AN EARLY MIOCENE PLANT ASSEMBLAGE SEVERELY INFLUENCED BY A VOLCANIC ERUPTION, STYRIA, AUSTRIA by JOHANNA KOVAR-EDER, MARGIT HAAS, CHRISTA-CHARLOTTE HOFMANN and BARBARA MELLER ABSTRACT. Sediment and organic facies as well as leaves, pollen, and diaspores co-occurring in a tuffite exposed in Oberdorf (Ko ¨flach/Voitsberg lignite area, Styria, Austria) were investigated. The tuffite is distinguishable from the under- and overlying clastic sediments and shows locally normally-graded lapilli, which is assumed to be a sign of direct airfall. The plant assemblage partly resulted from a volcanic eruption that coincided with the season in which many woody plants were flowering; deciduous ones were leafless or in the state of opening their buds; diaspores were not yet developed. Leaves of evergreen woody species, buds/bud scales, and clusters of immature pollen partly with adhering anther material were stripped off their parent plants and quickly embedded in the tuffite. The leaves of deciduous taxa and herbaceous monocotyledons, diaspores, partly also pollen, and organic facies components indicate progressive degradation. Such degraded plant debris probably accumulated in the backswamp sediments prior to the volcanic eruption. The tephra can only be correlated to the ‘Lower Rhyolite Tuffs’ in the Pannonian Basin (Hungary), indicating a (Late) Eggenburgian/Early Ottnangian age. The new species Ternstroemites waltheri is described. Distylium fergusonii Knobloch and Kvac ˇek and Laurophyllum rugatum Kvac ˇek and Bu ˚z ˇek are first reports from Austria. KEY WORDS: palaeobotany, taphonomy, volcanism. T HE Ko ¨ flach/Voitsberg lignite mining area is located about 30 km west of Graz, Styria (Text-fig. 1A). The Oberdorf Basin-fill was the subject of a recently published, detailed geological, sedimentological, palaeontological, and stratigraphical study (Steininger, ed. 1998). Micro-mammal-bearing vertebrate assemblages from the middle part of the hanging wall sequence (Text-fig. 1C– D) have been dated to the Mammal Neogene Zone 4, correlated to the Ottnangian, Early Miocene. Combined with the palaeomag- netic results, this part of the Oberdorf section has been correlated to chron 5Dn of the Geomagnetic Polarity Time Scale, currently 17·6–17·2 Ma (Daxner-Ho ¨ck et al. 1998; Steininger et al. 1998; Mauritsch and Scholger 1998). The tuffite was briefly exposed in 1997 in the eastern sub-basin of the Oberdorf opencast mine. Sedimentary and organic facies, as well as fossil leaf, diaspore, and pollen assemblages were investigated. The results presented here clearly reflect the synergetic effects of the multidisciplinary investigations. SEDIMENTARY ENVIRONMENT AND STRATIGRAPHY Sedimentary environment The 0·04–0·08-m-thick tuffite occurred as a distinct layer (sharp boundaries with the over- and underlying sediments; Pl. 3, fig. 12) within clastic and lignitic overbank deposits. It was exposed laterally for about 100 m in a north–south direction in the eastern part of the eastern sub-basin. Despite the thorough field investigations (Haas 1998; Kovar-Eder et al. 1998; Meller 1998; Meller et al. 1999), the tuffite has not been observed in any other part of either sub-basin, nor was it recovered in cores. The underlying units are characterized by a general fining-upward succession from medium/coarse- grained fluvial gravels to fine-grained overbank deposits indicating a decrease of current energy; this resulted in the accumulation of floodplain, crevasse, and levee sediments (Haas 1998, 1999). [Palaeontology, Vol. 44, Part 4, 2001, pp. 575–600, 4 pls] q The Palaeontological Association

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A N E A R L Y M I O C E N E P L A N T A S S E M B L A G E S E V E R E L Y
I N F L U E N C E D B Y A V O L C A N I C E R U P T I O N , S T Y R I A ,
A U S T R I A
by J O H A N N A K O V A R - E D E R , M A R G I T H A A S , C H R I S T A - C H A R L O T T E H O F M A N N andB A R B A R A M E L L E R
ABSTRACT. Sediment and organic facies as well as leaves, pollen, and diaspores co-occurring in a tuf®te exposed inOberdorf (Koȯach/Voitsberg lignite area, Styria, Austria) were investigated. The tuf®te is distinguishable from theunder- and overlying clastic sediments and shows locally normally-graded lapilli, which is assumed to be a sign ofdirect airfall. The plant assemblage partly resulted from a volcanic eruption that coincided with the season in whichmany woody plants were ¯owering; deciduous ones were lea¯ess or in the state of opening their buds; diaspores werenot yet developed. Leaves of evergreen woody species, buds/bud scales, and clusters of immature pollen partly withadhering anther material were stripped off their parent plants and quickly embedded in the tuf®te. The leaves ofdeciduous taxa and herbaceous monocotyledons, diaspores, partly also pollen, and organic facies components indicateprogressive degradation. Such degraded plant debris probably accumulated in the backswamp sediments prior to thevolcanic eruption. The tephra can only be correlated to the `Lower Rhyolite Tuffs' in the Pannonian Basin (Hungary),indicating a (Late) Eggenburgian/Early Ottnangian age. The new species Ternstroemites waltheri is described. Distyliumfergusonii Knobloch and KvacÏek and Laurophyllum rugatum KvacÏek and BuÊzÏek are ®rst reports from Austria.
KEY WORDS: palaeobotany, taphonomy, volcanism.
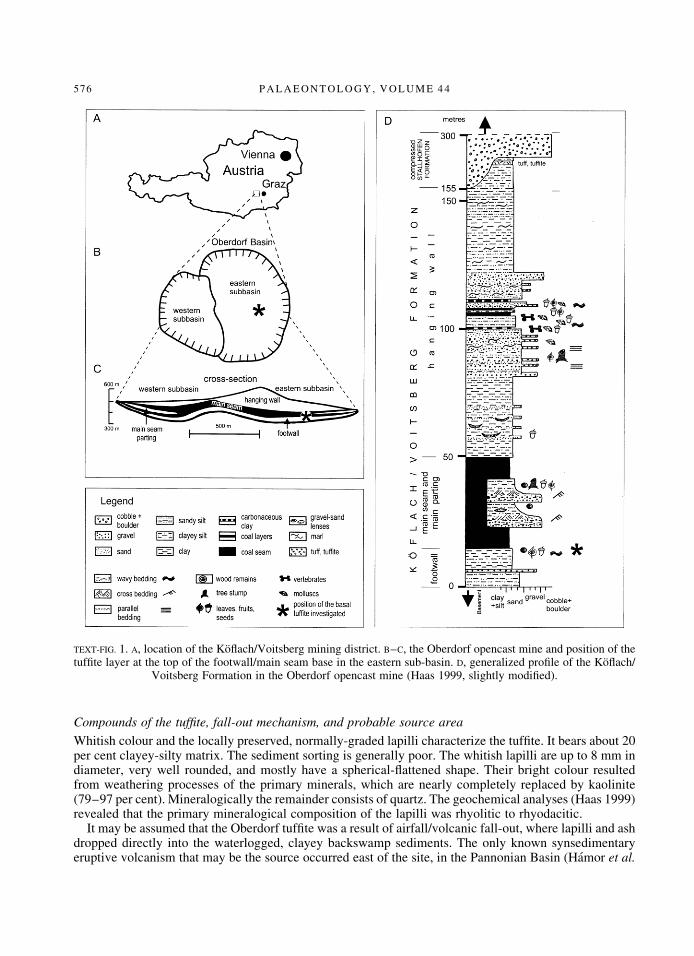
T H E Koȯach/Voitsberg lignite mining area is located about 30 km west of Graz, Styria (Text-®g. 1A). TheOberdorf Basin-®ll was the subject of a recently published, detailed geological, sedimentological,palaeontological, and stratigraphical study (Steininger, ed. 1998). Micro-mammal-bearing vertebrateassemblages from the middle part of the hanging wall sequence (Text-®g. 1C±D) have been dated to theMammal Neogene Zone 4, correlated to the Ottnangian, Early Miocene. Combined with the palaeomag-netic results, this part of the Oberdorf section has been correlated to chron 5Dn of the GeomagneticPolarity Time Scale, currently 17´6±17´2 Ma (Daxner-HoÈck et al. 1998; Steininger et al. 1998; Mauritschand Scholger 1998).
The tuf®te was brie¯y exposed in 1997 in the eastern sub-basin of the Oberdorf opencast mine.Sedimentary and organic facies, as well as fossil leaf, diaspore, and pollen assemblages were investigated.The results presented here clearly re¯ect the synergetic effects of the multidisciplinary investigations.
S E D I M E N T A R Y E N V I R O N M E N T A N D S T R A T I G R A P H Y
Sedimentary environment
The 0´04±0´08-m-thick tuf®te occurred as a distinct layer (sharp boundaries with the over- and underlyingsediments; Pl. 3, ®g. 12) within clastic and lignitic overbank deposits. It was exposed laterally for about100 m in a north±south direction in the eastern part of the eastern sub-basin. Despite the thorough ®eldinvestigations (Haas 1998; Kovar-Eder et al. 1998; Meller 1998; Meller et al. 1999), the tuf®te has notbeen observed in any other part of either sub-basin, nor was it recovered in cores.
The underlying units are characterized by a general ®ning-upward succession from medium/coarse-grained ¯uvial gravels to ®ne-grained overbank deposits indicating a decrease of current energy; thisresulted in the accumulation of ¯oodplain, crevasse, and levee sediments (Haas 1998, 1999).
[Palaeontology, Vol. 44, Part 4, 2001, pp. 575±600, 4 pls] q The Palaeontological Association
Compounds of the tuf®te, fall-out mechanism, and probable source area
Whitish colour and the locally preserved, normally-graded lapilli characterize the tuf®te. It bears about 20per cent clayey-silty matrix. The sediment sorting is generally poor. The whitish lapilli are up to 8 mm indiameter, very well rounded, and mostly have a spherical-¯attened shape. Their bright colour resultedfrom weathering processes of the primary minerals, which are nearly completely replaced by kaolinite(79±97 per cent). Mineralogically the remainder consists of quartz. The geochemical analyses (Haas 1999)revealed that the primary mineralogical composition of the lapilli was rhyolitic to rhyodacitic.
It may be assumed that the Oberdorf tuf®te was a result of airfall/volcanic fall-out, where lapilli and ashdropped directly into the waterlogged, clayey backswamp sediments. The only known synsedimentaryeruptive volcanism that may be the source occurred east of the site, in the Pannonian Basin (HaÂmor et al.
576 P A L A E O N T O L O G Y , V O L U M E 4 4
TEXT-FIG. 1. A, location of the Koȯach/Voitsberg mining district. B±C, the Oberdorf opencast mine and position of thetuf®te layer at the top of the footwall/main seam base in the eastern sub-basin. D, generalized pro®le of the Koȯach/
Voitsberg Formation in the Oberdorf opencast mine (Haas 1999, slightly modi®ed).

