This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Applied Soil Ecology 49 (2011) 187– 196
Contents lists available at ScienceDirect
Applied Soil Ecology
journa l h o me page: www.elsev ier .com/ locate /apsoi l
Amendment of soils with fresh and post-extraction lavender (Lavandulaangustifolia) and lavandin (Lavandula × intermedia) reduce inoculum ofVerticillium dahliae and inhibit wilt in strawberry
David Yohalem ∗, Thomas PasseyEast Malling Research, East Malling, Kent ME19 6BJ, UK
a r t i c l e i n f o
Article history:Received 7 December 2010Received in revised form 12 May 2011Accepted 13 May 2011
Keywords:HydrosolsTerpenoidsMonoterpenesInoculum densityGrossoMaillette
a b s t r a c t
Soils that were naturally infested with high levels of Verticillium dahliae, the causal agent of wilt diseasein strawberry, were amended with fresh and waste lavender, fresh and waste lavandin, hydrosols gener-ated during essential oil production, BioFenceTM (a mustard-based defatted seedmeal pellet), or water inmicrocosms to estimate their efficacy against microslecrotia of the pathogen. Single chemicals and mix-tures of the chemicals detected from the substrata were also effective in microcosms. The mixtures weremore effective than were the individual chemicals. Microplot evaluation of fresh and waste lavender andlavandin was also made in comparison to BioFenceTM and water controls. Lavandin waste was comparedto BioFenceTM and untreated controls at three sites in field plots that were subsequently planted to straw-berry. Disease incidence and severity were measured over time in the field. All of the Lavandula-basedmaterials could be associated with large reductions in the numbers of viable of microsclerotia recoveredin all but one experiment with greatest effect in microcosms and smaller effects in microplots and fieldplots, as could BioFenceTM pellets. Due to the high levels of inoculum found at all field sites the reductionin pathogen inoculum density was not necessarily associated with a corresponding reduction in diseaseincidence, nor with severity as determined by yield. The monoterpenoids associated with the Lavandulaspp. are of lower volatility than the isothiocyanates associated with crucifer decomposition and weredetected for more than one week after materials were incorporated in soil. This suggests both differencesin mode of action and the possibility of combining either the chemicals or the materials that producethem in order to further enhance efficacy. Several non-target effects were considered: numerosity anddiversity of bacterial and fungal populations; infection by arbuscular mycorrhizal fungi; and functionaldiversity of soil microflora. No persistent non-salient effects were detected.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
With the loss of methyl bromide and other fumigants, straw-berry (Fragaria × annanasa) production has come under increasingthreat of losses due to wilt caused by Verticillium dahliae Kleb. Bio-fumigation using decay products of green manures is one approachthat has been investigated as showing potential for management ofthe disease (Matthiessen and Kierkegaard, 2006), as has breedingfor resistance in the host (Wilhelm, 1981). Microbially-mediatedbiological control has also been observed as having beneficialeffects (Martin and Bull, 2002; Müller et al., 2004). In addition to dis-ease inhibition, additions of organic wastes have positive effects onsoil structure, nutrient availability and inoculum viability (Yohalemet al., unpublished data). Disease threat from V. dahliae is most fre-quently assessed by culturing infested soils and determination of
∗ Corresponding author. Tel.: +44 01732220193.E-mail address: [email protected] (D. Yohalem).
colony forming units of the pathogen per gram of soil. Disease inci-dence and severity in the crop are also used to determine treatmentefficacy.
Earlier work (Yohalem and Hall, 2009) has shown that freshEnglish lavender (Lavandula angustifolia P. Mill.) could reduceinoculum to below the detection limit of the method of Harriset al. (1993). However, the use of fresh lavender is not an eco-nomically viable approach, nor is lavender production consistentwith strawberry agronomy. Spike lavender, L. stoechas, has alsobeen shown to reduce populations of V. dahliae (Mwanza andBlanco-López, 2001; López-Escudero et al., 2007). Lavandin (Lavan-dula × intermedia Emeric ex Loisel), a sterile hybrid of L. angustifoliaand L. latifolia, is grown more abundantly and is more prolific than L.angustifolia. Its chemical profile is similar to, but distinct from thatof lavender (Moon et al., 2007). Both plants are most commonlygrown for the essential oils they produce and the post-extractionwaste material is most frequently burned or composted. Also pro-duced during the extraction of lavender and lavandin oils are lowervalue aqueous hydrosols of different concentrations depending
0929-1393/$ – see front matter © 2011 Elsevier B.V. All rights reserved.doi:10.1016/j.apsoil.2011.05.006

Author's personal copy
188 D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196
on the methods used in the extraction process (primarily varyingwith whether or not the water used in distilling the oil is recycledthrough the distillation apparatus or collected as run-off). Freshand waste lavandin materials all contain monoterpenoid volatilecomponents, albeit in different ratios. Monoterpenes are reportedto act upon membrane integrity, being either directly lethal tomicroorganisms or rendering them susceptible to physical, chemi-cal or biological attack (Mourey and Canillac, 2002; Trombetta et al.,2005).
We report here on microcosm, microplot and field experimentson the chemistry and effects of English lavender, lavandin and theirwastes on viability of microsclerotia and on disease expression insusceptible cultivars of strawberry, as well as on the detection oftheir volatile components from microcosms and from field exper-iments. They are compared to a Brassica-based seedmeal whichhas demonstrated efficacy against the pathogen (Yohalem andHall, 2009; Yohalem et al. Unpublished data). We also examinenon-target effects of soil amendment including mycorrhizal col-onization, functional diversity and phylogenetic diversity in orderto determine if there are positive or negative effects on populationssalient to strawberry production.
2. Methods
2.1. Microcosms
2.1.1. Lavender, lavandin and post-distillation plant materialsExperiments with four replicates were conducted to evalu-
ate the efficacy of lavender (L. angustifolia cv. Maillette), lavandin(L. × intermedia cv. Grosso) and waste materials that had beenextracted for their essential oils from both. These were comparedto unamended treatments in 200 g microcosms using naturallyinfested Marlow Series typical paleo-argillic silt loam. For eachmicrocosm 10 g of the material was cut into small pieces between0.5 and 1.0 cm in length with scissors, briefly ground in a pestle andmortar and mixed with 190 g dry weight of soil moistened with25 ml water in 350 ml glass jars (Down et al., 2004). Each mate-rial was evaluated for microscelerotial population numbers by thewet-sieve method of Harris et al. (1993) after 28 d incubation. Inbrief, 10 g of air-dried sieved (2 mm mesh) soil were shaken for1 h at 175 rpm and the 20–160 �m fraction obtained. This fractionwas resuspended in 20 ml sterile water and distributed over thesurfaces of 20 Petri dishes (9 cm, single vent) into which a selec-tive medium has been poured. The medium contained 2 g NaNO3,1 g KH2PO4, 0.5 g MgSO4·7H2O, 0.5 g KCl, 0.01 g FeSO4·7H2O, 2 gPGA, 1 ml tergitol NPX (Sigma, UK) and 15 g agar (Fluka, UK) perlitre water, adjusted to pH 6.4 prior to addition of the agar. Afterautoclaving, 1% by volume sterile filtered antibiotic solution wasadded (0.06 g streptomycin, 0.06 g chloramphenicol, 0.06 g chlorte-tracycline, 6 mg biotin). Aliquots (1 ml) of the soil suspension werepoured and spread on the surface of the medium and incubated for28 d at 22 ◦C. After washing the soil from the surface of the plate,the microsclerotia were enumerated under a dissecting micro-scope. The method gives a detection threshold of 0.1 colony formingunit g−1.
