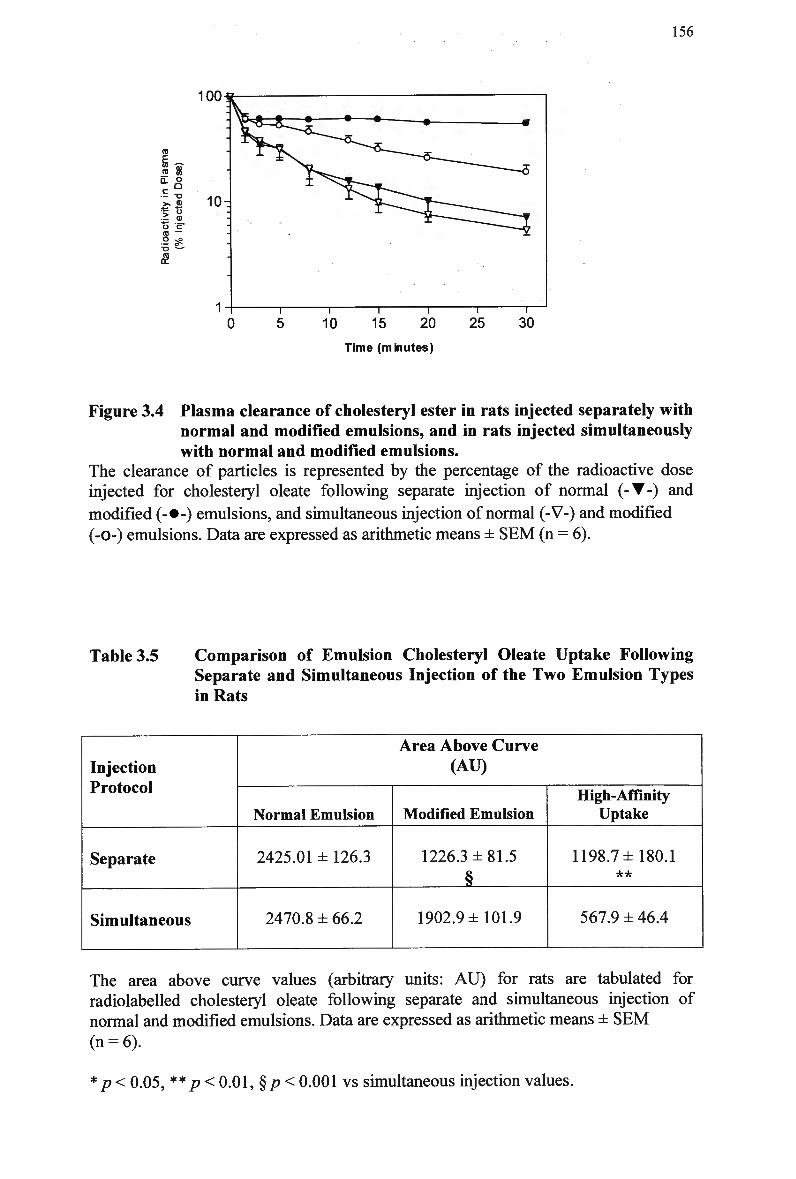
156
100
is
H >> s
1 S o —
CO — O o
&
Time (minutes)
Figure 3.4 Plasma clearance of cholesteryl ester in rats injected separately with normal and modified emulsions, and in rats injected simultaneously with normal and modified emulsions.
The clearance of particles is represented by the percentage of the radioactive dose injected for cholesteryl oleate following separate injection of normal (-T-) and
modified (-•-) emulsions, and simultaneous injection of normal (-V-) and modified
(-0-) emulsions. Data are expressed as arithmetic means ± S E M (n = 6).
Table 3.5 Comparison of Emulsion Cholesteryl Oleate Uptake Following Separate and Simultaneous Injection of the T w o Emulsion Types in Rats
Injection Protocol
Separate
Simultaneous
Area Above Curve (AU)
Normal Emulsion
2425.01 ± 126.3
2470.8 ± 66.2
Modified Emulsion
1226.3 ±81.5
1902.9 ±101.9
High-Affinity Uptake
1198.7 ±180.1 **
567.9 ± 46.4
The area above curve values (arbitrary units: A U ) for rats are tabulated for radiolabeled cholesteryl oleate following separate and simultaneous injection of normal and modified emulsions. Data are expressed as arithmetic means ± S E M
(n = 6).
* p < 0.05, **p< 0.01, §/? < 0.001 vs simultaneous injection values.

157
3.3.6 Removal from Plasma of Injected Emulsion Lipids
Following Separate Injection of the Two Emulsion Types in
Rabbits
The clearances of normal and modified chylomicron-like emulsions were studied in
rabbits to confirm the previous observations in rats, that the absence of unesterified
cholesterol in emulsions delayed the clearance of remnant particles. The rabbit is used
as an animal model for human studies as they utilise C E T P for transfer of lipoprotein
cholesterol. In m a n and rabbits, C E T P plays a central role in the transfer of
cholesteryl esters from H D L to other lipoproteins and it was necessary to establish
that cholesteryl ester transfer is minimal during this process.
Figures 3.5 and 3.6 compare the clearance from rabbit plasma of normal and
modified emulsion triglyceride and cholesteryl oleate, respectively. Consistent with
previous findings (Redgrave et al, 1987), clearance of modified emulsion particles
was impaired (as assessed by A A C ) , compared with normal emulsion clearance. At 8
min post-injection, 9 0 % and 7 5 % of triolein was remaining in plasma for normal and
modified emulsion, respectively. The difference between the triolein removal in
normal and modified emulsions was significant (Table 3.6). In addition, the absence
of cholesterol in the emulsions significantly delayed cholesteryl oleate clearance, with
6 0 % removed from the plasma 30 min after injection compared with 8 3 % for normal
emulsion.
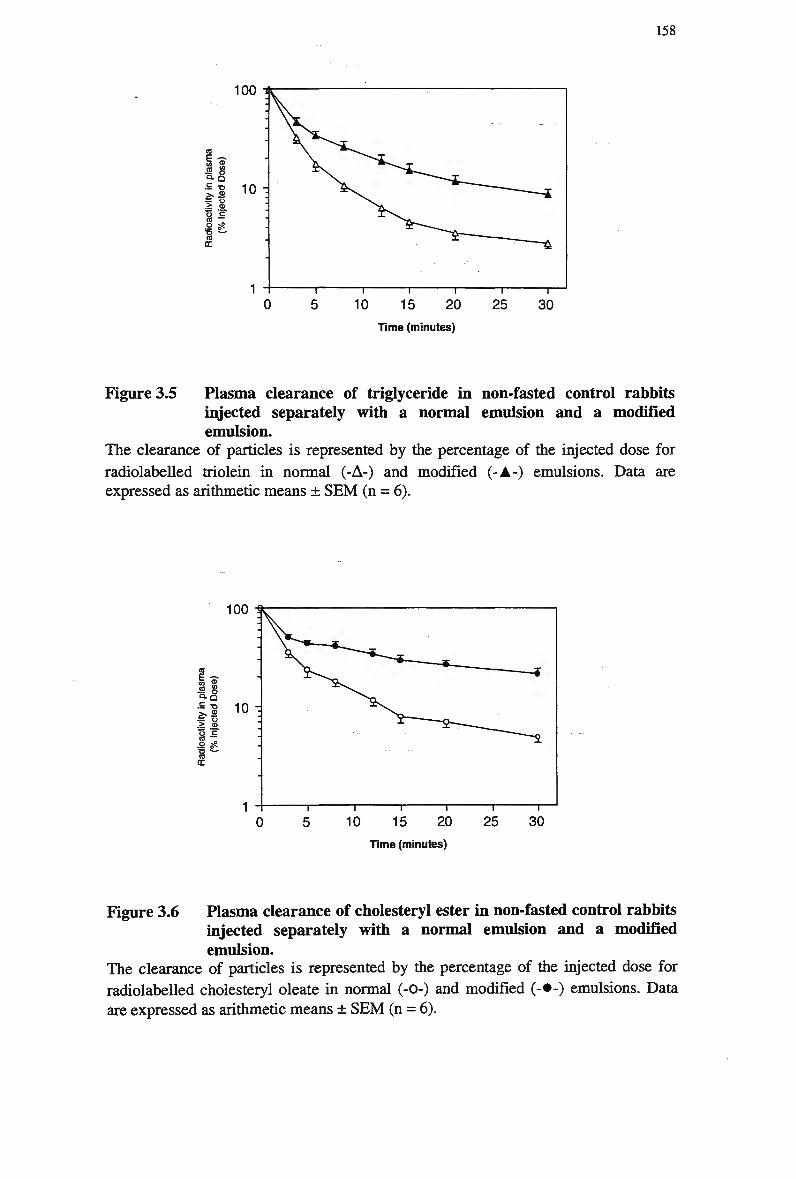
158
100
•S 6
£ -
> o> " _ •
CO —
° 5? "O — CO CC
10 -
10 15
Time (minutes)
Figure 3.5 Plasma clearance of triglyceride in non-fasted control rabbits injected separately with a normal emulsion and a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for
radiolabeled triolein in normal (-A-) and modified (-A-) emulsions. Data are expressed as arithmetic means ± S E M (n = 6).
100
Q.Q
& o > CD
10 -
Time (minutes)
Figure 3.6 Plasma clearance of cholesteryl ester in non-fasted control rabbits injected separately with a normal emulsion and a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for
radiolabeled cholesteryl oleate in normal (-0-) and modified (-•-) emulsions. Data are expressed as arithmetic means ± S E M (n = 6).
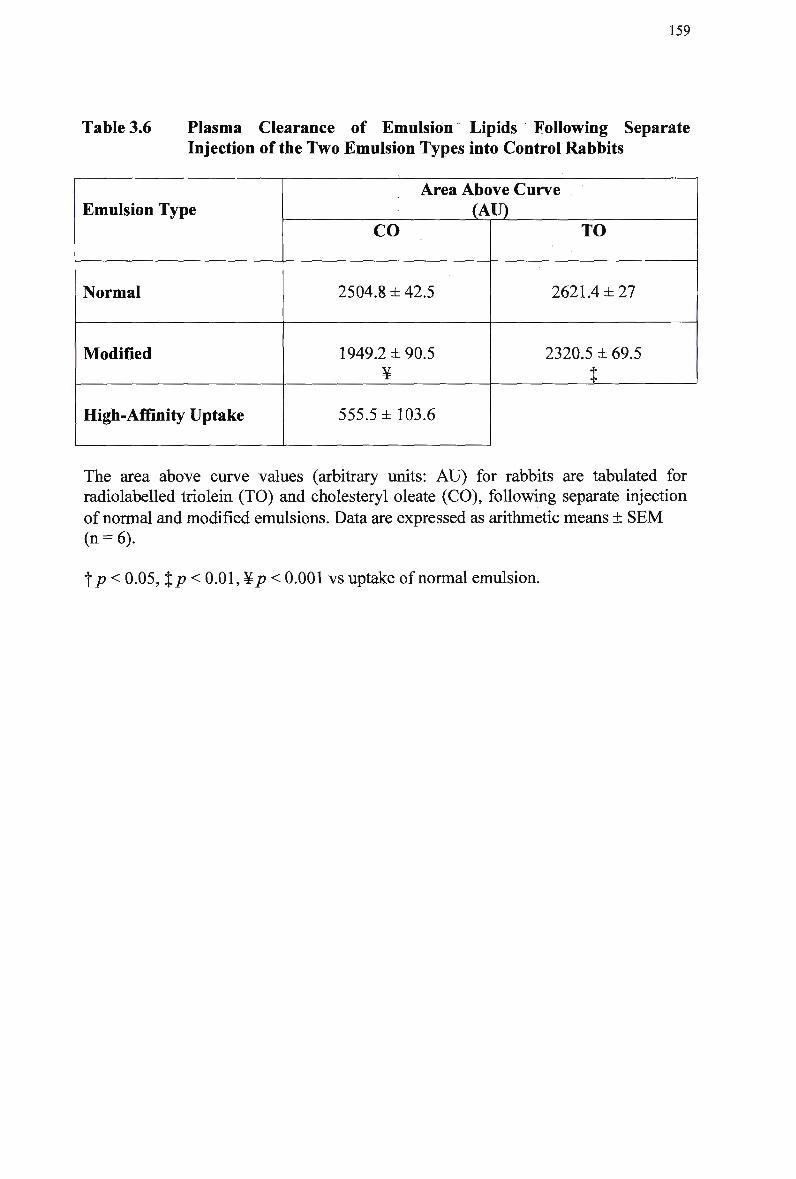
159
Table 3.6 Plasma Clearance of Emulsion Lipids Following Separate Injection of the T w o Emulsion Types into Control Rabbits
Emulsion Type
Normal
Modified
High-Affinity Uptake
Area Above Curve
J_VU) CO
2504.8 ± 42.5
1949.2 ±90.5 ¥
555.5 ±103.6
TO
2621.4 ±27
2320.5 ± 69.5
t
The area above curve values (arbitrary units: A U ) for rabbits are tabulated for radiolabeled triolein (TO) and cholesteryl oleate (CO), following separate injection
of normal and modified emulsions. Data are expressed as arithmetic means ± S E M (n = 6).
f p < 0.05, %p < 0.01, ¥/? < 0.001 vs uptake of normal emulsion.

160
3.3.7 Plasma Clearance of Emulsion Particles Following
Simultaneous Injection of the Two Emulsion Types in
Control and W H H L Rabbits
The in vivo kinetics of chylomicron-like emulsions in control and homozygous
W H H L rabbits were assessed. W H H L rabbits lack functioning L D L receptors and
take up chylomicron remnants via a mechanism genetically distinct from the L D L -
receptor. Normal and modified chylomicron-like emulsions were injected
simultaneously into unanaesthetised rabbits. Emulsions were differentially
radiolabeled with cholesteryl oleate to allow particle uptake to be monitored during
plasma clearance. As explained in Section 3.3.4, the simultaneous injection mode was
employed, as it is the preferred method for use in humans and provides an indication
of receptor activity at a single point in time. In addition, previous studies have not
assessed this approach to quantitating receptor uptake.
3.3.7.1 Plasma lipid concentrations for control and WHHL rabbits
The plasma lipid concentration profiles of control and homozygous WHHL rabbits
are provided in Table 3.7. Total plasma cholesterol and triglyceride concentrations
were determined before experimentation to detect any relationships between
lipoprotein clearance characteristics and plasma lipids. Plasma cholesterol
concentrations ranged from 0.64 to 1.8 mmoLl"1 in control rabbits, and from 8.7 to
13.5 mmoLl"1 in W H H L rabbits. Similarly, plasma triglyceride concentrations ranged
between 0.6 to 1.1 mmoLl"1 and 2.2 to 3.9 mmol.r1, in control and W H H L rabbits,
respectively. Compared with control rabbits, homozygous W H H L rabbits were
hypertriglyceridemic and hypercholesterolemic, and their cholesterol and triglyceride
concentrations were increased by 9.1 and 3.5 times, respectively. All lipid
concentrations tested in homozygous W H H L rabbits were significantly different from
control.
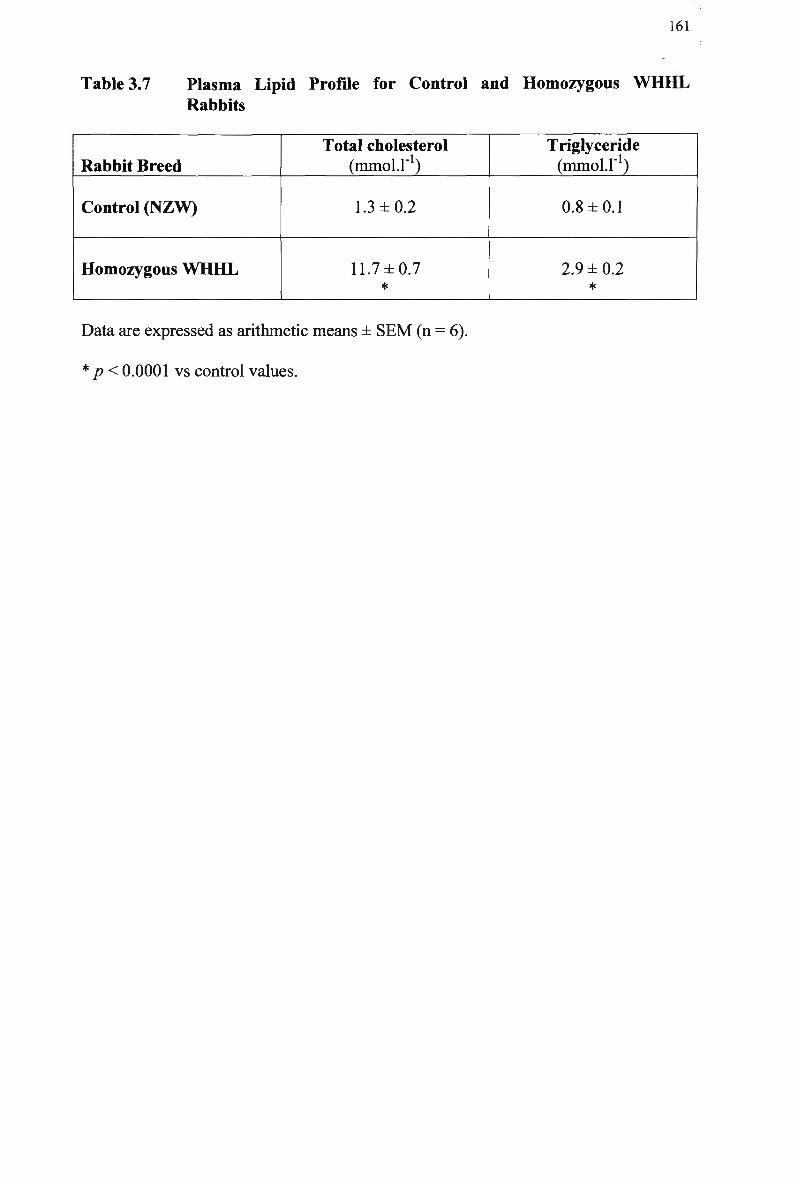
161
Table 3.7 Plasma Lipid Profile for Control and Homozygous W H H L
Rabbits
Rabbit Breed
Control (NZW)
Homozygous W H H L
Total cholesterol (mmol.r1)
1.3 ±0.2
11.7 ±0.7 *
Triglyceride (mmol.r1)
0.8 ±0.1
2.9 ±0.2 *
Data are expressed as arithmetic means ± SEM (n = 6).
* p< 0.0001 vs control values.

162
3.3.7.2 Removal from plasma of injected emulsion cholesteryl oleate following simultaneous injection of the two emulsion types in
rabbits
Table 3.8 lists the AAC derived from the clearances of the emulsion cholesteryl oleate
in control and homozygous W H H L rabbits. The clearance of emulsion cholesteryl
ester fitted the known mechanism by which chylomicrons are catabolised. In control
rabbits normal emulsion cholesteryl oleate was cleared more rapidly in the first 3 min
compared with the clearance of modified emulsion cholesteryl oleate (Figure 3.7).
The pattern of modified emulsion clearance was consistent with a defect in remnant
clearance, after normal depletion of emulsion triolein by the action of LPL. This is
reflected in the A A C data, which shows that the amount of cholesteryl oleate in
normal emulsions cleared from plasma was significantly greater compared with
modified emulsions (2437.3 and 2020 A.U., respectively). The clearance of normal
emulsion particles in W H H L rabbits was linear and varied little from the clearance of
modified emulsions (1858.1 and 1784.1 A.U., respectively).
Figure 3.7 compares the plasma clearance of a normal and modified emulsion
cholesteryl oleate in control and W H H L rabbits. At 30 min post injection, more than
9 0 % of the normal emulsion was cleared from the plasma of the control rabbits. In
contrast, only 3 5 % of the injected emulsion was cleared from the plasma of the
W H H L rabbits. The differences in clearance were statistically significant (p < 0.001).
In contrast, the plasma clearance of modified chylomicron-like emulsion cholesteryl
oleate in control and W H H L rabbits was similar (83% and 6 9 % , respectively).
To establish if modified emulsions utilised receptor uptake pathways in vivo,
receptor-mediated uptake was calculated as described (Table 3.8). Whilst there was
substantial receptor-mediated clearance of chylomicron-like emulsion cholesteryl
oleate by control rabbits, there was little clearance of chylomicrons by high affinity
uptake pathways in W H H L rabbits. Despite the larger contribution of receptor uptake
to particle uptake in control rabbits compared with W H H L rabbits (5.6 times), the
difference was not significant. W h e n calculated as a percent of the total emulsion
uptake, high affinity mechanisms accounted for 1 7 % of emulsion clearance in control
rabbits. Consequently, uptake of emulsion remnants via low affinity pathways
accounted for 8 3 % of emulsion clearance. In W H H L rabbits, there was no high
affinity mechanism evident ( 5 % high affinity mechanism was not significantly
different from zero).
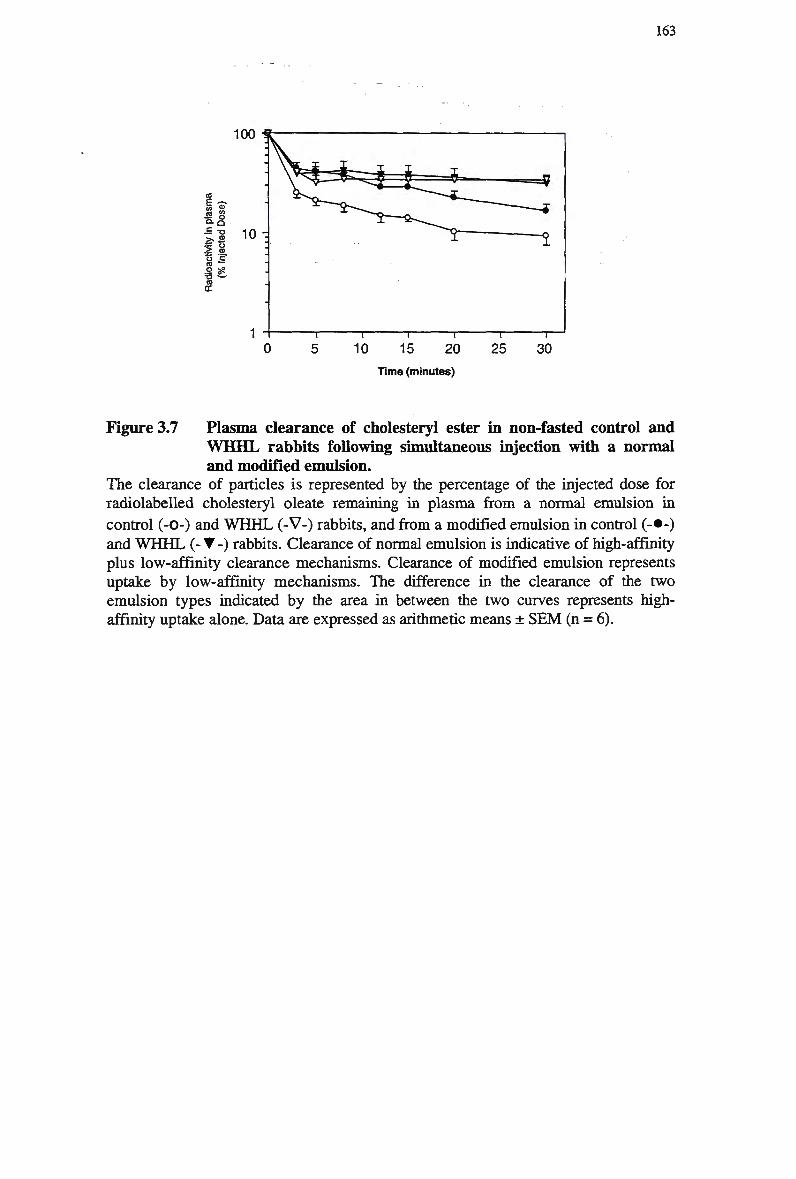
163
15 20 Time (minutes)
Figure 3.7 Plasma clearance of cholesteryl ester in non-fasted control and W H H L rabbits following simultaneous injection with a normal and modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate remaining in plasma from a normal emulsion in
control (-0-) and W H H L (-V-) rabbits, and from a modified emulsion in control (-•-) and W H H L (-•-) rabbits. Clearance of normal emulsion is indicative of high-affinity plus low-affinity clearance mechanisms. Clearance of modified emulsion represents uptake by low-affinity mechanisms. The difference in the clearance of the two emulsion types indicated by the area in between the two curves represents high-affinity uptake alone. Data are expressed as arithmetic means ± S E M (n = 6).
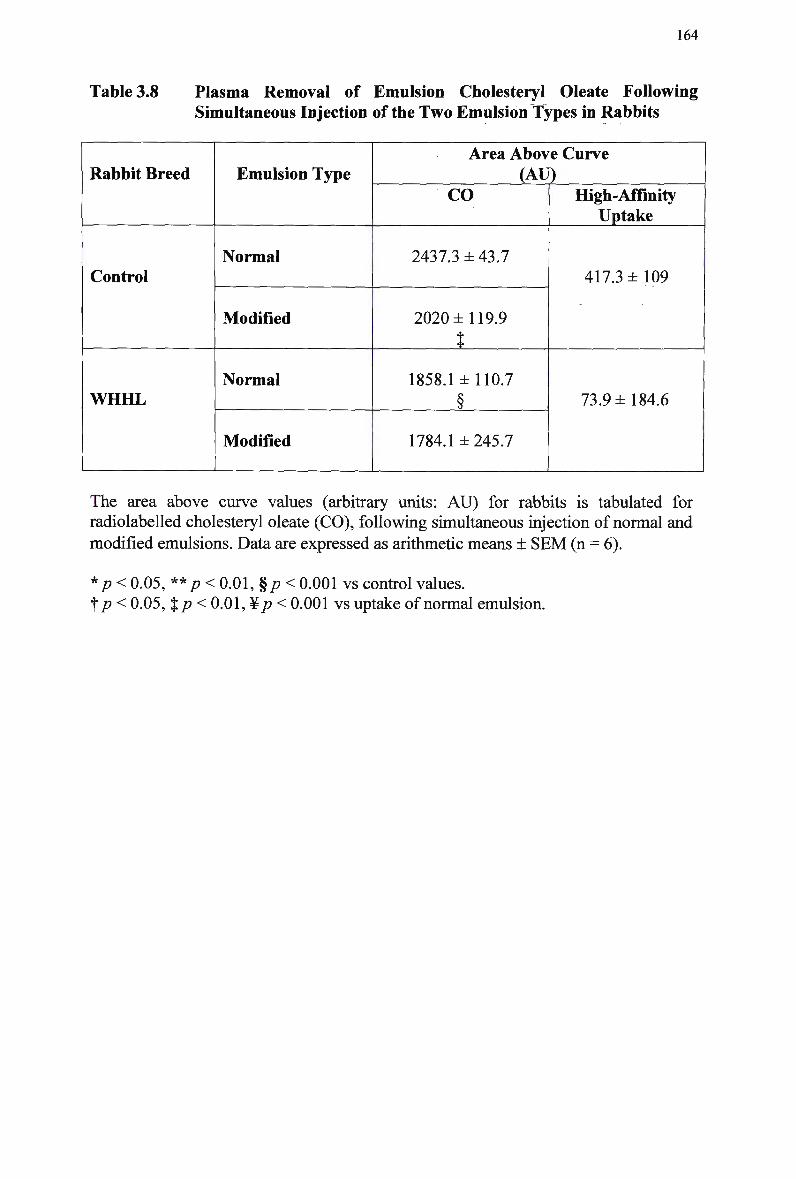
164
Table 3.8 Plasma Removal of Emulsion Cholesteryl Oleate Following Simultaneous Injection of the T w o Emulsion Types in Rabbits
Rabbit Breed
Control
WHHL
Emulsion Type
Normal
Modified
Normal
Modified
Area Above Curve
(AU) CO
2437.3 ± 43.7
2020 ±119.9
t
1858.1 ±110.7
§
1784.1 ±245.7
High-Affinity Uptake
417.3 ±109
73.9 ±184.6
The area above curve values (arbitrary units: A U ) for rabbits is tabulated for radiolabeled cholesteryl oleate (CO), following simultaneous injection of normal and
modified emulsions. Data are expressed as arithmetic means ± S E M (n = 6).
*p < 0.05, **p < 0.01, %p < 0.001 vs control values. f p < 0.05, %p< 0.01, ¥p < 0.001 vs uptake of normal emulsion.

165
3.3.8 Comparison of Emulsion Cholesteryl Ester Uptake
Following Separate versus Simultaneous Injection of the
Two Emulsion Types in Rabbits
The results from 3.3.6 and 3.3.7 for control rabbits are compared (Table 3.9). The
rationale for comparing separate and simultaneous injection was to determine if
simultaneous injection of the two emulsion types would alter the in vivo clearance
kinetics of chylomicron-like emulsion lipids. Figure 3.8 shows the clearance of
emulsion cholesteryl oleate following separate and simultaneous injection of the two
emulsion types.
As with previous clearance studies, the clearance of cholesteryl oleate is
indicative of remnant removal. Under both conditions of injection, normal emulsion
particles were cleared from plasma in a bi-exponential manner. The amount of
modified emulsion cholesteryl oleate cleared in the initial stages (~ 3 min) was always
less compared with normal emulsions (Figure 3.8), resulting in significantly less
modified emulsion cholesteryl oleate cleared over 30 min. The slower phase of
removal of modified emulsions represents a delay in the uptake of remnant lipoprotein
particles.
The amount of emulsion cholesteryl oleate cleared by high affinity pathways
( A A C data) following simultaneous injection was lower compared with separate
injection mode, however this was not considered significant. High affinity
mechanisms accounted for 2 2 % and 1 7 % of the total emulsion uptake for separate and
simultaneous injection in control rabbits, respectively. Consequently, uptake of
emulsion remnants via low affinity pathways accounted for 7 7 % following separate
and 8 3 % following simultaneous injection. The decrease was primarily attributed to
by a reduced proportion of normal emulsion removed via receptor mediated uptake
mechanisms, indicating that normal emulsion remnant removal following
simultaneous injection was facilitated by low affinity pathways to a larger extent.
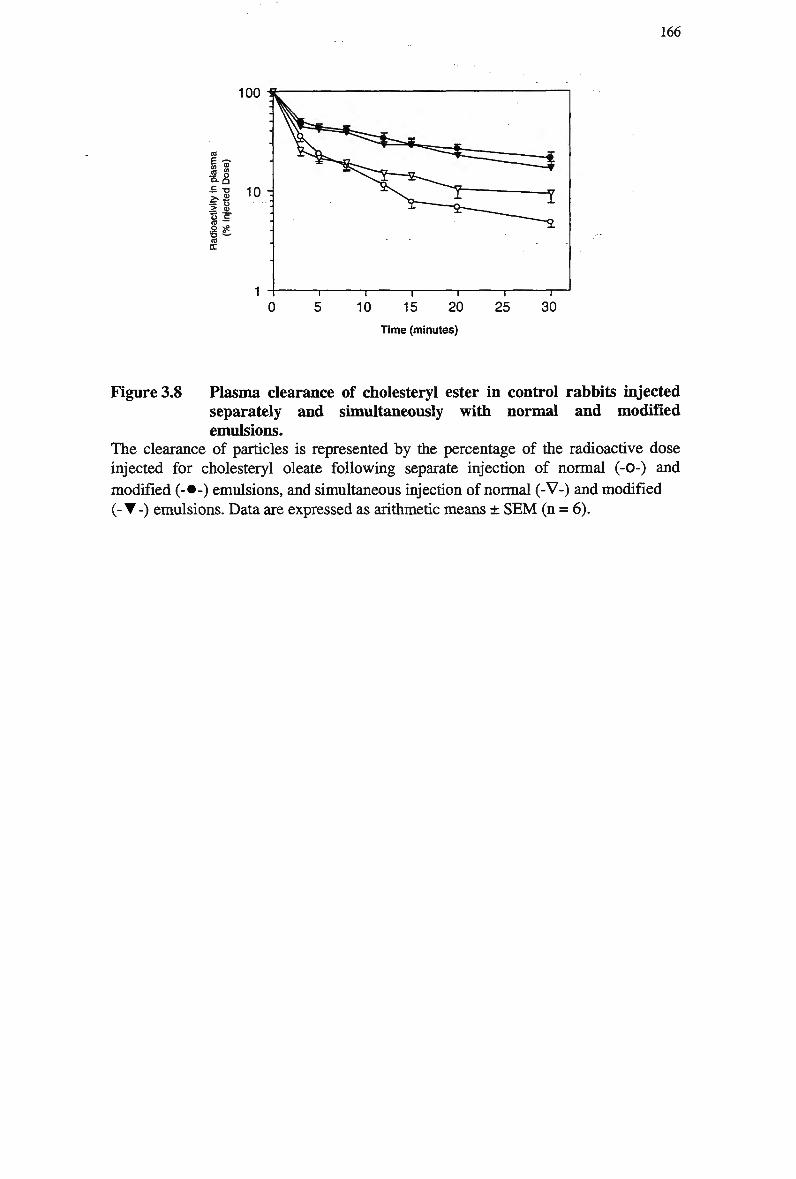
166
100
£ -£ o .2 cu ts _• 10 .9 S?
10 -
20 Time (minutes)
T 25
T 30
Figure 3.8 Plasma clearance of cholesteryl ester in control rabbits injected separately and simultaneously with normal and modified emulsions. The clearance of particles is represented by the percentage of the radioactive dose injected for cholesteryl oleate following separate injection of normal (-0-) and
modified (-•-) emulsions, and simultaneous injection of normal (-V-) and modified (- T -) emulsions. Data are expressed as arithmetic means ± SEM (n = 6).
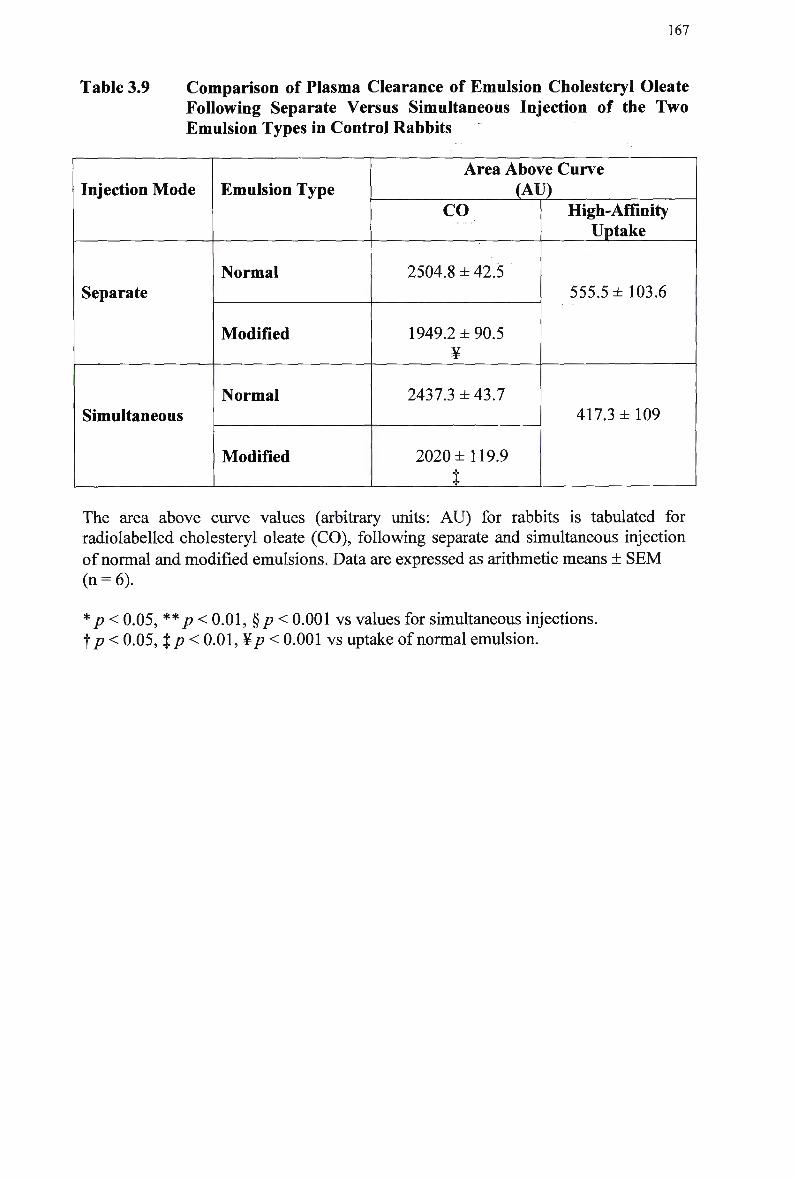
167
Table 3.9 Comparison of Plasma Clearance of Emulsion Cholesteryl Oleate Following Separate Versus Simultaneous Injection of the Two Emulsion Types in Control Rabbits
Injection Mode
Separate
Simultaneous
Emulsion Type
Normal
Modified
Normal
Modified
Area Above Curve (AU)
CO
2504.8 ± 42.5
1949.2 ±90.5 ¥
2437.3 ±43.7
2020 ±119.9
t
High-Affinity Uptake
555.5 ±103.6
417.3 ±109
The area above curve values (arbitrary units: AU) for rabbits is tabulated for radiolabeled cholesteryl oleate (CO), following separate and simultaneous injection
of normal and modified emulsions. Data are expressed as arithmetic means ± SEM (n = 6).
* p< 0.05, ** p<0.01,§p< 0.001 vs values for simultaneous injections. f p < 0.05, %p< 0.01, ¥/? < 0.001 vs uptake of normal emulsion.

168
3.4 Discussion
The aim of this study was to validate the use of normal and modified chylomicron-
like emulsions in measuring high affinity uptake in vivo. T o establish an appropriate
animal model for the two-emulsion method, the clearance studies were repeated in
rats and rabbits. The metabolism of chylomicron-like emulsions in control and
homozygous W H H L rabbits was also investigated, to confirm that chylomicron
clearance is delayed when the LDL-receptor is not present, and to verify that modified
emulsions are taken up via low affinity mechanisms. To investigate the suitability of
using simultaneous injection of the two emulsion types for quantitating receptor
activity in vivo, the clearance of normal and modified chylomicron-like emulsions
was studied following separate and simultaneous injection in rabbits and rats.
Redgrave et al. (Redgrave et al, 1987) established that cholesterol is
necessary for chylomicron-like emulsions to mimic the metabolism of lipoproteins,
including their clearance from plasma. Clearance of normal emulsions is indicative of
high affinity (receptor) plus non-specific uptake (nsu) mechanisms. A second
emulsion particle was synthesised, with the usual component of unesterified
cholesterol omitted. The modified emulsion interacts efficiently with hydrolytic
enzymes in vivo and is thus converted to a remnant at the same rate, however the
particles do not interact with high affinity uptake pathways (receptors) (Redgrave et
al, 1987). The clearance from plasma of the modified emulsion particles is therefore
representative of nsu mechanisms. A measure of high-affinity clearance is determined
as the difference in clearance of normal emulsion (receptor + nsu) and modified
emulsion (nsu) particles.
It should be noted that in the initial experiments utilising for triglyceride-
phospholipid emulsions, the lipid ratio was different from more recent studies
(Martins et al, 1996), (Redgrave et al, 1993) and that used in this set of experiments.
The modified emulsion lipid values (% total) for triglyceride, cholesteryl ester and
phospholipid were 72.4, 9.5 and 18.1, respectively (Redgrave et al, 1987) while the
above studies utilised a ratio of 71.4, 3.1 and 25.5, respectively. In addition, The
normal emulsion lipid values (% total) for triglyceride, cholesteryl ester, cholesterol
and phospholipid were 71.6, 9.5, 1 and 17.9, respectively (Redgrave et al, 1987) and
70, 3, 2 and 25, respectively for the above studies. All values provided are pre-
sonication.

169
Particle size is an important factor in regulating the distribution of
exchangeable lipoproteins (Tajima et al, 1983), and the modified emulsion was found
to be consistently similar in size to the normal emulsion. Hence the absence of
cholesterol alters receptor-binding capacity independent of particle size.
A major objective of this set of experiments was to study the metabolism of
normal and modified chylomicron-like emulsions, to compare the clearance of
chylomicron-like emulsion triglyceride and cholesteryl oleate, and to confirm
previous observations that the absence of unesterified cholesterol in emulsions delays
the clearance of remnant particles. The clearance of normal emulsions fitted the
known mechanism by which chylomicrons are catabolised in rat and rabbit animal
models. That is, there was greater amount of radiolabeled triolein was cleared
compared with radiolabeled cholesteryl oleate, consistent with triglyceride lipolysis
by endothelial lipases and consequent remnant formation within the plasma
compartment and uptake. The triglycerides from modified emulsions in this study
were removed in a similar manner to normal emulsion, indicating that modification of
particles did not impair lipolysis by L P L and remnant formation in rats or rabbits.
In this study, as with previous chylomicron clearance studies the clearance of
cholesteryl oleate should be indicative of remnant removal. For both rat and rabbit
clearance studies, there was a delay in the remnant particle uptake of the modified
emulsion compared with the normal emulsion, suggesting that the modified emulsions
may be taken up via alternate, non-specific mechanisms. High-affinity uptake
mechanisms operate more rapidly than those of non-specific (low-affinity)
mechanisms, and the clearance of the majority of the normal emulsion in the first 5
min supports its' removal via high-affinity (receptor) pathways.
In the rat studies, hepatic uptake of normal emulsion cholesteryl oleate was
greater than that of modified emulsion during separate injection of the two emulsion
types. The data suggests that the modified emulsion could not interact as efficiently
with hepatic receptors (which are comprised mainly of LDL-receptors) and peripheral
tissues were utilised as alternate uptake pathways. The uptake of modified emulsion
remnants via the spleen was greater, compared with normal emulsion. These data
support the clearance data, and indicate that the modified emulsion does not interact
with hepatic receptors, thus facilitating uptake via alternate routes.
Chylomicron remnants and L D L are primarily cleared via the LDL-receptor,
therefore their concentration in plasma is indicative of receptor activity. T o further

170
investigate the validity of these two emulsions as a model for monitoring receptor-
mediated uptake, the clearance of normal and modified chylomicron-like emulsions
was compared in control and homozygous W H H L rabbits. As discussed previously,
W H H L rabbits lack LDL-receptors thus any receptor-mediated chylomicron-like
emulsion removal should indicate the presence of an alternate receptor and decreased
clearance through competition with other lipoproteins. In addition, a lack of receptor-
mediated chylomicron remnant clearance in W H H L rabbits would suggest that the
LDL-receptor deficiency had a direct effect on chylomicron remnant metabolism.
Modified emulsions also bind apo E but are not recognised by LDL-receptors due to
changes in conformation (Redgrave et al, 1987). These modified emulsions were
used to determine the amount of non-receptor mediated (low-affinity) chylomicron
clearance. Receptor-mediated clearance was calculated as the difference in clearance
of the two emulsion types in control and W H H L rabbits.
The clearance of chylomicron-like emulsion cholesteryl oleate fitted the
known mechanism by which chylomicron remnants are catabolised, for normal and
modified emulsions, in control and W H H L rabbits. In W H H L rabbits, the total
amount of normal emulsion cholesteryl oleate cleared was significantly less compared
with the amount cleared in control rabbits, indicating delayed remnant removal. In all
rabbits, modified emulsion cholesteryl oleate was cleared in a linear fashion, which is
indicative of uptake via a low affinity pathway. In keeping with the hypothesis that
chylomicron remnants are cleared in vivo via the LDL-receptor, there was a
significant difference between the total clearance of normal and modified emulsion
cholesteryl oleate in control rabbits, however this difference was not observed for
W H H L rabbits.
These results are consistent with previous findings of defective plasma
clearance (Bowler et al, 1991), (Mamo et al, 1991) and metabolism (Redgrave et al,
1995) of injected chylomicrons and chylomicron-like emulsions in W H H L rabbits. It
is noteworthy that the previous studies assessed the clearance of the two emulsion
types on consecutive days, whereas the present studies injected both emulsions
simultaneously. LDL-receptors are defective in W H H L rabbits, consequently, the
clearance of chylomicron remnants is slow and the concentration of chylomicron
remnants in plasma is increased, potentially leading to the development of
arteriosclerosis. Furthermore, the clearance of remnants of triglyceride-rich

171
lipoproteins is defective in W H H L , reflective of the role of LDL-receptors acting as a
ligand for the apo E associated with remnants (Redgrave et al, 1987).
The difference in clearance of the two emulsion types is indicative of receptor-
mediated (high affinity) uptake, and was essentially absent in W H H L rabbits,
indicating that chylomicron remnant particle uptake in W H H L rabbits occurred via
low affinity mechanisms. High-affinity uptake was greater for control rabbits
compared with W H H L rabbits (417.3 and 73.9, respectively), however this difference
was not considered significant. In the W H H L rabbits the clearance of normal
emulsions was very linear and varied little from the clearance of modified emulsions,
indicating that there was no appreciable receptor-mediated clearance of chylomicrons
in the W H H L rabbits compared with controls. Despite the lack of significance, the
results suggest that W H H L rabbits utilise an alternate, low-affinity pathway with high
capacity compared with normal control rabbits. Regarding the suggestion that the
LDL-receptor is necessary for the normal uptake of chylomicron remnants, the
impaired removal of chylomicron remnants by W H H L rabbits indicated that this
impairment is a direct result of the LDL-receptor deficiency.
The present findings do not support the role of second receptor (high affinity)
mechanism responsible for remnant clearance, as there was no difference in the
uptake of normal and modified emulsions in homozygote W H H L rabbits. M a m o et al.
(Mamo et al, 1991) found that plasma devoid of apolipoproteins with a density of
less than 1.006 gm/ml from W H H L and control rabbits transferred similar amounts of
apolipoproteins, including apo E to emulsions. Thus it would appear that the plasma
clearance of chylomicron-like emulsions in W H H L rabbits is limited by the absence
of LDL-receptors, rather than a reduction in receptor binding ligands.
Despite the finding that normal chylomicron clearance in W H H L rabbits was
reported by Kita et al. (Kita et al, 1982-b), the authors also found reduced high
affinity binding of chylomicron remnants to W H H L rabbit liver membranes using in
vitro studies. This supports the proposed hypothesis that W H H L rabbits metabolise
chylomicron remnants via a non-receptor, low-affinity pathway. A number of other in
vitro studies demonstrate that the LDL-receptor is responsible for the high-affinity
binding of other apo E-containing lipoproteins, including V L D L , L D L and IDL
(Windier et al, 1980a), and chylomicrons (Wade et al, 1986). Internalisation of
chylomicron remnants by cultured human fibroblasts occurs mainly via the LDL-
receptor as determined by the ability of chylomicron remnants to regulate intracellular

172
esterification and expression of LDL-receptors. Floren et al. (Floren et al, 1981) also
demonstrated that chylomicron remnants compete with L D L for uptake and both the
rate of cholesteryl esterification and chylomicron remnant uptake in vivo. Other
researchers support these findings (Choi et al, 1991), (Nagata et al, 1988), (Chen et
al, 1991), (Windier et al, 1988), as discussed in Chapter 1.
Having established that receptor activity can be quantitated in vivo, it was
necessary to determine if the two emulsion types could be injected on consecutive
days (i.e., separately), or simultaneously, by mixing the two emulsion types prior to
injection. The advantages of simultaneous injection for use in humans are two fold.
This approach would provide a time saving measure and patients would only need to
visit a clinic on one occasion. Secondly, this method would reduce the day-to-day
variation in lipid metabolism and provide a measure of receptor activity at a single
point in time. The clearance of emulsion cholesteryl oleate following simultaneous
and separate injection of chylomicron-like emulsions was compared in rat and rabbit
animal models.
The data from clearance studies in rats showed no difference in the
metabolism of normal emulsion cholesteryl oleate between the two modes of
injection. However, the clearance of the modified emulsion was greater following
simultaneous injection of the two emulsion types, with approx. 2 0 % remaining in
plasma at 30 mins compared with 5 5 % for separate injection, suggesting that the
amount of emulsion injected has had an effect on the clearance. Compared with the
separate injection mode, there was an increase in the amount of emulsion cholesteryl
oleate removed via low affinity pathways following simultaneous injection (52% and
7 7 % , respectively). Consequently, the contribution of high affinity mechanisms to the
total clearance of emulsion cholesteryl oleate was decreased (48% and 2 3 % for
separate and simultaneous injection, respectively). In addition, the amount of
modified emulsion in plasma following simultaneous injection continued to decrease
over the 30 min period. This pattern was not apparent for the modified emulsion
following separate injection, suggesting that it did not interact with uptake
mechanisms to any extent after 1-3 min. The present results suggest that low affinity
uptake mechanisms contributed to a larger extent to normal and modified emulsion
particle uptake following simultaneous injection of the two emulsion types.
The differences observed in the clearance of emulsion cholesteryl oleate
following simultaneous injection may be attributed to variation between rats or

173
batches, as all rats were treated similarly prior to experimentation. For separate
injection studies, different groups of rats were used to assess clearance of normal and
modified emulsions, as the rats were sacrificed after each study. In contrast, the same
set of rats was used to assess the clearance of normal and modified emulsions
following simultaneous injection. The triglyceride mass injected was increased from
20 to 3 0 % of the total triglyceride pool for simultaneous injection, therefore was not
expected to alter the clearance kinetics of emulsion particles. Saturation of receptor-
mediated uptake mechanisms is unlikely as the decrease in high affinity uptake values
for the simultaneous injection study was primarily due to an increase in the uptake of
both emulsions by low affinity mechanisms. The clearance kinetics of the modified
emulsion was altered to a greater extent than the normal emulsion, suggesting a
greater capacity of low affinity pathways rather than a decrease in receptor activity.
The two modes of injection were also compared using the rabbit model. The
data indicated that the clearance of normal and modified emulsion cholesteryl oleate
was similar whether the emulsions were injected separately or simultaneously. In
addition, the values for high affinity uptake were similar, and the amount of modified
emulsion cholesteryl oleate cleared was significantly less compared with normal
emulsion cholesteryl oleate, for both injection protocols. This supports the earlier
assertion that no significant saturation of the lipoprotein uptake pathway(s) had
occurred due to the increase in lipid mass accompanying simultaneous injection of the
two emulsion types.
T o determine whether the simultaneous injection approach is suitable for
quantitating high affinity uptake in animal models and humans, it was necessary to
determine if the clearance kinetics had been altered, compared with separate injection.
As described above, the clearance of modified emulsion cholesteryl oleate was
significantly greater, and high affinity uptake of emulsion particles was significantly
less following separate versus simultaneous injection in rats. However, the high
affinity uptake values for simultaneous injection studies in rats were similar to those
obtained for both separate and simultaneous injection studies in rabbits (23%, 1 7 %
and 2 2 % of total uptake, respectively). The results therefore indicate that
simultaneous injection of the two emulsion types is a suitable method for quantitating
receptor uptake in vivo.

174
Chapter 4: Characterisation and Analysis of Chylomicron Remnants
4.1 Introduction
The objective of this study to characterise the size and lipid composition of
endogenous chylomicron remnants, with the aim of synthesising replica remnant-like
emulsion particles, based on the ratio of triglyceride, cholesteryl esters, phospholipid
and unesterified cholesterol from the characterisation data. Remnant-like emulsion
particles should have a diameter consistent with endogenous remnants (approx. 50
nm). Remnant-like emulsions are currently synthesised and utilised in animals and
man, however the data varies considerably and it is not known if the size and lipid
composition of these remnant particles is representative of remnants derived from
nascent chylomicrons.
Isolating chylomicron remnants from postprandial plasma pose some
difficulties because of contamination from lipoproteins (particularly V L D L and
chylomicrons) and low yields ( M a m o et al, 1996). In the past, chylomicron remnants
have been 'made' in vitro by treating human and animal lymph chylomicrons with
lipoprotein-deficient serum (to lipolyse chylomicrons) but this method gave
inconsistent sizes of chylomicron remnants and contamination with excessive free
fatty acids (Floren et al, 1981). Our laboratory has solved these problems by devising
a novel technique to make 'pure' chylomicron remnants ( M a m o et al, 1996). The
method utilised by our laboratory involves injecting donor chylomicrons into rabbits,
where they circulate in plasma for 3 hr (Ma m o et al, 1996). A large dose of

175
triglyceride (about 20 times the normal plasma pool) is injected to dilute residual
V L D L . Chylomicron remnants (density < 1.006 gm/ml) are then isolated from plasma
by ultracentrifugation (Redgrave et al, 1975). The rabbits are functionally
hepatectomised, therefore there is no entry of newly secreted V L D L into plasma, and
any circulating V L D L is converted to IDL and/or L D L (density = 0.019-1.063
gm/ml).
Chylomicrons consist of an oily core that contains mainly cholesteryl ester and
triacylglycerol with some free cholesterol (Zilversmit, 1965) surrounded by a
monolayer of polar lipids and protein. However, cholesteryl esters have been shown
to be located in the surface of the lipoprotein (Janiak et al, 1979), (Bhattacharya and
Redgrave, 1981). In model systems, Hamilton and Small (Hamilton and Small, 1981)
have shown that the surface of chylomicron particles may contain up to 3 %
triacylglycerol. The chylomicron mass in animals and man consists of approx. 75-
8 5 % triacylglycerol, 10-20% phospholipids, 0.5-2% cholesteryl esters, 0.5-2%
unesterified cholesterol, and 2 % protein (Redgrave, 1999). However, the composition
of lipoprotein particles changes with size (Fraser, 1970). Particles with diameters of
20-80 n m predominate in fasting lymph, while after a fatty meal, they range in
diameter from < 50nm to more than 300 n m (Tso et al, 1984), (Hayashi et al, 1990).
The triacylglycerols present in chylomicrons are rapidly hydrolysed by L P L at
the endothelial surface of capillaries in adipose tissue, skeletal muscle, cardiac muscle
and other sites (Havel, 1986b). W h e n lipolysis has removed about 70-90% of
triacylglycerol, residual triglycerides (approx. 3 0 % of the original) and most of the
particle cholesteryl esters remain in the circulation to form a chylomicron remnant
(Redgrave, 1999), (Mjos et al, 1975), (Nestel et al, 1963b). The remnant lipoprotein
is then removed by the liver via a receptor-mediated process, utilising apo E as the
ligand. The two-step model for chylomicron metabolism was directly established by
Redgrave (Redgrave, 1970), who demonstrated the accumulation of cholesteryl ester-
rich particles in the blood of functionally hepatectomised rats, and coined the term
chylomicron remnants.
Less is known about the lipid composition of chylomicron remnant particles.
Jeffrey and Redgrave (Jeffery and Redgrave, 1982) showed that chylomicron
remnants contain approx. 7 0 % triacylglycerol, 6-7% cholesteryl ester and 5-7%
protein. There are immediate changes in the composition of chylomicron
phospholipids and proteins due to exchanges with other lipoproteins in the plasma,

176
with a major fraction of phospholipids and soluble apolipoproteins reported to transfer
to the H D L fraction (Redgrave and Dunne, 1975). Other essential changes in
chylomicrons following lipolysis include loss of apolipoprotein A-I and C, and
acquisition of apo E, hence the proteins present on remnants are apo B-48, E and C
(Redgrave, 1970), (Jeffery and Redgrave, 1982), (Mjos et al, 1975). The diameter of
chylomicron remnants ranges from 50-64 n m (Redgrave, 1999).
Miller and Small (Miller and Small, 1982) developed artificial emulsions that
simulate the size, density, and composition of chylomicrons without apolipoproteins.
Following injection emulsions rapidly incorporate apo C-II to facilitate hydrolysis of
triacylglycerols by L P L in peripheral tissues, and apo E to enable recognition and
uptake of the emulsion remnant particles by the liver. This incorporation occurs by
transfer of apolipoproteins from plasma lipoproteins or by association of
apolipoproteins present in plasma (Maranhao et al, 1986), (Redgrave and Maranhao,
1985), (Martins et al, 2000a). Several other studies have validated the use of
chylomicron-like emulsions in mimicking the metabolism of natural chylomicrons
(Bowler et al, 1991), (Redgrave and Zech, 1987), (Redgrave and Callow, 1990),
(Zerbinatti et al, 1991), (Redgrave and Small, 1979). Furthermore, the metabolism of
these particles has been shown to be similar in rats and man (Redgrave et al, 1993).
Metabolism is characterised by slower plasma removal rates and hepatic recoveries
for cholesteryl ester than triglyceride. Hence the clearance of chylomicron
triglycerides reflects the sum of two processes, lipolysis and particle uptake, while
clearance of chylomicron cholesteryl ester reflects tissue uptake from the plasma of
remnant particles in various states of delipidation (Redgrave, 1999).
Manipulation of cholesteryl ester and cholesterol components of lipid-like
emulsions has furthered the understanding of chylomicron and chylomicron remnant
metabolism. The effect of lipid content on the metabolism of emulsions was first
investigated by Redgrave and Maranhao (Redgrave and Maranhao, 1985), who altered
the ratios of cholesterol: phospholipid and cholesteryl ester: triglyceride. The final
emulsion preparation showed larger proportions of core components than were
present in the starting mixtures. There was consistent discrimination against
phosphatidylcholine for incorporation into the final emulsions, whereas cholesterol
was approx. proportional to its starting concentration. Plasma clearance of all
emulsions was rapid and similar to the behaviour of natural chylomicrons, however as
the cholesteryl ester content increased, so did the uptake of triglyceride and

177
cholesteryl ester uptake into the liver. There was also evidence for regulation by the
size of the emulsion particle and by the content of cholesteryl esters.
Further studies by Maranhao et al. (Maranhao et al, 1986) showed that
emulsions poor in cholesteryl ester but rich in free cholesterol showed remnant-like
behaviour, whereas emulsions rich in cholesteryl ester but poor in free cholesterol
were metabolised like nascent chylomicron particles. The amount of free cholesterol
appeared to regulate metabolism by affecting the binding of apolipoproteins to the
particle surface. These observations are consistent with the metabolic differences
between chylomicrons and remnant particles, suggesting the amount of unesterified
cholesterol exerts a regulatory effect on chylomicron metabolism. High cholesterol
content may facilitate binding to apo E and hence promote emulsion uptake by the
liver, or it may change the packing arrangements of the surface lipids. Miller and
Small (Miller and Small, 1982), (Miller and Small, 1983a) studied the phase
behaviour of lipids, and found that the addition of cholesteryl esters to emulsions
promoted the distribution of more cholesterol to the core, and cholesteryl ester to the
surface.
Remnant-like emulsions have also been developed and are utilised in animals
and humans (Martins and Redgrave, 1998), (Martins et al, 2000b), (Redgrave et al,
2001). Hirata et al. (Hirata et al, 1987) compared the clearances of chylomicron and
remnant-like emulsions, and found small but significant changes in triacylglycerol
removal, but no changes in cholesteryl ester removal. More specifically, Oliveira et
al. (Oliveira et al, 1988) implemented a number of studies to determine if
chylomicron and remnant-like emulsions compete for plasma removal and uptake of
emulsion particles from their tissue receptor sites. The data showed that remnants
derived from natural chylomicrons or chylomicron-like emulsions strongly competed
with the remnant-like emulsion, although the liver trapped remnant-like emulsion
particles faster than the chylomicron-like emulsions.
The use of remnant-like emulsions offers the advantage of short-circuiting the
step of lipolysis and monitoring remnant particle clearance and metabolism directly,
thereby reducing the number of confounding variables. Accumulation of
chylomicrons in blood is ascribed to defects in the delipidation process, in the uptake
of remnant particles, or to variable degrees of combinations of both disturbances
(Nakandakare et al, 1994). Insufficiently delipidated chylomicrons may be poorly
recognised by hepatic receptors for these particles and the consequent delay in plasma

178
clearance m a y be interpreted as a defect in uptake mechanisms. Triglyceride fatty
acids from remnant-like particles are a substrate for lipolysis and can still be
metabolised, albeit to a smaller degree (Martins et al, 2000a). However, when
lipolysis is prevented with Triton W R 1339, remnant-like emulsion particles are not
displaced from their site of removal from the plasma (Oliveira et al, 1988), indicating
that lipolysis does not prevent the uptake of remnant-like emulsions in vivo.
The pattern of clearance of chylomicrons varies considerably between
individuals following a vitamin A fat load, and may make it difficult to ascertain
therapeutic effects on lipid metabolism. Chylomicron size affects clearance from
plasma (Martins et al, 1996) and the fatty acid composition of an oral load affects
chylomicron size in humans (Sakr et al, 1997). Chylomicrons are more
heterogeneous in composition and size that model emulsion particles. The emulsions
are prepared from four pure lipids, while all lipid classes in the chylomicrons are
complex mixtures due to the processes of intestinal absorption and formation in the
mucosal cells. Emulsions offer advantages for the study of specific aspects of
chylomicron metabolism because the chemical composition and particle size can be
controlled and varied. Possible immunological problems due to heterologous
apolipoproteins injected with natural chylomicrons are avoided because of the
absence of apolipoproteins on emulsions (Redgrave et al, 1993).
Monitoring clearance of remnant-like emulsions may offer stability
advantages over chylomicron-like emulsion particles because of their smaller size,
greater density and lower content of triglyceride relative to phospholipid. The
emulsions do not require lipolysis before associating with apo E, which mediates
uptake by the liver. The overall aim of this project was to determine chylomicron
remnant uptake via high-affinity processes, however receptor quantification utilising
chylomicron-like emulsions requires that the particles are first hydrolysed to remnants
by interaction with endothelial lipases. It would therefore be preferable to assess
remnant clearance independent of remnant formation and to determine high affinity
uptake using emulsions that resemble remnants in size and composition. Remnant-like
emulsions are currently used to monitor remnant metabolism, however, there are
numerous variations in the lipid composition, in particular cholesterol/phospholipid
molar ratio. T o facilitate the synthesis of a remnant-like emulsion that would
physiologically mimic chylomicron remnants in vivo, it was essential to establish the
lipid composition of chylomicron remnants derived from nascent chylomicrons.

179
Therefore, it was the aim of this study to characterise chylomicron remnant
lipid composition and size, and to synthesise replica emulsion particles based on this
data. L y m p h was collected from lymph cannulated donors infused with Intralipid via
an intestinal cannula. The chylomicrons were then separated by density gradient
ultracentrifugation and injected into a hepatectomised rabbit for conversion to
chylomicron remnants. The remnant fraction was isolated and characterised and the
data applied for the synthesis of a remnant-like emulsion.

180
4.2 Special Methods
4.2.1 Preparation of Chylomicron Remnants
Preparation and isolation of nascent lymph chylomicrons and chylomicron remnants
is described in Chapter 2.
4.2.2 Extraction of Lipids Using Thin Layer Chromatography
Prior to lipid extraction, chylomicron remnants were assayed for triglyceride and
cholesterol and the size of the chylomicron remnant particles was determined.
Lipids from chylomicron remnant were separated using TLC. Chylomicron
remnants were frozen at -80°C for 4 h, and saponified under vacuum to eliminate all
fluids. Lipids were extracted by a modified method of Folch et al. (Folch et al, 1957).
Each chylomicron remnant sample was suspended in methanol: chloroform (1:2 v/v)
in a glass kimble tube, and mixed gently for 30 min. One quarter of the total volume
of the solvent of 0.03 M HC1 in water was added; the mixture was shaken thoroughly
and allowed to settle. Water was then added to produce a 20-fold excess of solvent.
The tubes were mixed and allowed to stand overnight for adequate separation of the
phases. To complete separation, samples were centrifuged at 1500 rpm for 5 min, at
25°C. The lower phase containing the purified lipid was removed from under the
methanol-water layer with a glass syringe and a long needle. The lipid mixture was
transferred to a glass kimble tube and the solvent was evaporated completely under
nitrogen. The remaining sample was then resuspended in 1 ml chloroform: methanol
mixture (2:1, v/v).
The resuspended lipids were applied in a continuous narrow band to thin layer
plates precoated with 0.2 m m layers of silica gel (Merck). Purified lipid standards
were spotted alongside the sample. The samples were developed for one hr in a glass
tank in the solvent system petroleum ether (40-60°C)/ether/acetic acid (90:10:1, v/v).
The bands of different lipids were visualised using iodine vapour and separated in
sequence. The separated triglyceride, cholesteryl ester, unesterified cholesterol and
phospholipid bands were scraped into glass kimble tubes. Triglyceride and cholesterol
samples were suspended in 5 ml chloroform, and phospholipid samples were
suspended in 5 ml methanol and left for 4-5 hr for lipid extraction. All samples were

181
spun down in a centrifuge at 3000 rpm for 5 min to settle the silica gel before
removing the solvent layer with a pipette. This process was repeated until 25 ml
solvent had been collected.
4.2.3 Lipid Assays
The compositions of chylomicron remnant triglyceride and cholesterol were
determined using commercially available test kits in accordance with the
manufacturer's instructions, as described in Chapter 2. All lipid samples were
analysed within 24 hr of extraction. Prior to assaying, sample sizes were calculated
and aliquoted and the solvent evaporated completely under nitrogen. Triglyceride and
cholesterol samples were sonicated after the addition of respective assay reagents, to
disrupt all sample residues from the assay tubes. The reaction colour was determined
via spectrophotometric analysis at a wavelength of 600 and 500 n m for triglyceride
and cholesterol, respectively, against a reaction blank. The sample content determined
via a standard curve for the assay using the appropriate pure lipid standard.
Phosphorus was measured on chylomicron remnant extracts by a modification of a
method described by Bartlett (Bartlett, 1959).
4.2.4 Determination of Particle Size of Chylomicron Remnants
Lipoprotein particle size was determined to assess the consistency of both
chylomicron and chylomicron remnant size, as described in Chapter 2.
4.2.5 Two Methods of Preparation of Remnant-Like Emulsions
a) Remnant-like emulsions were prepared by sonication and purified by
ultracentrifugation. Triolein (34.6 mg), cholesteryl oleate (26.4 mg), cholesterol (22.4
mg) (Nu Chek Prep, Elysian, M N ) and egg phosphatidylcholine (16.6 mg) (Lipid
Products, Surrey, U K ) , each greater than 99 % pure, were dispensed from stock
solutions into vials. The emulsions were labelled with radioactive
cholesteryl[14C]oleate (Amersham, Surry Hills, N S W , Australia) to allow calculation
of recovery. Solvents were then dried under a stream of nitrogen before overnight
desiccation to eliminate residual solvent traces. The lipids were sonicated in 8.5 ml of

182
0.154 M NaCl/10 m M H E P E S (pH 7.4), at 55-56°C (monitored by a thermocouple in
the vessel) with the atmosphere above the mixture purged with nitrogen to prevent
lipid oxidation. Sonication was for 1 h using a 1cm probe with continuous output of
90-110 W with a Vibra-Cell high intensity ultrasonic processor (Sonics and Materials,
Inc., Danbury, C N ) . The crude emulsion was placed at the bottom of two centrifuge
tubes, then 2.5 ml of NaCl solutions of densities 1.065, 1.040 and 1.020 gm/ml were
sequentially layered above. The tubes were then centrifuged in a S W 4 1 rotor of a
Beckman L8-70M ultracentrifuge for 10 min at 10 000 rpm (12 342 gav) and 22°C.
The large, coarsely emulsified particles were removed from the top of the gradient
and replaced with 1.006 gm/ml solution. This was followed by a second
centrifugation at 30 000 rpm (111 081 gav) for 16 h and 22°C. The emulsion particles
that floated to the surface were removed, then analysed.
b) In this second method, remnant-like emulsions were prepared by sonication
and purified by ultracentrifugation, according to the method described by Hirata et al.
(Hirata et al, 1987). The emulsions were prepared from mixtures of triolein (40.7
mg), egg phosphatidylcholine (24.5 mg), cholesteryl ester (10.3 mg) and cholesterol
(24.5 m g ) and labelled with cholesteryl[14C]oleate. Solvents were then dried under a
stream of nitrogen before overnight desiccation to eliminate residual solvent traces.
The lipid mixture was sonicated in 8 ml of 2.785 M NaCl, density 1.101 gm/ml, at 55-
56°C (monitored by a thermocouple in the vessel) with the atmosphere above the
mixture purged with nitrogen to prevent lipid oxidation. Sonication was for 20 min
using a 1cm probe with continuous output of 70-80 W with a Vibra-Cell high
intensity ultrasonic processor. The crude emulsion was placed at the bottom of two
centrifuge tubes, and then 2.5 ml of NaCl solutions of densities 1.065, 1.040 and
1.020 gm/ml were sequentially layered above. The tubes were then centrifuged in a
S W 4 1 rotor of a Beckman L8-70M ultracentrifuge for 20 min at 10 000 rpm (12 342
gav) and 20°C. The large, coarsely emulsified particles were removed from the top of
the gradient and replaced with 1.006 gm/ml solution. This was followed by a second
centrifugation for 30 min at 38 000 rpm (178 224 gav) and 20°C. The particles that
floated to the surface were removed and used for injection studies.

