Cell Calcium 49 (2011) 184–190 Contents lists available at ScienceDirect Cell Calcium journal homepage: www.elsevier.com/locate/ceca Amyloid peptide oligomers directly activate NMDA receptors Laura Texidó a,b , Mireia Martín-Satué a,b , Elena Alberdi a,c , Carles Solsona a,b , Carlos Matute a,c,∗ a Centro de Investigación Biomédica en Red en Enfermedades Neurodegenerativas (CIBERNED), Spain b Department of Pathology and Experimental Therapeutics and Institut d’Investigació Biomèdica de Bellvitge (IDIBELL), Faculty of Medicine, Universidad de Barcelona, E-08907 Hospitalet de Llobregat, Spain c Departamento de Neurociencias, Universidad del País Vasco, E-48940 Leioa, Spain article info Article history: Received 2 December 2010 Received in revised form 1 February 2011 Accepted 3 February 2011 Available online 23 February 2011 Keywords: Amyloid NMDA receptors Xenopus oocytes Neuron death Alzheimerˇ ıs disease abstract Amyloid beta (A) oligomers accumulate in the brain tissue of Alzheimer disease patients and are related to disease pathogenesis. The precise mechanisms by which A oligomers cause neurotoxicity remain unknown. We recently reported that A oligomers cause intracellular Ca 2+ overload and neuronal death that can be prevented by NMDA receptor antagonists. This study investigated whether A oligomers directly activated NMDA receptors (NMDARs) using NR1/NR2A and NR1/NR2B receptors that were het- erologously expressed in Xenopus laevis oocytes. Indeed, A oligomers induced inward non-desensitizing currents that were blocked in the presence of the NMDA receptor antagonists memantine, APV, and MK- 801. Intriguingly, the amplitude of the responses to A oligomers was greater for NR1/NR2A heteromers than for NR1/NR2B heteromers expressed in oocytes. Consistent with these findings, we observed that the increase in the cytosolic concentration of Ca 2+ induced by A oligomers in cortical neurons is prevented by AP5, a broad spectrum NMDA receptor antagonist, but slightly attenuated by ifenprodil which blocks receptors with the NR2B subunit. Together, these results indicate that A oligomers directly activate NMDA receptors, particularly those with the NR2A subunit, and further suggest that drugs that attenuate the activity of such receptors may prevent A damage to neurons in Alzheimerˇ ıs disease. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction Soluble oligomers of the amyloid- (A) peptide are neuro- toxins that contribute to Alzheimer’s disease (AD) pathology. A oligomers form in mouse models of AD [1,2] and accumulate in the cerebrospinal fluid (CSF) [3,4] and brain tissue of AD patients [4–6]. The abundance of A oligomers also correlates with disease pro- gression [7–9].A oligomers exert neurotoxic effects by disrupting the integrity of both plasma and intracellular membranes [10] and by accumulating at excitatory synapses, impairing synapse func- tion [11,12].A oligomers may be toxic due to calpain activation following Ca 2+ influx mediated by NMDA receptors [13] and due to oxidative stress and mitochondrial damage initiated by NMDA receptor activation [14]. Notably, A oligomer neurotoxicity is neu- tralized by NMDA receptor antagonists, both in dissociated neurons and in organotypic hippocampal cultures [14]. Since A peptides can regulate the release of glutamate [15], the deleterious effects of A oligomers could be caused by over- activation of NMDA receptors due to excessive glutamate at synapses. However, silencing neuronal activity with tetrodotoxin ∗ Corresponding author at: Departamento de Neurociencias, Universidad del País Vasco, E-48940 Leioa, Spain. Tel.: +34 94 601 3244; fax: +34 94 601 3400. E-mail address: [email protected] (C. Matute). and removal of extracellular Ca 2+ to minimize transmitter release does not have any effect on the NMDA receptor-mediated cur- rents induced by A oligomers [14]. Because of that observation, we hypothesized that A oligomers may directly activate NMDA receptors, in particular those formed by the NR2A and NR2B sub- units which are abundantly expressed in the cerebral cortex and hippocampus. In the current study, we use a Xenopus oocyte expression system to show that A oligomers activate recombi- nant NMDA receptors formed by NR1 and NR2A or NR2B subunits. These subunits are abundantly expressed in the cerebral cortex and hippocampus, two regions that are particularly vulnerable to AD. 2. Materials and methods 2.1. Animals and solutions Oocytes were obtained from mature Xenopus laevis females (Centre d’Elevage des Xénopes, Montpellier, France). Stage V and VI oocytes were collected and maintained at 16–17 ◦ C in Barth’s solution (88 mM NaCl, 1 mM KCl, 0.33 mM Ca(NO 3 ) 2 , 0.41 mM CaCl 2 , 0.82 mM MgSO 4 , 2.40 mM NaHCO 3 , and 20 mM HEPES at pH 7.5) supplemented with penicillin (100 IU/ml) and streptomycin (0.1 mg/ml). 0143-4160/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.ceca.2011.02.001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A
La
b
Fc
a
ARRAA
KANXNA
1
tocTgtbtftrta
tas
V
0d
Cell Calcium 49 (2011) 184–190
Contents lists available at ScienceDirect
Cell Calcium
journa l homepage: www.e lsev ier .com/ locate /ceca
myloid � peptide oligomers directly activate NMDA receptors
aura Texidóa,b, Mireia Martín-Satuéa,b, Elena Alberdia,c, Carles Solsonaa,b, Carlos Matutea,c,∗
Centro de Investigación Biomédica en Red en Enfermedades Neurodegenerativas (CIBERNED), SpainDepartment of Pathology and Experimental Therapeutics and Institut d’Investigació Biomèdica de Bellvitge (IDIBELL),aculty of Medicine, Universidad de Barcelona, E-08907 Hospitalet de Llobregat, SpainDepartamento de Neurociencias, Universidad del País Vasco, E-48940 Leioa, Spain
r t i c l e i n f o
rticle history:eceived 2 December 2010eceived in revised form 1 February 2011ccepted 3 February 2011vailable online 23 February 2011
eywords:myloid �
a b s t r a c t
Amyloid beta (A�) oligomers accumulate in the brain tissue of Alzheimer disease patients and are relatedto disease pathogenesis. The precise mechanisms by which A� oligomers cause neurotoxicity remainunknown. We recently reported that A� oligomers cause intracellular Ca2+ overload and neuronal deaththat can be prevented by NMDA receptor antagonists. This study investigated whether A� oligomersdirectly activated NMDA receptors (NMDARs) using NR1/NR2A and NR1/NR2B receptors that were het-erologously expressed in Xenopus laevis oocytes. Indeed, A� oligomers induced inward non-desensitizingcurrents that were blocked in the presence of the NMDA receptor antagonists memantine, APV, and MK-
MDA receptorsenopus oocyteseuron deathlzheimerıs disease
801. Intriguingly, the amplitude of the responses to A� oligomers was greater for NR1/NR2A heteromersthan for NR1/NR2B heteromers expressed in oocytes. Consistent with these findings, we observed that theincrease in the cytosolic concentration of Ca2+ induced by A� oligomers in cortical neurons is preventedby AP5, a broad spectrum NMDA receptor antagonist, but slightly attenuated by ifenprodil which blocksreceptors with the NR2B subunit. Together, these results indicate that A� oligomers directly activateNMDA receptors, particularly those with the NR2A subunit, and further suggest that drugs that attenuate
tors m
the activity of such recep. Introduction
Soluble oligomers of the amyloid-� (A�) peptide are neuro-oxins that contribute to Alzheimer’s disease (AD) pathology. A�ligomers form in mouse models of AD [1,2] and accumulate in theerebrospinal fluid (CSF) [3,4] and brain tissue of AD patients [4–6].he abundance of A� oligomers also correlates with disease pro-ression [7–9]. A� oligomers exert neurotoxic effects by disruptinghe integrity of both plasma and intracellular membranes [10] andy accumulating at excitatory synapses, impairing synapse func-ion [11,12]. A� oligomers may be toxic due to calpain activationollowing Ca2+ influx mediated by NMDA receptors [13] and dueo oxidative stress and mitochondrial damage initiated by NMDAeceptor activation [14]. Notably, A� oligomer neurotoxicity is neu-ralized by NMDA receptor antagonists, both in dissociated neuronsnd in organotypic hippocampal cultures [14].