1978; HaÂmor 1979; SzeÂky-Fux et al. 1987; A rva-SoÂs and MaÂthe 1992). In contrast, the heavy mineraldistributions of the ¯uvial sediments in Oberdorf de®ne their source area in the north/north-west/west/south-west (Haas 1999, p. 93). This largely excludes a ¯uvial input of the lapilli.
Stratigraphy
Owing to the strong weathering and alteration of the lapilli, K/Ar-dating was not possible. However, theposition of the tuf®te in the Oberdorf sequence classi®es it as a product of a volcanic activity older than17´6 Ma (Early Miocene, see above and Text-®g. 2).
The rare earth elements (REE) were investigated to compare them with those from tuffs in thePannonian Basin (Haas 1999, p. 100). The REE-patterns resemble those described from Tokaj by Downeset al. (1995). However, these tuffs are younger (Badenian/Sarmatian). Unfortunately, REE patterns fromthe Middle and Lower Rhyolite Tuffs (MRT, LRT) are not available. Therefore, only crude comparisonsare possible. The MRT are younger (average age 16´4 6 0´8 Ma; HaÂmor et al. 1978; HaÂmor 1979) whilstthe LRT constitute the oldest volcanic deposits in the Pannonian Basin. K/Ar dating indicates 19´6 6 1´4Ma (HaÂmor et al. 1978; HaÂmor 1979). Even more accurate dating from the rhyolites of the Central Trans-Tisza region provides an age of 18´25 6 0´3 Ma (SzeÂky-Fux et al. 1987; A rva-SoÂs and MaÂthe 1992). The
K O V A R - E D E R E T A L . : M I O C E N E P L A N T S 577
TEXT-FIG. 2. Stratigraphic scheme of the Early Miocene (from Steininger et al. 1998).

correlation of the tuf®te from Oberdorf with the LRT in the Pannonian Basin is the only reasonableexplanation, thus indicating a (Late) Eggenburgian/Early Ottnangian age.
The rhyolitic tuffs/tuf®tes within the basal parts of the overlying Stallhofen Formation on theTregistsattel (Text-®g. 1D), east of Oberdorf, cannot be directly compared to a speci®c volcanic activityor source area either. Geochemical comparisons with Styrian volcanics of Karpatian/Early Badenian ageshowed similarities to the volcanic rocks of the Gleichenberg area. However, tuffs from Hungary have notbeen compared. The age of the Stallhofen Formation (Badenian) is based on regional geology only (Ebneret al. 1998, p. 427).
M A T E R I A L A N D M E T H O D S O F P L A N T I N V E S T I G A T I O N S
All the material investigated co-occurs in the 0´04±0´08 m thick tuf®te described above.
Leaves
The botanical determinations of the leaf remains are based on cuticular analysis (standard procedure HF,Schulze's reagent followed by KOH) owing to the lack of gross morphologically signi®cant features. Thecuticular terminology follows Dilcher (1974).
Palynomorphs, organic facies
Three samples taken from the tuf®te at some lateral distance from one another were processed in thestandard way with HCl and HF for palynological and organic facies analysis. One-half of the resultingextracts was kept for organic facies analyses; the other half was acetolysed to reduce the organic contentand concentrate the palynomorphs (Klaus 1987, p. 232). At least two slides each of the organic facies andpalynomorph extract were point counted. For the organic facies 250±300 counts per slide are suf®cient fortotal percentages. Palynomorph counts should be considerably higher, at least 350. In poorer samples itwas necessary to count through several slides (e.g. ®ve slides for sample 2) to reach these numbers.Average percentages of the relative numerical abundance of organic facies components and averaged totalpercentages of the palynomorphs of all three samples investigated are presented here.
Pollen clusters (see below) were transferred from the counting slide into a drop of glycerine mounted ona clean slide after the method of Zetter (1989) to obtain high-quality photographs. Unfortunately, some ofthe delicate clusters, mostly of the rare Alnus pollen, disintegrated into individual grains. The Alnus pollenappeared to be mature and the adhesion of the pollenkit was probably minimal. Massive pollen clusters ofFagaceae revealed little in LM photographs; SEM images were necessary to demonstrate their denselypacked nature. Five to 11 per cent of the counted palynomorphs could not be identi®ed because of poorpreservation.
Diaspores
About ®ve litres of sediment were analysed palaeocarpologically (sieved and washed). Based onexperience of other ®ne-grained layers in Oberdorf (Meller 1998, p. 506), this sample quantity shouldinclude the main diaspore spectrum.
The material was collected under the sample OENE-Me 97/3. It is kept in the palaeobotanical collection of the NaturalHistory Museum Vienna, under collection nos 1998B0005 (leaves), 1999B0053 (diaspores), and 1999B0054 (pollen).
T A P H O N O M Y A N D P R E S E R V A T I O N O F T H E D I F F E R E N T P L A N T O R G A N S
Leaves
During the ®eldwork at Oberdorf the densely packed leaf remains appeared to be mechanically heavilydamaged and fragmented. Almost complete leaves are exceptional and these are relatively small. It is
578 P A L A E O N T O L O G Y , V O L U M E 4 4

dif®cult to judge to what extent this re¯ects taphonomic processes and/or the complicated splitting of theirregularly strati®ed lapilli layer.
The good cuticle preservation of evergreen foliage contrasts strongly with the poor cuticle preservationof presumably deciduous taxa. Although not unusual, this is nonetheless a noteworthy phenomenon.While the cuticle preservation of deciduous foliage is worse, that of evergreen foliage is better than usualin the Oberdorf samples. Evergreen taxa dominate qualitatively and quantitatively (species number andspecimens) over deciduous ones.
Palynomorphs
Within the palynomorph assemblage some pollen types show little or no degradation, while others displayobvious signs of mechanical and/or biochemical destruction, resulting in high numbers of `unidenti®edpalynomorphs'. No patterns are recognizable except the generally good preservation of pollen packedtogether in clusters. Many of these pollen grains look quite immature, such as those of the Fagaceae andTaxodiaceae, and are often still attached to anther material (Pl. 4; Text-®g. 3).
Diaspores/buds, bud scales, etc.
The preservation of the diaspores is generally poor; they largely lack anatomical features. This contrastswith the high abundance and mostly excellent preservation of buds and bud scales. The sediment residuealso contained Taxodiaceae twig fragments, fusain splinters, and small pieces of resin. Fusain splinters arecommon in all parts of the Oberdorf section and we lack evidence whether those from the tuf®te are relatedto volcanic activity.
L E A F S P E C T R U M
More than 150 leaf remains have been investigated, of which about 20 fragments remain undeterminedowing to poor preservation (Table 1, pls 1±3). Thirteen leaf taxa have been determined, most of themrepresenting presumably evergreen foliage of mesophytic forest elements. Therefore, the tuf®te is morespecies rich than any other of the numerous megafossil-bearing samples from Oberdorf that have beeninvestigated (Kovar-Eder 1996; Kovar-Eder et al. 1998; Meller et al. 1999; Kovar-Eder and Meller inpress a, b). In contrast to all other leaf- and diaspore-bearing samples, taxa of zonal versus azonal habitatsare more diverse in the leaf than in the diaspore record. Leaves of deciduous woody plants are far lessabundant and their determinations are questionable: ?Alnus/Betulaceae, ?Fraxinus, ?Salix. This alsocontrasts with the previously investigated leaf-bearing Oberdorf samples.
Trigonobalanopsis rhamnoides (Pl. 3, ®g. 3) is by far the most dominant species, followed byDaphnogene polymorpha (Pl. 3, ®g. 2). The Lauraceae are represented by three species: D. polymorpha,Laurophyllum pseudoprinceps (Pl. 3, ®g. 1), and L. rugatum (two fragmentary leaves: Pl. 1, ®gs 3±7; Pl. 2,®g. 8; Pl. 3, ®g. 5). The latter and Distylium fergusonii (nine leaf fragments: Pl. 1, ®gs 1±2; Pl. 3 ®gs 7±8)have not been recorded from Austria before. The co-occurrence of Distylium uralense (diaspores) in thetuf®te is remarkable. Ternstroemites waltheri (Pl. 2, ®gs 1±7; Pl. 3, ®g. 6) is a new Theaceae species(seven leaf fragments), increasing the Theaceae foliage record in Oberdorf to three species (?Gordoniaoberdorfensis and ?Cleyera schilcheriana have been described from the hanging-wall sediments; Kovarand Meller in press a). Some poorly preserved monocotyledonous leaf fragments (Pl. 3, ®g. 10) arenoteworthy, as such remains are extremely rare in Oberdorf.
P A L Y N O S P E C T R U M
Approximately 65 taxa have been distinguished (Table 2, Text-®g. 3A±E, Pl. 4), representing about 62 percent of the micro¯oral spectrum present in the Oberdorf samples investigated by Zetter (1998). The tuf®teis, therefore, considered to be reasonably diverse in taxa.
K O V A R - E D E R E T A L . : M I O C E N E P L A N T S 579
Taxodiaceae (15±22 per cent), Alnus sp. (13±16 per cent), and bisaccates (Pinus spp. plusCathaya) (19±21 per cent) are the important taxa. Less abundant are Lithocarpus and Trigonobalanopsis(both 2±4 per cent), Quercus (1±4 per cent), Carya (4±7 per cent), Platycarya and Engelhardia (both 2±4 per cent), while Pterocarya reaches only 2 per cent in sample 1. In some cases a particular taxon maybe only <0´5 per cent in one sample, but amount to >2 per cent in the others; e.g. Ilex, Pterocarya,and Sparganium/Typha. These differences are not considered to be signi®cant. The main facieselements, Alnus, Taxodiaceae, and bisaccates, are represented in the tuf®te by relatively similarpercentages.
Frequently occurring pollen packed together in clusters, occasionally attached to anther material, appearto be quite immature (Text-®g. 3, Pl. 4). They derive from wind-pollinated taxa such as Taxodiaceae,Alnus, Engelhardia, Fagus, Lithocarpus, Quercus, Trigonobalanopsis, and Poaceae, as well as frominsect-pollinated forms such as Ilex, and Sparganium/Typha. The statistical-numerical abundance ofpollen clusters in the three samples investigated is about 2´5 per cent. Some taxa, such as Alnus, Pinaceae,and Taxodiaceae, are represented by both well- and poorly preserved pollen grains.
Pollen clusters, with or without attached anther tissue, are not such an unusual phenomenon with insect-pollinated ¯owers, because vigorous insect activity may lead to the destruction of ¯oral structures. Buttheir abundance in the tuf®te together with clusters (at least eight taxa) of wind-pollinated forms (at leastseven taxa) is quite unusual. Walther and Zetter (1993) described Trigonobalanopsis pollen clusters stuckto the underside of Trigonobalanopsis rhamnoides (RossmaÈssler) KvacÏek and Walther leaves. They
580 P A L A E O N T O L O G Y , V O L U M E 4 4
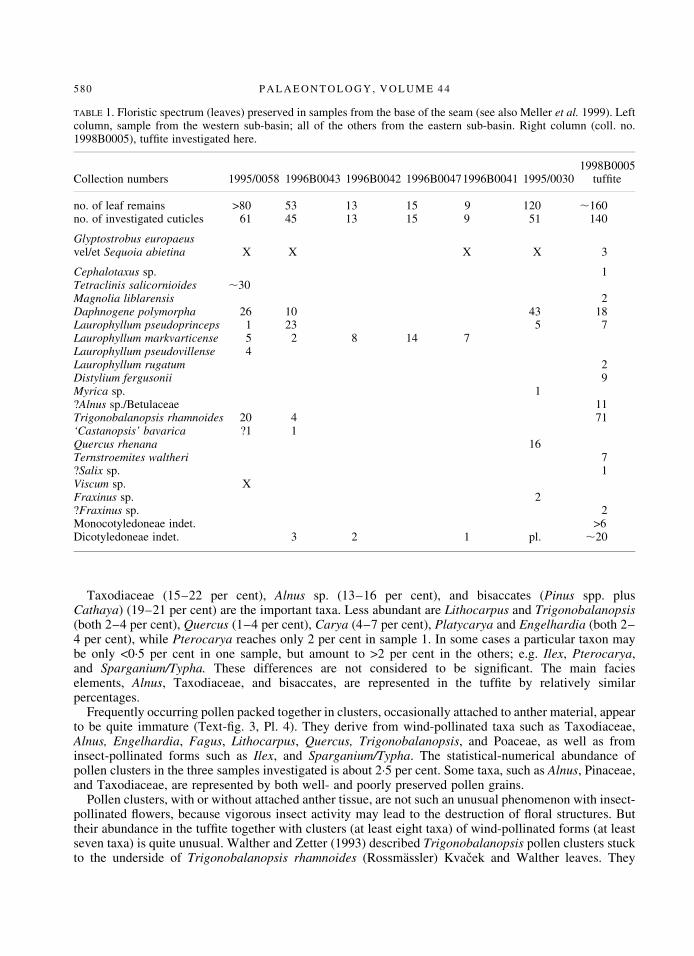
TABLE 1. Floristic spectrum (leaves) preserved in samples from the base of the seam (see also Meller et al. 1999). Leftcolumn, sample from the western sub-basin; all of the others from the eastern sub-basin. Right column (coll. no.1998B0005), tuf®te investigated here.
1998B0005Collection numbers 1995/0058 1996B0043 1996B0042 1996B00471996B0041 1995/0030 tuf®te
no. of leaf remains >80 53 13 15 9 120 ,160no. of investigated cuticles 61 45 13 15 9 51 140
Glyptostrobus europaeusvel/et Sequoia abietina X X X X 3
Cephalotaxus sp. 1Tetraclinis salicornioides ,30Magnolia liblarensis 2Daphnogene polymorpha 26 10 43 18Laurophyllum pseudoprinceps 1 23 5 7Laurophyllum markvarticense 5 2 8 14 7Laurophyllum pseudovillense 4Laurophyllum rugatum 2Distylium fergusonii 9Myrica sp. 1?Alnus sp./Betulaceae 11Trigonobalanopsis rhamnoides 20 4 71`Castanopsis' bavarica ?1 1Quercus rhenana 16Ternstroemites waltheri 7?Salix sp. 1Viscum sp. XFraxinus sp. 2?Fraxinus sp. 2Monocotyledoneae indet. >6Dicotyledoneae indet. 3 2 1 pl. ,20

