Amelioration of collagen-induced arthritis by Salix nigra bark extract via suppression of pro-inflammatory cytokines and oxidative stress Shikha Sharma a , Debasis Sahu b , Hasi Rani Das b , Deepak Sharma a,⇑ a School of Life Sciences, Jawaharlal Nehru University, New Delhi 110067, India b Proteomics and Structural Biology Division, Institute of Genomics and Integrative Biology, Mall Road, Delhi 110007, India article info Article history: Received 23 May 2011 Accepted 5 August 2011 Available online 29 September 2011 Keywords: Salix nigra Anti-inflammatory Antioxidant Collagen induced arthritis abstract Our study goals to investigate the anti-arthritic potential of Salix nigra bark methanol extract (SNME) against both inflammation and oxidative stress in the collagen-induced arthritis (CIA) rat model. Results showed that SNME exhibited maximum scavenging activity against superoxide, hypochlorous acid and hydrogen peroxide radicals along with the suppression of lipid peroxidation. Female wistar rats were immunized with porcine type II collagen and treated with SNME (100 mg/kg body weight) for 15 days starting on day 20. SNME significantly inhibited the paw swelling and arthritic score; exhibited maxi- mum CIA inhibition of 93.7% by the end of the experimental period. Administration of SNME to arthritic rats significantly improved the histological findings in joints as evident by reduced infiltration of poly- morphonuclear cells and smooth synovial lining. Roentgenograms of tibiotarsal joints of both SNME and indomethacin-treated rats showed protection against osteophyte formation, soft tissue swelling and bone resorption. Furthermore, levels of inflammatory mediators (nitric oxide, TNF-a, IL-1b, IL-6) measured in both plasma and joint exudates were significantly reduced by SNME treatment. Increased oxidative stress observed in the arthritic animals was also found to be significantly restored in SNME- treated rats. Taken together, our studies clearly indicate the potential of S. nigra as an anti-arthritic agent. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Rheumatoid arthritis (RA) is a chronic and debilitating autoim- mune disorder mainly affecting joints and characterized by synovial hyperplasia, inflammatory cell recruitment, and progressive destruction of cartilage and bone (Choi and Kim, 2008). Collagen-in- duced arthritis (CIA) is a T-cell dependent animal model of RA in which rats develop experimental arthritis after immunization with heterologous type II collagen (Rioja et al., 2004). Both CIA and RA share many clinical, histological and immunological features. This model is widely used to identify and validate the potential therapeu- tic agents for RA (Choi, 2007). Activation of cell mediated immunity in CIA results in the secretion of Th1-related cytokines and free rad- icals that further contribute to articular degeneration. This local and systemic inflammatory response also induce and activate oxidant generating enzymes like NADPH oxidase, xanthine oxidase, myelo- peroxidase, etc. Furthermore, these enzymes produce reactive oxy- gen (superoxide anion) and nitrogen species like nitric oxide which can exaggerate the pathogenesis through initiation of lipid peroxi- dation, alteration of antioxidant enzymes and depletion of glutathi- one (Fay et al., 2006). Thus, it follows that suppression of these inflammatory mediators and oxidative stress can be used to treat RA. Currently prescribed anti-arthritic drug regimen mainly relies on non-steroidal anti-inflammatory drugs (NSAIDs) and disease- modifying anti-rheumatic drugs (DMARDs), which can effectively reduce the symptoms of the disease but simultaneously pose poten- tial side-effects (Yeom et al., 2006). So, there is a strong interest in the development of better anti-inflammatory therapeutic agents for RA that can prevent the progression of disease and confer safe prolonged treatment. One solution to this problem is the use of her- bal therapies, which have been considered safe and effective in alle- viating chronic pain associated with arthritis (Soeken et al., 2003). Willows, from the genus Salix include a number of species of deciduous trees and shrubs, sharing many morphological and phy- tochemical similarities as well as variations, which render them exploited in a much wider therapeutic applications. Salix bark is a rich and inexpensive source of phenolic glycosides (Zaugg et al., 1997), flavonoids, condensed tannins, and polyphenols (Nahrstedt et al., 2007). Salicylates (calculated as salicin) are found in all members of Salix species but Salix daphnoides, Salix fragilis along with Salix purpurea contain the greatest yield (Meier et al., 1988). These constituents are also reported to possess anti- rheumatic, antipyretic, hyperglycemic/hypoglycemic, uricosuric/ antiuricosuric activities, increases prothrombin time, and plas- ma–albumin binding (Khayyal et al., 2005; Minirth et al., 2005). Salicin is believed to be decomposed into salicylic acid in the 0278-6915/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.fct.2011.08.013 ⇑ Corresponding author. Tel.: +91 11 26704508; fax: +91 11 26187338. E-mail address: [email protected] (D. Sharma). Food and Chemical Toxicology 49 (2011) 3395–3406 Contents lists available at SciVerse ScienceDirect Food and Chemical Toxicology journal homepage: www.elsevier.com/locate/foodchemtox

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Food and Chemical Toxicology 49 (2011) 3395–3406
Contents lists available at SciVerse ScienceDirect
Food and Chemical Toxicology
journal homepage: www.elsevier .com/locate / foodchemtox
Amelioration of collagen-induced arthritis by Salix nigra bark extract viasuppression of pro-inflammatory cytokines and oxidative stress
Shikha Sharma a, Debasis Sahu b, Hasi Rani Das b, Deepak Sharma a,⇑a School of Life Sciences, Jawaharlal Nehru University, New Delhi 110067, Indiab Proteomics and Structural Biology Division, Institute of Genomics and Integrative Biology, Mall Road, Delhi 110007, India
a r t i c l e i n f o a b s t r a c t
Article history:Received 23 May 2011Accepted 5 August 2011Available online 29 September 2011
Keywords:Salix nigraAnti-inflammatoryAntioxidantCollagen induced arthritis
0278-6915/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.fct.2011.08.013
⇑ Corresponding author. Tel.: +91 11 26704508; faxE-mail address: [email protected] (D.