Since A� peptides can regulate the release of glutamate [15],he deleterious effects of A� oligomers could be caused by over-ctivation of NMDA receptors due to excessive glutamate atynapses. However, silencing neuronal activity with tetrodotoxin
∗ Corresponding author at: Departamento de Neurociencias, Universidad del Paísasco, E-48940 Leioa, Spain. Tel.: +34 94 601 3244; fax: +34 94 601 3400.
E-mail address: [email protected] (C. Matute).
143-4160/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.ceca.2011.02.001
ay prevent A� damage to neurons in Alzheimerıs disease.© 2011 Elsevier Ltd. All rights reserved.
and removal of extracellular Ca2+ to minimize transmitter releasedoes not have any effect on the NMDA receptor-mediated cur-rents induced by A� oligomers [14]. Because of that observation,we hypothesized that A� oligomers may directly activate NMDAreceptors, in particular those formed by the NR2A and NR2B sub-units which are abundantly expressed in the cerebral cortex andhippocampus. In the current study, we use a Xenopus oocyteexpression system to show that A� oligomers activate recombi-nant NMDA receptors formed by NR1 and NR2A or NR2B subunits.These subunits are abundantly expressed in the cerebral cortexand hippocampus, two regions that are particularly vulnerableto AD.
2. Materials and methods
2.1. Animals and solutions
Oocytes were obtained from mature Xenopus laevis females(Centre d’Elevage des Xénopes, Montpellier, France). Stage V and
◦
VI oocytes were collected and maintained at 16–17 C in Barth’ssolution (88 mM NaCl, 1 mM KCl, 0.33 mM Ca(NO3)2, 0.41 mMCaCl2, 0.82 mM MgSO4, 2.40 mM NaHCO3, and 20 mM HEPES at pH7.5) supplemented with penicillin (100 IU/ml) and streptomycin(0.1 mg/ml).
alcium
2N
npSDluCaiio0Cs
2
rartg(FtfatatE
2
atfiMoapNcflwccoa
wbr1
2
od
L. Texidó et al. / Cell C
.2. Expression of NMDA receptors comprised of NR1/NR2A andR1/NR2B subunits
The plasmids containing the cDNA for human NR1 (pCI-eo), NR2A (HumNR2A-pcDNAI/Amp) and NR2B (HumNR2B6-cDNAI/Amp) subunits were provided by Dr. Paul Whiting, Ph.D.,enior Director, Neuroscience Research Centre (Merck Sharp &ohme Research Laboratories, Harlow, Essex, UK). Plasmids were
inearized with XbaI (NR1) or EcoRV (NR2A, NR2B), and the prod-cts were used for cRNA synthesis in vitro using the mCAP RNAapping Kit (Stratagene). The resulting capped cRNA was aliquotednd stored at −80 ◦C. The cRNA was injected (50 nl, 1.5 �g/�l)nto Xenopus oocytes 48–72 h before electrophysiological record-ng was performed. The follicular cell layer was removed from theocytes by enzymatic digestion [incubation of the oocytes with.25 mg/ml collagenase type 1A; (Sigma)] 24 h before recording.ontrol oocytes were injected with 50 nl of ultrapure water andubjected to the same enzymatic digestion procedure.
.3. Preparation of Aˇ oligomers
A�1-42 (ABX, Radeberg, Germany) was dissolved in hexafluo-oisopropanol (Sigma, St Louis, MO, USA) to obtain a 1 mM solutionnd then aliquoted in sterile microcentrifuge tubes. The hexafluo-oisopropanol was removed under vacuum using a SpeedVac andhe peptide film was stored (desiccated) at −80 ◦C. For the aggre-ation protocol, the peptide was first resuspended in dry DMSOSigma, St Louis, MO, USA) to a concentration of 5 mM, then Hams-12 (PromoCell, Labclinics, Barcelona, Spain) was added to bringhe peptide to a final concentration of 100 �M and incubated at 4 ◦Cor 24 h. The preparation was centrifuged at 14,000 × g for 10 mint 4 ◦C to remove insoluble aggregates, and the supernatants con-aining soluble A�1–42 were transferred to clean tubes and storedt 4 ◦C. A� peptide aggregation was evaluated by SDS–PAGE elec-rophoresis and Western blotting using 6E10 antibodies (Covance,meryville, California) (Supplementary Fig. S1).
.4. Electrophysiology
All recordings were made at room temperature (20–22 ◦C)nd performed using a two-microelectrode voltage-clamp sys-em (Axoclamp-2A, Axon Instruments, USA). Microelectrodes werelled with KCl (3 M) and had resistances ranging from 1 to 2 M�.embrane currents were low-pass filtered at 10 Hz and recorded
n a PC using the Whole Cell Analysis program (WinWCP v3.2.8nd v3.3.3) at twice the rate of the filter frequency. The membraneotential of the oocytes was clamped at −80 mV. Currents due toMDAR expression were evoked by bath application of a solutionontaining 100 �M l-glutamic acid and 10 �M glycine for 25 s. Theow rate was 8 ml/min. Oocytes were exposed to A� oligomers thatere manually applied to the surface of the cells for 25 s. In some
ases, manual delivery induced a change in the slope of the inwardurrent that was considered an artifact. The oligomers were washedut by re-establishing the flow; this, too, sometimes induced a smallrtifact current.
