Original article An assessment of the physiological properties of the so-called compatible solutes using in vitro experiments with leaf discs François Robert Larher *, Aziz Aziz, Yves Gibon, Patricia Trotel-Aziz, Ronan Sulpice, Alain Bouchereau Équipe Osmoadaptation et Métabolismes de Stress, Université de Rennes 1, (CNRS, UMR 6026 “ICM”), Campus Scientifique de Beaulieu, 263, avenue du Général Leclerc, 35042 Rennes cedex, France Received 15 November 2002; accepted 26 February 2003 Abstract Despite the considerable efforts directed over the past four decades to investigate, via correlative physiology and genetic engineering, the contribution of osmolytes to osmotolerance of higher plants, their physiological function(s) still remain a matter of debate. Here, we summarize the functions of proline and glycine betaine, recently documented in our group by using canola (Brassica napus L. var. oleifera), leaf discs treated in vitro with medium of low or high osmolarities. These discs accumulate proline when subjected to osmotic upshifts, whereas proline is rapidly consumed when the discs are further treated under non-stressing conditions. The period of time needed to osmoregulate the amount of proline is assumed to be too long to assign to the proline response a relevant function in turgor regulation which can be achieved with carbohydrates. In addition, proline accumulation is suppressed by supplying the osmotically stressing medium with polyamines, phyto-oxylipins and glycine betaine with no detrimental effect on the capacity to cope with osmotic stress. In opposition to proline, glycine betaine does not occur in canola. We take advantage of this situation to test the compatibility of exogenously supplied betaine in this plant material. Comparisons are done with spinach (Spinacia oleracea L.) leaf discs, which contain an endogenous amount of betaine and also exhibit the capacity to absorb a large amount of it. The betaine absorbed is not degraded and thus cannot be considered in this situation as a true osmoregulator. In turgid tissues of plants that do not produce betaine, this compound induces an elevated level of glutamine, a decreased level of protein, and photo-dependent glycine accumulation. This can rely on senescent-like effects and on a blockage in the glycolate cycle. Such damages to nitrogen metabolism are not observed in leaf explants treated simultaneously with betaine and the stressing agent. We also find that the treatments of turgid canola leaf discs with proline or betaine induce important changes in polyamine contents, mimicking, but with a higher order of magnitude, the effects of osmotic stress. Interestingly, this inducing activity is not observed when osmolytes are supplied under osmotic upshift conditions. Antagonistic effects between osmolytes and osmotic stress are described for the first time in higher plants. They suggest that proline and betaine can act as signal molecules in counteracting destabilizing effects to polyamine metabolism and lead us to decipher other counter-stress effects of nitrogenous osmolytes which, obviously, do not behave just as inert osmobalancers. © 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved. Keywords: Apparent toxicity; Brassica napus; Compatible solutes; Counter-stress effect; Glycine betaine; Proline; Spinacia oleracea 1. Introduction The compatible solutes “sensu stricto” [10], presently known on the basis of investigations performed with archea, bacteria, yeasts, algae and higher plants (as well as with various animals), can be brought together in a small number of biochemical “families” such as (a) polyols and their het- erosides, (b) amino acid such as proline, citrulline, glutamine and its derivatives, (c) N-acetylated amino acids such as N-D-acetylornithine and ectoines (d) betaines (trimethylam- monium compounds) and thetines (dimethylsulfonium com- pounds) and (e) the disaccharides sucrose and trehalose. Novel organic solutes have been recently characterized in methanogen bacteria [12] and marine algae [39]. The rationale for the selection of these compounds is not yet elucidated and it is suggested that the solute amassed in Abbreviations: ABA, abscissic acid; CLD, canola leaf disc; GB, glycine betaine; GS, glutamine synthetase; I 50 , concentration of an effector giving 50% inhibition; Me-JA, methyl jasmonate; 12-OPDA, 12-oxophytodienoic acid; PRO, proline; PRODH, proline dehydrogenase; PUT, putrescine; P5CS, pyrroline 5-carboxylate synthetase; SLD, spinach leaf disc; SPD, spermidine; SPM, spermine . * Corresponding author. E-mail address: [email protected] (F.R. Larher). Plant Physiology and Biochemistry 41 (2003) 657–666 www.elsevier.com/locate/plaphy © 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved. DOI: 10.1016/S0981-9428(03)00076-7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Original article
An assessment of the physiological properties of the so-calledcompatible solutes using in vitro experiments with leaf discs
François Robert Larher *, Aziz Aziz,Yves Gibon, Patricia Trotel-Aziz, Ronan Sulpice,Alain Bouchereau
Équipe Osmoadaptation et Métabolismes de Stress, Université de Rennes 1, (CNRS, UMR 6026 “ICM”), Campus Scientifique de Beaulieu,263, avenue du Général Leclerc, 35042 Rennes cedex, France
Received 15 November 2002; accepted 26 February 2003
Abstract
Despite the considerable efforts directed over the past four decades to investigate, via correlative physiology and genetic engineering, thecontribution of osmolytes to osmotolerance of higher plants, their physiological function(s) still remain a matter of debate. Here, wesummarize the functions of proline and glycine betaine, recently documented in our group by using canola (Brassica napusL. var.oleifera),leaf discs treated in vitro with medium of low or high osmolarities. These discs accumulate proline when subjected to osmotic upshifts,whereas proline is rapidly consumed when the discs are further treated under non-stressing conditions. The period of time needed toosmoregulate the amount of proline is assumed to be too long to assign to the proline response a relevant function in turgor regulation whichcan be achieved with carbohydrates. In addition, proline accumulation is suppressed by supplying the osmotically stressing medium withpolyamines, phyto-oxylipins and glycine betaine with no detrimental effect on the capacity to cope with osmotic stress. In opposition toproline, glycine betaine does not occur in canola. We take advantage of this situation to test the compatibility of exogenously supplied betainein this plant material. Comparisons are done with spinach (Spinacia oleraceaL.) leaf discs, which contain an endogenous amount of betaineand also exhibit the capacity to absorb a large amount of it. The betaine absorbed is not degraded and thus cannot be considered in this situationas a true osmoregulator. In turgid tissues of plants that do not produce betaine, this compound induces an elevated level of glutamine, adecreased level of protein, and photo-dependent glycine accumulation. This can rely on senescent-like effects and on a blockage in theglycolate cycle. Such damages to nitrogen metabolism are not observed in leaf explants treated simultaneously with betaine and the stressingagent. We also find that the treatments of turgid canola leaf discs with proline or betaine induce important changes in polyamine contents,mimicking, but with a higher order of magnitude, the effects of osmotic stress. Interestingly, this inducing activity is not observed whenosmolytes are supplied under osmotic upshift conditions. Antagonistic effects between osmolytes and osmotic stress are described for the firsttime in higher plants. They suggest that proline and betaine can act as signal molecules in counteracting destabilizing effects to polyaminemetabolism and lead us to decipher other counter-stress effects of nitrogenous osmolytes which, obviously, do not behave just as inertosmobalancers.