183
4.3 Results
4.3.1 Lipid Analysis of Chylomicron Remnants
Table 4.1 shows the raw data for the composition of chylomicron remnants derived
from injection of nascent lymph chylomicrons into functionally hepatectomised
rabbits. The remnant particles were relatively small (mean 81.1 ± 12.2 nm ) , indicating
a cholesterol-rich, dense particle. Remnants were composed of approx. 3 6 %
triglyceride, 2 3 % unesterified cholesterol, 2 5 % cholesteryl ester and 1 7 %
phospholipid (Table 4.1). However, the values varied considerably for triglyceride
(28-56%), unesterified cholesterol (13-29.1%), cholesteryl ester (15-36%), and
phospholipid (8-26%).
The ratio of triglyceride: cholesteryl ester was calculated to be 1.63 ± 0.26,
and the cholesteryl ester: triglyceride ratio was 0.75 ± 0.105, for chylomicron
remnants. The ratio of unesterified cholesterol: phospholipid in remnants was 1.48 ±
0.16. The composition of chylomicron remnants as a percent of total lipid mass is
shown in Figure 4.1. W h e n the lipid mass was converted to moles and calculated as a
percent of total lipid mass, there was a redistribution of lipid ratios (Figure 4.2). From
the molar data, triglyceride was lowered from 35.6% to 26.2%, while the cholesterol
content was increased from 22.8% to 37.2%, and cholesteryl ester and phospholipid
ratios remained relatively unchanged from the calculation of lipid weight as a percent
of total (Table 4.1).
Table 4.2 compares the lipid composition of lymph chylomicrons and their
remnants. Remnants derived from injection of lymph chylomicrons into functionally
hepatectomised rabbits differed from their precursors with respect to a 6 0 % decrease
in the triglyceride content, an increase in free and esterified cholesterol by 93-95%,
respectively, and a 4 3 % increase in phospholipid content. O n analysis, the removal of
triglycerides from chylomicrons during lipolysis resulted in a smaller, denser particle,
with approximately half of the particle comprised of cholesteryl ester and cholesterol.
W h e n expressed as a percent of total lipid mass, remnant triglyceride was
significantly correlated with particle size (p = 0.0111). Of all lipids analysed, the
relative quantity of phospholipid was the least altered by the process of lipolysis. The
size and chemical composition of chylomicrons and their remnant products (% total
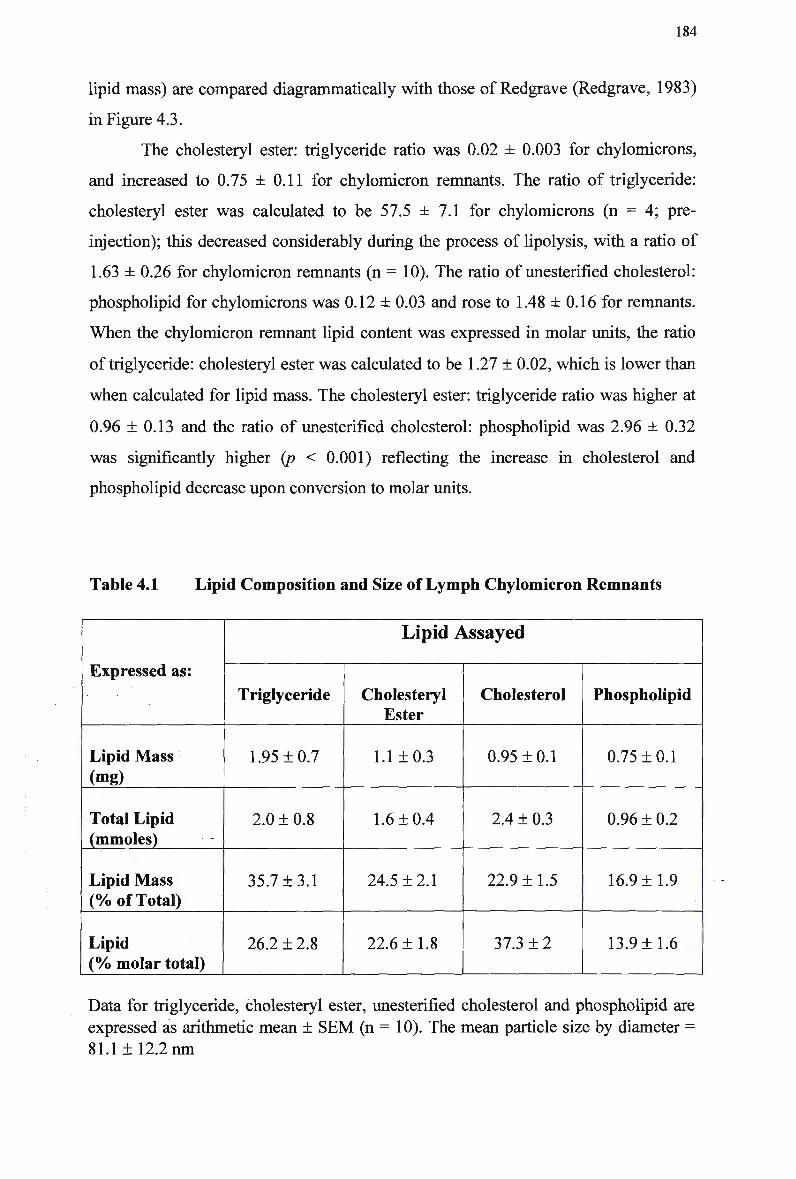
184
lipid mass) are compared diagrammatically with those of Redgrave (Redgrave, 1983)
in Figure 4.3.
The cholesteryl ester: triglyceride ratio was 0.02 ± 0.003 for chylomicrons,
and increased to 0.75 ±0.11 for chylomicron remnants. The ratio of triglyceride:
cholesteryl ester was calculated to be 57.5 ±7.1 for chylomicrons (n = 4; pre-
injection); this decreased considerably during the process of lipolysis, with a ratio of
1.63 ± 0.26 for chylomicron remnants (n = 10). The ratio of unesterified cholesterol:
phospholipid for chylomicrons was 0.12 ± 0.03 and rose to 1.48 ± 0.16 for remnants.
When the chylomicron remnant lipid content was expressed in molar units, the ratio
of triglyceride: cholesteryl ester was calculated to be 1.27 ± 0.02, which is lower than
when calculated for lipid mass. The cholesteryl ester: triglyceride ratio was higher at
0.96 ±0.13 and the ratio of unesterified cholesterol: phospholipid was 2.96 ± 0.32
was significantly higher (p < 0.001) reflecting the increase in cholesterol and
phospholipid decrease upon conversion to molar units.
Table 4.1 Lipid Composition and Size of Lymph Chylomicron Remnants
Expressed as:
Lipid Mass
(mg)
Total Lipid (mmoles)
Lipid Mass (% of Total)
Lipid (% molar total)
Lipid Assayed
Triglyceride
1.95 + 0.7
2.0 ± 0.8
35.7 ±3.1
26.2 ± 2.8
Cholesteryl Ester
1.1 ±0.3
1.6 ±0.4
24.5 ±2.1
22.6 ±1.8
Cholesterol
0.95 ±0.1
2.4 ± 0.3
22.9 ±1.5
37.3 ± 2
Phospholipid
0.75 ±0.1
0.96 ± 0.2
16.9 ±1.9
13.9 ±1.6
Data for triglyceride, cholesteryl ester, unesterified cholesterol and phospholipid are
expressed as arithmetic mean ± S E M (n = 10). The mean particle size by diameter =
81.1 ± 12.2 n m
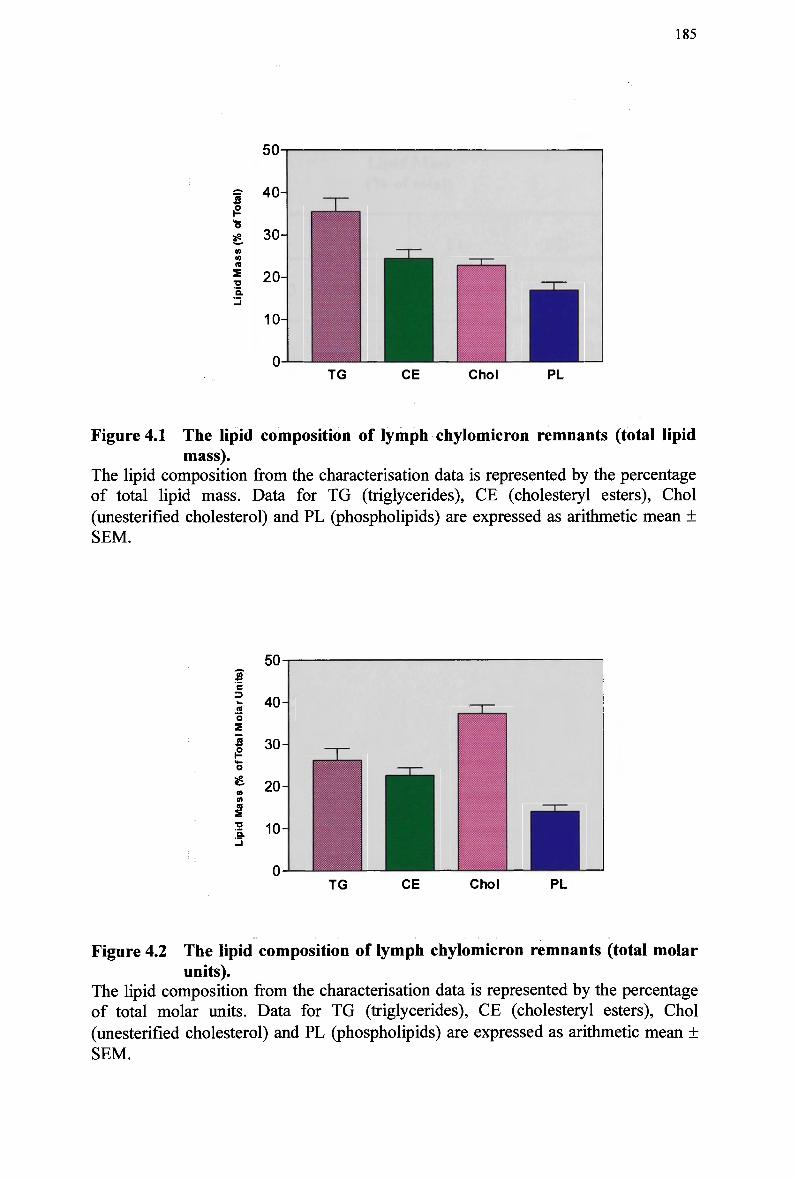
185
s P
CO CO (0
s Q.
Figure 4.1 The lipid composition of lymph chylomicron remnants (total lipid mass).
The lipid composition from the characterisation data is represented by the percentage of total lipid mass. Data for T G (triglycerides), C E (cholesteryl esters), Choi
(unesterified cholesterol) and PL (phospholipids) are expressed as arithmetic mean ± SEM.
c ~ _ 5 S
a & <*> o CO 10 IS
E '5.
Figure 4.2 The lipid composition of lymph chylomicron remnants (total molar
units). The lipid composition from the characterisation data is represented by the percentage of total molar units. Data for T G (triglycerides), C E (cholesteryl esters), Choi
(unesterified cholesterol) and PL (phospholipids) are expressed as arithmetic mean ±
SEM.
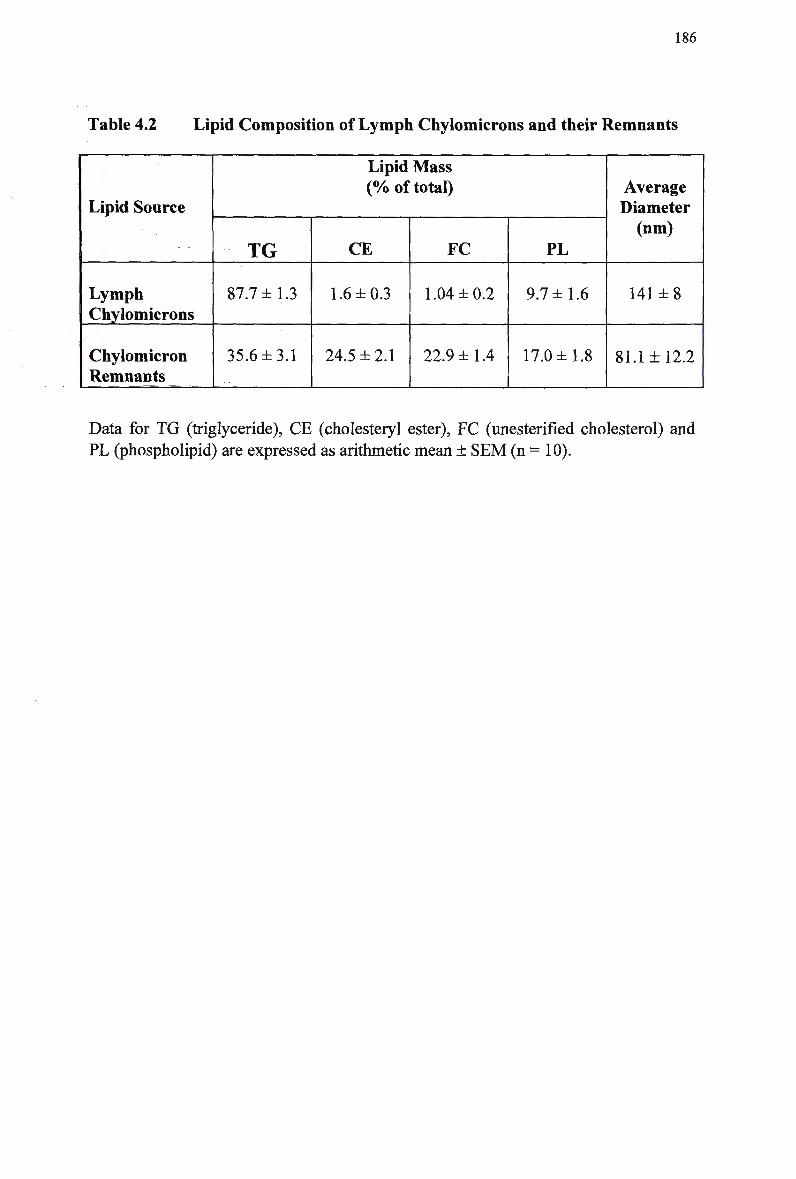
186
Table 4.2 Lipid Composition of Lymph Chylomicrons and their Remnants
Lipid Source
Lymph Chylomicrons
Chylomicron Remnants
Lipid Mass (% of total)
TG
87.7 ±1.3
35.6±3.1
CE
1.6 ±0.3
24.5 ±2.1
FC
1.04 ±0.2
22.9 ±1.4
PL
9.7 ±1.6
17.0 ±1.8
Average Diameter (nm)
141 ±8
81.1 ±12.2
Data for TG (triglyceride), CE (cholesteryl ester), FC (unesterified cholesterol) and
PL (phospholipid) are expressed as arithmetic mean ± SEM (n = 10).
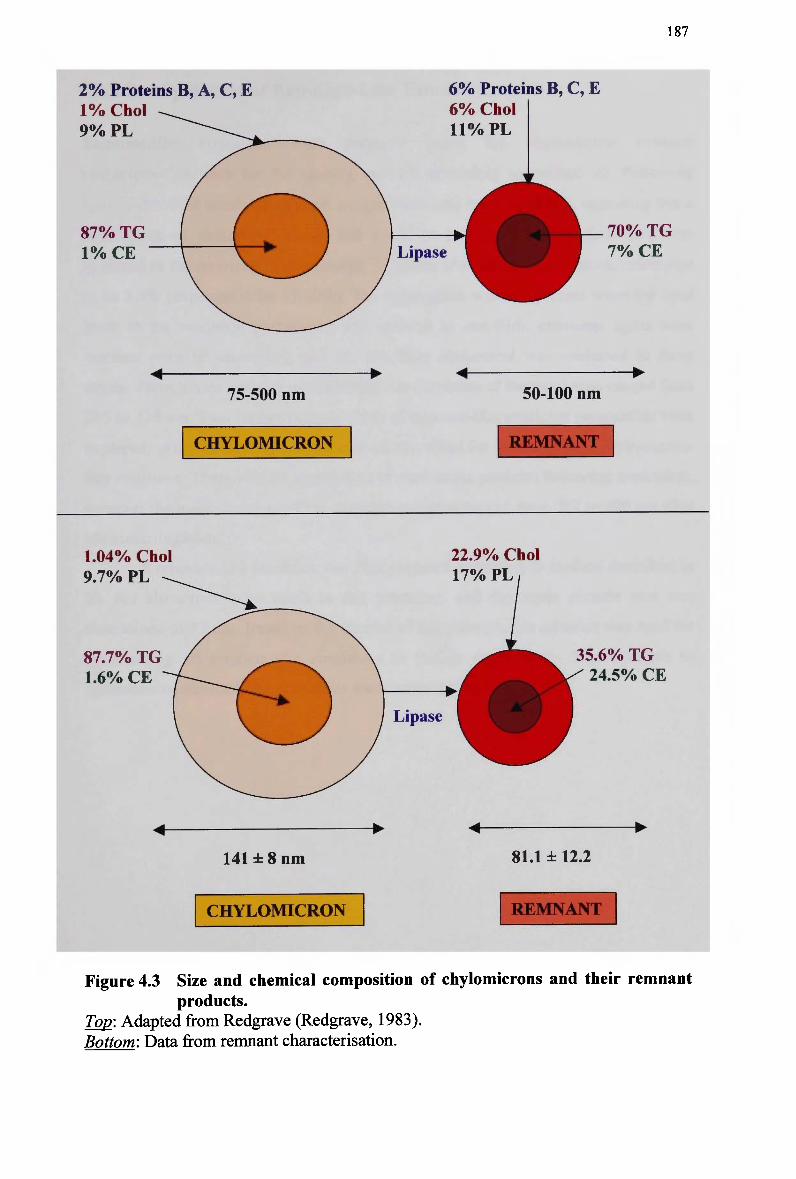
187
2 % Proteins B, A, C, E 1 % Choi 9% PL
87% TG 1%CE
6% Proteins B, C, E 6% Choi 11% PL
Lipase 70% TG 7% CE
75-500 nm 50-100 nm
CHYLOMICRON REMNANT
1.04% Choi 9.7% PL
87.7% TG 1.6% CE
22.9% Choi 17% PL
Lipase
35.6% TG 24.5% CE
141±8nm 81.1 ±12.2
CHYLOMICRON REMNANT
Figure 4.3 Size and chemical composition of chylomicrons and their remnant
products. Top: Adapted from Redgrave (Redgrave, 1983). Bottom: Data from remnant characterisation.

188
4.3.2 Synthesis of Remnant-Like Emulsion
Remnant-like emulsions were prepared using the chylomicron remnant
characterisation data for the starting mixture, according to method a). Following
sonication of the emulsion mixture, a significant lipid mass remained, indicating that a
large mass of cholesteryl oleate had not been sufficiently emulsified. This was
reflected in the recovery of cholesteryl[14C]oleate after ultracentrifugation, calculated
to be 2.4% (expected to be 15-30%). The aggregation was still present when the lipid
mass in the emulsion preparation was reduced to one-fifth, emulsion lipids were
warmed prior to sonication, and the emulsion cholesterol was sonicated in three
stages. Particle size analysis indicated that the diameters of the emulsions ranged from
595 to 733 nm. T w o further permeatations of remnant-like emulsion preparation were
explored, using method b) and the method described for synthesis of a chylomicron-
like emulsion. There was no aggregation of cholesterol particles following sonication,
however the mean diameter of the emulsion particles ranged from 397 to 409 n m after
ultracentrifugation.
A remnant-like emulsion was then prepared according to method described in
b). N o alterations were made to this procedure and the mean particle size was
determined as 77 nm. Based on the success of this attempt, this criterion was used for
synthesising all remnant-like emulsions in further experiments. The attempts to
synthesise of remnant-like emulsions are summarised in Table 4.3.
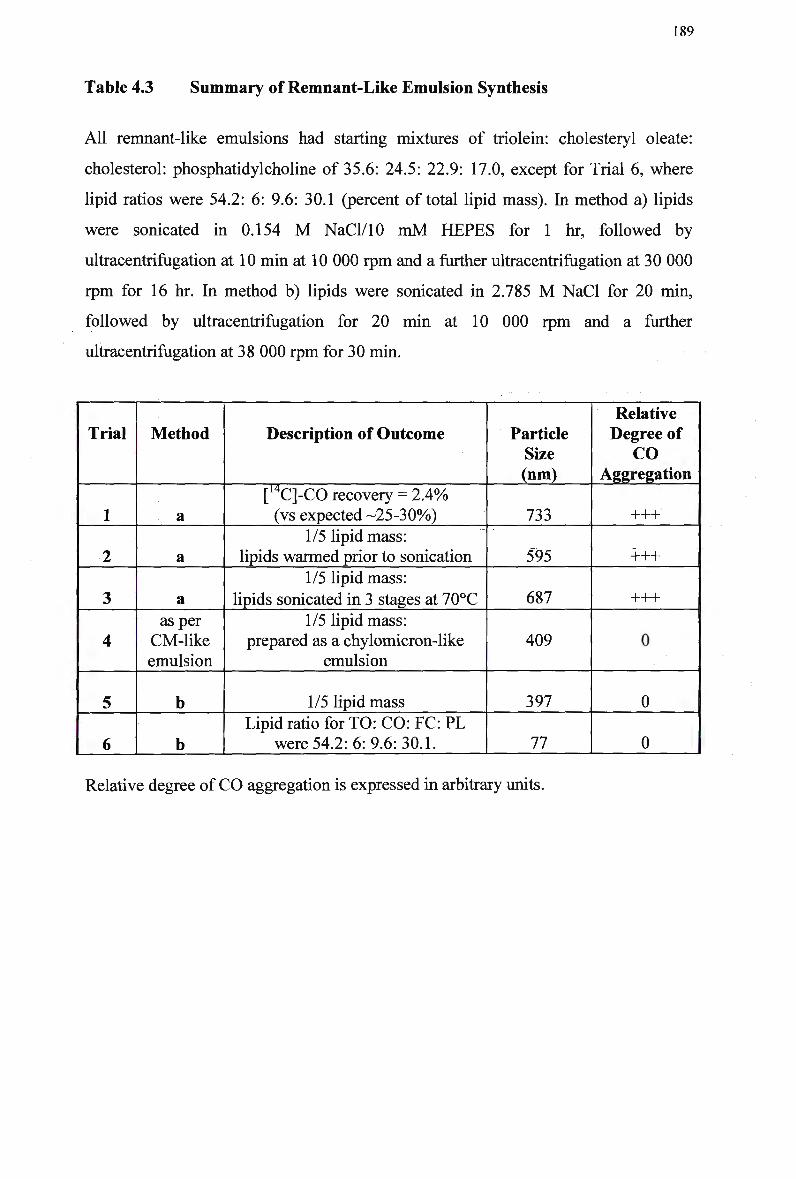
189
Table 4.3 Summary of Remnant-Like Emulsion Synthesis
All remnant-like emulsions had starting mixtures of triolein: cholesteryl oleate:
cholesterol: phosphatidylcholine of 35.6: 24.5: 22.9: 17.0, except for Trial 6, where
lipid ratios were 54.2: 6: 9.6: 30.1 (percent of total lipid mass). In method a) lipids
were sonicated in 0.154 M NaCl/10 m M H E P E S for 1 hr, followed by
ultracentrifugation at 10 min at 10 000 rpm and a further ultracentrifugation at 30 000
rpm for 16 hr. In method b) lipids were sonicated in 2.785 M NaCl for 20 min,
followed by ultracentrifugation for 20 min at 10 000 rpm and a further
ultracentrifugation at 38 000 rpm for 30 min.
Trial
1
2
3
4
5
6
Method
a
a
a as per
CM-like emulsion
b
b
Description of Outcome
[14C]-CO recovery = 2.4% (vs expected -25-30%)
1/5 lipid mass: lipids warmed prior to sonication
1/5 lipid mass:
lipids sonicated in 3 stages at 70°C
1/5 lipid mass: prepared as a chylomicron-like
emulsion
1/5 lipid mass
Lipid ratio for TO: CO: FC: PL were 54.2: 6: 9.6: 30.1.
Particle Size (nm)
733
595
687
409
397
77
Relative Degree of
CO Aggregation
+++
+++
+++
0
0
0
Relative degree of C O aggregation is expressed in arbitrary units.

190
4.4 Discussion
4.4.1 Chylomicron Remnant Composition
The characterisation of remnants in this study provides the first extensive
documentation of the composition of 'pure' chylomicron remnants (i.e., apo B-100-
containing lipoproteins were specifically excluded). The data is specific to
chylomicron remnants derived from hepatectomised rabbits. Previous studies have
utilised various methods of converting chylomicrons to remnants, including
incubation with lipase-rich plasma, purified LPL, and HDL-rich plasma (Redgrave
and Maranhao, 1985). Chylomicron remnants have also been prepared in vitro by
incubating lymph chylomicrons with lipoprotein-deficient serum, however this
method produced chylomicron remnants of varying sizes, and were contaminated with
excessive free fatty acids (Floren et al, 1981).
A technique was developed in our laboratory to make chylomicron remnants,
free from contamination with other lipoproteins (Mamo et al, 1996), (Redgrave et al,
1975), (Mamo and Wheeler, 1994). Lymph chylomicrons are collected from donor
animals and injected as a bolus dose (1.5 g of triglyceride/3 kg B W ) into functionally
hepatectomised rabbits. This protocol saturates the system with apo B-48-containing
lipoproteins, which are hydrolysed over 3 hr. Saturation of the system dilutes the
residual pool of V L D L and enhances the conversion of V L D L to remnants, ensuring
little or no contamination of apo B-100-containing lipoproteins in the isolated
remnant (d < 1.006 gm/ml) fraction. Previous studies have assessed the purity of
chylomicron remnants, based on the absence of apo B-100 containing lipoproteins
following SDS-polyacrylamide gel electrophoresis (Mamo et al, 1996), (Yu et al,
1997), (Proctor and M a m o , 1996), (Yu and M a m o , 1997a). The apolipoprotein
profiles showed significant quantities of apo E (-52%) and lesser quantities of apo C
(-18%), B-48 (-16%), and A-l (-7%), 3), and a constant diameter of 40 to 50 n m
compared to greater than 200 n m for nascent lymph chylomicrons as determined by
laser light scattering.
The study confirmed that triglycerides, the major core component of lymph
chylomicrons, are removed from the blood of functionally hepatectomised rabbits,
whereas the core component, cholesteryl ester, remains. O n average the triglyceride
content of chylomicron remnants had been depleted by approximately 6 0 % of the

191
initial chylomicron injected (Table 4.4). As a result of hydrolysis by lipases, the
chylomicron becomes smaller and denser but retains essentially all of the cholesteryl
esters, thereby raising the relative contribution of cholesteryl ester and cholesterol to
the total lipid mass, with a small increase in phospholipid. This trend has previously
been shown in rats (Redgrave, 1970), (Bezman-Tarcher et al, 1965) and dogs (Nestel
etal, 1983), (Bergman etal, 1971).
The extent of triglyceride depletion relative to cholesteryl esters and the
efficiency of lipolysis have been shown to vary considerably for chylomicrons.
Redgrave (Redgrave, 1970) calculated that that 9 0 % of triglyceride was removed
from chylomicrons injected into hepatectomised rats, to form a remnant particle.
Using the same method of conversion, a number of studies (Mjos et al, 1975),
(Redgrave and Callow, 1990) have demonstrated a decrease in the mass of
chylomicron-like emulsion triglyceride during their conversion to remnants (78.9% to
71.7%, respectively), when expressed as a percent of total lipid mass. In contrast,
Redgrave and Maranhao (Redgrave and Maranhao, 1985) reported an increase in
triglyceride content, from 6 3 % to 6 6 % of the total lipid mass, following incubation of
chylomicron-like emulsion in lipase-rich rat plasma. However, all studies report the
formation of particles enriched in cholesterol and depleted of triglycerides.
The triglyceride content varied between samples, despite injection of the same
lipid mass into each rabbit. This variation was attributed to the extent of chylomicron
hydrolysis, based on circulation time in hepatectomised rabbits (2-3 hours) and
possibly the effect of the anaesthetic. Given that the conditions of chylomicron
remnant preparation were similar for all trials, it is realistic to assume that there is
some variability in the animal species used. Other findings also show considerable
variation, depending on the method of synthesis, animal species, lipid composition,
and the method utilised. The extent of hydrolysis increases with circulation time and
is dependent on the amount and composition of lipids injected. Furthermore, the
particles were prepared in functionally hepatectomised animals therefore the removal
of triglyceride might have been more extensive than in the intact animal where
partially degraded particles would be continuously removed by the liver. These
factors support the variation in the size and composition of chylomicron remnants in
this study, despite the large number of trials and the attention paid to the preparation
and isolation of remnants.

192
The present data show that the ratio of triglyceride: cholesteryl ester decreased
considerably during the process of lipolysis, from approx. 54 for lymph chylomicrons
to 1.63 for chylomicron remnants. Previous studies have shown a reduction in the
ratio of triglyceride to cholesterol from approximately 20 to less than 3 during the
conversion of lymph chylomicrons to chylomicron remnants ( M a m o et al, 1996), (Yu
et al, 1997), (Yu and M a m o , 1997a). The triglyceride: cholesteryl ester ratios as
calculated for lipid compositions from other sources were much larger compared with
the characterisation data (Figure 4.4). The ratios ranged from 15.9 for chylomicron
remnants (Zeng et al, 1998) to 2.2 for remnant-like emulsion (Hirata et al, 1987).
The cholesteryl ester: triglyceride ratio increased (0.75 ± 0.110) during the conversion
of chylomicrons to remnants, which is comparable with findings reported by Oliveira
et al (Oliveira et al, 1988).
During the degradation of chylomicrons by LPL, marked changes in surface
chemistry occur. Remnants are relatively enriched in cholesteryl esters when
compared with the nascent chylomicron and ratio of cholesterol to phospholipid
contained in the surface area is increased (Quinn et al, 1982). The ratio of
unesterified cholesterol: phospholipid for remnants and chylomicrons was 1.48 and
0.116, respectively, for the present characterisation data. The ratios of unesterified
cholesterol to phospholipid reported by other studies ranged from 0.06 for lymph
chylomicron remnants (Ly et al, 1992) to 1.13 for remnant-like emulsion (Hirata et
al, 1987). Previous studies have also shown a molar ratio of unesterified cholesterol:
phospholipid in remnants of approx. 1.0 (Redgrave, 1983), (Jeffery and Redgrave,
1982), (Redgrave and Small, 1979), compared with the molar ratio of 2.96 ± 0.32
(unesterified cholesterol: phospholipid) in the present study.
W h e n related to particle size, the composition of remnants, like that of their
precursors, has been found to be consistent with the "pseudomicellar" model of
lipoproteins, in which a core of nonpolar lipids is covered by a monolayer of polar
lipids and protein (Mjos et al, 1975). Hence the loss of triglycerides from the core of
the particles is accompanied by removal of polar components (phospholipid and
unesterified cholesterol) from the surface and transfer of unesterified cholesterol to
the H D L fraction of plasma (Quinn et al, 1982). Chylomicrons decrease in size from
approximately 500nm to 50nm and increase in density due to the loss of the loosely
packaged core lipids (Redgrave, 1988). The data show that the changes in lipid
composition were accompanied by a reduction in mean diameter from 141 n m for

193
chylomicrons to 81 n m for remnants. Other studies report smaller diameters for
chylomicron remnants of 40-47 n m ( M a m o et al, 1996), (Yu et al, 1997). The
particle size showed a highly significant positive correlation with the triglyceride
content, when expressed as a percent of total lipid mass (r = 0.7909; p = 0.0111),
suggesting that the extent of hydrolysis of the remnant particle determined its size.
Previous data on chylomicron remnant characterisation has been inconsistent,
however whether this variation was a result of the different methods employed to
convert chylomicrons to remnants, or whether they were in fact a population of
heterogeneous particles, is not clear. Our method of chylomicron remnant isolation
and preparation has enabled the characterisation of a pure remnant particle. The
characterisation data suggests that size and composition of chylomicron remnant is
varied, which supports the assertion that chylomicron remnants are heterogeneous in
nature. This has not previously been reported, and therefore this data sheds new light
on the inherent properties of remnant particles. Several factors influence the
heterogeneity of chylomicron remnants, including lipases, C E T P , H D L and
postprandial triglyceride-rich lipoproteins. This is reflected in the range of values in
remnant lipids, the extent of triglyceride hydrolysis and particle size.
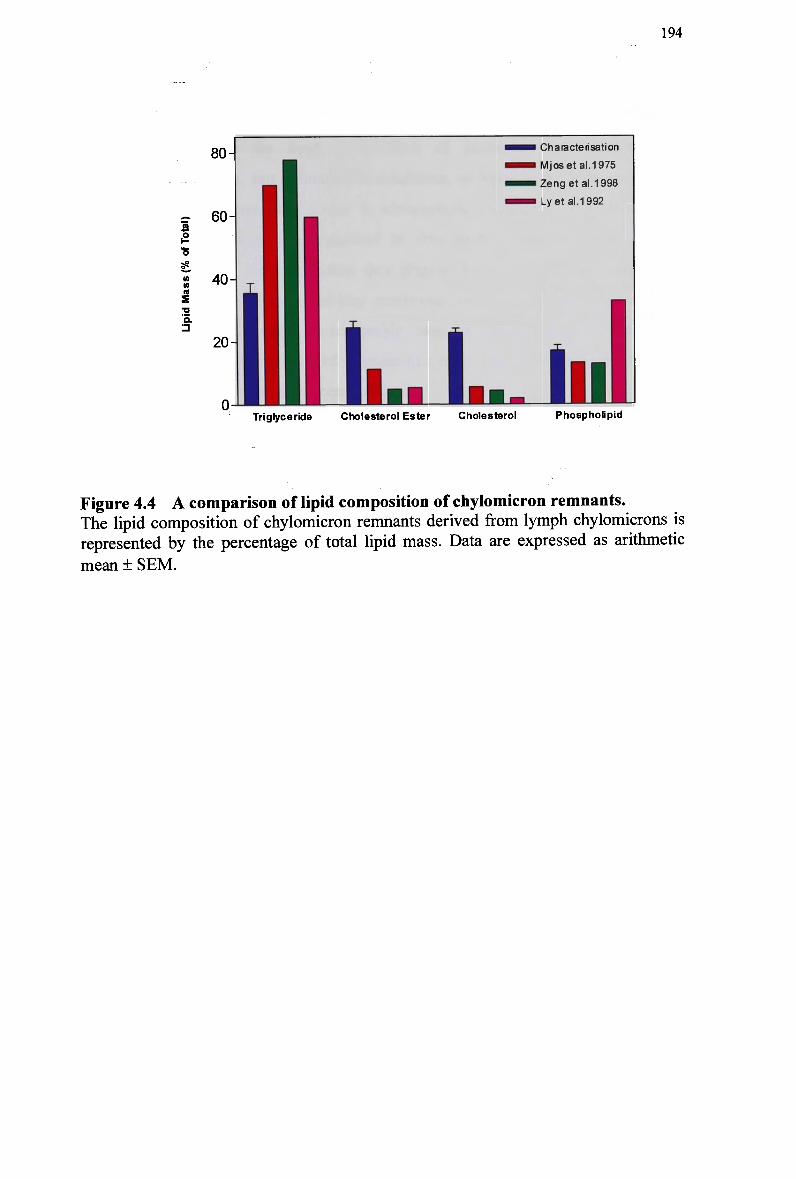
194
80-
_• 60-
I 40-CO
20-
0-
•••• Characterisation
mmmm Mjos et al.1 975
^^m Lyet al.1 992
Triglyceride Cholesterol Ester Cholesterol Phospholipid
Figure 4.4 A comparison of lipid composition of chylomicron remnants. The lipid composition of chylomicron remnants derived from lymph chylomicrons is represented by the percentage of total lipid mass. Data are expressed as arithmetic
mean + SEM.

195
4.4.2 Comparison and Synthesis of Remnant-Like Emulsions
Table 4.4 compares the lipid composition of nascent lymph chylomicrons,
chylomicron remnants, and remnant-like emulsions, as reported by different sources.
Remnant-like emulsions were closer in composition and size to the chylomicron
remnant characterisation data presented in this study, compared with previous
chylomicron remnant characterisation data (Figure 4.4). The starting mixtures and
final composition of the remnant-like emulsions vary considerably, however the
values for all lipids fall within an acceptable range for chylomicron remnant particle
composition. Final compositions of remnant-like emulsions differed quite markedly,
despite starting mixtures of same composition.
The data suggests that chylomicrons and their remnants are more
heterogeneous in composition and size than model emulsion particles. Emulsions are
prepared from four pure lipids, while lymph chylomicrons contain a complex mixture
of lipids. Chylomicron remnants also vary in size and composition according to the
method of preparation. The chemical composition and particle size of emulsions can
be controlled and varied for studying specific aspects of chylomicron metabolism, and
offer the advantage of stability. Initial attempts to synthesise a remnant-like emulsion
based on the characterisation data were inconclusive. The size of the remnant particles
was attributed primarily to the large mass of free and esterified cholesterol, which
could not be solubilised during the sonication process or incorporated into the
emulsion particles. Compared with previous starting lipid ratios for remnant-like
emulsions, the ratios of cholesteryl ester: triglyceride and cholesterol: phospholipid in
the present study are considerably higher.
The composition of chylomicron and remnant-like emulsions has been
manipulated (Table 4.4) to synthesise emulsion particles of a required size and
composition. This is evidenced by the success of the final remnant-like emulsion
preparation in this study, using the method of Hirata et al. (Hirata et al, 1987) which
contained lower and higher proportions of cholesteryl ester and phospholipid,
respectively, in the starting mixture compared with the proportions of these lipids in
the final remnant-like emulsion preparation. Further studies are required to explore
and refine the preparation of remnant-like emulsions, and m a y need to consider
relative recovery rates of individual lipids, in order to arrive at a composition similar
to nascent chylomicron remnants.
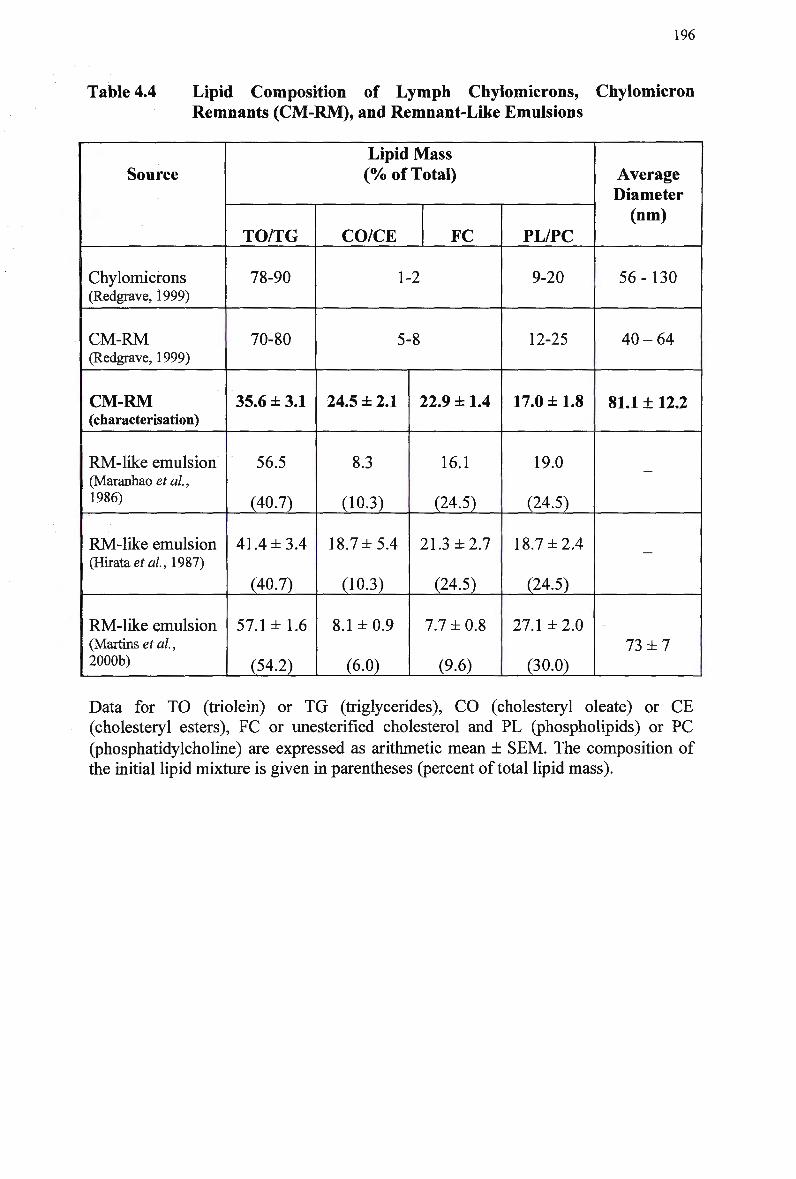
196
Table 4.4 Lipid Composition of Lymph Chylomicrons, Chylomicron Remnants (CM-RM), and Remnant-Like Emulsions
Source
Chylomicrons (Redgrave, 1999)
CM-RM (Redgrave, 1999)
CM-RM (characterisation)
RM-like emulsion (Maranhao et al., 1986)
RM-like emulsion (Hirata etal, 1987)
RM-like emulsion (Martins etal, 2000b)
Lipid Mass (% of Total)
TO/TG
78-90
70-80
35.6 ± 3.1
56.5
(40.7)
41.4 ±3.4
(40.7)
57.1 ±1.6
(54.2)
CO/CE FC
1-2
5-8
24.5 ± 2.1
8.3
(10.3)
18.7 ±5.4
(10.3)
8.1 ±0.9
(6.0)
22.9 ± 1.4
16.1
(24.5)
21.3 ±2.7
(24.5)
7.7 ±0.8
(9.6)
PL/PC
9-20
12-25
17.0 ± 1.8
19.0
(24.5)
18.7 ±2.4
(24.5)
27.1 ±2.0
(30.0)
Average Diameter (nm)
56-130
40-64
81.1 ± 12.2
-
—
73 ± 7
Data for T O (triolein) or T G (triglycerides), C O (cholesteryl oleate) or CE (cholesteryl esters), FC or unesterified cholesterol and PL (phospholipids) or PC
(phosphatidylcholine) are expressed as arithmetic mean ± SEM. The composition of the initial lipid mixture is given in parentheses (percent of total lipid mass).

197
Chapter 5: Hepatic Uptake of Chylomicron and Remnant-Like Emulsions in Mice
5.1 Introduction
Following lipolysis, chylomicron remnants are rapidly cleared from circulation. The
liver removes the large majority of chylomicron remnants where the large pores of the
hepatic sinusoids permit their passage (Redgrave, 1970), (Bergman et al, 1971). The
parenchymal cells of the liver account for the majority of remnant uptake (Jones et al,
1984), (Lippiello et al, 1981), (Floren and Nilsson, 1977b) and degradation of the
cholesteryl ester portion (Stein et al, 1969), (Nilsson and Zilversmit, 1971). This
second phase of rapid hepatic clearance is thought to involve several different steps.
The initial processes involve sequestration of chylomicron remnants into the space of
Disse, utilising apo E on the lipoprotein particles (Havel, 1995), (Windier et al,
1980a), (Windier et al, 1996), (Arbeeny and Rifici, 1984). Subsequent steps appear to
involve H S P G on the cell surface (Ji et al, 1994a), (Ji et al, 1995), where further
lipolysis of the remnants by H L (Shafi et al, 1994), (Sultan et al, 1990) and
acquisition of apo E may recur (Hamilton et al, 1990), (Shimano et al, 1994). The
final step of chylomicron remnant clearance involves the cellular internalisation of the
lipoproteins, mainly by receptor-mediated endocytosis.
Apo E appears to serve as a ligand for receptor-mediated recognition and
uptake of chylomicron remnants from plasma (Wilson et al, 1991), (Windier et al,
1988), (Plump et al, 1992), (Cooper et al, 1982a), (Mortimer et al, 1995a), (Havel,
1998). The receptor primarily responsible for chylomicron removal is considered to

198
be the LDL-receptor, accounting for approx. 8 0 % of total remnant removal from the
plasma (Bowler et al, 1991), (Brown and Goldstein, 1983). In mammals, including
the mouse, the LDL-receptor has high affinity for apo E (Schneider, 1989), (Mamo,
1995), (Floren et al, 1981), (Floren and Chait, 1981), which binds to distinct pockets
within the general binding domain of the LDL-receptor (Brown et al, 1991), (Francke
era/., 1984).
Uptake mechanisms other than the LDL-receptor, which may be involved in
the plasma removal of chylomicron remnants, include; the scavenger cell pathway,
fluid endocytosis, the L R P (Herz et al, 1988), (Willnow et al, 1994) and the
lipolysis-stimulated receptor (Yen et al, 1994). H S P G have also been proposed to
contribute to the process of remnant uptake (Zeng et al, 1998), (Ji et al, 1993), (Ji
and Mahley, 1994), (Ji et al, 1995). Such uptake mechanisms are suggested to be
low-affinity. The consensus appears to be that under normal circumstances the LDL-
receptor is the primary mechanism of uptake for chylomicron remnants, and that in
the absence of the LDL-receptor the L R P may become a more important uptake
pathway (Cooper, 1997), (Mortimer et al, 1995a), (Ishibashi et al, 1994b), (Mahley
et al, 1994). Apo E binding characteristics for chylomicron remnants have been
found to be similar for both receptors (Martins et al, 2000b), (Yu et al, 2000).
As discussed in Chapter 1, knockout animals provide unique models and have
been widely used to demonstrate the contribution of specific receptors and ligands to
the metabolism of chylomicron remnants. In the present study, mice deficient in apo E
or the LDL-receptor were used to confirm that apo E is an essential ligand for the
normal, rapid catabolism of chylomicron remnants by high affinity pathways, i.e.,
hepatic LDL-receptors. Emulsions do not contain apolipoproteins, therefore receptor-
mediated uptake of the remnant particles derived from emulsions relies on their
association in plasma with apo E. In addition, LDL-receptor-deficient mice were used
to establish than normal emulsions were taken up via receptor pathways, and that
modified emulsions do not interact with high affinity mechanisms. The establishment
of apo E as a ligand for uptake via the LDL-receptor will assist in validating the two-
emulsion technique as a means of quantifying high affinity uptake of chylomicron
remnants in vivo.
The data from Chapter 3 showed that normal emulsion particles were rapidly
removed from plasma following injection into recipient animals. W h e n modified
emulsions were injected, emulsion clearance curve was delayed compared with the

199
normal emulsion plasma kinetics. Thus it has been confirmed that chylomicron-like
emulsions can mimic chylomicron metabolism in vivo, and that the amount of
emulsion removed from plasma for the two emulsion types is significantly different.
However, this data does not verify whether the normal and modified emulsions were
cleared via high or low affinity uptake pathways, or both. To provide the basis for the
two-emulsion method to quantify receptor uptake, it was therefore necessary to
unequivocally demonstrate that the plasma kinetics observed in vivo was reflected at a
cellular level. It was also essential to establish that modified emulsions did not
interact with high-affinity (receptor) pathways.
To observe the hepatic clearance of normal and modified emulsion remnant
particles, a series of uptake experiments were undertaken in control and knockout
mice, using emulsions labelled with a fluorescent cholesteryl ester probe. The
emulsions were injected and liver sections from mice homozygous for deficiency in
either apo E or LDL-receptors processed and assayed by laser confocal microscopy.
Micrographs were compared with those from control mice. This technique has been
previously used to successfully monitor the hepatic uptake of emulsion lipids
(Mortimer et al, 1995a), (Zeng et al, 1998), (Martins et al, 2000b).
It was necessary to eliminate the possibility that the simultaneous injection of
normal and modified emulsions would interfere with the kinetics of particle uptake of
each emulsion type. A second fluorescent probe for cholesteryl ester was synthesised
to allow both emulsion types to be fluorescently labelled for simultaneous detection
by confocal laser microscopy, however this marker proved to be unsuitable.
Therefore, control (C57BL/6J) mice were injected with a fluorescently labelled
normal remnant-like emulsion and liver sections were compared with the
simultaneous injection of fluorescently labelled normal remnant-like emulsion and
unlabeled modified remnant-like emulsion. The inverse procedure was repeated using
a fluorescently labelled modified remnant-like emulsion. Remnant-like emulsions
were used as it allowed for lower doses of triglyceride to be injected into recipient
mice, and reduced the chance of saturation of clearance kinetics in vivo.

200
5.2 Special Methods
5.2.1 Animals
Colonies of apo E knockout mice and LDL receptor knockout mice where the genes
for apo E or LDL-receptor were nullified by homologous recombination were
established from progenitor stocks obtained from the Jackson Laboratories (Bar
Harbor, M E ) . The mice were bred by sibling matings to obtain animals homozygous
for the null mutation. C57BL/6J and 129/SV (hybrid) mice were obtained from the
Animal Resources Centre (Murdoch, Western Australia) and were used as controls for
LDL-receptor and apo E-deficient mice, respectively. Male mice ranging in age from
8 to 12 weeks were used for this study and weighed between 18-22 gm. Animals were
fed a pelleted diet containing approx. 5 % fat.
5.2.2 Materials
Egg yolk phosphatidylcholine was purchased from Lipid Products (Surrey, UK).
Cholesterol, cholesteryl oleate, and triolein were from Nu-Chek-Prep. (Elysian, M N ) .
The fluorescent probe cholesteryl-4,4-difluoro-5,7-dimethyl-4-bora-3a,4a-diaza-5-
indacene-3-dodecanoate (cholesteryl B O D I P Y ® FL C u) was purchased from
Molecular Probes, Inc. (Eugene, OR).
5.2.3 Chylomicron-Like Emulsion Preparation
Chylomicron-like emulsions were prepared by sonication and purified by
ultracentrifugation, as described previously (Zeng et al, 1998). Triolein (35 mg),
cholesteryl oleate (1.4 mg), cholesterol (1 mg) and egg phosphatidylcholine (12.5
mg), each greater than 9 9 % pure were dispensed from stock solutions into vials. This
was followed by 0.2 m g BODIPY-CE (substituting 0.4% cholesteryl oleate content).
The fluorescent probe was dissolved in ethanol and added to the lipid at a probe: lipid
ratio of 1: 200. Emulsions with and without the fluorescent ester were radiolabeled
with [14C]triolein and cholesteryl[3H]oleate to trace the lipolysis and remnant removal
of the emulsions, respectively. For modified emulsions, unesterified cholesterol was
omitted from the original lipid mixture to form particles with modified clearance

201
characteristics. Care was taken to keep emulsion preparation away from light, due to
the photosensitivity of the fluorescent label. Preparation of chylomicron-like
emulsions was described in Chapter 2. The emulsion particles that floated to the
surface were removed and made up to a volume of 3 ml with 0.15 M NaCl solution.
Emulsions were then analysed and injected into mice within 1 day.
5.2.4 Remnant-Like Emulsion Preparation
Remnant-like emulsions were prepared by sonication and purified by
ultracentrifugation, as previously described (Martins et al, 1998), (Martins et al,
2000b). This is method currently used to synthesise remnant-like emulsions for use in
animals. The emulsions were prepared from mixtures of triolein (4.5 mg), egg
phosphatidylcholine (2.5 mg), cholesteryl ester (0.5 mg) and cholesterol (0.8 mg) and
labelled with cholesteryl[3H]oleate to trace emulsion remnant removal. For
preparation of the fluorescently labelled particles, 0.1 m g of a fluorescent probe
(BODIPY-CE) was added. For modified emulsions, unesterified cholesterol was
omitted from the original lipid mixture. After a 1 hr sonication of the lipid mixture in
8.5 ml of 2.2% glycerol in water, the crude emulsion was made up to 8.5 ml with
D D W , then adjusted to 1.1 gm/ml with KBr. The emulsion was then placed at the
bottom of two centrifuge tubes, and then 2.5 ml of NaCl solutions of densities 1.065,
1.040 and 1.020 gm/ml were sequentially layered above. The tubes were then
centrifuged in a S W 4 1 rotor of a Beckman L8-70M ultracentrifuge for 60 min at 30
000 rpm (111 071 gav) and 20°C. The particles that floated to the surface were
removed and used for injection studies.
5.2.5 Operative Procedures
Anaesthesia was induced in the mice by intraperitoneal injection of tribromoethanol
(Avertin; Aldrich; 0.3-0.4 gm/kg B W ) . Exactly 50 pi emulsion, containing approx.
250-300 pg of total lipid, was injected via a 30 G needle (Sofijet, Mazamet, France)
into the tail vein as a bolus dose, for separate injection studies. This mass was
equivalent to 130-140 pg triglyceride (14-16% of the recipient animals' total plasma
triglyceride pool). For simultaneous injection of normal and modified remnant-like
emulsions, the emulsions were mixed immediately prior to injection, and a total of

202
270 pg emulsion triglyceride in a volume of 100 pi was injected into the tail vein
(equivalent to 28-33% of the recipient animals' total plasma pool). Plasma volume
was calculated as (5.09 ml/100 g m B W ) for mice. At 5 min, 20 min, and 2 hr after
injection of the emulsion, mice were anaesthetised with Avertin, and the abdomen
was opened to expose the portal vein. Ice-cold saline was perfused through the portal
vein and the liver was excised. Emulsion injection studies for each emulsion type in
control, and LDL-receptor- and apo E-knockout mice were done on the same day,
using the same emulsion for all mice. Parallel studies after injection of 50 pi 0.5%
NaCl in control mice were used to control for background fluorescence in the liver
samples at specified time points. The average plasma concentrations for triglyceride
were 0.8, 0.78 and 0.94 mg/ml for control, and apo E- and LDL-receptor-deficient
mice, respectively. The average plasma concentrations for cholesterol were 0.9
mg/ml, 4.2 mg/ml and 2.9 mg/ml for control, and apo E- and LDL-receptor-deficient
mice, respectively.
5.2.6 Experimental Procedure for the Preparation of Liver
Samples
Liver pieces (3-4 mm3) were fixed in 4% paraformaldehyde in 0.1 M cacodylate
buffer fixing solution and left overnight. The liver was then blotted, snap-frozen with
isopentane in liquid nitrogen, and sectioned (approximately 10-15 p m ) by cryostat
(Bright, U K ) . Sections were collected on gelatin-coated slides and mounted in an
aqueous medium.
5.2.7 Confocal Laser Scanning Microscopy
Digital images of liver sections were generated by confocal laser fluorescent scanning
microscopy (CLSM, BioRad M R C 1000) on an upright Nikon Microscope
(OptiopHot-2) on the same day as mounting, and viewed by Comos™ software. Nikon
phase lenses (Plan Apo 40 X lens, N.A., 1.0, Bio-Rad Microscience, Hemel
Hempstead, U K ) were used which enabled generation of both fluorescent and light
transmission scans. Fluorescence was visible at an absorbance of 412 nm, as a single
band. Qualitative ratings of fluorescent intensity were utilised to describe hepatic
uptake of emulsion particles.

203
5.3 Results
5.3.1 Lipid Composition of Fluorescently-Labelled Chylomicron-
Like Emulsions
The average size and compositions of normal and modified chylomicron-like
emulsion particles were similar, as shown in Table 5.1. The lipid composition of the
normal emulsions was similar to that previously reported (Redgrave and Callow,
1990), (Martins et al, 2000a).
The introduction of the fluorescent label into the emulsion particle had no
effect the composition of chylomicron-like emulsions. The size of fluorescently
labelled chylomicron-like emulsions was consistent with the size of non-labelled
chylomicron-like emulsions, suggesting that the fluorescent labelling procedure or the
incorporation of the label, had not altered the physical dimensions of the particle or
caused particle aggregation (Table 5.1).
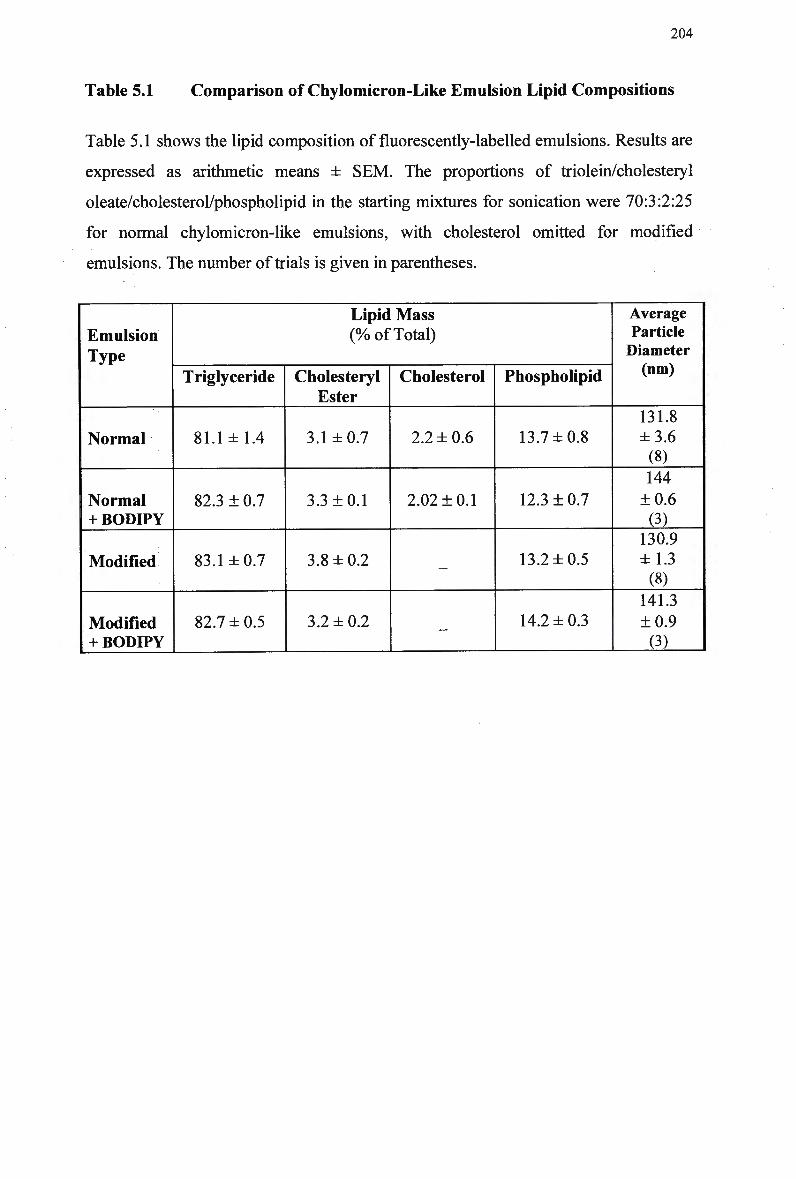
204
Table 5.1 Comparison of Chylomicron-Like Emulsion Lipid Compositions
Table 5.1 shows the lipid composition of fluorescently-labelled emulsions. Results are
expressed as arithmetic means ± SEM. The proportions of triolein/cholesteryl
oleate/cholesterol/phospholipid in the starting mixtures for sonication were 70:3:2:25
for normal chylomicron-like emulsions, with cholesterol omitted for modified
emulsions. The number of trials is given in parentheses.
Emulsion Type
Normal
Normal + BODIPY
Modified
Modified + BODIPY
Lipid Mass (% of Total)
Triglyceride
81.1 ±1.4
82.3 ± 0.7
83.1 ±0.7
82.7 ± 0.5
Cholesteryl Ester
3.1 ±0.7
3.3 ±0.1
3.8 ±0.2
3.2 ±0.2
Cholesterol
2.2 ± 0.6
2.02 ±0.1
—
—
Phospholipid
13.7 ±0.8
12.3 ±0.7
13.2 ±0.5
14.2 ±0.3
Average Particle Diameter (nm)
131.8 ±3.6 (8) 144
±0.6 (3)
130.9 ±1.3 (8)
141.3
±0.9 (3)

205
5.3.2 Hepatic Uptake of Normal Chylomicron-Like Emulsion in
LDL-Receptor-Deficient Mice
Preliminary studies in rats found that the plasma clearance and net organ uptakes of
[14C]triolein and cholesteryl[3H]oleate in emulsions with and without the fluorescent
cholesteryl ester were similar, indicating that the lipolysis and clearance kinetics were
not altered by the introduction of the fluorescent label into the emulsion particle (data
not shown).
The metabolism of normal chylomicron-like emulsions was studied in control
(C57BL/6J) and LDL-receptor deficient mice. To follow the intracellular pathway of
remnants in the liver, emulsions labelled with a fluorescent cholesteryl ester
(BODIPY) were injected, and liver sections were processed at given times and
assayed by laser confocal microscopy. The metabolism of emulsions in mice deficient
in LDL-receptor was compared with that of control C57BL/6J mice.
Figure 5.1 shows confocal images of the patterns of the uptake and
localisation of fluorescently labelled normal chylomicron-like emulsion remnants in
liver sections of control mice at 5 min (panel A ) , 20 min (panel B ) and 2 hr (panel C)
after intravenous injection into intact mice. The patterns of normal emulsion uptake
and metabolism in the liver of LDL-receptor deficient mice are also shown for 5 min
(panel D ) , 20 min (panel E) and 2 hr (panel F) after intravenous injection. The
intensity of hepatic fluorescence for control and LDL-receptor-deficient mice is
shown in Table 5.2, and indicates the amount of normal emulsion cholesteryl oleate
present.
There was significant hepatocyte uptake of remnants from normal
fluorescently labelled chylomicron-like emulsions in control mice within 5 mins (A),
peaking at approx. 20 min. However, in the LDL-receptor-deficient mice, there was
little or no accumulation of remnants in the hepatocytes up to 20 min (D). B y 20 min
after injection, the fluorescent label was evenly distributed in the hepatocytes of
control mice (B), and had accumulated to provide peak fluorescence. In contrast,
streaks of fluorescent remnants accumulated in the sinusoidal spaces of the L D L -
receptor-deficient mice at 20 min (E), however the net fluorescence produced by
labelled emulsion remnant particles was minimal compared with control mice.
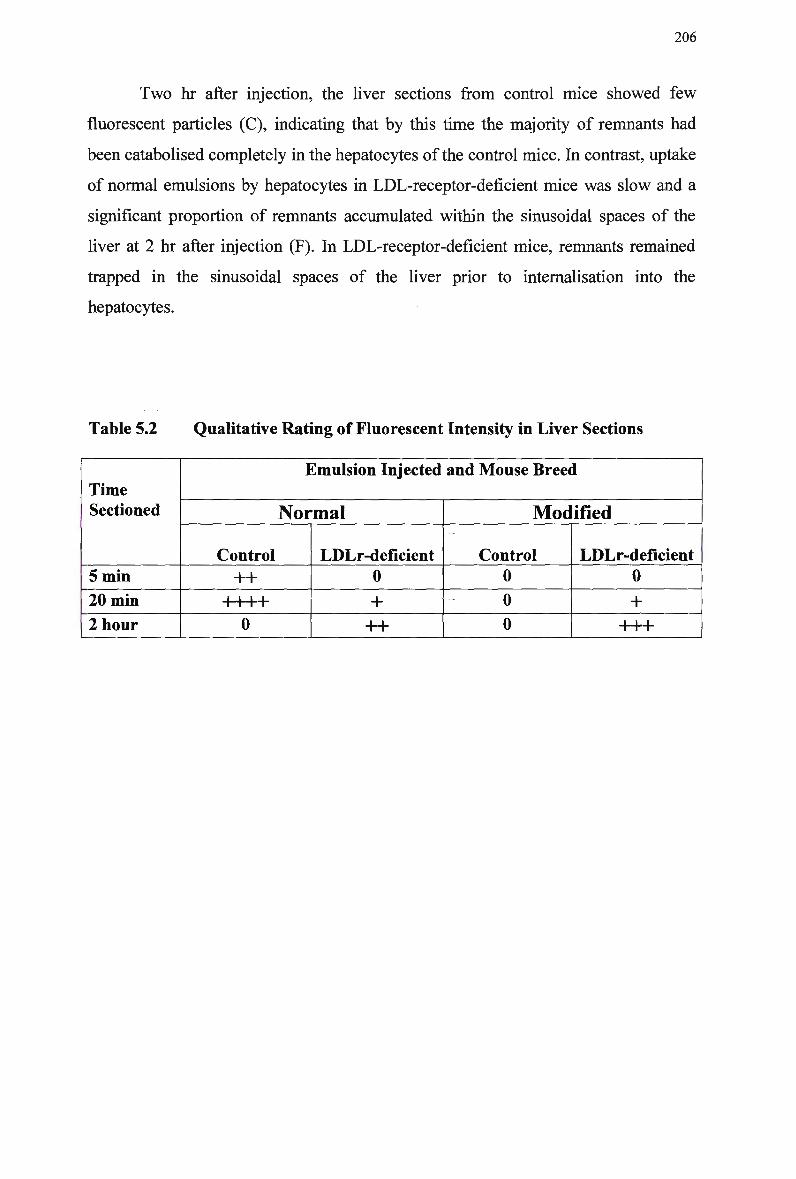
206
T w o hr after injection, the liver sections from control mice showed few
fluorescent particles (C), indicating that by this time the majority of remnants had
been catabolised completely in the hepatocytes of the control mice. In contrast, uptake
of normal emulsions by hepatocytes in LDL-receptor-deficient mice was slow and a
significant proportion of remnants accumulated within the sinusoidal spaces of the
liver at 2 hr after injection (F). In LDL-receptor-deficient mice, remnants remained
trapped in the sinusoidal spaces of the liver prior to internalisation into the
hepatocytes.
Table 5.2 Qualitative Rating of Fluorescent Intensity in Liver Sections
Time Sectioned
5 min
20 min
2 hour
Emulsion Injected and Mouse Breed
Normal
Control
++ i i i i T 1 1 P
0
LDLr-deficient
0
+
++
Modified
Control 0
0
0
LDLr-deficient
0
+ +++
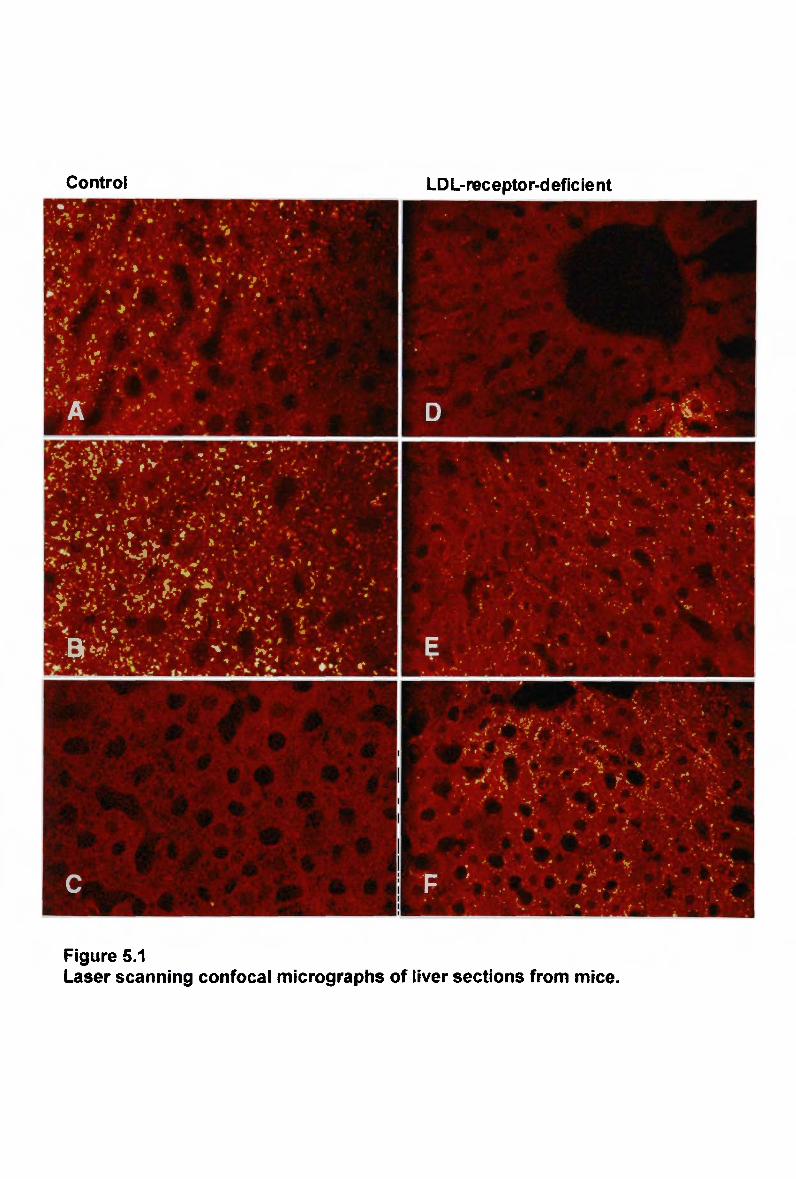
Control LDL-receptor-deficient
Figure 5.1 Laser scanning confocal micrographs of liver sections from mice.

208
5.3.3 Hepatic Uptake of Modified Chylomicron-Like Emulsion in
LDL-Receptor-Deficient Mice
The hepatic uptake of modified emulsion remnants was studied in control (C57BL/6J)
and LDL-receptor-deficient mice. The purpose of assessing the hepatic uptake was to
establish that this modified emulsion did not interact with high-affinity (receptor)
mechanisms.
Figure 5.2 shows the patterns of modified emulsion uptake and metabolism in
the livers of control mice at 5 min (panel A ) , 20 min (panel B ) and 2 hr (panel C) after
intravenous injection into intact mice. The patterns of modified emulsion uptake and
metabolism in the liver of LDL-receptor deficient mice are also shown for 5 min
(panel D ) , 20 min (panel E) and 2 hr (panel F) after intravenous injection. Qualitative
ratings of hepatic fluorescent intensity for control and LDL-receptor-deficient mice
are shown in Table 5.2, and indicate the amount of modified emulsion cholesteryl
oleate present.
The pattern of uptake of the modified emulsion was similar irrespective of
LDL-receptor expression. Critically, there was essentially no hepatic uptake of the
modified emulsion at 5 min in control (A) or LDL-receptor-deficient (D) mice. The
confocal micrographs of livers 20 min after injection of the modified chylomicron-
like emulsion showed considerably reduced uptake of fluorescent particles in control
mice (B) and minimal uptake in LDL-receptor-deficient mice (E).
T w o hr after injection of the modified emulsion, the liver sections from
control mice showed few fluorescent particles (C), indicating that remnants had been
taken up slowly. In contrast, in LDL-receptor-deficient mice, 2 hr after injection of
emulsion, fluorescence was located within the sinusoidal spaces, with some uptake by
hepatocytes (F).
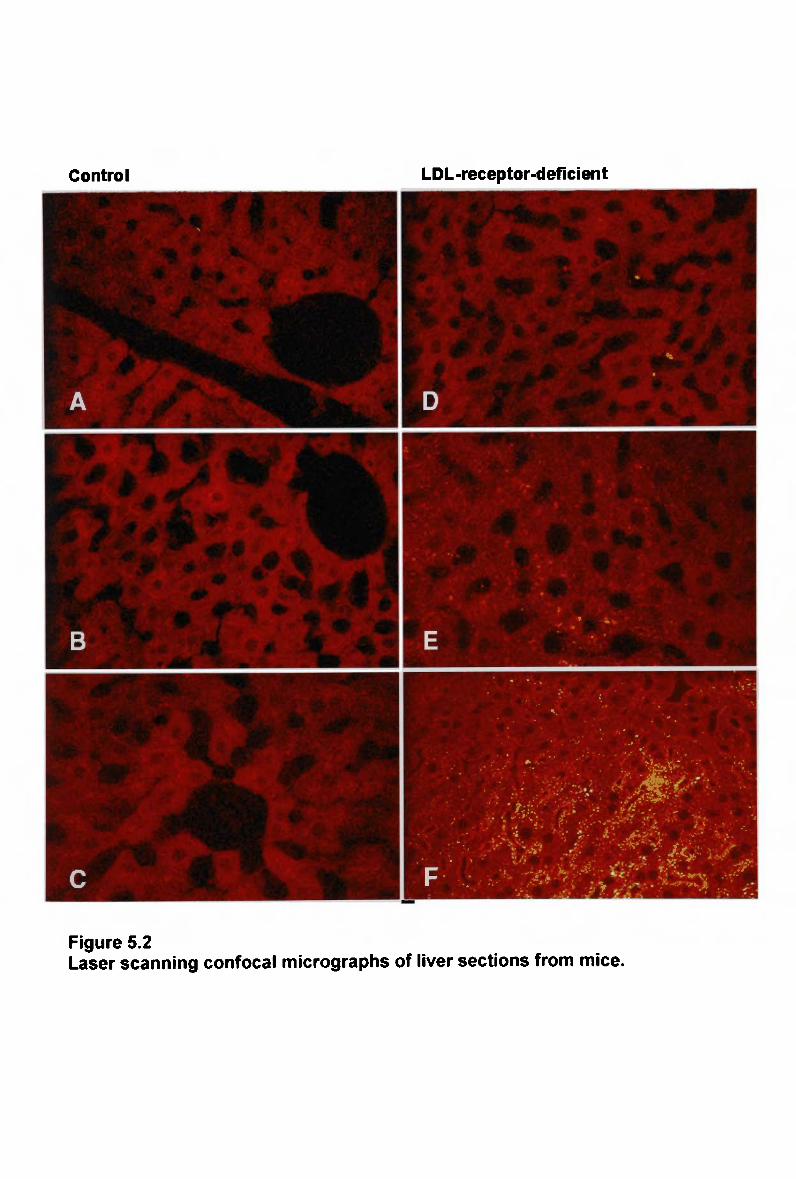
Control LDL -receptor-deficient
Figure 5.2 Laser scanning confocal micrographs of liver sections from mice.

210
5.3.4 Hepatic Uptake of Normal Chylomicron-Like Emulsion in
Apo E-Deficient Mice
To confirm the absolute requirement of apo E for the uptake of chylomicrons via high
affinity (receptor) pathways, the metabolism of normal and modified chylomicron-
like emulsions was studied in control (129/SV) and apo E-deficient mice. Emulsions
labelled with a fluorescent cholesteryl ester were injected into mice, and livers
sectioned at 5, 20 and 120 min.
Figure 5.3 shows confocal images of the patterns of normal emulsion uptake
and metabolism in the liver of control mice at 5 min (panel A ) , 20 min (panel B ) and
2 hr (panel C ) after intravenous injection into intact mice. The patterns of normal
emulsion uptake and metabolism in the liver of apo E-deficient mice are also shown
for 5 min (panel D ) , 20 min (panel E) and 2 hr (panel F) after intravenous injection.
Qualitative ratings of hepatic fluorescent intensity for control and apo E-deficient
mice are shown in Table 5.3, and indicate the amount of normal emulsion cholesteryl
oleate present.
Lipid remnants from normal emulsion were evenly distributed within the
hepatocytes of control mice as early as 5 min after injection of the emulsion (A), as
indicated by the presence of fluorescence on the cell surface. In contrast, fluorescently
labelled remnants were not associated with the liver cells of apo E-deficient mice (D).
Hepatocytes from control mice were stained intensely 20 min after injection of
fluorescent chylomicron-like emulsion (B), suggesting that more emulsion lipid had
been internalised and accumulated. In the apo E-deficient mice liver, fluorescence
was still undetectable (E).
T w o hours after injection, the fluorescent intensity decreased in the livers from
control mice (C), suggesting that most of the lipid emulsion had been metabolised into
undetectable products. In comparison with control mice the uptake of fluorescent
label in apo E-deficient mice was negligible (F). Staining in liver cells from apo E-
deficient mice was not evident at any time points following injection of the
fluorescence labelled emulsion, confirming the necessity of apo E as a ligand for
hepatic uptake of chylomicron remnants.
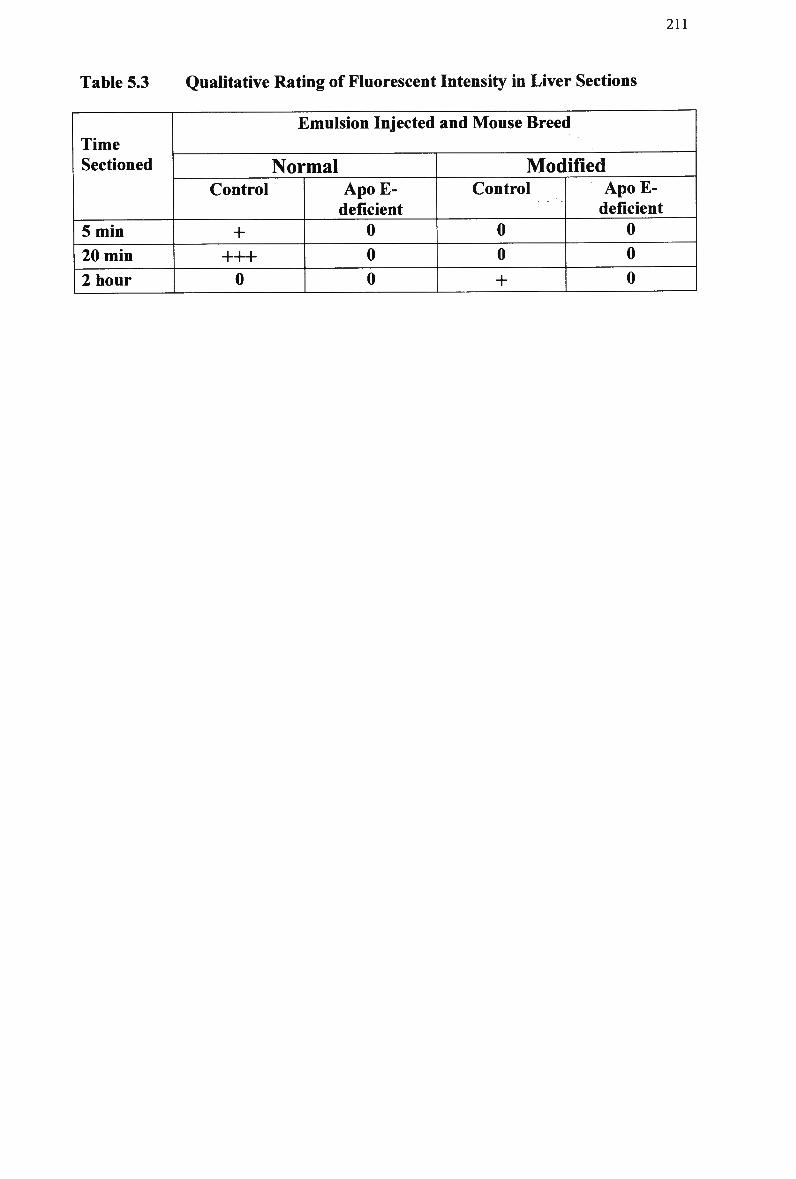
211
Table 5.3 Qualitative Rating of Fluorescent Intensity in Liver Sections
Time Sectioned
5 min
20 min
2 hour
Emulsion Injected and Mouse Breed
Normal Control
+ +++ 0
Apo E-deficient
0 0
0
Modified Control
0 0
+
ApoE-deficient
0 0 0
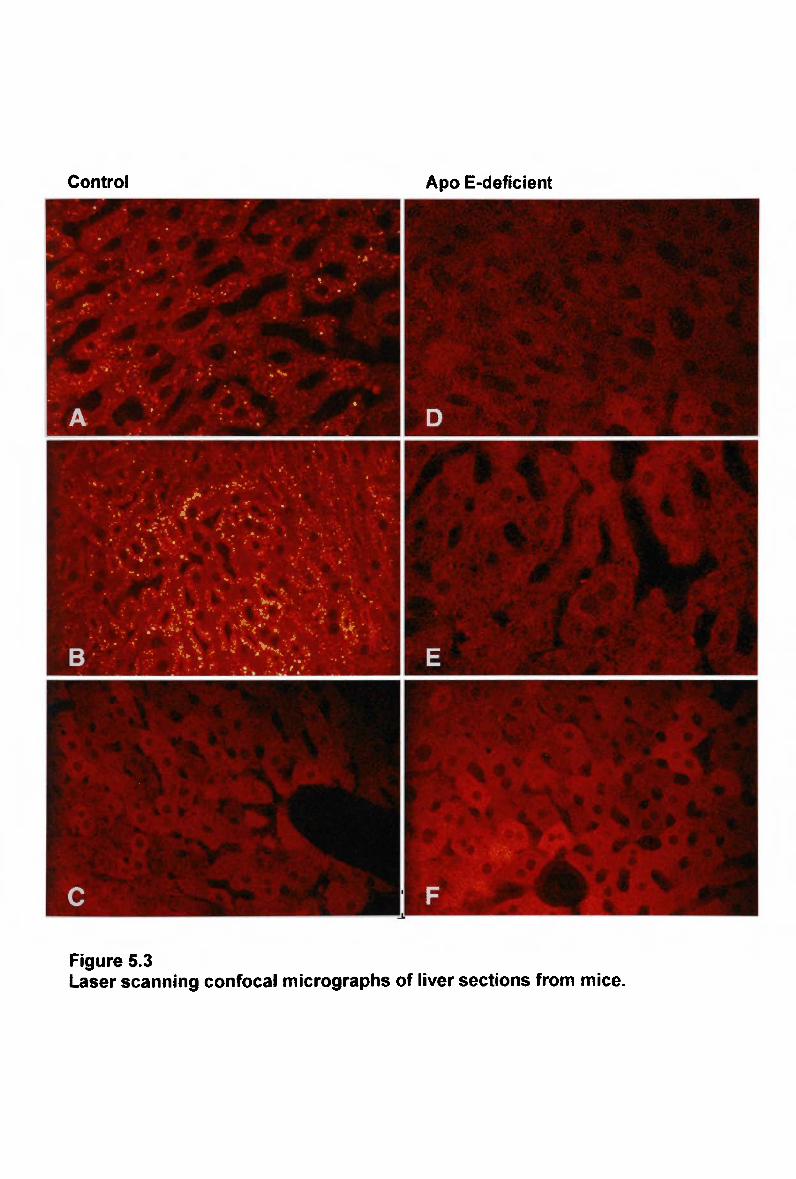
Control Apo E-deficient
Figure 5.3 Laser scanning confocal micrographs of liver sections from mice.