When investigating the effects of NMDAR antagonists, oocytesere preincubated with the NMDA receptor antagonists for 10 min
efore adding the A�. The total recording time was 2.5 min. Theecording solution was 115 mM NaCl, 2 mM KCl, 1.8 mM CaCl2, and0 mM HEPES at pH 7.4.
.5. Calcium imaging in cultured neurons from the cerebral cortex
Primary neuron cultures were obtained from the cortical lobesf E18 Sprague–Dawley rat embryos according to previouslyescribed procedures [14]. Cells were resuspended in B27 Neu-
49 (2011) 184–190 185
robasal medium plus 10% FBS (Sigma, St Louis, MO, USA) and thenseeded onto poly-l-ornithine-coated glass coverslips at 1 × 105
cells per coverslip (12 mm in diameter) and 48-well plates at1.5 × 105 per well. One day later, the medium was replaced byserum-free-, B27-supplemented Neurobasal medium. The cultureswere essentially free of astrocytes and microglia; they were main-tained at 37 ◦C and 5% CO2. Cultures were used 8–10 days afterplating.
[Ca2+]i was determined as previously described [14]. Oligoden-drocytes were loaded with fura-2 AM (5 �M; Molecular Probes,Invitrogen, Barcelona, Spain) in culture medium for 30 min at 37 ◦C.Cells were washed in HBSS containing 20 mM HEPES, pH 7.4, 10 mMglucose, and 2 mM CaCl2 (incubation buffer) for 10 min at roomtemperature. Experiments were performed in a coverslip cham-ber continuously perfused with incubation buffer at 4 ml/min. Theperfusion chamber was mounted on the stage of a inverted epiflu-orescence microscope (Zeiss Axiovert 35, Oberkochen, Germany)equipped with a 150 W xenon lamp Polychrome IV (T.I.L.L. Photon-ics, Martinsried, Germany) and a Plan Neofluar 40× oil immersionobjective (Zeiss). Cells were visualized with a high-resolution dig-ital black/white CCD camera (ORCA C4742-80-12 AG; HamamatsuPhotonics Iberica). [Ca2+]i was estimated by the 340/380 ratiomethod, using a Kd value of 224 nM. At the end of the assay, in situcalibration was performed with the successive addition of 10 mMionomycin and 2 M Tris/50 mM EGTA, pH 8.5. Data were analyzedwith Excel (Microsoft, Seattle, WA, USA) and Prism (GraphPad Soft-ware, San Diego, CA, USA) software.
2.6. Drugs
l-Glutamic acid and glycine were obtained from Sigma (St. Louis,MO, USA). The NMDA receptor antagonists D-AP5, MK-801, andmemantine were obtained from Tocris Bioscience (Cookson, Bristol,UK).
2.7. Calculations and statistics
Statistical analysis was performed using SigmaStat 3.2 soft-ware (SPSS Inc., Chicago, IL, USA). Values are reported as themean ± S.E.M. The Student’s t-test was used to compare the meansof two independent groups of normally distributed data.
3. Results
3.1. Effects of Aˇ oligomers on oocytes injected with water(control)
We first studied the interaction of soluble A� oligomers withNMDARs that were heterologously expressed in X. laevis oocytes.Defolliculated oocytes were injected with water and voltageclamped at −80 mV. To determine the response to NMDAR agonists,l-glutamic acid (100 �M) plus glycine (10 �M) or glycine (10 �M)alone was added. No currents were observed in the water-injectedoocytes (Fig. 1A). Similarly, no currents were observed when A�oligomers (1 �M) were added with glycine (Fig. 1A).
3.2. Aˇ oligomers elicit inward currents in NR1/NR2A-injectedoocytes
We next measured the effect of A� oligomers on NR1/NR2A-subtype NMDARs. When l-glutamic acid and glycine were added
to oocytes expressing NR1/NR2 NMDARs, there was a reversibleinward current with a mean maximal amplitude of 0.45 ± 0.07 �Aand a mean electrical charge of 14.70 ± 2.60 �C (n = 10) (Fig. 1B).Applied alone, glycine also evoked inward currents with a meanamplitude of 0.16 ± 0.04 �A and a mean charge or 4.45 ± 1.14 �C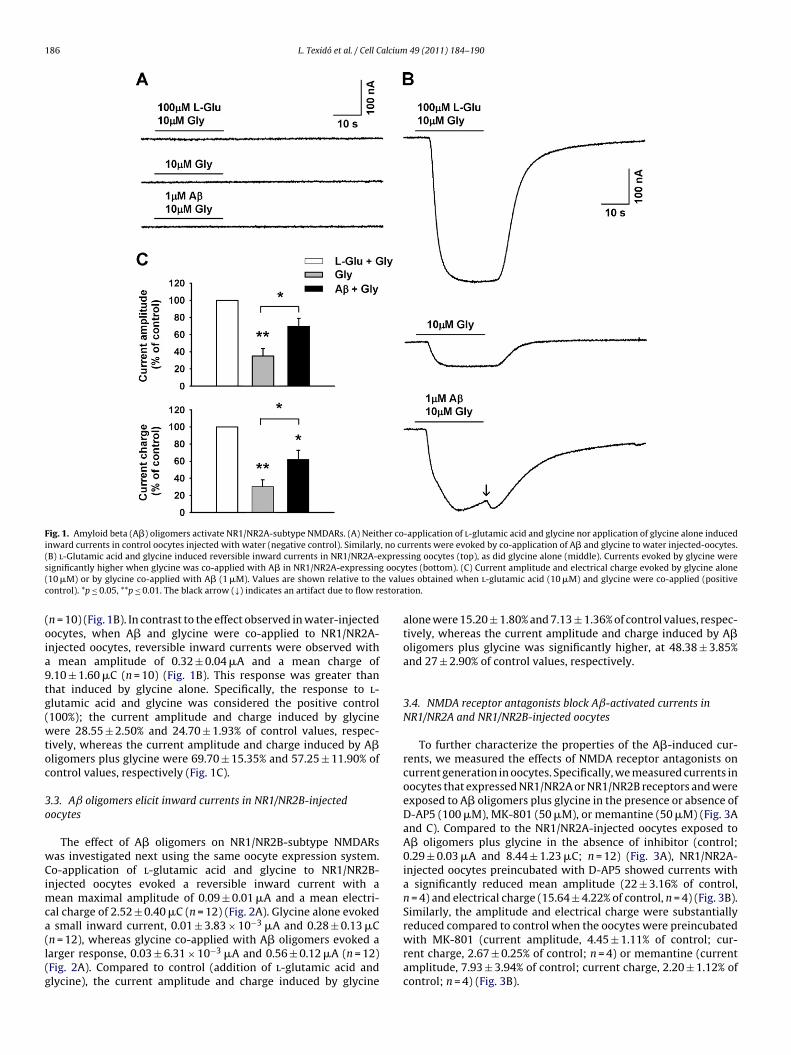
186 L. Texidó et al. / Cell Calcium 49 (2011) 184–190
Fig. 1. Amyloid beta (A�) oligomers activate NR1/NR2A-subtype NMDARs. (A) Neither co-application of l-glutamic acid and glycine nor application of glycine alone inducedinward currents in control oocytes injected with water (negative control). Similarly, no currents were evoked by co-application of A� and glycine to water injected-oocytes.( xpress oocy( e valuc estora