Our study goals to investigate the anti-arthritic potential of Salix nigra bark methanol extract (SNME)against both inflammation and oxidative stress in the collagen-induced arthritis (CIA) rat model. Resultsshowed that SNME exhibited maximum scavenging activity against superoxide, hypochlorous acid andhydrogen peroxide radicals along with the suppression of lipid peroxidation. Female wistar rats wereimmunized with porcine type II collagen and treated with SNME (100 mg/kg body weight) for 15 daysstarting on day 20. SNME significantly inhibited the paw swelling and arthritic score; exhibited maxi-mum CIA inhibition of 93.7% by the end of the experimental period. Administration of SNME to arthriticrats significantly improved the histological findings in joints as evident by reduced infiltration of poly-morphonuclear cells and smooth synovial lining. Roentgenograms of tibiotarsal joints of both SNMEand indomethacin-treated rats showed protection against osteophyte formation, soft tissue swellingand bone resorption. Furthermore, levels of inflammatory mediators (nitric oxide, TNF-a, IL-1b, IL-6)measured in both plasma and joint exudates were significantly reduced by SNME treatment. Increasedoxidative stress observed in the arthritic animals was also found to be significantly restored in SNME-treated rats. Taken together, our studies clearly indicate the potential of S. nigra as an anti-arthritic agent.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Rheumatoid arthritis (RA) is a chronic and debilitating autoim-mune disorder mainly affecting joints and characterized by synovialhyperplasia, inflammatory cell recruitment, and progressivedestruction of cartilage and bone (Choi and Kim, 2008). Collagen-in-duced arthritis (CIA) is a T-cell dependent animal model of RA inwhich rats develop experimental arthritis after immunization withheterologous type II collagen (Rioja et al., 2004). Both CIA and RAshare many clinical, histological and immunological features. Thismodel is widely used to identify and validate the potential therapeu-tic agents for RA (Choi, 2007). Activation of cell mediated immunityin CIA results in the secretion of Th1-related cytokines and free rad-icals that further contribute to articular degeneration. This local andsystemic inflammatory response also induce and activate oxidantgenerating enzymes like NADPH oxidase, xanthine oxidase, myelo-peroxidase, etc. Furthermore, these enzymes produce reactive oxy-gen (superoxide anion) and nitrogen species like nitric oxide whichcan exaggerate the pathogenesis through initiation of lipid peroxi-dation, alteration of antioxidant enzymes and depletion of glutathi-one (Fay et al., 2006). Thus, it follows that suppression of these
ll rights reserved.
: +91 11 26187338.Sharma).
inflammatory mediators and oxidative stress can be used to treatRA. Currently prescribed anti-arthritic drug regimen mainly relieson non-steroidal anti-inflammatory drugs (NSAIDs) and disease-modifying anti-rheumatic drugs (DMARDs), which can effectivelyreduce the symptoms of the disease but simultaneously pose poten-tial side-effects (Yeom et al., 2006). So, there is a strong interest inthe development of better anti-inflammatory therapeutic agentsfor RA that can prevent the progression of disease and confer safeprolonged treatment. One solution to this problem is the use of her-bal therapies, which have been considered safe and effective in alle-viating chronic pain associated with arthritis (Soeken et al., 2003).
Willows, from the genus Salix include a number of species ofdeciduous trees and shrubs, sharing many morphological and phy-tochemical similarities as well as variations, which render themexploited in a much wider therapeutic applications. Salix bark isa rich and inexpensive source of phenolic glycosides (Zaugget al., 1997), flavonoids, condensed tannins, and polyphenols(Nahrstedt et al., 2007). Salicylates (calculated as salicin) are foundin all members of Salix species but Salix daphnoides, Salix fragilisalong with Salix purpurea contain the greatest yield (Meier et al.,1988). These constituents are also reported to possess anti-rheumatic, antipyretic, hyperglycemic/hypoglycemic, uricosuric/antiuricosuric activities, increases prothrombin time, and plas-ma–albumin binding (Khayyal et al., 2005; Minirth et al., 2005).Salicin is believed to be decomposed into salicylic acid in the

3396 S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406
human body. The latter is the phytotherapeutic precursor of ace-tylsalicylic acid (ASA), the most important NSAID, commonlyknown as aspirin; hence willow bark is considered a natural formof ASA (Mahdi et al., 2006).
Salix nigra L., commonly called black willow is native to EasternNorth America. The bark of S. nigra is intensely bitter and has beenused as a substitute for quinine in the past. It is also an ingredientof spring tonics to purge the blood, as a remedy for sexual disor-ders and for the treatment of cold, asthma, gout and increased per-spiration. Tea prepared from bark is used to treat gangrene, ulcers,indigestion, and diarrhea (Pullaiah, 2006; Kane, 2006). Recently,the anti-inflammatory activity of the S. nigra bark extract has beeninvestigated in vitro using LPS-stimulated J774 murine macrophagecells (Verma et al., 2009).
To the best of our knowledge, there is no scientific evidence toprove the effectives of S. nigra on the development of CIA. To dem-onstrate the same, we first evaluated the antioxidant and free rad-ical scavenging activity of SNME in different cell-free modelsystems. Thereafter, the therapeutic effects of SNME were checkedin vivo in CIA rat model.
2. Materials and methods
2.1. Chemicals
Curcumin, rutin, 2,2-diphenyl-1-picrylhydrazyl (DPPH), 2,4,6-tris(2-pyridyl)-s-triazine (TPTZ), sodium nitroprusside, ferric chloride, L-ascorbic acid, gallic acidand indomethacin were purchased from Sigma Chemical Co. (St. Louis, MO).Antibodies of TNF-a, IL1b and IL6 were purchased from Peprotech Inc. All otherchemicals and solvents used were of analytical grade.
2.2. Collection of plant material and extract preparation
The bark of S. nigra was procured from Dr. Prakash Joshi, Homeopathic Pharma-copoeia Laboratory (HPL), Ghaziabad, India and authenticated by Dr. H.B. Singh, Na-tional Institute of Science Communication and Information Resources (NISCAIR),Delhi, India. A voucher specimen was deposited in the herbarium; code number:1557/155.