© 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.
Keywords:Apparent toxicity;Brassica napus; Compatible solutes; Counter-stress effect; Glycine betaine; Proline;Spinacia oleracea
1. Introduction
The compatible solutes “sensu stricto”[10], presentlyknown on the basis of investigations performed with archea,
bacteria, yeasts, algae and higher plants (as well as withvarious animals), can be brought together in a small numberof biochemical “families” such as (a) polyols and their het-erosides, (b) amino acid such as proline, citrulline, glutamineand its derivatives, (c) N-acetylated amino acids such asN-D-acetylornithine and ectoines (d) betaines (trimethylam-monium compounds) and thetines (dimethylsulfonium com-pounds) and (e) the disaccharides sucrose and trehalose.Novel organic solutes have been recently characterized inmethanogen bacteria[12] and marine algae[39].
The rationale for the selection of these compounds is notyet elucidated and it is suggested that the solute amassed in
Abbreviations:ABA, abscissic acid; CLD, canola leaf disc; GB, glycinebetaine; GS, glutamine synthetase; I50, concentration of an effector giving50% inhibition; Me-JA, methyl jasmonate; 12-OPDA, 12-oxophytodienoicacid; PRO, proline; PRODH, proline dehydrogenase; PUT, putrescine;P5CS, pyrroline 5-carboxylate synthetase; SLD, spinach leaf disc; SPD,spermidine; SPM, spermine .
* Corresponding author.E-mail address:[email protected] (F.R. Larher).
Plant Physiology and Biochemistry 41 (2003) 657–666
www.elsevier.com/locate/plaphy
© 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.DOI: 10.1016/S0981-9428(03)00076-7

one species should be highly congruous with the specificityof its cellular components. This may be related to coevolu-tion of the cells and their resident solutes throughout evolu-tionary changes. Such speculative suggestions can now beassessed through genetic engineering. For instance, cropspecies can be transformed to overexpress genes responsiblefor the biosynthesis of a given compatible solute [21]. Ifsynthesized and further tolerated, this novel substance isexpected to improve tolerance to osmotic stress in plantspecies that do not produce it naturally. A number of effortshave been devoted to demonstrate beneficial effects of suchnovel osmolytes in transgenic plants, but at the present time itis stressed that the practical outcomes of these studies remainmarginal [20]. The real interest of such biotechnologicalapproaches remains actually questionable, since tolerance toenvironmental stresses obviously results from quantitativetraits. As a consequence, it is illusive to hit the relevantgenetic target(s) by engineering a single gene or even arestricted number of genes. In this respect, the recent paper ofSeki et al. [38], is of outstanding importance since it high-lights a very large array of genes (and their products) re-quested for the responses of higher plants to drought, salinityand cold. Thus, it could be anticipated that the small numberof genes, which are involved in the biosynthesis of osmolytescould be only contributing to maintain internal osmotic ho-meostasis [48].
We are involved in the characterization of the physiologi-cal and metabolic processes associated with the accumula-tion of certain compatible solutes in halophytic higher plants[9]. We also extend our approach in seeking their physiologi-cal properties [26,27], and their usefulness when acting asmobile osmoregulators in vivo [29], giving a special attentionto proline (PRO) and glycine betaine (GB), presumably themost common osmolytes of higher plants. The changes inPRO metabolism responsible for the accumulation of thisamino acid are thoroughly studied, assuming that it coulddepend on the balance of increased de novo biosynthesis anddecreased consumption via oxidative processes and proteinsynthesis. This PRO accumulation is associated with relevantchanges in gene expression and enzyme amount on both thesides of PRO metabolism [13,14,45,47]. PRO accumulationmay also result from changes in the activity of the transportsystems involved in PRO distribution between the plant parts[34]. Surprisingly, the origin of the substrate(s) needed toaccumulate PRO in plant tissues is rarely investigated [17].Despite these very important efforts to elucidate the bio-chemical mechanisms of the PRO response, there is no clearagreement about its function in plant cells [20]. It is indeedconsidered as resulting alternatively from adaptive or detri-mental processes, reflecting induced disorders in amino acidmetabolism. It is stressed that an overaccumulation of PROthrough genetic engineering of the pathway responsible forits biosynthesis [25,28] has led authors to believe that it mayexert beneficial effects on growth under salt stress. In con-trast, other experiments demonstrate a damaging effect ofPRO [4,16,24].
In this paper, we bring together a number of experimentalevidences to demonstrate that osmotically induced PRO ac-cumulation can be suppressed under specific experimentalconditions, which suggests that it is not a prerequisite forimmediate water stress tolerance, but rather a symptomaticdisorder induced by dehydration.
The metabolic traits associated with GB production, aswell as its involvement as a true osmoregulator, in compari-son to that of PRO, are better argued. Its biosynthesis hasbeen studied in higher plants that naturally produce GB and itis well established that it is synthesized via a two-step path-way starting from choline and implying choline monooxyge-nase and betaine aldehyde dehydrogenase, which are stimu-lated under stress conditions [35]. However, while GBconcentration increased significantly under stress conditions(mainly salt stress), providing osmotically new particles tofulfil osmotic purposes, there is no evidence for an osmo-regulation of GB concentration when GB-accumulatingplants are transferred to non-stressing conditions. Thus, incontrast with what is well known for PRO [43], GB does notbehave as a true osmoregulator but rather as an inert end-product accumulated step by step in response to successiveosmotic stresses. Some of the biological properties of GBhave been elucidated through experiments performed withGram negative bacteria [11,37] where it was found to beactively absorbed and accumulated, suppressing the inhibi-tory effect of sodium chloride on the growth of bacteria. Thishas led to the paradigm that GB could be a universal compat-ible solute in any living cell, providing enhanced tolerance toenvironmental stresses. With this in mind and consideringthat the conversion of choline to GB is governed by a smallnumber of genes, authors have thought that GB productioncould be amenable to genetic engineering. This has actuallybeen achieved in certain higher plants such as Arabidopsis,rice and tobacco which do not naturally produce GB, throughtheir transformation with the single gene of choline oxidase(Cod A) from Arthrobacter globiformis. This enzyme cataly-ses both steps in the conversion of choline to GB [22].Indeed, these authors have obtained Arabidopsis transfor-mants, which contained in their shoots 1.2 µmol GB.g–1 freshweight. This amount appears to be very low in comparisonwith that present in the shoot of natural GB-accumulatingplants [9]. However, it has been claimed to be sufficient toenhance significantly the tolerance of Arabidopsis to variousstresses [23], suggesting that GB could be active directly orindirectly as an osmoprotectant rather than as a typical osmo-regulator.