(oia9tg(wtoc
3o
wCimca(l(g
B) l-Glutamic acid and glycine induced reversible inward currents in NR1/NR2A-eignificantly higher when glycine was co-applied with A� in NR1/NR2A-expressing10 �M) or by glycine co-applied with A� (1 �M). Values are shown relative to thontrol). *p ≤ 0.05, **p ≤ 0.01. The black arrow (↓) indicates an artifact due to flow r
n = 10) (Fig. 1B). In contrast to the effect observed in water-injectedocytes, when A� and glycine were co-applied to NR1/NR2A-njected oocytes, reversible inward currents were observed with
mean amplitude of 0.32 ± 0.04 �A and a mean charge of.10 ± 1.60 �C (n = 10) (Fig. 1B). This response was greater thanhat induced by glycine alone. Specifically, the response to l-lutamic acid and glycine was considered the positive control100%); the current amplitude and charge induced by glycineere 28.55 ± 2.50% and 24.70 ± 1.93% of control values, respec-
ively, whereas the current amplitude and charge induced by A�ligomers plus glycine were 69.70 ± 15.35% and 57.25 ± 11.90% ofontrol values, respectively (Fig. 1C).
.3. Aˇ oligomers elicit inward currents in NR1/NR2B-injectedocytes
The effect of A� oligomers on NR1/NR2B-subtype NMDARsas investigated next using the same oocyte expression system.o-application of l-glutamic acid and glycine to NR1/NR2B-
njected oocytes evoked a reversible inward current with aean maximal amplitude of 0.09 ± 0.01 �A and a mean electri-
al charge of 2.52 ± 0.40 �C (n = 12) (Fig. 2A). Glycine alone evoked
small inward current, 0.01 ± 3.83 × 10−3 �A and 0.28 ± 0.13 �Cn = 12), whereas glycine co-applied with A� oligomers evoked aarger response, 0.03 ± 6.31 × 10−3 �A and 0.56 ± 0.12 �A (n = 12)Fig. 2A). Compared to control (addition of l-glutamic acid andlycine), the current amplitude and charge induced by glycine
sing oocytes (top), as did glycine alone (middle). Currents evoked by glycine weretes (bottom). (C) Current amplitude and electrical charge evoked by glycine alonees obtained when l-glutamic acid (10 �M) and glycine were co-applied (positivetion.
alone were 15.20 ± 1.80% and 7.13 ± 1.36% of control values, respec-tively, whereas the current amplitude and charge induced by A�oligomers plus glycine was significantly higher, at 48.38 ± 3.85%and 27 ± 2.90% of control values, respectively.
3.4. NMDA receptor antagonists block Aˇ-activated currents inNR1/NR2A and NR1/NR2B-injected oocytes
To further characterize the properties of the A�-induced cur-rents, we measured the effects of NMDA receptor antagonists oncurrent generation in oocytes. Specifically, we measured currents inoocytes that expressed NR1/NR2A or NR1/NR2B receptors and wereexposed to A� oligomers plus glycine in the presence or absence ofD-AP5 (100 �M), MK-801 (50 �M), or memantine (50 �M) (Fig. 3Aand C). Compared to the NR1/NR2A-injected oocytes exposed toA� oligomers plus glycine in the absence of inhibitor (control;0.29 ± 0.03 �A and 8.44 ± 1.23 �C; n = 12) (Fig. 3A), NR1/NR2A-injected oocytes preincubated with D-AP5 showed currents witha significantly reduced mean amplitude (22 ± 3.16% of control,n = 4) and electrical charge (15.64 ± 4.22% of control, n = 4) (Fig. 3B).Similarly, the amplitude and electrical charge were substantially
reduced compared to control when the oocytes were preincubatedwith MK-801 (current amplitude, 4.45 ± 1.11% of control; cur-rent charge, 2.67 ± 0.25% of control; n = 4) or memantine (currentamplitude, 7.93 ± 3.94% of control; current charge, 2.20 ± 1.12% ofcontrol; n = 4) (Fig. 3B).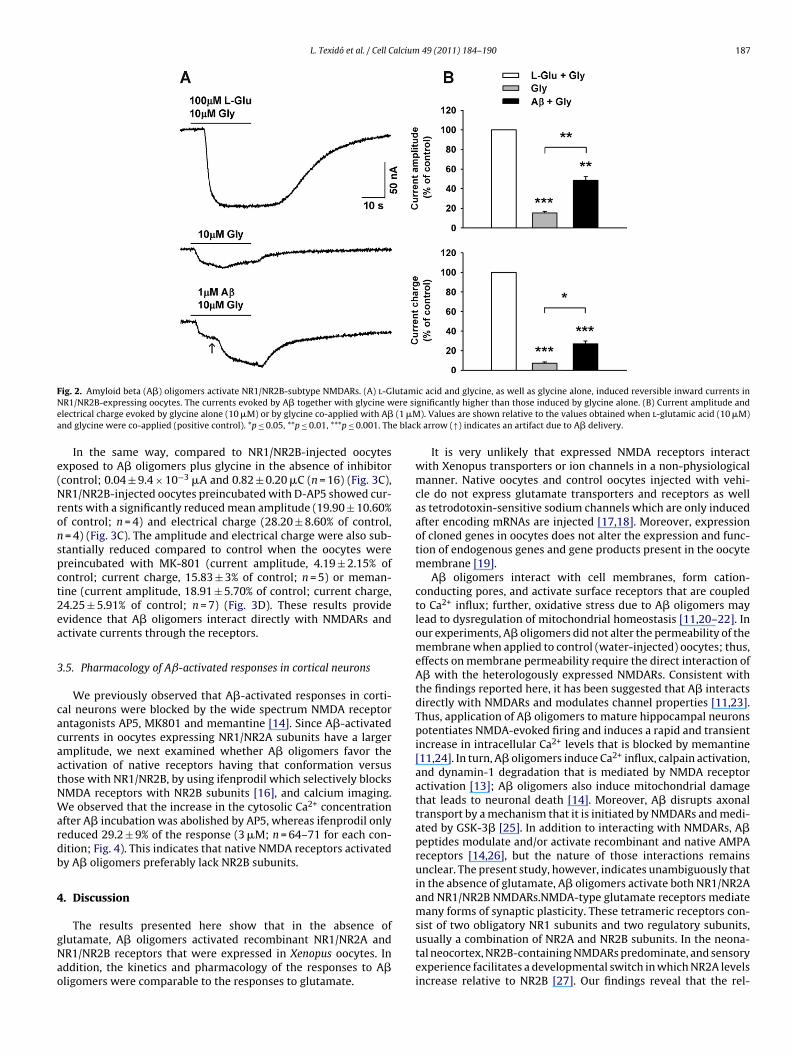
L. Texidó et al. / Cell Calcium 49 (2011) 184–190 187
F lutamN ere sie (1 �Ma e blac
e(Nronspct2ea
3
cacaatNWardb
4
gNao