The dried plant material (500 g) was ground to powder form and exhaustivelyextracted thrice with 100% methanol (10 times w/v) at room temperature (RT) for72 h in an orbital shaker. The supernatant was passed through Whatman filter pa-per No. 1, concentrated under reduced pressure on rotary evaporator and lyophi-lized to get the dry dark-brown product (yield 7.5%). The dried extract was thenstored at �20 �C till further use and designated as S. nigra methanolic extract(SNME).
2.3. Preliminary phytochemical screening of the plant extract
A small portion of SNME was used for phytochemical screening in accordancewith the standard procedures of (Ayoola et al., 2008; Dahiru et al., 2006) with someminor modifications.
2.4. Determination of in vitro antioxidant activity
After performing the initial phytochemical screening, SNME was further evalu-ated for antioxidant capability using various in vitro assay systems. In all the exper-iments, curcumin and rutin were taken as reference/standard antioxidants.
Superoxide anion radical scavenging activity of SNME was measured accordingto the previously reported method (Kaur and Arora, 2011). The method of Ogunlanaand Ogunlana (2008) was employed to determine the hydrogen peroxide radicalscavenging activity of SNME. Hypochlorous acid (HOCl) scavenging activity ofSNME was carried out according to the procedure of Hazra et al. (2008). Effect ofvariable concentrations of SNME on inhibition of lipid peroxidation induced byiron/ascorbate system in young rat brain cytosolic fraction was assessed accordingto the method of Kizil et al. (2008). The nitric oxide radical scavenging activity ofSNME was estimated using the method of Temraz and El-tantawy (2008). The abil-ity of SNME to scavenge DPPH radicals was determined by the method of Sanja et al.(2009). The reducing power of SNME was determined according to a previously re-ported method (Mathew and Abraham (2006). Total antioxidant capacity of SNMEwas evaluated by the phosphomolybdenum method described by Prieto et al.(1999). FRAP assay was carried out according to the method of Sharma et al. (2009).
2.5. Determination of polyphenolic content
Total phenolic content of the extract was determined by the Folin–Ciocalteumethod (Wolfe et al., 2003). Total flavonoid content was determined by thealuminium chloride colorimetric method (Lin and Tang, 2007). Method reportedby Afolayan et al. (2008) was used to determine the total proanthocyanidincontent.
2.6. Animals
Female wistar rats of 4–6 weeks of age, weighing 130–150 g were obtainedfrom the central animal facility. Rats were housed separately in cages, fed withstandard rodent chow (Hindustan Lever Limited, India) and water ad libitum. Pho-toperiods equaled 12 h of light and 12 h of darkness daily, with the environmen-tal temperature and humidity maintained at 25 �C ± 2 and 42% ± 5, respectively.All experimental protocols were approved by the Committee for the Purpose ofControl and Supervision of Experimental Animals (CPCSEA) and the InstitutionalAnimal Ethical Committee (IAEC) of Jawaharlal Nehru University, New Delhi,India.
2.7. Dosage
Our preliminary studies on adjuvant-induced arthritic rats with graded doses(100, 250 and 500 mg/kg body weight) of SNME revealed that the arthritic rats trea-ted with 500 mg/kg body weight of extract showed mortality signs after 5 days oforal treatment. Group of rats treated with 250 mg/kg body weight of SNME didnot show any mortality, rather remnants of unmetabolized S. nigra were observedin the peritoneum (data not shown). Upon comparison, 100 mg/kg body weightof SNME was found to be the optimum dose for further evaluation of anti-arthriticefficacy in CIA rats.
2.8. Experimental design
Animals were randomly allocated into the four groups of five animals each:
a) Group I: Animals served as healthy ‘‘control’’ with no collagen immuniza-tion and administered with PBS only.
b) Group II: ‘‘CIA (vehicle only)’’: treated with an equivalent amount of vehiclefor 15 days starting on day 20.
c) Group III: CIA rats treated every 24 h orally with 100 mg/kg body weight ofSNME, dissolved in PBS for 15 days starting on day 20; served as‘‘CIA + SNME’’ group
d) Group IV: CIA rats treated daily with 1 mg/kg body weight of indomethacin,a standard anti-inflammatory drug, dissolved in PBS for 15 days starting onday 20; served as ‘‘CIA + indomethacin’’ groups – positive treatmentcontrol.
2.9. Induction of CIA and SNME treatment
The induction of arthritis in animals was performed as described by Biswaset al. (2009). The gradual onset of arthritis normally started approximately 9 daysafter primary immunization and rats were observed daily for clinical signs of arthri-tis. SNME (100 mg/kg body weight) and indomethacin (1 mg/kg body weight) wereorally administered once daily for 15 days starting from day 20 of the experimentalperiod.
2.10. Macroscopic scoring of CIA
Clinical score, a composite index of disease severity and the number oflimbs affected, was assessed every 4 days to monitor disease progression as de-scribed previously (Cuzzocrea et al., 2000). The scores from each of the fourlimbs were then added to obtain an arthritic score. Since, CIA is more pro-nounced in the hind-limbs, so, the scoring of rat hind-limbs only has been pre-sented here (data of arthritic score is not shown of all the four limbs). Twoindependent observers, blinded to the experimental groups, performed thescoring.
2.11. Paw diameter measurement and Arthritic Index (AI)
Hind paw edema of the rats was monitored every 4th day throughout theexperiment, using a dial thickness gauge. The CIA inhibition and AI were calculatedas described by Coelho et al. (2004).
2.12. Body weight examination
Body weight was measured every 4th day. The change in body weight (%) ofeach individual animal after the onset of arthritis on day 12 was calculated asfollows:

S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406 3397
Change in body weightð%Þ ¼ Body WeightðdayXÞ � Body Weightðday12ÞBody Weightðday12Þ � 100
2.13. Radiological evaluation of CIA
After sacrificing the rats, hind limbs were removed and right hind leg was fixedin 10% buffered formalin for radiological evaluation. All radiographs were takenwith X-ray film (Kodak Diagnostic Film) using MBR-1505R (Hitachi Medical Corpo-ration, Tokyo, Japan). Settings for radiography were 5 mA, 40 kV and 1 s exposure.Films were placed 60 cm below the X-ray source.