Analysis of variance (ANOVA) was performed and contrastswere calculated to separate treatment effects (Genstat 9.1, UK).Contrasts were calculated for: the comparison of source materials(lavender vs. lavandin); for waste materials versus fresh; and fortheir interaction. A further experiment comparing effects of freshlavandin and untreated controls after 28 d and 56 d incubations forlavandin and analyzed by repeated measures. Evaluation of resultswas made at = 0.05.
Elemental nutrient analysis of lavandin waste was accomplishedin the following manner: initial drying and grinding of the sample
as described in Faithfull (1986); followed by a wet digest processfor all elements with the exception of boron. This latter process isbased on the digestion process in Faithfull (1986) [Method 43 forNitrogen in Plant Material]: a subsample of the ground material wassubjected to a wet micro-Kjeldahl type digestion process to destroyall organic matter for subsequent ICP-AES emission spectroscopicanalysis of key elements in the plant material, and for the separateanalysis of nitrogen by Fiastar 5000 Flow Injection Analysis (Foss,UK) instrument with the same digest. A separate dry ashing of thepowdered sample was undertaken to determine boron content.
Volatile chemicals were detected from head spaces above thesoils in microcosms. They were sampled by solid phase microex-traction using a polydimethylsiloxane fibre (75 �m; Supelco, UK)inserted through the cover of the jar for 5 min. Samples were takenat intervals up to 14 d. The volatiles were analyzed by gas chro-matography coupled to mass spectrometry (GC–MS) on a HP6890GC and HP5973 Mass Selective Detector (Agilent) fitted with a fusedsilica capillary GC column (30 m × 0.25 mm i.d.) coated with SPB5(250 �m; Supelco). The fibre was desorbed in the injector (220 ◦C)for 0.75 min, and the GC oven temperature programmed at 50 ◦Cfor 2 min then increased at 6 ◦C min−1 to 240 ◦C. Carrier gas washelium (1 ml min−1). Data were captured and processed with theChemstation software.
2.1.2. Evaluation of individual synthetic compoundsTen of the chemicals detected in head spaces above soils
amended with candidate biofumigant materials were evaluatedfor their effects on V. dahliae microsclerotial populations incomparison with water amended controls. The chemicals weredimethyldisulphide, dipropyldisulphide, dimethyltrisulphide, allylisothiocyanate (ITC), phenyl ITC, phenethyl ITC, �- and �-pinene(1:1), 1,8-cineole, camphor and borneol. All chemicals wereobtained from SigmaAldrich (Gillingham, Dorset, UK) except fordimethyltrisulphide (Oxford Chemicals, Billingham, Teeside, UK;now Frutarom (UK) Ltd.). A sterile filter paper (1 cm × 1 cm) placedat the bottom of a glass sample tube (70 mm × 25 mm diameter)was treated with 1 �l of a chemical and covered with 30 g dryweight of naturally heavily infested Fyfield Series 2, a typical argillicbrown earth moistened with 2.5 ml of water. The tube was coveredwith polythene sheet and incubated at 20 ◦C as above. There werefour replicates of each treatment and the control in a completelyrandomized design and samples were taken for estimation of V.dahliae microsclerotial populations at 14 d and 28 d.
2.1.3. Evaluation of mixtures of synthetic compoundsTwo synthetic mixtures were prepared to simulate the chemical
profiles detected from the Brassicaceous materials and L. angusti-folia, respectively. The former was made up of dimethylsulphide(1 �l), dimethyldisulphide (2 �l), allyl ITC (2 �l) and dimethyltrisul-phide (1 �l). The latter contained �-pinene (1 �l), �-pinene (1 �l),3-carene (1 �l), 1,8-cineole (10 �l), camphor (4 �l) and borneol(2 �l). Each mixture was applied to a filter paper, covered withmoistened Fyfield Series 2 soil naturally infested with V. dahliaeand incubated as above. The two mixtures of chemicals were com-pared with soil amended with 1.5 g BioFenceTM and soil with wateronly. There were four replicates of each treatment and the controlin a completely randomized design and estimations of V. dahliaemicrosclerotial populations were made after 14 d and 28 d.
2.1.4. Lavender and lavandin hydrosolsIn separate experiments, lavandin waste (10 g microcosm−1)
that had been stored for 7 months prior to initiation of theexperiment, three lavandin hydrosols, two lavender hydrosols andunamended water controls (23 ml microcosm−1) were comparedin three randomized complete blocks. To each microcosm, 23 mlof hydrosol was added to 200 g soil in a 360 ml glass jar, as above.

Author's personal copy
D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196 189
After treatment, microscosms were incubated for one month andthen assessed for V. dahliae microsclerotial populations. ANOVAwas performed in order to separate mean responses and evaluatedat = 0.05. Single degree of freedom contrasts were calculated andevaluated at = 0.05 for: all treatments against control; hydrosolsversus lavandin waste; and lavandin hydrosols versus lavenderhydrosols.
The composition of the hydrosols was determined by taking thehydrosol (5 ml), saturating it with sodium chloride and extractingthree times with a 1:1 mixture of diethyl ether and petroleum spirit(b.p. 40–60 ◦C; 2 ml). The combined extracts were dried with mag-nesium sulphate, made up to 10 ml and dodecyl acetate (0.1 mg)added as internal standard. The extracts were analyzed by GC–MSon a Saturn 2200 (Varian Ltd., UK) instrument fitted with a fused sil-ica capillary column (30 m × 0.25 mm i.d.) coated with polar DBWax(Supelco) with helium carrier gas (1 ml-min−1) and oven temper-ature held at 50 ◦C for 2 min and then programmed to 250 ◦C at6 ◦C min−1. The injector was at 220 ◦C and injection was splitless.Quantification was by comparison of peak areas with that of theinternal standard.
2.2. Microplots
An experiment was conducted to evaluate the efficacy oflavandin (L. intermedia cv. Grosso), lavender (L. angustifolia cv. Mail-lette) and waste materials from both that had been extracted fortheir essential oils on reducing populations of V. dahliae. Thesewere compared to unamended treatments in similar soils and toBioFenceTM organic fertilizer pellets (Lazzeri et al., 2004) composedof 6% N, 7% P2O5, 2.6% K2O, 4.4% SO3, 0.9% MgO, plus trace elementsand is 84.2% organic matter (Plant Solutions, UK). Five randomizedcomplete blocks with the six treatments were evaluated 28 d afterincorporation of the treatments.
Microplots, 1 m long × 90 cm wide, were established in raisedbeds at East Malling Research in naturally infested Marlow Seriessoil. Approximately 5 cm of material per plot was added to a 20 cmhigh raised bed and manually mixed with a fork. This thickness oflavandin waste represents approximately 3.7 kg with a moisturecontent of approximately two-thirds. An unamended control anda plot to which was added approximately 850 g m−2 of BioFenceTM
pellets were prepared in each bed. Beds were covered and sealedwith PTFE plastic film. Treatments were assigned in a completelyrandom block design, with each bed used as a blocking factor. Pop-ulations of V. dahliae were estimated from bulked soil cores for eachexperimental unit one month after treatment application. ANOVAwas performed for each date and for the entire experiment, wheneach date was treated as a major plot within a split plot experiment.Single degree of freedom contrasts were calculated for: the compar-ison of source materials (lavender vs. lavandin); for waste materialsversus fresh; and for their interaction. Evaluation of results wasmade at = 0.05.