The potential harmful effects of transformation with theCod A gene have been investigated, with the assumption thatit could affect directly choline metabolism and (or) inducetoxicity via an increased level of H2O2 in the leaf tissuessince this peroxide is a by-product of choline oxidation. Theauthors could not demonstrate such deleterious effects[1,22], which suggest promising prospects for this “ap-proach” of increasing GB content in plants through geneticengineering. However, we could assume that the GB effects
658 F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666

in situ were underestimated given its very low concentrationin the leaves of the transformed plants.
We have tried to assess the compatibility of GB in leafexplants of a non-natural GB-accumulating species (Bras-sica napus var. oleifera) and those of a natural accumulator(Spinacia oleracea) when it is absorbed from the externalmedium and further accumulated at very high levels. Theresults shown focus on the changes induced by GB in thenitrogen status in the leaf tissues of these two species.
Given the outstanding examples constituted by PRO andGB, we assess the physiological interest of the compatiblesolutes in stress tolerance of higher plants.
2. Results
2.1. Proline accumulation and its suppression by variouseffectors
2.1.1. Time course of osmoregulation of the PRO contentWhen treated with PEG (–1.69 MPa), canola leaf discs
(CLDs) accumulated PRO at a high rate after a 4 h lag phase(Fig. 1A). The final amount stabilized at a very high level inCLDs osmotreated for 20 h. After this preliminary osmotictreatment, when CLDs were transferred to the referencemedium without PEG (–0.05 MPa), their PRO content de-creased rapidly and then remained stable at a value higherthan that of freshly cut tissues. The successive changes ob-served could be relevant to osmoregulation of the PRO levelto achieve osmotic adjustment of the leaf explants. However,the timescale of PRO concentration and dilution could beunsuited to fulfil important functions in turgor restoration inresponse to osmotic upshift or osmotic downshift at the celllevel. We have actually found that turgor is partly re-established via changes in the amount of carbohydrate accu-mulated, within the 4 h following the onset of these treat-ments (Fig. 1B) and that PRO accumulated later on (Fig. 1C),i.e. when water deficit was partly alleviated [18].
2.1.2. Suppression of the osmoinduced PRO responseThe amount of accumulated PRO depends not only on the
osmotic potential of the osmoticum added to the externalmedium, but also on its chemical structure [30]. For instanceGB, at isoosmolar concentration with other osmotica(–2 MPa), behave as a very poor inducer of the PRO re-sponse. As a consequence, it was suspected to exert aninhibitory effect on this response. When added in comple-ment to stressing media that consisted of PEG (–3 MPa), GBbehaved indeed as a suppressor of PRO accumulation (Fig.2A), its I50 being close to 50 mM. Other possibilities tosuppress the PRO response were investigated through theextension of our pharmacological approach to various phy-tochemicals, whose concentrations are known to be modifiedunder stress conditions. This demonstrated the suppressingeffect of polyamines (Fig. 2B) and that of the phytooxy-lipines 12-oxophytodienoic acid and methyl jasmonate(Me-JA) (Fig. 2C). Other miscellaneous compounds such as
methylsalicylate, epibrassinolide, phloridzin also exhibitedsignificant suppressing effect with I50s of 0.25, 15,15.102 µM, respectively.
2.1.3. Changes in channelling of glutamate to PRO inpresence of suppressors
When the suppressing effect of polyamines was investi-gated we found that the spermine effect could be mimickedwith methionine sulfoximine, the inhibitor of glutamine syn-thetase [31]. Accordingly, the inhibitory effect of sperminewas reversed when glutamine was added to the incubatingmedium. This may constitute an indirect evidence to theinvolvement of GS and its product glutamine in the biosyn-thesis of the glutamate needed for the synthesis of accumu-lated PRO.
Similar effects were also observed in osmotically stressedCLDs supplied with Me-JA. Under such conditions the PROlevel decreased, as compared to that of discs treated onlywith the stressing agent while that of glutamine was found toincrease (Table 1). This could be related to a preferentialchannelling of glutamate to glutamine rather than to PRO, thebalance in favour of glutamine being promoted by Me-JA.
2.1.4. The counter-stress effect of PRO at the polyaminelevel
As shown in Fig. 3, CLDs incubated without osmoticstress but in the presence of increasing concentrations ofPRO exhibited important changes in their free polyamineprofiles, the amounts of putrescine and spermidine beingstrongly enhanced. Surprisingly, in CLDs supplemented withPRO while experiencing osmotic stress, the polyamine pro-
Fig. 1. Changes in PRO and total soluble carbohydrate (TSC) levels. (A)PRO levels in CLDs subjected for 20 h to a hyperosmotic shock with PEG6000 (–1.69 MPa) before being transferred (arrow) for 40 h to the referencemedium (–0.04 MPa). (B) total soluble carbohydrate levels in CLDs subjec-ted for 32 h to a hyperosmotic shock with PEG 6000 (–3 MPa). Control discswere maintained on the reference medium (–0.04 MPa). (C) PRO in CLDstreated as in B. Data are means of three replicates and bars indicate S.E.
659F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666

file was found to be stabilized giving a picture similar to thatobserved in either the freshly cut tissues or in those incubatedfor the same period of time under control conditions.
2.2. GB accumulation induces damages to nitrogenmetabolism
2.2.1. GB absorbed behaves as a dead-end productWhen incubated in presence of GB, CLDs accumulated
large amounts of this betaine, its final level depending on GBconcentration in the external medium and on the duration ofthe incubating period (Fig. 4). When CLDs were transferredto the reference medium without GB after an uptake periodof 6 h, their GB internal level remained constant. Moreover,feeding of CLDs with 14C-GB demonstrated that radioactiv-ity was quantitatively maintained under its original formeven after a long time of incubation (data not shown). Thecapacity to accumulate exogenous GB was found to be muchhigher in spinach leaf discs (SLDs), which already contain amild endogenous amount of GB at the onset of the experi-ment, the spinach plant being a natural producer of thisbetaine. GB accumulated was no longer consumed by SLDsafter their transfer to the reference medium without GB (datanot shown).