2.14. Assessment of arthritis damage through histology
Knee joint was excised from the fixed hind leg, its skin and muscular parts weretrimmed off and subjected to decalcification in 5% nitric acid for 7–10 days, pro-cessed for paraffin embedding, sectioned at 8 l followed by staining with hematox-ylin-eosin and studied using light microscopy.
2.15. Measurement of cytokines in plasma and joints
Blood was collected in EDTA-coated vials followed by centrifugation at2000 rpm for 10 min. Clear plasma aliquots were stored at �80 �C for estimationof cytokines.
For joint cytokine measurements, whole joints were pulverized using a mortarand pestle filled with liquid nitrogen. Powdered tissue was weighed and homoge-nized (1:10 w/v) in 0.02 M Tris–HCl buffer (pH 7.4) containing 20 mM MgCl2,1 mM PMSF, 10 lg leupeptin and aprotinin, 1 mM ATP, 5 mM EDTA, 50 mM DTTand 3 ll protease inhibitor cocktail per 10 ml buffer. Homogenate was first centri-fuged at 4000 rpm, 4 �C for 15 min to remove the debris and bony portions. Super-natant was centrifuged at 16,000 rpm for 15 min and resultant clear supernatantwas collected and stored at �80 �C for estimation of cytokines. Concentrations ofplasma and joint cytokines, TNF-a, IL1-a and IL6 were measured by indirect ELISA.Plates (Nunc) were coated with 10 lg protein/well diluted in carbonate buffer at37 �C for 1–2 h. The plates were washed thrice with PBST (0.2% Tween20 in 1XPBS) and blocked using blocking buffer (1% skimmed milk in 1X PBS) for 1 h at37 �C. After washing, 100 ll/well (0.2 lg/ml) anti-rat polyclonal antibodies againstTNF-a, IL1-b, and IL6 were added and incubated at 37 �C for 1 h. This was followedby washing the wells and then 100 ll/well (1:1000 diluted) anti-rabbit HRP labeledIgG secondary antibody was added. After incubation followed by washing, develop-ing buffer (10 mg/ml orthophenylene diamine dissolved in 150 mM citrate–phos-phate buffer, pH 5.6 with 30% H2O2) was added. The plates were incubated for 5–10 min at RT and the reaction was stopped with the stop solution (2 N H2SO4)and plate was read at 490 nm using Spectra Max Plus from Molecular Devices usingthe SoftMax-Pro software. Bovine serum albumin, at a concentration of 5 lg/ml(100 ll/well) was included as a negative control in each assay performed and thewells incubated with coating buffer instead of plasma were used as the blank.The level of cytokines TNF-a, IL1-b and IL6 were expressed in pg/ml. Each samplewas measured in duplicates and values were derived from the calibration curve.
2.16. Determination of in vivo antioxidant activity
Biochemical studies in joints, liver, spleen and kidney along with plasma.
2.16.1. Preparation of tissue homogenateTo prepare tissue homogenate, dissected organs were removed, rinsed quickly
in cold saline solution and homogenized in 0.05 M Tris–HCl buffer (pH 7.4) to yielda 10% (w/v) homogenate. An aliquot of the homogenate (0.5 ml) was used for assay-ing soluble sulfhydryl group (–SH) while the remainder was centrifuged at 1000gfor 10 min. The resultant supernatant was transferred into pre-cooled centrifuga-tion tubes and centrifuged at 12,000g for 30 min. The supernatant (cytosolic frac-tion), after discarding any floating lipid layer and appropriate dilution, was usedfor assaying the total cytosolic glutathione-S-transferase (GST), lipid peroxidation,protein oxidation, superoxide dismutase (SOD), catalase (CAT), glutathione peroxi-dase (GPx), glutathione reductase (GR) and lactate dehydrogenase (LDH) antioxi-dant enzymes. Homogenates were kept at �80 �C and thawed just before thestart of biochemical estimation.
2.16.2. Estimation of acid soluble sulfhydryl (–SH) groupThe level of acid soluble sulfhydrl group was estimated as total non-protein
sulfhydryl group by the method as described by Singh et al. (2000). Reduced gluta-thione was used as a standard to calculate lM of –SH content/g tissue.
2.16.3. Estimation of lipid peroxidation and protein oxidationThe levels of malondialdehyde and protein carbonyl content, as an index of lipid
peroxidation and protein oxidation, were measured according to the method of Jyo-ti et al. (2009).
2.16.4. Determination of SOD and CAT activitySOD and CAT were assayed utilizing the technique of Singh et al. (2000).
Enzyme activity was expressed as units/mg protein.
2.16.5. Determination of glutathione S-transferase, glutathione peroxidase andglutathione reductase activity
The cytosolic GST, GPx and GR activity was determined spectrophotometricallyat 37 �C according to the procedure of Singh et al. (2000).
2.16.6. Determination of lactate dehydrogenase activityLDH was assayed by measuring the rate of oxidation of NADH at 340 nm,
according to the method of Singh et al. (2000).
2.17. Statistical Analysis
Results of the in vitro antioxidant tests are expressed as the mean ± SD of threeparallel measurements. The IC50 value (the concentration required to inhibit radicalformation by 50%) was calculated using Microsoft excel 2003 and % inhibition wascalculated using the formula:
%Inhibition ¼ ½A0 � AtÞ=A0� � 100
where A0 was the absorbance of the control and At was the absorbance of SNME andstandards. Likewise, the results of animal studies are expressed as mean ± SD (n = 6).Statistical analysis was performed using GraphPad Prism software version 5.00. p-values were calculated using either paired t-tests or one-way ANOVA as applicable.A p-value of 60.05 was regarded as statistically significant.
3. Results
3.1. Preliminary phytochemical screening of SNME
Qualitative phytochemical analyses conducted on SNMEshowed appreciable amount of different types of active constitu-ents, namely saponins, alkaloids, phenols, flavonoids, steroids, tan-nins, terpenoids and moderate amount of hydrolysable tannins,glycosides and cardiac glycosides. The tests were negative for ami-no acids, anthraquinones and volatile oils.