2.3. Field experiments
2.3.1. Effects on V. dahliae and strawberriesField experiments were performed at three locations with dif-
ferent levels of infestation and soil types: Marlow Series typicalpaleo-argillic silt loam; Hamble 1 series, a typical argillic brownearth; and Fyfield 2 series sandy clay loam, a typical argillic brownearth. The first two fields are conventionally managed, the last isorganically managed. V. dahliae populations were estimated fromsoils sampled from six weeks to several months after incorporation.Strawberry runners, most often of the moderately wilt susceptiblecv. Elsanta (class A 17–29 mm) (Hargreaves, UK), were planted intothe beds, usually at least one month post-incorporation of the mate-rials. In one experiment, the ever-bearing cv. Camarillo (Driscoll’s
Genetics Ltd., UK) was grown. Treatments were post-extractionlavandin waste (3.7 kg fresh material m−2, no amendment andBioFenceTM pellets (0.85 kg m−2). All beds were sealed with PTFEplastic. Yields were assessed throughout the production period.Disease incidence as determined by characteristic yellowing ofleaves was observed throughout the strawberry growing season. Onseveral occasions leaves were harvested and petiole pieces platedon 50% potato dextrose agar (Difco, USA) to confirm diagnosis.Repeated measures ANOVA evaluated at = 0.05 was performed foreach of the four experiments and contrasts between untreated andtreated plots and between BioFenceTM and lavandin waste werecalculated. Area under the disease progress curve was estimatedas ˙[(DIt − DIt+1)/2] * [(t + 1) − t)] ((Genstat 9.1); where DI = diseaseincidence and t and t + 1 are two consecutive observation dates.Proportional data were subjected to angular transformation andcount data were log transformed when deemed necessary becauseof inequality of variance.
2.3.2. Detection of chemicals in soilsAllyl isothiocyanate (ITC) and camphor were used as indica-
tors of volatile release from the BioFenceTM and lavandin soilamendments, respectively. Allyl ITC is among the major productsof glucosinolate hydrolysis involved in biofumigation sensu strict(Matthiessen and Kierkegaard, 2006), while camphor is the mostabundant of the monoterpenoids released during lavandin decay.Volatiles were sampled and analyzed, as above. Each of the fiveexperimental units per treatment were sampled and analyzed foreach date at the East Malling Research site (conventional manage-ment, no tunnel).
A sample (50–100 g) of soil from just below the surface to adepth of approx 15 cm was taken from each plot on days 1, 2,3, 6, 9 and 15 after incorporation. The sample was mixed withenough ethyl acetate to cover it (approx 50 ml) and butyl isoth-iocyanate (100 �g) added as internal standard. The mixture wasshaken and then allowed to settle for 4 h. Samples were analyzed byGC–MS on a HP6890 GC and HP5973 Mass Selective Detector (Agi-lent) fitted with a fused silica capillary GC column (30 m × 0.25 mmi.d.) coated with SPB5 (250 �m; Supelco). Injection (220 ◦C) wassplitless and the GC oven temperature was programmed at 50 ◦Cfor 2 min then increasing at 6 ◦C min−1 to 240 ◦C. Carrier gas washelium (1 ml min−1). Data were captured and processed with theChemstation software.
2.3.3. Non-target effectsThree independent methods were used to assess non-target
effects of treatments in the field experiments. Roots of one healthyappearing strawberry plant per experimental unit were clearedwith hot 10% KOH, acidified with 2% HCl and stained with try-pan blue in lactoglycerol (Phillips and Hayman, 1970). Per centcolonization of secondary and tertiary roots was estimated by thegrid method at 40× magnification under a dissecting microscope(Giovannetti and Mosse, 1980) and checked for characteristic struc-tures of arbuscular mycorrhizal infection, primarily vesicles, at 200and 400× magnification under a binocular microscope.
One gram of soil from the composite sample used to evalu-ate V. dahliae microsclerotial populations was dispersed in 50 mlsterile water by maceration (Biomaster 80 Stomacher®, Seward,UK). Aliquots were plated onto semi-selective media: R2A (Difco)amended with 0.1% cycloheximide for oligotrophic bacteria; andPDA (Difco) amended with 0.005% rose bengal and 0.013% chloram-phenicol for culturable fungi (RB). After incubation at 25 ◦C for 24 h,spiral plated (Eddy Jet, IUL Instruments, Spain) populations wereestimated from the plates of R2A, while spread plates of RB wereincubated at 25 ◦C for 3 d in the dark and then enumerated. The bac-terial and fungal population estimates were log-transformed priorto ANOVA and evaluated at = 0.05.
Author's personal copy
190 D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196
Suspensions were centrifuged at low speed (500 g) to removeparticulates and aliquots (120 �L) of the supernatants were intro-duced into each of the 96 wells of BIOLOG ECO plates (Techno-PathDistribution Ltd., Ireland) and incubated at 25 ◦C for communitylevel physiological profiling (CLPP). Readings were made at 590 nmby a Fluostar Galaxy 10 (BMG Labtechnologies Ltd., UK) plate readerstarting 24 h after samples were introduced and then twice daily(morning and evening) for 5 d. Readings were standardized bydividing each reading by the average for the plate and subtractingthe mean absorbance of the three replicate control wells from eachreading; adjusted values < 0 were recorded as 0. Data were sub-jected to multivariate analysis of variance (MANOVA) and principalcomponent analyses (Genstat 9.1). These analyses were evaluatedat = 0.10.
DNA was extracted from 0.5 g subsamples of the compositedbulk soil samples for each experimental unit using a FastDNATM
Spin Kit for Soil (MP Biomedicals, LLC, USA) extraction kit, aftercell disruption using a FastPrepTM apparatus (QBiogene, USA),cleaned using the GeneCleanTM Turbo (QBiogene, USA) witha further removal of PCR inhibitors with polyvinylpyrrolidone(AppliChem GmbH, Germany) in a spin column (Micro Bio SpinChromatograhy Columns, Bio-Rad, USA). Subsequent to the aboveprotocols, extracted DNA was amplified for automated ribosomalintergenic spacer analysis (ARISA) using labelled universal primersand the resulting amplicons were visualized on a transillumina-tor in 1.5% agarose gels containing 0.01% ethidium bromide. Theprokaryote primer PkITSR: 6-FAM – TGACTGCCAAGGCATCCACC,based on the primers ITSReub and the ITSF primer were used toamplify the intergenic spacer regions of prokaryotes Cardinale et al.(2004). The eukaryote primer, Ek18f was replaced with the primerITS1F as suggested by Anderson and Cairney (2004) in order toexclusively amplify fungi among eukaryotes. The prokaryotes werelabelled with FAM and the fungi with VIC, enabling the productsto be combined for capillary electrophoresis. In order to mini-mize sampling biases, relatively large volumes of cleaned DNA(6 �L) were amplified for relatively few amplification cycles. Fora 50 �L PCR reaction the following amounts of reagents were used:0.4 �L TAQ polymerase (Qiagen, UK); 5 �L 10× Master Mix (Qia-gen); 2 �L MgCl2 (Qiagen); 2.5 �L each primer (Vh Bio, UK); 8 �LdNTPs (Invitrogen, UK); 23.6 �L water (Sigma); 6 �L extracted andcleaned DNA. Amplification conditions were: initial denaturationTAQ polymerase (Qiagen, UK) for 10 min at 94 ◦C; followed by 35cycles of 94 ◦C denaturation for 30 s, 57 ◦C amplification for 60 s and72 ◦C extension for 120 s, flowed by a final extension at 72 ◦C for10 min. After visualization, amplicons were separated by fragmentsize using capillary electrophoresis (ABI 3100, Applied Biosystems,
Table 1Colony forming units of Verticillium dahliae after incubation in soil microscosmsamended with fresh and waste English lavender (Lavandula angustifolia) and freshand waste lavandin (L. Intermedia) for 28 d.
Material cfu after 28 d Per cent reductiona
L. intermedia Fresh 0.03 (0.05)b 99.7L. intermedia Waste 0.05 (0.06) 99.4L. angustifolia Fresh 0.00 (0.00) 100L. angustifolia Waste 0.73 (1.36) 93.9
a Relative to water controls (12.1 cfu g−1 V. dahliae).b Mean with standard deviation in parentheses. N = 4 except for fresh L. angustifolia
for which five replicates were used.