ig. 2. Amyloid beta (A�) oligomers activate NR1/NR2B-subtype NMDARs. (A) l-GR1/NR2B-expressing oocytes. The currents evoked by A� together with glycine wlectrical charge evoked by glycine alone (10 �M) or by glycine co-applied with A�nd glycine were co-applied (positive control). *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001. Th
In the same way, compared to NR1/NR2B-injected oocytesxposed to A� oligomers plus glycine in the absence of inhibitorcontrol; 0.04 ± 9.4 × 10−3 �A and 0.82 ± 0.20 �C (n = 16) (Fig. 3C),R1/NR2B-injected oocytes preincubated with D-AP5 showed cur-
ents with a significantly reduced mean amplitude (19.90 ± 10.60%f control; n = 4) and electrical charge (28.20 ± 8.60% of control,= 4) (Fig. 3C). The amplitude and electrical charge were also sub-
tantially reduced compared to control when the oocytes werereincubated with MK-801 (current amplitude, 4.19 ± 2.15% ofontrol; current charge, 15.83 ± 3% of control; n = 5) or meman-ine (current amplitude, 18.91 ± 5.70% of control; current charge,4.25 ± 5.91% of control; n = 7) (Fig. 3D). These results providevidence that A� oligomers interact directly with NMDARs andctivate currents through the receptors.
.5. Pharmacology of Aˇ-activated responses in cortical neurons
We previously observed that A�-activated responses in corti-al neurons were blocked by the wide spectrum NMDA receptorntagonists AP5, MK801 and memantine [14]. Since A�-activatedurrents in oocytes expressing NR1/NR2A subunits have a largermplitude, we next examined whether A� oligomers favor thectivation of native receptors having that conformation versushose with NR1/NR2B, by using ifenprodil which selectively blocksMDA receptors with NR2B subunits [16], and calcium imaging.e observed that the increase in the cytosolic Ca2+ concentration
fter A� incubation was abolished by AP5, whereas ifenprodil onlyeduced 29.2 ± 9% of the response (3 �M; n = 64–71 for each con-ition; Fig. 4). This indicates that native NMDA receptors activatedy A� oligomers preferably lack NR2B subunits.
. Discussion
The results presented here show that in the absence oflutamate, A� oligomers activated recombinant NR1/NR2A andR1/NR2B receptors that were expressed in Xenopus oocytes. Inddition, the kinetics and pharmacology of the responses to A�ligomers were comparable to the responses to glutamate.
ic acid and glycine, as well as glycine alone, induced reversible inward currents ingnificantly higher than those induced by glycine alone. (B) Current amplitude and). Values are shown relative to the values obtained when l-glutamic acid (10 �M)
k arrow (↑) indicates an artifact due to A� delivery.
It is very unlikely that expressed NMDA receptors interactwith Xenopus transporters or ion channels in a non-physiologicalmanner. Native oocytes and control oocytes injected with vehi-cle do not express glutamate transporters and receptors as wellas tetrodotoxin-sensitive sodium channels which are only inducedafter encoding mRNAs are injected [17,18]. Moreover, expressionof cloned genes in oocytes does not alter the expression and func-tion of endogenous genes and gene products present in the oocytemembrane [19].
A� oligomers interact with cell membranes, form cation-conducting pores, and activate surface receptors that are coupledto Ca2+ influx; further, oxidative stress due to A� oligomers maylead to dysregulation of mitochondrial homeostasis [11,20–22]. Inour experiments, A� oligomers did not alter the permeability of themembrane when applied to control (water-injected) oocytes; thus,effects on membrane permeability require the direct interaction ofA� with the heterologously expressed NMDARs. Consistent withthe findings reported here, it has been suggested that A� interactsdirectly with NMDARs and modulates channel properties [11,23].Thus, application of A� oligomers to mature hippocampal neuronspotentiates NMDA-evoked firing and induces a rapid and transientincrease in intracellular Ca2+ levels that is blocked by memantine[11,24]. In turn, A� oligomers induce Ca2+ influx, calpain activation,and dynamin-1 degradation that is mediated by NMDA receptoractivation [13]; A� oligomers also induce mitochondrial damagethat leads to neuronal death [14]. Moreover, A� disrupts axonaltransport by a mechanism that it is initiated by NMDARs and medi-ated by GSK-3� [25]. In addition to interacting with NMDARs, A�peptides modulate and/or activate recombinant and native AMPAreceptors [14,26], but the nature of those interactions remainsunclear. The present study, however, indicates unambiguously thatin the absence of glutamate, A� oligomers activate both NR1/NR2Aand NR1/NR2B NMDARs.NMDA-type glutamate receptors mediatemany forms of synaptic plasticity. These tetrameric receptors con-