3.2. In vitro antioxidant assays
3.2.1. SNME acts as a superoxide radical (O2�) scavenger
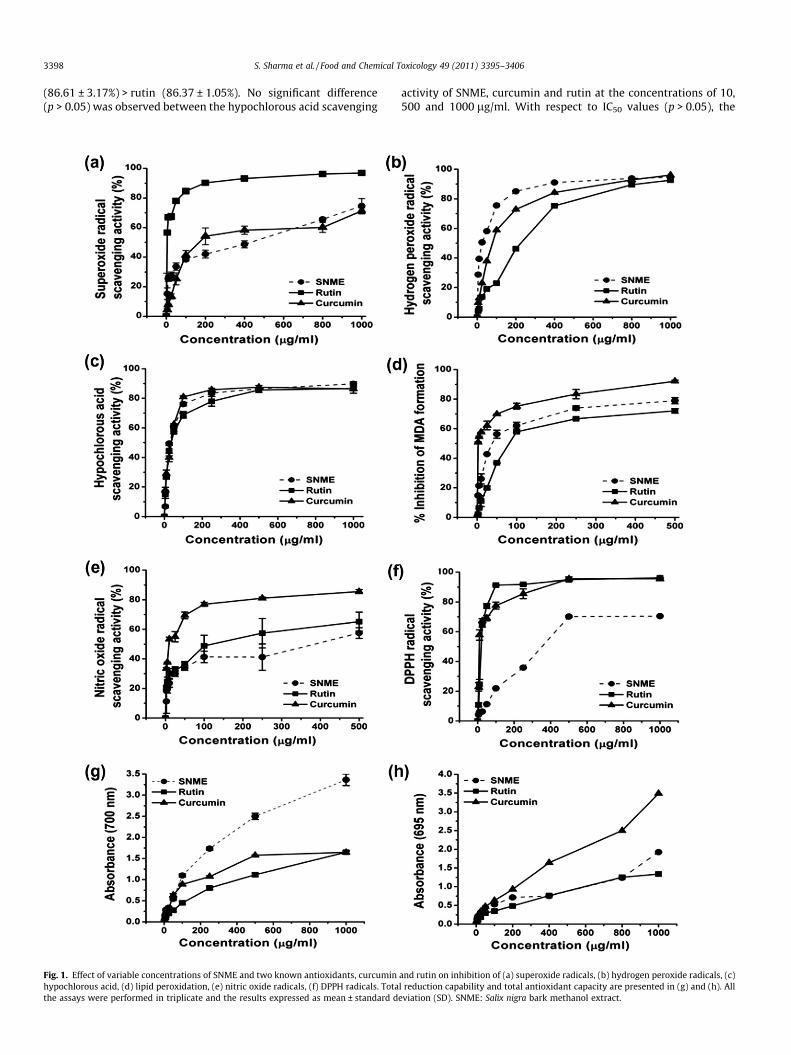
SNME significantly inhibited superoxide radicals in a dose-dependent manner (0–1000 lg/ml) and the results are shown inFig. 1a. At the highest concentration of 1000 lg/ml (p < 0.0001),SNME and curcumin exhibited 74.69 ± 4.97% and 71.27 ± 2.11%scavenging activities, respectively. Rutin (96.91 ± 1.65%) showedthe highest scavenging activities at all the concentrations studied.Significant differences (p < 0.0001) were also observed among theIC50 values and the order was found to be: rutin (1.54 ± 0.35 lg/ml) > SNME (225.52 ± 23.45 lg/ml) > curcumin (243.065 ± 19.76lg/ml).
3.2.2. SNME as a hydrogen peroxide radical scavengerHigh hydrogen peroxide radical scavenging activities of SNME
and the standards were observed at relatively lower concentra-tions (Fig. 1b). At 1000 lg/ml concentration, SNME, curcumin andrutin showed almost equal hydrogen peroxide radical scavengingactivity (p > 0.05) being 94.51 ± 1.43%, 96.08 ± 1.266% and92.67 ± 0.42%, respectively. Rutin showed the lowest activitythroughout, except at 5 lg/ml. With respect to IC50 values(p < 0.0001), the scavenging effect of SNME and both the standardsfollowed the order: SNME (21.86 ± 0.891 lg/ml) > curcumin(74.74 ± 1.58 lg/ml) > rutin (149.3 ± 3.89 lg/ml).
3.2.3. SNME as a hypochlorous acid scavengerAt all the concentrations of greater than 5 lg/ml, SNME exhib-
ited higher hypochlorous acid scavenging activity than curcumin(Fig. 1c). Percent inhibition (p > 0.05) of hypochlorous acid at1000 lg/ml followed the order: SNME (89.85 ± 1.38%) > curcumin
3398 S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406
(86.61 ± 3.17%) > rutin (86.37 ± 1.05%). No significant difference(p > 0.05) was observed between the hypochlorous acid scavenging
Fig. 1. Effect of variable concentrations of SNME and two known antioxidants, curcuminhypochlorous acid, (d) lipid peroxidation, (e) nitric oxide radicals, (f) DPPH radicals. Totathe assays were performed in triplicate and the results expressed as mean ± standard d
activity of SNME, curcumin and rutin at the concentrations of 10,500 and 1000 lg/ml. With respect to IC50 values (p > 0.05), the
and rutin on inhibition of (a) superoxide radicals, (b) hydrogen peroxide radicals, (c)l reduction capability and total antioxidant capacity are presented in (g) and (h). Alleviation (SD). SNME: Salix nigra bark methanol extract.

S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406 3399
scavenging potential followed the order: SNME (38.427 ± 1.107 lg/ml) > rutin (33.355 ± 0.304 lg/ml) > curcumin (41.789 ± 1.61 lg/ml).
3.2.4. Suppression of lipid peroxidation by SNMESNME inhibited lipid peroxidation in a concentration dependent
manner with 78.95% inhibition while rutin demonstrated 72.07%inhibition at a concentration of 500 lg/ml (Fig. 1d). Rutin showedthe lowest peroxidation inhibitory activity throughout. Curcuminexhibited the strongest lipid peroxidation inhibition of 92.14%among all. At the highest concentration, the lipid peroxidationinhibitory activity of SNME and reference antioxidants expressedas percentage inhibition (p < 0.0001) followed the order: curcu-min > SNME > rutin and was 92.14 ± 0.713%, 78.95 ± 0.51% and72.07 ± 1.59%, respectively. The lowest IC50 value (p < 0.0001) of3.22 ± 0.04 lg/ml was observed for curcumin followed by SNME(43.32 ± 4.72 lg/ml), whilst rutin had the highest IC50 value of100.95 ± 6.52 lg/ml indicating that SNME inhibited lipid peroxida-tion more strongly than rutin.