213
5.3.5 Hepatic Uptake of Modified Chylomicron-Like Emulsion in Apo E-Deficient Mice
The hepatic uptake of modified chylomicron-like emulsion remnants was studied in
control (129/SV) and apo E-deficient mice. Figure 5.4 shows the patterns of modified
emulsion uptake and metabolism in the liver of control mice at 5 min (panel A ) , 20
min (panel B ) and 2 hr (panel C ) after intravenous injection into intact mice. The
patterns of modified emulsion uptake and metabolism in the liver of apo E-deficient
mice are also shown for 5 min (panel D ) , 20 min (panel E) and 2 hr (panel F) after
intravenous injection. Qualitative ratings of hepatic fluorescent intensity for control
and apo E-deficient mice are shown in Table 5.3, and indicate the amount of modified
emulsion cholesteryl oleate present.
The pattern of uptake of the modified chylomicron-like emulsion was similar,
irrespective of apo E expression. There was essentially no hepatic uptake of the
modified emulsion by 5 min in control (A) or apo E-deficient (D) mice. The confocal
micrographs of livers 20 min after injection of emulsion showed considerably reduced
uptake of fluorescent particles in control mice (B) and no uptake in apo E-deficient
mice (E). T w o hours after injection, the increased fluorescent intensity within the liver
cells of control mice (C) suggested that some emulsion lipid had been internalised and
accumulated in the hepatocytes. However, few fluorescent particles from modified
emulsions were found to be located in hepatocytes. In contrast, liver sections from
apo E-deficient mice remained free of fluorescence (F).
5.3.6 NaCl
To observe background fluorescence, control (129/SV) mice were injected
intravenously with 50 pi saline (0.9% NaCl). Figure 5.5 shows the background
fluorescence in the liver of control mice at 5 min (panel A ) , 20 min (panel B) and 2 hr
(panel C ) after injection. There was no observable fluorescence at any time point.
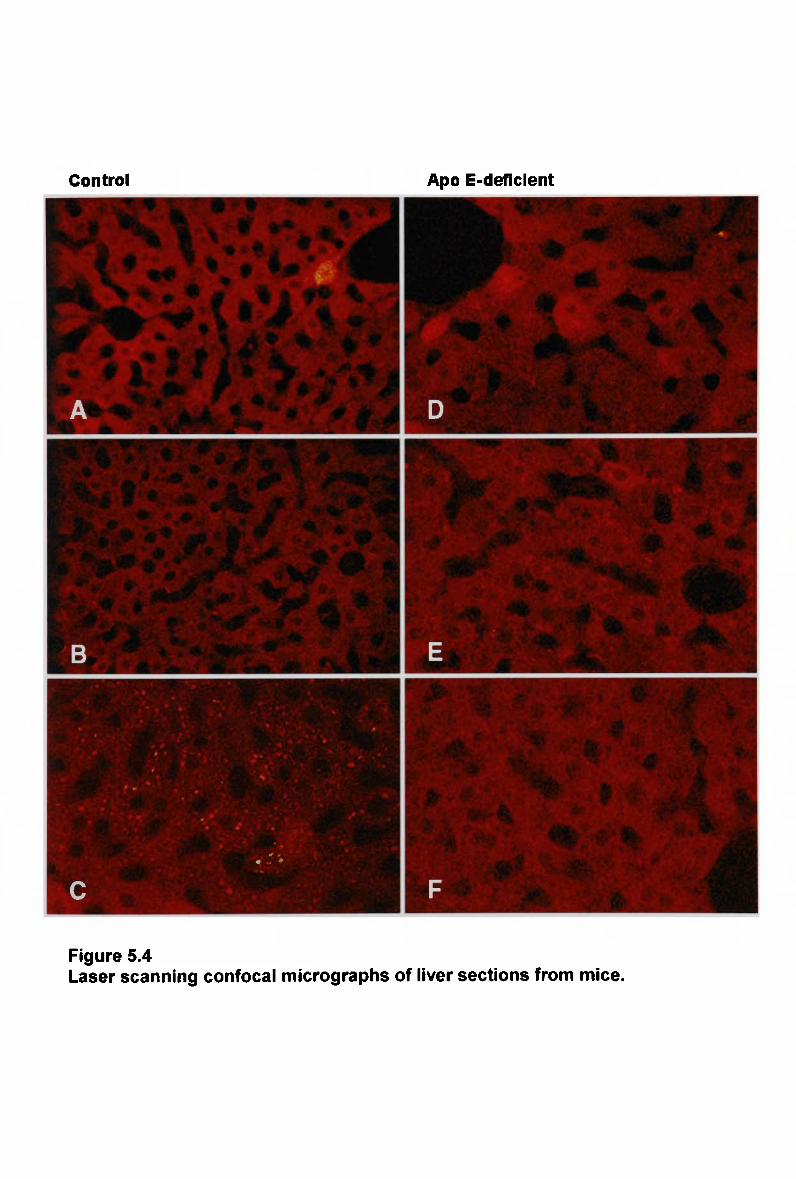
Control Apo E-deficient
| 1 • • :
Figure 5.4 Laser scanning confocal micrographs of liver sections from mice.
Figure 5.5 Laser scanning confocal micrographs of liver sections from control mice.
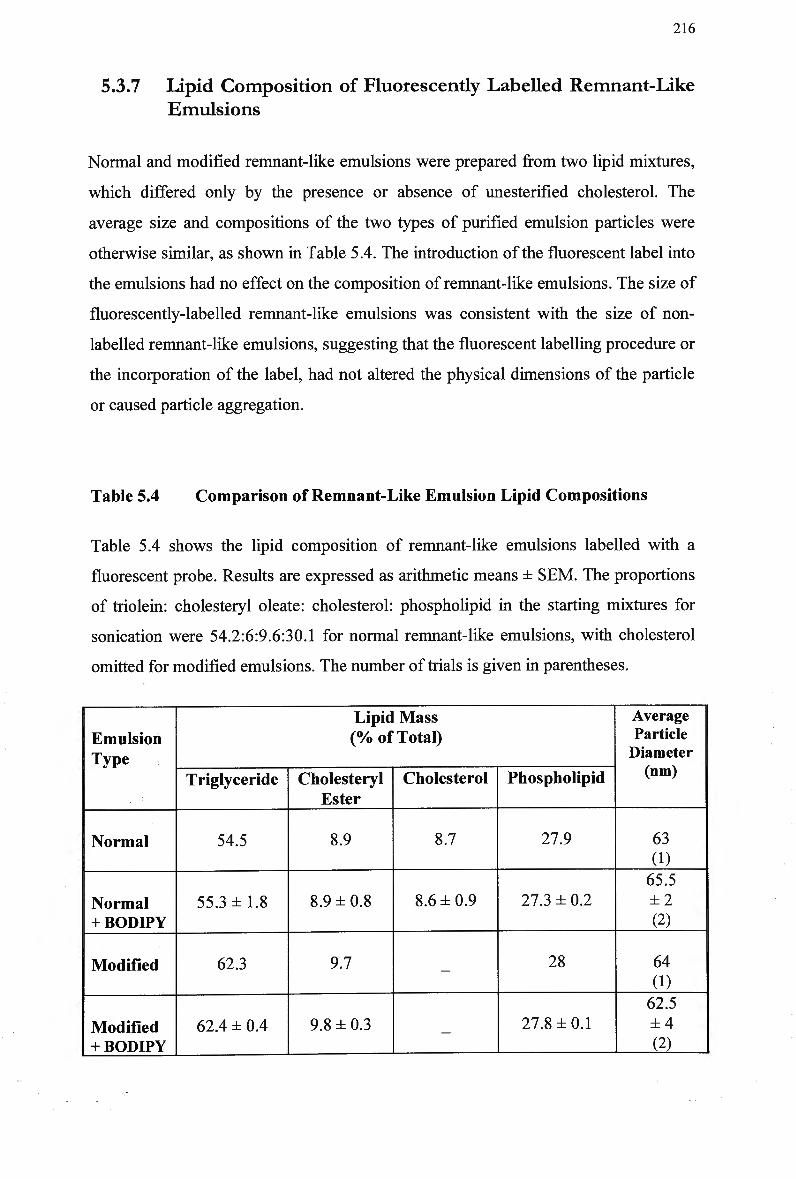
216
5.3.7 Lipid Composition of Fluorescently Labelled Remnant-Like
Emulsions
Normal and modified remnant-like emulsions were prepared from two lipid mixtures,
which differed only by the presence or absence of unesterified cholesterol. The
average size and compositions of the two types of purified emulsion particles were
otherwise similar, as shown in Table 5.4. The introduction of the fluorescent label into
the emulsions had no effect on the composition of remnant-like emulsions. The size of
fluorescently-labelled remnant-like emulsions was consistent with the size of non-
labelled remnant-like emulsions, suggesting that the fluorescent labelling procedure or
the incorporation of the label, had not altered the physical dimensions of the particle
or caused particle aggregation.
Table 5.4 Comparison of Remnant-Like Emulsion Lipid Compositions
Table 5.4 shows the lipid composition of remnant-like emulsions labelled with a
fluorescent probe. Results are expressed as arithmetic means ± SEM. The proportions
of triolein: cholesteryl oleate: cholesterol: phospholipid in the starting mixtures for
sonication were 54.2:6:9.6:30.1 for normal remnant-like emulsions, with cholesterol
omitted for modified emulsions. The number of trials is given in parentheses.
Emulsion Type
Normal
Normal + BODIPY
Modified
Modified + BODIPY
Lipid Mass (% of Total)
Triglyceride
54.5
55.3 ±1.8
62.3
62.4 ± 0.4
Cholesteryl Ester
8.9
8.9 ±0.8
9.7
9.8 ±0.3
Cholesterol
8.7
8.6 ±0.9
—
-
Phospholipid
27.9
27.3 ± 0.2
28
27.8 ±0.1
Average Particle Diameter (nm)
63
(1) 65.5 ±2 (2)
64 0) 62.5 ±4 (2)

217
5.3.8 Hepatic Uptake Following Simultaneous Injection of
Normal and Modified Remnant-Like Emulsions in Mice
The plasma clearance of remnant-like emulsion cholesteryl[3H]oleate was compared
in emulsions with and without fluorescent cholesteryl ester incorporated, in rats (data
not shown). Emulsions containing the fluorescent probe were cleared in a similar
manner compared with emulsions without the fluorescent probe, suggesting that the
clearance kinetics were not altered by the incorporation of the fluorescent probe into
the remnant-like emulsion particle.
The purpose of this set of experiments was to assess for competition of
emulsions for hepatic uptake mechanisms, at higher dose rates. This has repercussions
if the simultaneous injection of the two emulsion types is pursued. The metabolism of
normal remnant-like emulsions was studied in control (C57BL/6J) mice. Emulsions
labelled with a fluorescent cholesteryl ester were injected into control mice, and livers
sectioned at selected times. To determine if the metabolism of the normal emulsion
was altered or competed for by the simultaneous injection of modified emulsion (as
per proposed method), this was compared with the metabolism of normal emulsion
simultaneously injected with a modified emulsion (unlabeled). Control and knockout
mice were simultaneously injected with 250-280 pg triglyceride (30% of total
triglyceride pool). To follow the intracellular pathway of remnants in the liver, normal
emulsions were labelled with a fluorescent cholesteryl ester (BODIPY), injected, and
liver sections were processed and assayed by laser confocal microscopy.
Figure 5.6 shows the patterns of normal chylomicron remnant-like emulsion
uptake and metabolism in the liver of control mice at 5 min (panel A ) , 20 min (panel
B) and 2 hr (panel C ) after intravenous injection into intact mice. The patterns of
normal chylomicron remnant-like emulsion uptake and metabolism in the liver of
control mice following simultaneous injection of labelled normal emulsion plus
unlabeled modified emulsion are also shown for 5 min (panel D ) , 20 min (panel E)
and 2 hr (panel F). Qualitative ratings of hepatic fluorescent intensity for control mice
are shown in Table 5.5, and indicate the amount of normal emulsion cholesteryl oleate
present.
The normal chylomicron remnant-like emulsion became associated with
hepatocytes as early as 5 min after injection for both experimental regimes (A and D ) ,
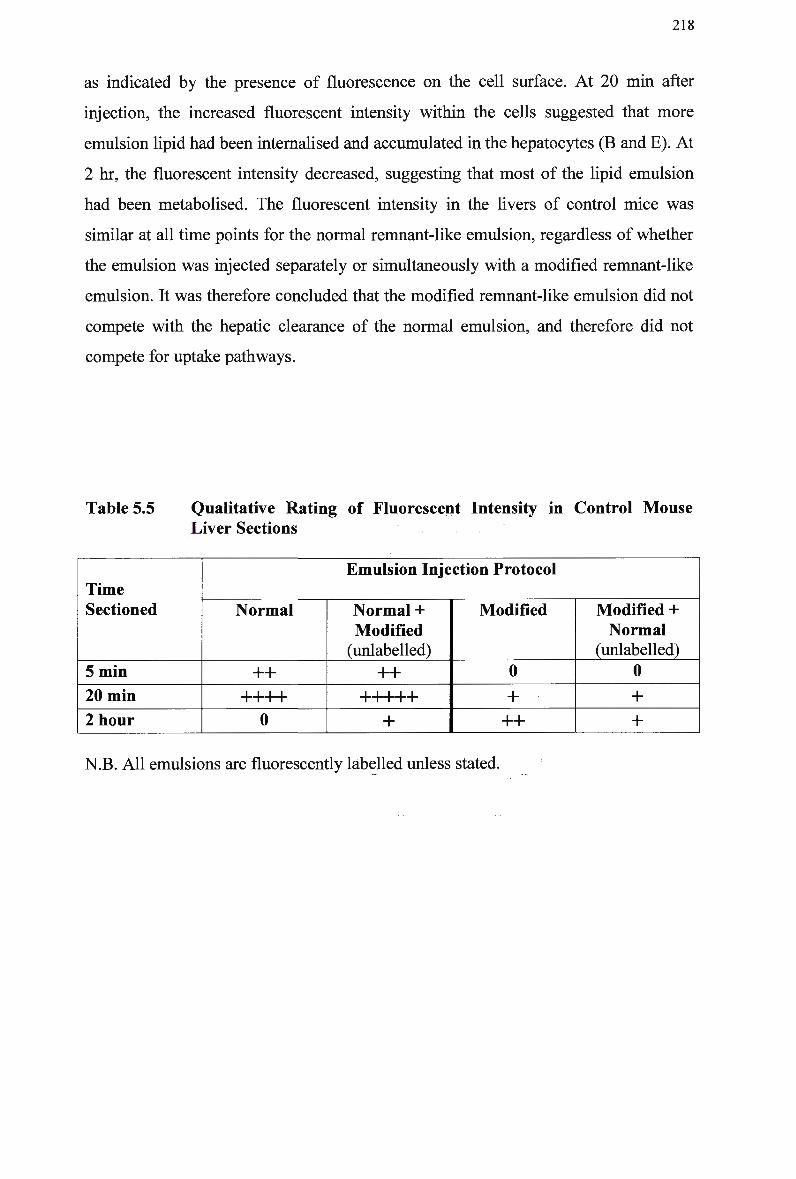
218
as indicated by the presence of fluorescence on the cell surface. At 20 min after
injection, the increased fluorescent intensity within the cells suggested that more
emulsion lipid had been internalised and accumulated in the hepatocytes (B and E). At
2 hr, the fluorescent intensity decreased, suggesting that most of the lipid emulsion
had been metabolised. The fluorescent intensity in the livers of control mice was
similar at all time points for the normal remnant-like emulsion, regardless of whether
the emulsion was injected separately or simultaneously with a modified remnant-like
emulsion. It was therefore concluded that the modified remnant-like emulsion did not
compete with the hepatic clearance of the normal emulsion, and therefore did not
compete for uptake pathways.
Table 5.5 Qualitative Rating of Fluorescent Intensity in Control Mouse Liver Sections
Time Sectioned
5 min
20 min
2 hour
Emulsion Injection Protocol
Normal
++ i i i i T 1 1 P
0
Normal + Modified (unlabeled)
++ i i i i i T 1 1 1 P
+
Modified
0
+ ++
Modified + Normal
(unlabeled)
0
+ +
N.B. All emulsions are fluorescently labelled unless stated.
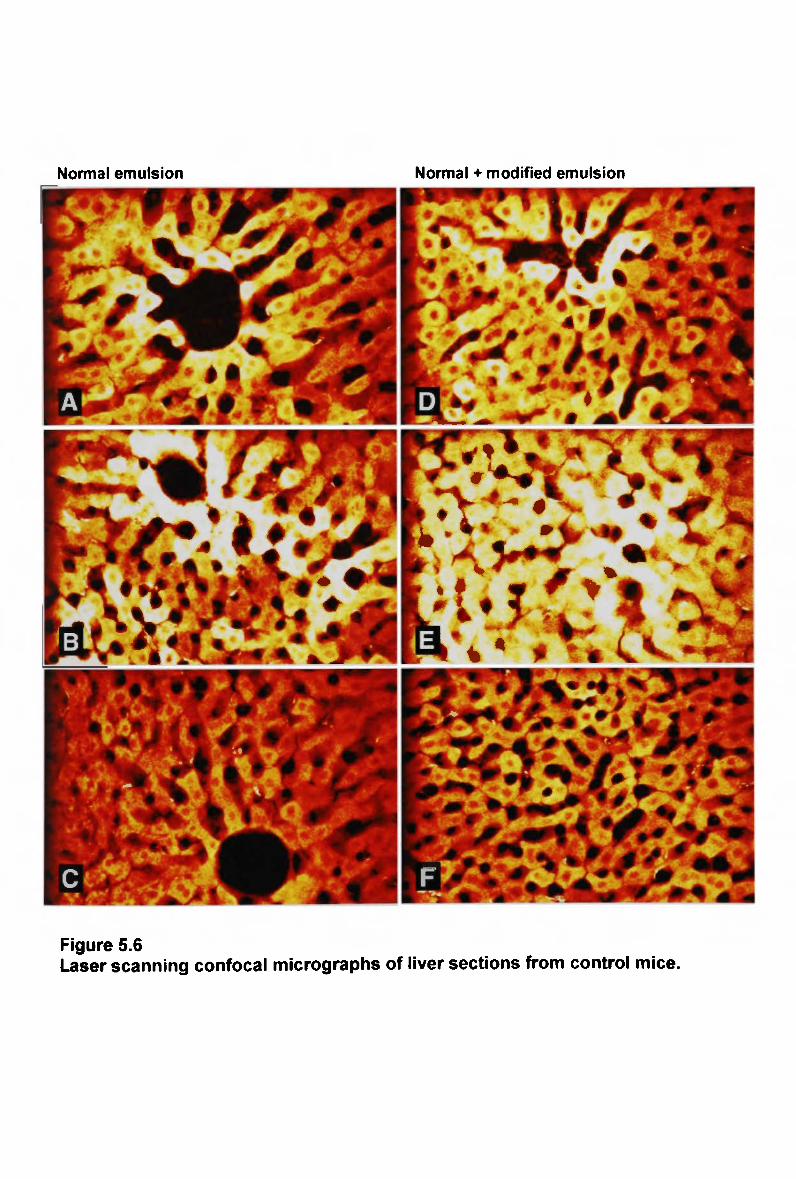
Normal emulsion Normal + modified emulsion
•••*•! * # * / v * ^ 1 *t> "SL: %
*
Figure 5.6 Laser scanning confocal micrographs of liver sections from control mice.

220
5.3.9 Hepatic Uptake Following Simultaneous Injection of
Modified and Normal Remnant-Like Emulsions in Mice
In this set of experiments, the metabolism of modified chylomicron remnant-like
emulsions was studied in control (C57BL/6J) mice. Emulsions labelled with a
fluorescent cholesteryl ester were injected, and livers sectioned at 5, 20 and 120 min.
To determine if the metabolism of the modified emulsion was altered by the
simultaneous injection of normal emulsion (as per proposed method), the uptake was
compared with the metabolism of fluorescently labelled modified emulsion
simultaneously injected with a normal emulsion (unlabeled).
Figure 5.7 shows the patterns of modified remnant-like emulsion uptake in the
liver of control mice at 5 min (panel A ) , 20 min (panel B) and 2 hr (panel C) after
intravenous injection. The patterns of modified emulsion uptake in the liver of control
mice following simultaneous injection of labelled modified emulsion and unlabeled
normal emulsion are also shown for 5 min (panel D ) , 20 min (panel E) and 2 hr (panel
F). Qualitative ratings of hepatic fluorescent intensity for control mice are given in
Table 5.5, and indicate the amount of modified emulsion cholesteryl oleate present.
There was essentially no hepatic uptake of the modified emulsion by 20 min in
control mice, irrespective of whether the emulsions were injected separately or
simultaneously with a normal emulsion. Few fluorescent particles from modified
emulsions were found in hepatocytes two hr after injection (C and F), suggesting that
a small amount of emulsion lipid had been internalised and accumulated in the
hepatocytes. The results suggest that the modified emulsions were metabolised in a
similar manner whether injected separately or simultaneously with a normal emulsion,
and the addition of the normal emulsion type did not appear to compete with the
uptake of modified emulsion by low affinity pathways in the liver.
5.3.10 NaCl
To observe background fluorescence, control (129/SV) mice were injected
intravenously with 50 pi saline (0.9% NaCl). Figure 5.8 shows the background
fluorescence in the liver of control mice at 5 min (panel A ) , 20 min (panel B) and 2 hr
(panel C) after injection. There was no observable fluorescence at any time point.
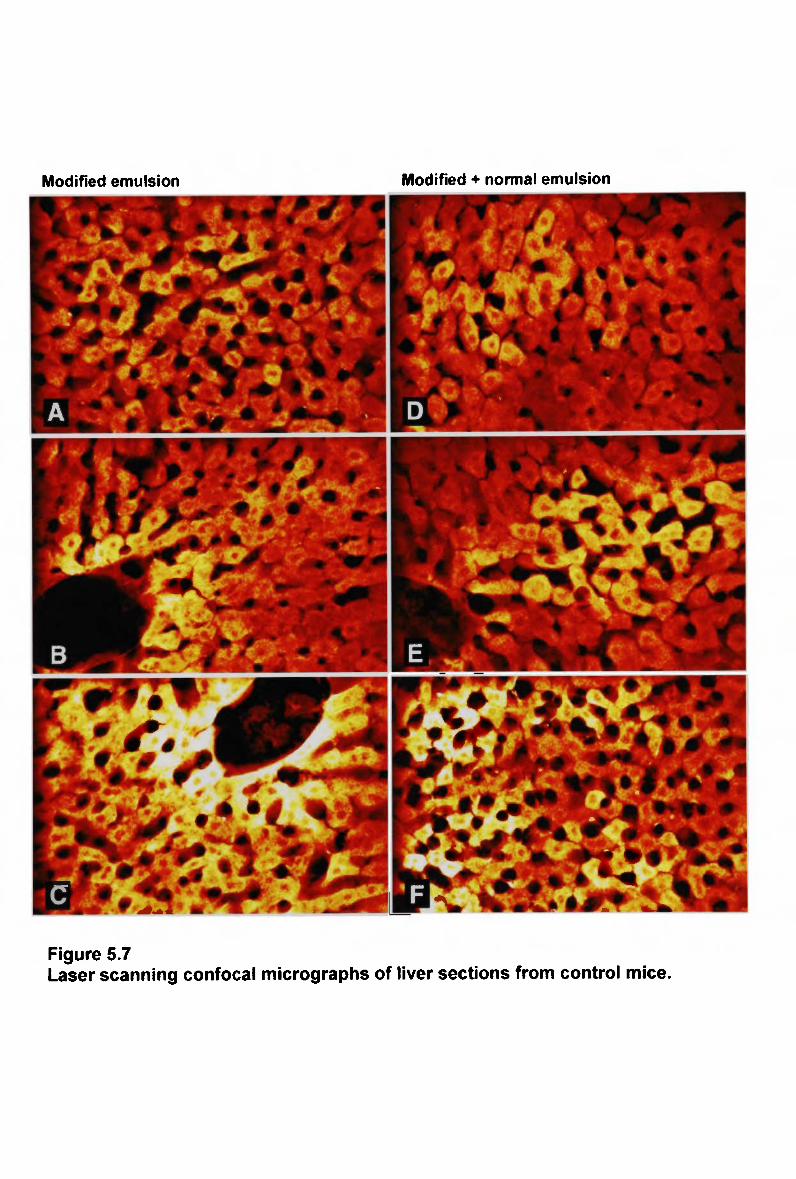
Modified emulsion Modified + normal emulsion
Figure 5.7 Laser scanning confocal micrographs of liver sections from control mice.
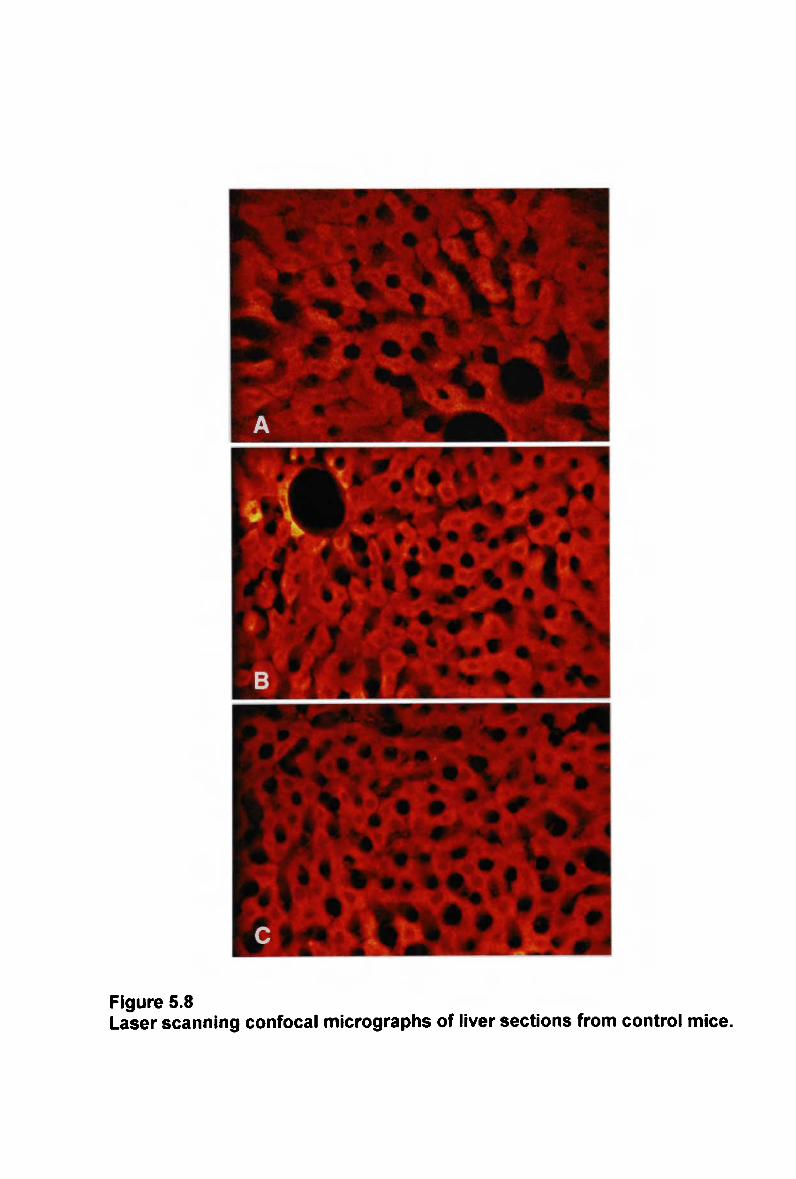
Figure 5.8 Laser scanning confocal micrographs of liver sections from control mice.

223
5.4 Discussion
In these experiments, a fluorescent cholesteryl ester label incorporated into
chylomicron-like emulsions was used to follow the uptake and metabolism of
remnants derived from the injected emulsions by the livers of gene targeted mice
homozygous for deficiency of LDL-receptor and apo E, as monitored by confocal
microscopy. The validity of mice as specific models of lipoprotein disorders and
atherosclerosis has been confirmed (Breslow, 1993). The LDL-receptor knockout
mouse provides a unique model for testing the contribution of receptors to
chylomicron remnant clearance in vivo, and at a cellular level (Mortimer et al,
1995a), (Martins and Redgrave, 1998). The C57BL/6J inbred strain of mouse is
susceptible to diet-induced arterial disease and apo E-knockout mice develop
extensive fatty streak lesions after consuming a fatty diet, characteristic of human
plaques (Stewart-Philips and Lough, 1991), (Zhang et al, 1992), (Plump et al, 1992),
(Paigen <?fa/., 1990).
Lipid emulsions were utilised in these experiments to study the kinetics in
vitro, and are advantageous because they do not contain any exogenous proteins or
apolipoproteins, which may nullify the defects of the knockout mice. The
chylomicron-like emulsions have been shown previously to become associated with
the endogenous apo E of the recipient animals after intravenous injection and
metabolised in a similar manner to lymph chylomicrons (Redgrave and Maranhao,
1985). The introduction of the fluorescent label into the emulsions had no effect on
mean particle size and composition of chylomicron- and remnant-like emulsions.
Preliminary studies showed that the incorporation of the fluorescent label into
chylomicron and remnant-like emulsions had no deleterious effect on the metabolism
of normal or modified emulsion clearance, as evidenced by plasma clearance and
organ uptake of emulsion radiolabeled cholesteryl oleate in rats. Previous studies in
rats have also verified that the metabolism of chylomicron-like emulsions is
unaffected by the presence of the fluorescent dye (Zeng et al, 1998). The clearance of
chylomicron remnants in mice is similar to remnant clearance in rats. Therefore, the
dose rates used in mice in these studies was equivalent to the dose rates previously
used in other rodent animal models, i.e., 1 5 % and 3 0 % of total triglyceride pool, for
separate and simultaneous injections, respectively. The dose rate was greater than
used in rabbit models and that predicted for use in human studies (approx. 1 0 %

224
triglyceride pool), and was designed to assess the possible competition effects on
hepatic uptake.
The LDL-receptor is the primary mechanism for the uptake of chylomicron
remnants from the plasma; therefore a delay in the hepatic uptake of normal emulsion
particles in LDL-receptor-deficient mice was expected, compared with control mice.
In contrast, the pattern of modified emulsion particle uptake is expected to be similar
for control and LDL-receptor-deficient mice, as the modified emulsion does not
interact with high affinity (receptor) mechanisms. In addition, the absence of
unesterified cholesterol does not allow apo E to associate with the particle surface or
reduces the affinity of apo E to bind the LDL-receptor, or both.
5.4.1 Patterns of Emulsion Uptake in LDL-Receptor Deficient
Mouse Liver
The results from the present study show that following injection of normal
chylomicron-like emulsion, there was measurable uptake of fluorescent label by the
liver of control mice at 5-20 min. In comparison, fluorescent remnants accumulated in
the sinusoidal spaces of the liver prior to internalisation into the hepatocytes in L D L -
receptor-deficient mice, and the endocytosis of fluorescent particles was markedly
impaired. The remnants also remained within the hepatocytes for a longer period (up
to 2 hr) in the LDL-receptor-deficient mice, compared with control mice. The
defective remnant uptake in LDL-receptor-deficient mice is consistent with the L D L
receptor being a mechanism of quantitative importance. These data also demonstrate
that when LDL-receptors are absent, hepatocytes can metabolise chylomicron
remnants by an alternate, slower uptake process.
The findings from this study confirm those of Mortimer et al. (Mortimer et al,
1995a), w h o also demonstrated a delay in uptake of fluorescently labelled
chylomicron-like emulsion particles by hepatocytes in LDL-receptor-deficient mice.
Confocal images of liver sections showed that remnants accumulated at the boundary
of the sinusoidal spaces in LDL-receptor-deficient mice, while remnants distributed
evenly in the hepatocytes of the control mice from 5 to 20 min after injection. T w o
hours after injection of the emulsions, no fluorescence was detected in the liver
sections obtained from control mice, suggesting catabolism of remnants, whereas in
mice deficient in LDL-receptors, fluorescent remnants were evenly distributed in

225
hepatocytes. Compared with control mice, fluorescently labelled remnants
accumulated at the boundary of the sinusoidal spaces. B y 3 hours emulsion particles
were evenly distributed within the hepatocytes, however endocytosis of the
fluorescent particles into hepatocytes was delayed. Their results suggested that in the
absence of the LDL-receptor, catabolism of the chylomicron remnants was probably
defective. The authors also found that the rates of chylomicron remnant clearance
from plasma were similar in control and LDL-receptor-deficient mice, with similar
liver uptakes of radiolabeled remnants. Evidently, the remnants were trapped in the
sinusoids but not internalised by hepatocytes. They concluded that the slow
catabolism of remnants was due to the slow internalisation of remnants via an
alternative apo E-dependent pathway.
Herz et al (Herz et al, 1995) studied the endosomal uptake of radioactive
labels to show a delay in the endocytosis of chylomicron remnants in LDL-receptor-
deficient mice, as evidenced by their accumulation in the endosomal fraction and by
the rate of hydrolysis of component cholesteryl esters. The rate of chylomicron
remnant removal by the livers of LDL-receptor-deficient mice was normal up to 30
min post-injection, leading the authors to conclude the initial hepatic removal of
chylomicron remnants is mediated by mechanisms that do not include the L D L -
receptor or the LRP. However, after the remnants bind to the hepatocytes, endocytosis
was primarily mediated by the LDL-receptor.
The hepatic uptake of modified emulsions was also investigated in control and
LDL-receptor-deficient mice. The metabolism of modified chylomicron-like
emulsions has not previously been investigated at a cellular level. The uptake of
modified emulsion particles was delayed in mice, regardless of LDL-receptor
expression. Fluorescent particles from modified emulsions were only found to be
located in hepatocytes of control mice 2 hr post-injection, confirming that the
modified emulsion did not interact significantly with rapid, high-affinity uptake
processes. In comparison, there was some accumulation of fluorescent remnants in the
sinusoidal spaces of the liver of LDL-receptor-deficient mice at 20 min, followed by
internalisation of remnant particles into the hepatocytes at 2 hr.
Thus the difference between hepatic uptake of normal and modified emulsion
was substantial in the control mice, but not in the LDL-receptor-deficient mice. If a
second receptor pathway for chylomicron remnants existed, a difference in uptake
between normal and modified emulsions when injected into LDL-receptor-deficient

226
mice would have been observed. Rather, the pattern of hepatic uptake was similar in
these mice, and resembled the pattern of uptake of modified emulsion in control mice.
These data suggest that the emulsion particles were sequestered via low affinity,
alternate pathways into the sinusoidal spaces of the liver in LDL-receptor-deficient
mice, and it appears that these mice are efficient at utilising this pathway. The results
support the initial assertion of this study that modified emulsion particles do not
interact with high affinity (receptor) pathways in the liver, therefore their uptake is
considered to be an appropriate indicator of remnant uptake via low-affinity uptake
mechanisms. The presence of emulsion cholesterol is suggested to be a critical
determinant of early metabolic events, as triglyceride-depleted emulsion remnant
particles have been found to remain in the plasma instead of being rapidly taken up by
the liver (Redgrave et al, 1987). The present study confirmed the assertion that while
modified emulsions remain substrates for L P L and undergo normal triglyceride
hydrolysis, the cholesterol component is necessary for hepatic uptake of the emulsion
remnants.
The results confirm previous findings of Redgrave et al. (Redgrave et al,
1995) and Martins et al. (Martins et al, 2000a), that modified emulsions were
metabolised much slower than normal emulsions, after injection of chylomicron-like
emulsions labelled with cholesteryl[14C]oleate into rats and remnant-like emulsions
labelled with cholesteryl! COJoleate into mice, respectively. The appearance of
14C02 and 1 3C02 in the expired breath of rats was significantly less following injection
of modified emulsion compared with normal emulsion, indicating a defect or delay in
the catabolism of remnants. In the absence of cholesterol, the removal of emulsion
triglyceride has been shown to be little affected, however, remnant particle clearance
was markedly delayed, with less than 3 0 % removed from the plasma by 12 min after
injection into conscious rats (Mortimer et al, 1995b). Compared with normal
emulsion remnant uptake of 7 0 % by the liver, only 2 7 % of modified emulsion
remnants were recovered. The metabolism of the modified emulsion was found to
increase during the course of the experiment and may be related to uptake by
alternative receptor pathways possibly involving the L R P pathway, phagocytic or
scavenger pathways.
Other studies have used antibodies to the LDL-receptor and shown that liver
membranes have apo E binding sites that are not the LDL-receptor (Cooper et al,
1987). The authors concluded that it is likely that in the absence of the LDL-receptor

227
at least some of these sites can play a role in remnant removal. Based on liver
perfusion studies in rats, Windier et al. (Windier et al, 1996) concluded that under
conditions of long-standing LDL-receptor deficiency, other receptors may take over
its function, and a somewhat longer circulation may alter these particles and facilitate
their interaction with other binding sites.
The current findings are in keeping with the data from Chapter 3, and previous
clearance data in W H H L rabbits (Bowler et al, 1991), ( M a m o et al, 1991). All
studies showed that the clearance of normal and modified emulsions was significantly
delayed in homozygous or heterozygous W H H L rabbits, compared to the clearance of
the normal emulsions in control rabbits. The clearance of modified emulsions in
W H H L and control rabbits was similar, confirming the use of the two emulsions as a
method for quantifying receptor uptake. Certainly the present findings that the hepatic
uptake of normal and modified emulsions in LDL-receptor-negative mice is similar,
suggesting that the modified emulsion did not interact with the LDL-receptor. The
results reflect the clearance data from Chapter 3 at a cellular level, i.e.; the clearance
of the modified emulsion was delayed in control mice, and taken up via a slower, low
affinity pathway. The difference in clearance of the two emulsion types is therefore
indicative of high affinity uptake.
It is therefore suggested that under normal circumstances, chylomicron-like
emulsion remnants are rapidly taken up by receptor pathways and internalised by
hepatocytes, and that LDL-receptor is the primary route for the uptake of chylomicron
remnants from plasma. High-affinity uptake mechanisms operate more rapidly than
those of non-specific (low-affinity) mechanisms therefore these data support the
removal of normal emulsion via high-affinity pathways. W h e n the LDL-receptor is
absent, chylomicron-like emulsions are taken up via a second pathway, first to the
sinusoidal space of the liver, with subsequent slow endocytosis and slow catabolism.
The modified emulsion does not appear to interact with receptor uptake mechanisms
in control mice, and the uptake observed occurs via non-specific mechanisms. W h e n
the LDL-receptor is absent, uptake of the modified emulsion is slowed, but was
greater in comparison to controls, suggesting that uptake via an alternate pathway
may be more efficient in LDL-receptor-deficient mice.

228
5.4.2 Patterns of Emulsion Uptake in Apo E-knockout Mouse
Liver
Apo E is utilised as the ligand for hepatic uptake of chylomicron remnants, therefore
the clearance of chylomicron-like emulsions was studied in apo E-knockout mice to
verify the essential requirement of apo E as a ligand for the hepatic uptake of
chylomicron remnants via high affinity pathways. A delay in the hepatic uptake of
normal emulsion particles in apo E-deficient mice was expected, compared with
control mice. In contrast, the pattern of modified emulsion particle uptake was
expected to be similar for control and apo E-deficient mice, as the absence of
unesterified cholesterol does not allow apo E to associate with the particle surface or
reduces the affinity of apo E to bind the LDL-receptor, or both.
Following injection of normal chylomicron-like emulsion, hepatocytes from
control mice stained intensely at 5 and 20 min. By 2 hr, there was little fluorescence
in the livers of control mice. Staining in liver cells from apo E-deficient mice was not
evident at any time points. These data demonstrate that when apo E is absent,
hepatocytes are not able to metabolise chylomicron remnants and confirm the critical
requirement for apo E in the hepatic clearance of remnants.
The findings from this set of experiments confirm those of Mortimer et al.
(Mortimer et al, 1995a), who found that micrographs from apo E-deficient mice
showed no fluorescent particles at any time after injection of chylomicron-like
emulsion, indicating that remnant removal from plasma was totally impeded in apo E-
deficient mice. In comparison, fluorescent particles were evenly distributed at 5-20
min post-injection in control mice. T w o hours after injection, little fluorescence was
detected in the liver sections of the control mice, indicating that by this time remnants
had been catabolised. Moreover, plasma removal of emulsion remnants was totally
impeded and measurement of expired radioactive C O 2 following injection of l C-
cholesteryl oleate-labelled emulsion indicated that remnant metabolism in apo E-
deficient mice was essentially nil. The slow catabolism of remnants by apo E-
deficient mice was confirmed by Mortimer et al. (Mortimer et al, 1997) who found
that over 6 0 % of injected emulsion cholesteryl oleate remained in the plasma of apo
E-deficient mice 30 min after injection of labelled emulsions. This was significantly
less than for control mice, suggesting defective remnant clearance in mice deficient in
apo E. In addition, over 6 0 % of the injected [3H]-cholesteryl oleate was recovered in

229
the livers of the control mice, compared with 5 % recovery in apo E-deficient mice
livers. The hepatic uptake of [14C]-triolein was similar in control and apo E-deficient
mice, indicating normal lipolysis of triglycerides.
In a study by Ishibashi et al. (Ishibashi et al, 1994b), apo E-deficient mice had
a marked elevation in apo B-48 but not apo B-100. The observation that apo B-48
increases more dramatically with apo E-deficiency than with LDL-receptor-deficiency
led the authors to conclude that apo E binds to a second receptor in addition to the
LDL-receptor. This was supported by the observation that in apo E/LDL-receptor-
deficient double homozygote mice, hypercholesterolemia does not increase beyond
the level observed in apo E-deficiency alone. However, the absence of apo E would
not allow the binding of chylomicron remnant particles to any receptor for which it is
a ligand, whereas LDL-receptor-deficient mice may be more efficient at utilising
alternate low-affinity pathways for initial uptake into the sinusoidal spaces of
hepatocytes. In later experiments, Ishibashi et al. (Ishibashi et al, 1996) administered
vitamin A fat-tolerance tests to mice deficient in the LDL-receptor, apo E, and both
apo E and the LDL-receptor. The area under the plasma retinyl ester curves were 4,
12 and 12 times larger in mice deficient in LDL-receptor, apo E, and both apo
E/LDL-receptor respectively, compared with control mice, and LDL-receptor-
deficient mice retained chylomicrons compared with control mice. The results suggest
that the LDL-receptor pathway plays a significant role in remnant metabolism,
however the larger retinyl ester excursion in apo E-deficient mice indicates the
presence of an apo E-dependent pathway for the clearance of retinyl ester. It appears
that when the quantity of particles to be removed exceeds the ability of receptors to
internalise them, they may become trapped in the space of Disse, or the sequestration
space. W h e n the quantity of particles that has been removed also exceeds the capacity
of this space and the ability of the L R P to clear them from this space, the L D L -
receptor defect becomes manifest as a delay in their removal (Cooper, 1997).
The patterns of uptake of the modified chylomicron-like emulsion were
similar in control and apo E-deficient mice. Critically, there was residual uptake of
fluorescent particles by hepatocytes by 20 min, with minimal fluorescent intensity
within the liver cells of control mice at 2 hr and no observed uptake by livers of apo
E-deficient mice. The micrographs indicate that some emulsion lipid had been
internalised and accumulated in the hepatocytes of control mice, indicating that
remnants were taken up via slower, low-affinity processes. There was essentially no

230
difference in uptake of modified emulsion in control and apo E-deficient mice,
suggesting that the modified emulsion did not interact with apo E.
The central role played by apo E in promoting the recognition and metabolism
of remnant lipoproteins by hepatic receptors has been established by several lines of
evidence (Wilson et al, 1991), (Windier et al, 1980a), (Sherrill et al, 1980),
(Shelbourne et al, 1980), (Sherrill and Dietschy, 1978). Small remnants carry fewer
apo E molecules, decreasing the likelihood and affinity of binding to LDL-receptors
in the liver (Redgrave et al, 1996). In contrast, the larger size of chylomicrons during
fat absorption by the intestine is associated with more rapid clearance of larger
particles from plasma (Rensen et al, 1997), (Chajek-Shaul et al, 1983), (Redgrave et
al, 1992-a), (Guldur and Mayes, 1992). These observations suggest that the rate of
clearance of apo E-containing lipoproteins from plasma is influenced by the amount
of apo E associated with each particle, and the atherogenicity of chylomicron
remnants appears to be increased in the absence of apo E.
Familial apo E-deficiency (type III hyperlipoproteinemia) impairs the plasma
clearance of lipoprotein particles that normally contain apo E, resulting in
hypertriglyceridemia, hypercholesterolemia and premature atherosclerosis (Mabuchi
et al, 1989). Homozygotes for apo E-deficiency have markedly retarded fractional
catabolism of apo B-48, and a low synthesis rate of apo E, indicating that apo E is
essential for the normal catabolism of triglyceride-rich lipoprotein constituents
(Schaefer et al, 1986). In type III hyperlipidemia, the presence of a particular apo E
phenotype that does not bind to liver receptors, or a rare cause of apo E deficiency,
leads to an accumulation of (3-VLDL (Mahley, 1988). These findings have confirmed
the critical role of apo E in directing the clearance of remnants from plasma and show
that the amount and type of apo E available modulate remnant clearance.
Remnant lipoproteins have been shown to accumulate in the plasma of apo E-
deficient mice, and the mice developed hypercholesterolemia and premature
atherosclerosis (Breslow, 1993), even when fed a low-fat chow diet (Zhang et al,
1992). The complexities of the atherosclerotic lesions that develop in apo E-deficient
mice are similar to those described in other species (Reddick et al (1994). Similar
results have been reported, whether apo E-deficient mice were fed a low fat, low-
cholesterol or a western-type diet (Plump et al, 1992), (Nakashima et al, 1994), (van
Vlijmen et al, 1996). As a result of feeding an atherogenic diet, Zhang et al. (Zhang

231
et al, 1994) found extensive deposition of lipid-filled plaques outside the
cardiovascular system in mice homozygous for apo E-deficiency. Heterozgyous apo
E-deficient mice also displayed hypercholesterolemia (Van Ree et al, 1994a) and
large foam cell lesions, despite the return of apo E in lipoproteins to normal (Zhang et
al, 1994). The authors found that a huge amount of lipid carrying particles
accumulate in apo E-deficient mice; the particles are cholesterol-rich, indicating that
they were large remnant lipoproteins.
In contrast, Y u et al. (Yu et al, 2000) found that the livers of mice that lacked
both apo E and the LDL-receptor had a similar rate of removal of chylomicron
remnants perfused through the liver at relatively low remnant concentrations,
compared with C57BL/6J (wild-type) mice. However, livers from apo E-knockout
mice had reduced capacity in removing remnants at a relatively high concentration of
chylomicron remnants. The authors findings suggest that hepatically localised apo E
is not a critical factor in the rapid initial removal of chylomicron remnants by either of
the major pathways, but do suggest that apo E can be added to lipoproteins to
accelerate their uptake.
In transgenic mice expressing high levels of rat apo E, the total plasma lipids
were found to be significantly reduced compared with controls after high fat feeding
(Shimano et al, 1992a). The authors also demonstrated enhanced clearance from
plasma of injected V L D L , L D L and chylomicron remnants (Shimano et al, 1992b).
Immunohistochemical studies by Shimano et al. (Shimano et al, 1994) showed that
after an injection of a large amount of chylomicrons into mice overexpressing apo E,
the density of cell surface apo E was markedly reduced. Vesicular staining in the
cytoplasm was also observed, suggesting that cell-surface apo E was utilised for the
hepatic endocytosis of chylomicrons and remnants. In other studies, LDL-receptor-
deficient mice had a mean plasma cholesterol level significantly lower and fatty
lesions were suppressed, when they were overexpressing the apo E transgene
compared with LDL-receptor-deficient mice, on a normal chow diet or when fed an
atherogenic diet (Osuga et al, 1998). Combined, these data support the
antiatherogenic properties of apo E and suggest that mice overexpressing apo E are
protected from diet-induced hypercholesterolemia.
In light of the current fluorescent emulsion studies and other findings, it would
appear that clearance and hepatic uptake of normal and modified emulsion remnants

232
in apo E-deficient mice is minimal, and that apo E is a necessary ligand for the uptake
of chylomicron remnants.
5.4.3 Comparison of Patterns of Uptake of Remnant-Like
Emulsions Injected Following Simultaneous versus Separate
Injection
The use of the two-emulsion technique was investigated in Chapter 3, and the
simultaneous injection of the two emulsions was found to have no deleterious effect
on the metabolism of either emulsion. This method has been used previously in
rabbits (Bowler et al, 1991), (Mamo et al, 1991). However, it was imperative to
establish that there were no interaction or competition effects for hepatic uptake, and
to demonstrate that plasma clearance kinetics were reflected at the cellular level. The
simultaneous injection of the two emulsion types addresses the importance of
assessing competition at higher dose rates. If the clearance of normal emulsions via
high affinity uptake pathways is saturated, further uptake may occur via low
affinity/high capacity sites over time. In this case, the clearance data would indicate
normal or slowed plasma clearance but may not affect total A A C if high affinity
uptake mechanisms were saturated.
The reduced triglyceride mass in remnant-like emulsions reduces the
dependency on L P L for lipolysis prior to uptake by the liver, thereby decreasing the
number of confounding variables affecting the metabolism of the two emulsion types.
The use of remnant-like emulsions also facilitated an increase the cholesteryl ester
and retinyl ester mass injected into recipient mice, to observe any saturation of
particle uptake via high affinity pathways. The removal rates of cholesteryl oleate
during plasma clearance have been found to be similar for remnant-like and
chylomicron-like emulsions (Hirata et al, 1987). Remnant-like emulsions have been
used extensively in animal studies of chylomicron remnant metabolism (Martins and
Redgrave, 1998), (Martins et al, 2000a), (Martins et al, 2000b), (Zeng et al, 1998).
Fluorescently labelled remnant-like emulsions were used to follow the uptake
of normal emulsion remnants by the livers of control mice, as monitored by confocal
microscopy. To study the effect of a second, simultaneous injection of modified
emulsion (as per proposed two-emulsion method), the procedure was repeated, but
with an addition of an injection of unlabeled modified emulsion. This procedure was

233
repeated by comparing the hepatic uptake of modified emulsion with the uptake of
modified emulsion remnants following simultaneous injection of fluorescently
labelled modified emulsion and unlabeled normal emulsion. Hepatic uptake
following separate and simultaneous injection procedures was compared to determine
if simultaneous injection of the two emulsion types altered the patterns of uptake by
liver cells. N o difference in the hepatic uptake of emulsion particles was anticipated,
following simultaneous or separate injection of the two emulsion types.
Qualitative analysis of fluorescent micrographs and fluorescent intensity
indicated that remnant-like emulsions were metabolised in a similar manner, whether
injected separately or simultaneously with a second emulsion. This suggests there was
no alteration of the pattern of emulsion lipids, saturation of lipid uptake pathways or
competition between the two emulsion types for hepatic uptake. In addition, the
normal emulsion did not compete for uptake with the modified emulsion via low
affinity pathways, and the hepatic uptake of modified emulsion was delayed
compared to normal emulsion, irrespective of the emulsion injection protocol utilised.
These results are in agreement with previous clearance data for chylomicron-like
emulsions (Chapter 3). It was therefore concluded that the difference in clearance of
the two emulsions provides a measure of receptor activity in vivo.
5.4.4 Conclusion
Under normal conditions, chylomicron remnants are rapidly sieved through the
endothelial fenestrae of the liver, allowing entrance into the space of Disse. Remnants
may then be removed directly by LDL-receptors, acquire additional apo E that is
secreted free into the space and then be removed directly by the LRP, or they may be
sequestered into the space of Disse (Cooper, 1997). The confocal micrographs
showed rapid uptake of normal emulsion remnant particles into the hepatocytes in
control mice, reflecting normal plasma clearance patterns. A delayed entrance of
remnant particles into the space of Disse and hepatocytes was observed in the liver
sections of LDL-receptor-deficient mice, suggesting uptake by low affinity pathways.
This uptake has been proposed to be due to interactions between apo E and H S P G and
between apo B and H L (Cooper, 1997). Sequestered particles may then undergo
further modification and be transferred to endocytotic receptors, including the LRP,
for internalisation. The metabolism of modified emulsions by control and L D L -

234
receptor-deficient mice was significantly delayed, reflecting the utilisation of an
alternate, apo E-dependent mechanism for hepatic uptake. There was no fluorescence
observed at any time in apo E-deficient mice for normal or modified emulsions.
The studies suggest that remnant-like emulsions m a y offer a viable alternative
to quantify receptor uptake. Simultaneous injection of remnant-like emulsions did not
appear to alter hepatic uptake kinetics, and suggest that the mass of lipid injected was
not sufficient to saturate lipolysis or hepatic uptake mechanisms.
The pattern of emulsion uptake in genetically manipulated animal models
supports the original proposal that chylomicron remnants require apo E as a ligand for
hepatic uptake primarily via the LDL-receptor, holds at this point. In contrast, the
modified emulsion does not appear to interact with the apo E or the LDL-receptor,
which is reflected in the similarity in uptake in control, apo E-deficient and L D L -
receptor-deficient mice. These results confirm at a cellular level, earlier observations
in Chapter 3 that the modified emulsion is delayed due to the inability to interact with
the LDL-receptor and are instead taken up via low-affinity pathways. From the
results, it can be concluded that the normal and modified emulsions are taken up via
high and low affinity pathways, respectively. Hence the difference in clearance of the
two emulsions can be used as a measure of quantitating rapid high-affinity uptake in
vivo. Furthermore, there was essentially no uptake of either emulsion in apo E-
deficient mice, compared to the delayed uptake in LDL-receptor-deficient mice,
suggesting that the pattern of uptake in these animal models is sufficiently unique to
allow differentiation between the two deficiencies.

235
Chapter 6: The Effect of Retinyl
Esters on Clearance Kinetics of
Chylomicron-Like Emulsions In
Vivo
6.1 Introduction
It is difficult to recognise or measure chylomicrons in plasma, first because the
amount of chylomicrons is low in relation to other lipoproteins and secondly because
the composition of chylomicrons is not unique, except for the presence of apo B-48.
The rapid metabolism of chylomicrons adds to the difficulty of measurement, as
chylomicrons have the fastest turnover or clearance from plasma of all lipoproteins.
Mainly for this reason, increased plasma contents of cholesterol in the form of
chylomicron remnants may escape attention, unless there is a very gross abnormality
such as absence of, or defective, apo E (Redgrave, 1999).
The intravenous fat tolerance test (versus the oral fat load test) has been used
historically as an index of chylomicron clearance (Hallberg, 1965). However, this test
provides no information about remnant clearance because the injected emulsion
contains no marker for remnants and the emulsion contains no cholesterol, (Redgrave
et al, 1987), thus the test provides a measure of triglyceride clearance rather than that
of remnant particles. A direct approach to measuring remnant clearance in humans
involves the injection of chylomicron-like emulsions, which provides an estimate of
remnant residence time. Lipid emulsion particles similar in size and composition to
nascent chylomicrons are made from purified lipids and rapidly acquire the

236
apolipoproteins necessary for metabolism when injected in vivo. Chylomicron-like
emulsions are traced with labelled cholesteryl oleate and triolein and have been used
to model chylomicron metabolism in rats, rabbits and man (Redgrave et al, 1993),
(Maranhao et al, 1996), (Nakandakare et al, 1994), (Bowler et al, 1991), (Redgrave
and Zech, 1987), (Redgrave and Callow, 1990). However, utilisation of this procedure
as a diagnostic assay is limited by the necessity to use potentially hazardous
radioisotopes, which are not suitable for use in human subjects.
In recent years, a breath test has been developed which has enabled stable
isotopes to be used as markers to monitor chylomicron remnant particle clearance in
humans (Redgrave et al, 2001), (Watts et al, 2001). However, this technique does
not measure plasma clearance, which is of vast relevance with regard to arterial 1 "X
exposure. Moreover, the appearance of C O 2 in the breath is a reflection of fatty acid
metabolism, which is regulated by mitochondrial oxidase activity, levels of carnitine,
intracellular fatty acid transporters and oxidation/esterification homeostasis. Most
importantly though, is that the test does not allow for quantitation of receptor activity.
Clinical interest in the evaluation of retinyl esters has increased in recent
years, owing to the possible roles of these nutrients as markers for detecting
deficiencies in dietary cholesterol clearance, which may be important in reducing risk
of a number of diseases including coronary heart disease. As discussed in Chapter 1,
endogenously-derived lipoproteins labelled with retinyl palmitate have been widely
used to estimate rates of clearance of chylomicrons and their remnants in subjects
(Wilson et al, 1983), (Wilson et al, 1985a), (Wilson et al, 1985b), (Berr and Kern
Jnr, 1984), (Berr et al, 1985), (Berr et al, 1986), (Cortner et al, 1987), (Sprecher et
al, 1991), (Weintraub et al, 1992a), (Weintraub et al, 1987a), (Weintraub et al,
1987b), (Ruotolo et al, 1992), (Krasinski et al, 1990b), (Rubinsztein et al, 1990).
Specifically, retinyl esters have been used to demonstrate impaired chylomicron
metabolism in subjects with type III hyperlipidemia (Hazzard and Bierman, 1976),
coronary heart disease (Groot et al, 1991), (Simpson et al, 1990), familial
hypercholesterolemia ( M a m o et al, 1998a), (Smith et al, 1997) and visceral obesity
(Smith etal, 1999).
Nonetheless, the validity of the vitamin A fat loading test has been questioned
because the retinyl ester may shift to plasma lipoproteins of higher densities to
chylomicrons, at later time points (Krasinski et al, 1990b), (Cohn et al, 1993).
Chylomicron retinyl esters have been shown undergo minimal transfer and negligible

237
intravascular hydrolysis when autologous retinyl ester-rich chylomicrons are
reinfused into the circulation and the clearance curve of retinyl ester measured over
time (Berr and Kern Jnr, 1984), (Berr, 1992). However the infused 'chylomicrons'
may contain a heterogeneous mixture of lipoproteins from different origins, and
therefore not represent chylomicron remnant clearance per se. In addition,
postprandial elevations in retinyl palmitate in fractions of triglyceride-rich
lipoproteins do not always coincide with elevations in apo B-48 (Karpe et al, 1995).
This suggests that exogenous retinyl ester may not be incorporated within the particle
at the same rate as the dietary triacylglycerol and the endogenous apo B-48
(Lovegrove et al, 1999), or that the two methods mark different populations of
chylomicrons and their remnants (Krasinski et al, 1990b).
To examine the extent to which L P L hydrolyses retinyl esters, radiolabeled
retinyl esters were used as a label for core lipids of chylomicrons and their remnants
(Hultin et al, 1996). N o detectable hydrolysis of retinyl esters was found, despite
hydrolysis of 7 0 % of triglycerides in vitro. Other studies have also found that retinyl
esters are not removed during particle lipolysis (Karpe et al, 1995), (Cohn, 1994). A
study of postprandial metabolism in dogs by Melchoir et al. (Melchoir et al, 1981)
used artificial chylomicrons labelled with radioactive retinyl ester and cholesteryl
ester, and found that the plasma radioactive decay curves and hepatic uptake of
natural chylomicrons were similar for both radiolabels. Another study by Ross and
Zilversmit (Ross and Zilversmit, 1977) found that the rate of disappearance of
radiolabeled cholesteryl oleate became faster than radiolabeled retinyl ester 25 min
after intravenous infusion. However, the chylomicrons infused contained only 2 % of
radioactive free retinol compared with 1 5 % radiolabeled cholesterol, which
exchanges rapidly among plasma lipoproteins.
Others have established that retinyl ester transfer to other lipoproteins in
human plasma is minimal during postprandial lipemia or in vitro incubation (Berr and
Kern Jnr, 1984), (Berr et al, 1985), (Martins et al, 1991), (Zilversmit et al, 1982),
(Ross and Zilversmit, 1977). M a n y reports disputing the reliability of retinyl ester
show transfer to other lipoprotein classes at later time points only, including Type I
hyperlipidemies with protracted residence time for chylomicrons (Sprecher et al,
1991). More recent studies suggest that vitamin A is not subject to transfer between
lipoproteins and that its isolation at densities greater than 1.006 gm/ml represents a
generation of small dense postprandial remnants ( M a m o et al, 1998a), (Peel et al,

238
1993). M a m o ( M a m o et al, 1998b) predicts that in situations where high affinity
uptake is compromised there would be increased appearance of remnants in more
dense plasma fractions as a consequence of increased interaction with lipolytic
enzymes. Together, these studies suggest that retinyl esters provide a suitable label for
core lipids and are not hydrolysed or transferred to other lipoproteins before the
chylomicrons or their remnants are removed from the circulation.
The aim of this project was to establish a non-radioisotopic marker for
chylomicrons and their remnants, to allow monitoring and quantitation of high affinity
uptake of particle clearance in humans. The approach was to assess whether
radioisotopes could be replaced by introducing retinyl esters into chylomicron-like
emulsions, without compromising the validity of the procedure. The proposed
technique required the two emulsion types to be labelled with retinyl esters (using a
different retinyl fatty acid for each). Retinyl ester concentration in plasma following
simultaneous injection of the emulsions will then used to trace particle uptake.
Quantitation of plasma retinyl esters is readily achievable in a clinical environment,
using a dedicated H P L C system.
Vitamin A (retinol) is present in food mainly as its palmitate ester, which like
cholesterol, is esterified in the intestinal mucosa to long-chain fatty acids by vitamin
A ester hydrolase of the pancreatic juice (Olson, 1969). Under normal conditions
vitamin A is absorbed almost exclusively as its free alcohol, retinol (Mahadevan et
al, 1963). Within intestinal cells retinol is reesterified mainly to its palmitate,
incorporated selectively into the hydrophobic core of chylomicrons in the mucosa,
and secreted into the intestinal lymph (Cortner et al, 1987), (Huang and Goodman,
1965), (Goodman, 1980). As chylomicrons undergo hydrolysis, they retain the retinyl
esters in their core (Goodman et al, 1965), (Goodman et al, 1966), thus acting as a
marker for chylomicron remnant particle uptake. The retinyl esters are relatively non-
exchangeable compounds of the chylomicrons (Ross and Zilversmit, 1977),
(Zilversmit et al, 1982) and their remnants (Melchoir et al, 1981). Under normal
conditions there is little extrahepatic removal of labelled chylomicron remnants
(Cooper, 1985), (Goodman, 1980), (Goodman et al, 1965), therefore chylomicron
remnants and retinyl esters are removed from plasma by the liver, primarily by the
LDL-receptor.
Hepatic uptake is followed by lysosomal hydrolysis of the cholesteryl esters
and retinyl esters to unesterified cholesterol and retinol, respectively. Dietary retinol

239
cannot be recycled into V L D L , nor can it be synthesised de novo, but is stored in the
liver or secreted into the plasma in complex with R B P for transport to peripheral
target tissues (Kanai et al, 1968), (Ross and Zilversmit, 1977), (Thompson et al,
1983), (Berr and Kern Jnr, 1984), (Goodman, 1980), (Blomhoff et al, 1988). Most of
the absorbed dietary vitamin A is delivered to hepatic parenchymal cells when
chylomicron remnants are metabolised by the liver, and transferred primarily to
stellate cells on R B P . Stellate cells are able to control the storage and mobilisation of
retinol, thus ensuring constant plasma retinol levels in spite of normal fluctuations in
daily vitamin A intake (Blomhoff et al, 1992), (Blomhoff et al, 1984), (Blomhoff,
1994), (Wingerath et al, 1997), (Hagen et al, 1999). As a result, there is virtually no
background level of plasma retinyl palmitate in fasting humans (Berr and Kern Jnr,
1984), or rabbits (Beaumont and Assadollahi, 1990). Retinyl esters were chosen as an
alternative to radioactive markers as they are safe, and have previously been used in
vitamin A fat load tests with minimal transfer to other lipoprotein fractions. The
proposed method involves intravenous injection of emulsion, which is expected to
clear from plasma rapidly.
The proposed method involves the development of retinyl esters as tracers for
two types of chylomicron-like emulsion particles. Hence two retinyl esters were
required for simultaneous injection of emulsion and subsequent detection. Retention
times of retinyl esters increase with increasing acyl chain length and degree of
saturation (Futterman and Andrews, 1964b). Palmitic and stearic acids were the fatty
acids of choice because their relative retention times are significantly divergent (Furr,
1990). Retinyl myristate was also investigated due to the degree of saturation, low
solubility in water and reduced triglyceride mobilisation. Retinyl stearate is 2 carbons
longer and retinyl myristate is 2 carbons shorter, than retinyl palmitate (18:0, 14:0 and
16:0, respectively). The esters of lipids such as glycerol, retinol and cholesterol
undergo slow changes in physical properties with respect to acyl chain length. For
example, thermal data for simple triacylglycerols (P form) indicates melting points of
57°C, 66.4°C and 73.1°C for trimyristin, tripalmitin and tristearin, respectively
(Small, 1986). Retinyl stearate was expected to be more stable under most
physiological conditions and less likely to disintegrate in preparation, however all
vitamin A esters are more stable to oxidation than retinol. Retinyl palmitate is
available commercially, however retinyl stearate and myristate were synthesised from

240
retinyl acetate by transesterification with the appropriate fatty acid chloride under
reduced pressure (Huang and Goodman, 1965).
The current study was concerned with establishing that the incorporation of
retinyl esters did not alter emulsion kinetics in animal models. To assess this,
clearance data for normal and modified emulsions, with and without retinyl esters,
was compared. Rats were selected as one model for this study because much of our
emulsion data that has formed the basis of this study was derived from rats (Redgrave
and Callow, 1990), (Redgrave et al, 1993), (Redgrave and Zech, 1987). However, the
two-emulsion procedure was first used to quantitate net receptor activity in rabbits
(Bowler et al, 1991), because like man, rabbits have active C E T P activity. Therefore,
comparative studies were carried out in rabbits as the C E T P activity may cause the
exchange or loss of the retinyl esters and cholesteryl esters in vivo, and it was
appropriate to select a model that may more closely resemble lipoprotein metabolism
in man.
Lipid emulsions were prepared from purified triolein, cholesteryl oleate,
cholesterol and phosphatidylcholine, and retinyl esters. Like cholesteryl esters, retinyl
esters are hydrophobic and will distribute within the core of the particles. Equal
masses of cholesteryl ester were substituted with retinyl esters to maintain the relative
proportion of core to surface components, and avoid disturbing the emulsion kinetics
in vivo. Based on the sensitivity of retinyl ester analysis by H P L C , w e estimated that
in a 70 kg normolipidemic individual, a mass of 2.5 m g retinyl esters would need to
be injected. With an estimated maximum dose of 225 m g triglyceride ( 5 % of
approximate plasma pool), 2 5 % of the cholesteryl ester mass will require substitution
with retinyl ester. For the purpose of these studies in animal models, 2 5 % of
cholesteryl ester mass was calculated to be sufficient for detection, however
substitution of up to 5 0 % of cholesteryl ester mass was assessed.

241
6.2 Special Methods
6.2.1 Animals
Male albino Wistar rats, weighing between 250 to 400 gm were obtained from the
Animal Resources Centre, Murdoch, Western Australia. Male N e w Zealand White
( N Z W ) and semi-lop (NZW/ - cross) rabbits, weighing between 1.8 to 3.5 kg, were
obtained from the Animal Resources Centre, Murdoch, Western Australia. Each
rabbit was used in a crossover design to reduce intra-individual variability.
6.2.2 Preparation of Retinyl Esters
Retinyl stearate and myristate were synthesised by transesterification by reaction of
retinol with the corresponding fatty acyl chlorides under reduced pressure, according
to the method of Huang and Goodman (Huang and Goodman, 1965). Briefly, 0.3 ml
of pyridine and 1.4 mmole of acyl chloride were added to 1.2 mmole of retinol. The
flask was flushed with nitrogen, briefly warmed to 50-60°C, and intermittently shaken
for 1 to 2 hr in the dark. The contents of the flask were then extracted with n-hexane
(50 ml), and the hexane solution was washed with 50 ml each of: 0.1 N N a O H in 5 0 %
ethanol, 0.1 N HC1, 0.03 N N a O H in 5 0 % ethanol, and water. After evaporation of the
hexane the oily residue was chromatographed on a column of alumina grade in. The
esters were pure as determined by the close agreement between the weight of the
product and the weight as calculated from the optical density at 328 nm.
Aliquots of the total lipid extracts were chromatographed on columns of 1 cm
diameter containing alumina of activity grade III by the method described by Olson
(Olson, 1961). The maximum load was 10 m g of lipid/gm of alumina. Fractions 1-5
were eluted from each column. The order of elution and volume of elutents/5 g m of
alumina were P-carotene, retinyl esters, retinal, retinol, and more polar compounds.
Retinoic acid was either eluted from the column terminally with methanol-25% acetic
acid, 3:1 (Fraction 6), or was extracted from an aliquot of the total lipid extract with
0.1 N K O H in 5 0 % ethanol.
Ultra-violet light inactivates vitamin A and its solutions. Therefore, all
procedures with retinol and its ester were performed in dim light and samples were
kept under an atmosphere of argon or nitrogen. The yields of retinyl esters varied

242
from 60-90% in different samples. Following synthesis, the presence of retinyl esters
was confirmed using flash chromatography on silica gel with petrol and ethyl acetate
as elutents. Proton and carbon nuclear magnetic resonance spectroscopy was used to
confirm the structure and purity of the retinyl esters. The esters were pure as
determined by the close agreement between the weight of the product and the weight
was calculated from the optical density at 325 nm. Retinyl ester purity was also
verified using T L C to identify retinyl esters as a single band. T L C plates were silica
on aluminium backing (Merck) were run in a solvent system of 95:5 (petrol (hexane):
ether, v/v), then developed by dipping in 95:5 (ethanol: sulfuric acid), and heating
with a hotplate or heatgun.
Prior to experimental use, the purity of all retinyl esters was re-confirmed by
examination of their ultraviolet absorption spectra and TLC. Each retinyl ester was
assayed on a Beckman D U 6 5 0 spectrophotometer, and the weight was calculated
from the optical density at 325 nm.
6.2.3 Preparation of Normal Chylomicron-Like Emulsions
Lipid emulsions resembling chylomicrons in composition and size were prepared as
described in Chapter 2. The following changes were implemented for emulsions
containing retinyl ester. For emulsions containing 2 5 % and 5 0 % retinyl ester, 2.25 m g
and 1.5 m g cholesteryl oleate, respectively was added to the emulsion mixture. To
substitute the cholesteryl oleate component, 0.75 m g or 1.5 m g retinyl palmitate
(Sigma Chemical Co., Australia), stearate or myristate, respectively, was suspended in
chloroform: methanol (2:1, v/v) and added to the mixtures. The emulsions were
labelled with radioactive cholesteryl[14C]oleate and [3H]triolein (Amersham, Surry
Hills, N S W , Aust).
6.2.4 Preparation of Modified Chylomicron-Like Emulsions
Modified chylomicron-like emulsion particles were prepared as described in Chapter
2, with the following changes. In emulsions containing retinyl ester, 0.75 m g or 1.5
m g retinyl palmitate (Sigma Chemical Co., Australia), stearate or myristate was
suspended in chloroform: methanol (2:1, v/v) and added to the lipid mixture,
substituting 2 5 % or 5 0 % of the cholesteryl oleate component, respectively. The

243
modified chylomicron-like lipid emulsions contained a different isotopic form of
cholesteryl oleate and triolein (cholesteryl[3H]oleate and [14C]triolein) so that
clearance from plasma of the two emulsion types could be distinguished.
6.2.5 Emulsion Clearance Studies in Rats
For clearance studies, emulsions containing < 2.5 mg triglyceride in a volume of 0.35-
4.0 ml were injected as a bolus into the bloodstream of non-fasted conscious rats
prepared with cannulas in the left common carotid artery and the left jugular vein. The
effect of retinyl ester on the clearance of normal and modified emulsions was assessed
on consecutive days, however control and experimental clearance studies for each
emulsion type were completed on the same day. Blood samples were taken and
processed as described in Chapter 2.
6.2.6 Emulsion Clearance Studies in Rabbits
Emulsions containing < 2.5 mg of triglyceride in a volume of 0.4-0.5 ml were injected
as a bolus into the bloodstream of non-fasted conscious rabbits via the marginal ear
vein. The effect of retinyl ester on the clearance of normal and modified emulsions
was assessed on consecutive days, however control and experimental clearance
studies for each emulsion type were completed on the same day. Details are described
in Chapter 2.
6.2.7 Determination of Radioactivity
Plasma was separated by centrifugation in a microcentrifuge (BHG Hermle Z 230M).
The plasma collected was then measured for radioactivity without extraction, by
adding 5 ml of Emulsifier-Safe™ (Packard) to plasma and counting in dual-label
mode with auto quench correction in a Beckman LS 3800 Liquid Scintillation counter
(Beckman Instruments Inc, U S A ) . Radioactivity in organs after lipid extraction was
also determined. Plasma samples of 150 ul and 400 ul were measured for radioactivity
in rats and rabbits, respectively.

244
6.2.8 O r g a n Extraction in Rats
After completion of clearance studies in rats, 20 mg sodium pentobarbitone
(Nembutal®) was injected and the liver and spleen were removed and washed
thoroughly in ice-cold 0.15 M NaCl. Liver and spleen were removed and extracted as
described in Chapter 2.
6.2.9 Calculations
Clearances of emulsion triglyceride and cholesteryl ester were determined from the
decline in plasma radioactivity, for normal emulsions and modified emulsions
(described in Chapter 2). To assess the effect of incorporating retinyl esters into
emulsions, the A A C data determined from emulsions with and without retinyl esters
was compared.
6.2.10 Statistical Analysis
Statistical analysis was performed as described in Chapter 2.
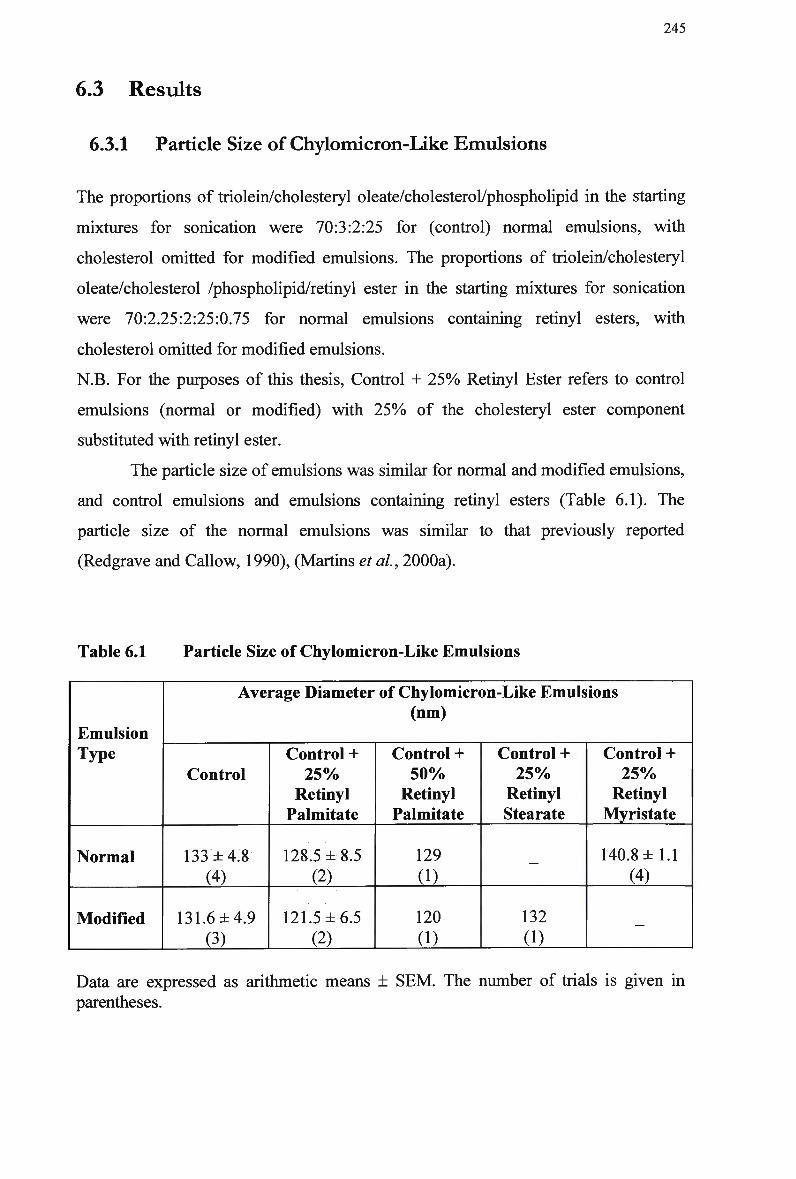
245
6.3 Results
6.3.1 Particle Size of Chylomicron-Like Emulsions
The proportions of triolein/cholesteryl oleate/cholesterol/phospholipid in the starting
mixtures for sonication were 70:3:2:25 for (control) normal emulsions, with
cholesterol omitted for modified emulsions. The proportions of triolein/cholesteryl
oleate/cholesterol /phospholipid/retinyl ester in the starting mixtures for sonication
were 70:2.25:2:25:0.75 for normal emulsions containing retinyl esters, with
cholesterol omitted for modified emulsions.
N.B. For the purposes of this thesis, Control + 25% Retinyl Ester refers to control
emulsions (normal or modified) with 25% of the cholesteryl ester component
substituted with retinyl ester.
The particle size of emulsions was similar for normal and modified emulsions,
and control emulsions and emulsions containing retinyl esters (Table 6.1). The
particle size of the normal emulsions was similar to that previously reported
(Redgrave and Callow, 1990), (Martins et al, 2000a).
Table 6.1 Particle Size of Chylomicron-Like Emulsions
Emulsion Type
Normal
Modified
Average Diameter of Chylomicron-Like Emulsions (nm)
Control
133 ±4.8
(4)
131.6±4.9
(3)
Control + 25 %
Retinyl Palmitate
128.5 ±8.5
(2)
121.5 ±6.5
(2)
Control + 5 0 %
Retinyl Palmitate
129
(1)
120
(1)
Control + 25 % Retinyl Stearate
—
132
(D
Control + 25 %
Retinyl Myristate
140.8 ±1.1
(4)
—
Data are expressed as arithmetic means ± SEM. The number of trials is given in
parentheses.