sist of two obligatory NR1 subunits and two regulatory subunits,usually a combination of NR2A and NR2B subunits. In the neona-tal neocortex, NR2B-containing NMDARs predominate, and sensoryexperience facilitates a developmental switch in which NR2A levelsincrease relative to NR2B [27]. Our findings reveal that the rel-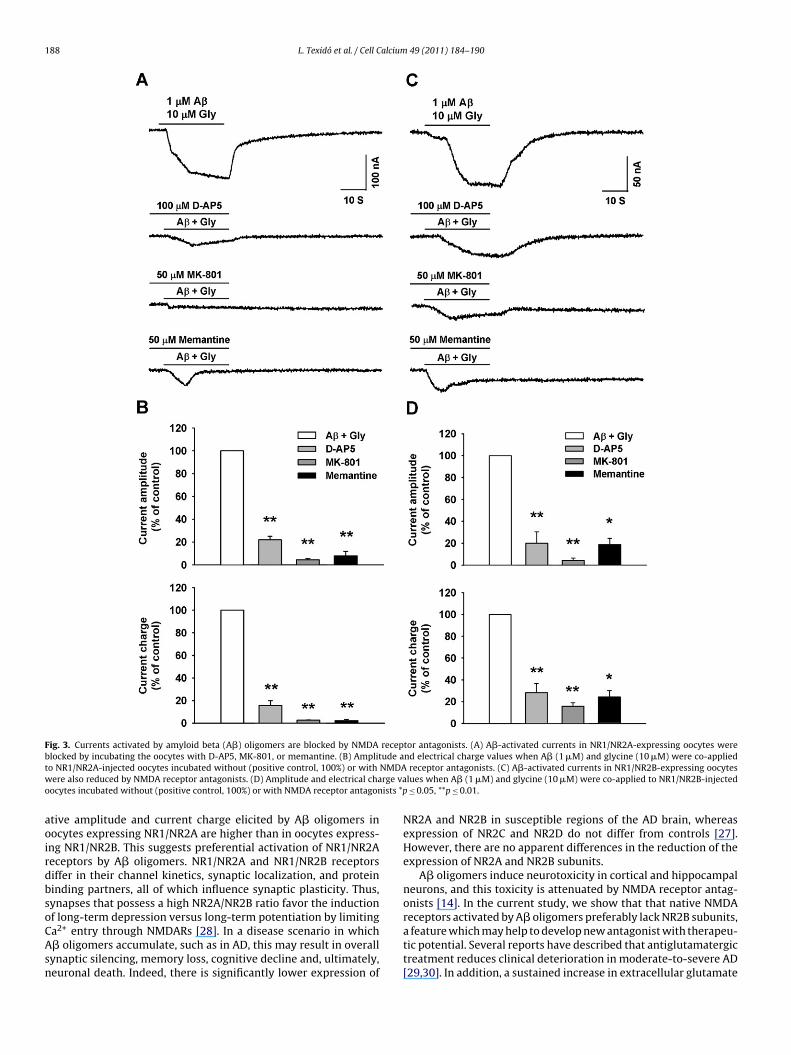
188 L. Texidó et al. / Cell Calcium 49 (2011) 184–190
Fig. 3. Currents activated by amyloid beta (A�) oligomers are blocked by NMDA receptor antagonists. (A) A�-activated currents in NR1/NR2A-expressing oocytes wereb ude at NMDAw rge vao ists *p
aoirdbsoCAsn
locked by incubating the oocytes with D-AP5, MK-801, or memantine. (B) Amplito NR1/NR2A-injected oocytes incubated without (positive control, 100%) or withere also reduced by NMDA receptor antagonists. (D) Amplitude and electrical cha
ocytes incubated without (positive control, 100%) or with NMDA receptor antagon
tive amplitude and current charge elicited by A� oligomers inocytes expressing NR1/NR2A are higher than in oocytes express-ng NR1/NR2B. This suggests preferential activation of NR1/NR2Aeceptors by A� oligomers. NR1/NR2A and NR1/NR2B receptorsiffer in their channel kinetics, synaptic localization, and proteininding partners, all of which influence synaptic plasticity. Thus,ynapses that possess a high NR2A/NR2B ratio favor the inductionf long-term depression versus long-term potentiation by limiting
a2+ entry through NMDARs [28]. In a disease scenario in which� oligomers accumulate, such as in AD, this may result in overallynaptic silencing, memory loss, cognitive decline and, ultimately,euronal death. Indeed, there is significantly lower expression ofnd electrical charge values when A� (1 �M) and glycine (10 �M) were co-appliedreceptor antagonists. (C) A�-activated currents in NR1/NR2B-expressing oocytes
lues when A� (1 �M) and glycine (10 �M) were co-applied to NR1/NR2B-injected≤ 0.05, **p ≤ 0.01.
NR2A and NR2B in susceptible regions of the AD brain, whereasexpression of NR2C and NR2D do not differ from controls [27].However, there are no apparent differences in the reduction of theexpression of NR2A and NR2B subunits.
A� oligomers induce neurotoxicity in cortical and hippocampalneurons, and this toxicity is attenuated by NMDA receptor antag-onists [14]. In the current study, we show that that native NMDAreceptors activated by A� oligomers preferably lack NR2B subunits,
a feature which may help to develop new antagonist with therapeu-tic potential. Several reports have described that antiglutamatergictreatment reduces clinical deterioration in moderate-to-severe AD[29,30]. In addition, a sustained increase in extracellular glutamate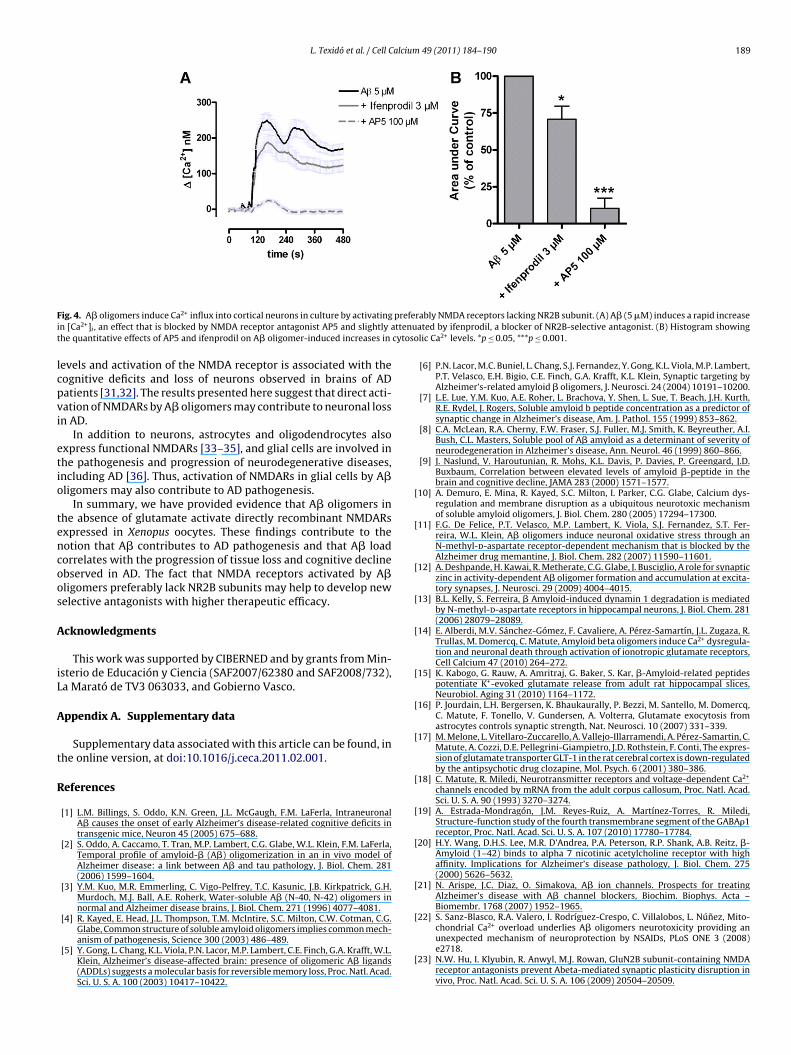
L. Texidó et al. / Cell Calcium 49 (2011) 184–190 189
F prefei ttenut cytoso
lcpvi
etio
tencoos
A
iL
A
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
ig. 4. A� oligomers induce Ca2+ influx into cortical neurons in culture by activatingn [Ca2+]i , an effect that is blocked by NMDA receptor antagonist AP5 and slightly ahe quantitative effects of AP5 and ifenprodil on A� oligomer-induced increases in
evels and activation of the NMDA receptor is associated with theognitive deficits and loss of neurons observed in brains of ADatients [31,32]. The results presented here suggest that direct acti-ation of NMDARs by A� oligomers may contribute to neuronal lossn AD.