K O V A R - E D E R E T A L . : M I O C E N E P L A N T S 581
TABLE 2. Palynomorph spectrum from the tuf®te.
sample 1 % sample 2 % sample 3 %
Alnus (2 forms?) 18 Taxodiaceae (2 forms) 22 Taxodiaceae (2 forms) 18Taxodiaceae (2 forms) 15 Aluns (2 forms?) 16 Alnus (2 forms?) 13Pinus (2 forms) 9 Pinus spp. (2 forms) 11 Pinus (2 forms) 12bisaccates 8 Cathaya 5 Carya 5Carya 7 bissaccates 4 bisaccates 5Lithocarpus 4 Carya 4 Cathaya 4Quercus (3 forms) 4 Polypodiaceae smooth 4 Lithocarpus 4Platycarya 4 Engelhardia (2 forms) 4 Trigonobalanopsis 4Polypodiaceae smooth 3 Osmunda 3 Quercus (3 forms) 3Engelhardia (2 forms) 3 Platycarya 2 Polypodiaceae smooth 2Cathaya 2 Trigonobalanopsis 2 Ulmus 2Trigonobalanopsis 2 Lithocarpus 2 Platycarya 2Pterocarya 2 Ulmus 2 Polypodiaceae rugulose 2Tilia 1 Sparganium/Typha 2 Ilex (2 forms) 2Osmunda 1 Zelkova 1 Engelhardia (2 forms) 2Ulmus 1 Quercus (2 forms) 1 Fraxinus (2 forms) 2Polypodiaceae rugulose 1 Parthenocissus 1Myrica 1 Fagus 1<1% <1% <1%Acer Pterocarya PterocaryaRosaceae gen. indet. Tilia Ericaceae (2 forms)Salix Tiliaceae gen. indet. TiliaNyssa Polypodiaceae rugulose Tiliaceae gen. indet.
Rosaceae (2 forms) OsmundaAcer (2 forms) fern gen. indet.Betula CeltisHamamelidaceae gen. indet. Platanus
Salixaccessory (1±2 grains) accessory (1±2 grains) accessory (1±2 grains)Reveesia Ilex (2 forms) CornusParthenocissus Sapotaceae gen. indet. Rosaceae (Rubus type)Picea Ericaceae (2 forms) Poaceae gen. indet.Celtis Carpinus ReveesiaPlatanus Platanus Rutaceae (2 forms)Sapotaceae Salix ZelkovaFraxinus (2 forms) Celtis PlatanusCarpinus Myrica MastixiaStyracaceae gen. indet. Styracaceae gen. indet. AbiesFagus Parthenocissus Anacardiaceae (Rhus type)Ilex Rutaceae (2 forms) Sapotaceae gen. indet.fern gen. indet. Poaceae gen. indet. PiceaHamamelidaceae gen. indet. Fraxinus NyssaOleaceae gen. indet. Apiaceae gen. indet. Sparganium/Typhaincertae sedis 11 Arecaceae Vitis
Mastixia Hamamelidaceae gen. indet.Monocotyledoneae gen. indet. Asteraceae gen. indet.incertae sedis 5 Monocotyledoneae gen. indet.
Pteridaceae gen. indet.incertae sedis 6
ON � 384 ON � 415 ON � 394
582 P A L A E O N T O L O G Y , V O L U M E 4 4

proposed that the pollen grains released from male ¯owers were slightly sticky and could thereforeadhere to the even stickier lower leaf surface. They appeared to be mature pollen grains, and anthermaterial was not observed there; this indicates ambophily (entomophilous in younger stages thenanemophilous because of the loss of stickiness of the pollenkit) of this particular pollen type (R. Zetter,pers. comm. 1999).
O R G A N I C F A C I E S
Translucent brown wood fragments (15±20 per cent), strongly coali®ed wood (12±22 per cent; counted assemi-opaque material), and amorphous organic matter (AOM) are important components in the organicfacies (Table 3). In particular, heterogeneous AOM (up to 30±40 per cent) indicates strong degradationbecause of fungal activity and other decompositional processes. Heterogeneous AOM probably resultedfrom the decomposition of weakly ligni®ed plant tissue, algal colonies, and completely fungi-infestedwoody tissue. Owing to the heavy degradation, structured materials other than wood, such as cuticles,membranes, different types of algae, algal cysts, and palynomorphs (all in the range of 3±7 per cent), areless common: most of these show signs of progressive decomposition. Isolated fungal spores occur onlyoccasionally, whereas isolated fungal hyphae are common (#9 per cent; all other fungal matter, such asmycelia, is hidden in the heterogeneous AOM). The relative numerical abundancies of the components do
K O V A R - E D E R E T A L . : M I O C E N E P L A N T S 583
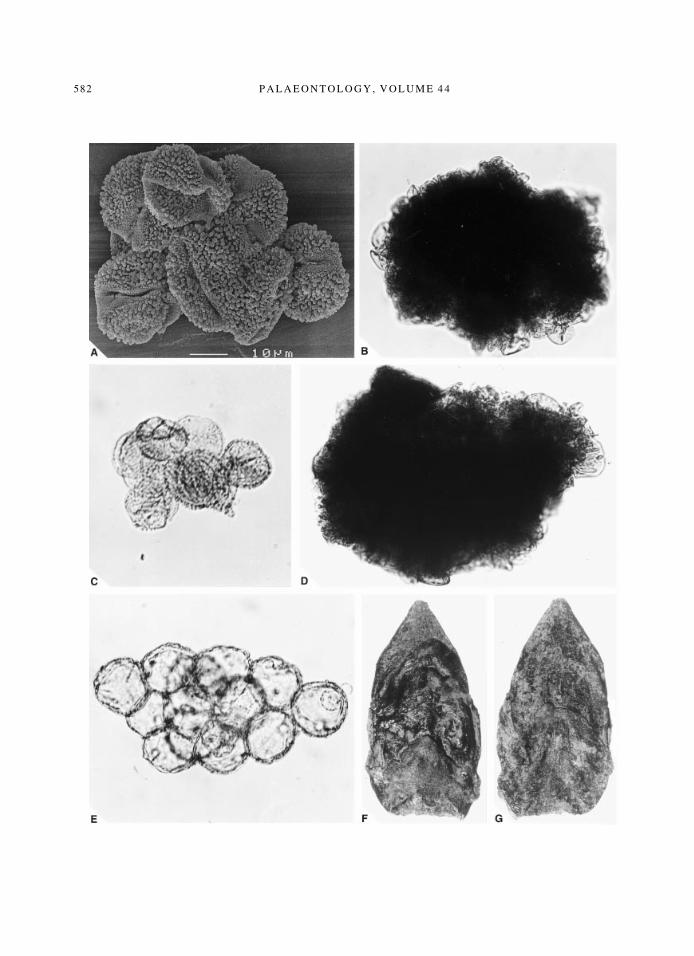
TEXT-FIG. 3. A±E, Coll. no. 1999B0054. A, Ilex sp. pollen cluster; pollen partly de¯ated and adpressed, micro-morphology fully developed, SEM; ´ 1100. B, anther of Taxodiaceae; pollen of very different sizes and maturity,very small and immature at the top, more mature and larger at the bottom, SEM; ´ 900. C, Ilex sp. pollen cluster;no anther material attached, LM; ´ 900. D, one of the numerous dense and solid anthers of no af®liation, LM;´ 900. E, Fagus sp. pollen cluster; most pollen grains still in the tetrahedron con®guration, most clearly visible inthe centre of the cluster, LM; ´ 900. F±G, ?Alnus sp.; complete bud, front and back side; coll. no. 1999B0053/12/A,
LM; ´ 5.
TABLE 3. Organic facies from the tuf®te. Heterogeneous AOM, degraded non-ligni®ed plant tissue and/or degradedalgae; homogeneous AOM, translucent, brownish, lath-shaped, thin sheets, possibly originating from ligni®ed cellwalls, but without diagnostic features; humic AOM, precipitated humic acids characterized by an oval to roundishshape; semi-opaque material, mostly stronger coali®ed wood and/or fungal infested/melanized tissue of both non-ligni®ed and ligni®ed nature.
components % sample 1 sample 2 sample 3
palynomorphs 2´5 <1 2brown wood 15 20 19fusain (mostly wood) <1 1 0cuticles 4´5 4 5membranes/translucent 6´5 4 4other plant parts 0 1´5 <1fungal hyphae 10 8 5´5fungal spores <1 1 1algae 2 3´5 5´5heterogeneous AOM 40 30 32homogeneous AOM 2 0 2humic AOM <1 <1 <1resin <1 <1 <1semi-opaque particles 12 22 21bituminous matter 4 3´5 1´5
N � 260 N � 271 N � 252

not vary signi®cantly. Sample 1 is much ®ner grained than the others; the wood and semi-opaquecomponents appear more rounded rather than lath-shaped, providing evidence for longer transport and/orresedimentation. Bitumen-like smears on the slides are relatively frequent, indicating the former presenceof oil-producing colonial algae such as Botryococcus, Pediastrum, and Zygnematales, which are mostlypoorly preserved and thus not easy to recognize. These features are quite common and typical for thesubstrate of backswamp environments, which often constitute a mosaic of puddles and herbaceous andwoody vegetation.
D I A S P O R E S P E C T R U M
About 90 specimens represent 11 taxa; four specimens remained undetermined (Table 4). The ¯oristicspectrum as well as the abundances of taxa ®t with other species-poor samples from the top of the footwall/base of the seam (Meller et al. 1999; Table 4, samples 2±3, 10). None of the taxa is new in the Oberdorfrecord and most may be regarded as representatives of azonal habitats: Sequoia abietina (Kovar-Eder andMeller in press a), Ampelopsis malvaeformis, A. rotundata, Sparganium, Carex, and possibly also Rubus.Distylium uralense, Manglietia germanica, and Poliothyrsis eurorimosa possibly represent mesophyticforest elements. Generally, the latter two are rare in the Oberdorf section. M. germanica, however, seemsto be relatively enriched in the tuf®te. The relatively low representation of presumably mesophyticelements strongly contrasts with the leaf and palynomorph record.
B U D S A N D B U D S C A L E S
The very well-preserved buds and isolated bud scales (Text-®g. 3F±G) probably all derive from Alnus (seesystematic palaeontology below). The occurrence of buds and bud scales is a common feature in washedand sieved samples from Oberdorf. However, none of the diaspore samples investigated hitherto was sorich in buds and bud scales as this material.
D I S C U S S I O N
Taphonomy
Because the tuf®te is developed as a distinct layer and displays locally well-preserved, normally-gradedlapilli, the volcanic sediments were probably deposited by direct airfall rather than by ¯uvial transport
584 P A L A E O N T O L O G Y , V O L U M E 4 4
TABLE 4. Diaspore spectrum from the tuf®te; *, seeds plus fragments of three adult and two juvenile cones.
diaspore taxa number %
Sequoia abietina 45* 44´4Manglietia germanica 9 10Distylium uralense 3 3´3Hamamelidaceae gen. et sp. indet. 1 1´1Poliothyrsis eurorimosa 4 4´4Rubus spp. 4 4´4Nyssa sp. 1 1´1Ampelopsis malvaeformis 2 2´2Ampelopsis rotundata 1 1´1Carex sp. 4 4´4Sparganium spp. (ex gr. elongatum-pusilloides-camenzianum) 17 18´9Carpolithus div. gen. et sp. 4 4´4total number of specimens/percentage 90 99´7