3.2.5. SNME acts as a nitric oxide radical scavengerFig. 1e shows that SNME exhibited a moderate dose-dependent
nitric oxide scavenging activity between 2.5 and 500 lg/ml with anIC50 value of 340.5 ± 11.03 lg/ml (p < 0.0001) compared to11.22 ± 0.5 lg/ml and 146.59 ± 9.835 lg/ml, which was the IC50
value for the references curcumin and rutin, respectively. The po-tency in terms of percentage inhibition (p < 0.01) was: curcumin(85.49 ± 1.36%) > rutin (65.21 ± 6.37%) > SNME (57.43 ± 3.61%).
3.2.6. DPPH free radical scavenging activity of SNMEResults presented in Fig. 1f shows that SNME inhibited DPPH rad-
icals in a dose-dependent manner with an IC50 value (p < 0.0001) of354.83 ± 4.62 lg/ml, higher to that of curcumin (11.91 ± 0.93 lg/ml) and rutin (22.96 ± 2.24 lg/ml). At the concentration of1000 lg/ml, scavenging abilities on DPPH radicals expressed as %inhibition (p < 0.0001) were in descending order: curcumin(95.18 ± 1.328) > rutin (95.87 ± 1.313) > SNME (76.23 ± 0.722).
3.2.7. Reducing power of SNMEThe reducing power of SNME expressed as absorbance values
increased from 0.16 ± 0.02 at 5 lg/ml to 3.36 ± 0.136 at 1000 lg/ml. Its reductive capability was highest at all the concentrationsstudied, except 50 lg/ml, where curcumin showed higher activity(Fig. 1g). SNME showed a rapid increase from 0.54 ± 0.03 to3.36 ± 0.14 in the concentration range of 50–1000 lg/ml. Rutinshowed lowest activity at every concentration used, excluding1000 lg/ml, where both the standards exhibited almost similaractivity (p > 0.05). No significant difference (p > 0.05) was observedbetween the reducing power of SNME and curcumin at 25 lg/mlconcentration. Reductive ability (p < 0.0001) of SNME and the stan-dards at 1000 lg/ml were in the following order: SNME(3.36 ± 0.136) > rutin (1.65 ± 0.04) � curcumin (1.647 ± 0.024).
3.2.8. Total Antioxidant CapacityFig. 1h shows that at relatively lower concentrations of 5–
50 lg/ml, SNME and curcumin exhibited almost similar totalantioxidant capacity (p > 0.05) whereas at the higher concentra-tions of 400 and 800 lg/ml, the activity of SNME and rutin wasfound to be statistically non-significant (p > 0.05). At 1000 lg/ml,this assay exhibited the following sequence of total antioxidantcapacity (p < 0.0001) in terms of optical density: curcumin(3.49 ± 0.047) > SNME (1.92 ± 0.026) > rutin (1.34 ± 0.02). Sincethis phosphomolybdenum method is quantitative, the total antiox-idant capacity of SNME was also expressed as the number of equiv-alents of ascorbic acid and gallic acid as 385.06 ± 0.026 lg/mg and326.32 ± 0.062 lg/mg SNME, respectively.
3.2.9. Total antioxidant activityThe total antioxidant capacity of SNME and reference antioxi-
dants was estimated from FRAP assay through its ability to reduceTPTZ-Fe(III) complex to TPTZ-Fe(II). Rutin (875.20 ± 12.81 lMFe(II)/g extract) exhibited highest antioxidant activity followedby curcumin (762.74 ± 55.41 lM Fe(II)/g extract) and SNME(564.81 ± 25.9 lM Fe(II)/extract).
3.2.10. Total phenolic, flavonoids and proanthocyanidins contentThe phenolic (104.04 ± 0.126 mg gallic acid equivalents per
gram of extract) and flavonoid (191.35 ± 0.012 mg rutin equiva-lents per gram of extract) contents of SNME were markedly higherthan that of the proanthocyanidin (36.95 ± 0.048 mg catechinequivalents per gram of extract) content.
3.2.11. Correlation analysisThe total flavonoid content of SNME was highest as compared to
phenolic and proanthocyanidin content. A positive linear correla-tion was observed between the total phenol and flavonoid contents(R2 = 0.9244, p < 0.0001) and also in between the total phenol andproanthocyanidin (R2 = 0.6888, p = 0.0056) contents, thus indicat-ing a significant positive relationship between the total phenols,flavonoids and proanthocyanidins.
The total phenolic contents were also found to be correlatedwell with the total antioxidant capacity (R2 = 0.9136, p = 0.0008),total reduction capability (R2 = 0.9472, p < 0.0001), in vitro lipidperoxidation (R2 = 0.8902, p = 0.0014) of SNME and its scavengingeffects on hypochlorous acid (R2 = 0.8309, p < 0.0006), superoxide(R2 = 0.8911, p = 0.0014), and hydrogen peroxide (R2 = 0.7917,p = 0.0073). The antioxidant activities of SNME measured asin vitro lipid peroxidation inhibition were significantly correlatedwith total antioxidant capacity (R2 = 0.9846, p < 0.0001), totalreduction capability (R2 = 0.7343, p = 0.0066) and its scavengingactivities on superoxide radical (R2 = 0.9299, p = 0.0019), hypochlo-rous acid (R2 = 0.9662, p < 0.0001), hydrogen peroxide (R2 = 0.9608,p = 0.0006) radicals.