246
6.3.2 Effect of Retinyl Palmitate Incorporation o n Plasma
Clearance of Chylomicron-Like Emulsions in Rats
6.3.2.1 Clearance of lipids in normal chylomicron-like emulsions
The clearance of control (normal and modified) emulsions in rats was compared with
emulsions with 2 5 % and 5 0 % of the cholesteryl ester mass substituted with retinyl
palmitate. To reduce variability, control and experimental clearance studies were
completed on the same day. Normal emulsions hydrolysed quickly, with 96%, 9 7 %
and 9 5 % of particle triolein disappearing from the plasma by 30 min after injection,
for control emulsions and emulsions containing 2 5 % and 5 0 % retinyl palmitate,
respectively (data not shown). The total amount of triglyceride cleared from plasma
(AAC) was similar for all normal emulsions.
The disappearance of triglyceride was followed by a rapid clearance of
particle remnants, with approx. 93%, 9 0 % and 9 2 % of labelled cholesteryl oleate
removed by 30 min for control emulsions and emulsions containing 2 5 % and 5 0 %
retinyl palmitate, respectively. Figure 6.1 compares the clearance of cholesteryl oleate
from plasma following injection of normal emulsion in rats. The clearance of normal
emulsion cholesteryl oleate from plasma of rats was similar for emulsions with or
without retinyl palmitate incorporated, with respect to the amount of cholesteryl ester
cleared from plasma (Table 6.2).
6.3.2.2 Clearance of lipids in modified chylomicron-like emulsions
Similar to the normal emulsion, mean AAC for modified emulsions was greater for
triglyceride than cholesteryl ester in rats. The removal of emulsion remnant particles
was delayed compared with removal of normal emulsion, consistent with a defect in
remnant clearance, after normal depletion of emulsion triolein by the action of LPL.
The modified emulsion hydrolysed rapidly, with 95%, 9 2 % , and 9 6 % of particle
triolein disappearing from the plasma by 30 min after injection, for control emulsions
and emulsions containing 2 5 % and 5 0 % retinyl palmitate, respectively (data not
shown). The amount of triglyceride cleared from plasma (AAC) was similar for
control emulsions or emulsions with 2 5 % and 5 0 % retinyl palmitate incorporated
(Table 6.2).

247
Figure 6.2 compares the clearance of modified emulsion cholesteryl oleate
from plasma in rats. Approx. 4 6 % , 5 4 % and 6 6 % of modified emulsion cholesteryl
oleate was removed by 30 min for control emulsions, and emulsions containing 2 5 %
and 5 0 % retinyl palmitate, respectively. The amount of cholesteryl ester cleared from
plasma of rats was similar for control emulsions and emulsions with 2 5 % retinyl
palmitate incorporated. However, the emulsion cholesteryl ester in modified
emulsions with 5 0 % retinyl palmitate incorporated cleared to a larger extent
compared with control and emulsions containing 2 5 % retinyl palmitate (Table 6.2).
For each emulsion, the disappearance of triglyceride preceded particle
clearance, indicating that incorporation of retinyl palmitate into emulsion particles did
not impair lipolysis or remnant formation.
6.3.2.3 High affinity uptake
Normal emulsions are cleared by non-specific mechanisms, similar to modified
emulsions, but they can also be cleared via receptor mechanisms. The difference
between the clearances of the two emulsions is indicative of high-affinity clearance.
High affinity uptake was calculated for control emulsions, and emulsions containing
2 5 % and 5 0 % retinyl palmitate, to determine if the incorporation of retinyl palmitate
into emulsions had any net effect on the amount of emulsion remnant cholesteryl
oleate cleared by receptor mechanisms.
The A A C data shows that while high affinity uptake for emulsions containing
2 5 % retinyl palmitate was similar compared with control emulsions, high affinity
uptake for emulsions containing 5 0 % retinyl palmitate was significantly less than
control emulsions (Table 6.2). This appears to be a combination of a greater amount
of modified emulsion cholesteryl oleate cleared from plasma, and a reduction in the
amount of normal emulsion cholesteryl oleate cleared from plasma, compared with
control emulsions.
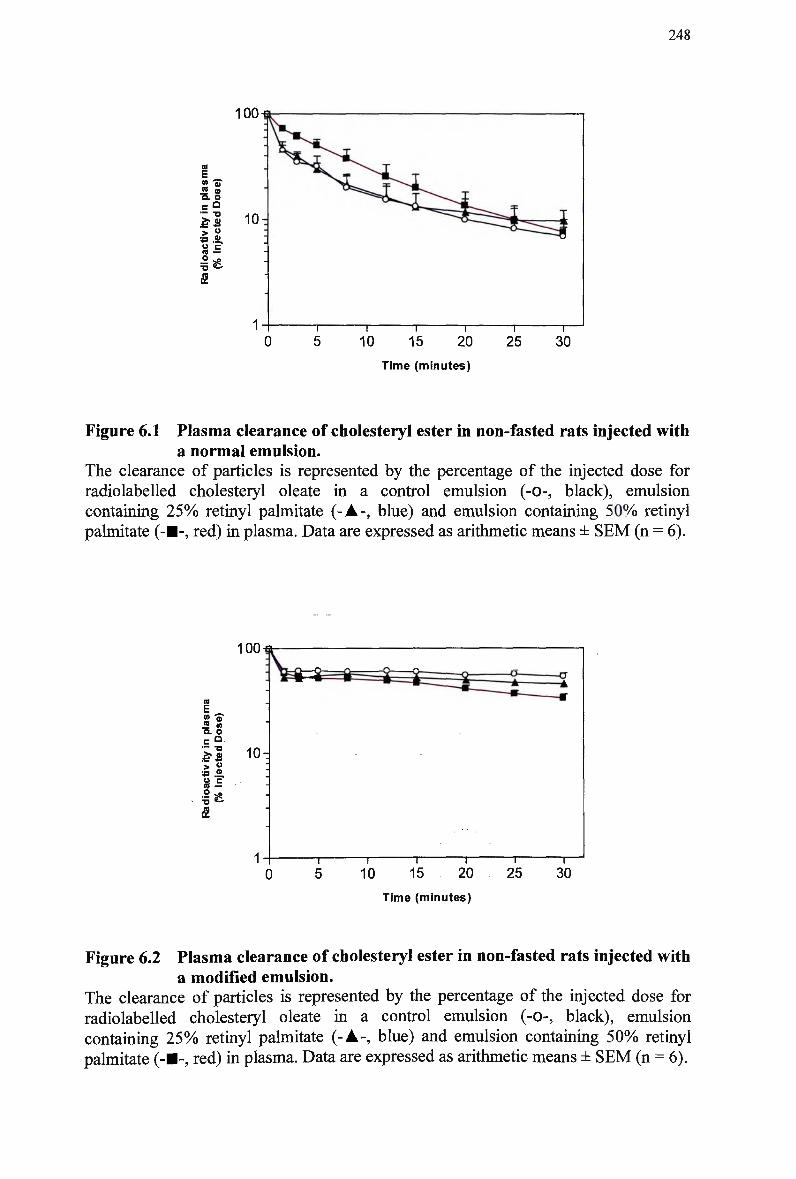
248
100
a
£ ™ to Q. O — •o
5 .2.
o ^o
Time (minutes)
Figure 6.1 Plasma clearance of cholesteryl ester in non-fasted rats injected with a normal emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate in a control emulsion (-0-, black), emulsion containing 2 5 % retinyl palmitate (-A-, blue) and emulsion containing 5 0 % retinyl palmitate (-•-, red) in plasma. Data are expressed as arithmetic means ± S E M (n = 6).
1 0 0 - 6
<S <» 3. o
c° — -a
> ° S.E
Time (minutes)
Figure 6.2 Plasma clearance of cholesteryl ester in non-fasted rats injected with a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate in a control emulsion (-0-, black), emulsion containing 2 5 % retinyl palmitate (-A-, blue) and emulsion containing 5 0 % retinyl palmitate (-•-, red) in plasma. Data are expressed as arithmetic means ± S E M (n = 6).
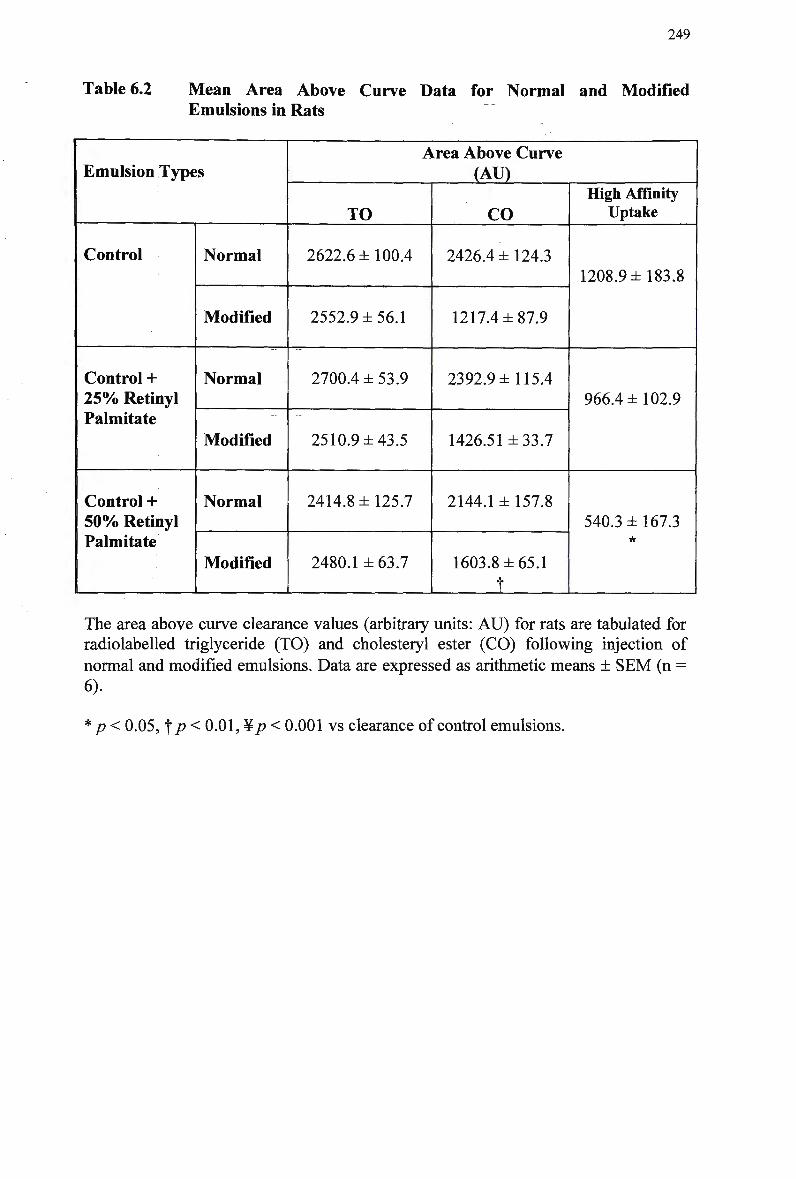
249
Table 6.2 Mean Area Above Curve Data for Normal and Modified Emulsions in Rats
Emulsion Types
Control
Control + 2 5 % Retinyl Palmitate
Control + 5 0 % Retinyl Palmitate
Normal
Modified
Normal
Modified
Normal
Modified
Area Above Curve (AU)
TO
2622.6 ±100.4
2552.9 ±56.1
2700.4 ±53.9
2510.9 ±43.5
2414.8 ±125.7
2480.1 ± 63.7
CO
2426.4 ± 124.3
1217.4 ±87.9
2392.9 ±115.4
1426.51 ±33.7
2144.1 ± 157.8
1603.8 ±65.1
f
High Affinity Uptake
1208.9 ±183.8
966.4 ±102.9
540.3 ±167.3 •k
The area above curve clearance values (arbitrary units: A U ) for rats are tabulated for radiolabeled triglyceride (TO) and cholesteryl ester (CO) following injection of
normal and modified emulsions. Data are expressed as arithmetic means ± S E M (n = 6).
p < 0.05, f p < 0.01, ¥ p < 0.001 vs clearance of control emulsions.

250
6.3.3 Organ Uptake
Normal and modified chylomicron-like emulsions were injected into unanaesthetised
rats. To examine the effect of incorporating retinyl esters into emulsions on organ
uptake patterns, organ uptake of control emulsions and emulsions with 2 5 % and 5 0 %
retinyl palmitate incorporated were compared. Liver uptake was measured at 30 min
post-injection. Splanchnic uptake was also monitored to exclude significant uptake by
the reticuloendothelial system. The organ uptake values for normal and modified
emulsions are presented in Table 6.3, as a percent of injected dose.
Hepatic recoveries of triglyceride of control emulsions (normal and modified)
were similar to the uptake of emulsions with 2 5 % and 5 0 % retinyl palmitate
incorporated. Recovery of cholesteryl oleate radiolabel in the liver was always less
for modified emulsions compared with normal emulsions (up to 4 times), but the
uptake of emulsions containing 2 5 % retinyl palmitate did not differ from control
emulsions for either emulsion type (Figures 6.3 and 6.4). The hepatic uptake of
normal and modified emulsion cholesteryl oleate containing 5 0 % retinyl palmitate
was higher than for control emulsions and emulsions containing 2 5 % retinyl
palmitate. The uptake of modified emulsion cholesteryl oleate was significantly
higher for emulsions containing 5 0 % retinyl palmitate, compared with control
emulsions.
Less than 1 % of triglyceride and cholesteryl ester radioactivities were
recovered in the spleen and the uptake of modified emulsion radioactivities by the
spleen was higher compared with normal emulsion uptake (Figures 6.5 and 6.6). The
splanchnic uptake of normal emulsion containing 5 0 % retinyl palmitate was higher
for both lipid radioactivities, compared with control emulsion uptake, and the uptake
of triolein was significantly higher compared with control emulsions.
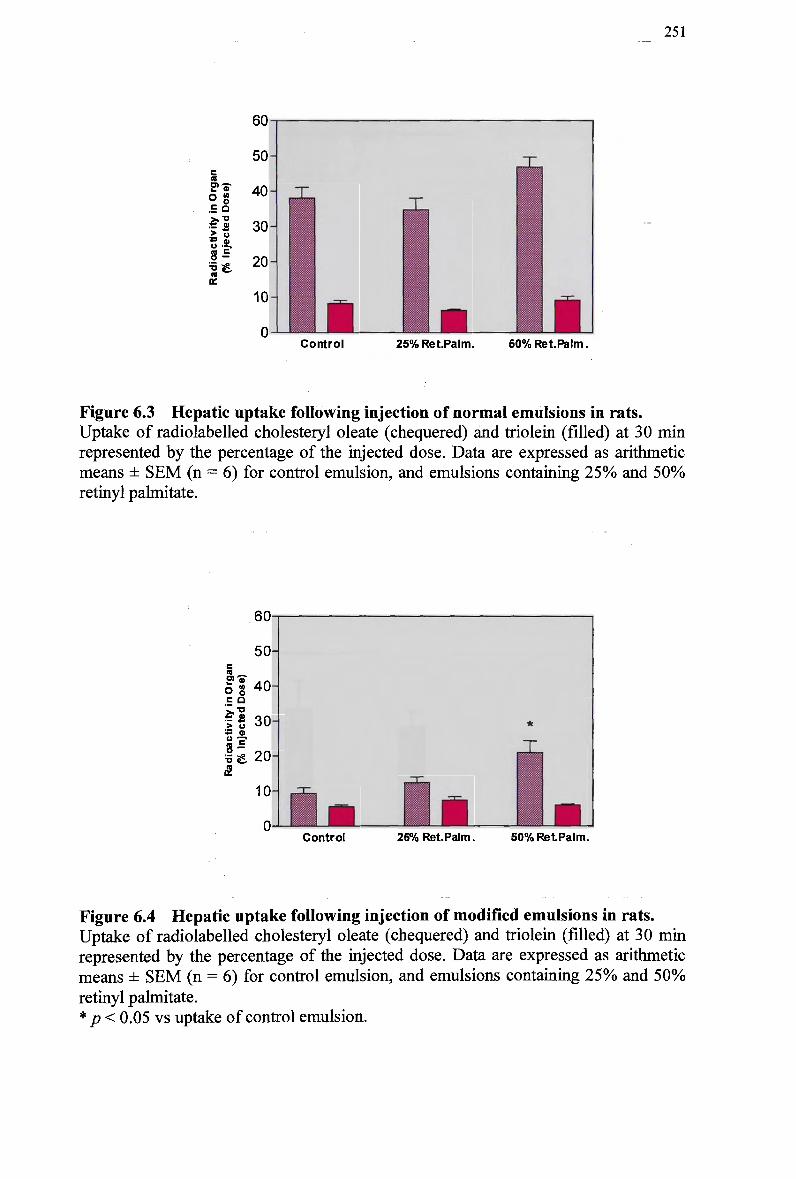
251
c « w-S-O g -H °
a:
60
50 H
40
30
20-
10-
0 a Control 25% RetPalm. S0%Ret.Palm.
Figure 6.3 Hepatic uptake following injection of normal emulsions in rats. Uptake of radiolabeled cholesteryl oleate (chequered) and triolein (filled) at 30 min represented by the percentage of the injected dose. Data are expressed as arithmetic means ± S E M (n = 6) for control emulsion, and emulsions containing 2 5 % and 5 0 % retinyl palmitate.
60
50
If 40
ll 30-IB 0
Ii. 20 &
10-
J L Control 25% Ret.Palm. 50% RetPalm.
Figure 6.4 Hepatic uptake following injection of modified emulsions in rats. Uptake of radiolabeled cholesteryl oleate (chequered) and triolein (filled) at 30 min represented by the percentage of the injected dose. Data are expressed as arithmetic means ± S E M (n = 6) for control emulsion, and emulsions containing 2 5 % and 5 0 %
retinyl palmitate. * p < 0.05 vs uptake of control emulsion.
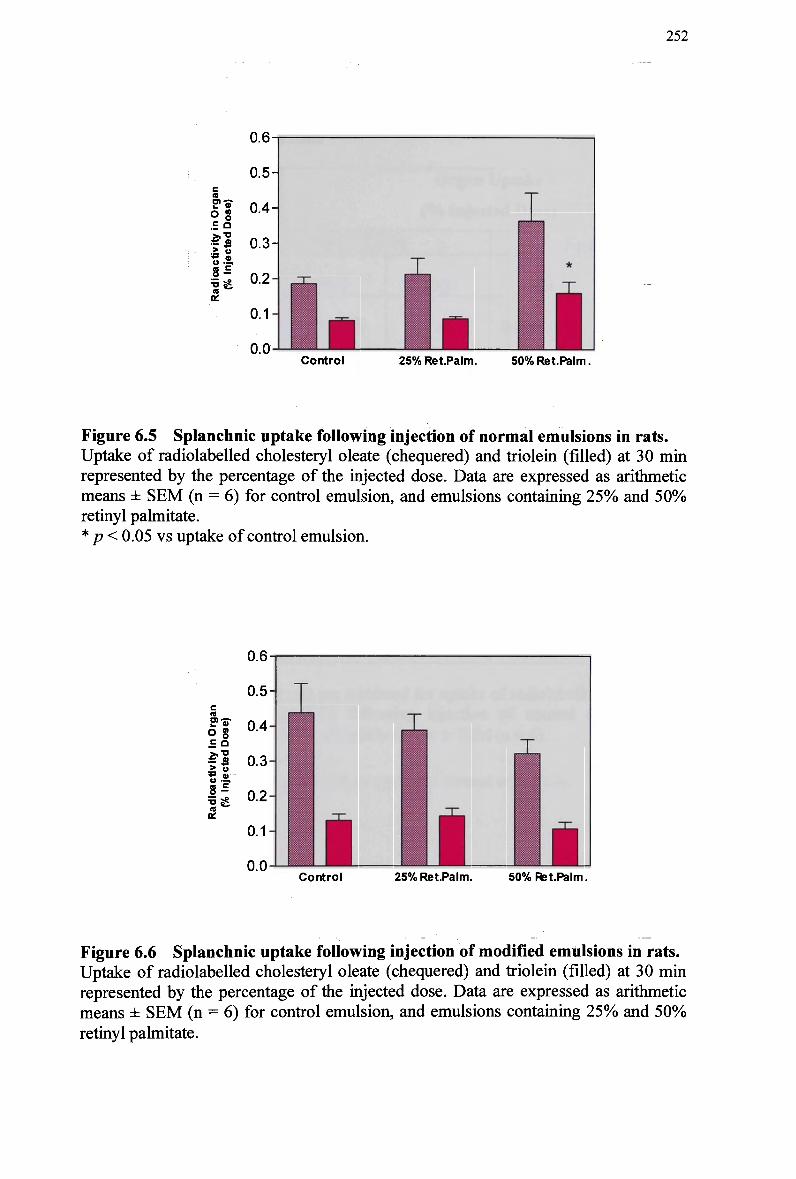
252
CO
ff-sr og
o •=.
1S£
a:
0.6-
0.5
0.4
0.3-
0.2-
0.1-
O.O-L Control 25% Ret.Palm. 50% Ret.Palm.
Figure 6.5 Splanchnic uptake following injection of normal emulsions in rats. Uptake of radiolabeled cholesteryl oleate (chequered) and triolein (filled) at 30 min represented by the percentage of the injected dose. Data are expressed as arithmetic means ± S E M (n = 6) for control emulsion, and emulsions containing 2 5 % and 5 0 % retinyl palmitate. * p < 0.05 vs uptake of control emulsion.
n
E"5T Og .E a •5 °
8^
5* cc
Control 25% RetPalm. 50% Ret.Palm.
Figure 6.6 Splanchnic uptake following injection of modified emulsions in rats. Uptake of radiolabeled cholesteryl oleate (chequered) and triolein (filled) at 30 min represented by the percentage of the injected dose. Data are expressed as arithmetic means ± S E M (n = 6) for control emulsion, and emulsions containing 2 5 % and 5 0 %
retinyl palmitate.
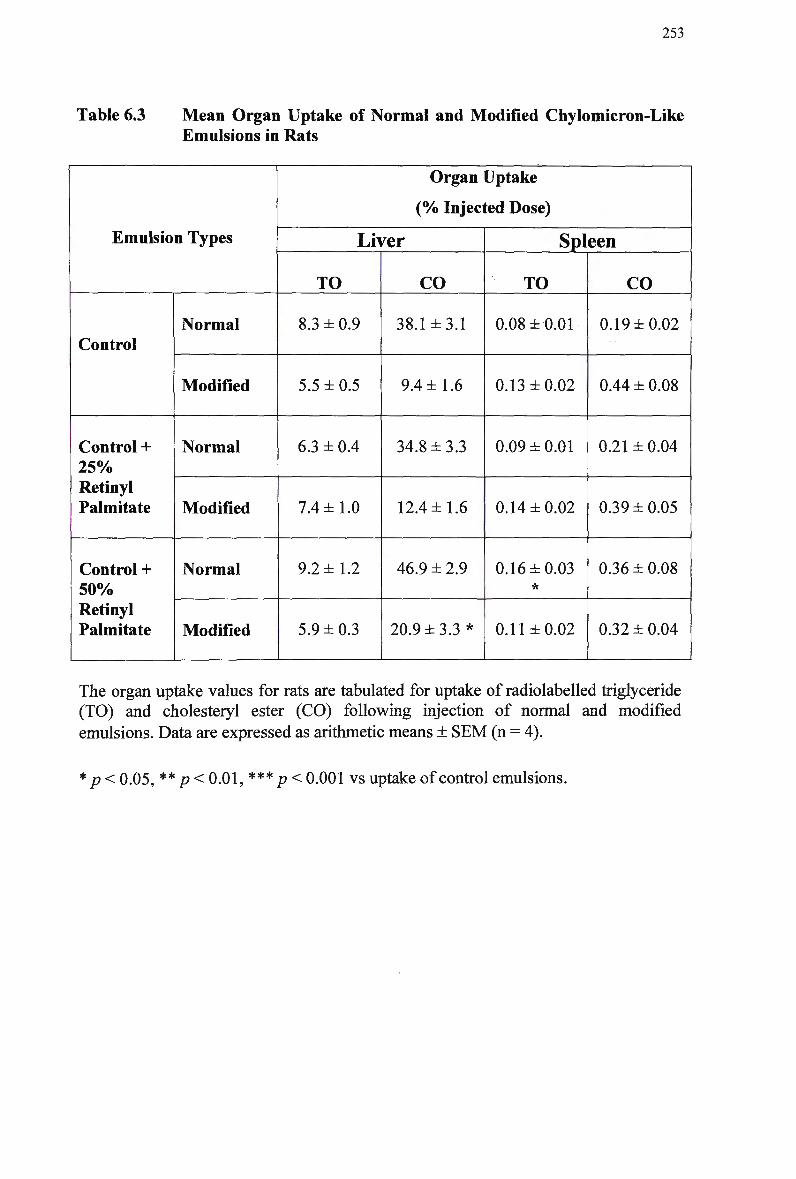
253
Table 6.3 Mean Organ Uptake of Normal and Modified Chylomicron-Like Emulsions in Rats
Emulsion Types
Control
Control + 2 5 % Retinyl Palmitate
Control + 5 0 % Retinyl Palmitate
Normal
Modified
Normal
Modified
Normal
Modified
Organ Uptake
(% Injected Dose)
Lii
TO
8.3 ±0.9
5.5 ±0.5
6.3 ± 0.4
7.4 ±1.0
9.2 ±1.2
5.9 ±0.3
fer
CO
38.1 ±3.1
9.4 ±1.6
34.8 ±3.3
12.4 ±1.6
46.9 ± 2.9
20.9 ±3.3*
Spleen
TO
0.08 ± 0.01
0.13 ±0.02
0.09 ± 0.01
0.14 ±0.02
0.16 ±0.03 *
0.11 ±0.02
CO
0.19 ±0.02
0.44 ± 0.08
0.21 ± 0.04
0.39 ±0.05
0.36 ±0.08
0.32 ±0.04
The organ uptake values for rats are tabulated for uptake of radiolabeled triglyceride (TO) and cholesteryl ester (CO) following injection of normal and modified
emulsions. Data are expressed as arithmetic means ± SEM (n = 4).
* p < 0.05, ** p < 0.01, *** p < 0.001 vs uptake of control emulsions.

254
6.3.4 Effect of Retinyl Palmitate Incorporation on Plasma
Clearance of Chylomicron-Like Emulsions in Rabbits
6.3.4.1 Clearance of lipids in normal chylomicron-like emulsions
The clearance of control (normal and modified) emulsions in rabbits was compared
with emulsions with 2 5 % of the cholesteryl ester mass substituted with retinyl
palmitate. This amount of emulsion retinyl ester should be sufficient for detection in
plasma by H P L C in future experiments. The same animals were re-used for all
experiments, thus acting as controls for both emulsion types and incorporation of
retinyl esters. Control and experimental clearance studies were completed on the same
day, to reduce variability.
The clearance profiles of normal emulsions in rabbits are presented in Table
6.4. Normal emulsions hydrolysed quickly, with 9 5 % of particle triolein disappearing
from the plasma by 15 min after injection, for control emulsions and emulsions
containing 2 5 % retinyl palmitate (data not shown). The amount of normal emulsion
triglyceride cleared from plasma (AAC) was similar for control emulsions and
emulsions with 2 5 % retinyl palmitate incorporated.
The disappearance of triglyceride from plasma was followed by a rapid
clearance of particle remnants, with approximately 9 5 % of labelled cholesteryl oleate
removed by 30 min for control emulsions and emulsions containing 2 5 % retinyl
palmitate (Figure 6.7). The clearance of normal emulsion cholesteryl oleate from
plasma in rabbits was similar for emulsions with or without retinyl palmitate
incorporated, with respect to amount of cholesteryl ester cleared from plasma.
6.3.4.2 Clearance of lipids in modified chylomicron-like emulsions
The clearance profiles for modified emulsions within rabbits are presented in Table
6.4. Similar to the normal emulsion, mean A A C was greater for triglyceride than
cholesteryl ester in rabbits. The modified emulsion also hydrolysed rapidly, with
approx. 9 0 % of particle triolein disappearing from plasma by 40 min, compared with
9 5 % triolein clearance for normal emulsion. Consistent with previous findings
(Redgrave et al, 1987), clearance of modified emulsion cholesteryl oleate was
impaired, consistent with a defect in remnant clearance, after normal depletion of
emulsion triolein by the action of LPL. For each emulsion, the disappearance of

255
triglyceride preceded particle clearance, indicating that incorporation of retinyl
palmitate into emulsion particles did not impair lipolysis or remnant formation.
Approx. 9 0 % of modified emulsion triolein disappeared from the plasma by
25 and 40 min after injection, for control emulsions and emulsions containing 2 5 %
retinyl palmitate, respectively (data not shown). The amount of triglyceride cleared
from plasma ( A A C ) was similar for control emulsions or emulsions with 2 5 % retinyl
palmitate incorporated.
Figure 6.8 compares the clearance of modified emulsion cholesteryl oleate
from plasma in rabbits. Approx. 8 9 % of labelled cholesteryl oleate removed by 40
min for control emulsions and emulsions containing 2 5 % retinyl palmitate. The
amount of cholesteryl ester cleared from plasma of rabbits was similar for control
emulsions and emulsions with 2 5 % retinyl palmitate incorporated.
6.3.4.3 High affinity uptake
Total uptake of chylomicron remnants via high affinity mechanisms (AAC data), was
calculated for control emulsions, and emulsions containing 2 5 % retinyl palmitate, to
determine whether the incorporation of retinyl palmitate into emulsions had a net
effect on the amount of emulsion remnant cholesteryl oleate taken up by receptor
mechanisms (Table 6.4).
High affinity uptake as calculated for A A C values was similar for control
emulsions, and emulsions with 2 5 % retinyl palmitate incorporated. Hence the overall
metabolism of normal and modified emulsion triglyceride and cholesteryl oleate in
rabbits was not altered by the incorporation of 2 5 % retinyl palmitate.
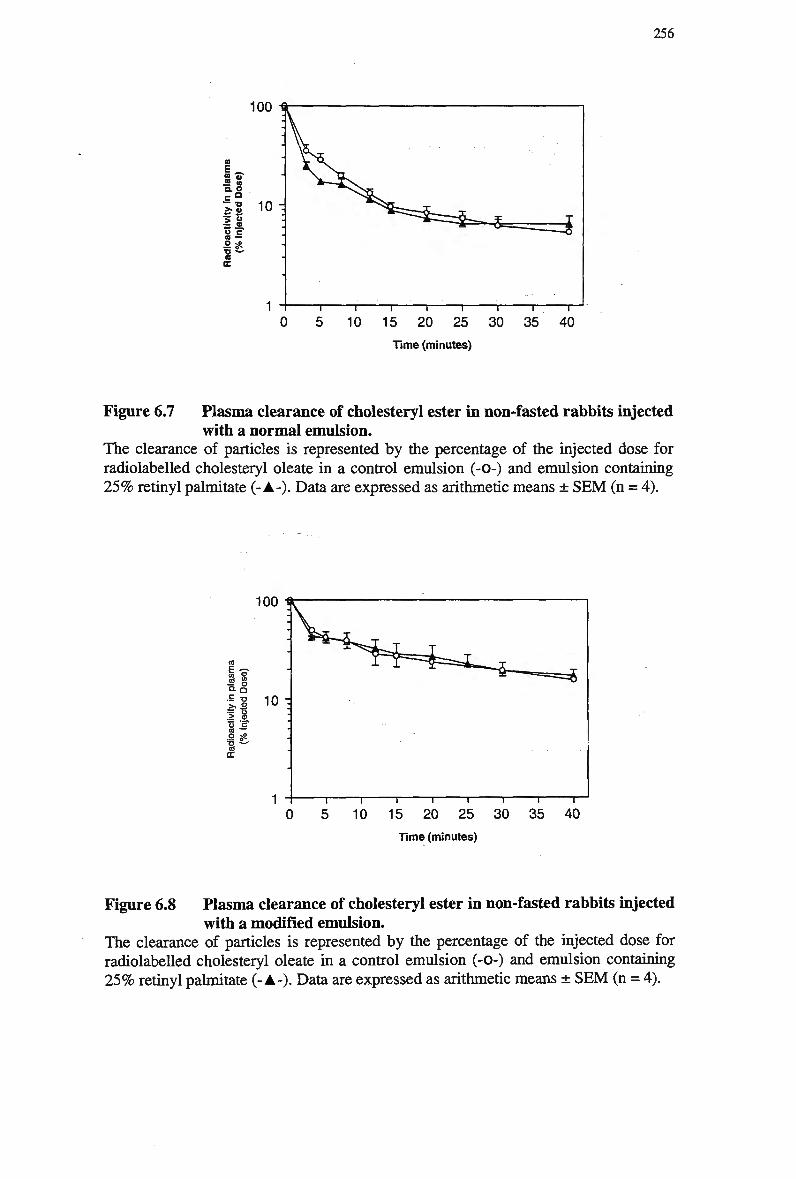
256
100
Time (minutes)
Figure 6.7 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a normal emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate in a control emulsion (-0-) and emulsion containing 2 5 % retinyl palmitate (- A-). Data are expressed as arithmetic means ± S E M (n = A).
10 15 20 25 30 35 40 Time (minutes)
Figure 6.8 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate in a control emulsion (-0-) and emulsion containing 2 5 % retinyl palmitate (- A-). Data are expressed as arithmetic means ± S E M (n = 4).
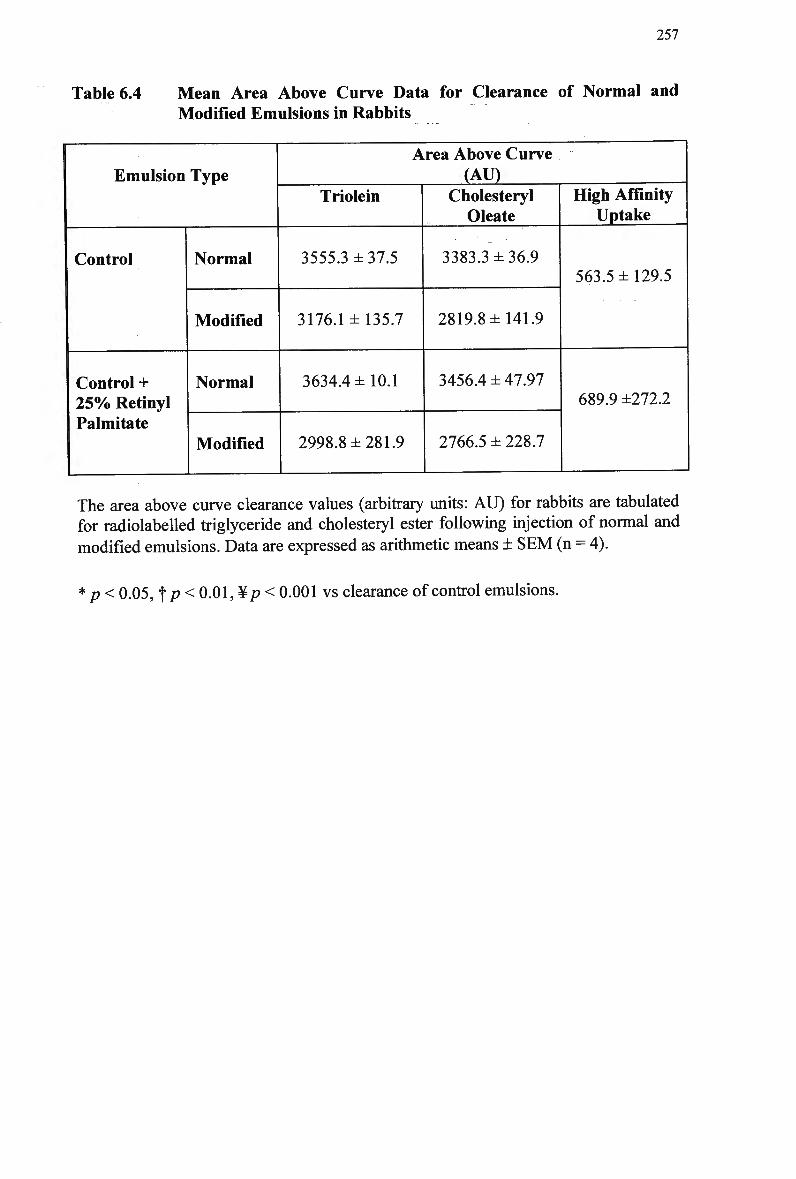
257
Table 6.4 Mean Area Above Curve Data for Clearance of Normal and
Modified Emulsions in Rabbits
Emulsion Type
Control
Control + 2 5 % Retinyl Palmitate
Normal
Modified
Normal
Modified
Area Above Curve (AV)
Triolein
3555.3 ±37.5
3176.1 ±135.7
3634.4 ±10.1
2998.8 ±281.9
Cholesteryl Oleate
3383.3 ±36.9
2819.8 ±141.9
3456.4 ± 47.97
2766.5 ± 228.7
High Affinity Uptake
563.5 ±129.5
689.9 ±272.2
The area above curve clearance values (arbitrary units: A U ) for rabbits are tabulated for radiolabeled triglyceride and cholesteryl ester following injection of normal and
modified emulsions. Data are expressed as arithmetic means ± S E M (n = 4).
* p< 0.05, f p < 0.01, ¥/? < 0.001 vs clearance of control emulsions.

258
6.3.5 Effect of Retinyl Stearate Incorporation on Plasma Clearance
of Modified Chylomicron-Like Emulsions in Rabbits
The clearance studies in Section 6.3.2 established that retinyl palmitate could be
incorporated into chylomicron-like emulsions without altering plasma clearance
kinetics of triglyceride clearance or particle uptake in rats. Measuring the clearance of
emulsions containing retinyl palmitate in rabbits assessed the effect of C E T P and
other differences influencing lipolysis between the two animal species, on emulsion
kinetics. The quantitation of high affinity (receptor) uptake is most accurately
assessed during simultaneous injection of normal and modified emulsions. Therefore,
it was necessary to assess whether a second restinyl ester would be a suitable
radioisotope replacement in modified emulsions, to allow simultaneous injection of
the two emulsion types without compromising the validity of the procedure. T o
determine this, retinyl stearate was synthesised and 2 5 % of the emulsion cholesteryl
oleate mass was substituted with retinyl stearate, as described in the Section 6.2.2 of
this chapter. Modified chylomicron-like emulsions with and without retinyl stearate
were injected into rabbits, and plasma clearance kinetics compared.
6.3.5.1 Clearance of lipids in modified emulsions
The clearance profiles for modified emulsions within rabbits are presented in Table
6.5. The modified emulsion hydrolysed rapidly, with > 9 6 % and 9 0 % of particle
triolein disappearing from plasma 12 min after injection, for control emulsions and
emulsions containing 2 5 % retinyl stearate, respectively (data not shown). The amount
of triglyceride cleared from plasma ( A A C ) was similar for control emulsions or
emulsions with 2 5 % retinyl stearate incorporated. Mean A A C values were greater for
triglyceride than cholesteryl ester in rabbits.
Figure 6.9 compares the clearance of modified emulsion cholesteryl oleate
from plasma in rabbits. Approx. 9 0 % of emulsion cholesteryl oleate cleared from
plasma by 30 min, for control emulsions and emulsions containing 2 5 % retinyl
stearate. The amount of cholesteryl ester cleared from plasma of rabbits was similar
for control emulsions and emulsions with 2 5 % retinyl stearate incorporated,
indicating that plasma clearance kinetics of triglyceride clearance or particle uptake in
rabbits is not altered by the incorporation of retinyl stearate into modified emulsions.
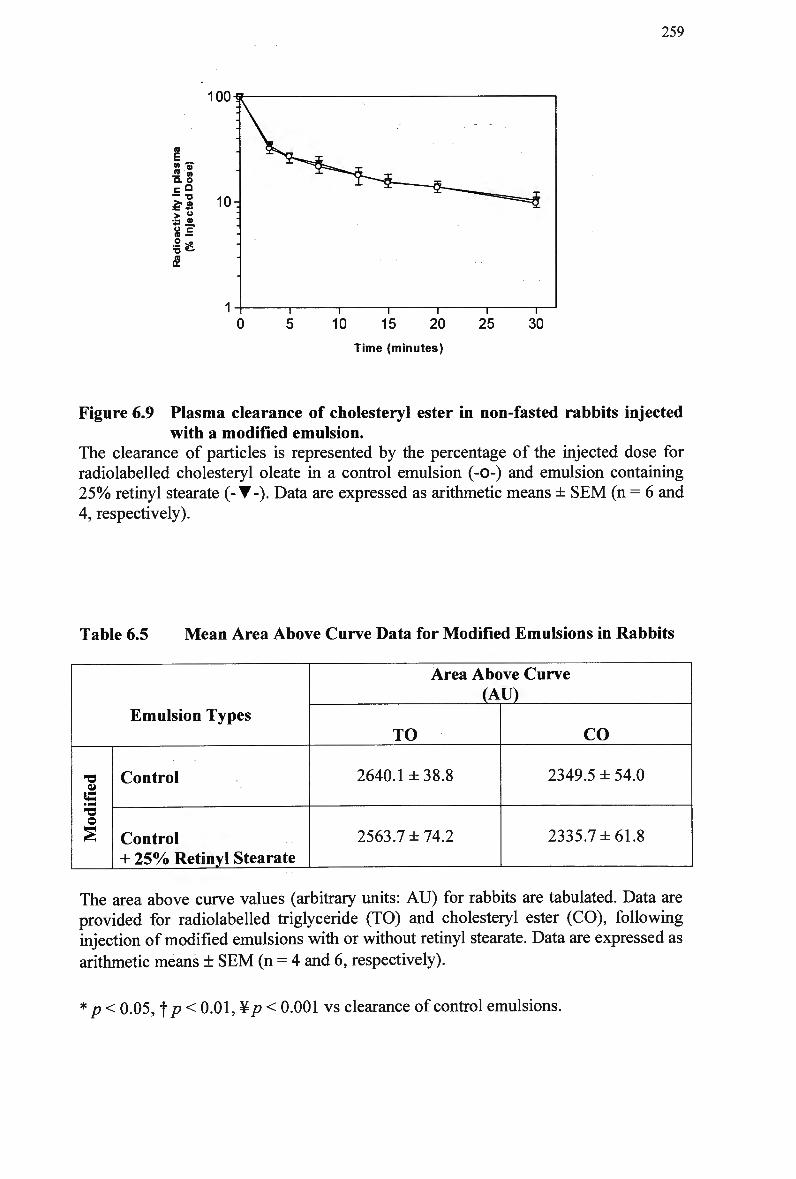
259
100
S » IS. o .EQ
> « 10-
iS
Time (minutes)
Figure 6.9 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate in a control emulsion (-0-) and emulsion containing 25% retinyl stearate (-•-). Data are expressed as arithmetic means ± SEM (n = 6 and 4, respectively).
Table 6.5 Mean Area Above Curve Data for Modified Emulsions in Rabbits
Emulsion Types
a (S O
Control
Control + 2 5 % Retinyl Stearate
Area Above Curve (AU)
TO
2640.1 ±38.8
2563.7 ±74.2
CO
2349.5 ± 54.0
2335.7 ±61.8
The area above curve values (arbitrary units: AU) for rabbits are tabulated. Data are provided for radiolabeled triglyceride (TO) and cholesteryl ester (CO), following injection of modified emulsions with or without retinyl stearate. Data are expressed as
arithmetic means ± SEM (n = 4 and 6, respectively).
*p< 0.05, f p < 0.01, ¥p < 0.001 vs clearance of control emulsions.

260
6.3.6 Effect of Retinyl Myristate Incorporation on Plasma
Clearance of Normal Chylomicron-Like Emulsions in
Rabbits
Retinyl myristate was investigated as a possible non-isotopic tracer for normal
emulsion particles. The aim of this study was to assess whether retinyl myristate
would alter the clearance kinetics of emulsion triglyceride and cholesteryl ester in
emulsions. Retinyl myristate was synthesised (Section 6.2.2) and 2 5 % of the
emulsion cholesteryl oleate mass was substituted with retinyl myristate. Normal
chylomicron-like emulsions with and without retinyl myristate were injected into
rabbits and their clearance parameters compared.
6.3.6.1 Clearance of lipids in normal emulsions
The clearance profiles of normal emulsions in rabbits are presented in Table 6.6. The
disappearance of triglyceride preceded particle clearance, indicating that
incorporation of retinyl myristate into the normal emulsion particles did not impair
lipolysis or remnant formation. The normal emulsion hydrolysed rapidly, with approx.
9 0 % of particle triolein disappearing from plasma by 8 min after injection, for control
emulsions and emulsions containing 2 5 % retinyl myristate (data not shown). The
A A C data for triglyceride in rabbits was similar for control emulsions or emulsions
with 2 5 % retinyl myristate incorporated.
The disappearance of triglyceride from plasma was followed by a rapid
clearance of particle remnants, with approx. 9 6 % of labelled cholesteryl oleate
removed by 30 min for control emulsions and emulsions containing 2 5 % retinyl
myristate (Figure 6.10). The clearance of normal emulsion cholesteryl oleate from
plasma in rabbits was similar for control emulsions and emulsions with 2 5 % of the
cholesteryl oleate mass substituted with retinyl myristate, with respect to the amount
cleared from plasma (Table 6.6).
The results indicate that plasma clearance kinetics of triglyceride clearance or
particle uptake in rabbits is not altered by the incorporation of retinyl myristate into
normal emulsions.
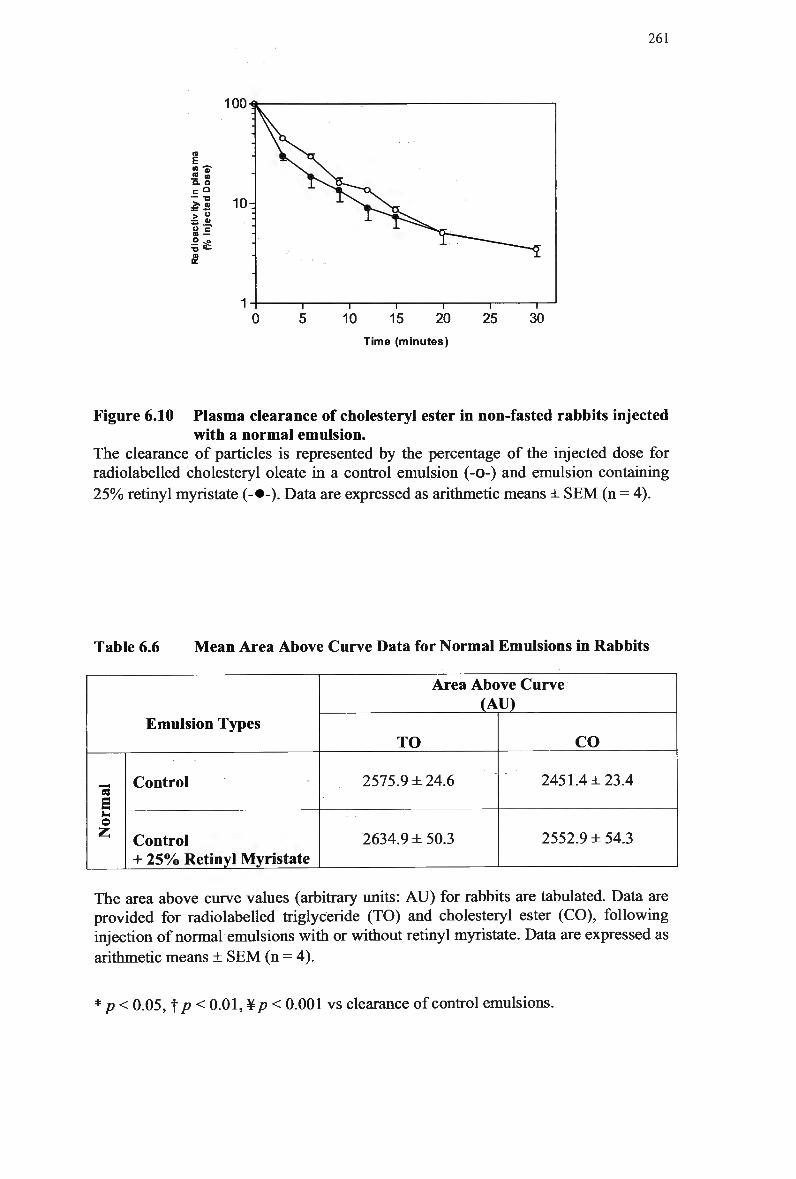
261
100
> °
10 15 20
Time (minutes)
25 30
Figure 6.10 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a normal emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate in a control emulsion (-0-) and emulsion containing
2 5 % retinyl myristate (-•-). Data are expressed as arithmetic means ± S E M (n = 4).
Table 6.6 M e a n Area Above Curve Data for Normal Emulsions in Rabbits
Emulsion Types
E s_
e Z
Control
Control + 25% Retinyl Myristate
Area Above Curve (AU)
TO
2575.9 ± 24.6
2634.9 ± 50.3
CO
2451.4 ±23.4
2552.9 ± 54.3
The area above curve values (arbitrary units: A U ) for rabbits are tabulated. Data are provided for radiolabeled triglyceride (TO) and cholesteryl ester (CO), following injection of normal emulsions with or without retinyl myristate. Data are expressed as
arithmetic means + S E M (n = 4).
* p< 0.05,1[p< 0.01, ¥/? < 0.001 vs clearance of control emulsions.

262
6.4 Discussion
The project involved the development of two lipid emulsions, which represent total
and non-receptor uptake of postprandial lipoproteins from plasma. The difference in
clearance represents removal via high-affinity (receptor) pathways. Utilisation of this
procedure is currently limited by the use of potentially hazardous radioisotopes
therefore it was necessary to develop non-isotopic tracers for use in human subjects.
As discussed in Chapter 1, it is well established that retinyl palmitate acts as a
marker for chylomicron remnant particles. Retinyl esters do not alter the kinetics of
chylomicrons and their remnants in vivo, because vitamin A is hydrophobic and is
secreted into the intestinal lymph as a chylomicron constituent under physiological
conditions, predominantly in the form of retinyl esters (Wilson et al, 1983).
However, it was not known whether retinyl esters would distribute within the core of
chylomicron-like lipid emulsions, and if so, whether the metabolism of these
emulsions would be disturbed under these conditions. The aim of this study was to
assess whether retinyl esters could be incorporated into chylomicron-like emulsions
without compromising the validity of the procedure.
In the present investigation, retinyl esters (palmitate, stearate and myristate)
were incorporated into normal and modified chylomicron-like emulsions. Equal
masses of the cholesteryl ester component of emulsions were substituted with retinyl
esters to maintain the relative proportion of core to surface components, without
disturbing the emulsion kinetics in vivo. Emulsions were then injected into recipient
animals and their clearance kinetics determined.
The effect of low (25%) and high (50%) doses of retinyl palmitate
incorporation on the clearance from plasma of emulsions was assessed in rats. At both
doses, clearance from plasma of normal and modified chylomicron-like emulsion
triglyceride, calculated as A A C , was not altered. Particle clearance, as determined by
emulsion cholesteryl oleate, was similar for normal emulsions with or without retinyl
palmitate incorporated. The A A C data for cholesteryl oleate in modified emulsions
was similar to control emulsions at low doses. However, the incorporation of 5 0 %
retinyl palmitate significantly increased the amount of cholesteryl oleate removed
from plasma, significantly reducing the amount of modified emulsion cleared by high
affinity pathways.

263
Assessment of radiolabeled cholesteryl oleate and triglyceride accumulation
in the liver and spleen suggests that the incorporation of 2 5 % retinyl esters in normal
and modified emulsions did not alter organ uptake of either lipid. However, the
incorporation of 5 0 % of retinyl ester increased the organ uptake of normal emulsion
cholesteryl oleate and triglyceride. In contrast, the inclusion of 5 0 % retinyl ester in
the modified emulsion decreased splanchnic uptake and significantly increased
cholesteryl oleate uptake by the liver. Thus the modified emulsion behaved more as a
chylomicron remnant with respect to the pattern of organ uptake.
To assess the effect of C E T P on emulsion kinetics, comparative studies were
carried out in rabbits. It was essential to eliminate the possibility that C E T P activity
would influence the clearance kinetics by causing exchange or loss of the retinyl
esters and cholesteryl esters in vivo. The clearance of normal chylomicron-like
emulsions containing low (25%) doses of retinyl palmitate and retinyl myristate and
modified chylomicron-like emulsions containing low (25%) doses of retinyl palmitate
and retinyl stearate was examined in rabbits. Organ uptake data was not collected, as
the rat data was considered conclusive and reflected the clearance data for both
emulsion types. In addition, each rabbit acted as its' own control for comparison of
the metabolism of the two emulsion types with and without retinyl esters incorporated
and therefore were not sacrificed. Retinyl esters or C E T P were not deleterious to the
fate of normal or modified emulsion in vivo, as assessed by A A C data. For each
emulsion type, the disappearance of triglyceride preceded particle clearance,
indicating that incorporation of retinyl palmitate into emulsion particles did not impair
lipolysis and remnant formation via lipolysis was concluded to be normal. This was
essential for the use of retinyl esters in the proposed technique, as a delay in
delipidation of chylomicron-like emulsions may result in a particle that is poorly
recognised by LDL-receptors.
These studies have established that the introduction of retinyl esters into
normal or modified chylomicron-like emulsions does not alter the size of these
emulsion particles or their metabolism in animal models. The findings permitted
continuation with the primary aim of the project, which involves the development of a
diagnostic assay using two of these retinyl esters as non-isotopic tracers for emulsion
particles. The development of this procedure will be investigated in the next chapter.

264
Chapter 7: Quantitation of Retinyl Esters in Chylomicron-Like Emulsions
7.1 Introduction
The proposed method involves the development of a diagnostic assay using retinyl
esters as tracers for chylomicron-like emulsion particles. Hence two suitable retinyl
esters were required for the detection of normal and modified emulsion following
simultaneous intravenous injection. The current study was concerned with the
detection of the retinyl esters (explored in Chapter 6), based on their inherent
characteristics. It was important to consider the relative retention times of retinyl
esters, to avoid co-elution and to ensure that the peak values were readily identifiable
for quantitation and subsequent calculation of clearance parameters. The differential
hydrophobicity of retinyl esters is determined by their fatty acyl chains, and like fatty
acids, the hydrophobicity and retention time of retinyl esters increases with increasing
acyl chain length and decreases with extent of unsaturation (Futterman and Andrews,
1964b), (Huang and Goodman, 1965). Retinyl oleate (Cig: i) is also two carbons
longer than retinyl palmitate (Ci6: o) therefore oleate is likely to be more active under
most conditions. However, retinyl oleate and retinyl palmitate have similar adsorption
characteristics and provide the greatest challenge for separation (Ross, 1990) due to
similarities in polarity (Ross, 1981b). The characteristics of retinyl stearate and
myristate were discussed in Chapter 6.
The aim of the current study was to develop a method for the detection of
plasma retinyl esters following injection of retinyl ester-labelled chylomicron-like

265
emulsions. The detection of retinyl esters was necessary to enable calculation of the
percent of injected dose remaining in plasma at any time. A method for detecting
plasma retinyl esters in human plasma has already been established for the fat load
test using large masses of retinyl palmitate, however this may not be suitable for a
small bolus emulsion injection in rabbit models. It was necessary to incorporate a
sufficient mass of retinyl ester into the emulsion particle for detection in plasma,
without saturating existing lipid pools or compromising the plasma kinetics of
emulsion particles.
The detection of retinyl esters in the plasma of human subjects (Weintraub et
al, 1987a), (Weintraub et al, 1987b), (Ross, 1990), (Berr et al, 1983), (Gylling et
al, 1996), (Richey Sharrett et al, 1995), (Krasinski et al, 1990b), (Karpe et al,
1995), (Wilson et al, 1983) and animal models (Bhat and Lacroix, 1983) plasma is
well documented. However, these studies utilised large doses of vitamin A (approx.
63 m g /70 kg subject) ingested as a fatty meal (approx. 58 g m lipid/70 kg subject), to
produce retinyl palmitate-rich chylomicrons. High fat meals enhance the intestinal
production of chylomicron particles, which potentially saturates the clearance
pathways (Bergeron and Havel, 1997). However, the strategy of Staprans et al.
(Staprans et al, 1992) in injecting increasingly larger doses of radiolabeled
chylomicron triglyceride seemed to prevent meaningful interpretation in terms of
tracing the physiological events of interest in rats. Clearance rate and hepatic uptake
were slowed as the injected triglyceride load increased, suggesting saturation of
clearance pathways in both control and diabetic rats. Ideally the tracer and the lipid
mass injected should be of the smallest possible quantity to avoid perturbation of the
existing pool (Martins et al, 199A).
The quantitation of high affinity uptake in vivo in animal models has been
previously reported ( M a m o et al, 199A), however radiolabeled cholesteryl oleate was
used as a marker for remnant particle uptake, allowing the mass of tracer and lipid to
remain relatively low. The present study was designed to investigate whether retinyl
esters were a suitable marker for remnant particles, for future application in humans.
The proposed method was concerned with monitoring plasma clearance of small
doses of emulsion retinyl esters, thus it was important to attain a sufficient
concentration of retinyl ester in plasma to enable detection via H P L C without altering
the altering emulsion kinetics in vivo. Equal masses of cholesteryl ester were
substituted with retinyl esters to maintain the relative proportion of core to surface

266
components. Perceived difficulties lie in the relative sensitivity of the H P L C system,
and inability to substitute > 2 5 % emulsion cholesteryl oleate with retinyl ester, as
confirmed by studies in the previous chapter.
The effect of increasing the emulsion lipid mass injected was also investigated
to facilitate the detection of retinyl esters in plasma, without saturating hydrolysis of
chylomicron triglyceride or compromising remnant uptake. For the purpose of
detecting plasma retinyl esters in rabbits, it was necessary to inject up to 1 5 % of the
total triglyceride pool, and up to 2 0 % is known to not saturate clearance kinetics in
vivo. Rabbits were used as an animal model, based on the presence of C E T P activity
and their susceptibility to developing atherosclerosis. The use of radioisotopes in
animal models is universally acceptable therefore dosage refinement will be
investigated in human subjects. Preliminary calculations predict that injection of 2.5
m g retinyl ester and 225 m g triglyceride ( 5 % of the total triglyceride pool) into a 70
kg normolipidemic individual would be sufficient for detection. For a dyslipidemic
subject, this mass of triglyceride would represent a lower percent of their total
triglyceride pool, therefore complications regarding competition or saturation of lipid
clearance are not anticipated.
Normal and modified chylomicron-like emulsions contained radiolabeled
triglyceride to monitor lipolysis, and radiolabeled cholesteryl oleate and retinyl ester
to monitor particle clearance. Our laboratory has previously established that > 9 8 % of
plasma radioactivities are cholesteryl esters associated with remnants of lipoproteins
with a density of <1.006 gm/ml. Emulsions were injected into the recipient animal
and their clearance determined from decline in plasma in cholesteryl ester
radioactivities. T o assess the suitability of utilising retinyl esters in emulsions for
quantitation of particle clearance, the clearance of cholesteryl oleate radioactivities
was compared with clearance parameters generated from the decline in plasma retinyl
ester concentrations, determined by H P L C . T o assess the suitability of utilising retinyl
esters for quantitation of receptor-mediated uptake, high affinity was calculated as the
difference in clearance of the two emulsion types, for A A C values. The high affinity
uptake values for the two types of tracees (cholesteryl oleate radioactivities and
retinyl esters) were compared.

267
7.2 Special Methods
7.2.1 Animals
Male New Zealand White (NZW) and semi-lop (NZW/ - cross) rabbits, weighing
between 1.8 to 3.5 kg, were obtained from the Animal Resources Centre, Murdoch,
Western Australia. The same animals were re-used for all experiments, thus acting as
controls for both emulsion types and detection of retinyl esters.
7.2.2 Preparation of Retinyl Esters
Retinyl stearate, oleate and myristate were synthesised from retinyl acetate by
transesterification with the appropriate fatty acyl chlorides under reduced pressure,
following the method of Huang and Goodman (Huang and Goodman, 1965), as
described in Chapter 6. Retinyl myristate was also prepared according to the methods
of Lentz etal. (Lentz et al, 1975), following oxidation of retinyl esters as synthesised
by the acyl chloride method. The known instability of acyl chlorides along with the
large excess of fatty acid used led us to attempt the acylation of retinol using the more
stable acid anhydrides. Since the anhydrides are known to be the mildest esterifying
agents, this procedure has already been suggested to be well suited for the
esterification of such labile compounds as the sterols (Lentz et al, 1975).
Briefly, anhydride acylation was carried out by mixing retinol and the desired
anhydride in C C U in a ratio of 1 mole of retinol: 3 moles of anhydride. After removal
of the organic solvent, the neat reaction was heated for the amount of time necessary
to optimise the yield. Retinol was dissolved in CCI4. To this solution in a 25 ml pear-
shaped flask was added a C C U solution of oleic anhydride (209 pmoles in 1.23 ml).
The solvent was removed from the mixture on a rotary evaporator under reduced
pressure. The flask containing the reaction mixture was then flushed with argon,
capped and rotated at the desired temperature in a heated oil bath. After the
appropriate reaction time the reaction mixture was dissolved in heptane. The yield of
retinyl ester was determined using TLC. The retinyl ester was purified on a silicic
acid column eluted first with heptane (~ 6 column volumes) and then with 1 %
benzene in heptane. The elution profile was followed by T L C analysis. Retinol ester
appeared in the initial 1 % benzene in heptane fraction. Late 1 % benzene in heptane

268
fractions contained traces of acid anhydride and was removed by chromatography on
the same column. Purity was checked using T L C with two developing systems: (a)
benzene: hexane 60:40 and (b) petroleum ether: ether 80:20. The pure product
produced a mass spectrum consistent with the fragmentation pattern expected for a
retinyl ester.
Prior to experimental use, each retinyl ester was assayed on a Beckman
DU650 spectrophotometer, at a wavelength of 325 nm. The purity of all retinyl esters
was confirmed regularly by examination of their ultraviolet absorption spectra
(Beckman) and TLC.
7.2.3 Preparation of Normal Chylomicron-Like Emulsions
Normal lipid emulsions resembling chylomicrons in composition and size were
prepared as described in Chapter 2, with the following changes. Triolein (70 mg),
cholesteryl oleate (2.25 mg), cholesterol (2 mg) (Nu Chek Prep, Elysian, M N ) and
egg phosphatidylcholine (25 mg) (Lipid Products, Surrey, U K ) , each greater than 99
% pure were dispensed from stock solutions into vials. To this mixture 0.75 m g
retinyl palmitate (Sigma Chemical Co., Australia), stearate, oleate or myristate was
added, substituting 2 5 % of emulsion cholesteryl oleate component. The ratio of lipids
remained the same for all experiments for normal and modified emulsions, regardless
of specific activity or the lipid mass injected. The emulsions were labelled with
[3H]triolein and cholesteryl[14C]oleate (Amersham, Surry Hills, N S W , Aust) to
monitor lipolysis and particle uptake, respectively.
7.2.4 Preparation of Modified Chylomicron-Like Emulsions
Modified emulsion particles were prepared according to the method described in
Chapter 2, with the following changes. Triolein (70 mg), cholesteryl oleate (2.25 mg)
(Nu Chek Prep, Elysian, M N ) and egg phosphatidylcholine (25 mg) (Lipid Products,
Surrey, U K ) , each greater than 99 % pure were dispensed from stock solutions into
vials. To the emulsion lipid mixture, 0.75 m g retinyl palmitate (Sigma Chemical Co.,
Australia), stearate, oleate or myristate was added, substituting 2 5 % of emulsion
cholesteryl oleate component. The modified chylomicron-like lipid emulsions were

269
labelled with [14C]triolein and cholesteryl[3H]oleate (Amersham, Surry Hills, N S W ,
Aust) to monitor lipolysis and particle uptake, respectively.
7.2.5 Clearance Studies in Rabbits
The protocol for clearance studies in rabbits is described in Chapter 2. Below are the
changes made for this study. Clearance studies using a modified emulsion were
carried out on the day prior to clearance studies using normal emulsions, to avoid any
accumulation or contamination of plasma with unesterified cholesterol.
7.2.5.1 1 x lipid mass
Chylomicron-like emulsions were injected into the lateral ear vein of conscious
rabbits. Approx. 2.5 m g of emulsion triglyceride (less than 5 % of total plasma
triglyceride pool) and 32 ng retinyl ester was injected, in a volume of 0.4-0.8 ml.
Blood samples of 1.0 ml were subsequently taken from the opposite ear vein at 3, 5, 8,
12, 15, 20 and 30 min.
7.2.5.2 3 x lipid mass
To increase the amount of retinyl ester available for detection by HPLC in plasma, the
total amount of lipid mass (and retinyl ester) injected into the animal was increased.
This was achieved by maintaining the total amount of radioactivity in the emulsion,
and increasing the lipid mass by three times. Hence the specific activity, lipid ratio
and injection volume all remained constant. To avoid putative complications in
interpretation of the data and to ensure sufficient retinyl ester detection by H P L C ,
approx. 12 -16 m g of emulsion triglyceride (20-30% of total plasma triglyceride pool)
was injected into recipient animals. This was equivalent to approx. 17-24 m g lipid, in
a volume of 0.4-0.8 ml. The amount of retinyl ester injected was 80 pg, to ensure a
plasma concentration of 0.5 pg/ml and detection up to 30 min. Chylomicron-like
emulsions were concentrated to reduce the volume injected into rabbits. Blood
samples of 1.0-2.0 ml were subsequently taken from the opposite ear vein at 3, 5, 8,
12, 15, 20 and 30 min.

270
7.2.6 Emulsion Clearance from Plasma
Determination of radioactivity in samples and calculation of emulsion clearance from
plasma was described in Chapter 6. Clearance of emulsions determined from decline
in plasma triolein and cholesteryl oleate radioactivities was compared with clearance
parameters generated from the decline in plasma retinyl ester concentrations.
7.2.7 Organ Extraction
Radioactivities in the liver and spleen were extracted using a modified procedure of
Folch et al. (Folch et al, 1957), as described in Chapter 2.
7.2.8 High Performance Liquid Chromatography (HPLC)
7.2.8.1 Properties of retinyl esters
Retinol and related fatty acid esters are characterised by strong UV absorption due to
their conjugated system of double bonds. Therefore, reversed phase H P L C coupled
with UV/visible detection is commonly used for isolation, separation and
identification of retinyl esters (Ross, 1981b), (Furr et al, 1986), (Bhat and Lacroix,
1983). Retinyl esters are readily detectable at Xm^ 325 nm, however the absorbance of
standard solutions is scanned over the wavelength 250-400 nm, so as to disclose
possible decomposition of standards (Furr, 1990). The selectivity of the H P L C system
depends on the fact that few other biological compounds absorb light significantly at
325 nm. Most other lipids are transparent at this wavelength, thus selective analyses
of retinyl esters is possible. As shown by Ross (Ross, 1981b), retinol and retinyl
esters have equal molar absorptivities and so standards may be quantitated by
absorbance. Because the retinyl esters have equal molar absorbance, retinyl acetate
may be used as quantitative standard for other esters if peak area is used.
Retinyl esters are among the most hydrophobic of retinoids and thus reversed-
phase H P L C columns of octadecyl- or octyl-substituted silica gel allow good
adsorption of these molecules, with desorption by solvents of low dielectric constant.
The differential hydrophobicity of retinyl esters is determined by their fatty acyl
chains, and, like fatty acids, the hydrophobicity increases with increasing acyl chain
length and decreases with extent of unsaturation (Ross, 1981b). Retention times of

271
retinyl esters also increase with increasing polarity (Futterman and Andrews, 1964b)
and retinyl esters are separated according to fatty acid chain length.
To prevent decomposition, retinyl esters were stored in the dark at -20°C under
argon either suspended in ethanol or as a dried lipid. The stock solutions were
prepared every month and fresh standards were prepared from the stock solutions for
each assay. Retinyl esters were measured using a modification of the method
described by Ross (Ross, 1981b). The concentrations of solutions of retinyl esters in
ethanol were calculated using the molar extinction coefficient of 52,275 M cm" at
their absorption maxima of 324-326 n m (Boldingh et al, 1951).
7.2.8.2 Materials
All-trans retinyl acetate, retinyl palmitate and fatty acyl chlorides were purchased
from Sigma Chemical Co. All chemicals were the highest grade commercially
available. Solvents were H P L C grade and were obtained from Merck (Darmstadt,
Germany) and filtered through Fluoropore filters (Millipore Corp.) and degassed
under vacuum before used. Water was house-distilled, then passed through a Milli-Q
purification system (Millipore, Milford, M A , U S A ) .
7.2.8.3 Standard curves and calculations
The standard curves were prepared by adding a constant amount of internal standard
to 7 different concentrations of the retinyl esters. The concentrations, in ethanol, were
determined spectrophotometrically using Beer's Law. The absorptivity coefficient
values (Ei%/cm) used were 1510 at 325 n m for retinyl acetate, and 975 at 325 n m for
the remaining retinyl esters. Duplicates of standards were prepared by adding 10 pi of
internal standard (6 pg/ml) to 90 pi standard. Computed areas were regressed against
the amount of standard injected. Retinoid standards gave a linear calibration curve
over the range of concentrations expected in serum (1.724 and 0.862 ng/pl for all
retinyl esters). The limit of detection was approx. 1.0 pmol (10 nmol/litre in 100 pi
sample). Best-fit linear regression lines were calculated using the method of least
squares. For all determinations, calculated values lay within the extreme limits of a
standard curve. Working calibration curves for retinyl acetate, myristate and oleate
(area to amount ratio) are shown in Figure 7.1. Serum concentrations in the unknown

272
samples were determined after measurement of the peak height ratios by use of these
calibration curves.
7.2.8.4 Extraction of blood samples
Following sampling, blood samples were placed directly into microfuge tubes, which
contained 20 pi heparin to prevent clotting. Samples were centrifuged at 1500 rpm for
10 min, and the plasma was transferred to glass kimble tubes for extraction. Retinyl
esters were extracted immediately after sample isolation with appropriate precautions
for exposure to light, using H P L C grade solvents. Whole plasma samples were used
due to the low volumes and concentrations of retinyl ester. Samples of 0.5-1.5 ml
were made up to a volume of 2.5 ml with D D W , and 3 pg retinyl acetate was added as
an internal standard. Samples were extracted with 2.5 ml ethanol and 5 ml hexane,
with mixing by inversion between each addition. The samples were mixed thoroughly
by inversion for 5 mins, then centrifuged at 1500 rpm for 5 min. T w o phases were
formed and 4 ml of the upper (hexane) phase was removed into graded glass tubes
and evaporated under nitrogen.
7.2.8.5 HPLC instrumentation
The residue from extraction was resuspended in 100 pi propan-2-ol, and an aliquot of
50 pi was injected automatically into an H P L C (Hewlett Packard Model 1050)
equipped with an ultraviolet absorbance monitor (variable wavelength). A reversed
phase Ci8 radial compression column (Hewlett Packard LiChrospher 100 RP-18; 5
p m ODS-18, cat. No. 799250D-564-3) and a Guardpak precolumn cartridge with
disposable inserts of 5 p m were used. The mobile phase was methanol (100%) and
was vacuum-filtered through a 45 p m Nylon filter (Millipore) prior to use, and
degassed automatically by the H P L C system. Flow rate was 1 ml/min at ambient
temperature, and pressures of 50 to 150 psi were generated.
Retinoid compounds were monitored with on-line U V detection at 320-330
n m and peaks were identified on the basis of their retention characteristics. Retention
times of the serum peaks matched those from a standard mixture containing the
different retinyl esters and retinyl esters were quantitated by measurement of peak
height and related to standard curves for purified retinyl esters. Results were

273
expressed as nmol of retinyl ester/litre of plasma, and area response of purified retinyl
ester. The relative retention times for retinyl acetate, palmitate, stearate, oleate and
myristate were 2.5, 13.6, 18.4, 13.9, and 10.1 min, respectively. Baseline separation
for most of the retinyl esters was achieved within 20 min. Chromatograms of such
standard mixtures for retinyl acetate (Ci: o), retinyl myristate (Ci4: o), retinyl palmitate
(Ci6: o), retinyl oleate (Cis: 0 and retinyl stearate (Cis: o) are shown in Figures 7.2 and
7.3. A typical chromatogram obtained from a plasma sample for retinyl palmitate is
shown in Figure 7.4.
7.2.8.6 Organ extraction
Retinyl esters of the liver and spleen were extracted within 4 h of removal from the
animals using a modified procedure of Folch et al (Folch et al, 1957), as described in
Chapter 2. The organs were weighed, minced and 0.5-1.0 g portions of the minced
organs extracted with 20 vol. of chloroform/methanol (2:1, v/v). After the chloroform
layer (infranatant) was removed from the extractant and dried under a stream of
nitrogen, one half of the volume was taken for retinyl ester detection via H P L C . To
this volume, 30 pg retinyl acetate was added, as an internal standard. The solvent was
then evaporated under a stream of nitrogen and 100 pi iso-propan-2-ol was added and
80 pi injected onto the H P L C column.
7.2.8.7 Retinyl Ester Calculations
Chylomicron retinyl ester was quantitated by the area ratio method, and data were
analysed by determining the A A C . Peak areas under the retinyl ester peaks were
measured using routine integration parameters. The ordinate represented the retinyl
ester peak area, while the abscissa represented the retinyl ester concentrations of the
solutions used in generating the standard curve. The concentration of retinyl ester in
each sample was determined by proportional comparison of the retinyl ester peak
height of the sample with the peak height of an internal standard. The total amount of
retinyl ester in plasma was calculated and corrected for the internal standard and
differences in analytical recovery between samples. The mass of emulsion retinyl
ester was also calculated, allowing retinyl ester at each time point to be expressed as a
percentage of the injected dose.