2.2.2. GB accumulation affects the protein and amino acidstatus
In CLDs treated for either 24 or 48 h with increasingconcentrations of GB, the total content of soluble proteinsdecreased (Fig. 5A), whereas in SLDs the same treatmentshad no effect on the protein level (not shown). In parallel, thefree amino acid contents increased in CLDs but not in SLDs
Fig. 2. The partial suppression of the osmoinduced PRO response by variouseffectors. (A) the time courses of PRO accumulation are shown for CLDstreated for 24 h with PEG 6000 (–2.5 MPa) added or not with 40 mM GB. (B,C) PRO accumulation in CLDs treated for 24 h with PEG 6000 (–2.5 MPa)added or not with putrescine, spermidine and spermine at various concen-trations (B) and Me-JA and 12-oxophytodienoic acid at various concentra-tions (C). Values in brackets are percentage of inhibition of PRO accumula-tion compared to that observed in CLDs treated with the sole osmoticum.
Table 1Changes in the amounts of free amino acids of CLDs subjected to a 20 hhyperosmotic treatment with PEG 6000 (–0.5 MPa), the stress mediumbeing supplied or not with 100 µM Me-JA
Amino acids (µmol g–1 DW) TreatmentsStress medium Stress medium +
Me-JAGlutamate 30.9 ± 4.7 29.2 ± 0.1Glutamine 8.8 ± 1.4 45.7 ± 4.3Proline 144.0 ± 15.4 103.2 ± 11.6Others 85.7 ± 7.4 84.4 ± 8.7
Fig. 3. Free polyamine contents of CLDs treated for 24 h with L-proline inthe light under either control or hyperosmotic conditions. Incubations wereperformed in the reference medium –0.1 MPa (control), supplemented withL-proline at concentrations ranging from 5 to 20 mM and added with PEG6000 (–1.7 MPa) (stress) or with PEG 6000 (–1.7 MPa) plus L-proline20 mM (stress + PRO).
660 F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666
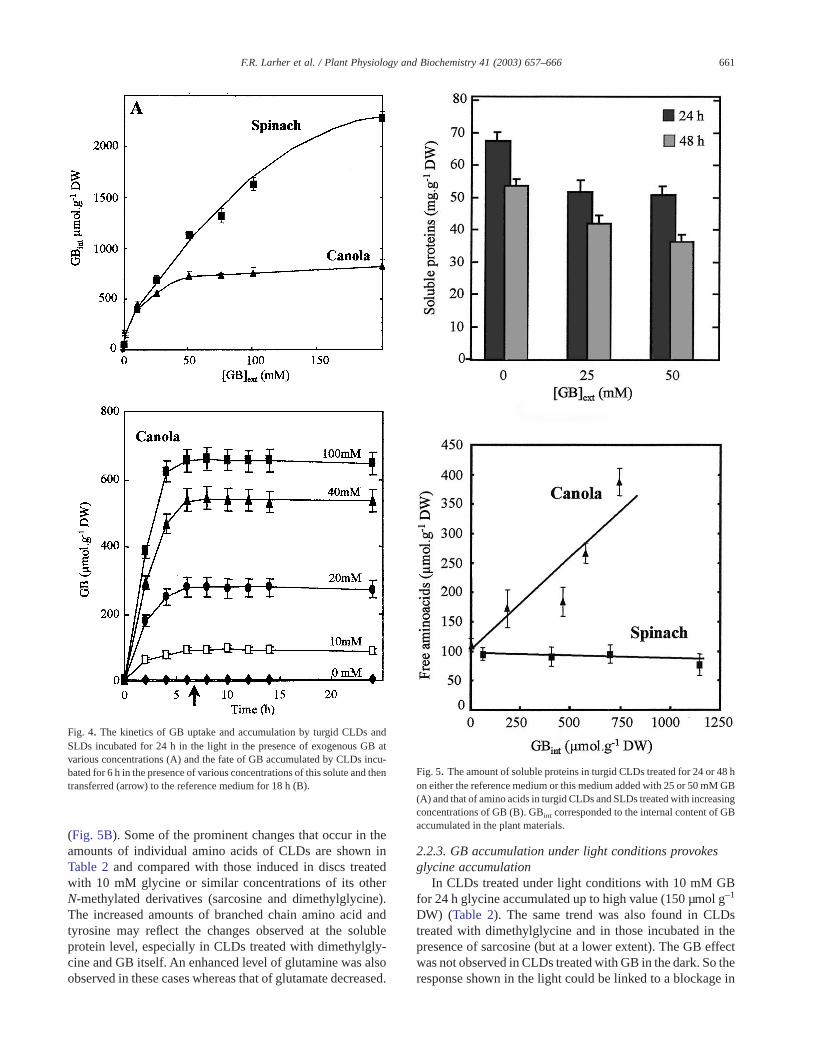
(Fig. 5B). Some of the prominent changes that occur in theamounts of individual amino acids of CLDs are shown inTable 2 and compared with those induced in discs treatedwith 10 mM glycine or similar concentrations of its otherN-methylated derivatives (sarcosine and dimethylglycine).The increased amounts of branched chain amino acid andtyrosine may reflect the changes observed at the solubleprotein level, especially in CLDs treated with dimethylgly-cine and GB itself. An enhanced level of glutamine was alsoobserved in these cases whereas that of glutamate decreased.
2.2.3. GB accumulation under light conditions provokesglycine accumulation
In CLDs treated under light conditions with 10 mM GBfor 24 h glycine accumulated up to high value (150 µmol g–1
DW) (Table 2). The same trend was also found in CLDstreated with dimethylglycine and in those incubated in thepresence of sarcosine (but at a lower extent). The GB effectwas not observed in CLDs treated with GB in the dark. So theresponse shown in the light could be linked to a blockage in
Fig. 4. The kinetics of GB uptake and accumulation by turgid CLDs andSLDs incubated for 24 h in the light in the presence of exogenous GB atvarious concentrations (A) and the fate of GB accumulated by CLDs incu-bated for 6 h in the presence of various concentrations of this solute and thentransferred (arrow) to the reference medium for 18 h (B).
Fig. 5. The amount of soluble proteins in turgid CLDs treated for 24 or 48 hon either the reference medium or this medium added with 25 or 50 mM GB(A) and that of amino acids in turgid CLDs and SLDs treated with increasingconcentrations of GB (B). GBint corresponded to the internal content of GBaccumulated in the plant materials.
661F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666

the metabolite fluxes through the photorespiratory glycolatecycle as demonstrated through GB treatment of leaf discsunder non-photorespiratory conditions (i.e. in the light and inthe presence of 1% O2). In these CLDs indeed the glycinelevel was stranded at a value similar to that of control CLDsin the light in the presence of 21% O2 [41].
2.2.4. The counter-stress effect of GB at the polyaminelevel
When CLDs were incubated under non-osmoticallystressing conditions in the presence of increasing concentra-tions of GB, they exhibited enhanced levels of putrescine andspermidine while that of spermine remained stable (Fig. 6).In contrast, when the GB treatment was carried out underosmotically stressing conditions (PEG –1.7 MPa), thepolyamine profile of CLDs was reset to that exhibited bythose kept under control conditions.