3.3. In vivo anti-inflammatory activity
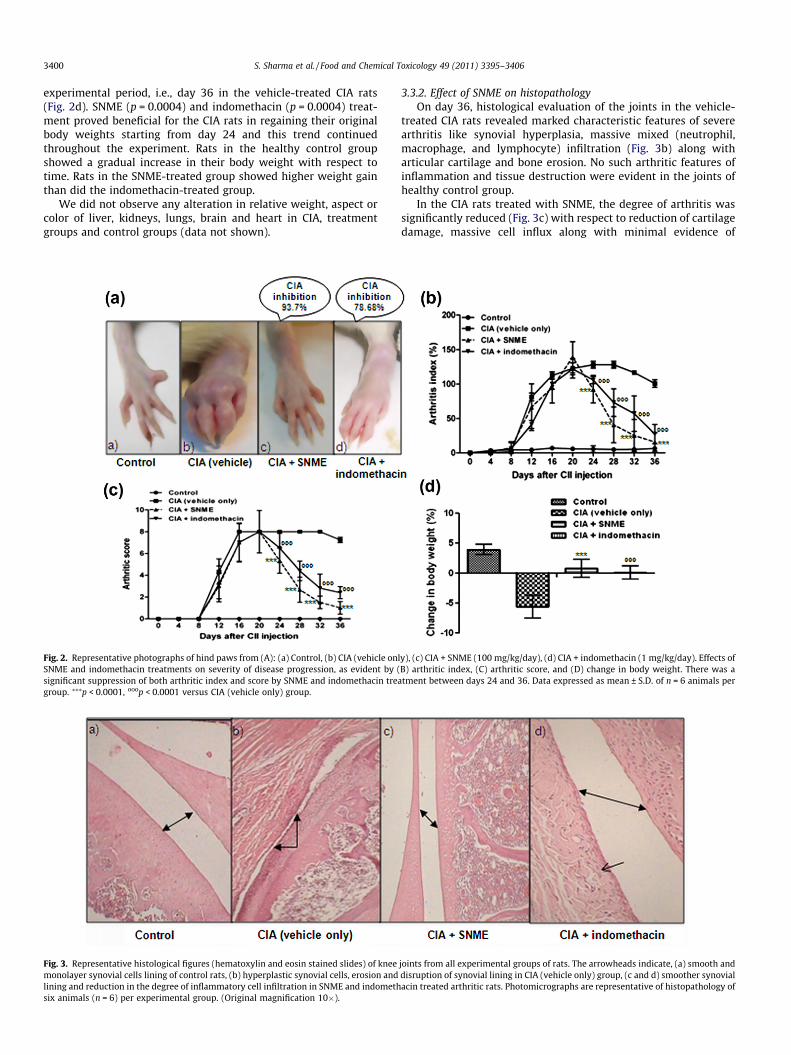
3.3.1. Effect of SNME on3.3.1.1. Kinetics of the clinical progression of CIA. Experimentalarthritis developed in the rats immunized with type II collagenas shown in Fig. 2a. Clinical signs of arthritis (mild periarticularerythema and edema) accompanied by the redness and swellingof the joints first appeared in hind paws approximately 10 dayspost challenge. The disease progression increased in frequencyand severity in a time-dependent manner and maintained a pla-teau of the peak of CIA response from day 20 to 28 (mean AI of126.16), and started decreasing gradually thereafter (Fig. 2b).SNME significantly (p < 0.0001) suppressed hind paw swellingfrom day 24 to 36 post-immunization. The CIA inhibition responseby the SNME treatment (100 mg/kg) was about 63.34–93.7% be-tween 24 and 36 days after the induction of CIA. In contrast, indo-methacin treatment inhibited CIA only by 78.68% till day 36 of theexperimental period. Normal rats showed no increase in hind pawdiameter over time.
Hind paw swelling and erythema showed a time-dependent in-crease in severity with maximum arthritic score of �8 observedbetween days 16 and 32 in rats immunized with CII (Fig. 2c). SNMEtreatment exerted a significant (p < 0.0001) suppression of the ar-thritic score between days 24 and 36 post-immunization. Therewas no macroscopic evidence of either hind paw erythema in thehealthy control rats.
3.3.1.2. Change in body weight. The onset stage of CIA was associ-ated with drastic body weight loss that continued till the end of
3400 S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406
experimental period, i.e., day 36 in the vehicle-treated CIA rats(Fig. 2d). SNME (p = 0.0004) and indomethacin (p = 0.0004) treat-ment proved beneficial for the CIA rats in regaining their originalbody weights starting from day 24 and this trend continuedthroughout the experiment. Rats in the healthy control groupshowed a gradual increase in their body weight with respect totime. Rats in the SNME-treated group showed higher weight gainthan did the indomethacin-treated group.
We did not observe any alteration in relative weight, aspect orcolor of liver, kidneys, lungs, brain and heart in CIA, treatmentgroups and control groups (data not shown).
Fig. 2. Representative photographs of hind paws from (A): (a) Control, (b) CIA (vehicle onlSNME and indomethacin treatments on severity of disease progression, as evident by (significant suppression of both arthritic index and score by SNME and indomethacin treagroup. ⁄⁄⁄p < 0.0001, ooop < 0.0001 versus CIA (vehicle only) group.
Fig. 3. Representative histological figures (hematoxylin and eosin stained slides) of kneemonolayer synovial cells lining of control rats, (b) hyperplastic synovial cells, erosion andlining and reduction in the degree of inflammatory cell infiltration in SNME and indomethsix animals (n = 6) per experimental group. (Original magnification 10�).
3.3.2. Effect of SNME on histopathologyOn day 36, histological evaluation of the joints in the vehicle-
treated CIA rats revealed marked characteristic features of severearthritis like synovial hyperplasia, massive mixed (neutrophil,macrophage, and lymphocyte) infiltration (Fig. 3b) along witharticular cartilage and bone erosion. No such arthritic features ofinflammation and tissue destruction were evident in the joints ofhealthy control group.