(Text-®gs 4±5). This is underlined by the prevailing trend of the transport directions from the north, north-west, west, and south-west of the underlying and surrounding clastic host sediments, which is in obviouscontrast to the presumably (south-) eastern source of the tephra.
Actuopalaeontological observations at El Chichon (Mexico) indicated that various factors, such asgrain size and parameters of atmospherical condition, contributed to form a variety of ash-fall depositsand to the preservation of different vegetational elements (Spicer 1989, p. 166). Several features indicatethat a volcanic eruption in¯uenced the composition of the different plant organ assemblages in theOberdorf tuf®te. Normally-graded lapilli are embedded in backswamp sediments and associated with plantremains there. On the one hand the plant organ assemblages consist of well-preserved pollen clusters,pollen, buds, bud scales, and cuticles of evergreen foliage, and on the other hand of poorly preservedpollen, herbaceous monocotyledons, cuticles of deciduous foliage, diaspores, and strongly degradedorganic matter.
Well-preserved plant organs
The clusters of obviously still immature but often well-preserved pollen of wind-pollinated taxa(Taxodiaceae, Alnus, Engelhardia, Fagus, Lithocarpus, Quercus, Trigonobalanopsis, and Poaceae),often still attached to anther tissue, do not re¯ect natural pollen dispersal. Their presence, as well as theabundance of pollen clusters of insect-pollinated taxa, point to a catastrophic event. However, no catkinsor other ¯oral structures have been discovered.
A volcanic eruption probably coincided with the season in which certain plant taxa such as Taxodiaceae,Alnus, Engelhardia, Fagus, Ilex, Lithocarpus, Quercus, Trigonobalanopsis, Poaceae, and Sparganium/Typha produced male ¯owers (Table 5). The abundance of well-preserved buds/bud scales of ?Alnus andthe pollen clusters perhaps indicate the beginning of the growing season when deciduous trees could stillhave been lea¯ess or in the state of opening their buds. This may explain why evergreen foliage (mainlyzonal ones) prevails qualitatively and quantitatively over deciduous (mainly azonal ones). Unfortunately,we cannot de®nitively conclude whether this predominance was caused by the scenario considered aboveor by selective taphonomic processes. However, the good cuticle preservation of evergreen foliage
K O V A R - E D E R E T A L . : M I O C E N E P L A N T S 585
Oberdorf tuf®tefacts interpretation
locally normally-graded lapilli in silty/clayey matrix,sharp contact to the over- and underlying sediments
a result of volcanic eruptiondirect airfall mechanismthe lapilli dropped into backswamp sediments
probable primary chemistry of the lapilli rhyolitic/rhyodacitic
source area: Pannonian Basinprobable correlation with the `Lower Rhyolite Tuffs'indicating a (Late) Eggenburgian/Early Ottnangian age
well-preserved cuticles of evegreen foliage, buds/budscales of ?Alnus sp.; pollen, clusters of juvenile pollenattached to anther tissue
eruption during ¯owering time: `springtime' deciduouswoody plants still lea¯essevergreen foliage, parts of ¯owers, buds/bud scalesripped off their parent plants and embedded quickly inthe backswamp sediment that contained alreadydecayed plant material
poorly preserved cuticles of deciduous foliage, herbac-eous monocotyledons; diaspores, pollen, dispersedorganic matter
deposition prior to the volcanic eruptiondeciduous foliage and diaspores were not yet developedat the time of the eruption; therefore, no well-preservedremains
TEXT-FIG. 4. Summary of facts and interpretations.

contrasts strongly with that of deciduous woody species and the cuticle fragments observed in the organicfacies (see below).
Poorly preserved plant remains
The monocotyledonous leaf fragments, as well as the questionable and rarer leaf fragments of deciduouswoody plants such as ?Alnus/Betulaceae, ?Salix, and ?Fraxinus, and the diaspores, were already depositedprior to the eruption and had already undergone some decay. The organic debris together with the poorlypreserved pollen are also thought to have been deposited within the surface layer of the backswampsediments earlier than the pollen clusters and the other well-preserved plant material.
Summarizing, we propose that when the lapilli and ash dropped directly into the backswamp sediments,the water-saturated and therefore soft surface layer of the sediment was incorporated, ®nally forming thematrix of the tuf®te. This `contamination' with a non-volcanic matrix and `old', poorly preserved plantmaterial contrasts strongly with the presumed syn-volcanically deposited, well-preserved pollen clustersand pollen, buds, bud scales, and evergreen foliage. At the time of the eruption, mature deciduous leavesand diaspores were perhaps not yet available to be deposited in the tuf®te.
586 P A L A E O N T O L O G Y , V O L U M E 4 4
TABLE 5. Flowering, leaf-¯ush and seed dispersal time of extant species (from Schopmeyer 1975; KruÈssmann 1976±1978; Nooteboom 1985; Tiner 1993; Wu and Raven 1999). Genera were selected based on the occurrence of pollenclusters, abundant leaves and fruits in the tuf®te. Roman numerals indicate the months of ¯owering/seed maturity/dispersal.
seed maturity/plant name origin ¯owering time leaf-¯ush time dispersal
Alnus formosana China, Asia V-VI deciduous ±/VII-IXAlnus serrulata USA II-IV deciduous ±/IX-XCleyera japonica Himalaya, Japan VI-VII evergreen ±Engelhardia roxburghiana China II-IV semi-evergreen I-XII/±Engelhardia spicata China XI-IV evergreen/deciduous I-XI/±Distylium myricoides China IV-V evergreen ±Distylium racemosum Japan III-IV evergreen ±/I-XIIFagus grandifolia USA III-V deciduous IX-XI/after frostFagus spp. China IV-V deciduous VIII-X/±Gordonia lasianthus USA V-VIII evergreen autumn/±Gordonia axillaris China, Taiwan XI-V evergreen ±Glyptostrobus pensilis China I-III deciduous IX-X/±Ilex latifolia Japan late spring evergreen ±/late autumnIlex vomitoria USA III-V evergreen autumn/±Laurus azorica Canary Islands IV evergreen ±Laurus nobilis Asia Minor III-VI evergreen autumn/±Lithocarpus densi¯orus W USA spring, summer, fall evergreen ±/autumnLithocarpus frenzilianus China II-IV evergreen ±/VIII-XIIManglietia glauca SE Asia I-XII evergreen I-XII/±Pinus elliottii USA II-III evergreen ±/XPinus roxburghii India II-IV evergreen ±/IV-VQuercus spp. USA II-early summer deciduous/evergreen ±Quercus spp. China III-IV deciduous IX-X/next yearQuercus spp. China V-VI evergreen IX-X/next yearSequoia sempervirens USA II evergreen ±/autumnSparganium americanum USA V-VIII perennial ±Sparganium simplex China IV-VI perennial VI-VII/±Trigonobalanus doichangensis China, Thailand XI evergreen III

C O M P A R I S O N W I T H O T H E R P L A N T - B E A R I N G V O L C A N I C L A S T I C D E P O S I T S
This multidisciplinary approach to unravel the event of the Oberdorf tuf®te deposition yielded a widerange of data. However, we were unable to ®nd any similar volcaniclastic deposit for comparison. Moststudies are con®ned to only one of the different plant organ assemblages (leaves, diaspores, petri®edwoods, pollen) or sedimentology. Various plant organs are not always preserved. Tuffs have been reportedto yield plant remains, e.g. in Ipolytarnoc, Lipovany, Stare Bystre; and some plant assemblages derivedfrom tuffs/tuf®tes of maar deposits e.g. `Hammerunterwiesenthal'. Based on the stratigraphic correlations,the tuf®tes from Oberdorf may have resulted from the eruptive cycle (LRT) that produced the tuffs ofIpolytarnoc and Lipovany.
Ipolytarnoc (north of Budapest on the Slovakian border, Hungary; Gyulaleskzi Rhyolite Tuff Formation,Lower Rhyolite Tuffs, Early Miocene, Eggenburgian/Ottnangian; Hably 1983).
The relatively widespread 0´2±0´4-m-thick tuf®tic volcaniclastics belong to the lowermost unit of theGyulaleskzi Rhyolite Tuff Formation. In contrast to Oberdorf the leaves are preserved mostly as isolated,gross-morphologically well-preserved impressions, and even some ¯owers have been described. The leaf¯ora is a result of autochthonous burial of litter from the forest growing on sands directly underlying thetuffs. The leaf-bearing parts of the tuffs do not yield lapilli as in Oberdorf. Information aboutpalynomorphs does not exist and diaspores (except Leguminocarpon and a pine cone) are not preserved(L. Hably, pers. comm. 2000).
Lipovany (SSE of Lucenec, South Slovakia at the Hungarian border, Bukovinka Formation, EarlyMiocene, Late Eggenburgian; Sitar and KvacÏek 1997). The plant-bearing volcaniclastics yielding the
K O V A R - E D E R E T A L . : M I O C E N E P L A N T S 587
TEXT-FIG. 5. Possible scenario that led to the formation of the plant assemblage recovered. 1, buds, bud scales; 2, pollen,pollen clusters; 3, leaves of evergreen taxa.