UK): 9 �L HiDi was combined with 0.25 �L of the GeneScanTM 2500ROX® Size Standard and 1.2 �L amplified DNA and denatured for3 min at 90 ◦C in each well (all reagents from Applied Biosystems).Amplicon separation was performed by Eurofins MedigenomixGmbH, Germany. Amplicons were scored and summarized inde-pendently for prokaryotes and fungi and ANOVA, evaluated at
= 0.05, was performed on the numbers of the different kindsand on the total numbers of amplicons detected by capillary elec-trophoresis. Correspondence analyses were performed using theSTATlab 3.0 program (SLP InfoWare, France).
3. Results
3.1. Microcosms
3.1.1. Effects on V. dahliae of fresh and waste lavender andlavandin
Comparison of waste and fresh lavender and lavandin. All treat-ments effected > 90% reductions in V. dahliae populations relativeto water controls (P < 0.001), and no significant differences werefound among the treatments after 28 d incubation (Table 1). Thegreatest variability in the assay was in extracted English lavenderwaste. No effect of time of incubation (P = 0.232) was detected whenthe microcosms with fresh lavandin were re-assessed after 56 dincubation.
Chemical analysis of lavandin waste revealed 1.9% N, 0.3% phos-phate, 2.2% K, 0.3% Mg and 1.3% Ca on a dry weight basis and29 mg kg−1 Mn, 32.6 mg kg−1 B, 11.6 mg kg−1 Cu, 31.1 mg kg−1 Znand 148 mg kg−1 Fe, also on a dry weight basis.
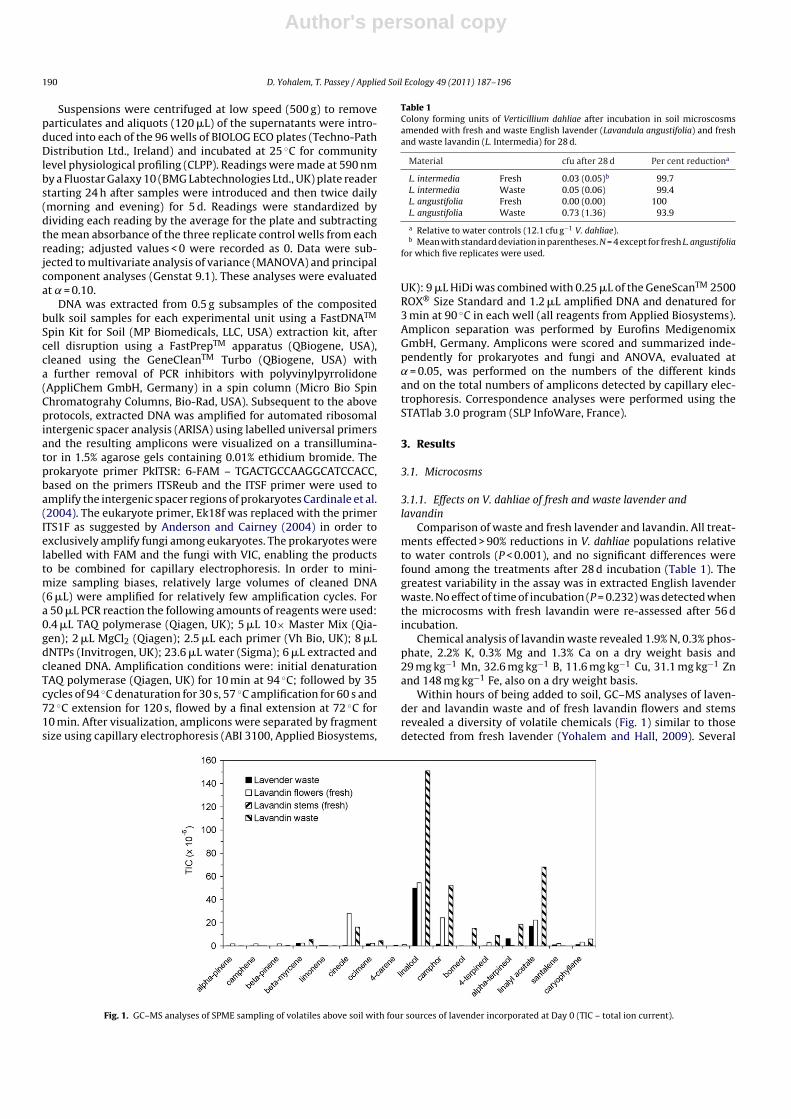
Within hours of being added to soil, GC–MS analyses of laven-der and lavandin waste and of fresh lavandin flowers and stemsrevealed a diversity of volatile chemicals (Fig. 1) similar to thosedetected from fresh lavender (Yohalem and Hall, 2009). Several
Fig. 1. GC–MS analyses of SPME sampling of volatiles above soil with four sources of lavender incorporated at Day 0 (TIC – total ion current).

Author's personal copy
D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196 191
LINALOOL
0
20
40
60
80
100
120
140
160
1614121086420Time (days)
TIC
(X 1
0-6) Lavender waste
Lavandin flowers (fresh)Lavandin waste
Fig. 2. Time course of detection of main volatile components from soil amendedwith lavender sampled by SPME and analyzed by GC–MS (TIC – total ion current;N = 2).
LAVANDIN
0.0
0.1
0.2
0.3
0.4
0.5
0.6
1614121086420Time (days)
TIC
(x 1
0-6)
cineolecamphor
Fig. 3. GC–MS analysis of SPME sampling of linalool above soil with three sources oflavender amendment over 14 d (Mean �g g−1 dry weight; 4 replicates) (TIC – totalion current).
monoterpenoids were detectable in head spaces above amendedsoils for several days after addition to soils (Fig. 2). The essentialoil, linalool, was detected in head spaces above lavandin wastein greater amounts than both lavender waste and fresh lavandinflowers for more than one week after mixing with soil, where theycould be detected for only a couple of days (Fig. 3). Both cineole andcamphor were detected above soils amended with stems of freshlavandin (Fig. 4); the camphor was detected for 5 d; the cineole wasinitially detected at a lower level and persisted for a shorter time.
LAVANDIN STEMS
0.0
0.1
0.2
0.3
0.4
0.5
0.6
1614121086420Time (days)
TIC
(x 1
0-6)
cineolecamphor
Fig. 4. GC–MS analysis of SPME sampling of cineole and camphor above soil withamended with stems of Lavandula × intermedia cv Grosso over 14 d (Mean �g g−1 dryweight; 4 replicates) (TIC – total ion current).
Table 2Microsclerotial populations of Verticillium dahliae in soil samples amended withindividual synthetic chemicals detected from lavender and BioFenceTM at initiationof a microcosm experiment and after 14 d and after 28 d incubation at 20 ◦C (1 �leach in 30 g soil; means of 5 replicates).
Compound V. dahliae population (cfu g−1 soil) (% reduction)
14 d 28 d
Dimethyldisulphide 32.5 (9.2)[24%]b 28.2 (6.2)[34%]Dipropyldisulphide 8.3 (6.3)[81%] 15.1 (8.3)[65%]Dimethyltrisulphide 3.9 (3.2)[91%] 5.1 (3.9)[88%]Allyl ITCa 8.2 (3.7)[81%] 13.8 (3.8)[68%]�- + �-Pinenes 27.5 (11.8)[35%] 18.0 (4.6)[58%]1,8-Cineole 18.8 (6.8)[56%] 10.9 (3.7)[74%]Camphor 15.2 (2.5)[64%] 16.7 (15.2)[61%]Borneol 20.9 (3.9)[51%] 20.7 (4.2)[51%]Pre-treatment populationsc 42.6 (4.0)s.e.d.d (time, 30 df) 3.0s.e.d.d (treatment, 50 df) 4.0
a ITC = isothiocyanate.b Mean (standard deviation) [% reduction relative to control].c Water controls assessed after 14 d incubation.d s.e.d. = standard error of the difference (values determined from an analysis with
a slightly larger suite of chemicals than those presented in the table).