3. Discussion
It was anticipated that a wide range of physiological andmetabolic processes should be induced, suppressed or ad-justed in CLDs subjected in vitro to a sudden osmotic upshift.These changes were supposed to mimic those which occur inshoots of the whole plant challenged by water stress, to be
collectively responsible for re-establishment of the osmoticand ionic homeostasis and to repair the damages occurring atvarious levels of plant cells. They were expected to counter-act the lesions caused by the increase of external osmolarityand injuries due to secondary stresses and to allow for sur-vival and further growth. It is known that a great number ofthe responses which confer osmotolerance are primarily de-pendent on changes in gene expression [8], whereas otherscould be more directly related to changes in enzymes andmembrane stability and activity. Some of these responseswere devoted to turgor restoration that is assumed to partlydepend on production and storage of compatible osmolytes.In the present study, we have assessed some of the functionsof PRO accumulated in leaf discs of canola in response toosmotic stress and that of GB which is not produced by thisplant species but that can be absorbed and accumulated by itsleaf discs when made available in their incubating medium.
Canola leaf tissues actually behave as strong accumulatorsof PRO when challenged in vitro with the non-permeantosmoticum PEG 6000 at a suitable osmotic potential. This issupported by the conversion to PRO of a significant part ofthe amino acids stored in RubisCO, which could rely oncoordinated changes in the turnover of this enzyme [17]. Italso relies on changes in activity of enzymes involved in bothPRO synthesis (P5CS) and PRO degradation (PRODH) [44].Interestingly, when leaf discs subjected for 20–24 h to anhyperosmotic shock were transferred to hypo osmotic condi-tions, PRO was rapidly consumed to sustain both amino acidand protein synthesis as well as energetic metabolism [43].On these basis, we suggest that the pool of accumulated PRObehave as a reservoir, whose level of filling is osmoregulated.Incidentally, PRO may also help to preserve the structuralintegrity of components of the plant cells [46]. Regarding theinvolvement of PRO in turgor restoration of the stressed leafdiscs, it seems likely that its increase in concentration arisestoo late to play a primary role in this adjustment but this doesnot preclude its contribution to postponed osmotic processeswhich can take place later on at recovery of internal osmotichomeostasis.
In contrast with osmoinduced PRO accumulation whichcan be reversed by rehydration, the functions of GB in osmo-
Table 2Changes in glycine and some other free amino acid contents in turgid CLDs treated for 24 h, under light conditions, with 10 mM glycine or 10 mM of theN-methylated derivatives of this amino acid. DMG, dimethyl glycine; GB, glycine betaine; GLY, glycine; SARC, sarcosine
Amino acids (µmol g–1 DW) Control 10 mM GLY 10 mM SARC 10 Mm DMG 10 mM GBIsoleucine 0.9 ± 0.2 4.3 ± 0.4 1.1 ± 0.1 6.7 ± 0.2 6.5 ± 0.6Leucine 0.4 ± 0.0 0.9 ± 0.1 0.2 ± 0.0 8.3 ± 0.2 5.5 ± 0.5Valine 0.9 ± 0.3 3.2 ± 0.2 2.2 ± 0.0 9.0 ± 0.3 8.1 ± 1.3Tyrosine 0.4 ± 0.2 1.0 ± 0.1 1.0 ± 0.1 2.8 ± 0.4 2.2 ± 0.4Glutamate 33.4 ± 6.4 37.0 ± 0.9 29.4 ± 4.1 15.7 ± 1.8 5.4 ± 0.4Proline 1.4 ± 0.7 4.9 ± 0.9 5.1 ± 1.6 3.3 ± 0.6 4.3 ± 0.4Glutamine 3.9 ± 0.1 192.2 ± 12.4 6.7 ± 0.7 28.1 ± 0.3 26.5 ± 5.5Glycine 5.0 ± 0.3 41.0 ± 2.7 13.5 ± 3.1 79.2 ± 7.1 149.9 ± 6.8Others 60.1 ± 10.3 246.7 ± 19.8 71.4 ± 10.2 93.5 ± 7.5 74.4 ± 7.5
Total 106.4 ± 18.4 531.2 ± 37.5 130.6 ± 19.9 246.6 ± 18.4 282.8 ± 23.4
Fig. 6. Free polyamine contents of CLDs treated for 24 h with GB in thelight under either control or hyperosmotic conditions. Incubations wereperformed in the reference medium –0.1 MPa (control), supplemented withGB at concentrations ranging from 5 to 20 mM and added with PEG 6000(–1.7 MPa) (stress) or with PEG 6000 (–1.7 MPa) plus GB 20 mM (stress +GB 20 mM).
662 F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666

tolerance give rise to a number of different questions. Firstly,it has been shown that GB has a much narrower distributionthan PRO in higher plants [9]. Secondly, when present, itssignificance in osmotic adjustment could be questioned ifone consider that GB already occurs at a high level withoutany challenge by osmotic stresses. This could be consideredas an advantage if GB is acting more as an osmoprotectantthan as an osmoregulator. However, when the amounts of GBwere also found to increase in response to salt stress, accord-ing to a time-scale similar to that of PRO accumulation [19],they were not downregulated when stress conditions werewithdrawn. This could be merely reflecting the lack of acatabolic pathway for GB in higher plants and as demon-strated in this study, the GB absorbed from the externalmedium by the leaf tissues of producers or that of non-producers of GB behave as a perfect dead-end product. Thus,it is inferred that the typical GB pattern in higher plants doesnot conform to that of true osmoregulators whose concentra-tions at the cell level rapidly fit to the external osmoticpotential.
As demonstrated by Briens et al. [9], neither GB nor PROaccumulation constitutes a specific response to salinity inhalophytic plants. Indeed GB production is restricted to thespecies of a small number of genus and families, whereasPRO metabolism is obviously constitutive in higher plantsincluding the stress sensitive glycophytes. As a consequence,we have hypothesized that non-accumulators of PRO synthe-size effectors able to counteract the changes in PRO metabo-lism induced under stress conditions. GB itself was firstsuspected because strong accumulators of this compound arepoor producers of PRO [40]. In these species indeed, GBaccumulated at low and mild salinities while the PRO levelenhanced at very high salinity suggesting that GB couldexert, until a threshold value of salinity, a negative effect onthe components of the PRO response. Accordingly, we foundthat exogenous GB supplied to canola leaf explants subjectedto in vitro osmotic stress, behave as an effective suppressor ofthe PRO response. Similar effects were further obtained withaliphatic polyamines (especially spermine), phytooxylipineslike 12-oxophytodienoic acid and Me-JA. Moreover, whenused in mixtures, these compounds induced additive restrict-ing effects on the amount of PRO accumulated by the leafdiscs (Larher et al. unpublished). If the properties detectedwith our in vitro leaf discs assay could be extrapolated totheir endogenous counterparts in the whole plant subjected towater stress, one can anticipate that these compounds mightalso participate in planta to the control of the PRO response.It is well known that the PRO response is highly variableaccording to the plant species and even according to thevarieties of a given crop plant [32]. This variability could bepartly relying on the actual concentrations of these negativeeffectors. Moreover, the unexpected data obtained with thesenegative effectors strongly suggest that PRO accumulation isnot an escapable metabolic diversion in higher plants experi-encing water stress. In fact, it can be experimentally sup-pressed without any apparent damage.