¯ora of Lipovany are closely related to those of Ipolytarnoc, as they are not only geographically close butbelong to the same volcanic complex. The 1´5-m-thick tuf®tic sediments investigated by NeÆmejc andKnobloch (1973, p. 694) included a supposed lens-shaped plant-bearing layer of 0´19±0´5 m thick. Thegood gross-morphological leaf preservation resembles that of Ipolytarnoc and, therefore, differs fromOberdorf. In Lipovany, leaves are sometimes carbonized, but cannot be macerated because they arefusinized.
Stare Bystre (Nowy Targ-Orawa Basin, Poland, Late Miocene; Worobiec 1994). A tuff/tuf®te horizon (1±2 m thick) is intercalated in ¯uvial deposits, in this respect resembling the situation in Oberdorf. From thishorizon an assemblage of mostly fragmented leaves was interpreted to be the litter of a ¯oodplain forest.Diaspores have not been found and palynological investigations have not been carried out in thepyroclastic sediments at Stare Bystre (G. Worobiec, pers. comm. 1999).
Hammerunterwiesenthal (Sachsen, Germany, Early Oligocene; Walther 1998). Within a maar of about2000 m in diameter, a 5´5-m-thick sequence of basaltic tuf®tes includes a 0´3-m-thick layer of a ®nelylaminated tuf®te rich in plant remains. The carbonized plant material and the pollen show signs of heating.As a result, cuticle preparations of the leaves were unsuccessful. All of these features differ from thematerial from Oberdorf. Although the leaves are better preserved in gross morphology than at Oberdorf,the splitting of the sediment for leaves entailed similar dif®culties (Walther 1998, p. 240). Only leavesallow botanical determinations in Hammerunterwiesental; the preservation of diaspores and pollen is toopoor. The ¯oristic spectrum is indicative of mixed mesophytic forests.
C O N C L U S I O N S
1. An approximately 0´04±0´08-m-thick tuf®te layer was found within backswamp sediments of theOberdorf opencast mine. Locally well-developed, normally-graded lapilli and sharp contacts with theunder- and overlying clastic sediments indicate a direct fall-out mechanism of volcanic tephra. Lapilli sizeranges from 1±8 mm in diameter. Based on its geochemical composition and the age of micro-mammal-bearing strata in Oberdorf, the tephra can only be correlated to the `Lower Rhyolite Tuffs' in thePannonian Basin (Hungary), indicating a (Late) Eggenburgian/Early Ottnangian age.2. The tuf®te contains well-preserved plant remains: cuticles of evergreen leaf compressions; pollen (proparte); pollen clusters partly attached to anther tissue of both wind- and insect-pollinatedplants (Taxodiaceae, Alnus, Engelhardia, Fagus, Ilex, Lithocarpus, Quercus, Trigonobalanopsis, andSparganium/Typha); buds, and bud scales. This contrasts strongly with the poorly preserved plant remains:cuticles of deciduous taxa, herbaceous monocotyledons, palynomorphs (pro parte), diaspores, andstrongly decomposed organic facies components.3. In the tuf®te, taxa of zonal versus azonal habitats are better documented in the leaf than in the diasporerecord, thus contrasting with all other Oberdorf samples. Foliage of Distylium fergusonii and Lauro-phyllum rugatum is recorded for the ®rst time from Austria, and Ternstroemites waltheri is a new species.All three are considered to be mesophytic elements.4. The palynomorph assemblage represents a mixture of different habitats known from Oberdorf.Unlike the generally more facies-biased assemblages from Oberdorf, the tuf®te assemblage is moretaphonomically biased.5. The diaspore assemblage is poor in both diversity and state of preservation, while buds and bud scales of?Alnus sp. are very well preserved and numerous.6. The diaspore and palynomorph assemblages contain only taxa already known from Oberdorf.7. The following interpretation is proposed: The fall-out of volcanic lapilli and ash occurred at a time whenmany woody plants were ¯owering and most of the deciduous ones were lea¯ess or at the stage of openingtheir buds. The volcanic eruption caused the evergreen leaves, fragments of male ¯oral structures (pollenclusters), and buds to be ripped off and deposited in the backswamp together with ash and lapilli. Otherplant material had already accumulated in the backswamp before the tephra fall-out, such as diaspores,
588 P A L A E O N T O L O G Y , V O L U M E 4 4

palynomorphs, leaf fragments of deciduous woody plants, and organic debris. Because they had alreadyundergone decomposition, a curious assemblage of well- and poorly preserved plant material is nowrecovered from the tuf®te.
S Y S T E M A T I C P A L A E O N T O L O G Y
We restrict this section to new species in the Oberdorf record.
Family HAMAMELIDACEAE Brown, in Abel, 1818
Genus DISTYLIUM Siebold and Zuccarini, 1841
Distylium fergusonii Knobloch and KvacÏek, 1976
Plate 1, ®gures 1±2; Plate 3, ®gures 7±8
1971 XXIX; Ferguson, p. 159, pl. 23; text-®g. 28A±D.1976 Distylium fergusonii sp. nov.; Knobloch and KvacÏek 1976, p. 45, pl. 17, ®gs 1, 3, 20; pl. 20, ®gs 1, 7,
13±14; pl. 22, ®gs 3±8; text-®g. 18.
Material. Collection nos 1998B0005/1A, F, 2E, G, 6E, 9B, 18B, 20A, 44.
Description. Fragments of ovate to elliptical leaves (specimens 1A, 2G, 9B), best-preserved specimens long/wide: 45/>35 mm (1A), 45/25 mm (9B), 35/23 mm (2G); apex acuminate (20A), base acute (2G), lamina slightly asymmetrical(2G); leaf margin entire, in specimen 20A ?one tooth present close to the apex; midvein straight, slightly curved,secondaries thin, widely spaced (1A, 9B).
Adaxial and abaxial cuticular membranes moderately thick; cuticular ¯anges of the non-modi®ed epidermal cellsstrongly omega-shaped undulate, forming cell outlines of extremely irregular shape that appear strongly intermeshed;size 30±60 mm; elongated cell outlines upon the veins, undulation of the cuticular ¯anges there with smalleramplitudes, abaxially often changing into pitted cuticular ¯anges, abaxially distinct striation present; trichomebases very prominent, more common abaxially than adaxially, star-like in shape, elevated, built of short and thickcells; poral rim extremely thickened, porus diameter 8±15 mm, striation around trichome bases sometimes present;stomatal complexes con®ned to the abaxial cuticular membrane, inconspicuous, coarsely and irregularly scattered,paracytic; subsidiary cells indistinct, stomata oval, small 10±20 (25) mm long, outer front cavity (sometimes wide)oval, 5±10 mm long.
Remarks. This is the ®rst record of D. fergusonii from Austria. Particularly the stomata and trichome basedensity match better the leaves from Kreuzau than those from Oder 2a, Oberpfalz (Knobloch and KvacÏek1976, p. 45). Considering the trichome base density alone, the similarity is also close to the leaf remainsfrom the Cypris Shale (BuÊzÏek et al. 1996, p. 23). The length of the stomata apparently varies considerablyin all localities: Oder 2a 20±22 (28) mm, Kreuzau 15±37 mm, Haselbach 19±26 mm (Mai and Walther1978, p. 53). In Oberdorf, they are partly even smaller [10±20 (25) mm]. However, BuÊzÏek et al. (1996,p. 23) also mentioned one specimen with smaller stomata from the Cypris Shale.
Distylium heinickei Walther (Mai and Walther 1991, p. 46) differs in having single teeth in the apicalthird of the lamina and a lower density of trichome bases. Within extant species of Distylium the leafserration is variable; teeth may be absent or present in the same species (Walker 1944, p. 321). The densityof trichome bases is variable in extant species and in D. fergusonii. Because only rather tiny cuticlefragments of Distylium are usually available in the fossil record, the original hair base presence/density isdif®cult to estimate, especially when they are scattered.
Herbarium material investigatedDistylium chinense (Fr.) Diels, Handel-Mazzetti, Iter sinense 1914±18, Nr. 10692, Kweitschou, leg. 11.7.1917, WU
[� D. buxifolium (Hance) Merr.]Distylium chinense Handel-Mazzetti, Iter sinense 1914±18, Nr. 10272, Prov. Kweitschou, ad viam Tschenning-
Huang-tsauba-YuÈnnan, alt. 1250 m, leg. 14.6.17, WU [� D. buxifolium (Hance) Merr.]
K O V A R - E D E R E T A L . : M I O C E N E P L A N T S 589

Distylium guatemalense Radlkofer sp. nov., Flora v. Guatemala, Coban, Coll. H. v. TuÈrckheim 1910, WDistylium myricoides Hemst., Plants of Anhwei Province, China, Coll. C. S. Fan and Y. Y. Li No. 128, 17.6.1936, WDistylium racemosum (Fr.) Diels, Handel-Mazzetti, Iter sinense 1914±18, Nr. 10810, Prov. Kweitschou, leg.
17.7.1919, WUDistylium racemosum S. and Z., Prov. Osumi, Japan, March 13, 1891, WUDistylium racemosum S. and Z., ex herb. horti bot. Petropolitani, Maximowicz; iter secundum, Japonia Nagasaki,
1863, WDistylium tsiangii Chun sp. nov., Waichai, Tuhshan, South-Kweichow, Border of Kwangsi, Y.Tsiang No. 6692, det.
W. Y. Chun, Coll. 25.8.1939, W
Family LAURACEAE Jussieu de, 1789
Genus LAUROPHYLLUM Goeppert, 1854
Laurophyllum rugatum KvacÏek and BuÊzÏek, 1966
Plate 1, ®gures 3±7; Plate 2, ®gure 8; Plate 3, ®gure 5
1966 Laurophyllum rugatum sp. nov.; KvacÏek and BuÊzÏek, p. 292, pl. 1, ®gs 3±5; pl. 4, ®g. 4.
Material. Collection nos 1998B0005/25, 34A.
Description of fragment 34A. No gross-morphological features preserved. Cuticular membranes of both surfaces thick;adaxial one thicker than abaxial one, lense-shaped secretory bodies present, diameter 16±54 mm; adaxial cuticularmembrane: anticlinal walls of the non-modi®ed epidermal cells thick, almost straight, forming irregularly polygonalcell outlines, size 12±24 mm; occasionally simple trichome bases present upon veins, consisting of an irregularpolygonal pore, 4-5 mm in diameter, surrounded by a somewhat thickened poral rim; abaxial cuticular membrane:anticlinal walls slender, but clearly visible, straight to slightly curved; size of non-modi®ed epidermal cells 11±22 mm,most of them border one to several trichome bases and stomatal complexes; stomatal complexes densely spaced,rounded, oval, oblique or rhomboidal, paracytic; the subsidiary cells bear thickened epidermal walls and largely coverthe guard cells; stomatal ledges/poral walls of the subsidiary cells distinct, reaching the stomatal poles; length ofstomatal complexes 12±22 mm, width (11) 15±22 (27) mm; trichome bases very dense, sometimes even joining eachother, situated at the joins of 4±6 (8) cells, bordered by very thick and high poral rim/trichome foot remains, innerdiameter of trichome pores 2±5 mm, external diameter of the trichome bases (poral rim included) 8±12 mm; faintstriation upon veins sometimes present.
Description of fragment 25. 65 mm (completed probably $ 100 mm) long, $55 mm wide, shape ?elliptical, wideelliptical or somewhat obovate, neither apex nor base preserved, entire margined; venation brochidodromous, midveinstraight, secondaries arising in distances of up to 20 mm (in the central part of the lamina), running in wide curvesacross the lamina, one of them bifurcating; secondaries forming loops; intersecondaries present; ab- and adaxialcuticular membranes thick, adaxially thicker than abaxially; secretory bodies present, 37±58 mm in diameter; adaxial
590 P A L A E O N T O L O G Y , V O L U M E 4 4
E X P L A N A T I O N O F P L A T E 1
Figs 1±2. Distylium fergusonii Knobloch and KvacÏek, 1976. Abaxial cuticular membrane, interference contrast. 1,characteristically prominent trichome base above vein, coll. no. 1998B0005/0044/2; ´ 200. 2, paracytic stomatalcomplexes, strong undulation of non-modi®ed epidermal cells, striation, coll. no. 1998B0005/0044/1; ´ 400.
Figs 3±7. Laurophyllum rugatum KvacÏek and BuÊzÏek, 1966. 3, abaxial cuticular membrane, trichome bases veryabundant, coll. no. 1998B0005/0034A/1, interference contrast; ´ 400. 4, abaxial cuticular membrane, rhomboidalstomatal complexes with thickened, well-stained epidermal walls of subsidiary cells, trichome bases scarce, coll. no.1998B0005/0025, interference contrast; ´ 400. 5, abaxial cuticular membrane, stomatal complexes, trichome base,coll. no. 1998B0005/0025, SEM, internal view; ´ 900. 6, faint striation, wrinkles around trichome pore, above veinor leaf margin, coll. no. 1998B0005/0025, SEM, external view; ´ 900. 7, faint striation, trichome pore, above vein,coll. no. 1998B0005/0025, SEM, external view; ´ 400.
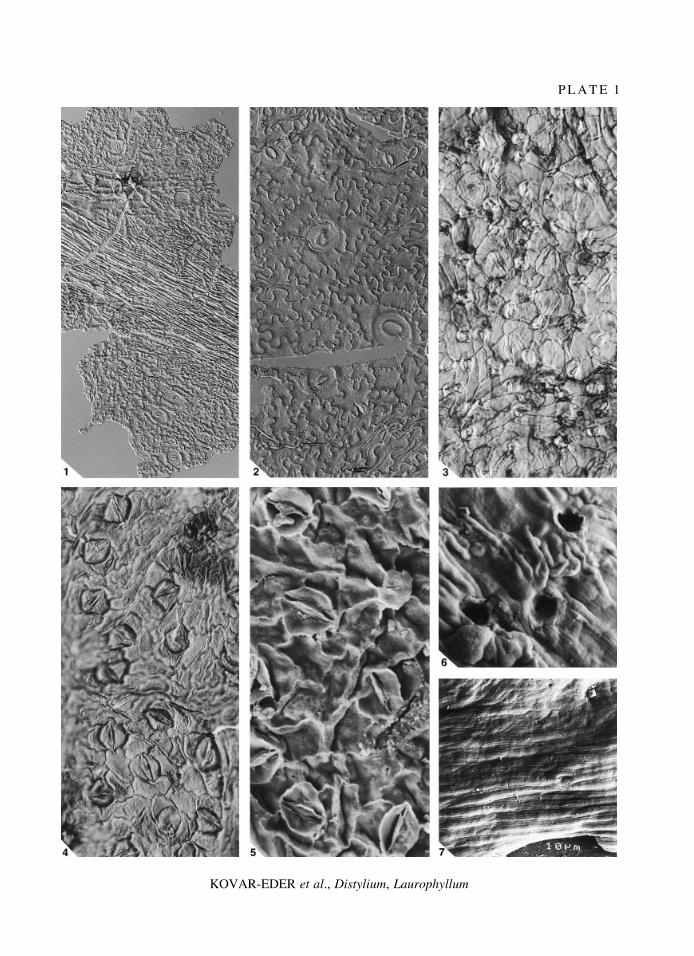
P L A T E 1
KOVAR-EDER et al., Distylium, Laurophyllum