274
7.2.9 Calculations
Calculations for AAC are detailed in Chapter 2. The emulsion clearance parameters
determined from decline in plasma triolein and cholesteryl oleate radioactivities were
compared with clearance parameters generated from the decline in plasma retinyl
ester concentrations, for the two emulsion types. To assess the suitability of utilising
retinyl esters in emulsions for quantitation of receptor-mediated uptake, the A A C r e c
determined from the two types of tracees (cholesteryl esters and retinyl esters) were
compared. Normal and modified chylomicron-like emulsions contained labelled
triglyceride and cholesteryl oleate to monitor lipolysis and particle uptake,
respectively.
7.2.10 Statistical Analysis
To determine the suitability of retinyl esters as tracees of chylomicron remnant
particle clearance, statistical analysis was performed as described in Chapter 2.
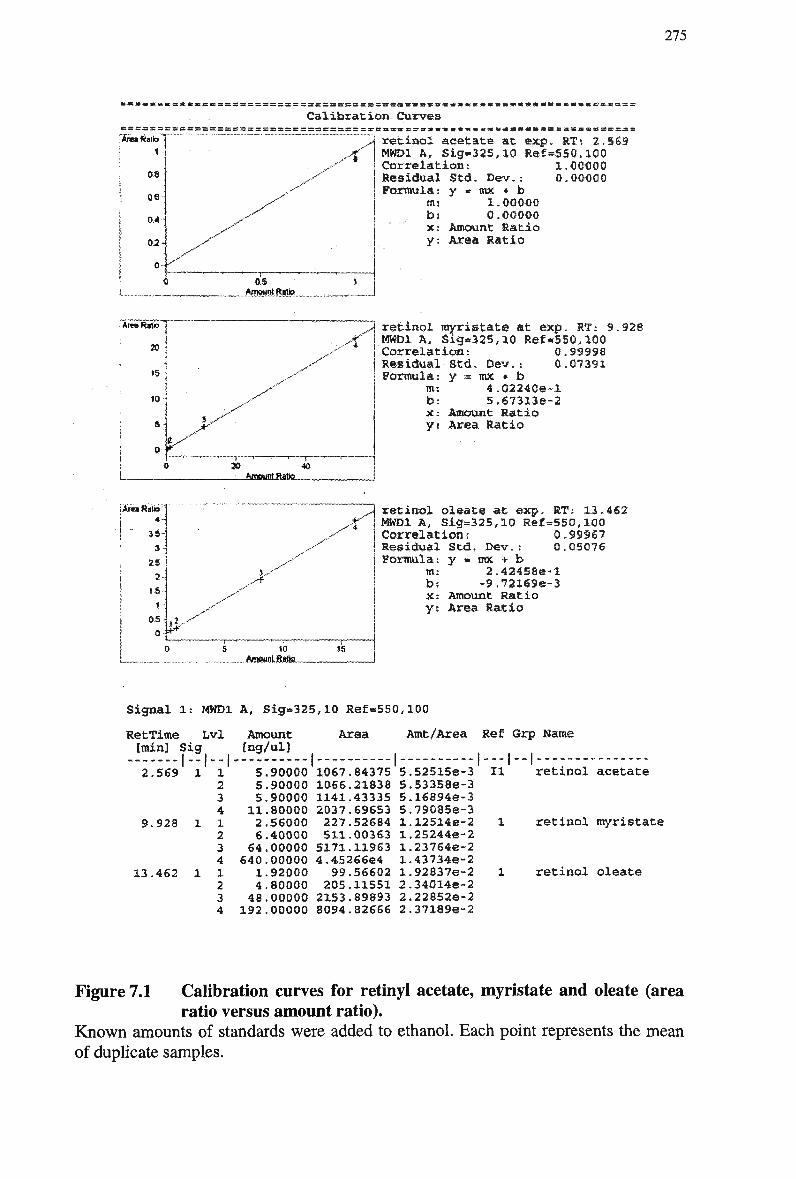
. « » » « « . £ £ 1 M . . . . . K « K t . M H l l H » H « » C S a S S s SS ss =r =: ss =: s K = = =: K = =: =:^ = s = = = — - s r s » « w 3
Calibration Curves
retinol acetate at exp. RT: 2.56:9 MWD1 A, Sig-325,10 Re€=5S0.1QO Correlation: 1.00000 Residual Std. Dev.: 0.00000 Formula: y » «ix * b
Bis 1.00000 b! 0.00000 x: Atsiount Ratio y: Area Ratio
alio
1
oa
oe-:
0A-
02-
0-<
y"
i
^
,y
&S ! _......Assspt. 8«§K! .„ _
;Are«Rs1io j
10 •<
3 ,
.X* ,,-r
—r~ 30
— i —
-&KSMSLBMI9L.
retinol myristate at exp. RT: 9 MWD1 A, Sig*325,10 Ref*550,2Q0 Correlation: 0.S3998 Residual-Std. Dev, s 0.07391 Formula: y = mx * b
m: 4.Q2240e~l b: 5,673136-2 x: Amount Ratio y: Area Ratio
928
I Area ftatb 1
S*
/
>
«0 15
retinol oleate at exp. RT: 13.462 MWD1 A, Sig=325,10 Ref=550,10Q Correlationr 0,99967 Residual Std. Dev. : 0.QS076 Formula: y « mx + b
m: 2.424S8e«l bs -9.72*696-3 X: Amount Ratio y: Area Ratio
Signal l: MWDl A, Sig*325,l0 Ref=550,100
RetTime Lvl Amount [min] Sig [ng/ul]
I-I--I
Area Amt/Area Ref Grp Name
1 "I — ! 2.569 1
9.928 1
13.462 1
1 2 3 4 1 2 3 4 1 2 3 4
5 5 5
11 2 6
€4 640 1 4
48 192
.90000
.90000 90000 .80000 .56000 .40000 .00000 00000 92000 80000 00000 00000
1067 1066 1141 2037 227 511
5171
84375 21 o J o 43335 69653 52684 00363 11963
4.45266e4 99
205 2153 8094
56602 11551 89893 82666
5 5 5 5 1 1 1 1 1 2 2 2
,52515e-3 .53358e-3 .16894e-3 79085e-3 12514e-2 .25244e-2 .23764e-2 437346-2 92837e-2 340146-2 .228526-2 .37189e-2
II retinol acetate
retinol myristate
retinol oleate
Figure 7.1 Calibration curves for retinyl acetate, myristate and oleate (area ratio versus amount ratio).
Known amounts of standards were added to ethanol. Each point represents the mean of duplicate samples.
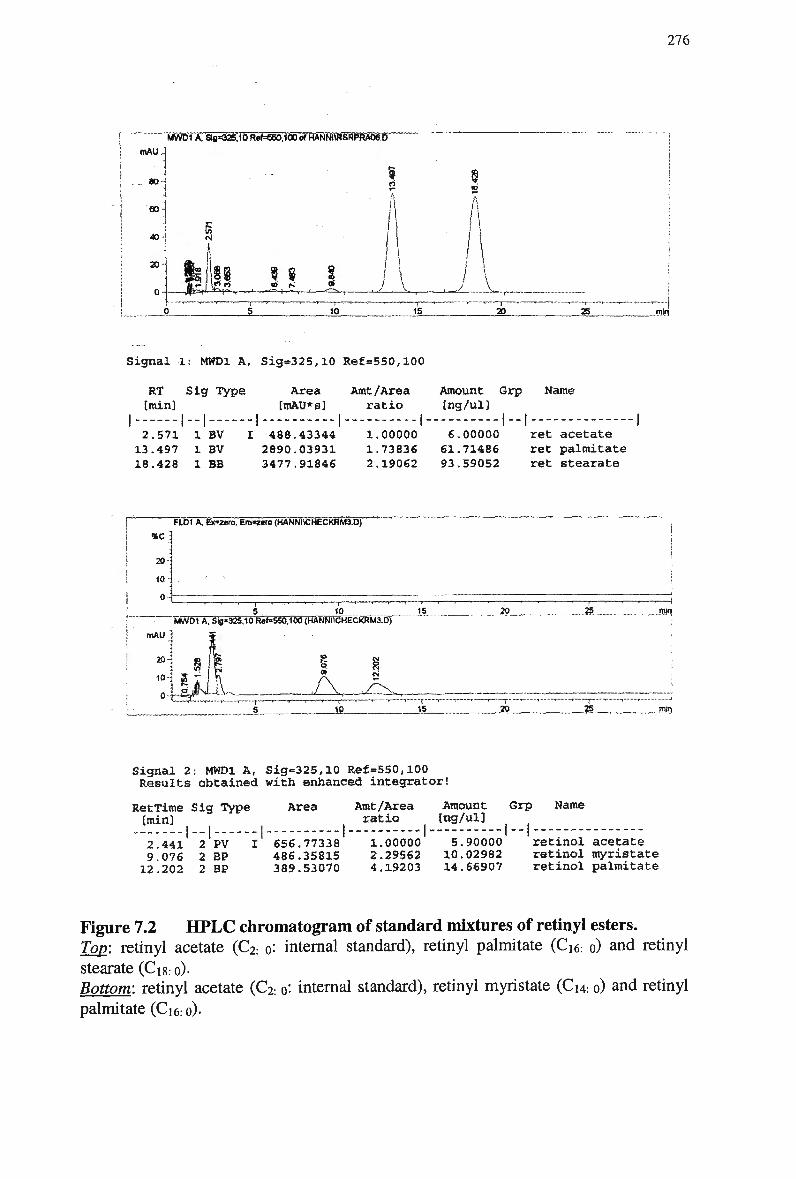
276
[ "' TSBBfOSF^KilO RaWW.IOD Of HANNIWSRPRA06.D | mAU-
! ao-i ! 1
"1 • i
40 j
20
0
Ei CM
B«2 f \Sf rt §
o is
«0
A
A to 16 20 25 -ffil.r|
Signal 1: MWDl A, Sig*325,10 Ref=550,100
RT Sig Type train]
I
Area Amt/Area Amount Grp Name EmAU*s] ratio [ng/ul]
2.571 1 BV I 488.43344 1,00000 6.00000 ret acetate 13.497 1 BV 2890.03931 1.73836 61.71486 ret palmitate 18.428 1 BB 3477.91846 2.19062 93.59052 ret stearate
FUJI A, Bezeia, em-zera <HANNI«HECKRT56xfr
10 15 20 jam MWDl A. Sig*325.10 R**-S50,100 (HANNUCHECKRM3.D)
ItlAU 1
j i
1fli sin 10
"T" 15 25
Signal 2: MWDl A, Sig=325,10 Ref=550,100 Results obtained with enhanced integrator!
RetTime Sig Type (min]
l"| — 2.441 2 PV 9.076 2 BP 12.202 2 BP
Area Amt/Area Amount Grp Name ratio (ng/ul]
I 656.77338 486.35815 389.53070
1.00000 5.90000 retinol acetate 2.29562 10.02982 retinol myristate 4.19203 14.66907 retinol palmitate
Figure 7.2 H P L C chromatogram of standard mixtures of retinyl esters. Top: retinyl acetate (C2: o: internal standard), retinyl palmitate (Ci6. o) and retinyl
stearate (Ci8:o)-Bottom: retinyl acetate (C2: „: internal standard), retinyl myristate (Cu. o) and retinyl palmitate (Ci6: o)-
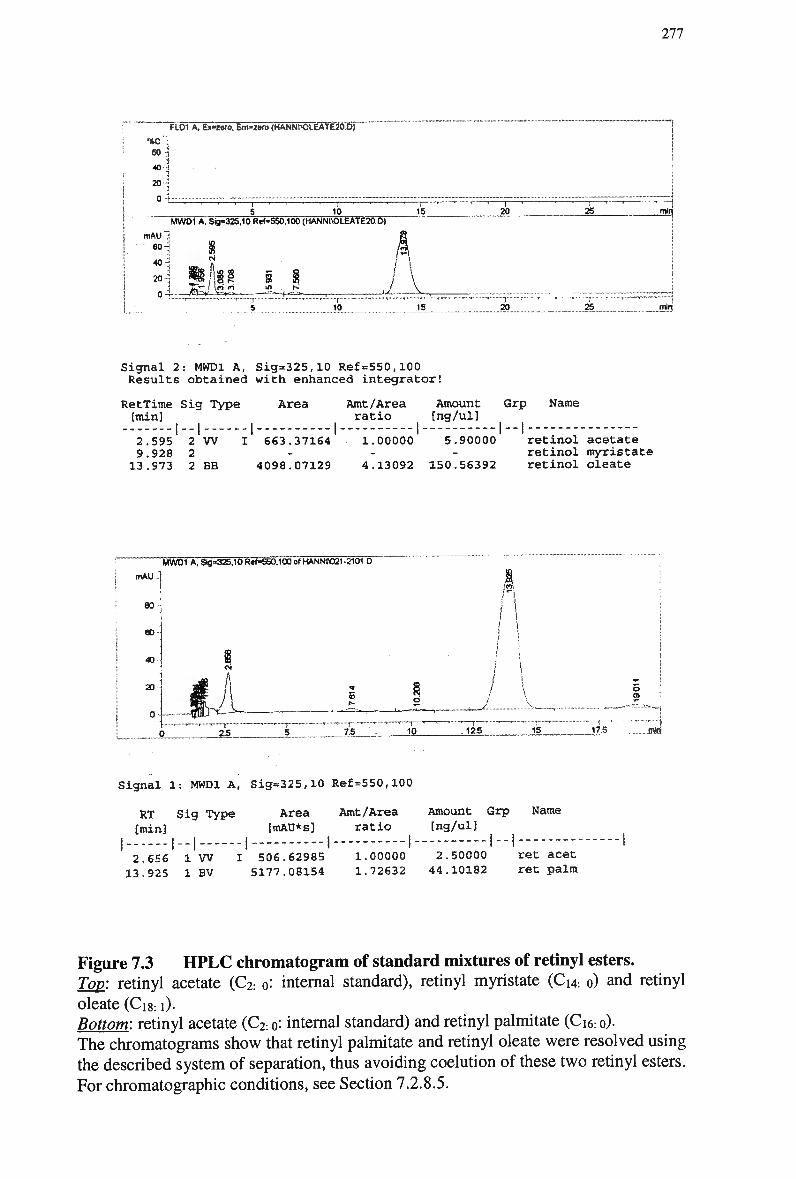
277
FID1 A. Ex=Mio, Em*2e<o(HANNUOLEATE20.D)
%C : 60 •
40 -1
70J 0 ' • ' < • 1 ' ''^ ' 1 ' ' • ' 1 " - —
5 10 15 MWOi A, Sig«325,10fte*=>650,lb0 (HANNM)LEATE20.D)
mAUl 8
" 1 8 A ' — • ' r •*•• "• •—••' r—""—•* """—'—l -"' ""'"
5 10 15
20
'"' "I" ' ''
20
2S
25
min
min
Signal 2: MWDl A, Sig=325,10 Ref=550,100 Results obtained with enhanced integrator!
RetTime Sig Type (min]
Area
I
Amt/Area Amount Grp Name ratio [ng/ul]
1-1 2.595 2 W 9.928 2 13.973 2 BB
I 663.37164
4098.07129
1.00000 5.90000
4.13092 150.56392
retinol acetate retinol myristate retinol oleate
M W D 1 A, $lg=325,10R«f«550.100of HANNIO21-2101 D
tnAU.3
80-
60
40
20
0
25 15 10
(I / \ I i
12.5 15 17.5 ..ran
Signal 1: MWDl A, Sig=325,10 Ref=S50,100
RT Sig Type [min]
Area Amt/Area fmAU*s] ratio
I
Amount Grp Name [ng/ul]
2.656 1 W I 506.62985 1.00000 2.50000 ret acet 13.925 1 BV 5177.08154 1.72632 44.10182 ret palm
Figure 7.3 H P L C chromatogram of standard mixtures of retinyl esters. Top: retinyl acetate (C2: o: internal standard), retinyl myristate (Ci4: o) and retinyl
oleate (Ci8:1). Bottom: retinyl acetate (C2: o: internal standard) and retinyl palmitate (Ci6: o). The chromatograms show that retinyl palmitate and retinyl oleate were resolved using the described system of separation, thus avoiding coelution of these two retinyl esters.
For chromatographic conditions, see Section 7.2.8.5.
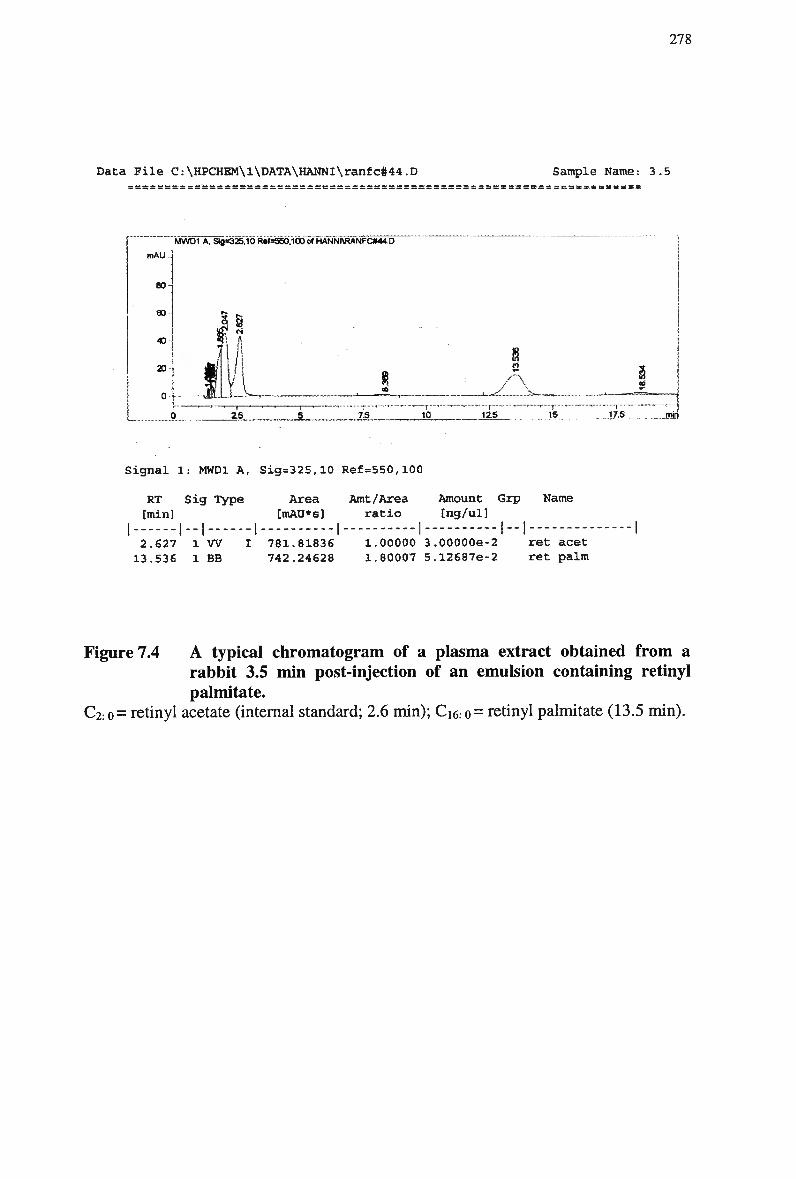
278
Data File C:\HPCHEM\l\DATA\HANNl\ranfc#44.D Sample Name: 3.5
MWD1 A, Sg*325.10 R»f=560.100 of HANNI\RANFC#44.D
mAU .
80
60
40
20
0 y
1 2S 7.S 10 125
r 15 17,5
Signal 1: MWDl A, Sig=325,10 Ref=550,l00
RT Sig Type (min]
Area Amt/Area Amount Grp Name [mAU*s] ratio [ng/ul]
l - l I I 2.627 1 W I 781.81836 1.00000 3.00000e-2 ret acet 13.536 1 BB 742.24628 1.80007 5.12687e-2 ret palm
Figure 7.4 A typical chromatogram of a plasma extract obtained from a rabbit 3.5 min post-injection of an emulsion containing retinyl palmitate.
C2: o= retinyl acetate (internal standard; 2.6 min); Ci6: o= retinyl palmitate (13.5 min).
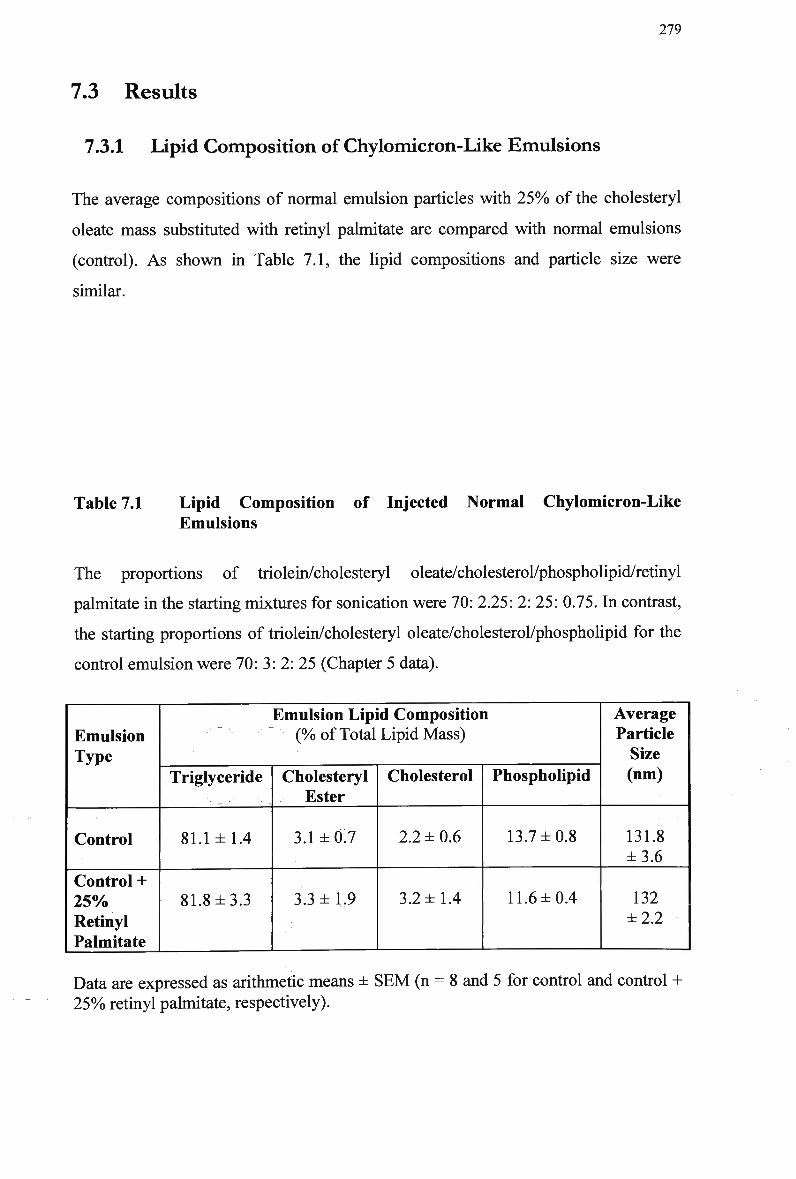
279
7.3 Results
7.3.1 Lipid Composition of Chylomicron-Like Emulsions
The average compositions of normal emulsion particles with 25% of the cholesteryl
oleate mass substituted with retinyl palmitate are compared with normal emulsions
(control). As shown in Table 7.1, the lipid compositions and particle size were
similar.
Table 7.1 Lipid Composition of Injected Normal Chylomicron-Like Emulsions
The proportions of triolein/cholesteryl oleate/cholesterol/phospholipid/retinyl
palmitate in the starting mixtures for sonication were 70: 2.25: 2: 25: 0.75. In contrast,
the starting proportions of triolein/cholesteryl oleate/cholesterol/phospholipid for the
control emulsion were 70: 3: 2: 25 (Chapter 5 data).
Emulsion Type
Control
Control + 25% Retinyl Palmitate
Emulsion Lipid Composition (% of Total Lipid Mass)
Triglyceride
81.1 ±1.4
81.8 ±3.3
Cholesteryl Ester
3.1 ±0.7
3.3 ±1.9
Cholesterol
2.2 ± 0.6
3.2 ±1.4
Phospholipid
13.7 ±0.8
11.6 ±0.4
Average Particle Size (nm)
131.8 ±3.6
132 ±2.2
Data are expressed as arithmetic means ± S E M (n = 8 and 5 for control and control +
2 5 % retinyl palmitate, respectively).

280
7.3.2 Clearance of Normal Emulsion Lipids and Retinyl Palmitate
(1 x lipid mass)
Approximately 2.5 mg of emulsion triglyceride (less than 5% of triglyceride plasma
pool) was injected into the lateral ear vein of rabbits, in a volume of 0.4-0.5 ml.
Normal emulsions hydrolysed quickly, with more than 9 6 % of particle triolein
disappearing from the plasma by 40 min after injection, followed by a rapid
disappearance of particle remnants. The A A C data was significantly greater for
triglyceride compared with the amount of cholesteryl oleate and retinyl palmitate.
However, the amount of cholesteryl oleate removed from plasma was significantly
less than that of retinyl palmitate (data not shown).
Retinyl palmitate in plasma samples was undetectable after 12 min. Similar
findings were made for the modified emulsion (data not shown). The amount of
retinyl palmitate mass injected was based on the following calculations. The mass of
retinyl palmitate in a single emulsion preparation was 0.21 m g and a total of 32 ng
injected into animal. At 8 min, with approx. 1 5 % remaining in plasma, total retinyl
palmitate is 4.9 pg, and at 14 min, with approx. 7.5% remaining in plasma, total
retinyl palmitate is 9.8 x 10" mg. In addition, the amount of retinyl palmitate
extracted was 38 ng, with approx. 1.9 ng injected onto the H P L C . Calculations were
based on the retinyl palmitate standard curve.

281
7.3.3 Plasma Kinetics and Emulsion Clearance of Control Normal
and Modified Emulsions (comparison of 1 and 3 x lipid mass)
Due to the inability to detect retinyl palmitate in plasma via HPLC, the amount of
retinyl palmitate injected into rabbit was increased. To achieve this, the following
changes were made. The total mass of emulsion lipid in each emulsion preparation
was increased (3 times), and the volume of emulsion injected was doubled. This
ensured an increase in the total lipid mass injected, whilst maintaining a constant lipid
ratio. The total mass of triglyceride injected was 1 5 % of the total triglyceride pool,
and it is known that up to 2 0 % of the total triglyceride pool can be injected without
saturating clearance kinetics in the rabbit.
It was therefore necessary to determine whether the increased lipid mass
would effect control emulsion clearance kinetics. Retinyl palmitate was not added to
the normal or modified emulsions, to reduce the number of variables being studied.
The same animals were re-used for all experiments, thus acting as controls for both
emulsion types and the mass of lipid injected. Control and experimental clearance
studies were completed on the same day, to reduce variability.
7.3.3.1 Clearance of lipids in normal chylomicron-like emulsions
The clearance profiles of normal emulsions in rabbits are presented in Table 7.2 for
A A C values. The amount of triglyceride cleared from plasma was similar for all
normal emulsions. The clearance of emulsion cholesteryl oleate from plasma in
rabbits is shown in Figure 7.5, and was similar following injection of 1 x lipid mass
compared with 3 x lipid mass, with respect to the amount of radiolabeled cholesteryl
ester cleared from plasma.
7.3.3.2 Clearance of lipids in modified chylomicron-like emulsions
The mean A A C clearance data for modified emulsion triglyceride in rabbits was
similar following injection of 1 x lipid mass and 3 x lipid mass. The clearance of
emulsion cholesteryl oleate from plasma in rabbits (Figure 7.6) was not significantly
different following injection of 1 x lipid mass or 3 x lipid mass, with respect to
amount of radiolabeled cholesteryl ester cleared from plasma.

282
Table 7.2 also compares the clearance data for normal and modified
emulsions. Following injection of emulsions containing 1 x lipid mass, the amount of
modified emulsion triolein and cholesteryl oleate remaining in plasma was
significantly different to normal emulsion triolein and cholesteryl oleate. W h e n 3 x
lipid mass was injected into rabbits, the A A C for modified emulsion triolein and
cholesteryl oleate was significantly different compared with the same parameters for
normal emulsion triolein and cholesteryl oleate.
7.3.3.3 High affinity uptake
Receptor-mediated clearance of chylomicron-like emulsions, calculated as the
difference between the clearance of normal and modified emulsion cholesteryl oleate,
is presented in Table 7.2. High affinity uptake of emulsions as calculated for A A C
values was similar following injection of 1 x lipid mass and 3 x lipid mass. Hence the
metabolism of control normal and modified emulsion triglyceride and cholesteryl
oleate in rabbits was not altered by an increase in lipid mass.
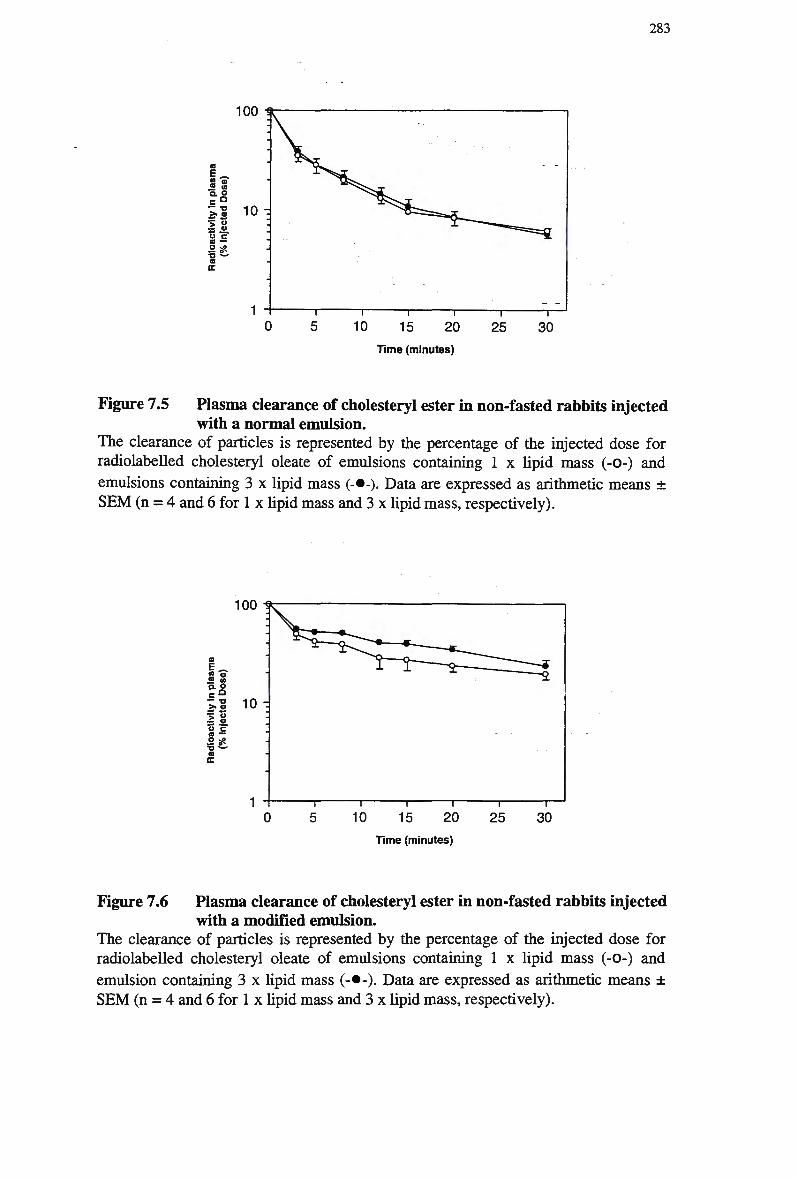
283
Time (minutes)
Figure 7.5 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a normal emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate of emulsions containing 1 x lipid mass (-0-) and
emulsions containing 3 x lipid mass (-•-). Data are expressed as arithmetic means ± S E M (n = 4 and 6 for 1 x lipid mass and 3 x lipid mass, respectively).
100
m
18 o c" ca —
it
10 15 20
Time (minutes)
Figure 7.6 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a modified emulsion.
The clearance of particles is represented by the percentage of the injected dose for radiolabeled cholesteryl oleate of emulsions containing 1 x lipid mass (-0-) and
emulsion containing 3 x lipid mass (-•-). Data are expressed as arithmetic means ± S E M (n = 4 and 6 for 1 x lipid mass and 3 x lipid mass, respectively).
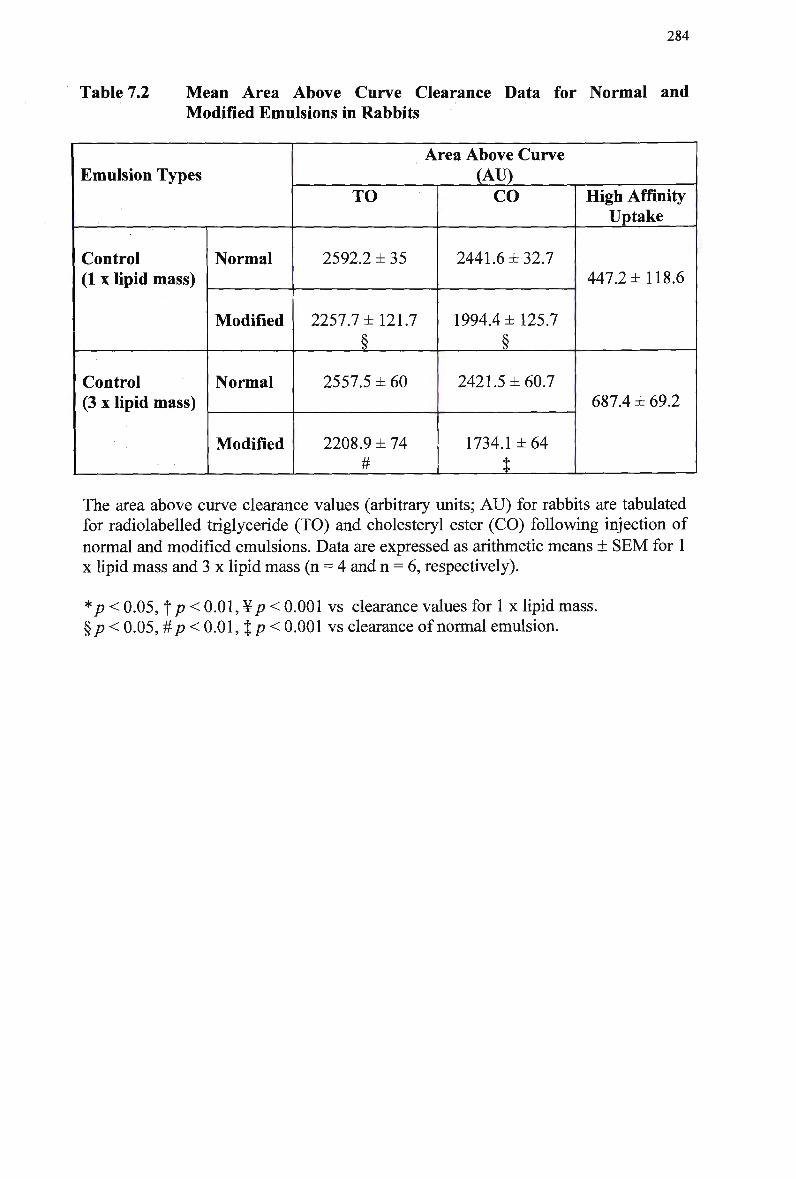
284
Table 7.2 Mean Area Above Curve Clearance Data for Normal and Modified Emulsions in Rabbits
Emulsion Types
Control (1 x lipid mass)
Control (3 x lipid mass)
Normal
Modified
Normal
Modified
Area Above Curve (AU)
TO
2592.2 ± 35
2257.7 ±121.7
§
2557.5 ± 60
2208.9 ± 74 #
CO
2441.6 ±32.7
1994.4 ±125.7
§
2421.5 ±60.7
1734.1 ±64
t
High Affinity Uptake
447.2 ±118.6
687.4 ± 69.2
The area above curve clearance values (arbitrary units; AU) for rabbits are tabulated for radiolabeled triglyceride (TO) and cholesteryl ester (CO) following injection of
normal and modified emulsions. Data are expressed as arithmetic means ± SEM for 1 x lipid mass and 3 x lipid mass (n = 4 and n = 6, respectively).
*p< 0.05, t p < 0.01, ¥ p < 0.001 vs clearance values for 1 x lipid mass. § p < 0.05, #p < 0.01, %p< 0.001 vs clearance of normal emulsion.

285
7.3.4 Organ Uptake of Lipids (comparison of 1 and 3 x lipid mass)
To avoid possible contamination of modified emulsions with unesterified cholesterol,
clearance studies using modified emulsions were completed prior to clearance studies
using normal emulsions. Therefore, at the completion of this set of clearance studies,
four rabbits were sacrificed and the liver and spleen uptake of radiolabeled
cholesteryl oleate was measured at 30 min following injection of a normal emulsion.
To determine if there was any effect of the injection of an increased lipid mass on
organ uptake, the data was compared with previous organ uptake data using 1 x lipid
mass (Chapter 3). The organ uptake values for rabbits with 1 x lipid mass and 3 x
lipid mass injected are compared in Figure 7.7.
Less than 1 % of radioactivities were recovered in the spleen, and there was a
significantly reduced amount of cholesteryl oleate taken up by the spleen following
injection of 3 x emulsion lipid mass. Mean splanchnic uptake in rabbits following
injection of 3 x lipid mass was approximately half the value for injection of the 1 x
lipid mass. Hepatic recoveries of radiolabeled cholesteryl oleate in normal emulsions
were similar following injection of the two lipid doses.
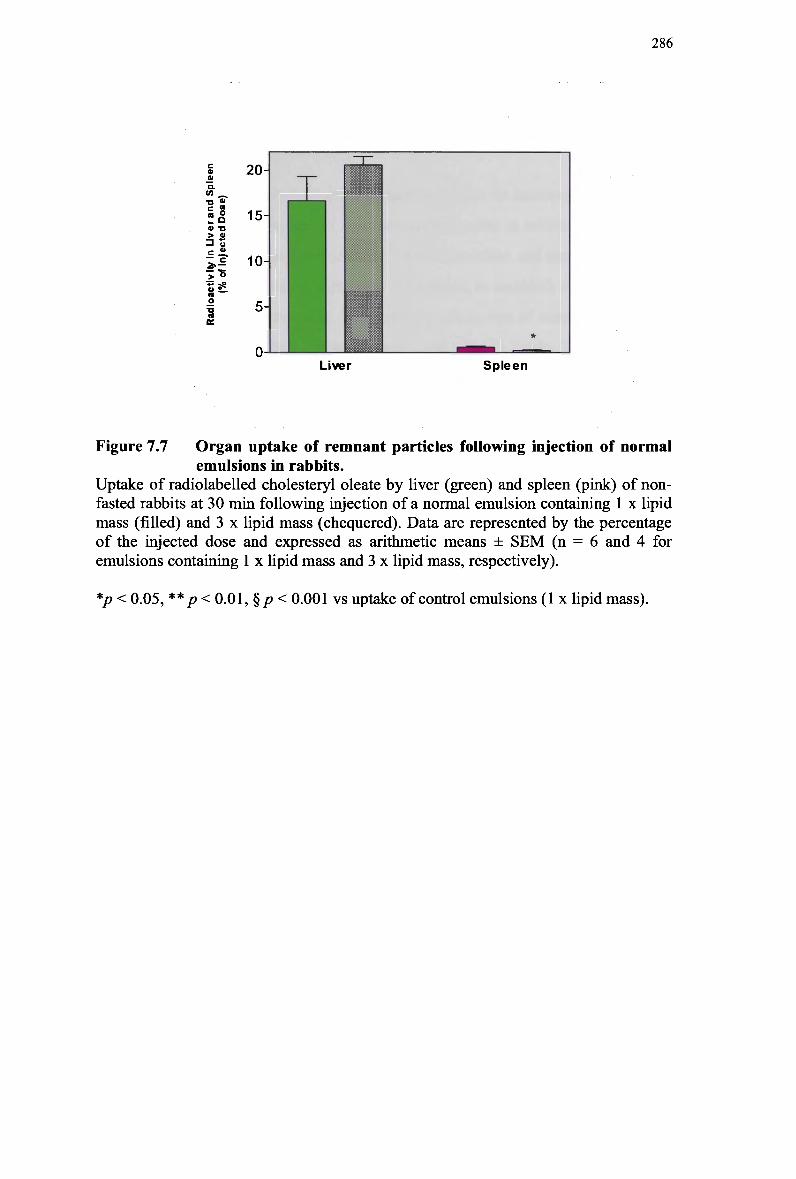
286
20-
15-
10-
5-
0 ' ™ ' ' ^ Liver Spleen
Figure 7.7 Organ uptake of remnant particles following injection of normal
emulsions in rabbits. Uptake of radiolabeled cholesteryl oleate by liver (green) and spleen (pink) of non-fasted rabbits at 30 min following injection of a normal emulsion containing 1 x lipid mass (filled) and 3 x lipid mass (chequered). Data are represented by the percentage of the injected dose and expressed as arithmetic means ± SEM (n = 6 and 4 for
emulsions containing 1 x lipid mass and 3 x lipid mass, respectively).
CO _
a -a > £ 3 o
o o
*p < 0.05, ** p < 0.01, §/> < 0.001 vs uptake of control emulsions (1 x lipid mass).

287
7.3.5 Clearance of Emulsion Lipids for Normal and Modified
Control Emulsions and Emulsions Containing 25% Retinyl
Palmitate (3 x lipid mass)
It was established in the previous set of experiments that an increase in lipid mass by
a factor of three did not alter the in vivo clearance kinetics in rabbits. The purpose of
this study was to compare the clearance of control emulsions and emulsions with 2 5 %
of cholesteryl oleate substituted with retinyl palmitate, to establish that an increase in
injected lipid mass would not alter the clearance parameters of emulsions containing
retinyl ester. T o achieve this, the following changes were made. The total mass of
emulsion lipid and retinyl palmitate in each emulsion preparation was increased by a
factor of three, and the volume of emulsion injected was doubled. Therefore, the total
amount of retinyl palmitate injected would increase, whilst maintaining a constant
retinyl palmitate: lipid ratio. The total mass of lipid injected into each rabbit was 30-
40 mg, for normal and modified emulsions.
7.3.5.1 Clearance of lipids in normal chylomicron-like emulsions
The clearance profiles for normal emulsions with and without retinyl palmitate in
rabbits are presented in Table 7.3. The normal emulsion hydrolysed rapidly, with
approximately 9 5 % of particle triolein disappearing from plasma by 30 min, however
clearance of cholesteryl oleate was slower, with more than 9 0 % of cholesteryl oleate
radioactivity remaining in plasma at 30 min (Figure 7.8). A greater amount of triolein
was cleared from plasma during the 30 min period, compared with cholesteryl oleate
(Table 7.3).
The clearance pattern of triglyceride in rabbits was not significantly different
in control emulsions or emulsions with 2 5 % retinyl palmitate incorporated, with
respect to A A C data. The amount of normal emulsion cholesteryl oleate cleared from
plasma was similar for control emulsions and emulsions with 2 5 % retinyl palmitate
incorporated (Figure 7.8).

288
7.3.5.2 Clearance of lipids in modified chylomicron-like emulsions
The clearance profiles for modified emulsions with and without retinyl palmitate in
rabbits are presented in Table 7.3. Mean A A C data for triglyceride was greater than
cholesteryl ester A A C data in rabbits. The modified emulsion hydrolysed rapidly,
with approximately 9 0 % of particle triolein disappearing from plasma by 30 min,
however clearance of modified emulsion cholesteryl oleate was impaired, with
approximately 7 5 % cholesteryl oleate cleared from plasma at 30 min (Figure 7.9).
The amount of triglyceride cleared from plasma ( A A C ) was similar for both
modified emulsions. The clearance of modified emulsion cholesteryl oleate from
plasma in rabbits was similar for control emulsions and emulsions with 2 5 % retinyl
palmitate incorporated, with respect to amount cleared from plasma.
The clearance data for normal and modified emulsions is also compared.
Following injection of control emulsions containing 3 x lipid mass, cholesteryl oleate
clearance was significantly different when comparing A A C data for normal and
modified emulsions. Following injection of emulsions containing 2 5 % retinyl
palmitate, only emulsion cholesteryl oleate clearance (AAC) was significantly less for
modified emulsion compared with normal emulsion.
7.3.5.3 High affinity uptake
Receptor-mediated clearance of chylomicron-like emulsions, calculated as the
difference between the clearance of normal and modified emulsion cholesteryl oleate
(AAC), is presented in Table 7.3. High affinity uptake was similar for control
emulsions, and emulsions with 2 5 % retinyl palmitate incorporated, following
injection of 3 x lipid mass.
Hence the overall metabolism of normal and modified emulsion triglyceride
and cholesteryl oleate in rabbits was not altered by the incorporation of 2 5 % retinyl
palmitate.
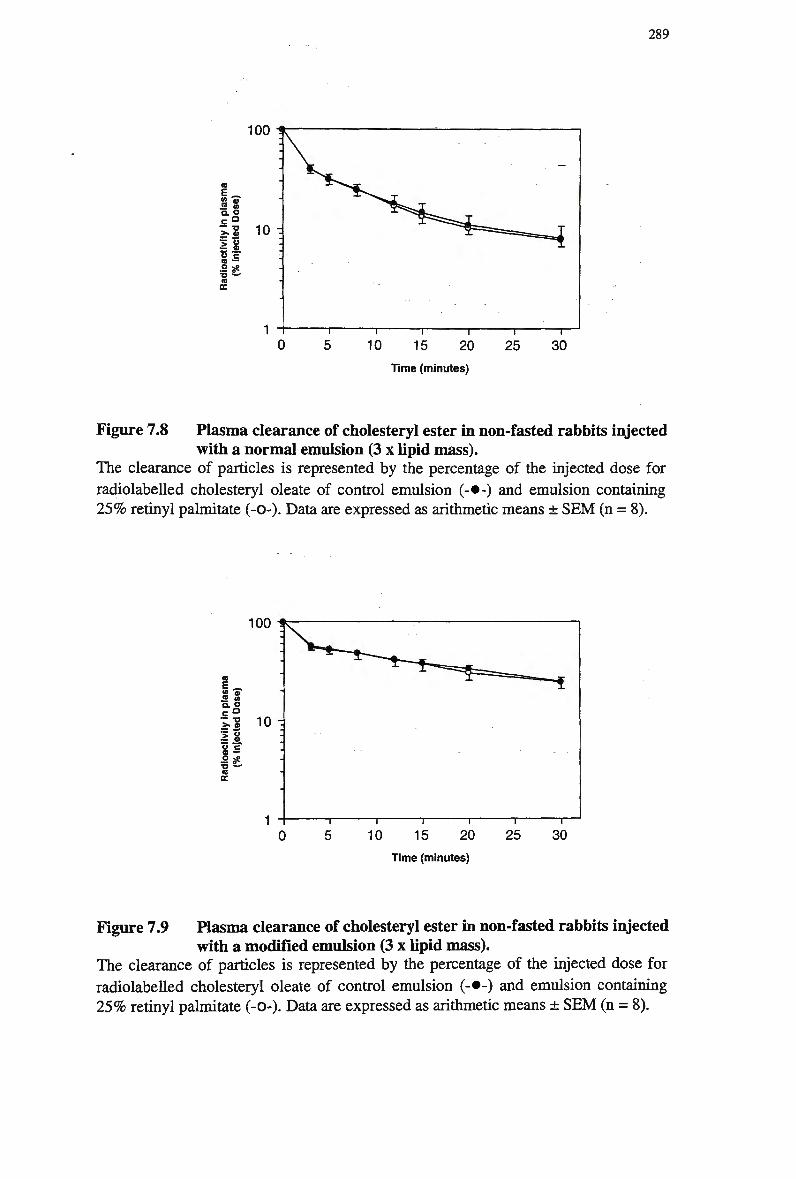
289
100
s « c a — "a
CO •••
^ • * - '
a IE
20
Time (minutes)
Figure 7.8 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a normal emulsion (3 x lipid mass).
The clearance of particles is represented by the percentage of the injected dose for
radiolabeled cholesteryl oleate of control emulsion (-•-) and emulsion containing 2 5 % retinyl palmitate (-0-). Data are expressed as arithmetic means ± S E M (n = 8).
100
£ a' S <A e- O
I £ CO —
•o -^ a CE
15
Time (minutes)
Figure 7.9 Plasma clearance of cholesteryl ester in non-fasted rabbits injected with a modified emulsion (3 x lipid mass).
The clearance of particles is represented by the percentage of the injected dose for
radiolabeled cholesteryl oleate of control emulsion (-•-) and emulsion containing 2 5 % retinyl palmitate (-0-). Data are expressed as arithmetic means ± S E M (n = 8).
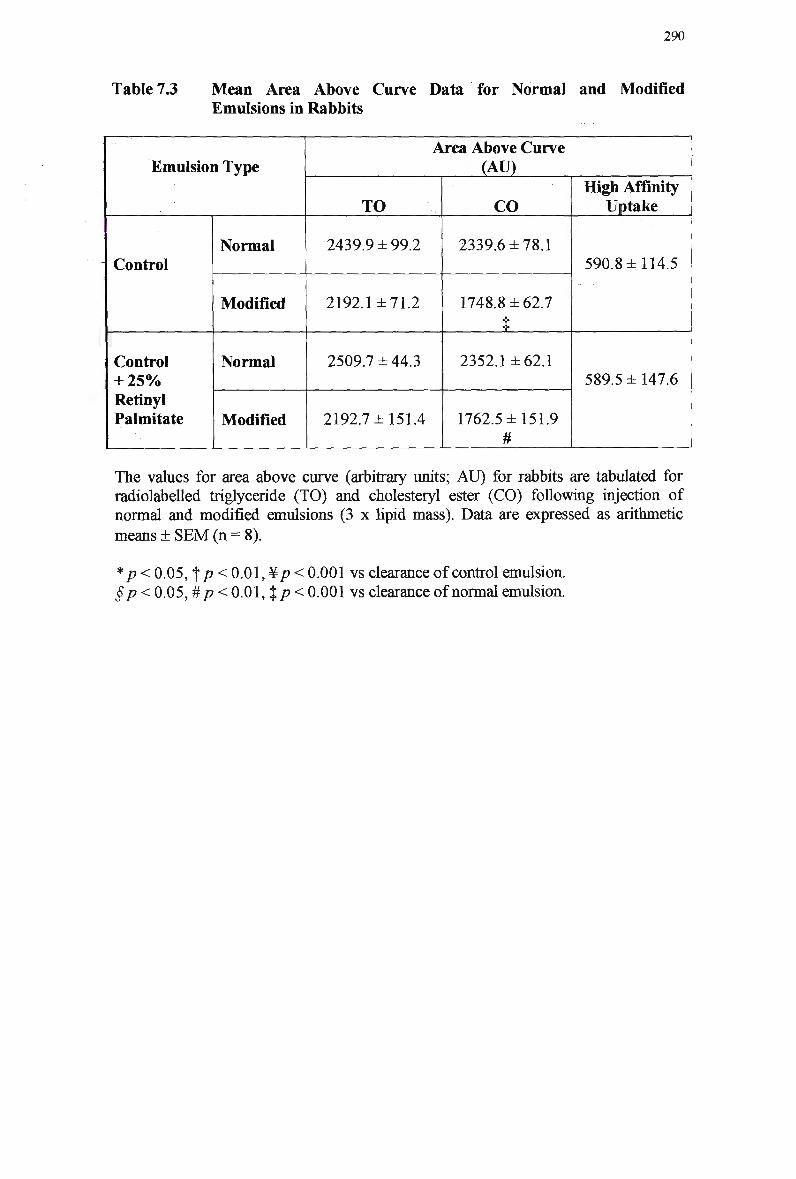
290
Table 7.3 Mean Area Above Curve Data for Normal and Modified Emulsions in Rabbits
Emulsion Type
Control
Control + 2 5 % Retinyl Palmitate
Normal
Modified
Normal
Modified
Area Above Curve (AU)
TO
2439.9 ±99.2
2192.1 ±71.2
2509.7 ±44.3
2192.7 ±151.4
CO
2339.6 ±78.1
1748.8 ±62.7
i
2352.1 ±62.1
1762.5 ±151.9 #
High Affinity Uptake
590.8 ±114.5
589.5 ±147.6
The values for area above curve (arbitrary units; AU) for rabbits are tabulated for radiolabeled triglyceride (TO) and cholesteryl ester (CO) following injection of normal and modified emulsions (3 x lipid mass). Data are expressed as arithmetic means ± S E M (n = 8).
* p < 0.05, f/? < 0.01, ¥/? < 0.001 vs clearance of control emulsion. §p < 0.05, #p< 0.01, %p< 0.001 vs clearance of normal emulsion.

291
7.3.6 Efficiency of Retinyl Ester Extraction
To date, the reliability of retinyl palmitate as a marker for chylomicron-like emulsion
remnant particles had been questionable. It appeared that although the mass in
emulsion and plasma was sufficient for detection, the concentration in plasma was
variable. At this point, the efficiency of extraction procedures and the stability of
retinyl palmitate in solution were examined.
The net recovery of retinyl esters in plasma and blood was compared with the
recovery from pure retinyl ester samples suspended in solvent. Following extraction
of the retinyl ester in all samples, it was observed that the recovery of pure retinyl
ester was much higher than recovery of retinyl ester from plasma samples, as analysed
by H P L C . The efficiency of retinyl ester extraction from emulsions containing 1 x
lipid mass and 3 x lipid mass was also investigated. Following H P L C analysis, the
extraction of retinyl palmitate and retinyl acetate was similar for emulsions containing
1 x lipid mass and 3 x lipid mass (~ 4 5 % ) .
The stability of retinyl esters over time was also investigated. Retinyl ester
was added to a fresh plasma sample immediately following venipuncture, duplicate
extractions were made and the samples were analysed. This procedure was repeated in
plasma that had been kept at -20°C and shielded from light for 48 hr. Following
H P L C analysis, the retinyl ester did not appear stable and there was evidence of
breakdown of the retinyl ester compound. Thus it appeared that retinyl esters required
immediate extraction and analysis to prevent possible breakdown.
To investigate the possibility that the suspended retinyl esters may adsorb to
the walls of the polypropylene tubes, retinyl ester samples with retinyl acetate added
as an internal standard were extracted in polypropylene and glass kimble tubes and
the extracted samples were injected onto the H P L C . The recoveries of retinyl esters
were more efficient and reliable when the extraction procedure was completed using
glass tubes (Table 7.4).
To facilitate retinyl ester detection in plasma for future clearance studies,
calculations were made to determine the mass of emulsion retinyl ester required, to
produce consistent extraction and H P L C detection. Based on the amount of
radiolabeled cholesteryl oleate remaining in plasma at specific time points and
extraction efficiency of retinyl ester in plasma, the amount of retinyl ester at the same
time points was calculated. The recovery for radiolabeled cholesteryl oleate is

292
approx. 3 0 % , thus a total of 0.18 m g retinyl palmitate in a volume of 200 pi was
injected into each rabbit. For an extraction efficiency of 8 0 % , the mass of retinyl
palmitate per plasma sample at 3 min would be ~ 0.03 pg and 0.003 pg at 20 min.
With a detection range of 0.03 - 0.003 pg and based on the calibration curve, these
amounts were calculated to be readily detectable via H P L C . However, experimental
studies of emulsion extractions via H P L C revealed that the amount of retinyl
palmitate at 5 min was 0.3 pg (~ 45 area units), representing the lower limit of
detection.
Sensitivity was increased by increasing the volume of emulsion injected, to
increase the mass of retinyl palmitate available for detection in rabbit plasma at later
time points. However, care was taken to avoid saturation of the plasma triglyceride
pool. Thus, -12 m g triglyceride (-30% triglyceride pool size) was injected, which
was equivalent to 0.14 m g retinyl palmitate. Emulsion clearance studies were repeated
with varying amounts of lipid mass and retinyl palmitate injected, to ascertain the
appropriate amounts to inject, for sufficient detection.
To further improve retinyl ester detection, blood samples of 2 ml were taken at
each time point to ensure a sufficient mass of retinyl palmitate was available for
extraction, glass tubes were used for all extractions, and the H P L C calibration curve
was adjusted to include lower concentrations. To improve the resolution of the peak
height and stabilise the retention time of retinyl esters, differing solvent solutions
were investigated as mobile phases for pure retinyl palmitate and acetate standards
injected onto H P L C . T L C confirmed the purity of all lipids and retinyl esters prior to
experimentation.
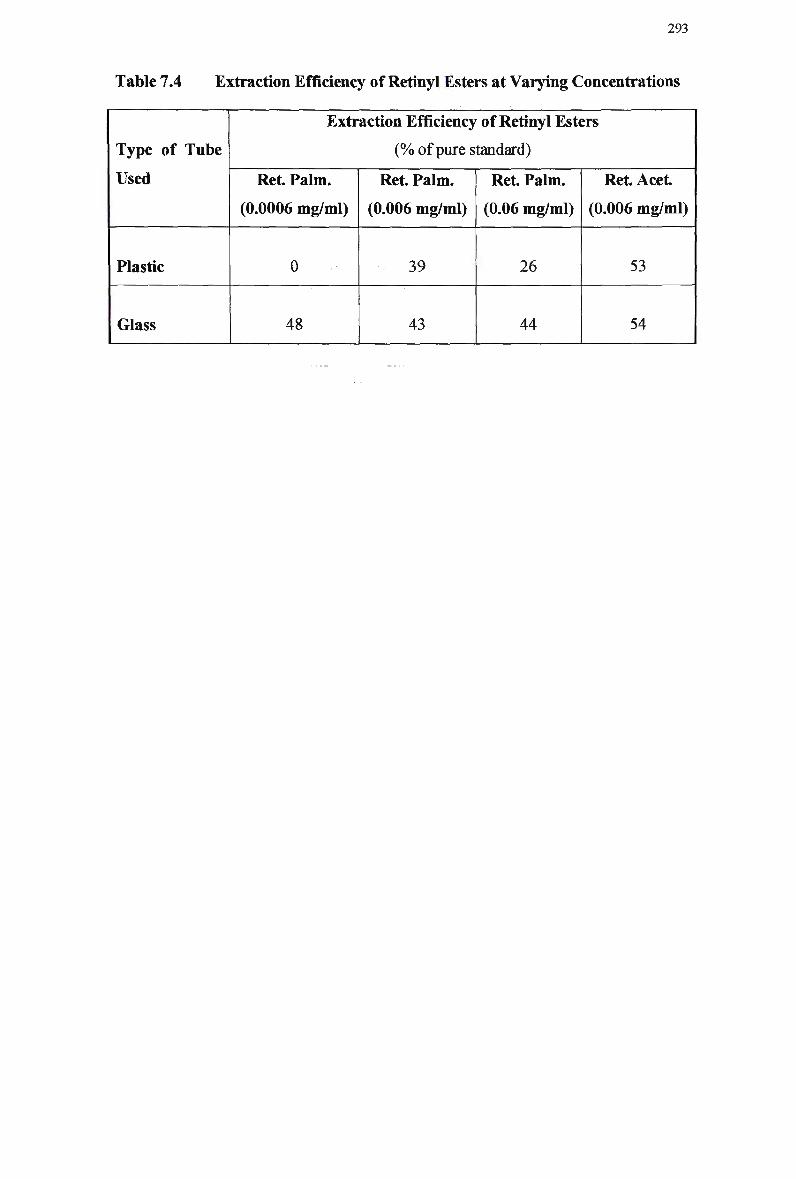
293
Table 7.4 Extraction Efficiency of Retinyl Esters at Varying Concentrations
Type of Tube
Used
Plastic
Glass
Extraction Efficiency of Retinyl Esters
(% of pure standard)
Ret. Palm.
(0.0006 mg/ml)
0
48
Ret. Palm.
(0.006 mg/ml)
39
43
Ret. Palm.
(0.06 mg/ml)
26
44
Ret. Acet
(0.006 mg/ml)
53
54

294
7.3.7 Detection of Retinyl Palmitate in Plasma Samples Using
Retinyl Acetate as an Internal Standard (3 x lipid mass)
The previous set of experiments established that an increase in the amount of lipid and
retinyl palmitate mass injected did not alter the clearance kinetics of emulsion
triglyceride or cholesteryl ester. T o assist in the detection of retinyl palmitate in
plasma, the calibration curves for retinyl palmitate were re-calculated to include lower
concentrations for detection. A calibration curve for retinyl acetate was calculated to
include concentrations similar to those of retinyl palmitate. A total of 0.3 pg retinyl
acetate was added to each plasma sample prior to extraction. A mobile phase of
methanol (100%) was used for H P L C , to reduce the retention time of retinyl palmitate
and the refine the retinyl palmitate peak. A smaller column pore size was also
employed.
7.3.7.1 Clearance of lipids in normal chylomicron-like emulsions
The clearance profiles of normal emulsions in rabbits are presented in Table 7.5. The
normal emulsion hydrolysed rapidly with approx. 9 8 % of particle triolein
disappearing from plasma by 30 min, however clearance of cholesteryl oleate and
retinyl palmitate was slower, with approx. 9 4 % remaining in plasma at 30 min. The
amount of triolein cleared from plasma was greater compared with cholesteryl oleate
and retinyl palmitate, however this was not significant (Figure 7.10).
7.3.7.2 Clearance of lipids in modified chylomicron-like emulsions
The mean AAC clearance data for modified emulsion triglyceride in rabbits was
significantly greater than for cholesteryl oleate and retinyl palmitate (Figure 7.11),
indicating a greater amount removed from plasma. The modified emulsion hydrolysed
rapidly, with approx. 9 8 % of particle triolein disappearing from plasma by 30 min. In
contrast, the clearance of cholesteryl oleate and retinyl palmitate was delayed, with
approx. 9 1 % cleared from plasma at 30 min.
Table 7.5 also compares the clearance data for normal and modified
emulsions. The amount of modified emulsion cholesteryl oleate remaining in plasma
was significantly greater to normal emulsion. The A A C data for modified emulsion

295
retinyl palmitate was less, however this was not significant. The amount of triolein
cleared was similar for both emulsions.
7.3.7.3 High affinity uptake
Receptor-mediated clearance of chylomicron-like emulsions, calculated as the
difference between the clearance of normal and modified emulsion cholesteryl oleate
and retinyl palmitate, is presented in Table 7.5. There was no significant difference in
the high affinity uptake of emulsion cholesteryl oleate and retinyl palmitate. Hence
the metabolism of normal and modified emulsion cholesteryl oleate and retinyl
palmitate in rabbits was similar, and retinyl palmitate appears to act as an appropriate
marker for chylomicron remnant particle clearance for both emulsion types.
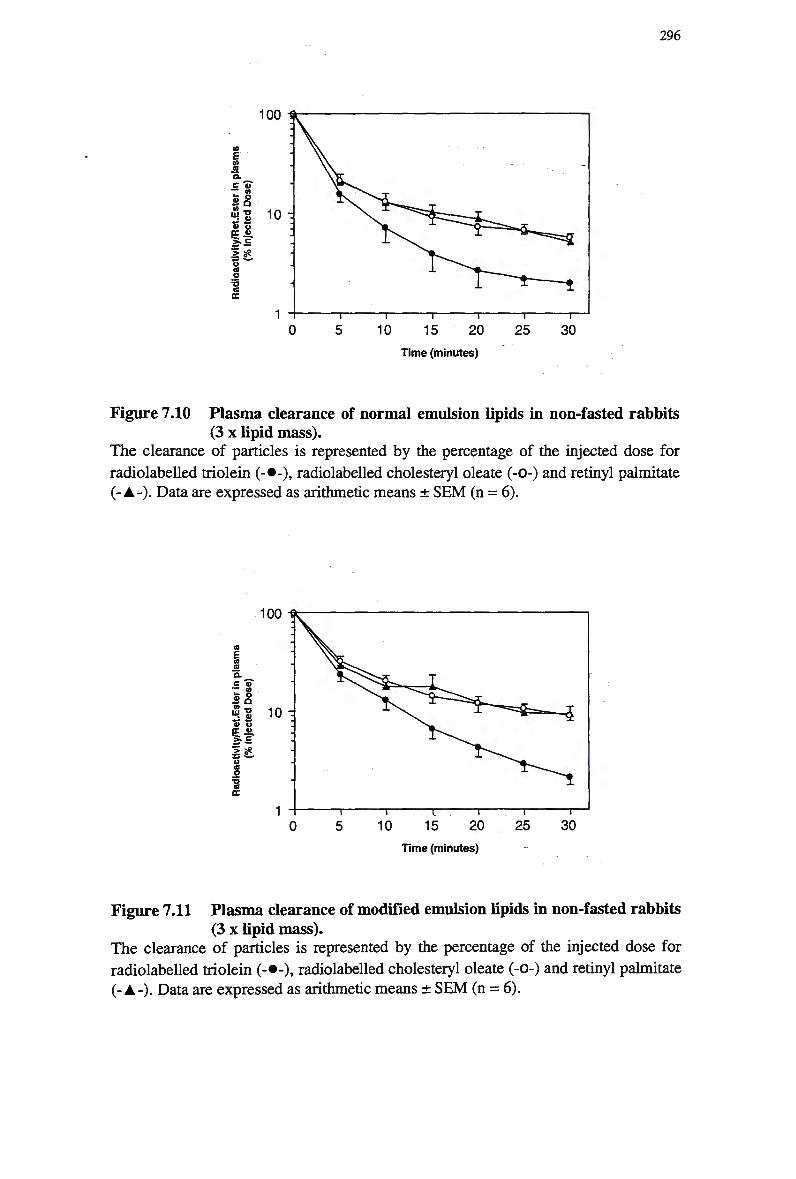
296
Time (minutes)
Figure 7.10 Plasma clearance of normal emulsion lipids in non-fasted rabbits (3 x lipid mass).
The clearance of particles is represented by the percentage of the injected dose for
radiolabeled triolein (-•-), radiolabeled cholesteryl oleate (-0-) and retinyl palmitate (-A-). Data are expressed as arithmetic means ± S E M (n = 6).
Time (minutes)
Figure 7.11 Plasma clearance of modified emulsion lipids in non-fasted rabbits (3 x lipid mass).
The clearance of particles is represented by the percentage of the injected dose for
radiolabeled triolein (-•-), radiolabeled cholesteryl oleate (-0-) and retinyl palmitate (-A-). Data are expressed as arithmetic means ± S E M (n = 6).
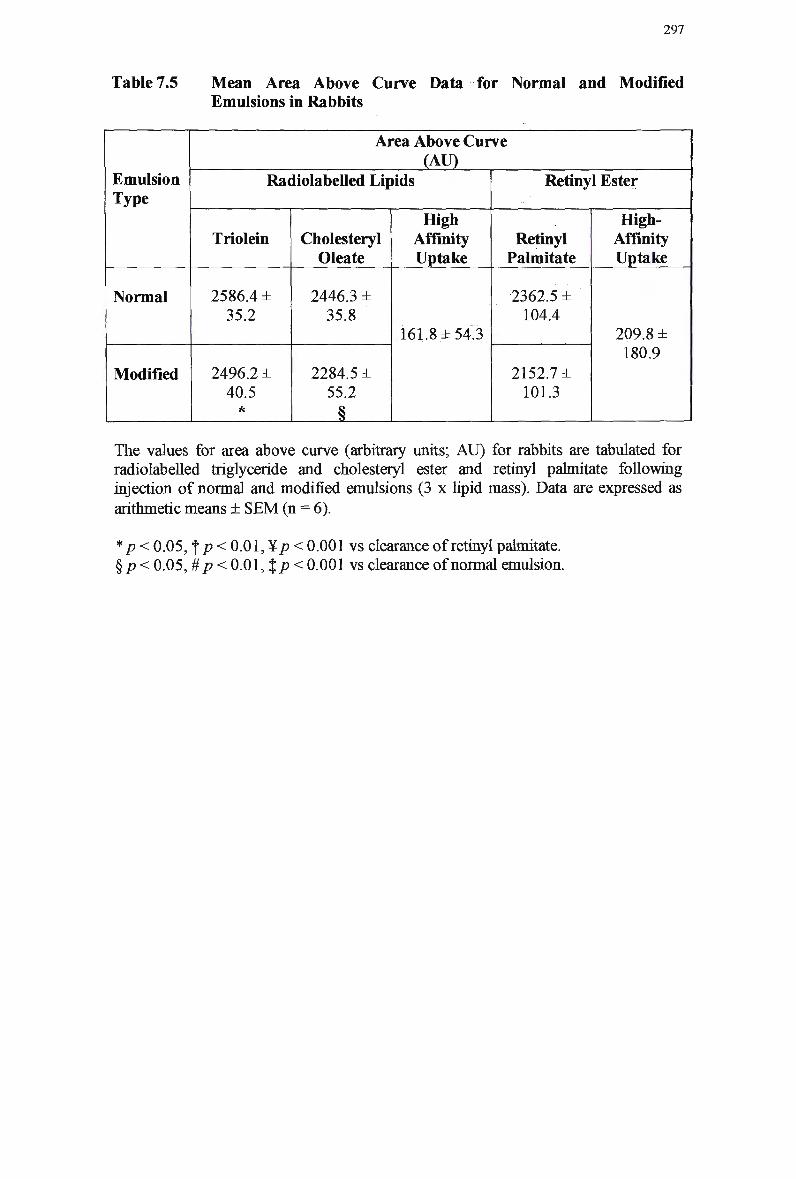
297
Table 7.5 Mean Area Above Curve Data for Normal and Modified Emulsions in Rabbits
Emulsion Type
Normal
Modified
Area Above Curve (AU)
Radiolabeled Lipids
Triolein
2586.4 ± 35.2
2496.2 ± 40.5
Cholesteryl Oleate
2446.3 ± 35.8
2284.5 ± 55.2
High Affinity Uptake
161.8 ±54.3
Retinyl Ester
Retinyl Palmitate
2362.5 ± 104.4
2152.7 ± 101.3
High-Affinity Uptake
209.8 ± i 180.9 ;
The values for area above curve (arbitrary units; AU) for rabbits are tabulated for radiolabeled triglyceride and cholesteryl ester and retinyl palmitate following injection of normal and modified emulsions (3 x lipid mass). Data are expressed as arithmetic means ± S E M (n = 6).
*p< 0.05, f p < 0.01, ¥/? < 0.001 vs clearance of retinyl palmitate. § p < 0.05, #p < 0.01, % p < 0.001 vs clearance of normal emulsion.