3.1.2. Synthetic chemicals and mixturesPure synthetic chemicals, when added to the soils in concen-
trations similar to those detected from crucifers and lavender, allreduced populations of V. dahliae (P < 0.001) with only dimethyl-trisulphide approaching the efficacy of the natural materials(Table 2). In general, the reductions were stable over time, withlittle evidence for a time × treatment interaction (P = 0.132). Smallincrease in efficacy due to longer incubation was observed for1,8-cineole, dimethyldisulphide and the pinene mixture; a slightrebound effect was observed for dipropyldislphide.
Mixtures of the chemicals in ratios similar to those detectedfrom the parent materials enhanced their efficacy relative to theabove treatments and were nearly as effective (P < 0.001) as theslightly more complex naturally occurring mixtures with a strongimprovement in the efficacy of the lavender mixture from 14 d to28 d incubation (P < 0.001) (Table 3). None of the other treatmentsshowed evidence of a time × treatment interaction.
3.1.3. HydrosolsHydrosols of lavender and lavandin were added directly to soils
because these rates approximated those detected in head spacesabove solid waste amended soils (data not shown). The compo-sition of the hydrosols is given in Table 4. The response of V.dahliae microslcerotia to addition of hydrosol to soil is given inTable 5. No difference was found between aged lavender waste andthe hydrosols (P = 0.819), nor between the lavender and lavandin-derived hydrosols (P = 0.695). All materials significantly reducedpopulations of V. dahliae relative to controls (P < 0.001).
Table 3Microsclerotial populations of Verticillium dahliae in soil samples amended withmixtures of synthetic chemicals simulating those from Brassicaceous materialsa orL. angustifolia in comparison to BioFenceTM and a water control after 14 d and 28 dincubation at 20 ◦C (means of 4 replicates).
Mixture Amount V. dahliae population (cfu g−1 soil) [% reduction]a
14 d 28 d
Brassica mix 0.2 �l g−1 0.0 (0)[100%] 0.1 (0.1)[99%]Lavender mix 0.63 �l g−1 9.9 (1.2)[6%] 0.2 (0.3)[98%]BioFenceTM 50 mg g−1 0.8 (0.4)[92%)] 0.0 (0)[100%]Water 10.5 (0.7) 11.6 (2.3)
a Mean (standard deviation) [% reduction relative to water control].

Author's personal copy
192 D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196
Table 4Retention time and composition of lavandin (1–3) and lavender (4,5) hydrosols ana-lyzed by GC–MS (calculated by comparison with the peak area of dodecyl acetateinternal standard).
RT (min) Compound % Composition
1 2 3 4 5
5.61 (Z)-3-hexenol 0.7 0.1 0.1 0.1 0.05.84 1-hexanol 0.6 0.4 0.3 0.8 0.38.58 3-octen-1-ol 0.6 0.7 0.6 4.3 1.99.55 Cineole 8.2 21.8 16.3 2.3 2.5
11.02 cis-linalool oxide 2.7 2.7 1.2 0.4 0.511.42 trans-linalool oxide 0.9 0.4 0.3 0.4 0.311.69 Linalool 49.0 42.3 50.1 82.5 69.312.81 Camphor 22.4 24.0 21.7 0.8 1.813.34 Borneol 5.7 2.8 3.5 1.8 1.913.67 4-terpineol 5.1 2.4 2.7 5.8 19.513.99 �-terpineol 4.2 2.4 3.2 0.8 2.0
Table 5Colony forming units of Verticillium dahliae after incubation in soil microscosmsamended with lavender and lavandin hydrosols or waste lavandin for 28 d.
Treatment cfu after 28 da Per cent reductionb
Water 9.7 (2.0)L. angustifolia hydrosol 1 3.9 (0.6) 59.8L. angustifolia hydrosol 2 4.4 (2.3) 54.6L. intermedia hydrosol 1 2.5 (3.0) 75.3L. intermedia hydrosol 2 5.0 (1.7) 48.5L. intermedia hydrosol 3 3.9 (1.9) 59.8L. intermedia waste 3.7 (1.7) 61.9s.e.d. 1.44F-prob 0.010
a Mean for 3 completely randomized blocks. Standard deviation in parentheses.b Relative to water controls.
3.2. Microplots
Comparison of colony forming units of V. dahliae in response toamendment of soils with fresh and post-extraction lavender andlavandin was made in microplots (Table 6). After one month in themicroplots, no difference was found between the BioFenceTM seed-meal pellets, the fresh lavender and the waste lavender treatments(P = 0.626), between the fresh and the waste lavender (P = 0.472),nor between the fresh and waste lavandin (P = 0.741) and all treat-ments were associated with significant decreases in viability ofmicrosclerotia (P < 0.001). The lavandin was more effective than thelavender in reducing inoculum density of the pathogen (P = 0.044).
3.3. Field experiments
3.3.1. Effects on V. dahliae populations, disease incidence andseverity in strawberries
Probably because of the higher variability found in field scaleexperiments, significant differences were found between controls,lavandin waste amended and BioFenceTM amended treatments in
Table 6Colony forming units (cfu) of Verticillium dahliae one month after incorporation oflavender (fresh and waste), lavandin (fresh and waste), BioFenceTM seedmeal pelletsand water in microplots.
Treatment V. dahliae cfua Per cent reductionb
Control 12.4 (3.7)BioFenceTM 7.4 (3.3) 41.4L. angustifolia (fresh) 5.0 (2.1) 59.7L. angustifolia (waste) 6.4 (7.1) 48.4L. intermedia (fresh) 1.1 (1.7) 91.2L. intermedia (waste) 2.0 (2.2) 83.9
a Mean for 5 completely randomized blocks. Standard deviation in parentheses.b Per cent reduction relative to control.
0.00
0.05
0.10
0.15
0.20
0.25
181614121086420
Mea
n (μ
g g-
1±S
E)
Time (days)
CONTROLallyl ITCcamphor
0
1
2
3
181614121086420
Mea
n ca
mph
or (μ
g g-
1±S
E)
Time (days)
LAVANDIN
Fig. 5. GC–MS of soil extracts collected from a field experiment (East Malling)amended with Lavandula × intermedia cv Grosso, BioFenceTM seedmeal pellets orunamended over 16 d (Mean �g g−1 dry weight; 3 replicates) (TIC – total ion current).Note the differences in scale of the y-axes.
the numbers of colony forming units of V. dahliae in only one of theexperiments (Table 7). However, in all three experiments, signifi-cant differences were found among treatments in terms of diseaseincidence (P = 0.048; P = 0.022; P = 0.021), with lavandin reducingthe numbers of diseased plants in two of three trials and BioFenceTM
pellets in two of three. The reduction in wilt incidence was not sup-ported by corresponding increases in yield, where differences weredetected in only one experiment. Areas under the disease progresscurve were significantly reduced by lavandin amendments underorganic management, but not under conventional management.
Soils were sampled from the day after incorporation for twoweeks. Samples were analyzed for camphor and ITC concentrations.Allyl ITC was detected in control plots for three d after incorpora-tion of BioFenceTM (Fig. 5) indicating some lateral diffusion throughthe soil matrix, but at very low concentrations. Much higher con-centrations were detected in soils from BioFenceTM amended plots,in which an initial burst of activity disappeared after three d in themoist soil. Camphor was also detected in the controls, although thequantity was even lower than for the more volatile ITCs, with anincrease at d four after incorporation. In lavandin-amended soils,the quantity of camphor increased over time, and peaked around12 d after amendment.