cuticular membrane: non-modi®ed epidermal cells 18±24 (32) mm in size, anticlinal walls thick, straight to somewhatwavy; simple trichome bases scarce, appearing as small, irregular star-like openings between adjacent cells, sometimessurrounded by faint, radial cuticular striation, upon veins parallel striation, wrinkles around trichome bases; abaxialcuticular membrane: non-modi®ed epidermal cells (17) 19±28 (37) mm in size, anticlinal walls wavy, not alwaysclearly traceable, more delicate than those on the upper leaf surface; cell surfaces centrally thickened and thereforedeeper staining; stomatal complexes densely spaced, paracytic, subsidiary cells mostly triangular, giving the stomatalcomplexes a rhomboidal but asymmetrical outline; subsidiary cells with often distinctly thickened, epidermal wallslargely cover the guard cells, stomatal ledges/poral walls of the subsidiary cells thickened, reaching the stomatal poles;length of stomatal complexes 18±22 (26) mm, width 16±22 mm; scattered simple trichome bases with thickened poralrim and basal remains of the trichome still attached, more common upon veins, but present also in the intercostal areas;trichome porus of irregular 5±8-sided shape, inner diameter 7±10 mm.
Remarks. This is the ®rst record of L. rugatum from the Paratethys region. It is already well known fromNorth and South Bohemia, Saxony, Leipzig, and the Oberpfalz area. L. rugatum is characterized by theconsistent cuticular features with high trichome density on the abaxial leaf surface as well as their scarcityon the adaxial. However, in specimen 25 the indumentum of the lower leaf surface is far less dense thanusual. Nonetheless, stomatal shape and size, the thickened epidermal walls of the subsidiary cells, theirthickened poral walls/stomatal ledges reaching the stomatal poles, and the faint striation on the upper leafsurface (see KvacÏek 1971, p. 62; Pl. 1, ®g. 7 herein) are distinct features of this taxon.
Family THEACEAE Don, 1825
Genus TERNSTROEMITES Berry, 1916
Ternstroemites waltheri Kovar-Eder sp. nov.
Plate 2, ®gures 1±7; Plate 3, ®gure 6
Derivation of name. In honour of Harald Walther, who contributed to detailed cuticular studies of Palaeogene/Neogeneplants and recently celebrated his 70th birthday.
Holotype. Collection no. 1998B0005/34B.
Other material. Collection nos 1998B0005/6A, 10B=10C, 43; all paratypes. Collection nos 1998B0005/19A, 50D.
Locality and horizon. Tuf®te at the top of the footwall/base of the main seam in the eastern sub-basin of the Oberdorfopencast mine, Koȯach/Voitsberg Formation. This tuf®te is correlated with the `Lower Rhyolite Tuffs' in thePannonian Basin, indicating a (Late) Eggenburgian/Ottnangian (Early Miocene) age.
Diagnosis. Ad- and abaxial cuticular membranes thick, simple trichome bases rare, with radial striation;
592 P A L A E O N T O L O G Y , V O L U M E 4 4
E X P L A N A T I O N O F P L A T E 2
Figs 1±7. Ternstroemites waltheri sp. nov. 1±5, coll. no. 1998B0005/0034B, holotype. 1, abaxial cuticular membrane,stomata surrounded by a narrow ring of somewhat thickened subsidiary cells, parallel striation, interference contrast;´ 400. 2, abaxial cuticular membrane, parallel striation, unequally developed, SEM, external view; ´ 400. 3, abaxialcuticular membrane, stomatal complex, epidermal walls of guard cells and polar I-shaped thickenings more or lessdistinctly developed, SEM, internal view; ´ 800. 4, adaxial cuticular membrane, interference contrast; ´ 200. 5,adaxial cuticular membrane, SEM, internal view; ´ 400. 6±7, adaxial cuticular membrane, strong, parallel striation,coll. no. 1998B0005/0010B�C, paratype. 6, interference contrast; ´ 400. 7, SEM, external view; ´ 600.
Fig. 8. Laurophyllum rugatum KvacÏek and BuÊzÏek, 1966; adaxial cuticular membrane, coll. no. 1998B0005/0025,SEM, internal view; ´ 600.
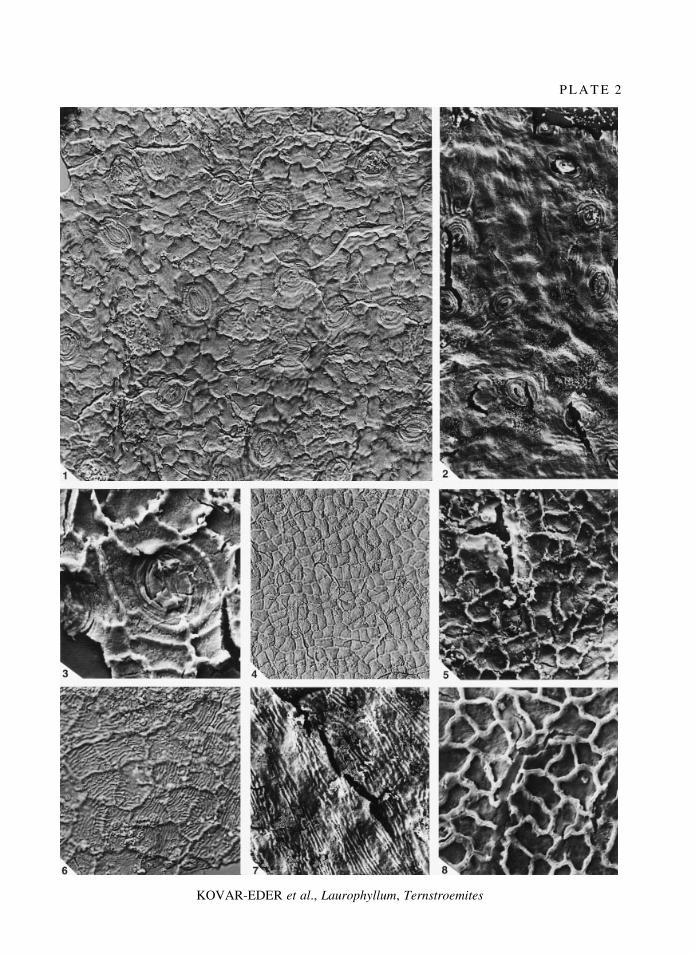
P L A T E 2
KOVAR-EDER et al., Laurophyllum, Ternstroemites

adaxial cuticular membrane: size of non-modi®ed epidermal cells 20±27 (42) mm, their anticlinal wallssmooth, slightly wavy, intensity of cuticular striation variable; abaxial cuticular membrane: size of non-modi®ed epidermal cells 22±38 mm, anticlinal walls U-shaped undulating, smooth; stomatal complexes ofthe Gordonia-type: subsidiary cells short, occasionally tangentially elongated, forming one and sometimesan incomplete second ring around the stomata; stomatal size (13) 16±22 (26) mm long, (12) 16±20 (21) mmwide; epidermal walls of guard cells more or less distinct, outer cuticular ledges thickened and smooth,enclosing the (wide) oval to spindle-shaped outer front cavity; I-shaped polar thickenings sometimespresent; cuticular striation of variable intensity running in bundles parallel over longer distances, butaround larger stomata appearing more radially orientated.
Description. Leaf fragments largely lacking diagnostic gross-morphological features; specimen 43 is a 30-mm-longfragment, from the midvein to the margin 18 mm wide, complete size of this leaf unknown, leaf margin unclear;cuticular membranes of all leaf fragments preserved; ad- and abaxial ones thick; simple trichome bases rare, withthickened poral rim and radial striation, present adaxially, ?present abaxially; adaxial cuticular membrane: size of non-modi®ed epidermal cells 20±27 (42) mm, their cuticular ¯anges smooth, slightly wavy, but less distinctly undulatingthan on the lower surface; cuticular striation sometimes strong, sometimes indistinct to almost missing; abaxialcuticular membrane: non-modi®ed epidermal cells 22±38 mm in size, their anticlinal walls U-shaped undulating,smooth; stomatal complexes of the Gordonia-type: cyclocytic, sometimes not only one but an incomplete second ringof subsidiary cells is developed; subsidiary cells mostly short but occasionally tangentially elongated, more stronglycutinized than the non-modi®ed epidermal cells; size of stomata variable, (13) 16±22 (26) mm long, (12) 16±20 (21)mm wide, guard cells with thickened and smooth outer cuticular ledges enclosing the oval to wide-oval, sometimesspindle-shaped outer front cavity; epidermal walls of guard cells more or less distinct; polar I-shaped thickenings(more or less well developed) sometimes present, outer surface with cuticular striation of varying intensity (distinct tosometimes faint to even almost missing) within one specimen, running in bundles parallel over longer distances;around larger stomata the striation appears more radially developed.
Distinctions and comparisons. ?Gordonia oberdorfensis Kovar-Eder (Kovar-Eder and Meller in press b)differs in having larger, suborbicular to orbicular stomata [diameter 24±(30) 37 mm], less distinctepidermal walls of the guard cells and anticlinal walls of the subsidiary cells, more concentric striationaround the stomata, and more common trichome bases.
?Cleyera schilcheriana Kovar-Eder differs in having larger non-modi®ed epidermal cells on both leafsurfaces [abaxially (31) 36±60 (70) mm, adaxially 37±60 mm], larger stomata [stomatal length 24±31 mm
594 P A L A E O N T O L O G Y , V O L U M E 4 4
E X P L A N A T I O N O F P L A T E 3
Fig. 1. Laurophyllum pseudoprinceps Weyland and Kilpper, 1963; abaxial cuticular membrane, coll. no. 1998B0005/0047/1, interference contrast; ´ 400.
Fig. 2. Daphnogene polymorpha (A. Braun, 1845) Ettingshausen, 1851; abaxial cuticular membrane, coll. no.1998B0005/0062B/1, interference contrast; ´ 200.
Fig. 3. Trigonobalanopsis rhamnoides (RossmaÈssler, 1840) KvacÏek and Walther, 1988; abaxial cuticular membrane,coll. no. 1998B0005/0001H/1, interference contrast; ´ 400.
Fig. 4. Magnolia liblarensis (KraÈusel and Weyland, 1959) KvacÏek, 1978; abaxial cuticular membrane, trichome basesrare; coll. no. 1998B0005/0001E/1, interference contrast; ´ 200.
Fig. 5. Laurophyllum rugatum KvacÏek and BuÊzÏek, 1966; coll. no. 1998B0005/0025.Fig. 6. Ternstroemites waltheri sp. nov.; coll. no. 1998B0005/0043, paratype.Figs 7±8. Distylium fergusonii Knobloch and KvacÏek, 1976. 7, coll. no. 1998B0005/0001A. 8, coll. no. 1998B0005/
0020A.Fig. 9. Trigonobalanopsis rhamnoides (RossmaÈssler, 1840) KvacÏek and Walther, 1988; coll. no. 1998B0005/0026.Fig. 10. Monocotyledoneae gen. et sp. indet.; coll. no. 1998B0005/0010A.Fig. 11. Surface view of a specimen showing lapilli embedded in the backswamp matrix and leaf fragments of (A)
Trigonobalanopsis rhamnoides and (B) Daphnogene polymorpha, coll. no. 1998B0005/0024.Fig. 12. Vertical section of a specimen showing the distinct layer of lapilli in the clayey/silty backswamp matrix.All ´ 1 except where indicated for 1±4.
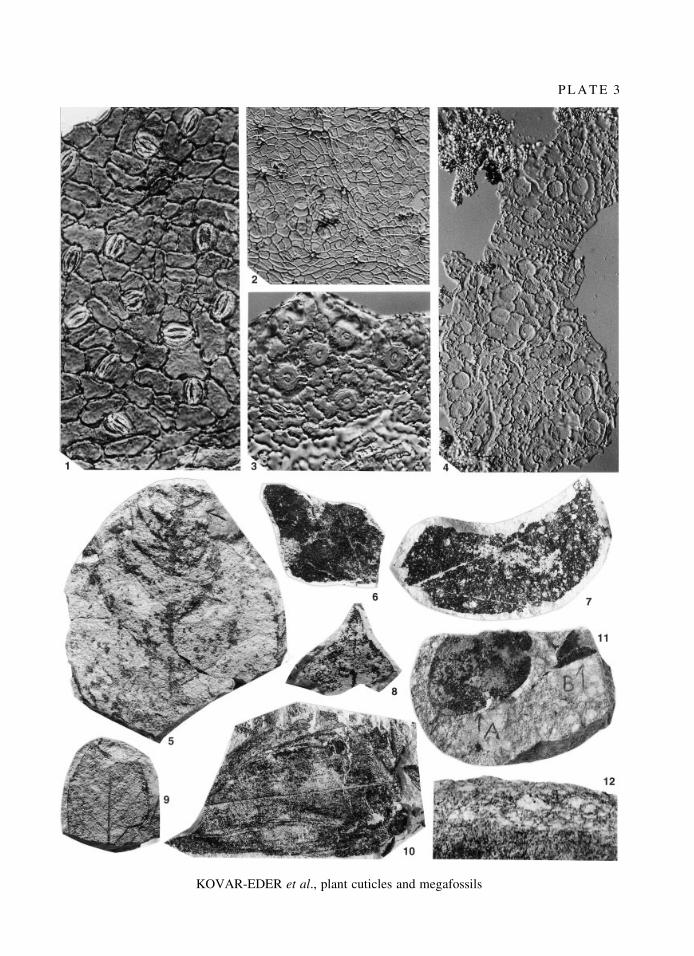
P L A T E 3
KOVAR-EDER et al., plant cuticles and megafossils