298
7.3.8 Investigation of Alternate Retinyl Esters as Tracees for
Chylomicron-Like Emulsions
7.3.8.1 Retinyl stearate as a marker for chylomicron-like emulsions
Retinyl stearate was synthesised by the Department of Chemistry at the University of
Western Australia according to the acyl chloride method of Huang and Goodman
(Huang and Goodman, 1965), as described in Chapter 6. The mass and purity of
retinyl stearate was confirmed by examination of their ultraviolet absorption spectra
and TLC. Commercially available retinyl palmitate contains butylated
hydroxytoluene (BHT) to prevent oxidation, however the commissioned retinyl ester
compounds did not contain B H T . The addition of B H T to retinyl ester preparations
during synthesis was not investigated due to time restraints. Chylomicron-like
emulsions with 2 5 % of cholesteryl oleate substituted with retinyl stearate were
synthesised as described in Chapter 6. Emulsions were similar to those previously
synthesised, with respect to particle size (134 ± 3 nm).
Data from clearance studies in rabbits, using retinyl stearate as a marker for
normal and modified emulsions clearly showed that the clearance of retinyl stearate
did not reflect the clearance of radiolabeled cholesteryl oleate (n = 9 and 5 for normal
and modified emulsions, respectively; data not shown). Despite repeated purification
of the retinyl stearate compound, the sample continued to disintegrate, as confirmed
by examination of their ultraviolet absorption spectra and TLC. In view of the
previous retinyl palmitate clearance data, the results suggest that retinyl stearate was
an unsuitable marker for monitoring remnant particle uptake.
7.3.8.2 Retinyl oleate as a marker for chylomicron-like emulsions
The suitability of retinyl oleate as an appropriate and reliable alternative marker for
particle uptake was explored. Retinyl oleate was synthesised by the Department of
Chemistry at the University of Western Australia, using the method described above.
Appropriate calibration curves were set up and the retinyl ester assayed for purity and
mass. Ongoing difficulties with the rapid disintegration of retinyl oleate were faced as
assessed by T L C and U V absorptive spectra, regardless of storage or handling
methods. In addition, the clearance of retinyl oleate did not reflect the clearance of

299
normal emulsion cholesteryl oleate in rabbits (n = 6, data not shown). Thus it does not
appear that retinyl oleate is a suitable marker for chylomicron-like emulsion particles.
7.3.8.3 Retinyl myristate as a marker for modified chylomicron-like emulsions
Retinyl myristate was synthesised by the Department of Chemistry at the University
of Western Australia, using the method of Lentz et al. (Lentz et al, 1975) described
in Chapter 6. Appropriate calibration curves were set up and the retinyl ester assayed
for purity and mass. The newly synthesised retinyl myristate was incorporated into
modified chylomicron-like emulsions, as described in Chapter 6. The clearance of
emulsions containing retinyl myristate was studied in rabbits, following injection of
approx. 12 m g triglyceride.
The clearance of retinyl myristate did not reflect the clearance of radiolabeled
cholesteryl oleate, as assessed in rabbits (n = 4, data not shown). To investigate the
properties of retinyl myristate, a comprehensive analysis of retinyl myristate was
undertaken, and compared with the properties of emulsion lipids.
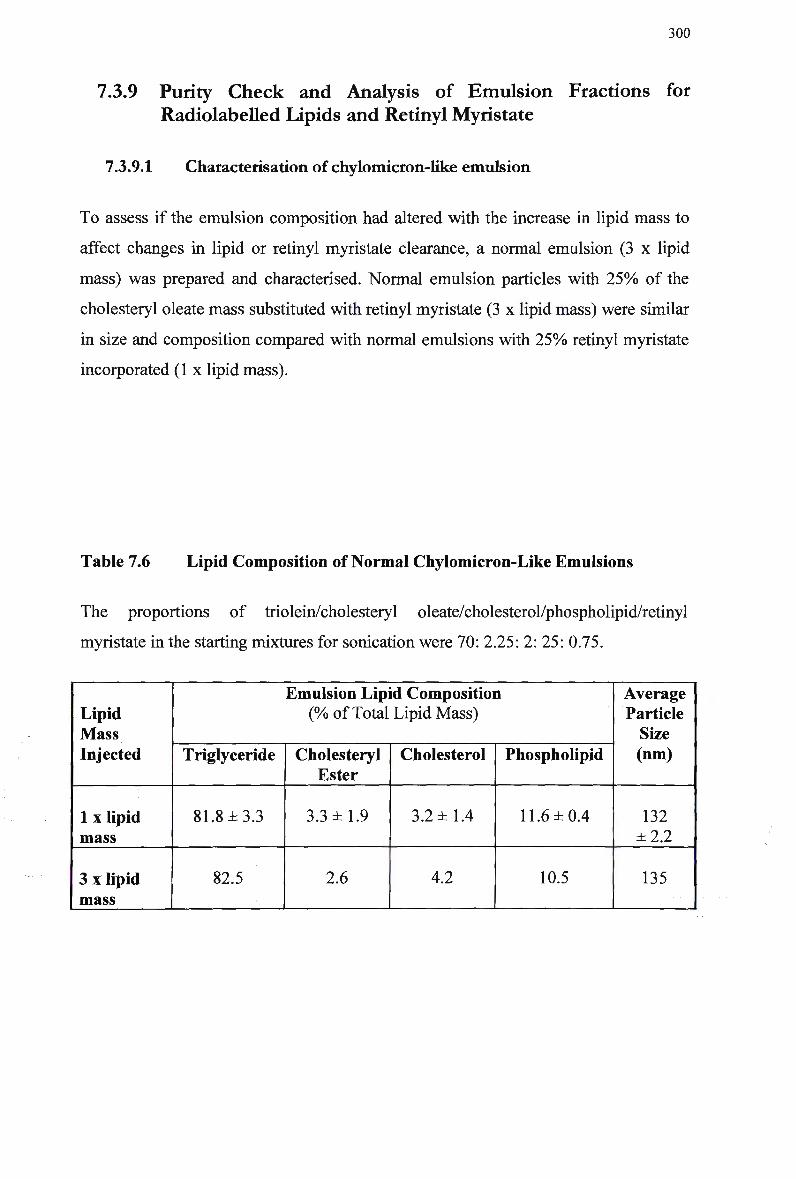
300
7.3.9 Purity C h e c k and Analysis of Emulsion Fractions for
Radiolabeled Lipids and Retinyl Myristate
7.3.9.1 Characterisation of chylomicron-like emulsion
To assess if the emulsion composition had altered with the increase in lipid mass to
affect changes in lipid or retinyl myristate clearance, a normal emulsion (3 x lipid
mass) was prepared and characterised. Normal emulsion particles with 2 5 % of the
cholesteryl oleate mass substituted with retinyl myristate (3 x lipid mass) were similar
in size and composition compared with normal emulsions with 2 5 % retinyl myristate
incorporated (1 x lipid mass).
Table 7.6 Lipid Composition of Normal Chylomicron-Like Emulsions
The proportions of triolein/cholesteryl oleate/cholesterol/phospholipid/retinyl
myristate in the starting mixtures for sonication were 70: 2.25: 2: 25: 0.75.
Lipid Mass Injected
1 x lipid mass
3 x lipid mass
Emulsion Lipid Composition (% of Total Lipid Mass)
Triglyceride
81.8 ±3.3
82.5
Cholesteryl Ester
3.3 ±1.9
2.6
Cholesterol
3.2 ±1.4
4.2
Phospholipid
11.6 ±0.4
10.5
Average Particle Size (nm)
132 ±2.2
135

301
7.3.10 Efficiency of Retinyl Ester Extraction
To determine the net recovery of retinyl esters in plasma and compare the efficiency
of extraction with the recovery from pure retinyl ester samples suspended in solvent,
retinyl myristate and retinyl acetate were added to fresh plasma samples. Following
H P L C analysis, the extraction of retinyl myristate and retinyl acetate from plasma
samples was 6 6 % and 7 4 % of pure standards, respectively. Analysis of retinyl esters
by T L C showed retinyl myristate on a number of lipid bands, suggesting that the
sample was not pure. This finding was confirmed by purity analysis of a range of
retinyl myristate concentrations by examination of their ultraviolet absorption spectra
and calculation of the retinyl ester mass. All samples displayed evidence of extensive
disintegration (1/3 - 1/10 original concentration), compared with retinyl acetate,
which was pure. Retinol suspended in free alcohol solutions is sensitive to oxidation,
therefore all assays were repeated using a range of fresh solutions, with similar
results. A sample chromatogram of an emulsion containing retinyl myristate is shown
in Figure 7.12 and demonstrates possible disintegration of the compound (shown as
minor peaks).
7.3.11 Analysis of Emulsion Fractions During Synthesis
A clearance study in rabbits was undertaken using the same emulsion as above, and
retinyl myristate was undetectable in all plasma samples. Finally, emulsion fractions
were analysed for retinyl myristate content and compared with the recovery of
radiolabeled cholesteryl oleate, to determine the extent of retinyl myristate
incorporation into the emulsion and the distribution of emulsion tracees during
emulsion synthesis. The preliminary aspiration is the large, coarsely emulsified
particles that were removed from the top of the gradient and replaced with 1.006
gm/ml solution. The emulsion fraction represents the particles that floated to the
surface and were removed following a second centrifugation. The infranatant is the
solution remaining after the emulsion fraction was aspirated; this was frozen at -80°C
for 4 hr, and saponified under vacuum to eliminate all fluids, prior to extraction. For
each fraction, the radiolabeled cholesteryl oleate was determined by liquid-
scintillation counting in a toluene-based fluor with auto quench correction (Beckman
LS3800 liquid-scintillation counter) and retinyl myristate was extracted and assayed
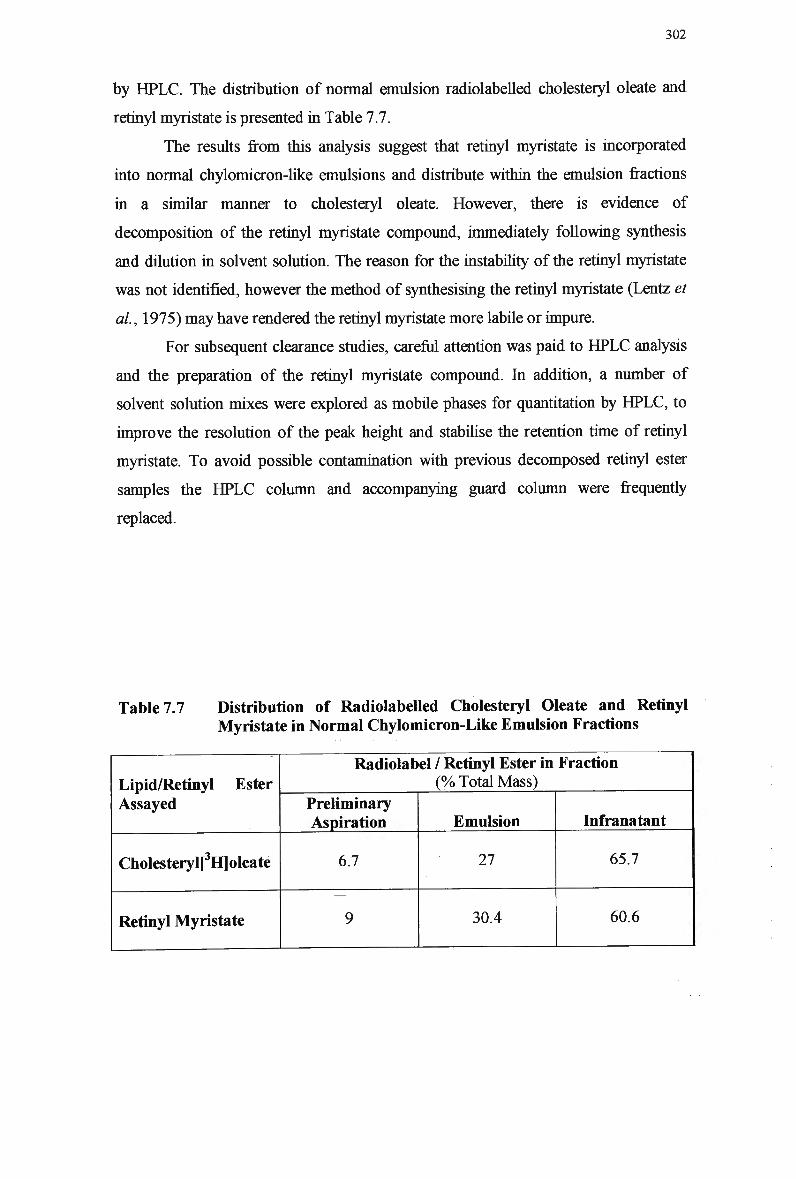
302
by H P L C . The distribution of normal emulsion radiolabeled cholesteryl oleate and
retinyl myristate is presented in Table 7.7.
The results from this analysis suggest that retinyl myristate is incorporated
into normal chylomicron-like emulsions and distribute within the emulsion fractions
in a similar manner to cholesteryl oleate. However, there is evidence of
decomposition of the retinyl myristate compound, immediately following synthesis
and dilution in solvent solution. The reason for the instability of the retinyl myristate
was not identified, however the method of synthesising the retinyl myristate (Lentz et
al., 1975) may have rendered the retinyl myristate more labile or impure.
For subsequent clearance studies, careful attention was paid to H P L C analysis
and the preparation of the retinyl myristate compound. In addition, a number of
solvent solution mixes were explored as mobile phases for quantitation by HPLC, to
improve the resolution of the peak height and stabilise the retention time of retinyl
myristate. To avoid possible contamination with previous decomposed retinyl ester
samples the H P L C column and accompanying guard column were frequently
replaced.
Table 7.7 Distribution of Radiolabeled Cholesteryl Oleate and Retinyl Myristate in Normal Chylomicron-Like Emulsion Fractions
Lipid/Retinyl Ester Assayed
Cholesteryl[3H]oleate
Retinyl Myristate
Radiolabel / Retinyl Ester in Fraction (% Total Mass)
Preliminary Aspiration
6.7
9
Emulsion
27
30.4
Infranatant
65.7
60.6

303
Data Pile C:\HPCHEM\1\DATA\HANNI\19199RM\19199014.D WFC eraul 50 ul
FLD1A. EX=Z«O, Em=2ero(HANNt\19199RM\l9169014.D)
30
20H 1
10 T 4 = 10
'TIS
MW01 A. Slg*32S,10R*f«5SO,100{HANNI\19199RM\1919»14.D)
mAO'l $ & * 30 i
Ana: 832.611
20 — j — , — T . . . . . . ^ . _ . . ^ _ . . . I
25 30 -jsi
o-fc: 10 —r-
15 20 ! ._T_
30
Signal 2: MWDl A, Sig=325,10 Ref-550,100 Results obtained with enhanced integrator!
RetTime Sig Type [min]
Area Amt/Area ratio
Amount [ng/ul]
Grp Name
1 - - I 2.S29 2 W 11.146 2 MM 15.437 2
617.82648 832.61127
1.00000 2.38143
5.90000 18.93497
retinol acetate retinol myristate retinol palmitate
Figure 7.12 A typical H P L C chromatogram of a normal emulsion containing retinyl myristate.
C2: o = retinyl acetate (internal standard; 2.5 min); Cu. 0 = retinyl myristate (11.1 min). Retinyl myristate was added to the emulsion during preparation, and pure standard was added during extraction of 50 pi emulsion. The chromatogram demonstrates possible disintegration of the compound (shown as minor peaks). For chromatographic conditions, see Section 7.2.8.5.

304
7.3.12 Clearance of Normal Emulsion Lipids and Retinyl Myristate
in Rabbits
Retinyl myristate was synthesised by the Department of Chemistry at the University
of Western Australia, using the method of Huang and Goodman (Huang and
Goodman, 1965) described in Chapter 6. Appropriate calibration curves were set up
and retinyl myristate assayed for purity and mass. The mass of emulsion lipid and
retinyl myristate to be injected in rabbits was recalculated, based on extraction
efficiencies of individual retinyl esters, and previous clearance data including H P L C
results.
The newly synthesised retinyl myristate was incorporated into normal
chylomicron-like emulsions. Sensitivity of retinyl ester detection was enhanced by
increasing the volume of emulsion injected, and hence the mass of retinyl myristate
available for detection in rabbit plasma (12-16 m g triglyceride). The mean diameter of
normal emulsions was 138 ± 3 nm.
7.3.12.1 Clearance of lipids in normal chylomicron-like emulsions
The mean AAC values for normal emulsions in rabbits are presented in Table 7.8.
Hydrolysis of the emulsion was rapid, with more than 9 6 % of particle triolein
disappearing from plasma by 30 min. The plasma clearance of retinyl myristate was
similar to radiolabeled cholesteryl oleate, with approx. 9 2 % of both markers cleared
from plasma at 30 min. The amount of triolein cleared from plasma was greater
compared with cholesteryl oleate and retinyl myristate, however this was not
significant (Figure 7.13).
The metabolism of normal emulsion cholesteryl oleate and retinyl myristate in
rabbits was similar, and retinyl myristate appears to act as an appropriate marker for
chylomicron remnant particle clearance. Retinyl myristate was not explored as a
marker for modified chylomicron-like emulsions, as it was previously established that
retinyl palmitate is suitable as a marker for these emulsion remnant particles (Section
7.3.5.2).
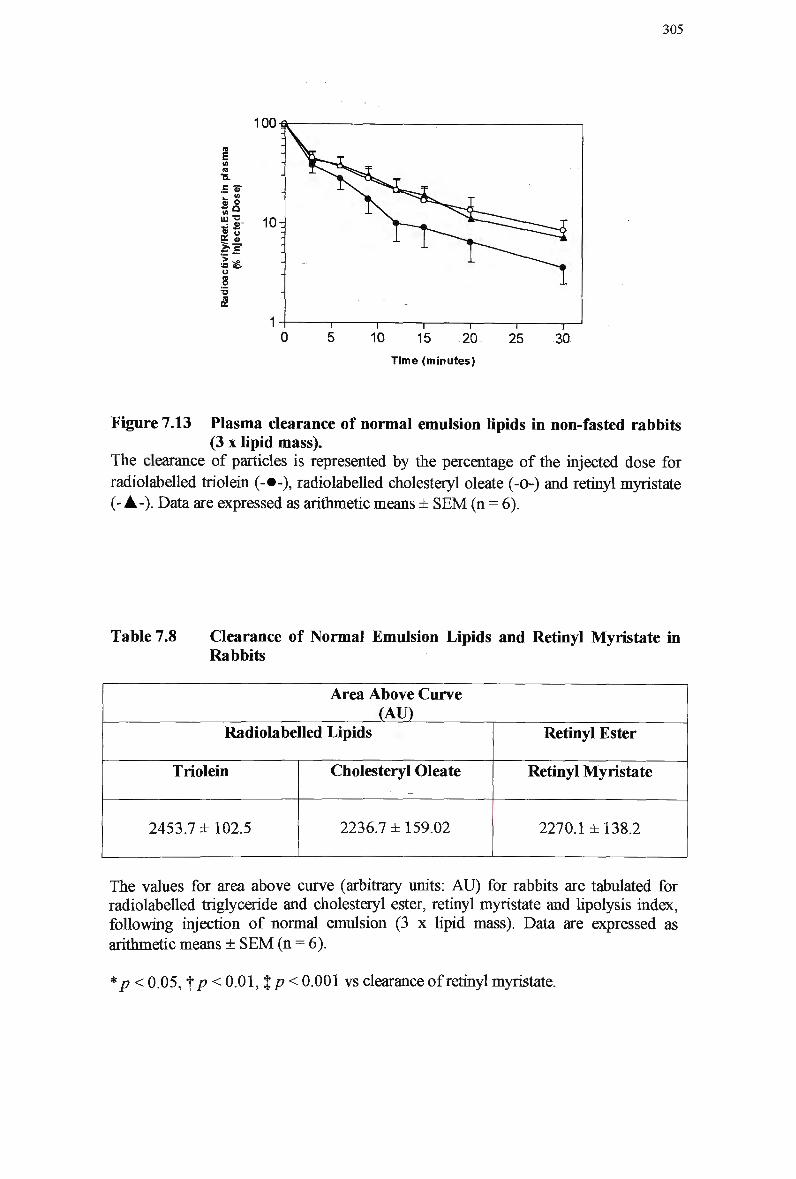
305
100
E ™
•- s SU O
»S 111 T3
« O
*=" > =
Time (minutes)
Figure 7.13 Plasma clearance of normal emulsion lipids in non-fasted rabbits (3 x lipid mass).
The clearance of particles is represented by the percentage of the injected dose for
radiolabeled triolein (-•-), radiolabeled cholesteryl oleate (-0-) and retinyl myristate (-A-). Data are expressed as arithmetic means ± S E M (n = 6).
Table 7.8 Clearance of Normal Emulsion Lipids and Retinyl Myristate in Rabbits
Area Above Curve (AU)
Radiolabeled Lipids
Triolein
2453.7 ± 102.5
Cholesteryl Oleate
2236.7 ±159.02
Retinyl Ester
Retinyl Myristate
2270.1 ± 138.2
The values for area above curve (arbitrary units: A U ) for rabbits are tabulated for radiolabeled triglyceride and cholesteryl ester, retinyl myristate and lipolysis index, following injection of normal emulsion (3 x lipid mass). Data are expressed as arithmetic means + S E M (n = 6).
*p < 0.05, f p < 0.01, % p < 0.001 vs clearance of retinyl myristate.

306
7.4 Discussion
The finding that the incorporation of various retinyl esters into chylomicron-like
emulsions did not alter clearance kinetics in vivo (Chapter 6) enabled the continuation
of the primary aim of the project, which involves the development of a diagnostic
assay using retinyl esters as tracers for normal and modified emulsion particles. In this
set of experiments, it was established that retinyl myristate and palmitate were
suitable tracees to monitor the plasma clearance of normal and modified chylomicron-
like emulsions, respectively. Retinyl stearate and retinyl oleate were also investigated
as potential emulsion tracees, however they proved unstable.
This set of studies summarises the quantitation of chylomicron remnant
clearance in rabbits using chylomicron-like emulsions with retinyl esters incorporated
and injected intravenously as a bolus dose into recipient animals. Retinyl myristate
and retinyl palmitate incorporated into normal and modified chylomicron-like
emulsions, respectively, paralleled the clearance of radiolabeled cholesteryl oleate in
rabbits. The clearance of retinyl ester-labelled emulsion remnant particles in rabbit
plasma was detectable up to 30 min, following injection of both emulsion types. The
initial rapid disappearance of retinyl ester (and cholesteryl oleate) on the clearance
curve could not be explained by intravascular processes such as remnant formation or
transfer of label, since such a process would not lower retinyl palmitate
concentrations. Instead, the rapid clearance of emulsions following intravenous
injection suggests that the retinyl esters remain with radiolabeled lipoproteins of d <
1.006 gm/ml and are distributed within the intravascular compartment prior to hepatic
uptake, and thus represent elimination of chylomicron remnants. Hence the
requirements of retinyl palmitate and retinyl myristate as markers of emulsions and
their remnants were fulfilled.
The clearance of normal emulsions fitted the known mechanism by which
chylomicrons are catabolised in rat and rabbit animal models. A delay in the remnant
particle uptake of the modified emulsion compared with the normal emulsion was
evident in the present clearance studies. High affinity (receptor) clearance was
quantitated by difference in clearance of normal and modified emulsions utilising
retinyl palmitate as a tracer, and confirmed by simultaneous monitoring of cholesteryl
oleate clearance. The refinement of retinyl myristate as a second tracer should allow

307
simultaneous assessment of the two emulsion types and quantitation of high-affinity
uptake processes in vivo.
The use of retinyl palmitate as an endogenous label of the core of
chylomicrons and their remnants has received wide recognition and acceptance (Berr,
1992), (Krasinski et al, 1990a), (Ross and Zilversmit, 1977), (Brenninkmeijer et al,
1987), (Hazzard and Bierman, 1976). While retinyl esters have previously been used
to monitor remnant particle clearance in humans via the vitamin A fat loading test, the
incorporation of retinyl esters into emulsions, bolus injection into the circulation and
their subsequent detection in plasma is a novel approach to monitoring chylomicron
metabolism. In addition, it allows for quantitation of receptor activity in vivo. Blaner
et al (Blaner et al, 1994) have previously used emulsions labelled with [3H]retinoid
to investigate whether L P L increases retinoid uptake from lipid-containing particles in
vitro, however the emulsion contained triolein and phosphatidylcholine and was not 1 O
representative of chylomicron composition. Similarly, tracer doses of stearic[ C]acid
(18:0) and palmitic[13C]acid (16:0) have been loaded into an emulsion and orally
administered (Rhee et al, 1997). The emulsions contained fatty acids administered as
free acids to facilitate the absorption of the saturated fatty acids in vivo, but did not
contain other components found in chylomicron particles.
Preliminary studies demonstrated that there was insufficient emulsion retinyl
ester present for detection in plasma samples. This inability to detect retinyl ester was
possibly due to inadequate retinyl palmitate mass in the chylomicron-like emulsion,
resulting in a small mass of retinyl palmitate injected into the rabbit. Consequently,
clearance studies were completed utilising varying quantities of chylomicron-like
emulsion lipid and retinyl ester, whilst retaining a constant lipid proportion. In
addition, changes were made to the H P L C apparatus and extraction procedures to
facilitate efficient detection.
The effect of an increase in retinyl ester and lipid mass on clearance kinetics
was investigated in rabbits. A lipid mass up to 16 m g triglyceride and 0.18 m g retinyl
palmitate was injected per rabbit (approx. 3 0 % triglyceride pool size). The proportion
of lipid cleared from plasma was not changed by the increase in lipid mass. Therefore,
it was concluded that the metabolism of normal or modified emulsions in rabbits was
not altered, and the lipid uptake pathways as evidenced by the clearance curves, did
not appear to be saturated. The formation of chylomicron remnant particles by L P L
did not appear rate limiting for remnant removal, even following injection of a higher

308
lipid mass. In fact, the pattern of removal reflected normal chylomicron metabolism,
i.e., triglyceride clearance is faster because removal is via two processes, lipolysis
within the plasma to form triglyceride depleted remnants, followed by remnant uptake
and removal of residual triolein, whilst cholesteryl ester clearance occurs during
remnant uptake.
Previous vitamin A fat-loading studies in rabbits have involved the
administration of 50,000 RJ (15 m g ) vitamin A via gastric intubation (Beaumont and
Assadollahi, 1990), with no apparent saturation of chylomicron clearance
mechanisms. Demacker et al. (Demacker et al, 1992) fed 13.7 m g vitamin A mixed
with 25 g m rabbit chow to W H H L and N Z W rabbits. In the same vitamin A fat
loading study, rabbits were given 27.4 m g vitamin A intragastrically. It is difficult to
compare the amount of emulsion retinyl palmitate injected per rabbit with vitamin A
fat loading studies, as emulsion clearance is independent of digestion, vitamin A
processing and chylomicron synthesis. However it appears that the increased lipid
mass did not alter the clearance kinetics by saturating the triglyceride hydrolysis or
remnant uptake pathways.
In contrast, Berr (Berr, 1992) found that physiological chylomicron catabolism
by liver appears to be saturable by ordinary lipid intake in healthy humans following
intravenous injection of autologous plasma containing retinyl palmitate-labelled
chylomicrons and their remnants. In earlier studies (Berr and Kern Jnr, 1984), (Berr et
al, 1985), the half-time of the major, rapid plasma disappearance in healthy humans
increased with the administered dose, suggesting saturable elimination. The
disappearance of retinyl palmitate after intravenous injection of labelled chylomicrons
in autologous plasma complies with the assumption that the lipoproteins distributed
rapidly within the intravascular space, and the elimination step apparently is rate-
limiting for plasma removal of retinyl palmitate (Berr and Kern Jnr, 1984), (Berr et
al, 1985), (Berretal, 1986).
The organ uptake data provide further evidence that the incorporation of
retinyl esters into chylomicron-like emulsions did not alter uptake processes.
Additionally, the increase in lipid mass did not alter hepatic uptake of cholesteryl
oleate. Splanchnic uptake was always low (<1%), and this was reduced following
injection of an increased emulsion lipid mass. It was not possible to detect retinyl
palmitate in organ extracts, due to the fact that the organs were simultaneously
extracted for the measurement of radiolabeled lipids. Retinyl esters were

309
undetectable by H P L C analysis, possibly due to the reduced mass of retinyl ester
available for extraction and oxidation of retinyl esters.
Retinol and its derivatives are hydrophobic compounds that are unstable in the
presence of oxygen and yield a mixture of dehydrated and double-bonded
rearrangement products in acids. Light catalyses double-bond isomerization of most
retinoids. With light of higher intensity, other photochemical reactions take place,
leading to dimerization and the formation of kitol and other polymers. These
properties require that retinoids be handled experimentally in an inert atmosphere,
avoiding contact with acids and under dim illumination (Blomhoff, 1994), to insure
sample stability and prevent the formation of artifacts (van Breeman et al, 1998).
Commercially synthesised retinyl palmitate contains the antioxidant B H T . This has
previously been added to preparations of tritiated retinyl acetate during the extraction
process to prevent oxidation (Thompson et al, 1983), retinol and other retinyl esters
(Zilversmit et al, 1982). During the present studies, the stability of retinyl stearate,
oleate and myristate was problematic, and the retinyl ester samples were continually
subject to disintegration shortly after synthesis, regardless of handling and storage
procedures. B H T was not added to retinyl ester preparations in these studies, as it was
not clear what effect, if any, it would have on clearance kinetics.
Separation and analysis of individual long-chain fatty acid esters of retinol are
difficult due to similarities in polarity among the major retinyl esters, especially
retinyl palmitate and retinyl oleate, and to susceptibility of all vitamin A molecules to
oxidation, especially during T L C procedures (Ross, 1981b). Reversed-phase H P L C is
the preferred separation and purification technique for retinoids (Furr et al, 1994),
(Wyss, 1995) because the mild conditions of H P L C are compatible with the heat, light
and oxygen sensitive properties of retinoids.
During the current studies, an individualised and appropriate method for the
analysis of retinyl esters by H P L C was developed, enabling the quantitation of retinyl
esters in plasma following intravenous injection of emulsions, and the analysis of
suitable retinyl esters for markers of particle uptake. The sensitivity of the H P L C
apparatus facilitated the detection of retinyl esters at concentrations where emulsion
plasma clearance was not compromised. It appears that the hydrophobic nature of
retinyl esters enables them to distribute within the core of the emulsion particles as
evidenced by the analysis of emulsion fractions, allowing them to be traced up to 30
min in rabbit plasma.

310
In this study, H P L C separations were carried out using a reversed-phase
column instead of G C separation, and retinyl palmitate was preserved for analysis, as
no sample hydrolysis or derivatisation was necessary. The disadvantages of H P L C
analysis can be eliminated by employing mass spectrometry (MS) which provides
both molecular weight information and characteristic fragment ion information
helpful for structural elucidation (Elliot and Waller, 1972), (van Berkel et al, 1996),
(Clifford et al, 1990). Recently, van Breeman et al. (van Breeman et al, 1998)
describe a method for the qualitative analysis of vitamin A compounds using H P L C -
atmospheric pressure chemical ionization-mass spectrometry (APCI-LC-MS). This
analysis utilises a simple hexane extraction of serum followed by on-line C3o-reversed
phase H P L C separation with A P C I mass spectrometric detection. The limit of
detection of APCI-LC-MS for all-trans retinol and all-trans retinyl palmitate was
approx. 34 fmol/ul and 36 fmol/ul, respectively. Subject to availability of APCI-LC-
M S apparatus, this method may provide an alternative for detection of emulsion
retinyl ester in vivo, in particular for later time samples.
In conclusion, the present studies suggest that retinyl myristate and retinyl
palmitate are reliable markers for normal and modified emulsion metabolism,
respectively. The criteria for establishing individual retinyl esters as suitable tracees
of remnant particle clearance were stability of the retinyl ester, definitive separate of
retinyl esters by H P L C , and plasma clearance paralleling that of emulsion cholesteryl
oleate. Receptor-mediated clearance of emulsion remnants was calculated as the
difference between the clearance of normal and modified emulsion cholesteryl oleate
(and retinyl ester).
A future priority is the refinement of the two-emulsion technique, to overcome
difficulties encountered with establishing protocol for lipid dosage. Further studies are
required to determine the lipid mass for injection into animal models, and to avoid
possible saturation of clearance kinetics. A wide range of lipid doses were explored,
however there appeared to be a tentative balance between sufficient retinyl ester in
plasma to enable detection, and the mass of lipid injected to saturate plasma clearance
kinetics. The increased substitution of cholesteryl oleate with retinyl ester as a percent
of total emulsion lipid content may also need to be explored.

311
Chapter 8: General Discussion
8.1 Introduction
In Western societies the increased risk of atherosclerosis may be closely related to the
presence in plasma of the remnant atherogenic lipoproteins, particularly since most
individuals are in a post-prandial state during most of their life span (van Vlijmen et
al, 1996). Clinical studies have shown a positive correlation between prolonged
postprandial lipemia and cardiovascular disease (Patsch et al, 1992), (Karpe and
Hamsten, 1995). The prolonged postprandial state is brought about by a delay in the
plasma removal of chylomicron remnants and current literature strongly supports the
notion that an individual is predisposed to C H D if their capacity to metabolise of
dietary lipids is impaired (Mamo, 1995). Impairment could relate to chylomicron
catabolism or removal of remnants. The extent of postprandial lipemia determines the
effective risk of developing C H D . Factors contributing to prolonged postprandial
lipemia and therefore an individual's capacity to metabolise dietary lipids are age,
genetics, and dietary habits. If the normal physiological uptake of remnants by the
liver is defective then remnants will persist in the circulation, be taken up to a greater
extent by extra-hepatic mechanisms of endocytosis, and thereby contribute to
pathology such as atheroma (Redgrave et al, 1992-a). Therefore, an improvement in
metabolic capacity of postprandial lipoproteins may reduce the risk associated with
coronary atherosclerosis.
Under normal circumstances then, the major route of clearance for
chylomicron remnants and L D L is via the apoB-100/E receptor (Bowler et al, 1991),
(Ishibashi et al, 1994b), (Mahley et al, 1994), (Carrella and Cooper, 1979), (Wade et
al, 1986), (Windier et al, 1988). Recent studies have also described the L R P (Herz et

312
al, 1988), (Herz et al, 1990), a protein that can bind lipoproteins enriched in apo E
(Kowal et al, 1989), (Kowal et al, 1990). The L R P is not subject to down regulation
and provides a potential alternate uptake pathway for chylomicrons (Brown et al,
1991), (Kutt et al, 1989), (De Villiers et al, 1994), (Krieger and Herz, 1994),
(Willnow et al, 1994). This is particularly apparent when the LDL-receptor is down
regulated or absent (Cooper, 1997), (Mortimer et al, 1995a), (Ishibashi etal, 1994b),
(Mahley et al, 1994), (Havel, 1996). Separate receptors and mechanisms for the
binding of remnants have been postulated but their physiological role in clearance of
chylomicrons remains unproven.
Delay or absence of remnant uptake could lead to accumulation of cholesterol-
rich particles in the plasma compartment, deposition of cholesterol into the artery
wall, increased arterial insult and the enhancement of plaque development.
Chylomicron remnant particles have been shown to preferentially accumulate in early
fatty lesions in animal studies, as a consequence of delayed clearance (Proctor and
M a m o , 1996). Elevated concentrations of plasma L D L and chylomicron remnants
occur because of decreased expression of the LDL-receptor. If receptor expression
were chronically suppressed for prolonged periods while chylomicron remnants
continued to enter the arterial wall, excessive accumulation of chylomicron remnants
would occur.
Consequently the primary aim of this project was to develop a diagnostic
assay, whereby receptor-mediated uptake of chylomicron remnants could be assessed
quickly and simply. The proposal is novel in that it may be used to monitor
lipoprotein particle clearance and provide a facile method for the quantitation of net
receptor expression in vivo. In addition, the procedure would be valuable in relating
high affinity uptake with chylomicron remnant clearance and L D L cholesterol
concentration. The procedure could improve screening procedures for individuals at
risk of developing vascular disease as a consequence of decreased receptor clearance
of proatherogenic postprandial lipoproteins from plasma, and possibly establishing
additional criteria for selecting individuals for improved dietary and/or drug
intervention.

313
8.2 The Use of Chylomicron-Like Emulsions to Monitor
Chylomicron Remnant Metabolism In Vivo
The project involved the development of two chylomicron-like emulsions which when
injected simultaneously gave a measure of receptor activity, based on their difference
in clearance. Chylomicron-like emulsion particles are similar in size and composition
to nascent chylomicrons and their plasma clearance is indistinguishable from native
chylomicrons because they rapidly acquire the apolipoproteins necessary for
metabolism (Mortimer et al, 1990), (Maranhao et al, 1986), (Redgrave and
Maranhao, 1985), (Grundy and Mok, 1976), (Cohen, 1989), (Hallberg, 1965). The
clearance of normal emulsions is indicative of high-affinity plus non-specific uptake
mechanisms. In contrast, modified emulsions are cleared via non-specific uptake
pathways only, as their clearance is blocked (confirmed in Chapter 5). Both emulsion
types interact efficiently with hydrolytic enzymes in vivo and are thus converted to a
remnant at the same rate (Chapter 3).
To confirm the use of the two-emulsion technique as a method of quantifying
high affinity uptake in vivo, the clearance of normal and modified emulsions was
studied in homozygous W H H L rabbits, as they provide a unique model for testing the
contributions by receptors for chylomicron remnant clearance in vivo. As predicted,
there was no difference in plasma clearance between the clearances of normal or
modified emulsions in W H H L rabbits, compared with control rabbits. The data from
Chapter 3 demonstrated that the modified emulsion did not interact with receptor
mechanisms in control rabbits and confirmed that the LDL-receptor is the primary
route of uptake of chylomicron remnants, and indicated that the procedure is able to
identify defective plasma clearance of lipoproteins via the LDL-receptor in vivo.
High affinity uptake may be determined by separate or simultaneous injection
of the two emulsion types, however the simultaneous injection approach offers the
advantage of controlling for day-to-day variability in postprandial lipoprotein
metabolism and provides an indication of receptor activity at any one time. There was
no deleterious effect of simultaneous injection on emulsion lipid kinetics compared
with separate injection of the two emulsion types. The comparison of the two modes
of injection in rat and rabbit clearance studies indicated that the lipolysis and particle
clearance was similar for emulsions when injected separately or simultaneously,

314
indicating that no significant saturation of the lipoprotein uptake pathway(s) had
occurred (Chapter 3). High affinity uptake was similar under both conditions.
If the two-emulsion technique was to be ultimately used as a diagnostic tool, it
was necessary to unequivocally establish that the modified emulsion did not interact
with high-affinity (receptor) mechanisms. The intracellular pathway of emulsion
particles labelled with fluorescent cholesteryl ester probes was followed in control,
LDL-receptor deficient mice (Chapter 5). The normal emulsion particles were evenly
distributed in the hepatocytes in control mice and were mainly located in the
sinusoidal spaces in LDL-receptor-deficient mice, indicating the primary role of the
LDL-receptor in chylomicron remnant uptake. Particles from modified emulsions
were not found in hepatocytes in either control or LDL-receptor-deficient mice, and it
was concluded that this emulsion did not interact with receptor mechanisms. In
addition, there was no particle uptake of normal or modified emulsions into the livers
of apo E-deficient mice, thereby confirming the necessity of apo E as a ligand for
receptor uptake. The fluorescent studies in mice deficient in apo E or the L D L
receptor suggest that apo E and the L D L receptor are essential for the normal, rapid
clearance of chylomicron remnants by the liver.
Despite the advantages of using chylomicron-like emulsions, measurement
and quantitation of remnant metabolism has been impeded because of unsatisfactory
procedures. One contributing factor is that the metabolism of chylomicrons is a two
step process, involving lipolysis of chylomicron triglycerides and the hepatic uptake
of remnant particles. Accumulation of chylomicrons in plasma is ascribed to defects
in the delipidation process, in the uptake of remnant particles, or to variable degrees
of combinations of both disturbances (Nakandakare et al, 1994). Insufficiently
delipidated chylomicrons may be poorly recognised by hepatic receptors for these
particles and the consequent delay in plasma clearance may be interpreted as a defect
in uptake mechanisms. However, if chylomicron remnant particles are to be
ultimately used in human subjects, then the established method of converting
chylomicron-like emulsions to remnant particles in vitro is unsatisfactory. The use of
remnant-like emulsions offers the advantage of bypassing the lipolytic pathway and
monitoring particle uptake as a single process, thus reducing confounding variables.
In Chapter 4, chylomicron-like remnants were extensively characterised, in an attempt
to synthesise a remnant-like emulsion based on the data.

315
The hepatic uptake of fluorescently labelled remnant-like emulsions was
compared following separate and simultaneous injection of normal and modified
emulsions in control mice. The particle uptake of both emulsions supported our
previous findings, that simultaneous injection of the two-emulsion types did not alter
the hepatic uptake characteristics, and suggested that the emulsions were taken up via
separate mechanisms. The assay was therefore considered an appropriate measure of
high-affinity uptake mechanisms.
8.3 Development of an Alternate Labelling Technique for Chylomicron-Like Emulsions
Previous experimental data has confirmed that information about remnant metabolism
in man can be obtained by analysis of plasma clearance data following the injection of
radiolabeled chylomicron-like emulsions (Bernardes-Silva et al, 1995), (Redgrave
and Callow, 1990), (Bowler et al, 1991). Emulsions labelled with radioisotopes or
stable isotopes have been utilised to monitor chylomicron metabolism in human
subjects (Redgrave et al, 1993), (Maranhao et al, 1996), (Martins et al, 1995), rats
(Redgrave and Maranhao, 1985), (Redgrave et al, 1991), (Martins et al, 2000b) and
rabbits (Bowler et al, 1991), (Redgrave et al, 1995). However, utilisation of this
procedure as a diagnostic assay is limited by the necessity to use potentially
hazardous radioisotopes that pose radiation problems and are thus less acceptable to
patients. Therefore it was necessary to develop non-isotopic tracers for emulsion
particles that were suitable for use in humans.
Retinyl palmitate has previously been utilised as a marker for chylomicron
remnants in the vitamin A fat load test (Borel et al, 1998), (Bitzen et al, 1994),
(Wilson et al, 1983), (Rassin et al, 1992), (Krasinski et al, 1990b), (Berr et al,
1983), (Berr et al, 1985). However this is a time-consuming procedure that includes
the processes of intestinal absorption and chylomicron synthesis. Radiolabeled retinyl
esters have also been used to monitor the clearance of chylomicron remnant particles
in vivo (van Bennekum et al, 1999a), (Zilversmit, 1979), but are not suitable for use
in humans. Retinyl esters were chosen as an alternative tracee to radiolabels, as they
are hydrophobic and distribute and remain within the core of the chylomicron remnant
during lipolysis and hepatic uptake (see Chapters 1 and 6 for details).

316
Preliminary experiments in established animal models assessed whether
radioisotopes could be replaced by retinyl esters without compromising the validity of
the procedure (Chapter 6). It was predicted that substitution of 25-50% of emulsion
cholesteryl ester mass would be sufficient for detection of retinyl esters in plasma by
H P L C , while maintaining the relative proportion of core to surface components. L o w
and high doses of retinyl esters were incorporated into chylomicron-like emulsions
without altering plasma clearance kinetics of triglyceride clearance or particle uptake
in rats. Measuring the clearance of emulsions containing low doses of retinyl esters in
rabbits also assessed the effect of C E T P on emulsion kinetics. None of the retinyl
esters were found to alter the kinetics of emulsions in rabbits. In addition, assessment
of radiolabeled cholesteryl oleate and triglyceride accumulation in liver suggested
that the retinyl esters did not alter organ uptake in either animal model.
Finally, to assess the use of retinyl esters as appropriate tracers, normal and
modified emulsions were labelled with cholesteryl oleate and retinyl ester.
Chylomicron-like emulsions were injected into recipient animals and their clearance
was determined from the decline in plasma cholesteryl ester radioactivities and retinyl
ester concentrations (Chapter 7). After considerable refinement, retinyl myristate and
retinyl palmitate were found to parallel the clearance of radiolabeled cholesteryl
oleate for normal and modified emulsions, respectively. High affinity uptake was
similar for both tracees.
There was initial concern over the possible transfer of the retinyl esters from
chylomicrons to other plasma lipoproteins. However, retinyl palmitate and retinyl
myristate traced radiolabeled cholesteryl oleate in plasma, suggesting that the retinyl
esters are suitable as appropriate tracees and do not transfer in vivo. Similarly, there
was no evidence of retinyl ester transfer following analysis of emulsion fractions.
Other researches have established that retinyl esters do not transfer to other
lipoproteins in human plasma (Martins et al, 1991), (Sprecher et al, 1991), (Hazzard
and Bierman, 1976). This is attributed to their tight association with chylomicrons and
their remnants (Berr and Kern Jnr, 1984) and lack of release by the liver (Thompson
et al, 1983), and the fact that they are not a substrate for C E T P . Additionally, the
time frame of the clearance studies was short (-30 min), with rapid clearance of
remnant particles. Therefore it is unlikely that a significant mass of retinyl ester will
be exchanged or transferred to plasma H D L or L D L pools, which have much slower
kinetics than chylomicron remnants.

317
During the process of refining the detection of retinyl esters by H P L C , it was
necessary to inject three times the usual lipid mass to facilitate detection of retinyl
esters by H P L C . The small concentrations of retinyl esters in the circulation during
the post-absorptive state have previously been reported as difficult to detect (De
Ruyter and D e Leenheer, 1978). The increase in lipid mass injected did not alter the
clearance kinetics of emulsion triglyceride or cholesteryl oleate (Chapter 7). A n
important issue is the effect of pool size on emulsion remnant clearance. It could be
argued that injecting an enlarged remnant triglyceride mass would cause dilution and
competition for the requirements of emulsion particle removal (e.g. apolipoproteins,
receptors), and potentially delay remnant clearance and increase the time for transfer
of retinyl esters. The dose rates used in rodents in these studies ( 3 0 % total triglyceride
pool) were similar to dose rates used in rabbits (up to 3 0 % total triglyceride pool) and
the dose predicted for human subjects (up to 1 5 % total triglyceride pool). Triglyceride
concentration and mass is traditionally utilised as the basis for calculating lipid
injections, to avoid competition for uptake pathways and saturation of the endogenous
triglyceride pools. However, it may not be appropriate to use the mass of injected
triglyceride as a reference when predicting the mass of retinyl ester required for
detection in plasma.
Collectively, this study represents the foundations of a diagnostic assay for
quantifying receptor activity in vivo, and has contributed significantly to establishing
retinyl esters as alternate markers for monitoring chylomicron remnant metabolism.
The data show that retinyl myristate and retinyl palmitate were suitable markers for
normal and modified emulsion remnant particles, respectively, as their clearance from
plasma paralleled that of radiolabeled cholesteryl oleate in vivo. The procedures
involved have been examined and refined, with respect to injection protocol and
retinyl ester detection.
8.4 Conclusion
It was concluded from clearance studies and fluorescent uptake studies that the L D L -
receptor is the primary receptor responsible for the hepatic uptake of chylomicron
remnants, and that apo E was the necessary ligand required for receptor-mediated
uptake. The application of the two-emulsion method in animal models was successful
in quantifying high affinity uptake, and the data also confirms the use of simultaneous

318
injection of the two emulsion types as an appropriate method for quantifying high
affinity uptake and will provide a dynamic indication of the effect of receptor
expression on lipoprotein metabolism in vivo.
8.5 Future Directions
It is anticipated that this study will provide a platform for continued refinement and
development of the two-emulsion technique for ultimate use in humans. Clinical
studies using F H subjects are required to establish the specificity and reliability of the
method in identifying receptor deficiency. This technique could also be used to
compare emulsion clearance in individuals diagnosed with hyperlipidemia (but not
hypertriglyceridemia), and in normolipidemic individuals. Defects in lipolysis can be
eliminated in these individuals, and the assay may provide specific information about
remnant metabolism.
The decomposition of retinyl esters proved problematic, despite precautions
with preparation and handling. These problems would need to be resolved if future
studies using the diagnostic assay were to be conducted in human subjects. To prevent
oxidation of the retinyl ester compounds, B H T may need to be incorporated during
the synthesis of emulsions and/or extraction of retinyl esters, as disintegration
occurred rapidly and considerable cost was incurred. B H T is used commercially as a
food additive and anti-oxidant, and has been demonstrated to have no link with
medical symptoms (Leclercq et al, 2000), (Reus et al, 2000). However, the
suitability of B H T for use in human subjects would need to be investigated prior to
injection of emulsions. Alternately, P-carotene or ubiquinone are naturally occurring
compounds and have been shown to prevent oxidation of lipids and may therefore
provide a viable alternative. Vitamin E is also a potential antioxidant to protect the
fatty acids in retinyl esters that should be considered. In addition, suitable storage and
handling conditions for retinyl esters requires further attention. The two-emulsion
technique was developed and studied using a dedicated H P L C system to analyse
retinyl ester in plasma, however more specific information may require the use of a
gas chromatography mass spectrometer to enable detection of minor defects in
receptor function.
The use of remnant-like emulsions was explored, however chylomicron-like
emulsions were ultimately utilised for the purposes of these experiments. Remnant-

319
like emulsions enable the assessment of remnant clearance independent of remnant
formation, and may offer stability advantages over chylomicron-like particles because
of their smaller size, greater density and lower content of triglyceride relative to
phospholipid. The 13C breath test utilises remnant-like emulsions and the stability and
procedures for the preparation of remnant-like emulsions has been rigorously assessed
for clinical use (Redgrave et al, 1995). Substitution of a larger percent of the
cholesteryl oleate component with retinyl esters may be less detrimental to clearance
kinetics due to the high proportion of cholesteryl oleate. Preliminary clearance studies
are required to determine if the clearance kinetics of remnant-like emulsions are
altered by the presence of retinyl esters.
Current techniques for quantifying LDL-receptor m R N A provide a means of
identifying individuals with LDL-receptor mutations or deficiencies, and to assess the
effect of therapeutic interventions on receptor activity. The expression of the L D L -
receptor is primarily regulated at the transcriptional level, therefore to define m R N A
translation, gene transcription, and LDL-receptor binding activity, the level of L D L -
receptor protein and m R N A can be measured in cells. The technique has been used to
measure the effects of dietary cholesterol on the expression of hepatic LDL-receptors
in rats (Roach et al, 1993a) and rabbits (Roach et al, 1993c), and to examine the
effect of simvastatin on the regulatory elements of cholesterol metabolism in
circulating mononuclear cells (Smith et al, 2000). Colloidal gold-LDL conjugates can
also be used to quantify tissue and cell LDL-receptor activity and mass (Roach et al,
1987). The in vivo expression and regulation of the LDL-receptor of circulating
mononuclear cells has been studied using a sensitive spectrophotometric assay with
colloidal gold-LDL conjugates in F H subjects (Roach et al, 1993b). Detection with
the colloidal gold-LDL conjugates has been shown to be as sensitive as
autoradiographic method with 125I-labeled L D L , and the biotinylated L D L method
could be used to quantify tissue and cell LDL-receptors down to attomolar levels.
A method described by Pal et al. (Pal et al, 2000) involves the binding of
colloidal gold-labelled chylomicron remnants to detect LDL-receptor binding activity
in fibroblasts, and suggests that changes in LDL-receptor expression might be more
readily identified using gold-labelled remnants. Recently, ICP-MS has been shown to
provide a more sensitive method for the detection and quantitation of LDL-receptor
binding and uptake of colloidal gold conjugates (Roach et al, 1993b), (Martin de
Llano et al, 1996), (Robenek et al, 1991), (Gierens et al, 2000). Further studies are

320
required to assess if LDL-receptor expression can be correlated with chylomicron
uptake using mononuclear cells.
Another method for monitoring chylomicron metabolism is the measurement
of the unique structural protein of intestinally derived lipoproteins, apo B-48 (Dane-
Stewart et al, 2001), (Watts et al, 2001), (Kane et al, 1980), (Phillips et al, 1997).
Apo B-48 is found exclusively with chylomicrons and their remnants, and the fasting
plasma concentration may provide an indication of the concentration of particles and
an indication of clearance and production. Fasting plasma concentration of apo B-48
appeared to be a good surrogate marker for the degree of postprandial lipidemia and
may circumvent the need for oral fat challenges (Smith et al, 1999).
Alternately, the stable isotope breath test provides a functional assessment of
chylomicron remnant metabolism. The rate of appearance of label in the breath can be
used to define the roles of certain disease states, e.g., LDL-receptor deficiency, on the
roles of lipid constituents on remnant clearance (Martins et al, 2000b). The breath
test provides an integrated and quantitative information assessment of capacity for
clearance and subsequent metabolism of the remnants of triglyceride-rich lipoproteins
in intact experimental animals and more recently in individuals with F H and familial
dyslipidemia (Watts et al, 2001), (Dane-Stewart et al, 2001), (Redgrave et al, 2001).
The breath test m a y prove to be easier to administer and interpret than presently
available chromatographic methods of measuring clearance of postprandial lipids,
particularly as stable isotope measurements become more routine (Redgrave et al,
2001). In particular, the metabolism of normal and modified emulsions can be
assessed using the breath test and a measure of high affinity uptake of remnant
particles could be obtained. The future usefulness of the breath test in clinical
investigation will depend on whether it identifies individuals at increased risk of
cardiovascular diseases in the absence of other conventional risk factors and whether
it allows the assessment of therapeutic interventions that decrease cardiovascular risk.

321
References
Adeli, K. (1994). Regulated intracellular degradation of apolipoprotein B in semipermeable HepG2 cells. The Journal of Biological Chemistry, 269 (12): 9166-9175.
Agellon, L. B., Walsh, A., Hayek, T., Moulin, P., Jiang, X. C, Shelanski, S. A., Breslow, J. L. and Tall, A. R. (1991). Reduced high density lipoprotein cholesterol in human cholesteryl ester transfer protein transgenic mice. The Journal of Biological Chemistry, 266 (17): 10796-801.
Alexander, C. A., Hamilton, R. L. and Havel, R. J. (1976). Subcellular localisation of B apoprotein of plasma lipoproteins in rat liver. Journal of Cell Biology, 69: 241-263.
Amar, M. J. A., Dugi, K. A., Haudenschild, C. C, Shamburek, R. D., Foger, B., Chase, M., Bensadoun, A., Hoyt, R. F., Brewer, H. B. and Santamarina-Fojo, S. (1998). Hepatic lipase facilitates the selective uptake of cholesteryl esters from remnant lipoproteins in apoE-deficient mice. Journal of Lipid Research, 39 (12): 2436-2442.
Andersen, J. M., Nervi, F. O. and Diestschy, J. M. (1977). Rate constants for the uptake of cholesterol from various intestinal and serum lipoprotein fractions by the liver of the rat in vivo. Biochimica et Biophysica Acta, 486: 298.
Andres, J., DeFalcis, D. and Massague, J. (1992). Binding of two growth factor families to separate domains of the proteoglycan betaglycan. The Journal of Biological Chemistry, 267: 5927-5930.
Angelin, B., Raviola, C. A., Innerarity, T. L. and Mahley, R. W. (1983). Regulation of hepatic lipoprotein receptors in the dog. Rapid regulation of apolipoprotein B,E receptors, but not of apolipoprotein E receptors by intestinal lipoproteins and bile acids. Journal of Clinical Investigation,
71: 816-831.
Anitschow, N. and Chalatow, S. (1913). Ueber experimentelle cholesterinseatose und ihre bedeutung fur die entstehung einiger pathologisher prozesse. Zentralbl. Allg. Pathol. Pathol. Anat, 24: 1-9.
Arbeeny, C. M. and Rifici, V. A. (1984). The uptake of chylomicron remnants and very low density lipoprotein remnants by the perfused rat liver. The Journal of Biological Chemistry, 259: 9662-9666.
Arem, R. and Patsch, W. (1990). Lipoprotein and apolipoprotein levels in subclinical hypothyroidism. Effect of levothyroxine therapy. Archives of Internal Medicine, 150 (10): 2097-2100.
Arnon, R., Sehayek, E., Vogel, T. and Eisenberg, S. (1991). Effects of exogenous apo-E-3 and of cholesterol-enriched meals on the cellular metabolism of human chylomicrons and their remnants. Biochimica et Biophysica Acta, 1085: 336-342.

322
Assman, G., von Eckardstein, A. and Brewer, H. B. (1995). In Familial High Density Lipoprotein Deficiency: Tangier Disease. The Metabolic and Molecular Bases of Inherited Disease., Vol. 2. (Eds, Scriver, C. R., Beaudet, A. L., Sly, W . S. and Valle, D.) McGraw-Hill Inc., N e w York, pp. 2053-2072.
Assmann, G. (1982). Lipid Metabolism and Atherosclerosis. Schattauer-Verlag, Germany.
Attie, A. D., Pittman, R. C, Watanabe, Y. and Steinberg, D. (1981). Low Density Lipoprotein Receptor Deficiency in Cultured Hepatocytes of the W H H L Rabbit. Further Evidence of Two Pathways for Catabolism of Exogenous Proteins. The Journal of Biological Chemistry, 256: 9789-9792.
Backer, J. M. and Dawidowicz, E. A. (1981). Mechanism of exchange of between phospholipid vesicles. Biochemistry, 20: 3805-3810.
Banuelos, S., Arrondo, J. L., Goni, F. M. and Pifat, G. (1995). Surface-core relationships in human low density lipoprotein as studied by infrared spectroscopy. The Journal of Biological Chemistry, 270 (16): 9192-9196.
Bar-on, H., Levy, E., Oschry, Y., Ziv, E. and Shafrir, E. (1984). Removal defect of very-low-density lipoproteins from diabetic rats. Biochimica et Biophysica Acta., 793 (1.): 115-118.
Barrett, P. H. R. (1998). Kinetics of triglyceride rich lipoproteins: chylomicrons and very low density lipoproteins. Atherosclerosis., 141 (Suppl. 1): S35-S40.
Barritt, D. W. (1956.). Alimentary lipaemia in men with coronary artery disease and controls. British Medical Journal., ii: 640-644.
Barter, P. and Rye, K.-A. (1994). Cholesteryl ester transfer protein: Its role in plasma lipid transport. Clinical and Experimental Pharmacology and Physiology., 21: 663-672.
Barter, P. J., Chang, L. B. F., Newnham, H. H., Rye, K.-A. and Rajaram, O. V. (1990). The interaction of cholesteryl ester transfer protein and unesterified fatty acids promotes a reduction in the particle size of high density lipoproteins. Biochimica et Biophysica Acta, 1045: 81-89.
Barter, P. J. and Lally, J. I. (1979). In vitro exchanges of esterified cholesterol between serum lipoprotein fractions: Studies of humans and rabbits. Metabolism., 28: 230-237.
Bartlett, G. R. (1959). Phosphorus assay In column chromatography. The Journal of Biological Chemistry., 234 (3.): 466-468.
Basu, S. K., Brown, M. S., Ho, Y. K., Havel, R. J. and Goldstein, J. L. (1981a). Mouse macrophages synthesize and secrete a protein resembling apolipoprotein E. Proc. Natl. Acad. Sci. U.S.A., 78 (12.):
7545-7549.
Basu, S. K, Goldstein, J. L., Anderson, R. G. and Brown, M. S. (1981b). Monensin interrupts the recycling of low density lipoprotein receptors in human fibroblasts. Cell, 24 (2): 493-502.
Beaumont, J.-L. and Assadollahi, F. (1990). Retinyl palmitate labeled intestinally derived lipoproteins accumulate in the circulation of W H H L rabbits. Atherosclerosis., 85: 103-111.
Beisegel, U., Weber, W. and Bengstsson-Olivecrona, G. (1991). Lipoprotein lipase enhanced the binding of chylomicrons to low density receptor-related protein. Proc. Natl Acad. Sci. U.S.A., 88.:
8342-8346.
Beisiegel, U., Weber, W., Ihrke, G., Herz, J. and Stanley, K. K. (1989). The LDL-receptor-related protein, LRP, is an apolipoprotein E-binding protein. Nature., 341: 162-163.
Benlian, P., De Gennes, J. L., Foubert, L., Zhang, H., Gagne, S. E. and Hayden, M. (1996). Premature atherosclerosis in patients with familial chylomicronemia caused by mutations in the lipoprotein lipase
gene. The New England Journal of Medicine., 335: 848-854.

323
Bennett Clark, S. (1979). Mucosal coenzyme A-dependent cholesterol esterification after intestinal perfusion of lipids in rats. The Journal of Biological Chemistry., 254: 1534-1536.
Bergeron, N. and Havel, R. J. (1995). Influence of Diets Rich in Saturated and Omega-6 Polyunsaturated Fatty Acids on the Postprandial Response of Apolipoproteins B-48, B-100, E, and Lipids in Triglyceride-Rich Lipoproteins. Arteriosclerosis Thrombosis and Vascular Biology, 15: 2111-2121.
Bergeron, N. and Havel, R. J. (1996). Prolonged postprandial response of lipids and apolipoproteins in triglyceride-rich lipoproteins of individuals expressing an apolipoprotein E4 allele. Journal of Clinical Investigation., 97: 65-72.
Bergeron, N. and Havel, R. J. (1997). Assessment of postprandial lipemia: nutritional influences. Current Opinion in Lipidology., 8: 43-52.
Bergeron, N., Kotite, L. and Havel, R. J. (1996). Simultaneous quantification of apolipoproteins B-100, B-48 and E separated by SDS-PAGE. Methods in Enzymology., 263: 82-94.
Bergman, E. N., Havel, R. J., Wolfe, B. M. and Bohmer, T. (1971). Quantitative studies of the metabolism of chylomicron triglycerides and cholesterol by liver and extrahepatic tissues of sheep and dogs. Journal of Clinical Investigation., 50: 1831-1839.
Bernardes-Silva, H., Toffoletto, O., Bortolotto, L. A., Latrilha, M. C. M., Krieger, E. M., Pileggi, F. and Maranhao, R. C. (1995). Malignant Hypertension Is Accompanied by Marked Alterations in Chylomicron Metabolism. Hypertension., 26 Q?art 2): 1207-1210.
Berr, F. (1992). Characterization of chylomicron remnant clearance by retinyl palmitate labed in normal humans. Journal of Lipid Research., 33: 915-930.
Berr, F., Eckel, R. and Kern Jnr, F. (1985). Plasma decay of chylomicron remnants is not affected by heparin-stimulated plasma lipolytic activity in normal fasting man. Journal of Lipid Research., 26: 852-859.
Berr, F., Eckel, R. H. and Kern Jnr, F. (1986). Contraceptive steroids increase hepatic uptake of chylomicron remnants in healthy young women. Journal of Lipid Research., 27: 645-651.
Berr, F. and Kern Jnr, F. (1984). Plasma clearance of chylomicrons labeled with retinyl palmitate in healthy human subjects. Journal of Lipid Research., 25: 805-812.
Berr, F., McKinley, C. and Kern Jnr, F. (1983). Plasma Clearance of Chylomicrons Labelled with Retinol Palmitate. Clinical Research., 31 (1): 29.
Bezman-Tarcher, A., Otway, S. and Robinson, D. S. (1965). The removal of triglyceride fatty acids from the circulation of the supradiaphragmatic portion of the rat. Proc. R. Soc. Lond. B. Biol. Sci., 162:
411-426.
Bhat, P. V. and Lacroix, A. (1983). Separation and estimation of retinyl fatty acyl esters in tissues of normal rat by high performance liquid chromatography. Journal of Chromatography., 272.: 269-278. Bhattacharya, S. and Redgrave, T. G. (1981). The content of apolipoprotein B in chylomicron particles. Journal of Lipid Research., 22: 820-828.
Bierman, E. L. and Glomset, J. A. (1985). In Disorders of lipid metabolism. Williams Textbook of Endocrinology., Qids, Wilson, J. D. and Foster, D. W.) W.B. Saunders., Philadelphia., pp. 1108-1136.
Bilheimer, D. (1987). In Familial Hypercholesterolaemia. Bailliere's Clinical Endocrinology and Metabolism. International Practice and Research., Vol. 1. (Ed, Shepherd, J.) Bailliere Tindall.,, pp.
581-601.

324
Bilheimer, D. W., Goldstein, J. L., Grundy, S. M., Starzl, T. E. and Brown, M . S. (1984). Liver transplantation to provide low density lipoprotein receptors and lower plasma cholesterol in a child with homozygous familial hypercholesterolemia. New England Journal of Medicine., 311: 1658-1664.
Bilheimer, D. W., Stone, N. J. and Grundy, S. M. (1979). Metabolic studies in familial hypercholesterolemia. Evidence for a gene-dosage effect in vivo. Journal of Clinical Investigation., 64: 524-533.
Bilheimer, D. W., Watanabe, Y. and Kita, T. (1982). Impaired receptor-mediated catabolism of low density lipoprotein in the W H H L rabbits, an animal model of familial hypercholesterolemia. Proc. Natl. Acad. Sci. U.S.A., 79: 3305-3309.
Bitzen, U., Winqvist, M., Nilsson-Ehle, P. and Fex, G. (1994). Retinyl Palmitate is a reproducible marker for chylomicron elimination from blood. Scand. J. Clin. Lab. Invest., 54: 611-613.
Bjorkegren, J., Packard, C. J., Hamsten, A., Bedford, D., Caslake, M., Foster, L., Shepherd, J., Stewart, P. and Karpe, F. (1996). Accumulation of large very low density lipoprotein in plasma during intravenous infusion of a chylomicron-like triglyceride emulsion reflects competition for a common lipolytic pathway. Journal of Lipid Research., 37: 76-86.
Black, D. D. (1995). Invited Review. Intestinal Metabolism. Journal of Pediatric Gastoenterology and Nutrition., 20: 125-147.
Blaner, S., Obunike, J. C, Kurlandsky, S. B., Al-Haideri, M., Piantedosi, R., Deckelbaum, R. J. and Goldberg, I. J. (1994). Lipoprotein Lipase Hydrolysis of Retinyl Ester. Possible Implications for Retinoid Uptake by Cells. The Journal of Biological Chemistry., 269 (24): 16559-16565.
Blomhoff, R. (1994). Transport and Metabolism of Vitamin A. Nutrition Reviews., 52 (2): S13-S23.
Blomhoff, R., Berg, T. and Norum, K. R. (1988). Transfer of retinol from parenchymal to stellate cells in liver is mediated by retinol-binding protein. Proc. Natl. Acad. Sci. U.S.A., 85: 3455-3458.
Blomhoff, R., Green, M. H., Green, J. B., Berg, T. and Norum, K. R. (1991). Vitamin A metabolism: new perspectives on absorption, transport, and storage. Physiological Reviews., 71 (4): 951-990.
Blomhoff, R., Green, M. H. and Norum, K. R. (1992). Vitamin A: physiological and biochemical processing. Annu. Rev. Nutr., 12: 37-57.
Blomhoff, R., Helgerud, P., Rasmussen, M., Berg, T. and Norum, K. R. (1982). In vivo uptake of chylomicron [3H]retinyl ester by rat liver: Evidence for retinol transfer from parenchymal to nonparenchymal cells. Proc. Natl. Acad. Sci. USA., 19: lUe-HW.
Blomhoff, R., Holte, K., Naess, L. and Berg, T. (1984). Newly Administered [3H]Retinol is Transferred from Hepatocytes to Stellate Cells in Liver for Storage. Experimental Cell Research., 150:
186-193.
Boldingh, J., Cama, H. R., Collins, F. D., Morka, R. A., Gridgeman, N. T., Isler, O., Kofler, M., Taylor, R. J., Welland, A. S. and Bradbury, T. (1951). Pure all-trans Vitamin A Acetate and the assessment of Vitamin A potency by spectrophotometry. Nature., 168 (Oct. 6): 598.
Bollman, J. L. (1948). A cage which limits the activity of rats. /. Lab. Clin. Med., 33.: 1348.
Boquist, S., Ruotolo, G., Tang, R., Bjorkegren, J., Bond, M. G., de Faire, U., Karpe, F. and Hamsten, A. (1999). Alimentary lipemia, postprandial triglyceride-rich lipoproteins, and common carotid intima-media thickness in healthy, middle-aged men. Circulation., 100 (7): 723-728.
Borchardt, R. A. and Davis, R. A. (1987). Intrahepatic assembly of very low density lipoproteins. Rate of transport out of the endoplasmic reticulum determines rate of secretion. The Journal of Biological
Chemistry., 262 (34): 16394-16402.

325
Borel, P., Mekki, N., Boirie, Y., Partier, A., Alexandre-Gouabau, M.-C, Grolier, P., Beaufrere, B., Portugal, H., Lairon, D. and Azais-Braesco, V. (1998). Comparison of the Postprandial Plasma Vitamin A Response in Young and Older Adults. Journal of Gerentology: Biological Sciences., 53A (2): B133-B140.
Borensztajn, J. and Kotlar, T. J. (1984). Liver uptake of chylomicron remnants with high and low apoprotein E:C ratios. Proc. Natl. Acad. Sci. U.S.A., 81 (18.): 5863-5866.
Bostrom, K., Boren, J., Wettesten, M., Sjoberg, A., Bondjers, G., Wiklund, O., Carlsson, P. and Olofsson, S.-O. (1988). Studies on the assembly of apo B-100-containing lipoproteins in HepG2 cells. The Journal of Biological Chemistry., 263 (9): 4434-4442.
Bowler, A., Redgrave, T. G. and Mamo, J. C. L. (1991). Chylomicron-remnant clearance in homozygote and heterozygote Watanabe-heritable-hyperlipidaemic rabbits is defective. Biochemical Journal, 276: 381-386.
Boyd, G. S. and Gorban, A. M. S. (1980). Newly Discovered Systems of Enzyme Regulation by Reversible Phosphorylation., Vol. 1. (Ed, Cohen, P.) Elsevier., North Holland, Amsterdam., pp. 95-134.
Brasaemle, D. L., Cornelymoss, K. and Bensadoun, A. (1993). Hepatic lipase treatment of chylomicron remnants increases exposure of apolipoprotein E. Journal of Lipid Research., 34: 455-465.
Braun, J. E. A. and Severson, D. L. (1992). Regulation of the synthesis, processing and translocation of lipoprotein lipase. Biochemical Journal, 287: 337-347.
Brenninkmeijer, B. J., Stuyt, P. M. J., Demacker, P. N. M., Stalenhoef, A. F. H. and van"t Laar, A. (1987). Catabolism of chylomicron remnants in normolipidemic subjects in relation to the apoprotein E phenotype. Journal of Lipid Research., 28: 361-370.
Breslow, J. L. (1993). Transgenic mouse models of lipoprotein metabolism and atherosclerosis. Proc. Natl Acad. Sci. USA., 90: 8314-8318.
Breslow, J. L. (1996). Mouse models of atherosclerosis. Science., 272: 685-688.
Brown, A. J. and Roberts, D. C. K. (1991). Moderate Fish Oil Intake Improves Lipemic Response to a Standard Fat Meal. A Study in 25 Healthy Men. Arteriosclerosis and Thrombosis., 11: 457-466.
Brown, M. S., Basu, S. K., Falck, J. R., Ho, Y. K. and Goldstein, J. L. (1980). The Scavenger Cell Pathway for Lipoprotein Degradation : Specificity of the Binding Site That Mediates the Uptake of Negatively-Charged L D L by Macrophages. Journal of Supramolecular Structure., 13: 67-81.
Brown, M. S. and Goldstein, J. L. (1975). Regulation of the activity of the low density lipoprotein receptor in human fibroblasts. Cell, 6. (3): 307-316.
Brown, M. S. and Goldstein, J. L. (1979). Receptor-mediated endocytosis: Insights from the lipoprotein receptor system. Proc. Natl. Acad. Sci. USA., 16 (7): 3330-3337.
Brown, M. S. and Goldstein, J. L. (1983). Lipoprotein Receptors in the Liver. Control Signals for Plasma Cholesterol Traffic. Journal of Clinical Investigation., 72: 743-747.
Brown, M. S. and Goldstein, J. L. (1984). How LDL receptors influence cholesterol and
atherosclerosis. Scientific American., 251: 58-67.
Brown, M. S. and Goldstein, J. L. (1986). A Receptor-Mediated Pathway for Cholesterol Homeostasis.
Science., 232: 34-47.
Brown, M. S., Herz, J., Kowal, R. C. and Goldstein, J. L. (1991). The low-density lipoprotein receptor-related protein: double agent or decoy? Current Opinion in Lipidology., 2: 65-72.

326
Brown, M . S., Kovanen, P. T. and Goldstein, J. L. (1981). Regulation of Plasma Cholesterol by Lipoprotein Receptors. Science., 212: 628-635.
Brown, M. S., Kovanen, P. T., Goldstein, J. L, Eeckels, R., Vandenberghe, K., van den Berghe, H„ Fryns, J. P. and Cassiman, J. J. (1978). Prenatal diagnosis of homozygous familial hypercholesterolemia. Lancet., 1 (8063): 526-529.
Brummer, D., Evans, D., Berg, D., Greten, H., Beisiegel, U. and Mann, W.A. (1998). Expression of Type III Hyperlipoproteinemia in patients homozygous for apolipoprotein E-2 is modulated by lipoprotein lipase and postprandial hyperinsulinemia. Journal of Molecular Medicine., 76 (5): 355-364.
Brunzell, J. D., Hazzard, W. R., Porte, D. and Bierman, E. L. (1973). Evidence for a Common, Saturable, Triglyceride Removal Mechanism for Chylomicrons and Very Low Density Lipoproteins in Man. Journal of Clinical Investigation., 52: 1578-1585.
Buhman, K. K., Accad, M., Novak, S., Choi, R. S., Wong, J. S., Hamilton, R. L., Turley, S. and Farese, R. V. (2000). Resistance to diet-induced hypercholesterolemia and gallstone formation in ACAT2-deficient mice. Nature Medicine., 6(12): 1341-1347.
Cabezas, M. C, De Bruin, T. W. A., Westerfeld, H. E., Meijer, E. and Erkelens, D. W. (1998). Delayed chylomicron remnant clearance in subjects with heterozygous familial hypercholesterolaemia. Journal of Internal Medicine., 244 (4): 1-9 or 299-307.
Cabezas, M. C, Erkelens, D. W., Kock, L. A. W. and De Bruin, T. W. A. (1994). Postprandial apolipoprotein B100 and B48 metabolism in familial combined hyperlipidaemia before and after reduction of fasting plasma triglycerides. European Journal of Clinical Investigation., 24: 669-678.
Carlson, L. A. and Hallberg, D. (1963). Studies on the Elimination of Exogenous Lipids from the Blood Stream. The Kinetics of the Elimination of a Fat Emulsion and of Chylomicrons in the Dog after a Single Injection. Acta. Physiol. Scand., 59: 52-61.
Carrella, M. and Cooper, A. D. (1979). High affinity binding of chylomicron remnants to rat liver plasma membranes. Proc. Natl. Acad. Sci. U.S.A., 76: 338-342.
Carroll, K. K. (1971). Plasma cholesterol levels and liver cholesterol biosynthesis in rabbits fed commercial or semisynthetic diets with and without added fats or oils. Atherosclerosis., 13: 67-76.
Castelli, W. A., Garrison, R. J., Wilson, P. W. F. and Abbott, R. P. (1986). Incidence of coronary heart disease and lipoprotein cholesterol level. /. Am. Med. Assoc, 256: 2835-2838.
Cavallero, E., Piolot, A. and Jacotot, B. (1995). Postprandial lipoprotein clearance in type 2 diabetes: fenofibrate effects. Diabete et Metabolisme., 21 (2): 118-120.
Chajek, T. and Eisenberg, S. (1978). Very low density lipoprotein. Metabolism of phospholipids, cholesterol and apolipoprotein C in the isolated perfused rat heart. Journal of Clinical Investigation.,
61: 1654.
Chajek-Shaul, T., Eisenberg, S., Oschry, Y. and Olivecrona, T. (1983). Metabolic heterogeneity of post-lipolysis rat mesenteric lymph small chylomicrons produced in vitro. Journal of Lipid Research.,
24: 831-840.
Chang, B. H., Liao, W., Li, L., Nakamuta, M., Mack, D. and Chan, L. (1999). Liver-specific inactivation of the abetalipoproteinemia gene completely abrogates very low density lipoprotein/low density lipoprotein production in a viable conditional knockout mouse. The Journal of Biological
Chemistry., 21A (10): 6051-6055.
Chang, T. Y., Chang, C. C. Y. and Cheng, D. (1997). Acyl-coenzyme A:cholesterol acyltransferase.
Ann. Rev. Biochem., 66: 613-638.
Chang, T. Y. and Doolitle, G. M. (1983). The Enzymes. Academic Press., New York., 16.

327
Chappell, D. A., Fry, G. L., Waknitz, M. A., Iverius, P.-H, Williams, S. E. and Strickland, D. E. (1992). The low density lipoprotein receptor-related protein/alpha-2-macroglobulin receptor binds and mediates catabolism of bovine milk lipoprotein lipase. The Journal of Biological Chemistry., 267: 25764-25767.
Chatterton, J. E., Phillips, M. L., Curtiss, L. K, Milne, R. W., Marcel, Y. L. and Schumaker, V. N. (1991). Mapping apolipoprotein B on the low density lipoprotein surface by immunoelectron microscopy. The Journal of Biological Chemistry., 266 (9): 5955-5962.
Chen, T., Floren, C. H., Nilsson, A. and Infante, R. (1991). Regulation of chylomicron remnant uptake in the human hepatoma cell-line Hep G2. Role of the low-density lipoprotein receptor. Biochimica et Biophysica Acta., 1083: 173-178.
Chen, Y. D., Swami, S., Skowronski, R., Coulston, A. and Reaven, G. M. (1993). Differences in postprandial lipemia between patients with normal glucose tolerance and noninsulin-dependent diabetes mellitus. Journal of Clinical Endocrinology and Metabolism., 76 (1): 172-177.
Choi, S. Y. and Cooper, A. D. (1993). A comparison of the roles of the low density lipoprotein (LDL) receptor and the L D L receptor-related protein/alpha-2-macroglobulin receptor in chylomicron remnant removal in the mouse in vivo. Journal of Cell Biology., 268: 15804-15811.
Choi, S. Y., Fong, L. G., Kirven, M. J. and Cooper, A. D. (1991). Use of an Anti-Low Density Lipoprotein Receptor Antibody to Quantify the Role of the L D L Receptor in the Removal of Chylomicron Remnants in the Mouse In vivo. Journal of Clinical Investigation., 88 (1173-1181).
Clarkson, T. B. (1963). Atherosclerosis - spontaneous and induced. Adv. Lipid. Res., 1: 211-252.
Clarkson, T. B. (1972). Animal models of atherosclerosis. Adv. Vet. Sci. Comp. Med., 16: 151-173.
Clarkson, T. B., Pritchard, R. W., Bullock, B. C, Lehner, N. D., Lofland, H. B. and St Clair, R. W. (1970). Animal models of atherosclerosis. Animal models for biomedical research., 3: 22-41.
Clifford, A. J., Jones, A. D. and Furr, H. C. (1990). Stable isotope dilution mass spectrometry to assess vitamin A status. Methods in Enzymology., 189: 94-104.
Clifton, P. M., Kestin, M., Abbey, M., Drysdale, M. and Nestel, P. J. (1990). Relationship between sensitivity to dietary fat and dietary cholesterol. Arteriosclerosis., 10 (3.): 394-401. Cohen, J. C. (1989). Chylomicron triglyceride clearance: comparison of three assessment methods. American Journal of Clinical Nutrition., 49: 306-313.
Cohn, J. S. (1994). Postprandial lipid metabolism. Current Opinion in Lipidology., 5: 185-190.
Cohn, J. S., Johnson, E. J., Millar, J. S., Cohn, S. D., Milne, R. W., Marcel, Y. L., Russell, R. M. and Schaefer, E. J. (1993). Contribution of apoB-48 and apoB-100 triglyceride-rich lipoproteins (TRL) to postprandial increases in the plasma concentration of T R L triglycerides and retinyl esters. Journal of Lipid Research., 34: 2033-2040.
Cohn, J. S., McNamara, J. R., Krasinski, S. D., Russell, R. M. and Schaefer, E. J. (1989). Role of triglyceride-rich lipoproteins from the liver and intestine in the etiology of postprandial peaks in plasma triglyceride concentration. Metabolism., 38 (5): 484-490.
Connelly, P. W., Maguire, G. F. and Little, J. A. (1987). Apolipoprotein CII St. Michael. Familial apolipoprotein C U deficiency associated with premature vascular disease. Journal of Clinical
Investigation., 80: 1597-1606.
Connelly, P. W., Vezina, C. and Maguire, G. F. (1996). Quantification of apolipoprotein C-II by immunochemical and chromatographic methods. Methods in Enzymology., 263: 188-208.

328
Connor, W . E. and Conner, S. L. (Eds.) (1986). Dietary cholesterol and fat and the prevention of coronary heart disease. Risks and benefits of nutritional change. Diet and prevention of coronary heart disease and cancer., Raven Press., N e w York., pp. 113-147.
Constantinides, P. (1961). Production of experimental atherosclerosis in animals. /. Atheroscler. Res., 1: 374-385.
Cooper, A. D. (1977). The metabolism of chylomicron remnants by isolated perfused rat liver. Biochimica et Biophysica Acta., 488: 464-474.
Cooper, A. D. (1985). Role of the liver in the degradation of lipoproteins. Gastroenterology., 88: 192-205.
Cooper, A. D. (1997). Hepatic uptake of chylomicron remnants. Journal of Lipid Research., 38: 2173-2192.
Cooper, A. D., Erickson, S. K., Nutik, R. and Shrewsbury, M. A. (1982a). Characterization of chylomicron remnant binding to rat liver membranes. Journal of Lipid Research., 23.: 42-52.
Cooper, A. D., Nutik, R. and Chen, J. (1987). Characterisation of the estrogen-induced lipoprotein receptor of rat liver. Journal of Lipid Research., 28: 59-68.
Cooper, A. D., Shrewsbury, M. A. and Erickson, S. K. (1982b). Comparison of binding and removal of remnants of triglyceride-rich lipoproteins of intestinal and hepatic origin by rat liver in vitro. American Journal of Physiology., 243 (5): G389-G395.
Cooper, A. D. and Yu, P. Y. S. (1978). Rates of removal and degradation of chyomicron remnants by isolated perfused rat liver. Journal of Lipid Research., 19: 635-643.
Cortner, J. A., Coates, P. M., Le, N.-A., Cryer, D. R., Ragni, M. C, Faulkner, A. and Langer, T. (1987). Kinetics of chylomicron remnant clearance in normal and in hyperlipoproteinemic subjects. Journal of Lipid Research., 28: 195-206.
Curtin, A., Deegan, P., Owens, D., Collins, P., Johnson, A. and Tomkin, G. H. (1994). Alterations in apo B-48 in the postprandial state in N J D D M . Diabetologia., 37: 1259-1264.
Cuthbert, J. A., East, C. A., Bilheimer, D. W. and Lipsky, P. E. (1986). Detection of Familial Hypercholesterolemia by Assaying Functional Low-Density-Lipoprotein Receptors on Lymphocytes. The New England Journal of Medicine., 314: 879-883.
Czarnecka, H. and Yokoyama, S. (1995). Lecithinxholesterol acyltransferase reaction on cellular lipid released by free apolipoprotein-mediated efflux. Biochemistry., 34 (13): 4385-4392.
Dallongeville, J. and Fruchart, J.-C. (1998). Postprandial Dyslipidemia: A Risk Factor for Coronary Heart Disease. Annals of Nutrition and Metabolism., 42 (1): 1-11.
Dane-Stewart, C. A., Watts, G. F., Mamo, J. C, Dimmitt, S. B., Barrett, P. H. and Redgrave, T. G. (2001). Elevated apolipoprotein B-48 and remnant-like particle-cholesterol in heterozygous familial hypercholesterolaemia. European Journal of Clinical Investigation., 31 (2): 113-117.
Dashti, N., Williams, D. L. and Alaupovic, P. (1989). Effects of oleate and insulin on the production rates and cellular m R N A concentrations of apolipoproteins in HepG2 cells. Journal of Lipid Research.,
30 (9): 1365-1373.
Davidson, N. O., Kollmer, M. E. and Glickman, R. M. (1986). Apolipoprotein B synthesis in rat small intestine: regulation by dietary triglyceride and biliary lipid. Journal of Lipid Research., 27. (1.): 30-39. Davidson, N. O , Powell, L. M., Wallis, S. C. and Scott, J. (1988). The rat and mouse liver also produces apo B-48. The Journal of Biological Chemistry, 263: 13482-13488.