The comparative study performed with the leaf discs ofspinach plants and that of canola plants, to study someaspects of the physiological interest of GB demonstrated thatthis betaine did not possess all the expected properties for auniversal compatible solute. Its natural occurrence in spinachwas already a definite proof for its harmlessness in thisspecies. This was confirmed by experiments demonstratingthat exogenous GB could be accumulated, in addition to theendogenous one, by SLDs without any apparent detrimentaleffect. In contrast, GB absorption by CLDs induces injuriesmade visible by a decrease of the total protein level and acoincidental increase of that of glutamine. This could bereflecting senescent-like processes [3] induced by GB. Inparallel, the CLDs treated with GB accumulated glycine in aphotodependent manner which revealed a negative effect atthe level of the glycolate cycle as shown by experimentsperformed under non-photorespiratory conditions. This re-sponse did not occur in SLDs which suggested that thetarget(s) needed to induce the GB effect could be different inthe producer and the non-producer of GB.
Given the fact that PRO and GB are nitrogenous solutes,additional changes could be expected in the N-status of theleaf discs treated in vitro with these compounds. We foundindeed that both compounds induced a strong increase ofputrescine and spermidine levels. These changes resembledthose provoked under osmotic stress and were suspected toreflect (or to be responsible for) non-optimal conditions forgrowth [5]. They could result for instance, from the “chemi-cal stress” provoked by these nitrogenous solutes when accu-mulated in plant cells at a toxic level. Further experiments areneeded to characterize the common target responsive to PROor GB, assuming that it could be located at the level ofenzymes involved in polyamine metabolism. It seems un-likely that changes in substrate-product relationships couldbe implicated since, contrary to PRO, GB cannot be con-verted to ornithine, the essential substrate for putrescinesynthesis. On the other hand, the suppressing effect exertedby exogenously supplied polyamines upon the PRO responsealso suggested regulatory effect(s), which specifically couldtake place under stress conditions. Such a remarkable inter-play was again underlying in experiments in which thesolute-mediated polyamine responses were abated, providedthat osmotic upshift and solute treatment were temporallyassociated. Alternatively, it becomes also likely that changesinduced directly or indirectly by osmotic stress (at least at thepolyamine profile level) could also be counteracted by PROor GB and that such a counter effect could be of paramountimportance if also effective in the whole plant experiencingosmotic stress. These unexpected observations could be re-flecting the “adaptive value” of compounds assumed to be-have, until the present time, as inert osmotic balancers whosemain function was assumed to provide cell turgor. Our resultsshow, for the first time, that they could play important func-tions in counteracting injuries induced by osmotic stress inshutting down or offsetting certain processes in a controlledway. The challenge now will be to elucidate the mechanisms
663F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666

of the counter-stress effects exerted by the so-called compat-ible solutes, which obviously are not inert osmolytes.
4. Methods
4.1. Plant material and growth conditions
Canola plants (Brassica napus L. var. oleifera cv. Samou-rai) and Spinach plants (Spinacia oleracea L. cv Junius)were grown for 6 weeks under controlled conditions as pre-viously described [30]. Discs (8 mm diameter) were preparedfrom fully expanded leaves using a cork borer, surface steril-ized (1% NaOCl) and immediately transferred for 2 h to areference medium (5 mM 2-[N-morpholino]-ethanesulfonicacid, 10 mM KCl, 1.5 mM CaCl2, pH 6.0).
4.2. Osmotic treatments
Hyperosmotic treatments were performed by adding PEG6000 at a specified concentration to the reference medium,and pH was readjusted to 6.0. CLDs were placed in 100 mlflasks, with each containing 15 discs in 15 ml of incubatingmedium. Flasks were closed with Petri dish covers, placedunder continuous light (22 W m–2 at the discs level) andshaken continuously at 29 °C for defined incubation time. Alltreatments were replicated three times concurrently withcontrol assays performed with CLDs incubated in flaskscontaining 15 ml of reference medium.
4.3. Treatments with PRO, GB and various effectors
CLDs were incubated either in the reference medium or inthe stress medium, supplemented with indicated concentra-tions of PRO, GB or effectors under continuous light asdescribed above. When effectors were solubilized in DMSO(i.e. Me-JA and 12-OPDA), control experiments were per-formed with CLDs incubated on the reference medium addedwith identical concentrations of this solvent. At the end of thetreatments, the discs were washed with distilled water andgently blotted on filter paper before extraction and analysis oforganic solutes.
4.4. Solute extraction
Treated discs were transferred into test tubes containing2 ml 95% ethanol (v/v) and kept in a water bath at 95 °C untilcomplete ethanol evaporation. Then, 1 ml of distilled waterwas added and the test tubes were vigorously shaken. Thecorresponding crude extracts were centrifuged (5 min,10,000 × g) in order to remove insoluble particles, the super-natants being kept for analysis.
4.5. Solute analysis and determination
4.5.1. Proline assayPRO contents were determined according to the method of
Troll and Lindsley [42], except that phosphoric acid wasomitted to avoid interference with sugars [33].
4.5.2. Total soluble carbohydrate determinationSoluble non-structural carbohydrate contents were deter-
mined with the anthrone reagent according to the method ofRoe [36].
4.5.3. Glycine betaine determinationGB was measured by high performance liquid chromatog-
raphy [17]. Briefly, the crude extracts were purified on aBioRad AG1-X8 anion exchanger. Then, GB was isolated ona 5 µm C18 (4.6 × 250 mm) reversed phase column (Spher-isorb 5 ODS2, Biochrom), using isocratic conditions of elu-tion with a 13 mM sodium 1-heptanesulphonate pH 3.7solution added with 5 mM Na2SO4. The GB peak was moni-tored by a UV detector (200 nm) and quantified in referenceto an external GB standard.