3.3.2. Non-target effectsNo differences were found in degree of arbuscular mycorrhizal
colonization among the treatments (P = 0.654; P = 0.237; P = 0.829)for the three experiments. Mean colonization was approximately53%, 58% and 41% for the three experiments across treatments asassessed by the grid intersect method.
No differences in log transformed numbers of bacteria weredetected for the experiments in the conventionally managed soils(P = 0.725; P = 0.331) or fungi enumerated on RB media in any

Author's personal copy
D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196 193
Table 7Colony forming units of Verticillium dahliae, disease incidence, area under the disease progress curves and disease severity (yields) for strawberries in field experiments.
Treatment V. dahliae cfua Angular transformationof percentage wiltedplants (17 June)b
Area under diseasecurvec
Total yield class 1fruit (g plant−1)
(a) Conventional management cv. Elsanta, no tunnel, East Malling Research, KentControl 6.9 (4.0) 16.0 (7.6) 3104 212BioFenceTM 4.5 (2.2) 10.6 (6.4) 3182 567L.intermedia waste 5.4 (3.6) 3.0 (0.3) 2375 101s.e.d. 2.3 4.74 471.6 76.5F-prob 0.595 0.048 0.229 <0.001
(b) Conventional management cv. Camarillo, tunnel, Faversham, KentControl 0.7 (0.4) 53.7 (64.9) 4072 74BioFenceTM 0.9 (0.5) 40.5 (42.2) 3300 85L. intermedia waste 1.8 (0.7) 46.8 (53.2) 3733 108s.e.d. 0.328 3.35 264.2 30.4F-prob 0.035 0.022 0.070 0.565
(c) Organic management cv. Elsanta, no tunnel, Tuesley, SurreyControl 7.3 (5.2) 18.9 (10.5) 2751 129BioFenceTM 3.7 (0.8) 21.2 (13.1) 2324 123L. intermedia waste 7.9 (1.9) 10.4 (3.3) 2331 21s.e.d. 1.8 3.15 356 38.8F-prob 0.091 0.021 0.543 0.041
a Means with standard errors in parentheses.b Means and s.e.d. presented on angular scale, with back-transformed (%) means alongside in parentheses.c Number of wilted plants over time.
of the soils (P = 0.093; P = 0.324; P = 0.482) were detected amongtreatments within experiments. Data are absent for the bacterialpopulation estimated for the organically managed soil.
Principal components analysis uses the correlations amongmany independent variables to develop a small set of componentswhich empirically summarize the correlations among them. Thus,many variables can be expressed in fewer dimensions in order tobetter grasp the distribution of the variation. In two of the threeexperiments, no differences in patterns or rates of metabolismwere detected for the CLPP analyses. After 96 h incubation, prin-cipal components analysis grouped the BioFenceTM treated soilsseparately from those treated with lavandin waste and those wereseparate from the untreated soils, in the light sandy loam soil ofthe Faversham experiment (Fig. 6). The first principal componentaccounted for 47.75% and the second for 23.29% of the total varia-tion giving 71.04% in these first two. The first principal component
Fig. 6. First two principal components of community level physiological profiles ofsoils amended with Lavandula × intermedia cv Grosso (upward facing triangle, andL for the centroid of the cluster), BioFenceTM seed meal pellets (circles and B for thecentroid of the cluster) and unamended soils (downward facing triangles and C forthe centroid of the cluster. Symbols are for the individual sample data, letters forthe treatment centroids. Ellipses represent approximate 95% confidence limits forthe clusters.
does little to distinguish among treatments (P = 0.916), however,the second does (P = 0.028). ANOVA of the 31 substrates in theBiolog Eco plate revealed that four were individually informa-tive: d-galactonic acid �-lactone (P = 0.035; control < BioFenceTM);�-d-lactose (P = 0.038; control < BioFenceTM = lavandin waste);Tween80 (P = 0.057; control < lavandin); and �-methyl-d-glucoside(P = 0.086; control < BioFenceTM).
Amplicons produced from universal primers were separated ona fragment analyzer An electropherogram of co-amplified opera-tional taxonomic units, illustrates the raw information producedby fragment analysis (Fig. 7). Correspondence analyses indicatedno separation of communities due to treatment in any of theexperiments; the community fingerprints show overlap and fail todistinguish differences (Fig. 8). Neither was there significant differ-ence in the numbers of operational taxonomic units in any of theexperiments (Table 8).
4. Discussion
Lavandula-based materials can reduce the number of viablemicrosclerotia of V. dahlia in soil and inhibit wilt in strawberry.While different aerial parts of Lavandula spp. differ in their chem-ical composition (Aburajai et al., 2005; Angioni et al., 2006) andwith season (Angioni et al., 2006), this was felt to be minor inour experiments with waste materials, since they represent a mix-ture of stems, leaves and flowers and are harvested at the time ofmaximum essential oil concentration, mid-summer in the UK. Theessential oils of Lavandula spp. have well documented antimicro-bial properties (Angioni et al., 2006; Moon et al., 2007; Bakkali et al.,2008; Romeo et al., 2008). Lavandin wastes have been shown tocontain high levels of coumarin (Tiliacos et al., 2008) and coumarinhas been implicated as having herbicidal properties (Haig et al.,2009). Indeed, lavandin extracts have been identified as havingherbicidal properties (Haig et al., 2009), however, to our knowl-edge this is the first report of waste materials having antimicrobialeffects. In addition to the linalool and linalyl acetate that areprevalent in and characterise the commercial essential oil usedin perfumery, waste Lavandula spp. are high in camphor and 1,8cineole, which are individually active against microsclerotia of V.dahliae and more so in combination than alone (Yohalem and Hall,2009). Plants that contain these oxygenated monoterpenes are

Author's personal copy
194 D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196
Fig. 7. Electropherogram of capillary electrophoretic separation of internally transcribed spacer regions of rDNA amplified from soil community DNA extractions. Blue trace:prokaryotes; green trace: eukaryotes. Colors may be seen in the online but not in the print edition of the journal.
abundant (Bakkali et al., 2008) and others, particularly Rosmarinusofficianalis (common rosemary) and Melaleuca alternifolia (tea tree)because of their relative abundance, should be examined for theirefficacy against soil-borne plant pathogens both as fresh materi-als and as wastes after processing. Monoterpenoid compounds actagainst microorganisms by disrupting the lipid fractions of plasma
membranes leading to alterations in permeability (Trombetta et al.,2005).
In addition to the effects of its essential oils the degradation oflignin from the perennial stems of the Lavandula spp. may affectthe survival of microsclerotia, as indicated by Debode et al. (2005).However, lignin content was not measured and this hypothesis was
Fig. 8. Correspondence plots for the first two coordinate axes derived from combined prokaryrotic and eukaryotic amplicons after automated ribosomal intergenic spacer anal-yses. (A) Organically managed plots at Surrey; (B) and (C) Spring and autumn sampling dates from conventionally managed plots at Faversham, Kent; and (D) Conventionallymanaged plots at East Malling. Symbols: diamond = control; circle = lavandin; square = BioFenceTM.

Author's personal copy
D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196 195
Table 8Numbers of operational taxonomic units (OTUs) from automated ribosomal intergenic spacer analysis (ARISA) data.