(42 mm in giant ones), stomatal width (19) 22±29 mm]; more strongly thickened outer cuticular ledges ofthe guard cells, tangentially elongated subsidiary cells, much stronger and more wrinkle-like striations,and globoid organic structures attached to the inner surface of the adaxial cuticular membrane.
Ternstroemites sokolovensis KvacÏek and Walther (1984, p. 339; Late Eocene) and Ternstroemitesbockwitzensis Walther (Mai and Walther 1991, p. 77; Late Oligocene) are both glabrous and differ by strongerundulation of the anticlinal walls of the non-modi®ed epidermal cells on the adaxial cuticular membrane.
Theaceae gen. et sp. indet. Walther (1999, p. 123) differs in having signi®cantly larger stomata [stomatallength 35´6 mm (32±40 mm), stomatal width 24 mm (20±36 mm)], and trichome bases have not beenreported (H. Walther, pers. comm. 2000).
Recent descriptions of Theaceae leaves (Bozukov and Palamarev 1995; Givulescu et al. 1997) are basedexclusively on gross morphology, which is largely not preserved in our remains. Therefore, a comparisonis not possible.
Remarks. Because a de®nitive assignment to one living genus within the Theaceae is not possible, theseremains have to be referred to Ternstroemites (Berry 1916). The complex of cuticular features pointstowards Cleyera and Gordonia (including Polyspora). However, in the former the epidermal walls of theguard cells are thickened, this feature being less distinct in the fossil remains. In Gordonia sensu stricto nopolar thickenings are developed but the striation may be strong. In former Polyspora (now included inGordonia) polar thickenings of variable shape are developed but the striation is more concentric aroundthe stomata and usually does not affect the non-modi®ed cells as in the fossil remains described here.
Family ?BETULACEAE Gray, 1821
Genus ALNUS Miller, 1754
?Alnus sp.
Text-®gure 3F±G
Material. Collection no. 1999B0053/12 (about 50 specimens).
Description. Buds and bud scales; buds up to 11 mm long and 6 mm wide, length/width ratio mostly (1´2) 1´5±1´8;shape almost triangular, base straight, apex acute, sides straight to slightly convex.
Remarks. The buds most probably represent one taxon and the scales clearly derive from these buds. Theycompare with Alnus (Godet 1995, p. 140).
Acknowledgements. These investigations were carried out within the framework of the projects 10337-GEO and10339-GEO ®nanced by the Austrian Science Fund. Cuticle comparisons were possible thanks to the collection of Z.
596 P A L A E O N T O L O G Y , V O L U M E 4 4
E X P L A N A T I O N O F P L A T E 4
Figs 1±2. Taxodiaceae pollen cluster. 1, juvenile intact pollen de¯ated and adpressed, SEM; ´ 620. 2, detail, de¯atedpollen with visible papillus, SEM; ´ 3200.
Figs 3±5. Myrica sp. 3, small pollen cluster; individual grains adpressed, LM; ´ 900. 4, pollen cluster; pollen grainsde¯ated and partly covered by small ¯akes of anther material, SEM; ´ 1200. 5, detail of pollen grain with fullydeveloped micromorphological surface features, SEM; ´ 10,500.
Figs 6±8. Fagus sp. 6, pollen cluster; immature pollen grains de¯ated, in polar view subtriangular, SEM; ´ 600. 7,detail of pollen cluster; surface micromorphology not fully developed, covered by small ¯akes of anther material,SEM; ´ 2700. 8, detail of pollen cluster; surface micromorphology not fully developed; small roundish globules(possibly Ubisch-bodies) adhere to the surface, SEM; ´ 7500.
All specimens coll. no. 1999B0054.
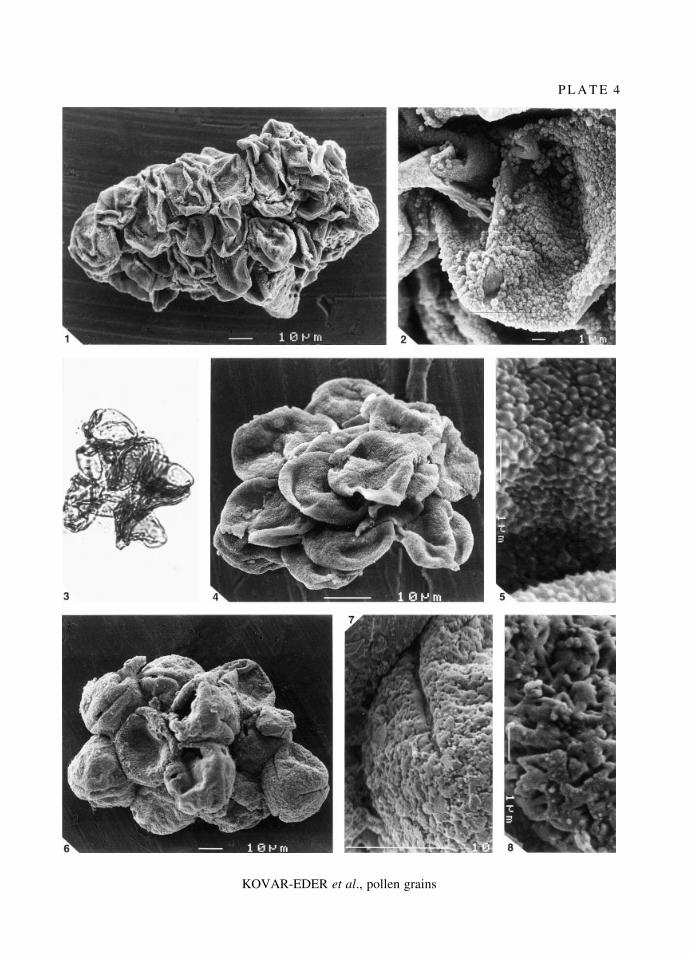
P L A T E 4
KOVAR-EDER et al., pollen grains