329
Davignon, J. and Cohn, J. S. (1996). Triglycerides: a risk factor for coronary heart disease.
Atherosclerosis., 124 (Suppl.): S57-S64.
Davis, R. A. (1991). In Lipoprotein structure and secretion. Biochemistry of lipids, lipoproteins and membranes., (Eds, Vance, D. E. and Vance, J.) Elseviers Science Publishers B.V., Amsterdam., pp.
403.
Davis, R. A. (1997). Evolution of processes and regulators of lipoprotein synthesis: From birds to
mammals. Journal of Nutrition., 127 (Suppl.): 795S-800S.
Davis, R. A. (1999). Cell and molecular biology of the assembly and secretion of apolipoprotein B-containing lipoproteins by the liver. Biochimica et Biophysica Acta., 1440 (1): 1-31.
Davis, R. A., Prewett, A. B., Chan, D. C. F., Thompson, J. J., Berchardt, R. A. and Gallaher, W. R. (1989). Intrahepatic assembly of very low density lipoproteins: immunologic characterization of apolipoprotein B in lipoproteins and hepatic membrane fractions and its intracellular distribution.
Journal of Lipid Research., 30(8): 1185-1196.
De Langen, C. D. (1916). Cholesterol exchange and pathology of race. Presse Med., 24: 332-333.
De Langen, C. D. (1922). Cholesterol content of blood in India. Geneeskundig Tijdschrift v Nederland
Indie., 62: 5.
De Lima, J. J., Latrilha, M. d. C, Toffoletto, O., Ianhez, L. E., Krieger, E. M. and Maranhao, R. C. (1998). Plasma kinetics of chylomicron-like emulsion in renal transplant patients receiving cyclosporin-based immunosuppression. Clinical Cardiology., 21 (6): 411-413.
De Man, F. H. A. F., Castro Cabezas, M., Van Barlingen, H. H. J. J., Erkelens, D. W. and De Bruin, T. W . A. (1996). Triglyceride-rich lipoproteins in non-insulin dependent diabetes mellitus: post-prandial metabolism and relation to premature atherosclerosis. European Journal of Clinical Investigation., 26:
89-108.
De Ruyter, M. G. M. and De Leenheer, A. P. (1978). Simultaneous Determination of Retinol and Retinyl Esters in Serum or Plasma by Reversed-Phase High-Performance Liquid Chromatography.
Clinical Chemistry., 24 (11): 1920-1923.
De Villiers, W. J. S., Coetzee, G. A. and Van der Weshuyzen, D. R. (1994). Lipoprotein receptors.
Handbook Exp. Pharmacol, 109: 53-63.
Demacker, P. N. M., van Hiejst, P. J. and Stalenhoef, A. F. H. (1992). A study of the chylomicron metabolism in W H H L rabbits after fat loading. Discrepancy between results based on measurement of
apoprotein B-48 or retinyl palmitate. Biochemical Journal, 285: 641-646.
Devaraj, S., Vega, G., Lange, R., Grundy, S. M. and Jialal, I. (1998). Remnant-like particle cholesterol levels in patients with dysbetalipoproteinemia or coronary artery disease. American Journal of
Medicine., 104 (5): 445-450.
Deykin, D. and Goodman, D.-W. S. (1962). The Hydrolysis of Long-chain Fatty Acid Esters of Cholesterol with Rat Liver Enzymes. The Journal of Biological Chemistry., 237 (12): 3649-3656.
Dietschy, J. M., Turley, S. D. and Spady, D. K. (1993). Role of liver in the maintenance of cholesterol and low density lipoprotein homeostasis in different animal species, including humans. Journal of
Lipid Research., 34(10): 1637-1659.
Dolphin, P. J. (1985). Lipoprotein metabolism and the role of apolipoproteins as metabolic programmers. Canadian Journal of Biochemistry and Cell Biology., 63 (8): 850-869.
Driscoll, D. M. and Cassanova, E. (1990). Characterisation of the apolipoprotein B mRNA editing activity in enterocyte extracts. The Journal of Biological Chemistry., 265: 21401-21403.

330
Du, E. Z., Wang, S.-L., Kayden, H. J., Sokol, R., Curtiss, L. K. and Davis, R. A. (1996). Translocation of apolipoprotein B across the endoplasmic reticulum is blocked in abetalipoproteinemia. Journal of Lipid Research., 37 (6): 1309-1315.
Durlach, V., Attia, N., Zabouani, A., Leutenagger, M. and Girard-Globa, A. (1996). Postprandial cholesteryl ester transfer and high density lipoprotein composition in normotriglyceridemic non-insulin-dependent diabetic patients. Atherosclerosis., 120: 155-165.
Dzau, V. J. (1994). Pathobiology of atherosclerosis and plaque complications. American Heart Journal, 128: 1300-1304.
Eisenberg, S. and Schurr, D. (1976). Phospholipid removal during degradation of rat plasma very low density lipoprotein in vitro. Journal of Lipid Research., 17: 578.
Ekman, R. and Nilsson-Ehle, P. (1975). Effects of apolipoproteins on lipoprotein lipase activity of human adipose tissue. Clinica Chimica Acta., 63 (1): 29-35.
Ekman, S., Derksen, A. and Small, D. M. (1988). The partitioning of fatty acid and cholesterol between core ans surfaces of phosphatidlycholine-triolein emulsions at pH 7.4. Biochimica et Biophysica Acta., 959: 343-348.
Elliot, W. H. and Waller, G. R. (1972). Biochemical Applications of Mass Spectrometry., (Ed, Waller, G. R.) Wiley., N e w York., pp. 499.
Ellsworth, J. L., Cooper, A. D. and Kraemer, F. B. (1986). Evidence that chylomicron remnants and B-V L D L are transported by the same receptor pathway in J774 murine macrophage-derived cells. Journal of Lipid Research., 27: 1062-1072.
Ellsworth, J. L., Cooper, A. D. and Kraemer, F. B. (1987). Transport of B-very low density lipoproteins and chylomicron remnants by macrophages is mediated by the low density lipoprotein receptor pathways. The Journal of Biological Chemistry., 262: 2316-2325.
Ellsworth, J. L., Fong, L. G., Kraemer, F. B. and Cooper, A. D. (1990). Differences in the processing of chylomicron remnants and beta-VLDL by macrophages. Journal of Lipid Research., 31: 1399-1411.
Elovson, J., Chatterton, J. E., Bell, G. T., Schumaker, V. N., Reuben, M. A., Puppione, D. L., Reeve, J. R. J. and Young, N. L. (1988). Plasma very low density lipoproteins contain a single molecule of apolipoprotein B. Journal of Lipid Research., 29 (11): 1461-1473.
Endemann, G., Stanton, L. W., Madden, K. S., Bryant, C. M., White, R. T. and Protter, A. A. (1993). CD36 is a receptor for oxidised low density lipoproteins. The Journal of Biological Chemistry., 268
(16): 11811-11816.
Erkelens, D. W., Chen, C, Mitchell, C. D. and Glomset, J. A. (1981). Studies of the Interaction Between Apolipoproteins A and C and Triacylglycerol-Rich Particles. Biochimica et Biophysica Acta.,
665: 221-223.
Eto, M., Watanabe, K, Iwashima, Y., Morikawa, A., Chonan, N., Oshima, E., Sekiguchi, M. and Ishii, K. (1987). Increased frequency of apolipoprotein epsilon 4 allele in type II diabetes with hypercholesterolemia. Diabetes., 36 (11): 1301-1306.
Fielding, B. A., Callow, J., Owen, R. M., Samra, J. S., Matthews, D. R. and Frayn, K. N. (1996). Postprandial lipemia: the origin of an early peak studied by specific dietary fatty acid intake during sequential meals. American Journal of Clinical Nutrition., 63: 36-41.
Fielding, C. J. (1978b). Metabolism of cholesterol-rich chylomicrons. Mechanism of binding and uptake of cholesteryl esters by the vascular bed of the perfused rat heart. Journal of Clinical
Investigation., 62: 141.

331
Fielding, C. J. (1992). Lipoprotein receptors, plasma cholesterol metabolism, and the regulation of cellular free cholesterol concentration. FASEB J., 6: 3162-3168.
Filip, D. A., Nistor, A., Bulla, A., Radu, A., Lupu, F. and Simionescu, M. (1987). Cellular events in the development of valvular atherosclerotic lesions induced by experimental hypercholesterolemia. Atherosclerosis., 67 (2-3): 199-214.
Fillios, L. C. and Mann, G. V. (1956.). The importance of sex in the variability of the cholesterolaemic response of rabbits fed cholesterol. Circulation Res., 4: 406-412.
Floren, C.-H., Albers, J. J. and Bierman, E. L. (1981). Uptake of chylomicron remnants causes cholesterol accumulation of cultured human arterial smooth muscle cells. Biochimica et Biophysica Acta., 663: 336-349.
Floren, C.-H. and Chait, A. (1981). Uptake of chylomicron remnants by the native LDL receptor in human monocyte-derived macrophages. Biochimica et Biophysica Acta., 665: 608-611.
Floren, C.-H. and Nilsson, A. (1977a). Degradation of Chylomicron Remnant Cholesterol Ester By Rat Hepatocyte Monolayers. Inhibition By Chloroquine and Colchicine. Biochemical and Biophysical Research Communications., 74 (2): 520-528.
Floren, C.-H. and Nilsson, A. (1977b). Binding, Interiorization and Degradation of Cholesteryl Ester-Labelled Chylomicron-Remnant Particles by Rat Hepatocyte Monolayers. Biochemical Journal, 168.: 483-494.
Foger, B. and Patsch, J. R. (1993). Strategies and methods for the assessment of disturbed postprandial lipid metabolism. Current Opinion in Lipidology., 4: 428-433.
Folch, J., Lees, M. and Sloane-Stanley, G. M. (1957). A simple method for the isolation and purification of total lipids from animal tissues. The Journal of Biological Chemistry., 226: 497-509.
National Heart Foundation.. (1999). Heart, Stroke and Vascular Diseases Australian Facts 1999 Highlights. In Secondary Heart, Stroke and Vascular Diseases Australian Facts 1999 Highlights., Sydney.
Francke, U., Brown, M. S. and Goldstein, J. L. (1984). Assignment of the human gene for the low density lipoprotein receptor to chromosome 19: synteny of a receptor, a ligand and a genetic disease. Proc. Natl. Acad. Sci. U.S.A., 81: 2826-2830.
Fraser, R. (1970). Size and lipid composition of chylomicrons of different Svedberg units of flotation.
Journal of Lipid Research., 11: 60-65.
Fraser, R., Cliff, W. J. and Courtice, F. C. (1968). The effect of dietary fat load on the size and composition of chylomicrons in the thoracic duct. Quarterly Journal of Experimental Physiology., 53:
390-398.
Frederickson, D. S. and Levy, R. I. (1972). In Familial Hyperlipoproteinemia. The Metabolic Basis of Inherited Disease., (Eds, Stanbury, J. B., Wyngaarden, J. B. and Frederickson, D. S.) McGraw-Hill.,,
pp. 545.
Fredrickson, D. S., Levy, R. I. and Lees, R. S. (1967). Fat transport in lipoproteins - an integrated approach to mechanisms and disorders. New England Journal of Medicine., 276: 34, 94, 148, 215,273.
Freeman, M. E., Rohrer, Y. L., Penman, M., Freedman, N. J., Chisholm, G. M. and Krieger, M. (1991). Expression of type I and type II bovine scavenger receptors in Chinese hamster ovary cells: Lipid droplet accumulation and nonreciprocal cross competition by acetylated and oxidised low density
lipoproteins. Proc. Natl. Acad. Sci. U.S.A., 88: 4931-4935.

332
Fujioka, Y., Cooper, A. D. and Fong, L. G. (1998). Multiple processes are involved in the uptake of chylomicron remnants by mouse peritoneal macrophages. Journal of Lipid Research., 39 (12): 2339-
2349.
Funahashi, T., Yokoyama, S. and Yamamoto, A. (1989). Association of apolipoprotein E with the low density lipoprotein receptor: demonstration of its co-operativity on lipid microemulsion particles. Journal of Biochemistry., 105: 582-587.
Furr, H. C. (1990). Reversed-Phase High-Performance Liquid Chromatography of Retinyl Esters.
Methods in Enzymology., 189: 85-95.
Furr, H. C, Barua, A. B. and Olson, J. A. (1994). In Analytical Methods. The Retinoids: Biology, Chemistry and Medicine., (Eds, Sporn, M . B., Roberts, A. B. and Goodman, D. S.) Raven Press., N e w
York., pp. 179-209.
Furr, H. C, Cooper, D. A. and Olson, J. A. (1986). Separation of retinyl esters by non-aqueous reversed-phase high-performance liquid chromatography. Journal of Chromatography., 378 (1): 45-53.
Futterman, S. and Andrews, J. S. (1964b). The composition of liver vitamin A ester and the synthesis of vitamin A ester by liver microsomes. The Journal of Biological Chemistry., 239: 4077-4080.
Gage, S. H. (1920). The free granules (chylomicrons) of fresh blood as shown by the dark-field microscope, and their dependence upon the kind of food ingested. Anat. Rec, 18: 233-234.
Gage, S. H. and Fish, P. A. (1924.). Fat digestion, adsorption, and assimilation in man and animals as determined by the dark field microscope, and a fat-soluble dye. American Journal of Anatomy., 34: 1-
86.
Gandarias, J. M., Lacort, M. and Ochoa, B. (1984). Cholesteryl Ester Hydrolysis in Rat Liver Cytosol.
Modulation by Female Sex Hormones. Lipids., 19 (12): 916-922.
Genest, J., Sniderman, A., Cianflone, K., Teng, B., Wacholder, S., Marcel, Y. and Kwiterovich, P. J. (1986). Hyperabobetalipoproteinemia. Plasma lipoprotein responses to oral fat load. Arteriosclerosis., 6
(3): 297-304.
Genest, J. J., McNamara, J. R., Salem, D. N. and Schaefer, E. J. (1991). Prevalence of risk factors in men with premature coronary artery disease. American Journal of Cardiology., 67 (15): 1185-1189.
Gennat, H. C, OHanlon, J. P. and Mamo, J. C. (unpublished observations). Hypertriglyceridemia in insulin deficiency is due to a decrease in high affinity clearance of triglyceride-rich lipoproteins, not a
decrease in hydrolysis or overproduction.
Gerrity, R. G., Goss, J. and Soby, L. (1985). Control of monocyte recruitment by chemotactic factor(s)
in lesion-prone areas of swine aorta. Arteriosclerosis., 5: 55-65.
Ghosh, S. and Grogan, W. M. (1991). Rapid three-step purification of a hepatic neutral cholesteryl
ester hydrolase which is not the pancreatic enzyme. Lipids., 26 (10): 793-798.
Ghosh, S., Kounnas, M. Z. and Grogan, W. M. (1990). Separation and differential activation of rat liver cytosolic' cholesteryl ester hydrolase, triglyceride lipase and retinyl palmitate hydrolase by
cholestyramine and protein kinases. Lipids., 25 (4): 221-225.
Gianturco, S. H. and Bradley, W. A. (1987). In Lipoprotein receptors. Plasma lipoproteins., (Ed, Gotto,
A. M.) Elsevier Science Publishers B.V., Amsterdam., pp. 190-191.
Gierens, H., Nauck, M., Roth, M., Schinker, R., Schurmann, C, Scharnagl, H., Neuhaus, G., Wieland, H. and Mare, W . (2000). Interleukin-6 stimulates L D L receptor gene expression via activation of sterol-responsive and Spl binding elements. Arteriosclerosis Thrombosis and Vascular Biology, 20:
1777-1783.

333
Gillotte, K. L., Zaiou, M., Lund-Katz, S., Anantharamaiah, G. M., Holvoet, P., Dhoest, A., Palgunachari, M . N., Segrest, J. P., Weisgraber, K. H., Rothblat, G. H. and Phillips, M. C. (1999). Apolipoprotein-mediated Plasma Membrane Microsolubilization. Role of lipid affinity and membrane penetration in the efflux of cellular cholesterol and phospholipid. The Journal of Biological Chemistry., 274 (4): 2021-2028.
Ginsberg, H. N. (1994). Lipoprotein metabolism and its relationship to atherosclerosis. Medical Clinics of North America., 78: 1-20.
Glickman, R. M. and Green, P. H. R. (1977). The intestine as a source of apo A-I. Proc. Natl. Acad. Sci. U.S.A., 74: 2569-2573.
Glickman, R. M. and Magun, A. M. (1986). High density lipoprotein formation in the intestine. Methods in Enzymology., 129: 519-536.
Glickman, R. M. and Sabesin, S. M. (1988). In The Liver: Biology and Pathobiology. Vol. 2nd. (Eds, Arias, I. M., Jackoby, W . B. and Popper, H.) Raven Press., N e w York., pp. 331-354.
Glomset, J. A. (1968). The plasma lecithinxholesterol acyltransferase reaction. Journal of Lipid Research., 9: 155-167.
Glomset, J. A. and Norum, K. R. (1973). The metabolic role of lecithimcholesterol acyl-transferase: perspective from pathology. Adv. Lipid Res., 11: 1-65.
Goldstein, J. L., Basu, S. K., Brunschede, G. Y. and Brown, M. S. (1976). Release of low density lipoprotein from its cell surface receptor by sulfated glycosaminoglycans. Cell, 7 (1): 85-95.
Goldstein, J. L. and Brown, M. S. (1973). Familial hypercholesterolemia: identification of a defect in the regulation of 3-hydroxy-3-methylglutaryl coenzyme A reductase activity associated with overproduction of cholesterol. Proc. Natl. Acad. Sci. U.S.A., 70 (10): 2804-2808.
Goldstein, J. L. and Brown, M. S. (1974). Binding and degradation of low density lipoproteins by cultured human fibroblasts: comparison of cells from a normal subject and from a patient with homozygous familial hypercholesterolemia. The Journal of Biological Chemistry., 249: 5153-5162.
Goldstein, J. L. and Brown, M. S. (1975). Lipoprotein receptors, cholesterol metabolism, and atherosclerosis. Archives of Pathology & Laboratory Medicine., 99 (4): 181-184.
Goldstein, J. L. and Brown, M. S. (1977). The low density lipoprotein pathway and its relation to
atherosclerosis. Ann. Rev. Biochem., 46: 897-930.
Goldstein, J. L. and Brown, M. S. (1983). In Familial Hypercholesterolemia. The Metabolic Basis of Inherited Disease., (Eds, Stanbury, J. B., Wyngaarden, J. B., Frederickson, D. S., Goldstein, J. L. and Brown, M. S.) McGraw-Hill., N e w York., pp. 672-712.
Goldstein, J. L., Ho, Y. K, Basu, S. K. and Brown, M. S. (1979). Binding site on macrophages that mediates uptake and degradation of acetylated low density lipoprotein, producing massive cholesterol
deposition. Proc. Natl. Acad. Sci. U.S.A., 76: 333-337.
Goodman, D. S. (1980). Vitamin A metabolism. Federation Proc, 39: 2716-2722.
Goodman, D.-W. S., Blomstrand, R., Werner, B., Huang, H. S. and Shiratori, T. (1966). The Intestinal Absorption and Metabolism of Vitamin A and B-Carotene in Man. Journal of Clinical Investigation.,
45: 1615-1623.
Goodman, D.-W. S., Huang, H. S. and Shiratori, T. (1965). Tissue distribution and metabolism of
newly absorbed vitamin A in the rat. Journal of Lipid Research., 6: 390-396.

334
Gordon, T., Castelli, W . P., Hjortland, M. C , Kannel, W . B. and Dawber, T. R. (1977). High Density Lipoprotein As a Protective Factor Against Coronary Heart Disease. The Framingham Study. The American Journal of Medicine., 62: 707-714.
Gordon, T., Kannel, W. B., Castelli, W. A. and Dawber, T. R. (1981b). Lipoproteins, cardiovascular diseases and death. The Framingham Study. Archives of Internal Medicine., 141: 1128-1131.
Gotto, A. M. (1983). High-density lipoproteins: biochemical and metabolic factors. American Journal of Cardiology., 52 (4): 2B-4B.
Gown, A. M., Tsukada, T. and Ross, R. (1986). Human atherosclerosis II. Immunocytochemical analysis of the cellular composition of human atherosclerotic lesions. Am. J. Pathol, 125: 191-200.
Greeve, J., Papaderou, M. and Windier, E. (1988). The Significance of the hepatic LDL-receptor for the metabolism of exogenous cholesterol. Agents and Actions., 26 (Suppl.): 173-179.
Greten, H. (1982). Function of hepatic triglyceride lipase in lipoprotein metabolism. Atheroscerosis Review.,: 15-22.
Groot, P. H. E., van Stiphout, W. A. H. J., Krauss, X. H., Jansen, H., van Tol, A., van Ramshorst, E., Chin-On, S., Hofman, A., Cresswell, S. R. and Havekes, L. (1991). Postprandial Lipoprotein Metabolism in Normolipidemic Men With and Without Coronary Artery Disease. Arteriosclerosis and Thrombosis., 11: 653-662.
Grundy, S. M. (1986). Cholesterol and coronary heart disease. A new era. JAMA., 256. (20.): 2849-2858.
Grundy, S. M. (1991). George Lyman Duff Memorial Lecture. Multifactorial etiology of hypercholesterolemia. Implications for prevention of coronary heart disease. Arteriosclerosis & Thrombosis, 11 (6): 1619-35.
Grundy, S. M., Bilheimer, D. W., Blackburn, H., Brown, W. V., Kwiterovich, P. O. J., Mattson, F., Schonfeld, G. and Weidman, W . H. (1982). Rationale of the Diet-heart Statement of the American Heart Association. Report of Nutrition Committee. Circulation., 65 (4): 839a-854a.
Grundy, S. M. and Denke, M. A. (1990). Dietary influences on serum lipids and lipoproteins. Journal of Lipid Research., 31 (7): 1149-1172.
Grundy, S. M. and Mok, H. Y. I. (1976). Chylomicron Clearance in Normal and Hyperlipidemic Man.
Metabolism., 25 (11): 1225-1239.
Guldur, T. and Mayes, P. A. (1992). Metabolic comparison of small chylomicrons (intestinal VLDL) and large chylomicrons. Biochemical Society Transactions., 20: 104S.
Gylling, H., Relas, H., Miettinen, H. E., Radhakrishnan, R. and Miettinen, T. A. (1996). Delayed postprandial retinyl palmitate and squalene removal in a patient heterozygous for apolipoprotein A-1FIN mutation (Leu 159 ® Arg) and low H D L cholesterol level without coronary artery disease.
Atherosclerosis., 127: 239-243.
Gylling, H., Relas, H. and Miettinen, T. A. (1995). Postprandial vitamin A and squalene clearances and cholesterol synthesis off and on lovastatin treatment in type III hyperlipoproteinaemia.
Atherosclerosis., 115: 17-26.
Ha, Y. C. and Barter, P. J. (1982). Differences in plasma cholesteryl ester transfer activity in sixteen
vertebrate species. Comp. Biochem. Physiol, 71B: 265-269.
Ha, Y. C, Chang, L. B. F. and Barter, P. J. (1984). Incubation of human lipid transfer protein with plasma from normal-fed and cholesterol-fed rats: effect on the distribution of constituents between
lipoprotein fractions. Comp. Biochem. Physiol, 78B: 675-682.

335
Ha, Y. C , Chang, L. B. F. and Barter, P. J. (1985). Effect of injecting exogenous lipid transfer protein into rats. Biochimica et Biophysica Acta., 833: 203-210.
Habenicht, A. J. R., JanBen-Timmen, U., Ziegler, R. and Schettler, G. (1994). Lipoproteins and their functions. Clinical Investigation., 72: 639-652.
Hagen, E., Myhre, A. M., Smeland, S., Halvorsen, B., Norum, K. R. and Blomhoff, R. (1999). Uptake of vitamin A in macrophages from physiologic transport proteins: Role of retinol-binding protein and chylomicron remnants. Journal of Nutritional Biochemistry, 10 (6): 345-352.
Haghpassand, M. and Moberly, J. B. (1995). 9-cis-retinoic acid increases apolipoprotein Al secretion and m R N A expression in HepG2 cells. Atherosclerosis, 117 (2): 199-207.
Hallberg, D. (1965). Studies on the Elimination of Exogenous Lipids from the Blood Stream. The Kinetics of the Elimination of a Fat Emulsion Studied by Single Injection Technique in Man. Acta. Physiol. Scand, 64: 306-313.
Hamilton, J. A. and Small, D. M. (1981). Solubilisation and localisation of triolein in phosphatidylcholine bilayers. A 13C N M R study. Proc. Natl. Acad. Sci. U.S.A., 78: 6878-6882.
Hamilton, R. L., Guo, L. S. S., Felker, T. E., Chao, Y.-S. and Havel, R. J. (1986). Nascent high density lipoproteins from liver perfusates of orotic acid-fed rats. Journal of Lipid Research, 27 (9): 967-978.
Hamilton, R. L., Moorehouse, A. and Havel, R. J. (1991). Isolation and properties of nascent lipoproteins from highly purified rat hepatocyte golgi fractions. Journal of Lipid Research, 32: 529-
543.
Hamilton, R. L., Wong, J. S., Guo, S. S., Krisan, S. and Havel, R. J. (1990). Apolipoprotein E localisation in rat hepatocytes by immunogold labeling of cryothin sections. Journal of Lipid Research,
31: 1589-1603.
Harris, K. L. and Felts, J. M. (1970). Kinetics of chylomicron triglyceride removal from plasma in rats: a comparison of the anesthetized and the unanesthetized states. Journal of Lipid Research, 11: 75-81.
Harrison, E. H. (2000). Lipases and carboxylesterases: Possible roles in the hepatic utilisation of vitamin A. Journal of Nutrition, 130 (2 (Suppl. S)): 340S-344S.
Harrison, E. H. and Gad, M. Z. (1989). Hydrolysis of retinyl palmitate by enzymes of rat pancreas and liver. Differentiation of bile salt-dependent and bile salt-independent neutral retinyl ester hydrolases in
rat liver. The Journal of Biological Chemistry, 264: 17142-17147.
Harrison, E. H., Gad, M. Z. and Ross, A. C. (1995). Hepatic uptake and metabolism of chylomicron retinyl esters: probable role of plasma membrane/endosomal retinyl ester hydrolases. Journal of Lipid
Research, 36: 1498-1506.
Hashimoto, S. and Fogelman, A. M. (1980). Smooth microsomes, a trap for cholesteryl ester formed in hepatic microsomes. The Journal of Biological Chemistry, 255 (18): 8678-8684.
Havel, R. (1994). McCollum Award Lecture, 1993: Triglyceride-rich lipoproteins and atherosclerosis -new perspectives. American Journal of Clinical Nutrition, 59: 795-799.
Havel, R. J. (1980). Lipoprotein biosynthesis and metabolism. Ann. N.Y. Acad. Sci., 348: 16-29.
Havel, R. J. (1986b). In Mechanisms of uptake and processing of macromolecules by the liver. Receptor-mediated uptake in the liver, (Eds, Greten, H., Windier, E. and Beisiegel, U.), pp. 1-5.
Havel, R. J. (1988). Role of the liver in reverse cholesterol transport. Agents and Actions, 26 (Suppl.):
125-132.

336
Havel, R. J. (1989). In Contrasts and similarities between the metabolism of intestinal and hepatic lipoproteins: the role of particle size. Intestinal Lipid and Lipoprotein Metabolism, (Eds, Windier, E. and Greten, H.) W . Zuckschwerdt Verlag, Munich, pp. 168-173.
Havel, R. J. (1992). Role of Liver in Hyperlipidemia. Seminars in Liver Disease, 12 (4): 356-363.
Havel, R. J. (1995). Chylomicron remnants: hepatic receptors and metabolism. Current Opinion in Lipidology, 6 (5): 312-316.
Havel, R. J. (1996). Dissection of steps mediating the hepatic uptake and catabolism of chymomicron remnants in rodents - removal from the blood and endocytosis into hepatocytes. Zeitschrift fur Gastroenterologie, 34 (Suppl. 3): 99-102.
Havel, R. J. (1997). Postprandial lipid metabolism: an overview. Proceedings of the Nutrition Society, 56 (2): 659-666.
Havel, R. J. (1998). Receptor and non-receptor mediated uptake of chylomicron remnants by the liver. Atherosclerosis, 141 (Suppl. 1): S1-S7.
Havel, R. J., Chao, Y.-S., Windier, E. E., Kotite, L. and Guo, L. S. S. (1980). Isoprotein specificity in the hepatic uptake of apolipoprotein E and the pathogenesis of familial dysbetalipoproteinemia. Proc. Natl. Acad. Sci. U.S.A. 11 (7): 4349-4353.
Havel, R. J., Fielding, C. J., Olivecrona, T., Shore, V. G., Fielding, P. E. and Egelrud, T. (1973b). Cofactor activity of protein components of human very low density lipoproteins in the hydrolysis of triglycerides by lipoprotein lipase from different sources. Biochemistry, 12: 1828-1833.
Havel, R. J. and Kane, J. P. (1994). In Structure and metabolism of plasma lipoproteins. The Metabolic Basis of Inherited Disease, Qids, Scriver, C. R., Beaudet, A. L., Sly, W . S. and al., e.) McGraw-Hill,
N e w York.
Havel, R. J., Kane, J. P. and Kashyap, M. L. (1973a). Interchange of apolipoprotein between chylomicron and high density lipoproteins during alimentary lipemia in man. Journal of Clinical
Investigation, 52: 32-38.
Havel, R. J., Kita, T., Kotite, L, Kane, J. P., Hamilton, R. L., Goldstein, J. L. and Brown, M. S. (1982). Concentration and Composition of Lipoproteins in Blood Plasma of the W H H L Rabbit. An Animal Model Human Familial Hypercholesterolemia. Arteriosclerosis, 2: 467-474.
Hayashi, H., Fujimoto, K, Cardelli, J. A., Nutting, D. F., Bergstedt, S. and Tso, P. (1990). Fat feeding increases size, but not number of chylomicrons produced by small intestine. American Journal of
Physiology, 259: G709-G719.
Hazzard, W. R. and Bierman, E. L. (1976). Delayed Clearance of Chylomicron Remnants Following Vitamin-A-containing Oral Fat Loads in Broad-B Disease (Type II Hyperlipoproteinemia).
Metabolism, 25 (7): 777-801.
Hegele, R. A., Little, J. A., Vezina, C, GF., M., L, T., TS., W., DJ., J. and PW., C. (1993). Hepatic lipase deficiency. Clinical, biochemical, and molecular genetic characteristics. Arteriosclerosis and
Thrombosis, 13 (5): 720-728.
Helgerud, P., Peterson, L. B. and Norum, K. R. (1983). Retinol esterification by microsomes from the mucosa of human small intestine. Evidence for acyl-coenzyme A retinol acyltransferase activity.
Journal of Clinical Investigation, 71: 747-753.
Hendler, R. W. (1964). Procedure for simultaneous assay of two beta-emitting isotopes with the liquid scintillation counting technique. Analytical Biochemistry, 7: 110-120.
Herbert, P. N., Assmann, G., Gotto, A. M. and Frederickson, D. S. (1982). In Familial lipoprotein deficiency (abetalipoproteinemia and Tangier disease). The Metabolic Basis of Inherited Disease, (Eds,

337
Stanbury, J. B., Wyngaarden, J. B., Frederickson, D. S., Goldstein, J. L. and Brown, M . S.) McGraw-Hill, Inc, N e w York, pp. 589-621.
Herz, J. (1993). The LDL-receptor-related protein - portrait of a multifunctional receptor. Current Opinion in Lipidology, 4: 107-113.
Herz, J., Clouthier, D. E. and Hammer, R. E. (1992). LDL receptor-related protein internalises and degrades uPA-PA-1 complexes and is essential for embryo implantation. Cell, 71: 411-421.
Herz, J., Hamann, U., Rogne, S., Myklebost, O, Gausepohl, H. and Stanley, K. K. (1988). Surface location and high affinity for calcium of a 500-kd liver membrane protein closely related to the L D L receptor suggest a physiological role as lipoprotein receptor. EMBO Journal, 7: 4119-4127.
Herz, J., Kowal, R. C, Goldstein, J. L. and Brown, M. S. (1990). Proteolytic processing of the 600 kD low density lipoprotein receptor-related protein (LRP) occurs in a trans-Golgi compartment. EMBO Journal, 9 (6): 1769-1776.
Herz, J., Qui, S.-Q., Oesterle, A., DeSilva, H. V., Shafi, S. and Havel, R. J. (1995). Initial hepatic removal of chylomicron remnants is unaffected but endocytosis is delayed in mice lacking the low density lipoprotein receptor. Proc. Natl. Acad. Sci. USA, 92: 4611-4615.
Herz, J. and Willnow, T. E. (1995). Functions of the LDL receptor gene family. Ann. NY. Acad. Sci., 737: 14-19.
Hesler, C. B., Swenson, T. L. and Tall, A. R. (1987). Purification and characterisation of a human plasma cholesteryl ester transfer protein. The Journal of Biological Chemistry, 262: 2275-2282.
Hiltunen, T. P., Luoma, J. S., Nikkari, T. and Yla-Herttuala, S. (1998). Expression of LDL receptor, V L D L receptor, L D L receptor-related protein, and scavenger receptor in rabbit atherosclerotic lesions: marked induction of scavenger receptor and V L D L receptor expression during lesion development. Circulation, 97 (11): 1079-1086.
Hirata, M. H., Oliveira, H. C. F., Quintao, E. C. R., Redgrave, T. G. and Maranhao, R. C. (1987). The effects of Triton WR-1339, protamine sulfate and heparin on the plasma removal of emulsion models of chylomicrons and remnants in rats. Biochimica et Biophysica Acta, 917: 344-346.
Hobbs, H. H., Brown, M. S., Goldstein, J. L. and Russell, D. W. (1986). Deletion of exon encoding cysteine-rich repeat of low density lipoprotein receptor alters its binding specificity in a subject with familial hypercholesterolemia. The Journal of Biological Chemistry, 261: 13114-13120.
Hobbs, H. H., Leitersdorf, E., Goldstein, J. L., Brown, M. S. and Russell, D. W. (1988). Multiple crm-mutations in familial hypercholesterolemia. Evidence for 13 alleles, including four deletions. Journal of Clinical Investigation, 81 (3): 909-917.
Hobbs, H. H., Leitersdorf, E., Leffert, C. C, Cryer, D. R., Brown, M. S. and Goldstein, J. L. (1989). Evidence for a dominant gene that suppresses hypercholesterolemia in a family with defective low density lipoprotein receptors. Journal of Clinical Investigation, 84: 656-664.
Hodges, P. and Scott, J. (1992). Apolipoprotein B mRNA editing: a new tier for the control of gene expression. Trends in Biochemical Sciences, 17 (2): 77-81.
Hoff, H. F., Heideman, C. L., Gaubatz, J. W., Titus, J. L. and Gotto, A. M. (1978). Quantitation of apo B in human aortic fatty streaks. A comparison with grossly normal intima and fibrous plaques.
Atherosclerosis, 30: 263.
Hoff, H. F., Heideman, C. L., Jackson, R. L., Bayardo, R. J., Kim, H. and Gotto, A. M. (1975b). Localisation patterns of plasma apolipoproteins in human atherosclerotic lesions. Circulation Research,
37: 72.

338
Hoff, H. F., Heideman, C. L., Noon, G. P. and Meyer, M . S. (1975a). Localisation of apo-lipoproteins in human carotid artery plaques. Stroke, 6: 531.
Holdsworth, G., Stocks, J., Dodson, P. and Galton, D. J. (1982). An abnormal triglyceride-rich lipoprotein containing excess sialylated apolipoprotein C-III. Journal of Clinical Investigation, 69 (4): 932-939.
Holt, P. R., Wu, A. L. and Clark, S. B. (1979). Apoprotein composition and turnover in rat intestinal lymph during steady state triglyceride absorption. Journal of Lipid Research, 20: 494-502.
Howard, B. V. (1987). Lipoprotein metabolism in diabetes mellitus. Journal of Lipid Research, 28: 613-628.
Huang, H. S. and Goodman, D.-W. S. (1965). Vitamin A and Carotenoids. I. Intestinal absorption and metabolism of 14C-labeled vitamin A alcohol and fi-carotene in the rat. The Journal of Biological Chemistry, 240 (7): 2839-2844.
Huettinger, M., Retzek, H., Hermann, M. and Goldenberg, H. (1992). Lactoferrin Specifically Inhibits Endocytosis of Chylomicron Remnants but Not alpha-2~Macroglobulin. The Journal of Biological Chemistry, 267: 18551-18557.
Hui, D. Y., Innerarity, T. L. and Mahley, R. W. (1984a). Defective Hepatic Lipoprotein Receptor Binding of (-Very L o w Density Lipoproteins from Type III Hyperlipoproteinemic Patients. Importance of Apolipoprotein E. The Journal of Biological Chemistry, 259 (2): 860-869.
Hui, D. Y., Innerarity, T. L., Milne, R. W., Marcel, Y. L. and Mahley, R. W. (1984b). Binding of chylomicron remnants and B-very low density lipoproteins to hepatic and extra-hepatic lipoprotein receptors. The Journal of Biological Chemistry, 259: 15060-15068.
Hultin, M., Savonen, R. and Olivecrona, T. (1996). Chylomicron metabolism in rat: lipolysis, recirculation of triglyceride-derived gatty acids in plasma FFA, and fate of core lipids as analyzed by compartmental modelling. Journal of Lipid Research, 37: 1022-1036.
Humphries, S. E. (1986). Familial hypercholesterolemia as an example of early diagnosis of coronary artery disease risk by D N A techniques. British Heart Journal, 56: 201-205.
Hungria, V. T., Brandizzi, L. I., Chiattone, C. S., Bydlowski, S. P. and Maranhao, R. C. (1999). Metabolism of an artificial emulsion resembling chylomicrons in patients with multiple myeloma. Leukemia Research, 23 (7): 637-641.
Hunninghake, D. B. (1988). Current and new therapy for hypercholesterolemia. Journal of the American Osteopathic Association, 88 (7): 865-869.
Hussain, M. M., Innerarity, T. L., W.J., B. and Mahley, R. W. (1995). Chylomicron Metabolism in Normal, Cholesterol-fed, and Watanabe Heritable Hyperlipidemic Rabbits. Saturation of the Sequestration Step of the Remnant Clearance Pathway. The Journal of Biological Chemistry, 270 (15):
8578-8587.
Hussain, M. M., Mahley, R. W., Boyles, J. K., Fainaru, M., Brecht, W. J. and Lindquist, P. A. (1989). Chylomicron-chylomicron remnant clearance by liver and bone marrow in rabbits: factors that modify dssue-specific uptake. The Journal of Biological Chemistry, 264: 9571-9582.
Hussain, M. M., Maxfield, F. R., Mas-Oliva, J., Tabas, I., Ji, Z. S., Innerarity, T. L. and Mahley, R. W. (1991). Clearance of chylomicron remnants by the low density lipoprotein receptor-related protein/alpha 2-macroglobulin receptor. The Journal of Biological Chemistry, 266 (21): 13936-13940.
Hussain, M. M., Strickland, D. K. and Bakillah, A. (1999). The mammalian low-density lipoprotein receptor family (Review). Annual Review of Nutrition, 19: 141-172.

339
Ignatowski, A. (1909). Uber die Wirkung des tierischen Eiweisses auf die Aorta and die parenchymatosen Organe der Kaninchen. Virchows Arch. path. Anat. Physiol, 198: 248-270.
Imaizumi, K., Fainaru, M. and Havel, R. J. (1978a). Composition of proteins of mesenteric lymph chylomicrons in rats and alterations produced upon exposure of chylomicrons to blood serum and serum proteins. Journal of Lipid Research, 19: 712-722.
Imaizumi, K., Havel, R. J., Fainaru, M. and Vigne, J.-L. (1978b). Origin and transport of the A-I and arginine-rich aplipoproteins in mesenteric lymph of rats. Journal of Lipid Research, 19: 1038-1046.
Inazu, A., Brown, M. L., Hesler, C. B., Agellon, L. B., Koizumi, J., Takata, K., Maruhama, Y., Mabuchi, H. and Tall, A. R. (1990). Increased high density lipoprotein caused by a common cholesteryl ester transfer protein gene mutation. New England Journal of Medicine, 323: 1234-1238.
Innerarity, T. L., Bersot, T. P., Arnold, K, Weisgraber, K. H., Davis, P. A., Forte, T. M. and Mahley, R. W . (1984). Receptor binding activity of high-density lipoproteins containing apoprotein E from abetalipoproteinemic and normal neonate plasma. Metabolism, 33 (2): 186-195.
Innerarity, T. L. and Mahley, R. W. (1978). Enhanced binding by cultured human fibroblasts of apo-E-containing lipoproteins as compared with low density lipoproteins. Biochemistry, 17: 1440-1447.
Isherwood, S. G., Williams, C. M. and Gould, B. J. (1997). Apolipoprotein B-48 as a marker for chylomicrons and their remnants: studies in the postprandial state. Proceedings of the Nutrition Society,
56: 497-505.
Ishibashi, S., Herz, J., Maeda, N., Goldstein, J. L. and Brown, M. S. (1994b). The two-receptor model of lipoprotein clearance: Tests of the hypothesis in "knockout" mice lacking the low density lipoprotein receptor, apolipoprotein E, or both proteins. Proc. Natl. Acad. Sci. USA, 91: 4431-4435.
Ishibashi, S., Perrey, S., Chen, Z., Osuga, J., Shimada, M., Ohashi, K, Harada, K., Yazaki, Y. and Yamada, N. (1996). Role of low density lipoprotein (LDL) receptor pathway in the metabolism of chylomicron remnants. A quantitative study in knockout mice lacking the L D L receptor, apolipoprotein E, or both. The Journal of Biological Chemistry, 271 (37): 22422-22427.
Jackie, S., Rinninger, R, Greeve, J., Greten, H. and Windier, E. (1992). Regulation of the hepatic removal of chylomicron remnants and B-very low density lipoproteins in the rat. Journal of Lipid
Research, 33: 429-429.
Jackie, S., Runquist, E., Brady, S., Hamilton, R. L. and Havel, R. J. (1991). Isolation and characterisation of three endosomal fractions from the liver of normal rats after lipoprotein loading.
Journal of Lipid Research, 32: 485-498.
James, R. W., Martin, B., Pometta, D., Fruchart, J. C, Duriez, P., Puchois, P., Farriaux, J. P., Tacquet, A., Demant, T. and Clegg, R. J. (1989). Apolipoprotein B metabolism in homozygous familial
hypercholesterolemia. Journal of Lipid Research, 30 (2): 159.
Janiak, M. J., Loomis, C. R., Shipley, G. G and Small, D. M. (1974). The ternary phase diagram of lecithin cholesterol lineoleate and water: Phase behaviour and structure. Journal of Molecular Biology,
86: 325-329.
Janiak, M. J., Small, D. M. and Shipley, G. G. (1979). Interactions of cholesteryl esters with phospholipids: cholesteryl myristate and dimyristoyl lecithin. Journal of Lipid Research, 20: 183-199. Jeffery, F. and Redgrave, T. G. (1982). Chylomicron catabolism differs between Hooded and albino
laboratory rats. Journal of Lipid Research, 23: 154-160.
Ji, Z.-S., Brecht, W. J., Miranda, R. D., Hussain, M. M., Innerarity, T. L. and Mahley, R. W. (1993). Role of Heparan Sulfate Proteoglycans in the Binding and Uptake of Apolipoprotein E-enriched Remnant Lipoproteins by Cultured Cells. The Journal of Biological Chemistry, 268 (14): 10160-10167.

340
Ji, Z.-S., Fazio, S., Lee, Y.-L. and Mahley, R. W . (1994a). Secretion-capture role for apolipoprotein E in remnant lipoprotein metabolism involving cell surface heparin sulfate proteoglycans. The Journal of Biological Chemistry, 269: 2764-2772.
Ji, Z. S., Lauer, S. J., Fazio, S., Bensadoun, A., Taylor, J. M. and Mahley, R. W. (1994b). Enhanced binding and uptake of remnant lipoproteins by hepatic lipase-secreting hepatoma cells in culture. The Journal of Biological Chemistry, 269 (18): 13429-13436.
Ji, Z.-S., Sanan, D. A. and Mahley, R. W. (1995). Intravenous heparinase inhibits remnant lipoprotein clearance from the plasma and uptake by the liver: in vivo role of heparan sulfate proteoglycans. Journal of Lipid Research, 36: 583-592.
Ji, Z. W. and Mahley, R. W. (1994). Lactoferrin binding to heparan sulphate proteoglycans and the L D L receptor-related protein: further evidence supporting the importance of direct binding to remnants lipoproteins to HSPG. Arteriosclerosis and Thrombosis, 14: 2025-2032.
Jonasson, L., Holm, J., Skalli, O., Bonders, G. and Hansson, G. K. (1986). Regional accumulation of T-cells, macrophages, and smooth muscle cells in the human atherosclerotic plaque. Arteriosclerosis, 6: 131-138.
Jones, A. L., Hradek, G. T., Hornick, H. C, Renaud, G., Windier, E. E. T. and Havel, R. J. (1984). Uptake and processing of remnants of chylomicrons and very low density lipoproteins by rat liver. Journal of Lipid Research, 25: 1151-1158.
Kanai, M., Raz, A. and Goodman, D.-W. S. (1968). Retinol-Binding Protein: the Transport Protein for Vitamin A in Human Plasma. Journal of Clinical Investigation, 47: 2025-2044.
Kane, J. P. (1983). Apoprotein B: structural and metabolic heterogeneity. Ann. Rev. Phys., 45: 637-650.
Kane, J. P. (1996). In Structure and Function of the Plasma Lipoproteins and their receptors. Atherosclerosis and Coronary Artery Disease, OEds, Fuster, V., Ross, R. and Topol, E. J.) Lippincott-Raven Publishers, Philadelphia, pp. 89-103.
Kane, J. P., Hardman, D. A. and Paulus, H. E. (1980). Heterogeneity of apolipoprotein B: Isolation of a new species from human chylomicrons. Proc. Natl. Acad. Sci. U.S.A, 77: 2465-2469.
Kane, J. P. and Havel, R. J. (1994). In Disorders of the biogenesis and secretion of lipoproteins containing the B-apolipoproteins. The Metabolic Basis of Inherited Disease, (Eds, Scriver, C. R., Beaudet, A. L., Sly, W . S. and al., e.) McGraw-Hill, N e w York.
Kannel, W. B. and Castelli, W. A. (1979). Cholesterol in the prediction of atherosclerotic disease. New perspectives from the Framingham Study. Annals of Internal Medicine, 90: 85-92.
Karpe, F. (1997b). Effects of diet on postprandial lipaemia: a suggestion for methodological standardization. Nutrition Metabolism and Cardiovascular Diseases, 7: 44-55.
Karpe, F. (1999). Postprandial lipoprotein metabolism and atherosclerosis. Journal of Internal
Medicine, 246 (4): 341-355.
Karpe, F., Bell, M., Bjorkegren, J. and Hamsten, A. (1995). Quantification of Postprandial Triglyceride-Rich Lipoproteins in Healthy M e n by Retinyl Ester Labeling and Simultaneous Measurement of Apolipoproteins B-48 and B-100. Arteriosclerosis Thrombosis and Vascular Biology,
15: 199-207.
Karpe, F. and Hamsten, A. (1995). Postprandial lipoprotein metabolism and atherosclerosis. Current
Opinion in Lipidology, 6: 123-129.
Karpe, F., Hamsten, A., Uffelman, K. and Steiner, G. (1996). Apolipoprotein B-48. Methods in
Enzymology, 263: 95-104.

341
Karpe, F., Olivecrona, T., Hamsten, A. and Hultin, M . (1997b). Chylomicron/chylomicron remnant turnover in humans: evidence for margination of chylomicrons and poor conversion of larger to smaller chylomicron remnants. Journal of Lipid Research, 38: 949-961.
Karpe, F., Steiner, G„ Uffelman, K., Olivecrona, T. and Hamsten, A. (1994). Postprandial lipoproteins and progression of coronary atherosclerosis. Atherosclerosis, 106: 83-97.
Kashyap, M. L., Hynd, B. A., Robinson, K. and Gartside, P. S. (1981). Abnormal preponderance of sialylated apolipoprotein CIII in triglyceride rich lipoproteins in type V hyperlipoproteinemia. Metabolism, 30(2): 111-118.
Kesaniemi, Y. A., Witzum, J. L. and Steinbrecher, U. P. (1983). Receptor-mediated catabolism of low density lipoprotein in man. Journal of Clinical Investigation, 71: 950.
Keys, A. (1970). Coronary heart disease in seven countries. Circulation, 41 (Suppl.): 1-211.
Keys, A. (1988). Dietary lipids and ischaemic heart disease - progress in 20 years. American Journal of Clinical Nutrition, 47: 965-969.
Keys, A., Menotti, A., Karvonen, M. J., Aravanis, C, Blackburn, H., Buzina, R., Djordjevic, B. S., Dontas, A. S., Fidanza, F. and Keys, M . H. (1986). The diet and 15-year death rate in the seven countries study. American Journal of Epidemiology, 124 (6): 903-915.
Khachadurian, A. K. (1971). A general review of clinical and laboratory features of familial hypercholesterolemia. Proc. Biol. Fluids., 19: 315-318.
Kinlay, S. (1988). High cholesterol levels: is mass screening the best option? Medical Journal of Australia, 148: 635-637.
Kinnunen, P. K. J., Virtanen, J. A. and Vainio, P. (1983). Lipoprotein lipase and hepatic lipase: Their roles in plasma metabolism. Atheroscerosis Review, 11: 65-105.
Kita, T., Brown, M. S., Bilheimer, D. W. and J.L., G. (1982-a). Delayed clearance of very low density and intermediate density lipoproteins with enhanced conversion to low density lipoproteins in W H H L rabbits. Proc. Natl. Acad. Sci. U.S.A, 19: 5693.
Kita, T., Brown, M. S., Watanabe, Y. and Goldstein, J. L. (1981). Deficiency of low density lipoprotein receptors in liver and adrenal gland of the W H H L rabbit, an animal model of familial hypercholesterolemia. Proc. Natl. Acad. Sci. U.S.A, 78: 2268.
Kita, T., J.L., G., Brown, M. S., Watanabe, Y., Hornick, C. A. and Havel, R. J. (1982-b). Hepatic uptake of chylomicron remnants in W H H L rabbits: A mechanism genetically distinct from the low density lipoprotein receptor. Proc. Natl. Acad. Sci. USA, 79: 3623-3627.
Kodama, T., Freeman, M., Rohrer, L., Zabrecky, J., Matsudaira, P. and Krieger, M. (1990). Type I macrophage scavenger receptor contains &-helical and collagen-like coiled coils. Nature, 343: 531-
535.
Koizumi, J., Inazu, A., Kuminas, Y., Ichiro, K, Uno, Y., Kajinami, K, Miyamoto, S., Moulin, P., Tall, A. R. and Mabuchi, H. (1991). Serum lipoprotein lipids concentrations and composition in homozygous and heterozygous patients with cholesteryl ester transfer protein deficiency.
Atherosclerosis, 90(2-3): 189-196.
Koo, C, Wernette-Hammond, M. E., Garcia, Z., Malloy, M. J., Uauy, R., East, C, Bilheimer, D. W., Mahley, R. W . and Innerarity, T. L. (1988). Uptake of cholesterol-rich remnant lipoproteins by human monocyte-derived macrophages is mediated by low density lipoprotein receptors. Journal of Clinical
Investigation, 81 (5): 1332-1340.

342
Koo, C , Wernette-Hammond, M . E. and Innerarity, T. L. (1986). Uptake of canine beta-very-low density lipoproteins by mouse peritoneal macrophages is mediated by a low density lipoprotein receptor. The Journal of Biological Chemistry, 261: 11194-11201.
Kostner, G. M. and Karadi, I. (1988). Lipoprotein alterations in diabetes mellitus. Diabetologia, 31: 717-722.
Kovanen, P. T., Brown, M. S., Basu, S. K., Bilheimer, D. W. and Goldstein, J. L. (1981). Saturation and suppression of hepatic lipoprotein receptors: a mechanism for the hypercholesterolemia of cholesterol-fed rabbits. Proc. Natl. Acad. Sci. U.S.A. 78 (3): 1396-1400.
Kowal, R. C, Herz, J., Goldstein, J. L., Esser, V. and Brown, M. S. (1989). Low density lipoprotein receptor-related protein mediates uptake of cholesteryl esters derived from apoprotein E-enriched lipoproteins. Proc. Natl. Acad. Sci. U.S.A, 86: 5810-5814.
Kowal, R. C, Herz, J., Weisgraber, K. H., Mahley, R. W., Brown, M. S. and Goldstein, J. L. (1990). Opposing effects of apolipoproteins E and C on lipoprotein binding to low density lipoprotein receptor-related protein. The Journal of Biological Chemistry, 265: 10771-10779.
Kraemer, F. B. (1987). Diabetes and lipoprotein receptors. Diabetes-Metabolism Reviews, 3 (3): 591-618.
Krapp, A., Ahle, S., Kersting, S., Hua, Y., Knoser, K., Nielsen, M., Gliemann, J. and Beisegel, U. (1996). Hepatic lipase mediates the uptake of chylomicrons and B - V L D L int cells via the L D L receptor-related protein 0-RP). Journal of Lipid Research, 37: 926-936.
Krasinski, S. D., Cohn, J. S., Russell, R. M. and Schaefer, E. J. (1990b). Postprandial Plasma Vitamin A Metabolism in Humans: A Reassessment of the Use of Plasma Retinyl Esters as Markers for Intestinally Derived Chylomicrons and Their Remnants. Metabolism, 39 (4): 357-365.
Krasinski, S. D., Cohn, J. S., Schaefer, E. J. and Russell, R. M. (1990a). Postprandial Plasma Retinyl Ester Response Is Greater in Older Subjects Compared with Younger Subjects. Evidence for Delayed Plasma Clearance of Intestinal Lipoproteins. Journal of Clinical Investigation, 85: 883-892.
Krieger, M. and Herz, J. (1994). Structures and functions of multiligand lipoprotein receptors: Macrophages, scavenger receptors, L D L receptor-related protein (LRP). Ann. Rev. Biochem., 63: 601-
637.
Krishnaiah, K. V., Walker, L. F., Borensztajn, J., Schonfeld, G. and Getz, G. S. (1980). Apolipoprotein B variant derived from rat intestine. Proc. Natl. Acad. Sci. U.S.A, 11 (7): 3806-3810.
Kristensen, T., Moestrup, S. K., Glieman, J., Bendtsen, L., Sand, O. and Sottrup-Jensen, L. (1990). Evidence that the newly cloned low density lipoprotein receptor-related protein (LRP) binds is the a2-
macroglobulin receptor. FEBS Letters, 276: 151-155.
Kritchevsky, D. (1969). Experimental atherosclerosis in primates and other species. Ann. NY. Acad.
Set, 162 (80-88.).
Kushwaha, R. S. and W.R., H. (1978). Catabolism of very low density lipoproteins in the rabbit. Effect of changing composition and pool size. Biochimica et Biophysica Acta, 528: 176.
Kutt, H., Herz, J. and Stanley, K. K. (1989). Structure of the low-density lipoprotein receptor-related protein (LRP) promoter. Biochimica et Biophysica Acta, 1009 (3): 229-236.
Kwiterovich, P. O., Frederickson, D. S. and Levy, R. I. (1974). Familial hypercholesterolemia (one form of familial type II hyperlipoproteinemia). A study of its biochemical, genetic, and clinical
presentation in childhood. Journal of Clinical Investigation, 53: 1237-1249.
Laird, C. W., Fox, R. R., Schultz, H. S., Mitchell, B. P. and Blau, E. M. (1970). Strain variations in rabbits: biochemical indicators of thyroid functions. Life Sciences, 9 (2): 203-214.

343
Langer, T., Strober, W . and Levy, R. I. (1972). The metabolism of low density lipoprotein in familial type II hypercholesterolemia. Journal of Clinical Investigation, 51: 1528-1536.
Lawn, R. M., Wade, D. P., Garvin, M. R., Wang, X., Schwartz, K., Porter, J. G., Seilhamer, J. J., Vaughan, A. M . and Oram, J. F. (1999). The Tangier disease gene product A B C 1 controls the cellular apolipoprotein-mediated lipid removal pathway. Journal of Clinical Investigation, 104 (8): R25-R31.
Le, N.-A., Coates, P. M., Gallagher, P. R. and Cortner, J. A. (1997). Kinetics of Retinyl Esters During Postprandial Lipemia in Man: A Compartmental Model. Metabolism, 46 (5): 584-594.
Le, N.-A., Gibson, J. C. and Ginsberg, H. N. (1988). Independent regulation of plasma apolipoprotein C-II and C-III concentrations in very low density and high density lipoproteins: implications for the regulation of the catabolism of these lipoproteins. Journal of Lipid Research, 29 (5): 669-677.
Leclercq, C, Arcella, D. and Turrini, A. (2000). Estimates of the theoretical maximum daily intake of erythorbic acid, gallates, butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT) in Italy: a stepwise approach. Food & Chemical Toxicology, 38 (12): 1075-84.
Lees, R. S. and Wilson, D. E. (1971). The treatment of hyperlipidemia. New England Journal of Medicine, 284: 186-195.
Leitersdorf, E., Van der Westhuyzen, D. R., Coetzee, G. A. and Hobbs, H. H. (1989). Two common low density lipoprotein receptor gene mutations cause familial hypercholesterolemia in Afrikaners. Journal of Clinical Investigation, 84 (3): 954-961.
Lenich, C. M. a. and Ross, A. C. (1987). Chylomicron remnant-vitamin A metabolism by the human hepatoma cell line HepG2. Journal of Lipid Research, 28: 183-194.
Lentz, B. R., Barenholz, Y. and Thompson, T. E. (1975). A simple method for the synthesis of cholesterol esters in high yield. Chemistry and Physics of Lipids, 15: 216-221.
Lenzo, N. P., Martins, I., Mortimer, B. C. and Redgrave, T. G. (1988). Effects of phospholipid composition on the metabolism of triacylglycerol, cholesteryl ester and phosphatidylcholine from lipid emulsions injected intravenously in rats. Biochimica et Biophysica Acta, 960: 111-118.
Lestavel, S. and Fruchart, J. C. (1994). Lipoprotein Receptors. Cellular and Molecular Biology, 40 (4):
461-481.
Levine, G. N., Keaney, J. F. and Vita, J. A. (1995). Cholesterol reduction in cardiovascular disease. New England Journal of Medicine, 332: 512-521.
Levine, Y. K. and Wilkins, M. H. F. (1971). Structure of oriented lipid bilayers. Nature (London), New
Biology, 230 (11): 69-72.
Levy, E., Shafrir, E., Ziv, E. and Bar-On, H. (1985). Composition, removal and metabolic fate of chylomicrons derived from diabetic rats. Biochimica et Biophysica Acta, 834: 376-385.
Levy, R. I., Fredrickson, D. S., Shulman, R., Bilheimer, D. W., Breslow, J. L., Stone, N. J., Lux, S. E., Sloan, H. R., Krauss, R. M . and Herbert, P. N. (1972). Dietary and drug treatment of primary hyperlipoproteinemia. NTH Conference Discussion. Annals of Internal Medicine, 11 (2): 267-294.
Lewis, G. F., OMeara, N. M., Soltys, P. A., Blackman, J. D., Iverius, P. H., Pugh, W. L., Getz, G. S. and Polonsky, K. S. (1991). Fasting Hypertriglyceridemia in Noninsulin-Dependent Diabetes Mellitus Is an Important Predictor of Postprandial Lipid and Lipoprotein Abnormalities. Journal of Clinical Endocrinology and Metabolism, 72: 934-944.
Li, Q.-T. and Sawyer, W. H. (1992). Effect of unesterified cholesterol on the compartmentation of a fluorescent cholesteryl ester in a lipoprotein-like lipid emulsion. Journal of Lipid Research, 33: 503-
512.

344
Li, Q.-T. and Sawyer, W . H. (1993). Effect of cholesteryl ester on the distribution of flourescent cholesterol analogues in triacylglycerol-rich emulsions. Biochimica et Biophysica Acta, 1166: 145-153.
Li, Q.-T., Tilley, L, Sawyer, W. H., Looney, F. and Curtain, C. C. (1990). Structure and dynamics of microemulsions which mimic the lipid phase of low-density lipoproteins. Biochimica et Biophysica Acta, 1042: 42-50.
Linder, C, Chernick, S. S., Fleck, T. R. and Scow, R. O. (1976). Lipoprotein lipase and uptake of chylomicron triglyceride by skeletal muscle of rats. American Journal of Physiology, 231: 1859-1867.
Lippiello, P. M., Dijkstra, J., van Galen, M., Scherphof, G. and Waite, B. M. (1981). The Uptake and Metabolism of Chylomicron-remnant Lipids by Nonparenchymal Cells in Perfused Liver and by Kupffer Cells in Culture. The Journal of Biological Chemistry, 256: 7454-7460.
Lombardi, P., Sijbrands, E. J., van de Giessen, K., Smelt, A. H., Kastelein, J. J., Frants, R. R. and Havekes, L. M . (1995). Mutations in the low density lipoprotein receptor gene of familial hypercholesterolemic patients detected by denaturing gradient gel electrophoresis and direct sequencing. Journal of Lipid Research, 36 (4): 860-867.
Lovegrove, J. A., Jackson, K. G., Murphy, M. C, Brooks, C. N., Zampelas, A., Knapper, J. M. E., Wright, J. W., Gould, B. J. and Williams, C. M . (1999). Markers of intestinally-derived lipoproteins: Application to studies of altered diet and meal fatty acid composition. Nutr. Metab. Cardiovasc. Dis., 9:9-18.
Lund, H., Takahashi, K., Hamilton, R. L. and Havel, R. J. (1989). Lipoprotein binding and endosomal itinerary of the low density lipoprotein receptor-related protein in rat liver. Proc. Natl. Acad. Sci. U.S.A, 86 (23): 9318-9322.
Luo, C.-C, Li, W.-H., Moore, M. N. and Chan, L. (1986). Structure and evolution of the apoliprotein multigene family. Journal of Molecular Biology, 187 (3): 325-340.
Lusis, A. J., Taylor, B. A., Quon, D., Zollman, S. and LeBoeuf, R. C. (1987). Genetic factors controlling structure and expression of apolipoproteins B and E in mice. The Journal of Biological Chemistry, 262 (16): 7594-7604.
Ly, H. L., Mortimer, B.-C, Baker, E. and Redgrave, T. G. (1992). Clearance from plasma of lymph chylomicrons and chylomicron remnants labelled with 1251-tyramine cellobiose. Biochemical Journal,
286 Cart 3): 937-943.
Mabuchi, H., Itoh, M„ Takeda, M., Kajinami, K., Wakasugi, T., Koizumi, J., Takeda, R. and Asagami, C. (1989). A young type III hyperlipoproteinemic patient associated with apolipoprotein E deficiency.
Metabolism, 38 (2): 115-119.
Mabuchi, H., Miyamoto, S., Ueda, K., Oota, M., Takegoshi, T., Wakasugi, T. and Takeda, R. (1986). Causes of death in patients with familial hypercholesterolemia. Atherosclerosis, 61 (1): 1-6.
MacDonald, P. N. and Ong, D. E. (1988). Evidence for a lecithin-retinol-acyltransferase activity in the rat small intestine. The Journal of Biological Chemistry, 263: 12478-12482.
Mackintosh, V. S., Phan, C. T., Mortimer, B.-C. and Redgrave, T. G. (1996). Vasoactive Mediators Affect the Clearance of Lipids from Emuslion Models of Plasma Lipoproteins in Rats. Journal of
Cardiovascular Pharmacology, 27: 447-454.
Mackness, M. I. and Durrington, P. N. (1992). Lipoprotein Separation and Analysis for Clinical Studies. Lipoprotein Analysis. A Practical Approach. IRL Press, Oxford University Press, Oxford,
N e w York, Tokyo.

345
Mahadevan, S., Seshadri Sastry, P. and Ganguly, J. (1963). Studies on the Metabolism of Vitamin A. 4. Studies of the mode of absorption of Vitamin A by rat intestine in vitro. Biochemical Journal, 88: 534-539.
Mahley, R. W. (1988). Apolipoprotein E: Cholesterol Transport Protein with Expanding Role in Cell Biology. Science, 240: 622-630.
Mahley, R. W. (1990). In Principles and Practice of Endocrinology and Metabolism. (Ed, Becker, K. L.) Lippincott Company, London, pp. 1219-1229.
Mahley, R. W. and Angelin, B. (1984). Type III hyperlipoproteinemia: recent insights into the genetic defect of familial dysbetalipoproteinemia. Adv. Int. Med., 29: 385-411.
Mahley, R. W. and Holcombe, K. S. (1977). Alterations of the plasma lipoproteins and apoproteins following cholesterol feeding in the rat. Journal of Lipid Research, 18 (3): 314-324.
Mahley, R. W., Hui, D. Y., Innerarity, T. L. and Beisegel, U. (1989). Chylomicron remnant metabolism: role of hepatic lipoprotein receptors in mediating uptake. Arteriosclerosis, 19 (Suppl.): 114-118.
Mahley, R. W., Hui, D. Y., Innerarity, T. L. and Weisgraber, K. H. (1981). Two Independent Lipoprotein Receptors on Hepatic Membranes of Dog, Swine, and Man. Journal of Clinical Investigation, 68: 1197-1206.
Mahley, R. W. and Hussain, M. M. (1991). Chylomicron and chylomicron remnant catabolism. Current Opinion in Lipidology, 2: 170-176.
Mahley, R. W. and Innerarity, T. L. (1983). Lipoprotein Receptors and Cholesterol Homeostasis. Biochimica et Biophysica Acta, 737: 197-222.
Mahley, R. W., Innerarity, T. L., Rail, S. C. and Weisgraber, K. H. (1984). Plasma lipoproteins: apolipoprotein structure and function. Journal of Lipid Research, 25: 1277-1294.
Mahley, R. W. and Ji, Z.-S. (1999). Remnant lipoprotein metabolism: key pathways involving cell-surface heparan sulfate proteoglycans and apolipoprotein E. Journal of Lipid Research, 40 (1): 1-16.
Mahley, R. W., Ji, Z.-S., Brecht, W. J., Miranda, R. D. and He, D. (1994). Role of heparan sulfate proteoglycan and the low-density-lipoprotein receptor-related protein in remnant lipoprotein
metabolism. Ann. NY. Acad. Sci., 737: 39-52.
Mahley, R. W. and Rail, S. C. (1989). In Type III hyperlipoproteinaemia (dysbetalipoproteinaemia): the role of apolipoprotein E in normal and abnormal lipoprotein metabolism. The Metabolic Basis of Inherited Disease, (Eds, Scriver, C. R., Deaudet, A. L., Sly, W . S. and Valle, D.) McGraw-Hill, N e w
York, pp. 1195-1213.
Mamo, J. C, Proctor, S. D. and Smith, D. (1998b). Retention of chylomicron remnants by arterial tissue; importance of an efficient clearance mechanism from plasma. Atherosclerosis, 141 (1): S63-
S69.
Mamo, J. C, Smith, D., Yu, K. C, Kawaguchi, A., Harada-Shiba, M., Yamamura, T. and Yamamoto, A. (1998a). Accumulation of chylomicron remnants in homozygous subjects with familial hypercholesterolaemia. European Journal of Clinical Investigation, 28 (5): 379-384.
Mamo, J. C. L. (1995). Atherosclerosis as a Post-prandial Disease. Endocrinology and Metabolism, 2:
229-244.
Mamo, J. C. L., Bowler, A., Elsegood, C. L. and Redgrave, T. G. (1991). Defective plasma clearance of chylomicron-like lipid emulsion in Watanabe heritable hyperlipidemic rabbits. Biochimica et
Biophysica Acta, 1081: 241-245.

346
Ma m o , J. C. L., Bowler, A., Redgrave, T. G. and Elsegood, C. L. (1994). Clearance of chylomicron-like lipid emulsions is increased in normal rabbits but not in heterozygous Watanabe Heritable Hyperlipidaemic rabbits following treatment with Cholestyramine or Pravastatin. Clinical and Experimental Pharmacology and Physiology, 21: 687-694.
Mamo, J. C. L., Elsegood, C. L., Gennat, H. C. and Yu, K. (1996). Degradation of Chylomicron Remnants by Macrophages Occurs via Phagocytosis. Biochemistry, 35: 10210-10214.
Mamo, J. C. L, Elsegood, C. L., Umeda, Y., Hirano, T. and Redgrave, T. G. (1993). Effect of Probucol on Plasma Clearance and Organ Uptake of Chylomicrons and VLDLs in Normal and Diabetic Rats. Arteriosclerosis and Thrombosis, 13: 231-239.
Mamo, J. C. L. and Wheeler, J. R. (1994). Chylomicrons or their remnants penetrate rabbit thoracic aorta as efficiently as do smaller macromolecules, including low-density lipoprotein, high-density lipoprotein, and albumin. Coronary Artery Disease, 5: 695-705.
Mamo, J. C. L., Yu, K. C. W., Elsegood, C. L., Smith, D., Vine, D., Gennat, H. C, Voevodin, M. and Proctor, S. D. (1997). Is atherosclerosis exclusively a postprandial phenomenon? Clinical and Experimental Pharmacology and Physiology, 24: 288-293.
Mann, C. J., Khallou, J., Chevreuil, O., Troussard, A. and Guermani, I. M. (1995). Mechanisms of activation and functional significance of the lipolysis-stimulated receptor: evidence for a role as chylomicron remnant receptor. Biochemical Journal, 34: 10421-10431.
Maranhao, R. C, Cesar, T. B., Pedroso-Mariani, S., Hirata, M. H. and Mesquita, C. H. (1993). Metabolic behaviour in rats of a nonprotein microemulsion resembling low density lipoprotein. Lipids, 28: 691-696.
Maranhao, R. C, Feres, M. C, Martins, M. T., Mesquita, C. H., Toffoletto, O, Vinagre, C. G. C, Gianinni, S. D. and Pileggi, F. (1996). Plasma kinetics of a chylomicron-like emulsion in patients with coronary artery disease. Atherosclerosis, 126: 15-25.
Maranhao, R. C, Garicocchea, B., Silva, E. L., Dorlheac-Llacer, P., Cadena, S. M. S., Coelho, I. J. C, Meneghetti, J. C , Pileggi, F. J. C. and Chamone, D. A. F. (1994). Plasma kinetics and biodistribution of a lipid emulsion resembling low density lipoprotein in patients with acute myeloid leukemia. Cancer Research, 54: 4660-4666.
Maranhao, R. C, Garicochea, B., Silva, E. L., Dorlheac-Llacer, P., Pileggi, F. and Chamone, D. A. F. (1992). Increased plasma removal of microemulsions resembling the lipid phase of low density lipoproteins in patients with acute myeloid leukemia: A possible new strategy for the treatment of the
disease. Braz J. Med. Biol. Res., 25: 68-79.
Maranhao, R. C, Roland, I. A., Toffoletto, O., Ramires, J. A., Goncalves, R. P., Mesquita, C. H. and Pileggi, F. (1997). Plasma kinetic behavior in hyperlipidemic subjects of a lipidic microemulsion that binds to low density lipoprotein receptors. Lipids, 32 (6): 627-633.
Maranhao, R. C, Tercyak, A. M. and Redgrave, T. G. (1986). Effects of cholesterol content on the metabolism of protein-free emulsion models of lipoproteins. Biochimica et Biophysica Acta, 875: 247-
255.
Maranhao, R. C, Vinagre, C. G., Arie, S., Guimaraes, J. B., da-Luz, P., Bellotti, G. and Pileggi, F. (1995). Lipoprotein (a) in subjects with or without coronary artery disease: relation to clinical history and risk factors. Brazilian Journal of Medical and Biological Research, 28 (4): 439-446.
Marcel, Y. L. (1982). Lecithin: cholesterol acyltransferase and intravascular cholesterol transport.
Advances in Lipid Research, 19: 85-136.

347
Martin de Llano, J. J., Andreu, E. J. and Knecht, E. (1996). Use of inductively coupled plasma-mass spectrometry for the quantitation of the binding and uptake of colloidal-gold low density lipoprotein conjugates by cultured cells. Analytical Biochemistry, 243: 210-217.
Martin, M. J., Hulley, S. B., Browner, W. S., Kuller, L. H. and Wentworth, D. (1986). Serum cholesterol, blood pressure, and mortality: implications of a cohort of 361 662 men. Lancet, ii: 933-936.
Martinez, M. J., Ruiz, J. I., Lacort, M. and Ochoa, B. (1991). Diurnal variations of rat liver enzymes catalyzing cholesterol ester hydrolysis. Biochimica et Biophysica Acta, 1085 (1): 106-111.
Martins, I., Vilcheze, C, Mortimer, B.-C, Bittman, R. and Redgrave, T. G. (1998). Sterol side chain length and structure affect the clearance of chylomicron-like lipid emulsions in rats and mice. Journal of Lipid Research, 39: 302-312.
Martins, I. J., Hone, E., Chi, C, Seydel, U., Martins, R. N. and Redgrave, T. G. (2000b). Relative roles of LDLr and L R P in the metabolism of chylomicron remnants in genetically manipulated mice. Journal of Lipid Research, 41 (2): 205-213.
Martins, I. J., Hopkins, L., Joll, C. A. and Redgrave, T. G. (1991). Interactions between model triacylglycerol-rich lipoproteins and high-density lipoproteins in rat, rabbit and man. Biochimica et Biophysica Acta, 1081: 328-338.
Martins, I. J., Lenzo, N. P. and Redgrave, T. G. (1989). Phosphatidylcholine metabolism after transfer from lipid emulsions injected intravenously in rats. Implications for high-density lipoprotein metabolism. Biochimica et Biophysica Acta, 1005: 217-224.
Martins, I. J., Mortimer, B.-C, Miller, J. and Redgrave, T. G. (1996). Effects of particle size and number on the plasma clearance of chylomicrons and remnants. Journal of Lipid Research, 37 (12): 2696-2705.
Martins, I. J., Mortimer, B.-C. and Redgrave, T. G. (1997). Effect of the ACAT Inhibitor CL 227,082 on Apolipoprotein B48 Transport in Mesenteric Lymph and on Plasma Clearance of Chylomicrons and Remnants. Arteriosclerosis Thrombosis and Vascular Biology, 17 (1): 211-216.
Martins, I. J. and Redgrave, T. G. (1998). A 13C02 breath test to assess the metabolism of triglyceride-rich lipoprotein remnants in mice. Journal of Lipid Research, 39 (3): 691-698.
Martins, I. J., Sainsbury, A. J., Mamo, J. C L. and Redgrave, T. G. (1994). Lipid and apolipoprotein B48 transport in mesenteric lymph and the effect of hyperphagia on the clearance of chylomicron-like emulsions in insulin-deficient rats. Diabetologia, 37: 238-246.
Martins, I. J., Vermeulen, R. and Redgrave, T. G. (2000a). Relative roles of mitochondrial and perixisomal fatty acid oxidation in the metabolism of chylomicron remnants in rats and mice as assessed by a stable-isotope breath test. Atherosclerosis, 150: 13-20.
Martins, M. C T., Pileggi, F. and Maranhao, R. C. (1995). Clearance of a chylomicron-like emulsion from plasma is delayed in patients with coronary artery disease. Brazilian Journal of Medical and
Biological Research, 28: 427-431.
McGee, D. L., Reed, D. M., Yano, K, Kagan, A. and Tillotson, J. (1984). Ten year incidence of coronary heart disease in the Honolulu Heart Program. American Journal of Epidemiology, 119: 733-
741.
McLean, R. and Phillips, M. C (1981). Mechanisms of cholesterol and phophatidylcholine exchange or transfer between unilamellar vesicles. Biochemistry, 20: 2893-2900.
McNamara, J. R., Campos, H., Ordovas, J. M., Peterson, J., Wilson, P. W. and Schaefer, E. J. (1987). Effect of gender, age, and lipid status on low density lipoprotein subtraction distribution. Results from the Framingham Offspring Study. Arteriosclerosis, 1 (5): 483-490.