4.5.4. Polyamine extraction and analysisFrozen CLDs (10 units) were powdered and mixed with
1 M HCl. The homogenates were kept for 1 h on ice andcentrifuged for 20 min at 24,000 × g to obtain freepolyamines. HPLC equipped with fluorescence spectropho-tometer detector was used to separate and quantifypolyamines prepared as their dansyl derivatives according tothe method of [15]. The column was a reversed phase Ultra-sphere C18 (particle size 5 µm, 4.6 × 250 mm, Beckman,USA). Dansylated polyamines were eluted with a pro-grammed methanol:water (v/v) solvent gradient as describedin [2,31].
4.5.5. Amino acid analysisFree amino acids of the crude aqueous extracts were
characterized and quantified with HPLC after pre-columnderivatization with 6-aminoquinolyl-N-hydroxysuccini-midyl carbamate (AQC) (WatersAccq-Tag amino acid analy-sis system) and reversed-phase liquid chromatographic sepa-ration as described in [6].
4.6. Protein extraction and determination
Leaf discs treated or not with GB were rinsed, gentlyblotted on filter paper, placed in centrifuge tubes and thenfinely ground in the presence of liquid nitrogen. The powdersobtained were homogenized in a 20 mM Tris-HCl buffer (pH7.5), shaken vigorously for 1 min and frozen at –20 °C. Thenthey were thawed at room temperature and centrifuged at1000 × g. The supernatants were directly used for proteindetermination according to the method of Bradford [7].
Acknowledgements
The help of P. Gadal, at various steps of this study, iswarmfully acknowledged. The authors thank L. Leport forcritical reading of the successive versions of this paper. Theywould like to acknowledge the expert help of P. Lemesle inpreparing the manuscript and the excellent technical assis-tance of C. Monnier and N. Raimbeaud.
664 F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666

References
[1] Alia, T.H.H. Chen, N. Murata, Transformation with a gene for cholineoxidase enhances the cold tolerance of Arabidopsis during germina-tion and early growth, Plant Cell Environ. 21 (1998) 232–239.
[2] A. Aziz, J. Martin-Tanguy, F. Larher, Plasticity of polyamine metabo-lism associated with high osmotic stress in rape leaf discs and withethylene, J. Plant Growth Regul. 21 (1997) 153–163.
[3] D. Bauer, K. Biehler, H. Fock, E. Carrayol, B. Hirel, A. Migge, T.W.Beeker, A role for cytosolic glutamine synthetase in the remobiliza-
tion of leaf nitrogen during water stress in tomato, Physiol. Plant. 99(1997) 241–248.
[4] C.A. Bonner, D.S. Williams, H.C. Aldrich, R.A. Jensen, Antago-nism by L-glutamine of toxicity and growth inhibition caused by otheraminoacids in suspension cultures of Nicotiana silvestris, Plant Sci.113 (1996) 43–58.
[5] A. Bouchereau, A. Aziz, F. Larher, J. Martin-Tanguy, Polyamines andenvironmental challenges: recent development, Plant Sci. (review)140 (1999) 103–125.
[6] A. Bouchereau, C. Duhazé, J. Martin-Tanguy, J.P. Guégan, F. Larher,Improved analytical methods for determination of nitrogenous stressmetabolites occurring in Limonium species, J. Chromatogr. A 836(1999) 209–221.
[7] M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding, Anal. Biochem. 72 (1976) 248–254.
[8] E.A. Bray, Classification of genes differentially expressed duringwater-deficit stress in Arabidopsis thaliana: an analysis usingmicroarray and differential expression data, Ann. Bot. 89 (2002)803–811.
[9] M. Briens, F. Larher, Osmoregulation in halophytic higher plants: acomparative study of soluble carbohydrates, polyols, betaines and freeproline, Plant Cell Environ. 5 (1982) 287–292.
[10] A.D. Brown, J.R. Simpson, Water relations of sugar tolerant yeasts:the role of intracellular polyols, J. Gen. Microbiol. 72 (1972)589–591.
[11] L.N. Csonka, A.D. Hanson, Prokaryotic osmoregulation, Annu. Rev.Microbiol. 45 (1991) 569–606.
[12] M.S. da Costa, H. Santors, E.A. Galinski, An overview of the role anddiversity of compatible solutes in bacteria and archaea,Adv. Biochem.Eng. Biotechnol. 61 (1998) 117–153.
[13] A. Delauney, D.P.S. Verma, Proline biosynthesis and osmoregulationin plants, Plant J. 4 (1993) 215–223.
[14] K. Deuschle, D. Funck, H. Hellman, K. Däschner, S. Binder, W.B.Frommer, A nuclear gene encoding mitochondrial D1-pyrroline-5-
carboxylate dehydrogenase and its potential role in protection fromproline toxicity, Plant J. 27 (2001) 345–355.
[15] H.E. Flores, A.W. Galston, Analysis of polyamines in higher plantsby high performance liquid chromatography, Plant Physiol. 69 (1982)701–706.
[16] A.B. Garcia, J. de Almeida Engler, S. Iyer, T. Gerats, M. VanMontagu, A.B. Caplan, Effects of osmoprotectants upon NaCl stressin rice, Plant Physiol. 115 (1997) 159–169.
[17] Y. Gibon, M.A. Bessières, F. Larher, Is glycine betaine a non-compatible solute in higher plants that do not accumulate it?, PlantCell Environ. 20 (1997) 329–340.
[18] Y. Gibon, Syndrome prolinique osmodépendant dans les explantsfoliaires de Colza (Brassica napus L.); une adaptation métaboliqueassociée à la protéolyse Thèse Doct. Univ. Rennes 1, France. 1988(147 p.).
[19] G. Goas, M. Goas, F. Larher, Accumulation of free proline and glycinebetaine in Aster tripolium subjected to a saline shock: a kinetic studyrelated to light period, Physiol. Plant. 55 (1982) 383–388.
[20] P.D. Hare, W.A. Cress, J. van Staden, Dissecting the roles ofosmolyte accumulation during stress, Plant Cell Environ. 21 (1998)535–553.
[21] P.M. Hasegawa, R.A. Bressan, J.K. Zhu, H.J. Bohnert, Plant cellularand molecular responses to high salinity, Annu. Rev. Plant Physiol.Plant Mol. Biol. 51 (2000) 463–499.
[22] H. Hayashi, Alia, L. Mustardy, P. Deshnium, M. Ida, N. Murata,Transformation of Arabidopsis thaliana with the codA gene for cho-line oxidase; accumulation of glycinebetaine and enhanced toleranceto salt and cold stress, Plant J. 12 (1997) 133–142.
[23] H. Hayashi, Alia, A. Sakamoto, H. Nonaka, T.H.H. Chen, N. Murata,Enhanced germination under high-salt conditions of seeds of trans-genic Arabidopsis with a bacterial gene (codA) for choline oxidase, J.Plant Res. 111 (1998) 357–362.