In addition to neurons, astrocytes and oligodendrocytes alsoxpress functional NMDARs [33–35], and glial cells are involved inhe pathogenesis and progression of neurodegenerative diseases,ncluding AD [36]. Thus, activation of NMDARs in glial cells by A�ligomers may also contribute to AD pathogenesis.
In summary, we have provided evidence that A� oligomers inhe absence of glutamate activate directly recombinant NMDARsxpressed in Xenopus oocytes. These findings contribute to theotion that A� contributes to AD pathogenesis and that A� loadorrelates with the progression of tissue loss and cognitive declinebserved in AD. The fact that NMDA receptors activated by A�ligomers preferably lack NR2B subunits may help to develop newelective antagonists with higher therapeutic efficacy.
cknowledgments
This work was supported by CIBERNED and by grants from Min-sterio de Educación y Ciencia (SAF2007/62380 and SAF2008/732),a Marató de TV3 063033, and Gobierno Vasco.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.ceca.2011.02.001.
eferences
[1] L.M. Billings, S. Oddo, K.N. Green, J.L. McGaugh, F.M. LaFerla, IntraneuronalA� causes the onset of early Alzheimer’s disease-related cognitive deficits intransgenic mice, Neuron 45 (2005) 675–688.
[2] S. Oddo, A. Caccamo, T. Tran, M.P. Lambert, C.G. Glabe, W.L. Klein, F.M. LaFerla,Temporal profile of amyloid-� (A�) oligomerization in an in vivo model ofAlzheimer disease: a link between A� and tau pathology, J. Biol. Chem. 281(2006) 1599–1604.
[3] Y.M. Kuo, M.R. Emmerling, C. Vigo-Pelfrey, T.C. Kasunic, J.B. Kirkpatrick, G.H.Murdoch, M.J. Ball, A.E. Roherk, Water-soluble A� (N-40, N-42) oligomers innormal and Alzheimer disease brains, J. Biol. Chem. 271 (1996) 4077–4081.
[4] R. Kayed, E. Head, J.L. Thompson, T.M. McIntire, S.C. Milton, C.W. Cotman, C.G.
Glabe, Common structure of soluble amyloid oligomers implies common mech-anism of pathogenesis, Science 300 (2003) 486–489.[5] Y. Gong, L. Chang, K.L. Viola, P.N. Lacor, M.P. Lambert, C.E. Finch, G.A. Krafft, W.L.Klein, Alzheimer’s disease-affected brain: presence of oligomeric A� ligands(ADDLs) suggests a molecular basis for reversible memory loss, Proc. Natl. Acad.Sci. U. S. A. 100 (2003) 10417–10422.
[
rably NMDA receptors lacking NR2B subunit. (A) A� (5 �M) induces a rapid increaseated by ifenprodil, a blocker of NR2B-selective antagonist. (B) Histogram showinglic Ca2+ levels. *p ≤ 0.05, ***p ≤ 0.001.
[6] P.N. Lacor, M.C. Buniel, L. Chang, S.J. Fernandez, Y. Gong, K.L. Viola, M.P. Lambert,P.T. Velasco, E.H. Bigio, C.E. Finch, G.A. Krafft, K.L. Klein, Synaptic targeting byAlzheimer’s-related amyloid � oligomers, J. Neurosci. 24 (2004) 10191–10200.
[7] L.E. Lue, Y.M. Kuo, A.E. Roher, L. Brachova, Y. Shen, L. Sue, T. Beach, J.H. Kurth,R.E. Rydel, J. Rogers, Soluble amyloid b peptide concentration as a predictor ofsynaptic change in Alzheimer’s disease, Am. J. Pathol. 155 (1999) 853–862.
[8] C.A. McLean, R.A. Cherny, F.W. Fraser, S.J. Fuller, M.J. Smith, K. Beyreuther, A.I.Bush, C.L. Masters, Soluble pool of A� amyloid as a determinant of severity ofneurodegeneration in Alzheimer’s disease, Ann. Neurol. 46 (1999) 860–866.
[9] J. Naslund, V. Haroutunian, R. Mohs, K.L. Davis, P. Davies, P. Greengard, J.D.Buxbaum, Correlation between elevated levels of amyloid �-peptide in thebrain and cognitive decline, JAMA 283 (2000) 1571–1577.
10] A. Demuro, E. Mina, R. Kayed, S.C. Milton, I. Parker, C.G. Glabe, Calcium dys-regulation and membrane disruption as a ubiquitous neurotoxic mechanismof soluble amyloid oligomers, J. Biol. Chem. 280 (2005) 17294–17300.
11] F.G. De Felice, P.T. Velasco, M.P. Lambert, K. Viola, S.J. Fernandez, S.T. Fer-reira, W.L. Klein, A� oligomers induce neuronal oxidative stress through anN-methyl-d-aspartate receptor-dependent mechanism that is blocked by theAlzheimer drug memantine, J. Biol. Chem. 282 (2007) 11590–11601.
12] A. Deshpande, H. Kawai, R. Metherate, C.G. Glabe, J. Busciglio, A role for synapticzinc in activity-dependent A� oligomer formation and accumulation at excita-tory synapses, J. Neurosci. 29 (2009) 4004–4015.