348
McNeish, J., Aiello, R. J., Guyot, D., Turi, T., Gabel, C , Aldinger, C , Hoppe, K. L., Roach, M . L., Royer, L. J., de Wet, J., Broccardo, C , Chimini, G. and Francone, O. L. (2000). High density lipoprotein deficiency and foam cell accumulation in mice with targeted disruption of ATP-binding cassette transporter-1. Proc. Natl. Acad. Sci. U.S.A, 91 (8): 4245-4250.
Melchoir, G. W., Mahley, R. W. and Buckhold, D. K. (1981). Chylomicron metabolism during dietary-induced hypercholesterolemia in dogs. Journal of Lipid Research, 22: 598-609.
Menotti, A., Keys, A., Kromhout, D., Aravanis, C, Bloemberg, B., Buzina, R., Dontas, A. S., Fidanza, F. and Giampaoli, S. (1993). Inter-cohort differences in coronary heart disease mortality in the 25 year follow up of the seven countries study. European Journal of Epidemiology, 9 (5): 527-536.
Meyer, E., Tineke Westerveld, H., de Ruyter-Meijstek, F. C, van Greevenbroek, M. M. J., Rienks, R, van Rijn, H. J. M., Willem Erkelens, D. and de Bruin, T. W . A. (1996). Abnormal postprandial apolipoprotein B-48 and triglyceride responses in normolipidemic women with greater than 7 0 % stenotic coronary artery disease: a case-control study. Atherosclerosis, 124: 221-235.
Miettinen, T. A. and Kasaiemei, Y. A. (1986). In Cholesterol balance and lipoprotein metabolism in man. Bile Acids and Atherosclerosis, Vol. 15. (Ed, Grundy, S. M.) Raven Press, N e w York, pp. 113-155.
Miller, K. W. and Small, D. M. (1982). The Phase Behaviour of Triolein, Cholesterol, and Lecithin Emulsions. Journal of Colloid and Interface Science, 89 (2): 466-478.
Miller, K. W. and Small, D. M. (1983a). Triolein-Cholesterol Oleate-Cholesterol-Lecithin Emulsions: Structural Models of Triglyceride-Rich Lipoproteins. Biochemistry, 22: 443-451.
Miller, K. W. and Small, D. M. (1983b). Surface-to-Core and Interparticle Equilibrium Distributions of Triglyceride-rich Lipoprotein Lipids. The Journal of Biological Chemistry, 258: 13772-13784.
Miller, K. W. and Small, D. M. (1987). In Structure of triglyceride-rich lipoproteins: An analysis of core and surface phases. Plasma Lipoproteins: New Comprehensive Biochemistry, (Ed, Gotto, A. M.) Elsevier, Amsterdam.
Mims, M. P., Soma, M. R. and Morrisett, J. D. (1990). Effect of particle size and temperature on the conformation and physiological behaviour of apo E bound to model lipoprotein particles. Biochemistry,
29:6639-6647.
Minari, O. and Zilversmit, D. B. (1963). Behaviour of dog lymph chylomicron lipid constituents during incubation with serum. Journal of Lipid Research, 4: 424-436.
Mishkel, M. A. and Crowther, S. M. (1977). Hypothyroidism, an important cause of reversible hyperlipidemia. Clinica ChimicaActa, 74 (2): 139-151.
Mjos, O. D., Faegerman, O., Hamilton, R. L. and Havel, R. J. (1975). Characterisation of remnants produced during the metabolism of triglyceride-rich lipoproteins of blood plasma and intestinal lymph
in the rat. Journal of Clinical Investigation, 56: 603-615.
Mokuno, H., Brady, S., Kotite, L., Herz, J. and Havel, R. J. (1994). Effect of the 39 kDa receptor-associated protein on the hepatic uptake and endocytosis of chylomicron remnants and low density lipoproteins in the rat. The Journal of Biological Chemistry, 269 (18): 13238-13243.
Mora, R., Simionescu, M. and Simionescu, N. (1990). Purification and partial characterization of extracellular liposomes isolated from the hyperlipidemic rabbit aorta. Journal of Lipid Research, 31
(10): 1793-1807.
Mortimer, B.-C, Beveridge, D. J., Martins, I. J. and Redgrave, T. G. (1995a). Intracellular Localization and Metabolism of Chylomicron Remnants in the Livers of Low Density Lipoprotein Receptor-deficient Mice and ApoE-deficient Mice. The Journal of Biological Chemistry, 270 (48): 28767-28776.

349
Mortimer, B.-C, Holthouse, D. J., Martins, I. J., Stick, R. V. and Redgrave, T. G. (1994a). Effects of triacylglycerol-saturated acyl chains on the clearance of chylomicron-like emulsions from the plasma of the rat. Biochimica et Biophysica Acta, 1211: 171-180.
Mortimer, B.-C, Martins, I., Zeng, B. J. and Redgrave, T. G. (1997). Use of Gene-Manipulated Models to Study the Physiology of Lipid Transport. Clinical and Experimental Pharmacology and Physiology, 24: 281-285.
Mortimer, B. C, Redgrave, T. G, Spangler, E. A., Verstuyft, J. G. and Rubin, E. M. (1994b). Effect of human apoE4 on the clearance of chylomicron-like lipid emulsions and atherogenesis in transgenic mice. Arteriosclerosis & Thrombosis, 14 (10): 1542-1552.
Mortimer, B.-C, Simmonds, W. J., Cockman, S. J., Stick, R. V. and Redgrave, T. G. (1990). The effect of monostearoylglycerol on the metabolism of chylomicron-like lipid emulsions injected intravenously in rats. Biochimica et Biophysica Acta, 1046: 46-56.
Mortimer, B.-C, Tso, P., Phan, C T., Beveridge, D. J., Wen, J. a. and Redgrave, T. G. (1995b). Features of cholesterol structure that regulate the clearance of chylomicron-like lipid emulsions. Journal of Lipid Research, 36: 2038-2053.
Mouzeyan, A., Choi, J., Allayee, H., Wang, X., Sinsheimer, J., Phan, J., Castellani, L. W., Reue, K, Lusis, A. J. and Davis, R. C. (2000). A locus conferring resistance to diet-induced hypercholesterolemia and atherosclerosis on mouse chromosome. Journal of Lipid Research, 41 (4): 573-582.
Myers, G. L., Cooper, G. R. and Sampson, E. J. (1994). Traditional lipoprotein profile: clinical utility, performance requirements, and standardization. Atherosclerosis, 108 (Suppl.): S157-S169.
Myhre, A. M., Hagen, E., Blomhoff, R. and Norum, K. R. (1998). Retinoylation of proteins in a macrophage tumor cell line J774, following uptake of chylomicron remnant retinyl ester. Journal of Nutritional Biochemistry, 9(12): 705-711.
Nagasaki, A., Kikuchi, T., Kurata, K., Masushige, S., Hasegawa, T. and Kato, S. (1994). Vitamin A regulates the expression of apolipoprotein Al and CIII genes in the rat. Biochemical and Biophysical Research Communications, 205 (3): 1510-1517.
Nagata, Y., Chen, J. and Cooper, A. D. (1988). Role of Low Density Lipoprotein Receptor-dependent and -independent Sites in Binding and Uptake of Chylomicron Remnants in Rat Liver. The Journal of Biological Chemistry, 263: 15151-15158.
Nakajima, K., Okazaki, M., Tanaka, A., Pullinger, C. R., Wang, T., Nakano, T., Adachi, M. and Havel, R. J. (1996). Separation and Determination of Remnant-like Particles in Human Serum Using Monoclonal Antibodies to Apo B-100 and Apo A-l. Journal of Clinical Ligand Assay, 19 (3): 177-183.
Nakajima, K, Saito, T., Tamwa, A., Suzuki, M., Nakano, T., Adachi, M., Tanaka, A., Tada, N„ Nakamura, H. and Campos, E. (1993). Cholesterol in remnant-like lipoproteins in human serum using monoclonal anti-apo B-100 and anti-apo A-l immunoaffinity mixed gels. Clinica Chimica Acta, 223
(1-2): 53-71.
Nakandakare, E. R., Lottenberg, S. A., Oliveira, H. C. F., Bertolami, M. C, Vasconcelos, K. S., Sperotto, G. and Quintao, E. C R. (1994). Simultaneous measurements of chylomicron lipolysis and remnant removal using a doubly labeled artificial lipid emulsion: studies in normolipidemic and
hyperlipidemic subjects. Journal of Lipid Research, 35: 143-152.
Nakashima, Y., Plump, A. S., Raines, E. W., Breslow, J. L. and Ross, R. (1994). ApoE-deficient mice develop lesions of all phases of atherosclerosis throughout the arterial tree. Arteriosclerosis &
Thrombosis, 14 (1): 133-40.

350
Nakazato, K., Ishibashi, T., Shindo, J., Shiomi, M . and Maruyama, Y. (1996). Expression of very low density lipoprotein receptor m R N A in rabbit atherosclerotic lesions. American Journal of Pathology, 149(6): 1831-1838.
Nestel, P. J., Billington, T. and Fidge, N. H. (1983). Slower removal of intestinal apolipoprotein B-48 than of apolipoprotein B-100 in severely hypertriglyceridemic subjects. Biochimica et Biophysica Acta, 751:422-427.
Nestel, P. J., Havel, R. J. and Bezman, A. (1963a). Metabolism of constituent perfused rat liver. Biochimica et Biophysica Acta, 488: 464-474.
Nestel, P. J., Havel, R. J. and Bezman, A. (1963b). Metabolism of constituent lipids of dog chylomicrons. Journal of Clinical Investigation, 42: 1313-1321.
Nestruck, A. C and Rubinstein, D. (1976). The synthesis of apoproteins of very low density lipoproteins isolated from the Golgi apparatus of rat liver. Canadian Journal of Biochemistry, 54 (7): 617-628.
Newman, T. C, Dawson, P. A., Rudel, L. L. and Williams, D. L. (1985). Quantitation of apolipoprotein E m R N A in the liver and peripheral tissues of nonhuman primates. The Journal of Biological Chemistry, 260 (4): 2452-2457.
Niemeier, A., Gafvels, M., Heeren, J., Meyer, N., Angelin, B. and Beisegel, U. (1996). VLDL receptor mediates the uptake of human chylomicron remnants in vitro. Journal of Lipid Research, 37: 1733-1742.
Nilsson, A. and Zilversmit, D. B. (1971). Distribution of chylomicron cholesterol ester between parenchymal and Kupffer cells of rat liver. Biochimica et Biophysica Acta, 248: 137-142.
Noel, S.-P., Dolphin, P. J. and Rubinstein, D. (1975). An In Vitro Model for the Catabolism of Rat Chylomicrons. Biochemical and Biophysical Research Communications, 63 (3): 764-772.
Nordestgaard, B. G. and Tybaerg-Hansen, A. (1992). IDL, VLDL, Chylomicrons and Atherosclerosis. European Journal of Epidemiology, 8 (Suppl.): 92-98.
Nordestgaard, B. G., Wootton, R. and Lewis, B. (1995). Selective retention of VLDL, IDL and LDL in the arterial intima of genetically hyperlipidemic rabbits in vivo. Molecular size as a determinant of fractional loss from the intima-inner media. Arteriosclerosis Thrombosis and Vascular Biology, 15:
534-542.
Nordoy, A. and Goodnight, S. H. (1990). Dietary Lipids and Thrombosis. Relationships to Atherosclerosis. Arteriosclerosis, 10: 149-163.
Norum, K. R. and Blomhoff, R. (1992). McCollum Award Lecture: Vitamin A absorption, transport, cellular uptake, and storage. American Journal of Clinical Nutrition, 56: 735-744.
O'Brien, K. D., Deeb, S. S., Ferguson, M., McDonald, T. O., Allen, M. D., Alpers, C. E. and Chait, A. (1994). Apolipoprotein E localisation in human coronary atherosclerotic plaques by in situ hybridisation and immunocytochemistry and comparison with lipoprotein lipase. American Journal of
Pathology, 144: 538-548.
Ochoa, B., Gee, A., Jackson, B. and Suckling, K. E. (1990). Regulation of cholesteryl ester metabolism in the hamster liver. Biochimica et Biophysica Acta, 1044 (1): 133-138.
Oliveira, H. C. F., Hirata, M. H., Redgrave, T. G. and Maranhao, R. C. (1988). Competition between chylomicrons and their remnants for plasma removal: a study with artificial emulsion models of chylomicrons. Biochimica et Biophysica Acta, 958: 211-217.

351
Olofsson, S.-O., Bjursell, G., Bostrom, K., Carlsson, P., Elovson, J., Protter, A. A., Rueben, M. A. and Bondjers, G. (1987). Apolipoprotein B: structure, biosynthesis and role in the lipoprotein assembly process. Atherosclerosis, 68: 1-17.
Olson, J. A. (1961). The conversion of radioactive B-carotene into Vitamin A by rat intestine in vivo. The The Journal of Biological Chemistry, 236 (2): 349-356.
Olson, J. A. (1969). Metabolism and function of vitamin A. Federation Proceedings, 28 (5): 1670-1677.
OMeara, N. M., Lewis, G. F., Cabana, V. G., Iverius, P. H., Getz, G. S. and Polonsky, K. S. (1992). Role of Basal Triglyceride and High Density Lipoprotein in Determination of Postprandial Lipid and Lipoprotein Responses. Journal of Clinical Endocrinology and Metabolism, 75: 465-471.
Oram, J. F., Lawn, R. M., Garvin, M. F. and Wade, D. P. (2000). ABCA1 is the cAMP-inducible apolipoprotein receptor that mediates cholesterol secretion from macrophages. The Journal of Biological Chemistry, 275 (44): 34508-34511.
Oram, J. F. and Yokoyama, S. (1996). Apolipoprotein-mediated removal of cellular cholesterol and phospholipids. Journal of Lipid Research, 37 (12): 2473-2491.
Organisation., W. H. (1988). World Health Statistics Annual. In Secondary World Health Statistics Annual, .Volume (Number), pp. 400-407.
Oschry, Y. and Eisenberg, S. (1982). Rat plasma lipoproteins: re-evaluation of a lipoprotein system in an animal devoid of cholesteryl ester transfer activity. Journal of Lipid Research, 23 (8): 1099-1106.
Osuga, J., Yonemoto, M., Yamada, N., Shimano, H., Yagyu, H., Ohashi, K., Harada, K, Kamei, T., Yazaki, Y. and Ishibashi, S. (1998). Cholesterol lowering in low density lipoprotein receptor knockout mice overexpressing apolipoprotein E. Journal of Clinical Investigation, 102 (2): 386-394.
Packard, C. J., Shepherd, J., Lindsay, G. M., Gaw, A. and Taskinen, M. R. (1993). Thyroid replacement therapy and its influence on postheparin plasma lipases and apolipoprotein-B metabolism in hypothyroidism. Journal of Clinical Endocrinology & Metabolism, 76 (5): 1209-16.
Paigen, B., Ishida, B. Y., Verstuyft, J. G, Winters, R. B. and Albee, D. (1990). Atherosclerosis susceptibility differences among progenitors of recombinant inbred strains of mice. Arteriosclerosis,
10: 316-323.
Pal, S., Elsegood, C L., Proctor, S. D. and Mamo, J. C. (2000). Detection of LDL receptor by ligand blotting with chylomicron remnants labelled with colloidal gold. Annals of Clinical Biochemistry, 37
fl>t4): 471-8.
National Cholesterol Education Program. Adult Treatment Panel., (1988). Report of the Expert Panel on Detection, Evaluation and Treatment of Blood Cholesterol in Adults. Archives of Internal Medicine,
148: 36.
Patsch, J. R. (1987). In Postprandial lipaemia. Bailliere's Clinical Endocrinology and Metabolism. International Practice and Research, Vol. 1. (Ed, Shepherd, J.) Bailliere Tindall,, pp. 551-580.
Patsch, J. R., Miesenbock, G., Hopferwieser, T., Muhlberger, V., Knapp, E., Dunn, J. K, Gotto, A. M. and Patsch, W . (1992). Relation of Triglyceride Metabolism and Coronary Artery Disease. Studies in
the Postprandial State. Arteriosclerosis and Thrombosis, 12: 1336-1345.
Patsch, J. R., Prasad, S., Gotto, A. M. and Bengtsson-Olivecrona, G. (1984). Postprandial lipemia: A key for the conversion of H D L 2 into H D L 3 by hepatic lipase. Journal of Clinical Investigation, 74:
2017-2023.

352
Peel, A. S., Zampelas, A., Williams, C M . and Gould, B. J. (1993). A novel antiserum specific to apolipoprotein B-48 in the investigation of postprandial lipaemia in humans. Clinical Science, 85: 521-524.
Phillips, M. L., Pullinger, C, Kroes, I., Kroes, J., Hardman, D. A., Chen, G., Curtiss, L. K., Gutierrez, M . M., Kane, J. P. and Shumaker, V. N. (1997). A single copy of apolipoprotein B-48 is present on the human chylomicron remnant. Journal of Lipid Research, 38: 1170-1177.
Phillips, M. L. and Schumaker, V. N. (1989). Conformation of apolipoprotein B after lipid extraction of low density lipoproteins attached to an electron microscope grid. Journal of Lipid Research, 30: 415-422.
Piedrahita, J. A., Zhang, S. H., Hagaman, J. R., Oliver, P. M. and Maeda, N. (1992). Generation of mice carrying a mutant apolipoprotein E gene inactivated by gene targeting in embryonic stem cells. Proc. Natl. Acad. Sci. U.S.A., 89: 4471-4475.
Pitas, R. E., Innerarity, T. L., Arnold, K. S. and Mahley, R. W. (1979). Rate and equilibrium constants for binding of apo-E H D L c (a cholesterol-induced lipoprotein) and low density lipoproteins to human fibroblasts: Evidence for multiple receptor binding of apo-E HDLc. Proc. Natl. Acad. Sci. U.S.A., 76: 2311-2315.
Pitman, R. C, Carew, T. E., Attie, A. D., Witzum, J. L., Watanabe, Y. and Steinberg, D. (1982). Receptor-dependent and receptor-independent degradation of low density lipoprotein in normal rabbits and in receptor-deficient mutant rabbits. The Journal of Biological Chemistry, 257: 7994.
Plump, A. S., Smith, J. D., Hayek, T., Aalto-Setala, K., Walsh, A., Verstuyft, J. G., Rubin, E. M. and Breslow, J. L. (1992). Severe Hypercholesterolemia and Atherosclerosis in Apolipoprotein E-Deficient Mice Created by Homologous Recombination in ES Cells. Cell, 71 (2): 343-353.
Potts, J. L, Fisher, R. M., Humphreys, S. M., Coppack, S. W., Gibbons, G. F. and Frayn, K. N. (1991). Chylomicrons are the preferred substrate for lipoprotein lipase in vivo. Biochemical Society
Transactions, 19 (3): 314S.
Powell, L. M., Wallis, S. C, Pease, R. J., Edwards, Y. H., Knott, T. J. and Scott, J. (1987). A novel form of tissue-specific R N A processing produces apolipoprotein B48 in intestine. Cell, 50: 831-840.
Poznansky, M. J. and Czekanski, S. (1979). Cholesterol exchange as a function of cholesterol/phospholipid mole ratios. Biochemistry Journal, 177: 989-991.
Presti, F. T., Pace, R. J. and Chan, S. I. (1982). Cholesterol-phospholipid interaction in membranes. 2. Stoichiometry and molecular packing of cholesterol-rich domains. Biochemistry, 21 (16): 3831-3835.
Proctor, S. D. and Mamo, J. C L. (1996). Arterial fatty lesions have increased uptake of chylomicron remnant but not low-density lipoproteins. Coronary Artery Disease, 1 (3): 239-245.
Proctor, S. D. and Mamo, J. C. L. (1998). Retention of fluorescent-labelled chylomicron remnants within the intima of the arterial wall. Evidence that plaque cholesterol may be derived from postprandial lipoproteins. European Journal of Clinical Investigation, 28 (6): 497-503.
Pullinger, C. R., North, J. D., Teng, B. B., Rifici, V. A., Ronhild de Brito, A. E. and Scott, J. (1989). The apolipoprotein B gene is constitutively expressed in HepG2 cells: Regulation of secretion by oleic acid, albumin, and insulin, and measurement of the m R N A half-life. Journal of Lipid Research, 30 (7):
1065-1077.
Qui, S. Q., Bergeron, N., Kotite, L., Krauss, R. M., Bensadoun, A. and Havel, R. J. (1998). Metabolism of lipoproteins containing apolipoprotein B in hepatic lipase-deficient mice. Journal of Lipid Research,
39 (8): 1661-1668.
Quinn, D., Shirai, K. and Jackson, R. L. (1982). Lipoprotein lipase: mechanism of action and role in
lipoprotein metabolism. Prog. Lipid. Res., 22: 35-78.

353
Rajaram, O. V. and Sawyer, W . H. (1996). Penetration of an emulsion surface by cholesteryl ester
transfer protein. European Biophysics Journal, 25: 31036.
Rail, S. C J. and Mahley, R. W. (1992). The role of apolipoprotein E genetic variants in lipoprotein
disorders. Journal of Internal Medicine, 231 (6): 653-659.
Rapp, J. H., Lespine, A., Hamilton, R. L., Colyvas, N., Chaumeton, A. H., Tweedie-Hardman, J., Kotite, L., Kunitake, S. T., Havel, R. J. and Kane, J. P. (1994). Triglyceride-rich lipoproteins isolated by selected affinity anti-apolipoproteinB immunosorption from human atherosclerotic plaque.
Arteriosclerosis and Thrombosis, 14(11): 1767-1774.
Rassin, T., Liron, M., Rubinstein, A., Arad, J. and Weintraub, M. (1992). Vitamin A Loading - An Indicator of Post-Prandial Lipoprotein Clearance in Healthy and Hypertriglyceridemic Subjects. Isr. J.
Med. Sci., 28: 706-710.
Reddick, R. L., Zhang, S. H. and Maeda, B. (1994). Athersoclerosis in Mice Lacking Apo E. Evaluation of Lesional Development and Progression. Arteriosclerosis and Thrombosis, 14: 141-147.
Redgrave, T. G. (1970). Formation of Cholesteryl Ester-Rich Particulate Lipid during Metabolism of Chylomicrons. Journal of Clinical Investigation, 49: 465-471.
Redgrave, T. G. (1983). In Formation and Metabolism of Chylomicrons. Gastrointestinal Physiology TV. International Review of Physiology, Vol. 28. (Ed, Young, J. A.) University Park Press, Baltimore,
pp. 103-130.
Redgrave, T. G. (1988). A New Approach to the Physiology of Lipid Transport. NIPS, 3: 10-13.
Redgrave, T. G. (1999). In Chylomicrons. Lipoproteins in Health and Disease, Current Book Contents
in Current Contents,, pp. 31-54.
Redgrave, T. G. and Callow, M. J. (1990). The Effect of Insulin Deficiency on the Metabolism of Lipid Emulsion Models of Triaclyglycerol-Rich Lipoproteins in Rats. Metabolism, 39 (1): 1-10.
Redgrave, T. G. and Dunne, K. B. (1975). Chylomicron formation and composition in unanaesthetised
rabbits. Atherosclerosis, 22: 389-400.
Redgrave, T. G., Dunne, K. B., Roberts, D. C. K. and West, C E. (1976). Chylomicron metabolism in rabbits fed diets with or without added cholesterol. Atherosclerosis, 24: 501-508.
Redgrave, T. G., Elsegood, C L., Mamo, J. C L. and Callow, M. J. (1991). Effects of hypothyroidism on the metabolism of lipid emulsion models of triacylglycerol-rich lipoproteins in rats. Biochemical
Journal, 273: 375-381.
Redgrave, T. G., Fidge, N. H. and Yin, J. (1982). Specific, saturable binding and uptake of rat chylomicron remnants by rat skin fibroblasts. Journal of Lipid Research, 23: 638-644.
Redgrave, T. G., Ly, G. L, Quintao, E. C. R., Ramberg, C. F. and Boston, R. C (1993). Clearance from plasma of triacylglycerol and cholesteryl ester after intravenous injection of chylomicron-like lipid emulsions in rats and man. Biochemical Journal, 290: 843-847.
Redgrave, T. G., Mamo, J. C L., Bowler, A. and Mackintosh, V. S. (1992-a). In Factors Regulating Hepatic Uptake of Remnant Lipoproteins. Hepatic Endocytosis of Lipids and Proteins, O^ds, Windier,
E. and Greten, H.) W . Zuckschwerdt Verlag, Munich, pp. 320-325.
Redgrave, T. G. and Maranhao, R. C (1985). Metabolism of protein-free lipid emulsion models of chylomicron in rats. Biochimica et Biophysica Acta, 835: 104-112.
Redgrave, T. G. and Martin, G. (1977). Effects of Chronic Ethanol Consumption on the Catabolism of Chylomicron Triacylglycerol and Cholesteryl Ester in the Rat. Atherosclerosis, 28: 69-80.

354
Redgrave, T. G., Martins, I. J. and Mortimer, B.-C. (1995). Measurement of expired carbon dioxide to assess the metabolism of remnant lipoproteins. Journal of Lipid Research, 36: 2670-2675.
Redgrave, T. G., Mortimer, B.-C. and Martins, I. J. (1996). Receptor-ligand interactions in the hepatic uptake of chylomicron remnants studied with genetically manipulated mice. Z. Gastroenterol, 34 (Suppl. 3): 105-107.
Redgrave, T. G., Rakic, V., Mortimer, B.-C. and Mamo, J. C L. (1992-b). Effects of sphingomyelin and phosphatidylcholine acyl chains in the clearance of triacylglycerol-rich lipoproteins from plasma. Studies with lipid emulsions in rats. Biochimica et Biophysica Acta, 1126: 65-72.
Redgrave, T. G., Roberts, D. C. K. and West, C E. (1975). Separation of Plasma Lipoproteins by Density-Gradient Ultracentrifugation. Analytical Biochemistry, 65: 42-49.
Redgrave, T. G. and Small, D. M. (1979). Quantitation of the Transfer of Surface Phospholipid of Chylomicrons to the High Density Lipoprotein Fraction during the Catabolism of Chylomicrons in the Rat. Journal of Clinical Investigation, 64: 103-130.
Redgrave, T. G. and Vakakis, N. (1976). Hepatic vitamin A fat storage cells and the metabolism of chylomicron cholesterol. Aust. J. Exp. Biol. Med. Sci., 54 (6): 519-525.
Redgrave, T. G., Vassiliou, G. G. and Callow, M. J. (1987). Cholesterol is necessary for triacylglycerol-phospholipid emulsions to mimic the metabolism of lipoproteins. Biochimica et Biophysica Acta, 921: 154-157.
Redgrave, T. G., Watts, G. R, Martins, I. J., Barrett, P. H. R., Mamo, J. C L., Dimmitt, S. B. and Marais, A. D. (2001). Chylomicron remnant metabolism in familial dyslipidemias studied with a remnant-like emulsion breath test. Journal of Lipid Research, 42: 710-715.
Redgrave, T. G. and Zech, L. A. (1987). A kinetic model of chylomicron core lipid metabolism in rats: the effect of a single meal. Journal of Lipid Research, 28: 473-482.
Rensen, P. C N., Herijgers, N., Netscher, M. H., Meskers, S. C. J., van Eck, M. and van Berkel, T. J. C. (1997). Particle size determines the specificity of apolipoprotein E-containing triglyceride-rich emulsions for the L D L receptor versus hepatic remnant receptor in vivo. Journal of Lipid Research, 38:
1070-1084.
Rensen, P. C. N. and van Berkel, T. J. C. (1996). Apolipoprotein E effectively inhibits lipoprotein lipase-mediated lipolysis of chylomicron-like triglyceride-rich lipid emulsions in vitro and in vivo. The Journal of Biological Chemistry, 271 (25): 14791-14799.
Reus, K. E., Houben, G. R, Stam, M. and Dubois, A. E. (2000). Food additives as a cause of medical symptoms: relationship shown between sulfites and asthma and anaphylaxis; results of a literature review. Nederlands Tijdschrift voor Geneeskunde, 144 (38): 1836-1839.
Reznik, Y., Pousse, P., Herrou, M., Morello, R., Mahoudeau, J., Drosdowsky, M. A. and Fradin, S. (1996). Postprandial Lipoprotein Metabolism in Normotriglyceridemic Non-Insulin-Dependent Diabetic Patients: Influence of Apolipoprotein E Polymorphism. Metabolism, 45 (1): 63-71.
Rhee, S. K, Kayani, A. J., Ciszek, A. and Brenna, J. T. (1997). Desaturation and interconversion of dietary stearic and palmitic acids in human plasma and lipoproteins. American Journal of Clinical
Nutrition, 65 (2): 451-458.
Richey Sharrett, A., Chambless, L. E., Heiss, G, Paton, C. and Patsch, W. (1995). Association of Postprandial Triglyceride and Retinyl Palmitate Responses With Asymptomatic Carotid Artery Atherosclerosis in Middle-aged M e n and Women. Arteriosclerosis Thrombosis and Vascular Biology,
15: 2122-2129.

355
Roach, P. D., Balasubramanium, S., Hirata, F., Abbey, M., Szanto, A., Simons, L. A. and Nestel, P. J. (1993a). The low density lipoprotein receptor and cholesterol synthesis are affected differently by
dietary cholesterol in the rat. Biochimica et Biophysica Acta, 1170: 165-172.
Roach, P. D., Hosking, J., Clifton, P. M., Bais, R., Kusenic, B., Coyle, P., Wight, M. B., Thomas, D. W . and Nestel, P. J. (1993b). The effects of hypercholesterolaemia, simvastatin and dietary fat on the low density lipoprotein receptor of unstimulated mononuclear cells. Atherosclerosis, 103 (2): 245-254.
Roach, P. D., Kerry, N. L., Whiting, M. J. and Nestel, P. J. (1993c). Coordinate changes in the low density lipoprotein receptor activity of liver and mononuclear cells in the rabbit. Atherosclerosis, 101
(2): 157-164.
Roach, P. D., Zollinger, M. and Noel, S. P. (1987). Detection of the low density lipoprotein (LDL) receptor on nitrocellulose paper with colloidal gold-LDL conjugates. Journal of Lipid Research, 28:
1515-1521.
Robenek, H., Harrach, B. and Severs, N. J. (1991). Display of low density lipoprotein receptors is clustered, not dispersed, in fibroblasts and hepatocyte plasma membranes. Arteriosclerosis
Thrombosis, 11 (2): 261-271.
Rodriguez, J. L., Catapano, A., Ghiselli, G. C and Sirtori, C. R. (1976). Very low density lipoproteins in normal and cholesterol-fed rabbits: lipid and lipoprotein composition and metabolism.
Atherosclerosis, 23: 85.
Rohlmann, A., Gotthardt, M., Hammer, R. E. and Herz, J. (1998). Inducible Inactivation of Hepatic L R P Gene by Cre-mediated Recombination Confirms Role of L R P in Clearance of Chylomicron
Remnants. Journal of Clinical Investigation, 101: 689-695.
Rohrer, L., Freeman, M., Kodama, T., Penman, M. and Krieger, M. (1990). Coiled-coil fibrous domains mediate ligand binding by macrophage scavenger receptor type II. Nature, 343 (6258): 570-
572.
Ross, A. C (1981b). Separation of Long-Chain Fatty Acid Esters of Retinol by High-Performance
Liquid Chromatography. Analytical Biochemistry, 115: 324-330.
Ross, A. C, Minick, C. R. and Zilversmit, D. B. (1978). Equal atherosclerosis in rabbits fed cholesterol-free, low-fat diet or cholesterol-supplemented diet. Atherosclerosis, 29: 161.
Ross, A. C. and Zilversmit, D. B. (1977). Chylomicron remnant cholesteryl esters as the major constituent of very low density lipoproteins in plasma of cholesterol-fed rabbits. Journal of Lipid
Research, 18: 169-181.
Ross, C (1990). Separation of Fatty Acid Esters of Retinol by High-Performance Liquid-
Chromatography. Methods in Enzymology, 189: 81-84.
Ross, R. (1981a). George Lyman Duff Memorial Lecture. Atherosclerosis: a problem of the biology of arterial wall cells and their interactions with blood components. Arteriosclerosis, 1 (5): 293-311.
Ross, R. (1986). The pathogenesis of atherosclerosis - An update. New England Journal of Medicine,
314:'488-500.
Ross, R. (1993a). The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature, 362: 801-
809.
Ross, R. (1993b). Atherosclerosis: A Defense Mechanism Gone Awry. Rous-Whipple Award Lecture.
American Journal of Pathology, 143 (4): 987-1002.
Ross, R. and Glomset, J. A. (1976). The pathogenesis of atherosclerosis. New England Journal of
Medicine, 295 (7-8): 369-377,420-425.

356
Ross, R., Masuda, J., Raines, E. W., Gown, A. M., Katsuda, S., Sasahara, M., Maiden, L. T., Masuko, H. and Sato, H. (1990). Localisation of PDGR-B protein in macrophages in all stages of atherosclerosis. Science, 248: 1009-1012.
Rubinsztein, D. C, Cohen, J. C, Berger, G. M., van der Westhuyyzen, D. R., Coetzee, G. A. and Gevers, W . (1990). Chylomicron Remnant Clearance from the Plasma is Normal in Familial Hypercholesterolemic Homozygotes with Defined Recepter Defects. Journal of Clinical Investigation, 86: 1306-1312.
Rudel, L. L., Bond, M. G. and Bullock, B. C. (1985). LDL heterogeneity and atherosclerosis in nonhuman primates. Anal. NY. Acad. Sci., 454: 248-253.
Rudling, M. (1992). Hepatic mRNA levels for the LDL receptor and HMG-CoA reductase show coordinated regulation in vivo. Journal of Lipid Research, 33 (4): 493-501.
Ruotolo, G., Zhang, H., Bentsianov, V. and Le, N.-A. (1992). Protocol for the study of the metabolism of retinyl esters in plasma lipoproteins during postprandial lipemia. Journal of Lipid Research, 33: 1541-1549.
Russell, D. W., Esser, V. and Hobbs, H. H. (1989). Molecular basis of familial hypercholesterolemia. Arteriosclerosis, 9 (Suppl. 1): 18-113.
Sabesin, S. M. and Frase, S. (1977). Electron microscopic studies of the assembly, intracellular transport, and secretion of chylomicrons by rat intestine. Journal of Lipid Research, 18 (4): 496-511.
Saito, H., Minamida, T., Arimoto, I., Handa, T. and Miyajima, K. (1996). Physical states of surface and core lipids in lipid emulsions and apolipoprotein binding to the emulsion surface. The Journal of Biological Chemistry, 271 (26): 15515-15520.
Sakai, J., Hishino, A., Takahashi, S., Miura, Y., Ishii, H., Suzuki, H., Kawarabayasi, Y. and Yamamoto, T. (1994). Structure, chromosome location, and expression of the human very low density lipoprotein receptor gene. The Journal of Biological Chemistry, 269: 2173-2181.
Sakata, K., Miho, N., Shirotani, M., Yoshida, H., Takada, Y. and Takada, A. (1998). Remnant-like particle cholesterol is a major risk factor for myocardial infarction in vasospasticl angina with nearly normal coronary artery. Atherosclerosis, 136: 225-231.
Sakr, S. W., Attia, N., Haourigui, M., Paul, J. L., Soni, T., Vacher, D. and Girard-Globa, A. (1997). Fatty acid composition of an oral load affects chylomicron size in human subjects. British Journal of
Nutrition,!! (1): 19-31.
Saltykow, S. (1908). Die experimentell erzeugten Arterienveranderungen in ihrer Beziehung zu Atherosklerose und verwandten Krankheiten des Menschen. Zentbl. allg. Path. path. Anat, 19 (II):
397-407.
Santamarina-Fojo, S. and Brewer, H. B. (1991). The familial hyperchylomicronemia syndrome. New insights into underlying genetic defects. Journal of the American Medical Association, 265: 904-908.
Schaefer, E. J. (1990). In Hyperlipoproteinemias and other lipoprotein disorders. Principals and practice of endocrinology and metabolism, (Eds, Becker, K. L., Bilezikian, J. P., Bremmer, W . J., Hung, W., Kahn, C R., Loriaux, D. L., Rebar, R. W., Robertson, G. L. and Wartofsky, L.) J.B. Lippencott Company,, pp. 1229-1241.
Schaefer, E. J., Gregg, R. E., Ghiselli, G., Forte, T. M., Ordovas, J. M., Zech, L. A. and Brewer, H. B. (1986). Familial Apolipoprotein E Deficiency. Journal of Clinical Investigation, 78: 1206-1219.
Schaefer, E. J., Wetzel, M. G., Bengtsson, G., Scow, R. O, Brewer, H. B. and Olivecrona, T. (1982). Transfer of human lymph chylomicron constituents to other lipoprotein density fractions during in vitro lipolysis. Journal of Lipid Research, 23: 1259-1273.

357
Schneeman, B. O., Kotite, L., Todd, K. M . and Havel, R. J. (1993). Relationships between the responses of triglyceride-rich lipoproteins in blood plasma containing apolipoproteins B-48 and B-100 to a fat-containing meal in normolipidemic humans. Proc. Natl. Acad. Sci. USA., 90: 2069-2073.
Schneider, W. J. (1989). The low density lipoprotein receptor. Biochimica et Biophysica Acta, 988: 303-317.
Schneider, W. J., Beisiegel, U., Goldstein, J. L. and Brown, M. S. (1982). Purification of the Low Density Lipoprotein Receptor, an Acidic Glycoprotein of 164,000 Molecular Weight. The Journal of Biological Chemistry, 257: 2664-2673.
Schneider, W. J., Kovanen, P. T., Brown, M. S., Goldstein, J. L., Utermann, G., Weber, W., Havel, R. J., Kotite, L., Kane, J. P., Innerarity, T. L. and Mahley, R. W . (1981). Familial Dysbetalipoproteinemia. Abnormal binding of mutant apoprotein E to low density lipoprotein receptors of human fibroblasts and membranes from liver and adrenal of rats, rabbits, and cows. Journal of Clinical Investigation, 68: 1075-1085.
Schumaker, V. N., Phillips, M. L. and Chatterton, J. E. (1994). Apolipoprotein B and low density lipoprotein structure: Implications for biosynthesis of triglyceride-rich lipoproteins. Adv. Protein. Chem., 45: 205-248.
Scow, R. O. and Blanchette-Mackie, E. J. (1992). Endothelium, the dynamic interface in cardiac lipid transport. Molecular and Cellular Biochemistry, 116(1-2): 181-191.
Scow, R. O, Blanchette-Mackie, E. J. and Smith, L. C. (1976). Role of capillary endothelium in the clearance of chylomicrons. A model for lipid transport from blood by lateral diffusion in cell membranes. Circulation Research, 39: 149.
Scrott, H. G., Goldstein, J. L., Hazzard, W. R., McGoodwin, M. M. and Motulsky, A. G. (1972). Familial hypercholesterolemia in a large kindred: Evidence for a monogenic mechanism. Annals of
Internal Medicine, 67: 711-718.
Segrest, J. P., Garber, D. W., Brouilette, C G., Harvey, S. C and Anantharamaiah, G. M. (1994-a). The amphipathic alpha-helix: A multifunctional structural motif in plasma apolipoproteins. Advances
in Protein Chemistry, 45: 303-369.
Segrest, J. P., Jones, M. K., De Loof, H., Brouillette, C. G., Venkatachalapthi, Y. V. and Anantharamaiah, G. M . (1992). The amphipathic helix in the exchangeable apolipoproteins: A review of secondary structure and function. Journal of Lipid Research, 33 (2): 141-166.
Segrest, J. P., Jones, M. K., Mishra, V. K., Anantharamaiah, G. M. and Garber, D. W. (1994-b). Apo B-100 has a pentapartite structure composed of three amphipathic alpha-helical domains alternating with two amphipathic beta-strand domains. Detection by the computer program L O C A T E . Arteriosclerosis and Thrombosis, 14: 1674-1685.
Serfaty-Lacrosniere, C, Civeira, F., Lanzberg, A., Isaia, P., Berg, J., Janus, E. D., Smith, M. P., Pritchard, P. H., Frohlich, J. and Lees, R. S. (1994). Homozygous Tangier disease and cardiovascular
disease. Atherosclerosis, 107 (1): 85-98.
Shafi, S., Brady, S. E., Bensadoun, A. and Havel, R. J. (1994). Role of hepatic lipase in the uptake and processing of chylomicron remnants in rat liver. Journal of Lipid Research, 35: 709-720.
Shelbourne, F., Hanks, J., Myers, W. and Quarfordt, S. (1980). Effect of proteins on hepatic uptake of triglyceride emulsions in the rat. Journal of Clinical Investigation, 65: 652-658.
Shen B. W., Scanu, A. M. and Kezdy, F. J. (1977). Structure of human serum lipoproteins inferred from compositional analysis. Proc. Natl. Acad. Sci. U.S.A., 74: 837-841.

358
Sherrill, B. C and Dietschy, J. M. (1978). Characterization of the Sinusoidal Transport Process Responsible for Uptake of Chylomicrons by the Liver. The Journal of Biological Chemistry, 253: 1859-1867.
Sherrill, B. C, Innerarity, T. L. and Mahley, R. W. (1980). Rapid Hepatic Clearance of the Canine Lipoproteins Containing Only the E Apoprotein by a High Affinity Receptor. Identity With the Chylomicron Remnant Transport Process. The Journal of Biological Chemistry, 255: 1804-1807.
Shimano, H., Namba, Y., Ohsuga, J., Kawamura, M., Yamamoto, K, Shimada, M., Gotoda, T., Harada, K , Yazaki, Y. and Yamada, N. (1994). Secretion-Recapture Process of Apolipoprotein E in Hepatic Uptake of Chylomicron Remnants in Transgenic Mice. Journal of Clinical Investigation, 93: 2215-2223.
Shimano, H., Yamada, N., Katsuki, M., Shimada, M., Gotoda, T., Harada, K., Murase, T., Fukazawa, C , Takaku, F. and Yazaki, Y. (1992a). Overexpression of apolipoprotein E in transgenic mice: marked reduction in plasma lipoproteins except high density lipoprotein and resistance against diet-induced hypercholesterolemia. Proc. Natl. Acad. Sci. U.S.A., 89: 1745-1750.
Shimano, H., Yamada, N., Katsuki, M., Yamamoto, K, Gotoda, T., Harada, K, Shimada, M. and Yazaki, Y. (1992b). Plasma lipoprotein metabolism in transgenic mice overexpressing apolipoprotein E - accelerated clearance of lipoproteins containing apolipoprotein B. British Journal of Clinical Investigation, 90: 2084-2091.
Simionescu, M. and Simionescu, N. (1991). Cellular interactions of lipoproteins with the vascular endothelium: endocytosis and transcytosis. Targital. Diagn. Ther., 5: 45-95.
Simionescu, N., Vasile, E., Lupu, F., Popescu, P. and Simionescu, M. (1986). Prelesional events in atherogenesis. Accumulation of extracellular cholesterol-rich liposomes in the arterial intima and cardiac valves of the hyperlipidemic rabbit. American Journal of Pathology, 123: 189-195.
Simons, L. A., Dwyer, T., Simons, J., Bernstein, L., Mock, P., Poonia, N. S., Balasubramaniam, S., Baron, D., Branson, J., Morgan, J. and Roy, P. (1987). Chylomicrons and chylomicron remnants in coronary artery disease: a case-control study. Atherosclerosis, 65: 181-189.
Simpson, H. S., Williamson, C M., Olivecrona, T., Pringle, S., Maclean, J., Lorimer, A. R., Bonnefous, R, Bogaievsky, Y., Packard, C J. and Shepherd, J. (1990). Postprandial lipaemia, fenofibrate and coronary heart disease. Atherosclerosis, 85 (2-3): 653-662.
Slyper, A. H. (1992). A fresh look at the atherogenic remnant hypothesis. The Lancet, 340: 289-291.
Small, D. M. (1977). Cellular mechanisms for lipid deposition in atherosclerosis. New England Journal
of Medicine, 297 (16-17): 873-877: 924-929.
Small, D. M. (1986). Handbook of Lipid Research 4. The physical chemistry of lipids. From alkanes to phospholipids. Plenum Press, N e w York and London.
Smith, D., Proctor, S. D. and Mamo, J. C. L. (1997). A highly sensitive assay for quantitation of apolipoprotein B48 using an antibody to human apolipoprotein B and enhanced chemiluminescence. Annals of Clinical Biochemistry, 34: 185-189.
Smith, D., Watts, G. F., Dane-Stewart, C. and Mamo, J. C. (1999). Post-prandial chylomicron response may be predicted by a single measurement of plasma apolipoprotein B48 in the fasting state. European
Journal of Clinical Investigation, 29 (3): 204-209.
Smith, J. E. and Goodman, D. S. (1979). Retinol-binding protein and the regulation of vitamin A transport. Federation Proceedings, 38: 2504-2509.
Smith, J. E., Muto, Y., Milch, P. O. and Goodman, D.-W. S. (1973). The Effects of Chylomicron Vitamin A on the Metabolism of Retinol-binding Protein in the Rat. The Journal of Biological
Chemistry, 248 (5): 1544-1549.

359
Smith, J. L., Roach, P. D., Wittenberg, L. N., Riottot, M., Pillay, S. P., Nestel, P. J. and Nathanson, L. K. (2000). Effects of simvastatin on hepatic cholesterol metabolism, bile lithogenicity and bile acid hydrophobicity in patients with gallstones. Journal of Gastroenterology & Hepatology, 15 (8): 871-879.
Sniderman, A. D., Pedersen, T. and Kjekshus, J. (1997). Putting low-density lipoproteins at center
stage in atherogenesis. American Journal of Cardiology, 79 (1): 64-67.
Soutar, A. K., Myant, N. B. and Thompson, G. R. (1977). Simultaneous measurement of apolipoprotein B turnover in very low and low-density lipoproteins in familial hypercholesterolemia. Atherosclerosis,
28: 147.
Soutar, A. K, Myant, N. B. and Thompson, G. R. (1982). The metabolism of very low density and intermediate density lipoproteins in patients with familial hypercholesterolemia. Atherosclerosis, 43:
217.
Spady, D. K., Turley, S. D. and Dietschy, J. M. (1985). Rates of low density lipoprotein uptake and cholesterol synthesis are regulated independently in the liver. Journal of Lipid Research, 26 (4): 465-
472.
Sprecher, D. L., Hoeg, J. M., Schaefer, E. J., Zech, L. A., Gregg, R. E., Lakatos, E. and Brewer, H. B. J. (1985). The association of L D L receptor activity, L D L cholesterol level, and clinical course in homozygous familial hypercholesterolemia. Metabolism, 34 (3): 294-299.
Sprecher, D. L., Knauer, S. L., Black, D. M., Kaplan, L. A., Akeson, A. A., Dusing, M., Lattier, D., Stein, E. A., Rymaszewski, M . and Wiginton, D. A. (1991). Chylomicron-Retinyl Palmitate Clearance in Type I Hyperlipidemic Families. Journal of Clinical Investigation, 88: 985-994.
Stalenhoef, A. R, Malloy, M. J., Kane, J. P. and Havel, R. J. (1984). Metabolism of apolipoproteins B-48 and B-100 of triglyceride-rich lipoproteins in normal and lipoprotein lipase-deficient humans. Proc
Natl. Acad. Sci. U.S.A., 81 (6): 1839-1843.
Stanton, L. W., White, R. T., Bryant, C. M., Protter, A. A. and Endemann, G. (1992). A macrophage Fc receptor for IgG is also a receptor for oxidised low density lipoprotein. The Journal of Biological
Chemistry, 267 (31): 22446-22451.
Staprans, I., Pan, X.-M., Rapp, J. H. and Feingold, K. R. (1992). Chylomicron and chylomicron remnant metabolism in STZ-induced diabetic rats. Diabetes, 41: 325-333.
Stein, O, Stein, Y., Goodman, D. S. and Fidge, N. H. (1969). The metabolism of chylomicron cholesteryl ester in rat liver. A combined radioautographic-electron microscopic and biochemical
study. Journal of Cell Biology, 43 (3): 410-431.
Steinberg, D. (1976). Interconvertible enzymes in adipose tissue regulated by cyclic AMP-dependent
protein kinase. Advances in Cyclic Nucleotide Research, 1: 157-198.
Steinberg, D. (1988). Metabolism of lipoproteins and their role in the pathogenesis of atherosclerosis.
Atheroscerosis Review, 18: 1-23.
Steinberg, D. (1991). Antioxidants in the Prevention of Human Atherosclerosis. (Summary of the Proceedings of a National Heart, Lung, and Blood Institute Workshop). Circulation, 85: 2337-2343.
Steinberg, D., Parthasarathy, S., Carew, T. E., Khoo, J. C. and Witztum, J. L. (1989). Beyond Cholesterol. Modifications of Low-Density Lipoprotein That Increase Its Atherogenicity. The New England Journal of Medicine, 320: 915-924.
Steiner, G. (1993). Triglyceride-rich Lipoproteins and Atherosclerosis, from Fast to Feast. Annals of
Medicine, 25: 431-435.

360
Steiner, G. (1997). Diabetes and atherosclerosis - a lipoprotein perspective. Diabetic Medicine, 14 (8,
Suppl 3): 44.
Steinmetz, A., Jakobs, C, Motzny, S. and Kaffarnik, H. (1989). Differential distribution of apolipoprotein E isoforms in human plasma lipoproteins. Arteriosclerosis, 9 (3): 405-411.
Stewart-Philips, J. L. and Lough, J. (1991). Pathology of atherosclerosis in cholesterol-fed, susceptible mice. Atherosclerosis, 90: 211-218.
Stokke, K. T. (1972). Subcellular distribution and kinetics of the acid cholesterol esterase in liver. Biochimica et Biophysica Acta, 280 (2): 329-335.
Stone, B. G., Evans, C. D., Fadden, R. J. and Schreiber, D. (1989). Regulation of hepatic cholesterol ester hydrolase and acyl-coenzyme A: cholesterol acyltransferase in the rat. Journal of Lipid Research, 30: 1681-1690.
Stone, N. J., Levy, R. I., Frederickson, D. S. and Verter, J. (1974). Coronary artery disease in 116 kindred with familial type II hyperlipoproteinemia. Circulation, 49: 476-488.
Stow, J. L., Kjellen, L., Unger, E., Hook, M. and Farquhar, M. G. (1985). Heparan sulfate proteoglycans are concentrated on the sinusoidal plasmalemmal domain and in intracellular organelles of hepatocytes. Journal of Cell Biology, 100: 975-980.
Strickland, D. K., Ashcom, J. D., Williams, S., Burgess, W. H., Migliorini, M. and Argraves, W. S. (1990). Sequence identity between the a2-macroglobulin receptor and low density lipoprotein receptor-related protein suggests that this molecule is a multifunctional receptor. The Journal of Biological Chemistry, 265: 17401-17404.
Suckling, K. E. and Jackson, B. (1993). Animal models of human lipid metabolism. Prog. Lipid. Res., 32 (1): 1-24.
Sudhof, T. C, Goldstein, J. L., Brown, M. S. and Russell, D. W. (1985). The LDL Receptor Gene: A Mosaic of Exons Shared with Different Proteins. Science, 228: 815-822.
Sultan, R, Lagrange, D., Jansen, H. and Griglio, S. (1990). Inhibition of hepatic lipase activity impairs chylomicron remnant-removal in rats. Biochimica et Biophysica Acta, 1042: 150-152.
Sultan, F., LaGrange, D., Le Liepvre, X. and Griglio, S. (1989). Chylomicron-remnant uptake by freshly isolated hepatocytes. Effect of heparin and of hepatic triacylglycerol lipase. Biochemical
Journal, 258: 587-594.
Tajima, S., Yokoyama, S. and Yamamoto, A. (1983). Effect of Lipid Particle Size on Association of Apolipoproteins with Lipid. The Journal of Biological Chemistry, 258: 10073-10082.
Takahashi, S., Kawarabayasi, Y., Nakai, T., Sakai, J. and Yamamoto, T. (1992). Rabbit very low density lipoprotein receptor: a low density lipoprotein receptor-like protein with distinct ligand specificity. Proc. Natl. Acad. Sci. U.S.A., 89: 9252-9256.
Tall, A. R. (1993). Plasma cholesteryl ester transfer protein. Journal of Lipid Research, 34: 1255-1274. Tall, A. R., Green, P. H. R., Glickman, R. W . and Riley, J. W . (1979). Metabolic fate of chylomicron phospholipids and apoproteins in the rat. Journal ofClincal Investigation, 64: 977-989.
Tan, M. H., Sata, T. and Havel, R. J. (1977). The significance of lipoprotein lipase in rat skeletal
muscles. Journal of Lipid Research, 18: 363-370.
Tanzawa, K., Shimada, Y., Kuroda, M., Tsujita, Y., Arai, M. and Watanabe, H. (1980). WHHL-rabbit: a low density lipoprotein receptor-deficient animal model for familial hypercholesterolemia. FEBS
Letters, 118(1): 81-84.

361
Tennyson, G. E., Sabatos, C A., Higuchi, K , Meglin, N. and Brewer, N. (1989). Apo B-48 produced by liver and intestine. Proc. Natl. Acad. Sci. U.S.A., 86: 500-505.
Thompson, G. R. (1989). A Handbook of Hyperlipidaemia. Current Science Ltd, London.
Thompson, H. K., Hughes, L. B. and Zilversmit, D. B. (1983). Lack of Secretion of Retinyl Ester by Livers of Normal and Cholesterol-Fed Rabbits. Journal of Nutrition, 113: 1995-2001.
Tolleshaug, H., Hobgood, K. K, Brown, M. S. and Goldstein, J. L. (1983). The LDL receptor locus in familial hypercholesterolemia: multiple mutations disrupt transport and processing of a membrane receptor. Cell, 32: 941.
Tomono, S., Kawazu, S., Kato, N., Ono, T., Ishii, C, Ito, Y., Shimizu, M., Shimoyama, M., Nakano, T. and Nakajima, K. (1994). Uptake of remnant like particles (RLP) in diabetic patients from mouse peritoneal macrophages. Journal of Atherosclerosis & Thrombosis, 1 (2): 98-102.
Tso, P., Drake, D. S., Black, D. D. and Sabesin, S. M. (1984). Evidence for separate pathways of chylomicron and very-low-density lipoprotein assembly and transport by the small intestine. American Journal of Physiology, 247: G559-G610.
Tsukada, T., Rosenfeld, M. E., Ross, R. and Gown, A. M. (1986). Immunocytochemical analysis of cellular componets in atherosclerotic lesions. Use of monoclonal antibodies with the Watanabe and fat-fed rabbits. Arteriosclerosis, 6: 601-613.
Umeda, Y., Redgrave, T. G., Mortimer, B. C. and Mamo, J. C. L. (1995). Kinetics and uptake in vivo of oxidatively modified lymph chylomicrons. American Journal of Physiology, 268.
Utermann, G., Kindermann, I., Kaffarnik, H. and Steinmetz, A. (1984). Apolipoprotein E phenotypes and hyperlipidemia. Human Genetics, 65 (3): 232-236.
van Bennekum, A. M., Kako, Y., Weinstock, P. H., Harrison, E. H., Deckelbaum, R. J., Goldberg, I. J. and Blaner, W . S. (1999a). Lipoprotein lipase expression level influences tissue clearance of chylomicron retinyl ester. Journal of Lipid Research, 40 (3): 565-574.
van Berkel, T. J. C, Bihain, B. E., Ziere, G. J., Kruyt, J. K., Bijsterbosch, M. K. and Kuiper, J. (1996). Ligand specificity of the LDL-receptor related protein. Z Gastroenterol, 34 (Suppl. 3): 116-117.
van Breeman, R. B., Nikolic, D., Xu, X., Xiong, Y., van Lieshout, M., West, C. E. and Schilling, A. B. (1998). Development of a method for quantitation of retinol and retinyl palmitate in human serum using high-performance liquid chromatography-atmospheric pressure chemical ionization-mass spectrometry. Journal of Chromatography, A (794): 245-251.
Van der Westhuyzen, D. R., Fourie, A. M. and Coetzee, G. A. (1990). The LDL receptor. Current
Opinion in Lipidology, 1: 128-135.
Van Dijk, M. C. M., Ziere, G. J., Boers, W., Linthorst, C, Bijsterbosch, M. K. and Van Berkel, T. J. C. (1991). Recognition of chylomicron remnants and B-migrating very-low-density lipoproteins by the remnant receptor of parenchymal liver cells is distinct from the liver x2-macroglobulin-recognition site.
Biochemical Journal, 219: 863-870.
Van Lenten, B. J., Fogelman, A. M., Hokom, M. M., Benson, L., Haberland, M. E. and Edwards, P. A. (1983). Regulation of the Uptake and Degradation of B-Very Low Density Lipoprotein in Human Monocyte Macrophages. The Journal of Biological Chemistry, 258: 5151-5157.
Van Lenten, B. J., Fogelman, A. M., Jackson, R. L., Shapiro, S., Haberland, M. E. and Edwards, P. A. (1985). Receptor-mediated Uptake of Remnant Lipoproteins by Cholesterol-loaded Human Monocyte-macrophages. The Journal of Biological Chemistry, 260: 8783-8788.
Van Ree, J. H., van den Broek, W. J. A. A., Dahlmans, V. E. H., Groot, P. H. E., Vidgeon-Hart, M., Frants, R. R., Wieringa, B., Havekes, L. M. and Hofker, M. H. (1994a). Diet-induced

362
hypercholesterolemia and atherosclerosis in heterozygous apolipoprotein E-deficient mice. Atherosclerosis, 111: 25-37.
van Vlijmen, B. J. M., van Ree, J. H., Frants, R. R., Hofker, M. H. and Havekes, L. M. (1996). Elevated levels of chylomicron and V L D L remnants leads to atherosclerosis in apoE transgenic mice. Z. Gastroenterol, 34 (Suppl. 3): 113-115.
Vigne, J.-L. and Havel, R. J. (1981). Metabolism of apolipoprotein A-I of chylomicrons in rats and humans. Canadian Journal of Biochemistry, 59: 613-618.
Wade, D. P., Knight, B. L. and Soutar, A. K. (1986). Binding of low-density lipoprotein and chylomicron remnants to the hepatic low-density lipoprotein receptor of dogs, rats and rabbits demonstrated by ligand blotting. Failure to detect a distinct chylomicron-remnant-binding protein by ligand blotting. European Journal of Biochemistry, 159: 333-340.
Walton, K. W. (1973). The validity of the lipid-fed rabbit as a model for the study of atherogenesis. J. Pathol, 109 (2).
Warshawsky, I., Broze, G. J. and Schwartz, A. L. (1994). The low density lipoprotein receptor-related protein mediates the cellular degradation of tissue factor pathway inhibitor. Proc. Natl. Acad. Sci. U.S.A., 91: 6664-6668.
Watanabe, T., Hirate, M., Yoshikawa, Y., Nagafuchi, Y., Toyoshima, H. and Watanabe, T. (1985). Role of Macrophages in Atherosclerosis: Sequential Observations of Cholesterol-Induces Rabbit Aortic Lesion by the Immunoperoxidase Technique using Monoclonal Antimacrophage Antibody. Laboratory Investigation, 53: 80-90.
Watanabe, Y. (1980). Serial breeding with hereditary hyperlipidemia (WHHL-rabbit). Atherosclerosis, 36: 261-268.
Watanabe, Y., Ito, T., Saeki, M., Kuroda, M., Tanzawa, K., Mochizuki, M., Tsujita, Y. and Arai, M. (1981). Hypolipidemic effects of CS-500 (ML-236B) in WHHL-rabbit, a heritable animal model for hyperlipidemia. Atherosclerosis, 38 (1-2): 27-31.
Watts, G. F., Barrett, P. H., Marais, A. D., Dane-Stewart, C, Martins, I. J., Dimmitt, S. B. and Redgrave, T. G. (2001). Chylomicron remnant metabolism in familial hypercholesterolemia studied with a stable isotope breath test. Atherosclerosis, 157 (2): 519-523.
Watts, G. F., Mamo, J. C L. and Redgrave, T. G. (1998). Postprandial dyslipidaemia in a nutshell: food for thought. Aust. N.Z. J. Med., 28: 816-823.
Weber, F. (1981). In XII International Congress of NutritionNew York, pp. 119-135.
Weinberg, R. B. and Scanu, A. M. (1982). In vitro reciprocal exchange of apoproteins and non polar lipids between human high density lipoproteins and an artificial triglyceride-phospholipid emulsion (Intralipid). Atherosclerosis, 44: 141-152.
Weinberg, R. B. and Spector, M. S. (1985). Human apolipoprotein A-IV: displacement from the surface of triglyceride-rich particles by HDL2-associated C-apoproteins. Journal of Lipid Research, 26
(1): 26-37.
Weintraub, M., Burstein, A., T., R., Liron, M., Ringel, Y., Cabili, S., Blum, M., Peer, G. and Iaina, A. (1992a). Severe defect in clearing postprandial chylomicron remnants in dialysis patients. Kidney
International, 42: 1247-1252.
Weintraub, M., Grosskopf, I., Trostanesky, Y., Charach, G., Rubinstein, A. and Stern, N. (1999). Thyroxine replacement therapy enhances clearance of chylomicron remnants in patients with hypothyroidism. Journal of Clinical Endocrinology and Metabolism, 84 (7): 2532-2536.

363
Weintraub, M . S., Eisenberg, S. and Breslow, J. L. (1987a). Dietary Fat Clearance in Normal Subjects Is Regulated by Genetic Variation in Apolipoprotein E. Journal of Clinical Investigation, 80: 1571-
1577.
Weintraub, M. S., Eisenberg, S. and Breslow, J. L. (1987b). Different Patterns of Postprandial Lipoprotein Metabolism in Normal, Type Ha, Type III, and Type IV Hyperlipoproteinemic Individuals. Effects of Treatment with Cholestyramine and Gemfibrozil. Journal of Clinical Investigation, 79: 1110-1119.
Weintraub, M. S., Grosskopf, I., Rassin, T., Miller, H., Charach, G, Rotmensch, H. H., Liron, M., Rubinstein, A. and Iaina, A. (1996). Clearance of chylomicron remnants in normolipidaemic patients with coronary artery disease: case control study over three years. British Medical Journal, 312: 935-939.
Wernette-Hammond, M. E., Garcia, Z., Arnold, K. S. and Innerarity, T. L. (1989). B-very low density lipoprotein uptake in cultured fibroblasts and smooth muscle cells from Watanabe Heritable hyperlipidemic rabbits. Arterosclerosis, 9: 501-510.
Wetterau, J. R. and Zilversmit, D. B. (1986). Localization of intracellular triacylglycerol and cholesteryl ester transfer activity in rat tissues. Biochimica et Biophysica Acta, 875 (3): 610-617.
Whitlock, M. E., Swenson, T. L., Ramakrishnan, R., Leonard, M. T., Marcel, Y. L., Milne, R. W. and Tall, A. R. (1989). Monoclonal antibody inhibition of cholesteryl ester transfer protein activity in the rabbit. Effects on lipoprotein composition and high density lipoprotein cholesteryl ester metabolism.
Journal of Clinical Investigation, 84(1): 129-137.
Wieland, E., Walli, A. K., Niedmann, P. D., Lohstoter, C, Kramer, A. and Seidel, D. (1990). Gestorte Remnant clearance als Risikofaktor fur die koronare Herzkrankheit.
Williams, K. J. and Tabas, I. (1995). The response-to-retention hypothesis of early atherogenesis. Arteriosclerosis Thrombosis and Vascular Biology, 15: 551-561.
Willnow, T. E., Goldstein, J. L., Orth, K, Brown, M. S. and Herz, J. (1992). Low density lipoprotein receptor-related protein and gp330 bind similar ligands including plasminogen activator complexes and lactoferrin, an inhibitor of chylomicron remnant clearance. The Journal of Biological Chemistry, 267:
26172-26180.
Willnow, T. E., Sheng, Z., Ishibashi, S. and Herz, J. (1994). Inhibition of hepatic chylomicron remnant uptake by gene transfer of a receptor antagonist. Science, 264: 1471-1474.
Wilson, C, Wardell, M. R., Weisgraber, K. H., Mahley, R. W. and Agard, D. A. (1991). Three-dimensional structure of the L D L receptor-binding domain of human apolipoprotein E. Science, 252:
1817-1822.
Wilson, D. E. and Chan, I.-F. (1983). In Vitro Transfer of Chylomicron Retinol and Retinyl Esters. Biochemical and Biophysical Research Communications, 115: 958-964.
Wilson, D. E., Chan, I.-F. and Ball, M. (1983). Plasma Lipoprotein Retinoids After Vitamin A Feeding in Normal Man: Minimal Appearance of Retinyl Esters Among Low-Density Lipoproteins.
Metabolism, 32 (5): 514-517.
Wilson, D. E., Chan, I.-F., Buchi, K. N. and Horton, S. C. (1985b). Postchallenge Plasma Lipoprotein Retinoids: Chylomicron Remnants in Endogenous Hypertriglyceridemia. Metabolism, 34 (6): 551-558.
Wilson, D. E., Chan, I.-F., Cheung, A. K., Dutz, W. and Buchi, K. N. (1985a). Retinyl Ester Retention in Chronic Renal Failure. Further Evidence for a Defect in Chylomicron Remnant Metabolism.
Atherosclerosis, 57: 189-197.
Wilson, P. W. R, Castelli, W. P. and Kannel, W. B. (1987). Coronary risk prediction in adults (the Framingham Heart Study). American Journal of Cardiology, 59: 91G-94G.

364
Windhaus, A. (1910). Uber den gehalt normaler und atheromatoser aorten an cholesterin und cholesterinestern. Hoppe-Seyler's Z Physiol. Chem., 67: 174.
Windier, E., Chao, Y.-S. and Havel, R. J. (1980a). Determinants of Hepatic Uptake of Triglyceride-rich Lipoproteins and Their Remnants in the Rat. The Journal of Biological Chemistry, 255 (11): 5475-5480.
Windier, E., Chao, Y.-S. and Havel, R. J. (1980b). Regulation of the hepatic uptake of triglyceride-rich lipoproteins in the rat. Opposing effects of homologous apolipoprotein E and individual C apoproteins. The Journal of Biological Chemistry, 255 (17): 8303-8307.
Windier, E., Greeve, J., Daerr, W. H. and Greten, H. (1988). Binding of rat chylomicrons and their remnants to the hepatic low-density-lipoprotein receptor and its role in remnant removal. Biochemical Journal, 252: 553-561.
Windier, E., Greeve, J., Jackie, S., Rinninger, F., Pox, C, Puchta, D., Petkova, D., Robenek, H., Daerr, W . and Greten, H. (1996). Endocytic mechanisms for uptake and metabolism of chylomicron remnants in the liver. Z Gastroenterol, 34 (Suppl. 3): 103-104.
Windier, E., Greeve, J., Levkau, B., Kolb-Bachofen, V., Daerr, W. and Greten, H. (1991). The human asialoglycoprotein receptor is a possible binding site for low-density lipoproteins and chylomicron remnants. Biochemical Journal, 276: 79-87.
Wingerath, T., Kirsch, D., Spengler, B., Kaufmann, R. and Stahl, W. (1997). High-Performance Liquid Chromatography and Laser Desorption/Ionization Mass Spectrometry of Retinyl Esters. Analytical Chemistry, 69 (19): 3855-3860.
Witzum, J. L. (1994). The oxidation hypothesis of atherosclerosis. Lancet, 344 (8925): 793-795.
Wu, A.-L. and Windmueller, H. G. (1979). Relative contributions by liver and intestine to individual plasma apolipoproteins in the rat. The Journal of Biological Chemistry, 254: 7316-7322.
Wu, X., Sakata, N., Lui, E. and Ginsberg, H. N. (1994). Evidence for a lack of regulation of the assembly and secretion of apolipoprotein B-containing lipoprotein from HepG2 cells by cholesteryl ester. The Journal of Biological Chemistry, 269 (16): 12375-12382.
Wyss, R. (1995). Chromatographic and electrophoretic analysis of biomedically important retinoids. Journal of Chromatography B: Biomedical Applications, 671 (1-2): 381-425.
Yamada, N., Inoue, I., Kawamura, M., Harada, K, Watanabe, Y., Shimano, H., Gotoda, T., Shimada, M., Kohzaki, K , Tsukada, T., Shiomi, M., Watanabe, Y. Y. and Yazaki, Y. (1992). Apolipoprotein E prevents the progression of atherosclerosis in Watanabe Heritable hyperlipidemic rabbits. Journal of
Clinical Investigation, 89: 706-711.
Yamamoto, A., Matsuzawa, Y., Yokoyama, S., Funahashi, T., Yamamura, T. and Kishino, B. (1986-a). Effects of probucol on xanthomata regression in familial hypercholesterolemia. American Journal of
Cardiology, 57 (16): 29H-35H.
Yamamoto, T., Bishop, R. W., Brown, M. S., Goldstein, J. L. and Russell, D. W. (1986-b). Deletion in cysteine-rich region of L D L receptor impedes transport to cell surface in W H H L rabbit. Science, 232:
1230.
Yamamoto, T., Davis, C. G., Brown, M. S., Schneider, W. J., Casey, M. L., Goldstein, J. L. and Russell, D. W . (1984). The human LDL-receptor: a cysteine-rich protein with multiple Alu sequences
in its m R N A . Cell, 39 (1): 27-38.
Yen, F. T., Mann, C. J., Guermani, L. M., Hannouche, N. F., Hubert, N., Hornick, C A., Bordeau, V. N., Agnani, G. and Bihain, B. E. (1994). Identification of lipolysis-stimulated receptor that is distinct from the L D L receptor and the L D L receptor-related protein. Biochemistry, 33: 1172-1180.

365
Yen, F. Y., R.J., D., Mann, C J., Marcel, Y. L., Milne, R. W . and Tall, A. R. (1989). Inhibition of cholesteryl ester transfer protein activity by monoclonal antibodies: effects on cholesteryl ester formation and neutral lipid mass transfer in human plasma. Journal of Clinical Investigation, 83: 2018-2024.
Yla-Herttuala, S., Jaakkola, O, Ehnholm, C, Tikkanen, M. J., Solakivi, T., Sarkioja, T. and Nikkari, T. (1988). Characterization of two lipoproteins containing apolipoproteins B and E from lesion-free human aortic intima. Journal of Lipid Research, 29 (5): 563-572.
Young, S. G. (1990). Recent progress in understanding apolipoprotein B. Circulation, 82: 1574-1594. Young, S. G., Peralta, R P., Dubois, B. W., Curtiss, L. K , Boyles, J. K. and Witztum, J. L. (1987). Lipoprotein B37, a naturally occurring lipoprotein containing the amino-terminal portion of apolipoprotein B100, does not bind to the apolipoprotein B,E (low density lipoprotein) receptor. The Journal of Biological Chemistry, 262 (34): 16604-16611.
Yu, K. C. and Cooper, A. D. (2001). Postprandial lipoproteins and atherosclerosis. Frontiers in Bioscience, 1 (6): D332-D354.
Yu, K. C, Jiang, Y., Chen, W. and Cooper, A. D. (1999). Evaluation of the components of the chylomicron remnant removal mechanism by use of the isolated perfused mouse liver. Journal of Lipid Research, 40 (10): 1899-1910.
Yu, K. C, Jiang, Y., Chen, W. and Cooper, A. D. (2000). Rapid initial removal of chylomicron remnants by the mouse liver does not require hepatically localized apolipoprotein E. Journal of Lipid Research, 41 (11): 1715-1727.
Yu, K. C. and Mamo, J. C. (1997a). Binding and uptake of chylomicron remnants by cultured arterial smooth muscle cells from normal and Watanabe-heritable-hyperlipidemic rabbits. Biochimica et Biophysica Acta, 23 (3): 212-220.
Yu, K. C. and Mamo, J. C (1997b). Regulation of cholesterol synthesis and esterification in primary cultures of macrophages following uptake of chylomicron remnants. Biochemistry & Molecular Biology International, 41 (1): 33-39.
Yu, K. C. and Mamo, J. C. (2000). Chylomicron-remnant-induced foam cell formation and cytotoxicity: a possible mechanism of cell death in atherosclerosis. Clinical Science, 98 (2): 183-192.
Yu, K. C.-W., Smith, D., Yamamoto, A., Kawaguchi, A., Harada-Shiba, M., Yamamura, T. and Mamo, J. C L. (1997). Phagocytic degradation of chylomicron remnants by fibroblasts from subjects with homozygous familial hypercholesterolemia. Clinical Science, 92 (2): 197-203.
Zannis, V. I., Cole, S. F., Jackson, C, Kurnit, D. M. and Karathanasis, S. K. (1985). Distribution of apolipoprotein A-I, C-II, CIII, and E m R N A in fetal human tissues. Time-dependent induction of apolipoprotein E m R N A by cultures of human monocyte-macrophages. Biochemistry, 24 (16): 4450-
4455.
Zeng, B.-J., Mortimer, B.-C, Martins, I. J., Seydel, U. and Redgrave, T. G. (1998). Chylomicron remnant uptake is regulated by the expression and function of heparan sulfate proteoglycan in hepatocytes. Journal of Lipid Research, 39 (4): 845-860.
Zerbinatti, C. V., Olivecrona, H. C. F., Wechesler, S. and Quintao, E. C. R. (1991). Independent Regulation of Chylomicron Lipolysis and Particle Removal Rates: Effects of Insulin and Thyroid Hormones on the Metabolism of Artificial Chylomicrons. Metabolism, 40 (11): 1122-1127.
Zhang, S. H., Reddick, R. L., Burkey, B. and Maeda, N. (1994). Diet-induced Atherosclerosis in Mice Heterozygous and Homozygous for Apolipoprotein E Gene Disruption. Journal of Clinical
Investigation, 94: 937-945.

366
Zhang, S. H., Reddick, R. L., Piedrahita, J. A. and Maeda, N. (1992). Spontaneous Hypercholesterolemia and Arterial Lesions in Mice Lacking Apolipoprotein E. Science, 258: 468-471.
Zhang, Z., Cianflone, K. and Sniderman, A. D. (1999). Role of cholesterol ester mass in regulation of secretion of ApoBlOO lipoprotein particles by hamster hepatocytes and effects of statins on that relationship. Arteriosclerosis Thrombosis and Vascular Biology, 19 (3): 743-752.
Zilversmit, D. B. (1965). The composition and structure of lymph chylomicrons in dog, rat and man. Journal of Clinical Investigation, 44: 1610-1622.
Zilversmit, D. B. (1978). In Assembly of Chylomicrons in the Intestinal Cell. Disturbances in Lipid and Lipoprotein Metabolism, (Eds, Dietschy, J. M., Gotto Jnr, A. M . and Outko, J. A.) Williams & Wilkins Company,, pp. 69-98.
Zilversmit, D. B. (1979). Atherogenesis: A Postprandial Phenomenon. Circulation, 60 (3): 473-485.
Zilversmit, D. B., Morton, R. E., Hughes, L. B. and Thompson, K. H. (1982). Exchange of retinyl and cholesteryl esters between lipoproteins of rabbit plasma. Biochimica et Biophysica Acta, 712: 88-93.
Zilversmit, D. B. and Shea, T. M. (1989). Quantitation of apoB-48 and apoB-100 by gel scanning or radio-iodination. Journal of Lipid Research, 30: 1639-1646.

Related Documents