Experiment # Prokaryotes # Eukaryotes Total
Control Lavandin BioFenceTM Control Lavandin BioFenceTM Control Lavandin BioFenceTM
Organic 19.0 15.8 21.8 8.8 4.4 8.2 27.8 24.0 26.2s.e.d 4.85 1.66 5.79F-prob 0.496 0.058 0.809EMR 2 19.3 16.1 26.6 8.7 8.1 5.8 28.0 24.2 32.3s.e.d 3.61 1.29 3.57F-prob 0.067 0.144 0.154Fav. 1a 10.3 6.8 9.8 8.0 9.5 8.5 16.0 22.8 9.5s.e.d 2.30 1.83 3.21F-prob 0.326 0.718 0.476Fav. 2 8.2 14.8 4.8 7.8 8.0 4.8 16.0 22.8 9.5s.e.d 4.75 3.80 8.07F-prob 0.183 0.656 0.328
a Fav. 1 was sampled in May 2009, Fav. 2 was sampled from the same experiment in October 2009.
not pursued in the present study. While sealing the beds with PTFEinhibits loss of volatiles, the amount of organic material added wasprobably insufficient to induce anaerobic conditions such as thosemonitored by Blok et al. (2000). Some lateral movement of ITCs wasobserved in other experiments (Yohalem et al., unpublished); fur-ther, the bed width would suggest that anaerobic conditions werenever obtained due to lateral diffusion of oxygen.
In both the microcosm and the microplot assays, all Lavandulasolid-based materials reduced inoculum density of V. dahliae by aminimum of 48%. Greater reductions were obtained in microcosmsthan in microplots, which is consistent with the lower variability ofthe systems. The lower volatility of the monoterpenoids comparedto the glucosinolate hydrolysis products and their persistence inthe soil matrix may be reflected by the delayed response of V.dahliae to several of the materials and the mixture. The lavandin,both waste and fresh, reduced inoculum density by at least 80%in the microplots; the waste material was then used for deter-mination of efficacy in larger-scaled field experiments. While thematerial reduced inoculum density in the field, as expected, andwas associated with reduced disease incidence and slower rate ofincrease in wilt, the experiments failed to show a reduction in dis-ease severity. There are several possible reasons for the observedfailure to increase yields. In order to detect changes in microscle-rotial populations, the soils in these experiments were initiallyhighly infested—indeed, so highly infested that one would notpropose growing strawberries on a commercial scale in them with-out conventional chemical fumigation treatment. Even a dramaticreduction, as observed in several of our experiments, left residualpopulations that were too high for strawberry production. Addi-tionally, the Lavandula waste material is fairly coarse and fibrousand did not mix uniformly throughout the beds. Further process-ing of the material and better distribution might have resulted ingreater efficacy. This is supported by the greater efficacy observedin microcosms relative to the microplots and greater reductionsobserved in the microplots compared to the field plots; smallerexperimental units permitted better distribution of material andgreater uniformity of response. Another explanation might involvethe development of anaerobic conditions in the microcosms (Bloket al., 2000), which were more tightly sealed than the microplots,which were better sealed than the field plots.
The biocidal effects of glucosinolate degradation products arewell known, particularly the isothiocyanates (Manici et al., 1997;Lazzeri and Manici, 2001; Matthiessen and Kierkegaard, 2006).BioFenceTM, which is derived from a de-fatted seedmeal of Brassicacarinata, contains the glucosinolate sinegrin, which decomposesinto allyl isothiocyanate upon hydrolysis in the presence of myrosi-nase (Lazzeri et al., 2004). As expected, this chemical was detectedfor several days after incorporation of BioFenceTM. In other exper-
iments (Yohalem and Hall, 2009) several alkyl sulphides weredetected in head spaces above Brassica amended soils, a finding inagreement with Wang et al. (2009). Several of the monoterpenoidsassociated with and detected from lavender and lavandin affectviability of fungi through interference with membrane integrity(Trombetta et al., 2005). Their presence and persistence were cor-related with the reduction in pathogen populations observed in thepresent studies.
That there were no detected differences in mycorrhizal infec-tion rates nor in CLPP profiles (with one exception) is either: areflection of the rapid recovery, resistance or resilience of the soilcommunities to perturbation (van Bruggen and Semenov, 2000); orindicative of the lack of robustness of the methods used to assessnon-target effects of the treatments. However, the similar lack ofobserved differences among treatments in the microbial diversityanalyses as determined by amplification of universal sequencessupports the rapid recovery/resilience hypothesis. Numerosity ofboth bacteria and fungi and lack of differences associated withthe treatments are in agreement with this finding. While there arenumerous reports of positive effects of soil amendment on popula-tions (e.g. Abadie et al., 1998; Borneman, 1999; Weller et al., 2002)and communities (e.g. Hoitink and Boehm, 1999; Weller et al., 2002)of antagonistic microorganisms, this was not found to be the casewith amendment with de-fatted seedmeal pellets nor with wasteof Lavandula × intermedia; it may be that such changes are eithernon-occurring, ephemeral or require higher levels of or repeatedperturbation to be detected by the methods employed in this study.
Further experimentation, particularly on rates of addition andformulation, is needed. Because of the fibrous nature of Lavan-dula spp. and their wastes, fine chopping or formulation as a pelletwould better help its distribution through the upper horizons ofthe soil profile. Mixing the lavandin with another (e.g. a defattedseedmeal) might aid in the pelletization process. If the seedmealis individually efficacious, as with BioFenceTM, the added ben-efit of mixing Lavandula waste to seedmeal would be multiplemodes or timing of action and could enhance overall microsclero-tial reductions. Another approach would be to micro-encapsulateoptimised combinations of the active chemicals, perhaps in a pul-lulan (Kshirsagar et al., 2009) or cyclodextrin (Mourtzinos et al.,2008) matrix. Besides the obvious benefit to growers with Verticil-lium infested field, using post-extraction Lavandula wastes wouldadd value and economically benefit essential oil producers.
Acknowledgements
We thank David Hall and Dudley Farman for detectionand analysis of the allelochemicals (Natural Resources Institute,University of Greenwich, Chatham Maritime, Chatham, Kent) and

Author's personal copy
196 D. Yohalem, T. Passey / Applied Soil Ecology 49 (2011) 187– 196
Gillian Arnold for statistical advice. Christine Lewis is thanked forher assistance in performing some of the microscelrotial viabilitywork. We thank the growers and owners of Langdon Manor, Faver-sham, Kent and Hall Hunter Partnership, Tuesely, Surrey for theuse of their fields. Anna Herrero Langreo is thanked for her assis-tance with the correspondence analyses. Nick Harvey re-designedthe primers used for the ARISA amplifications. The work was fundedas Horticulture LINK Project HL 0177, supported by the Departmentfor Environment, Food and Rural Affairs, the Horticultural Devel-opment Company, Berry World Ltd., KG Growers Ltd., Marks andSpencer PLC, East Malling Ltd., the East Malling Trust for Horticul-tural Research, Farm Advisory Services Team Ltd., Hugh Lowe FarmsLtd., Hall Hunter Partnership, and Plant Solutions Ltd.
References
Abadie, C., Edel, V., Alabouvette, C., 1998. Soil suppressiness to Fusarium wilt: influ-ence of a cover-plant on density and diversity of Fusarium populations. Soil Biol.Biochem. 30, 643–649.
Aburajai, T., Hudiab, M., Cavrini, V., 2005. Chemical composition of the essential oilfrom different aerial parts of lavender (Lavandula coronopofolia Poiert) (Lami-aceae) grown in Jordan. J. Essent. Oil Res. 17, 49–51.
Anderson, I.C., Cairney, J.W., 2004. Diversity and ecology of soil fungal communi-ties: increased understanding through the application of molecular techniques.Environ. Microbiol. 6, 769–779.