[24] H. Hellman, D. Funk, D. Rentsch, W.B. Frommer, Hypersensitivity ofan Arabidopsis sugar signaling mutant towar exogenous proline appli-cation, Plant Physiol. 123 (2000) 779–790.
[25] Z. Hong, K. Lakkineni, Z. Zhang, D.P.S. Verma, Removal of feedbackinhibition of D1-pyrroline-5-carboxylate synthetase results inincreased proline accumulation and protection of plants from osmoticstress, Plant Physiol. 122 (2000) 1129–1136.
[26] Y. Jolivet, J. Hamelin, F. Larher, Osmoregulation in halophytic higherplants: the protective effects of glycine betaine and other relatedsolutes against the oxalate destabilization of membranes in beet rootcells, Zeit. Pflanzenphysiol. 109 (1983) 171–180.
[27] Y. Jolivet, F. Larher, J. Hamelin, Osmoregulation in halophytic higherplants: the protective effect of glycine betaine against the heat desta-bilization of membranes, Plant Sci. Lett. 24 (1982) 193–201.
[28] P.B. Kavi Kishor, Z. Hong, G.H. Miao, C.A.A. Hu, D.P.S. Verma,Over-expression of D-pyrroline-5-carboxylate synthetase increasesproline production and confers osmotolerance in transgenic plants,Plant Physiol. 108 (1995) 1387–1394.
[29] F. Larher, Natural abundance 13C-nuclear magnetic resonance studieson the compatible solutes of halophytic higher plants, Plant. Physiol.Biochem. 26 (1988) 35–45.
[30] F. Larher, L. Leport, M. Petrivalski, M. Chappart, Effectors for theosmoinduced proline response in higher plants, Plant Physiol. Bio-chem. 31 (1993) 911–922.
[31] F. Larher, A. Aziz, C. Deleu, P. Lemesle, A. Ghaffar, F. Bouchard,M. Plasman, Suppression of the osmoinduced proline response ofrapeseed leaf discs by polyamines, Physiol. Plant. 102 (1998)139–147.
[32] L. Leport, Accumulations de proline associées aux contraintes envi-ronnementales et à la floraison chez le Colza (Brassica napus L.)Thèse Doct; Univ. Rennes 1, France. 1992 (260 p.).
[33] C. Magné, F. Larher, High sugar content of extracts interferes withcolorimetric determination of amino acids and free proline, Anal.Biochem. 200 (1992) 115–118.
[34] D. Rentsch, B. Himer, E. Schmelzer, W.B. Frommer, Salt stress-induced proline transporters and salt stress-repressed broad specificamino acid permeases identified by suppression of a yeast amino acidtargeting mutant, Plant Cell 8 (1996) 1437–1446.
[35] D. Rhodes, A.D. Hanson, Quaternary ammonium and tertiary sulfo-nium compounds in higher plants, Annu. Rev. Plant Physiol. PlantMol. Biol. 44 (1993) 357–384.
[36] J.H. Roe, The determination of sugar in blood and spinal fluid withanthrone reagent, J. Biol. Chem. 212 (1955) 335–343.
[37] D. Sauvage, J. Hamelin, F. Larher, Glycine betaine and other structur-ally related compounds improve the salt tolerance of Rhizobiummeliloti, Plant Sci. Lett. 31 (1983) 291–302.
[38] M. Seki, M. Narusaka, J. Ishida, T. Nanjo, M. Fujita, Y. Oono,A. Kamiya, M. Nakajima, A. Enju, T. Sakurai, M. Satou, K. Akiyama,T. Taji, K. Yamaguchi-Shinozaki, P. Carninci, J. Kawai, Y. Hayash-izaki, K. Shinozaki, Monitoring the expression profiles of 7000 Ara-bidopsis genes under drought, cold and high-salinity stresses using afull-length cDNA microarray, Plant J. 31 (2002) 279–292.
[39] C. Simon-Colin, N. Kervarec, R. Pichon, M.A. Bessières, E. Deslan-des, Characterization of N-methyl-L-methionine sulfoxide andisethionic acid from the red alga Grateloupia doryphora, Phycol. Res.50 (2002) 125–128.
665F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666

[40] G.R. Stewart, F. Larher, I. Ahmad, J.A. Lee, Nitrogen metabolismand salt-tolerance in higher plants halophytes, in: R.L. Jefferies, A.J.Davy (Eds.), Ecological Processes in Coastal Environments, Black-
well Scientific Publications, Oxford, 1979, pp. 211–227.[41] R. Sulpice, Y. Gibon, G. Cornic, F.R. Larher, Interaction between
exogenous glycine betaine and the photorespiratory pathway incanola leaf discs, Physiol. Plant. 116 (2003) 460–467.
[42] W. Troll, J. Lindsley, A photometric method for determination ofproline, J. Biol. Chem. 215 (1955) 655–660.
[43] P. Trotel, A. Bouchereau, M.F. Niogret, F. Larher, The fate of osmo-accumulated proline in leaf discs of Rape (Brassica napus L.) incu-bated in a medium of low osmolarity, Plant Sci. 118 (1996) 31–45.
[44] P. Trotel-Aziz, M.F. Niogret, C. Deleu, A. Bouchereau, F. Larher, Thecontrol of proline mobilization by abscissic acid during osmotic stressrecovery of canola leaf discs, Physiol. Plant. 116 (2003) 213–221.
[45] N. Verbruggen, X.J. Hua, M. May, M. Van Montagu, Environmentaland developmental signals modulate proline homeostasis: evidencefor a negative transcriptional regulator, Proc. Natl. Acad. Sci. USA 93(1996) 8787–8791.
[46] D.M. Vernon, H.J. Bonhert, A novel methyl transferase induced byosmotic stress in the facultative halophyte Mesembryanthemum crys-tallinum, EMBO J. 11 (1992) 2077–2085.
[47] Y. Yoshiba, T. Kiyosue, T. Katagiri, H. Ueda, T. Mizoguchi, K. Yama-gushi-Shinozaki, K. Wada, Y. Harada, K. Shinozaki, Correlationbetween the induction of a gene for d1-pyrroline-5-carboxylate syn-thetase and the accumulation of proline in Arabidopsis thaliana underosmotic stress, Plant J. 7 (1995) 751–760.
[48] J.K. Zhu, Salt and drought stress signal transduction in plants, Annu.Rev. Plant Biol. 53 (2002) 247–273.
666 F.R. Larher et al. / Plant Physiology and Biochemistry 41 (2003) 657–666
Related Documents