KvacÏek at the Palaeontological Department, Charles University, Prague. We are grateful to L. Hably (Budapest), Z.KvacÏek, H. Walther (Dresden), G. Worobiec (Krakow), and R. Zetter (Vienna) for discussions, and to Z. KvacÏek andH. Walther for critically reading the manuscript. The cuticle preparations were done by A. Taumberger. Thephotographs of the leaves and cuticles were provided by A. Schumacher.
R E F E R E N C E S
AÂ RVA-SOÂ S, E. and MAÂ THEÂ , Z. 1992. Mineralogical and petrographic study of some Neogene tuff layers of the Mecsekmountains (South Hungary) and their K/Ar dating. Acta Geologica Hungarica, 35, 177±192.
BERRY, E. W. 1916. The Lower Eocene ¯oras of southeastern North America. United States Geological Survey,Professional Paper, 91, 1±481.
BOZUKOV, V. and PALAMAREV, E. 1995. On the Tertiary history of the Theaceae in Bulgaria. Flora Mediterranea, 5,177±190.
BROWN, R. in ABEL, C. 1818. Narative of a journey in the interior of China. Longman, Hurst, London, 374±379.BUÊ ZÏ EK, C., HOLYÂ , F. and KVACÏ EK, Z. 1996. Early Miocene ¯ora of the Cypris shale (Western Bohemia). Acta Musei
Nationalis Pragae Series B., Historia Naturalis, 52, 1±72.DAXNER-HOÈ CK, G., HAAS, M., MELLER, B. and STEININGER, F. F. 1998. PalaÈooÈkologie, Sedimentologie und Stratigraphie.
195±224. In Wirbeltiere aus dem Unter-MiozaÈn des Lignit-Tagebaues Oberdorf (Weststeirisches Becken,OÈ sterreich). Annalen des Naturhistorischen Museums, 99/A, 1±224.
DILCHER, D. L. 1974. Approaches to the identi®cation of angiosperm leaf remains. The Botanical Review, 40 (1), 1±157.DON, D. 1825. Prodomus ¯orae nepalensis. J. Gale, London, 256 pp.DOWNES, H., PANTO, G., POKA, T., MATTEY, D. P. and GREENWOOD, P. B. 1995. Calc-alkaline volcanics of the Inner
Carpathian arc, northern Hungary: new geochemical and oxygen isotopic results. Acta Vulcanologica, 7, 29±41.EBNER, F., MALI, H., OBENHOLZER, J. H., VORTISCH, W. and WIESER, J. 1998. Pyroclastic deposits from the Middle Miocene
Stallhofen Formation. 425-428. In STEININGER, F. F. (ed.). The Early Miocene lignite deposit of Oberdorf N Voitsberg(Styria, Austria). Jahrbuch der Geologischen Bundesanstalt, 140, 395±496.
FERGUSON, D. K. 1971. The Miocene ¯ora of Kreuzau. Verhandelingen der Koninklijke Nederlandse Akademie vanWetenschappen, Afd. Natuurkunde, Tweede Reeks, 60 (1), 297 pp.
GIVULESCU, R., BARBU, O. and PETRESCU, I. 1997. Nouveaux taxa pour la ¯ore OligoceÁne de Roumanie. RomanianJournal Paleontology, 77, 15±20.
GODET, J.-D. 1995. Knospen und Zweige der einheimischen Baum- und Straucharten. Naturbuch-Verlag, Augsburg, 431pp.
GOEPPERT, H. R. 1854. Die TertiaÈr¯ora auf der Insel Java. C. W. Mieling, 'S Gravenhage, 170 pp.GRAY, S. F. 1821. A natural arrangement of British plants. London, 827 pp. and 757 pp.HAAS, M. 1998. Sedimentological investigations in the Early Miocene lignite opencast mine Oberdorf (N Voitsberg,
Styria, Austria). 413±424. In STEININGER, F. F. (ed.). The Early Miocene lignite deposit of Oberdorf N Voitsberg(Styria, Austria). Jahrbuch der Geologischen Bundesanstalt, 140, 395±496.
ÐÐ 1999. Die fruÈhmiozaÈnen Sedimente der Braunkohletagebaue Oberdorf und Zangtal - eine environmentanalytischeUntersuchung (N Voitsberg, Steiermark, OÈ sterreich). Unpublished PhD thesis, Formal-Naturwissenschaftliche FakultaÈtder UniversitaÈt Wien, Austria.
HABLY, L. 1983. Early Miocene plant fossils from Ipolytarnoc, N Hungary. Geologica Hungarica, Series Palaeonto-logica, 45, 77±255.
HAÂ MOR, G. 1979. K/Ar dating of Miocene pyroclastic rocks in Hungary. Annales GeÂologique Pays HelleÂnique, HorsSerie, 2, 491±500.
ÐÐ BALOGH, K. and RAVASZ-BARANYAI, L. 1978. Radiometric age of the Tertiary formations in North Hungary. AMagyar Allami FoÈldtani Intezet evi Jelentese, 1976, 61±76.
JUSSIEU, A. L., de 1789. Genera plantarum. V. Herissant, Paris, 498 pp.KLAUS, W. 1987. EinfuÈhrung in die PalaÈobotanik. Deuticke, Wien, 232 pp.KNOBLOCH, E. and KVACÏ EK, Z. 1976. MiozaÈne BlaÈtter¯oren vom Westrand der BoÈhmischen Masse. Rozpravy UÂ strÏedniho
U stavu Geologickeho, 42, 5±129.KOVAR-EDER, J. 1996. Eine bemerkenswerte BlaÈtter-Vergesellschaftung aus dem Tagebau Oberdorf bei Koȯach,
Steiermark (Unter-MiozaÈn). Mitteilungen der Abteilung fuÈr Geologie und Bergbau, Landesmuseum Joanneum, 54,147±171.
ÐÐ and MELLER, B. in press a. Plant assemblages from the hanging wall sequence of the opencast mine Oberdorf NVoitsberg, Styria (Austria, Early Miocene, Ottnangian). Palaeontographica, Abteilung B.
ÐÐ ÐÐ in press b. The plant assemblages from the main seam parting of the western subbasin of Oberdorf, NVoitsberg, Styria, Austria (Early Miocene). Courier Forschungsinstitut Senckenberg.
598 P A L A E O N T O L O G Y , V O L U M E 4 4

ÐÐ MELLER, B. and ZETTER, R. 1998. Comparative investigations on the basal fossiliferous layers at the opencast mineOberdorf (Koe¯ach-Voitsberg lignite deposit, Styria, Austria; Lower Miocene). Review of Palaeobotany andPalynology, 101, 125±145.
KRUÈ SSMANN, G. 1976±1978. Handbuch der LaubgehoÈlze I-III. Parey Verlag, Berlin-Hamburg.KVACÏ EK, Z. 1971. Fossil Lauraceae in the stratigraphy of the North-Bohemian Tertiary. SbornõÂk GeologickyÂch veÏd
Paleontologie, 13, 47±86.ÐÐ and BUÊ ZÏ EK, C. 1966. Einige interessante Lauraceen und Symplocaceen des nordboÈhmischen TertiaÈrs. ZvlaÂsÏtnõÂ
Otisk VeÏstnõÂku UÂ streÏdnõÂho UÂ stavu GeologickeÂho, 41, 291±294.ÐÐ and WALTHER, H. 1984. Nachweis tertiaÈrer Theaceen Mitteleuropas nach blatt-epidermalen Untersuchungen. 2.
Bestimmung fossiler Theaceen-Sippen. Feddes Repertorium, 95 (5±6), 331±346.MAI, H.-D. and WALTHER, H. 1978. Die Floren der Haselbacher Serie im Weisselster-Becken (Bezirk Leipzig, DDR).
Abhandlungen des Staatlichen Museums fuÈr Mineralogie und Geologie zu Dresden, 28, 200 pp., 50 pls.ÐÐ and WALTHER, H. 1991. Die oligozaÈnen und untermiozaÈnen Floren NW-Sachsens und des Bitterfelder Raumes.
Abhandlungen des Staatlichen Museums fuÈr Mineralogie und Geologie zu Dresden, 38, 230 pp., 48 pls.MAURITSCH, H. J. and SCHOLGER, R. 1998. Palaeomagnetismus und Magnetostratigraphie im untermiozaÈnen Braun-
kohlentagebau Oberdorf (N Voitsberg, Steiermark, OÈ sterreich). 429±432. In STEININGER, F. F. (ed.). The EarlyMiocene lignite deposit of Oberdorf N Voitsberg (Styria, Austria). Jahrbuch der Geologischen Bundesanstalt, 140,395±496.
MELLER, B. 1998. Systematisch-taxonomische Untersuchungen von Karpo-Taphocoenosen des Koȯach-VoitsbergerBraunkohlenrevieres (Steiermark, OÈ sterreich, UntermiozaÈn) und ihre palaÈooÈkologische Bedeutung. Jahrbuch derGeologischen Bundesanstalt, 140, 497±655.
ÐÐ KOVAR-EDER, J. and ZETTER, R. 1999. Lower Miocene leaf, palynomorph, and diaspore assemblages from the baseof the lignite-bearing sequence in the opencast mine Oberdorf, N Voitsberg (Styria, Austria) as an indication of`Younger Mastixioid' vegetation. Palaeontographica, Abteilung B, 252, 123±179.
MILLER, P. 1754. Gardener's dictionary. Fourth abridged edition. John and James Rivington, London, 1582 pp.NEÆ MEJC, F. and KNOBLOCH, E. 1973. Die Makro¯ora der Salgo tarjaÂner Schichtengruppe (Die Flora aus Lipovany),
694±757. In PAPP, A., ROÈ GL, F. and SENESÏ , J. (eds). Chronostratigraphie und Neostratotypen Bd.3. M2,Ottnangien. Die Innviertler, Salgo tarjaÂner, BaÂntapusztaer Schichtengruppe und die Rzehakia Formation.Vydavatel'tstvo Slovenskej AkadeÂmie vied Bratislava, 841 pp.
NOOTEBOOM, H. P. 1985. Notes on Magnoliaceae with a revision of Pachylarnax and Elmerrillia and the Malesianspecies of Manglietia and Michelia. Blumea, 31, 65±121.
SCHOPMEYER, C. S. (tech. coord.) 1974. Seeds of woody plants in the United States. Forest Service, United StatesDepartment of Agriculture, Washington DC, 883 pp.
SIEBOLD, P. F. and ZUCCARINI, J. G. 1841. Flora japonica, 1. Lugduni batavorum, Leiden, 193 pp.SITAR, V. and KVACÏ EK, Z. 1997. Additions and revisions to the Early Miocene ¯ora of Lipovany (southern Slovakia).
Geologica Carpathica, 48 (4), 263±280.SPICER, R. A. 1989. The formation and interpretation of plant fossil assemblages. Advances in Botanical Research, 16,
96±191.STEININGER, F. F. (ed.). 1998. The Early Miocene lignite deposit of Oberdorf N Voitsberg (Styria, Austria). Jahrbuch der
Geologischen Bundesanstalt, 140, 395±496.ÐÐ DAXNER-HOÈ CK, G., HAAS, M., KOVAR-EDER, J., MAURITSCH, H. J., MELLER, B. and SCHOLGER, R. M. 1998. Stratigraphy of
the `basin ®ll' in the Early Miocene lignite opencast mine Oberdorf (N Voitsberg, Styria, Austria). 491±496. InSTEININGER, F. F. (ed.). The Early Miocene lignite deposit of Oberdorf N Voitsberg (Styria, Austria). Jahrbuch derGeologischen Bundesanstalt, 140, 395±496.
SZEÂ KY-FUX, V., PEÂ CSKAY, Z. and BALOGH, K. 1987. Miocene volcanic rocks from boreholes in Transtibiscia(Hungary) and their K/Ar chronology. Bulletin de l'AcadeÂmie Serbe Sciences des Arts Classe Sciences Naturelles,92, 109±128.
TINER, R. W. 1993. Coastal wetland plants. University of Massachusetts Press, Amherst, 328 pp.WALKER, E. H. 1944. A revision of Distylium and Sycopsis (Hamamelidaceae). Journal of the Arnold Arboretum,
Harvard University, 25, 319±341.WALTHER, H. 1998. Die TertiaÈr¯ora von Hammerunterwiesenthal (Freistaat Sachsen). Abhandlungen des Staatlichen
Museums fuÈr Mineralogie und Geologie zu Dresden, 43/44, 239±264.ÐÐ 1999. Die TertiaÈr¯ora von Kleinsaubernitz bei Bautzen. Palaeontographica, Abteilung B, 249, 63±174.ÐÐ and ZETTER, R. 1993. Zur Entwicklung der palaÈogenen Fagaceae Mitteleuropas. Palaeontographica, Abteilung B,
239, 183±194.WOROBIEC, G. 1994. Upper Miocene fossil plants from the outcrop of Stare Bystre (Western Carpathians, Poland). Acta
Palaeobotanica, 34, 83±105.
K O V A R - E D E R E T A L . : M I O C E N E P L A N T S 599

WU ZHENG-YI and RAVEN, P. H. (main eds) 1999. Flora of China. Vol. 4. Cycadaceae through Fagaceae. Science PressBeijing-Missouri Botanical Garden Press, St. Louis, 453 pp.
ZETTER, R. 1989. Methodik und Bedeutung einer routinemaÈûig kombinierten lichtmikroskopischen Untersuchungfossiler Mikro¯oren. Courier Forschungsinstitut Senckenberg, 109, 41±50.
ÐÐ 1998. Palynological investigations from the Early Miocene lignite opencast mine Oberdorf (N Voitsberg, Styria,Austria). 461±468. In STEININGER, F. F. (ed.). The Early Miocene lignite deposit of Oberdorf N Voitsberg (Styria,Austria). Jahrbuch der Geologischen Bundesanstalt, 140, 395±496.
JOHANNA KOVAR-EDER
Natural History MuseumGeological-Palaeontological Department
Burgring 7A-1014 Vienna, Austria
e-mail [email protected]
MARGIT HAAS
Institut fuÈr PetrologieGEO-Zentrum
Althanstrasse 14A-1090 Vienna, Austriae-mail [email protected]
CHRISTA-CHARLOTTE HOFMANN,
BARBARA MELLER
Institut fuÈr PalaÈontologieGEO-Zentrum
Althanstrasse 14A-1090 Vienna, Austria
e-mail [email protected]@univie.ac.at
Typescript received 23 February 2000Revised typescript received 10 November 2000
600 P A L A E O N T O L O G Y , V O L U M E 4 4
Related Documents