13] B.L. Kelly, S. Ferreira, � Amyloid-induced dynamin 1 degradation is mediatedby N-methyl-d-aspartate receptors in hippocampal neurons, J. Biol. Chem. 281(2006) 28079–28089.
14] E. Alberdi, M.V. Sánchez-Gómez, F. Cavaliere, A. Pérez-Samartín, J.L. Zugaza, R.Trullas, M. Domercq, C. Matute, Amyloid beta oligomers induce Ca2+ dysregula-tion and neuronal death through activation of ionotropic glutamate receptors,Cell Calcium 47 (2010) 264–272.
15] K. Kabogo, G. Rauw, A. Amritraj, G. Baker, S. Kar, �-Amyloid-related peptidespotentiate K+-evoked glutamate release from adult rat hippocampal slices,Neurobiol. Aging 31 (2010) 1164–1172.
16] P. Jourdain, L.H. Bergersen, K. Bhaukaurally, P. Bezzi, M. Santello, M. Domercq,C. Matute, F. Tonello, V. Gundersen, A. Volterra, Glutamate exocytosis fromastrocytes controls synaptic strength, Nat. Neurosci. 10 (2007) 331–339.
17] M. Melone, L. Vitellaro-Zuccarello, A. Vallejo-Illarramendi, A. Pérez-Samartin, C.Matute, A. Cozzi, D.E. Pellegrini-Giampietro, J.D. Rothstein, F. Conti, The expres-sion of glutamate transporter GLT-1 in the rat cerebral cortex is down-regulatedby the antipsychotic drug clozapine, Mol. Psych. 6 (2001) 380–386.
18] C. Matute, R. Miledi, Neurotransmitter receptors and voltage-dependent Ca2+
channels encoded by mRNA from the adult corpus callosum, Proc. Natl. Acad.Sci. U. S. A. 90 (1993) 3270–3274.
19] A. Estrada-Mondragón, J.M. Reyes-Ruiz, A. Martínez-Torres, R. Miledi,Structure-function study of the fourth transmembrane segment of the GABA�1receptor, Proc. Natl. Acad. Sci. U. S. A. 107 (2010) 17780–17784.
20] H.Y. Wang, D.H.S. Lee, M.R. D’Andrea, P.A. Peterson, R.P. Shank, A.B. Reitz, �-Amyloid (1–42) binds to alpha 7 nicotinic acetylcholine receptor with highaffinity. Implications for Alzheimer’s disease pathology, J. Biol. Chem. 275(2000) 5626–5632.
21] N. Arispe, J.C. Diaz, O. Simakova, A� ion channels. Prospects for treatingAlzheimer’s disease with A� channel blockers, Biochim. Biophys. Acta –Biomembr. 1768 (2007) 1952–1965.
22] S. Sanz-Blasco, R.A. Valero, I. Rodríguez-Crespo, C. Villalobos, L. Núnez, Mito-
chondrial Ca2+ overload underlies A� oligomers neurotoxicity providing anunexpected mechanism of neuroprotection by NSAIDs, PLoS ONE 3 (2008)e2718.23] N.W. Hu, I. Klyubin, R. Anwyl, M.J. Rowan, GluN2B subunit-containing NMDAreceptor antagonists prevent Abeta-mediated synaptic plasticity disruption invivo, Proc. Natl. Acad. Sci. U. S. A. 106 (2009) 20504–20509.

1 alcium
[
[
[
[
[
[
[
[
[
[
[
Anat. 210 (2007) 693–702.[35] Y. Bakiri, V. Burzomato, G. Frugier, N.B. Hamilton, R. Káradóttir, D. Attwell,
Glutamatergic signaling in the brain’s white matter, Neuroscience 158 (2009)
90 L. Texidó et al. / Cell C
24] V. Szegedi, G. Juhasz, D. Budai, B. Penke, Divergent effects of A� 1–42 onionotropic glutamate receptor-mediated responses in CA1 neurons in vivo,Brain Res. 1062 (2005) 120–126.
25] H. Decker, K.Y. Lo, S.M. Unger, S.T. Ferreira, M.A. Silverman, Amyloid-beta pep-tide oligomers disrupt axonal transport through an NMDA receptor-dependentmechanism that is mediated by glycogen synthase kinase 3beta in primarycultured hippocampal neurons, J. Neurosci. 30 (2010) 9166–9171.
26] H. Tozaki, A. Matsumoto, T. Kanno, K. Nagai, T. Nagata, S. Yamamoto, T.Nishizaki, The inhibitory and facilitatory actions of amyloid-� peptides on nico-tinic ACh receptors and AMPA receptors, Biochem. Biophys. Res. Commun. 294(2002) 42–45.
27] K. Yashiro, B.D. Philpot, Regulation of NMDA receptor subunit expression andits implications for LTD, LTP, and metaplasticity, Neuropharmacology 55 (2008)1081–1094.
28] M.R. Hynd, H.L. Scott, P.R. Dodd, Differential expression of N-methyl-d-aspartate receptor NR2 isoforms in Alzheimer’s disease, J. Neurochem. 90(2004) 913–919.
29] B. Reisberg, R. Doody, A. Stöffler, F. Schmitt, S. Ferris, H.J. Möbius, for theMemantine Study Group, Memantine in moderate-to-severe Alzheimer dis-ease, N. Engl. J. Med. 348 (2003) 1333–1341.
[
49 (2011) 184–190
30] M.R. Farlow, NMDA receptor antagonists. A new therapeutic approach forAlzheimer’s disease, Geriatrics 59 (2004) 22–27.
31] M.P. Mattson, S.L. Chan, Neuronal and glial calcium signaling in Alzheimer’sdisease, Cell Calcium 34 (2003) 385–397.
32] M.R. Hynd, H.L. Scott, P.R. Dodd, Glutamate-mediated excitotoxicity andneurodegeneration in Alzheimer’s disease, Neurochem. Int. 45 (2004)583–595.
33] A. Verkhratsky, F. Kirchhoff, NMDA receptors in glia, Neuroscientist 13 (2007)28–37.
34] C. Matute, E. Alberdi, M. Domercq, M.V. Sánchez-Gómez, A. Pérez-Samartín, A.Rodríguez-Antigüedad, F. Pérez-Cerdá, Excitotoxic damage to white matter, J.
266–274.36] J.J. Rodríguez, M. Olabarria, A. Chvatal, A. Verkhratsky, Astroglia in dementia
and Alzheimer’s disease, Cell Death Differ. 16 (2009) 378–385.
Related Documents