Angioni, A., Barra, A., Coroneo, V., Dessi, S., Cabras, P., 2006. Chemical composition,seasonal variability, and antifungal activity of Lavandula stoechas L. ssp. stoechasessential oils from stem/leaves and flowers. J. Agric. Food Chem. 54, 4364–4370.
Bakkali, F., Averbeck, S., Averbeck, D., Idaomar, M., 2008. Biological effects of essen-tial oils: a review. Food Chem. Toxicol. 46, 446–475.
Blok, W.J., Lamers, J.G., Termorshuizen, A.J., Bollen, G.J., 2000. Control of soilborneplant pathogens by incorporating fresh organic amendments followed by tarp-ing. Phytopathology 90, 253–259.
Borneman, J., 1999. Culture-independent identification of microorganisms thatrespond to specific soil stimuli. Appl. Environ. Microbiol. 65, 3398–3400.
Cardinale, M., Brusetti, L., Quatrini, P., Borin, S., Puglia, A.M., Rizzi, A., Zanardini, E.,Sorlini, C., Corselli, C., Daffonchio, D., 2004. Comparison of different primer setsfor use in automated ribosomal intergenic spacer analysis of complex bacterialcommunities. Appl. Environ. Microb. 70, 6147–6156.
Debode, J., Clewes, E., DeBacker, G., Höfte, M., 2005. Lignin is involved in the reduc-tion of Verticillium dahliae var. longisporum inoculum in soil by crop residueincorporation. Soil Biol. Biochem. 37, 301–309.
Down, G.J., Harris, D.C., Murray, R.A., 2004. Destruction of Verticillium dahliae in soilfollowing the addition of sulphur containing volatile compounds potentiallyproduced from Brassica tissues. Agroindustria 3, 293–294.
Faithfull, N.T., 1986. Methods in Agricultural Chemical Analysis: A Practical Hand-book, third edition. CABI, UK, 266 pp.
Giovannetti, M., Mosse, B., 1980. An evaluation of techniques for measuring vesiculararbuscular mycorrhizal infection in roots. New Phytol. 84, 489–500.
Haig, T.J., Haig, T.J., Seal, A.N., Pratley, J.E., An, M., Wu, H., 2009. Lavender as a sourceof novel plant compounds for the development of a natural herbicide. J. Chem.Ecol. 35, 1129–1136.
Harris, D.C., Yang, J.R., Ridout, M.S., 1993. The detection and estimation of Verticiiliumdahliae in naturally infested soils. Plant Pathol. 42, 238–250.
Hoitink, H.A.J., Boehm, M.J., 1999. Biocontrol within the context of soil microbialcommunities: a substrate dependent phenomenon. Ann. Rev. Phytopathol. 37,427–446.
Kshirsagar, A.C., Yenge, V.B., Sarkar, A., Singhal, R.S., 2009. Efficacy of pullulan inemulsification of turmeric oleoresin and its subsequent microencapsulation.Food Chem. 113, 1139–1145.
Lazzeri, L., Leoni, O., Manici, L.M., 2004. Biocidal plant dried pellets for biofumigation.Industrial Crops and Products 20, 59–65.
Lazzeri, L., Manici, L.M., 2001. Allelopathic effect of glucosinilate-containing greenmanure on Pythium sp and total fungal population in soil. Hortscience 36,1283–1289.
López-Escudero, F.J., Mwanza, C., Blanco- López, M.A., 2007. Reduction of Verticilliumdahliae microsclerotia in soil by dried plant residues. Crop Prot. 26, 127–133.
Manici, L.M., Lazzeri, L., Palmieri, S., 1997. In vitro fungitoxic activity of some glu-cosinolates and their enzyme-derived products toward plant pathogenic fungi.J. Agric. Food Chem. 45, 2768–2773.
Martin, F.N., Bull, C.T., 2002. Biological approaches for control of root pathogens ofstrawberry. Phytopathology 92, 1356–1362.
Matthiessen, J.N., Kierkegaard, J.A., 2006. Biofumigation and enhanced biodegrada-tion: opportunity and challenge in soilborne pest and disease management. Crit.Rev. Plant Sci. 25, 235–265.
Moon, T., Cavanagh, H.M.A., Wilkinson, J.M., 2007. Antifungal activity of Australiangrown Lavandula spp. essential oils against Aspergillus nidulans, Trichophytonmentagrophytes Leptosphaeria maculans and Sclerotinia sclerotiorum. J. Essent.Oil Res. 19, 171–175.
Mourey, A., Canillac, N., 2002. Anti-Listeria mononcytogenes activity of essential oilcomponents of conifers. Food Control 13, 289–292.
Mourtzinos, I., Kalogeropoulos, N., Papadakis, S.E., Konstantinou, K., Karathanos,V.T., 2008. Encapsulation of nutraceutical monoterpenes in �-cyclodextrin andmodified starch. J. Food Sci. 73, S89–S94.
Müller, H., Meincke, R., Berg, G., 2004. Strategy to control Verticillium dahliae inoilseed rape using Serratia plymuthica HRO-C48. In: Proceedings of a Meetingof the IOBC/WPRS workin groups Management of Plant Diseases and ArthropodPests by BCAs and Their Integration in Agricultural Systems , Trentino, Italy.
Mwanza, C., Blanco-López, M.A., 2001. Effect of organic amendments on viability ofVerticillium dahliae microsclerotia in soil. In: Proceedings of 8th InternationalVerticillium Symposium, Cordoba , Spain, November 5–8.
Phillips, J.M., Hayman, D.S., 1970. Improved procedure for clearing roots and stainingparasitic and vesicular–arbuscular mycorrhizal fungi for rapid assessment ofinfection. Trans. Br. Mycol. Soc. 55, 157–161.
Romeo, F.V., De Luca, S., Piscopo, A., Poiana, M., 2008. Antimicrobial effect of someessential oils. J. Essent. Oil Res. 20, 373–379.
Tiliacos, C., Gaydou, E.M., Bessière, J.-M., Agnel, R., 2008. Distilled lavandin (Lavan-dula intermedia Emeric ex Loisel) wastes: a rich source of coumarin andherniarin. J. Essent. Oil Res. 20, 412–413.
Trombetta, D., Castelli, F., Grazia Sarpietro, M., Venuti, V., Cristani, M., Daniele,C., Saija, A., Mazzanti, G., Bisignado, G., 2005. Mechanisms of antibac-terial action of three monoterpenes. Antimicrob. Agents Chemother. 49,2474–2478.
van Bruggen, A.H.C., Semenov, A.H., 2000. In search of biological indicators for soilhealth and disease suppression. Appl. Soil Ecol. 15, 13–24.
Wang, D., Rosen, C., Kinkel, L., Cao, A., Tharayil, N., Gerik, J., 2009. Production ofmethyl sulphide and dimethyl sulphide from soil-incorporated plant mate-rials and implications for controlling soilborne pathogens. Plant Soil 324,185–197.
Weller, D.M., Raaijmakers, J.M., Gardener, B.B.M., Thomashow, L.S., 2002. Microbialpopulations responsible for specific suppressiveness to plant pathogens. Ann.Rev. Phytopathol. 40, 309–348.
Wilhelm, S., 1981. Sources and genetics of resistance in field and fruit crop. In: Mace,M.E., Bell, A.A., Beckman, C.H. (Eds.), Fungal Wilt Diseases of Plants. AcademicPress, NY, pp. 299–376.
Yohalem, D., Hall, D., 2009. Selection and partial characterisation of biofumigants formanagement of Verticillium wilt in strawberries. In: Steinberg, C., Edel-Hermann,V., Friberg, H., Alabouvette, C., Tronsmo, A. (Eds.), Multitrophic Interactions inSoil, vol. 42. IOBC/wprs Bulletin, pp. 201–206.
Related Documents