In the CIA rats treated with SNME, the degree of arthritis wassignificantly reduced (Fig. 3c) with respect to reduction of cartilagedamage, massive cell influx along with minimal evidence of
y), (c) CIA + SNME (100 mg/kg/day), (d) CIA + indomethacin (1 mg/kg/day). Effects ofB) arthritic index, (C) arthritic score, and (D) change in body weight. There was atment between days 24 and 36. Data expressed as mean ± S.D. of n = 6 animals per
joints from all experimental groups of rats. The arrowheads indicate, (a) smooth anddisruption of synovial lining in CIA (vehicle only) group, (c and d) smoother synovialacin treated arthritic rats. Photomicrographs are representative of histopathology of
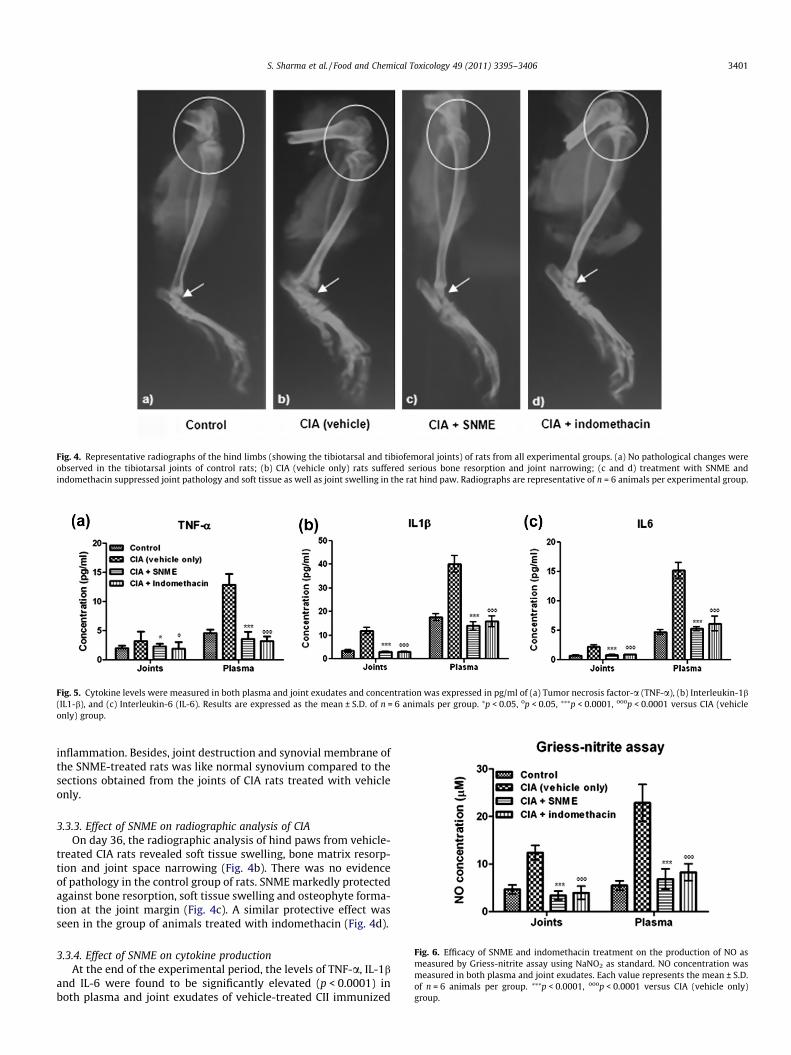
Fig. 4. Representative radiographs of the hind limbs (showing the tibiotarsal and tibiofemoral joints) of rats from all experimental groups. (a) No pathological changes wereobserved in the tibiotarsal joints of control rats; (b) CIA (vehicle only) rats suffered serious bone resorption and joint narrowing; (c and d) treatment with SNME andindomethacin suppressed joint pathology and soft tissue as well as joint swelling in the rat hind paw. Radiographs are representative of n = 6 animals per experimental group.
Fig. 5. Cytokine levels were measured in both plasma and joint exudates and concentration was expressed in pg/ml of (a) Tumor necrosis factor-a (TNF-a), (b) Interleukin-1b(IL1-b), and (c) Interleukin-6 (IL-6). Results are expressed as the mean ± S.D. of n = 6 animals per group. ⁄p < 0.05, op < 0.05, ⁄⁄⁄p < 0.0001, ooop < 0.0001 versus CIA (vehicleonly) group.
Fig. 6. Efficacy of SNME and indomethacin treatment on the production of NO asmeasured by Griess-nitrite assay using NaNO2 as standard. NO concentration wasmeasured in both plasma and joint exudates. Each value represents the mean ± S.D.of n = 6 animals per group. ⁄⁄⁄p < 0.0001, ooop < 0.0001 versus CIA (vehicle only)group.
S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406 3401
inflammation. Besides, joint destruction and synovial membrane ofthe SNME-treated rats was like normal synovium compared to thesections obtained from the joints of CIA rats treated with vehicleonly.
3.3.3. Effect of SNME on radiographic analysis of CIAOn day 36, the radiographic analysis of hind paws from vehicle-
treated CIA rats revealed soft tissue swelling, bone matrix resorp-tion and joint space narrowing (Fig. 4b). There was no evidenceof pathology in the control group of rats. SNME markedly protectedagainst bone resorption, soft tissue swelling and osteophyte forma-tion at the joint margin (Fig. 4c). A similar protective effect wasseen in the group of animals treated with indomethacin (Fig. 4d).
3.3.4. Effect of SNME on cytokine productionAt the end of the experimental period, the levels of TNF-a, IL-1b
and IL-6 were found to be significantly elevated (p < 0.0001) inboth plasma and joint exudates of vehicle-treated CII immunized
3402 S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406
rats in comparison to the healthy control rats [Fig. 5a–c]. Consis-tent with the joint swelling result, the levels of these pro-inflam-matory cytokines were significantly suppressed in CIA ratstreated with SNME (p < 0.0001) and indomethacin (p < 0.0001).
3.3.5. Inhibition of nitric oxide (NO) production by SNME in bothplasma and joint exudates
Fig. 6 depicts the effect of SNME on the inhibition of NO produc-tion in both the plasma and joint exudates of all the experimentalanimals. Significant elevation of NO production was observed inthe CIA rats (p < 0.0001) given vehicle alone compared to thehealthy controls. Administration of SNME (p < 0.0001) significantlyreduced the NO production in plasma and joints of arthritic rats.Similar results were obtained on treatment with indomethacin(p < 0.0001).
3.4. In vivo antioxidant activity
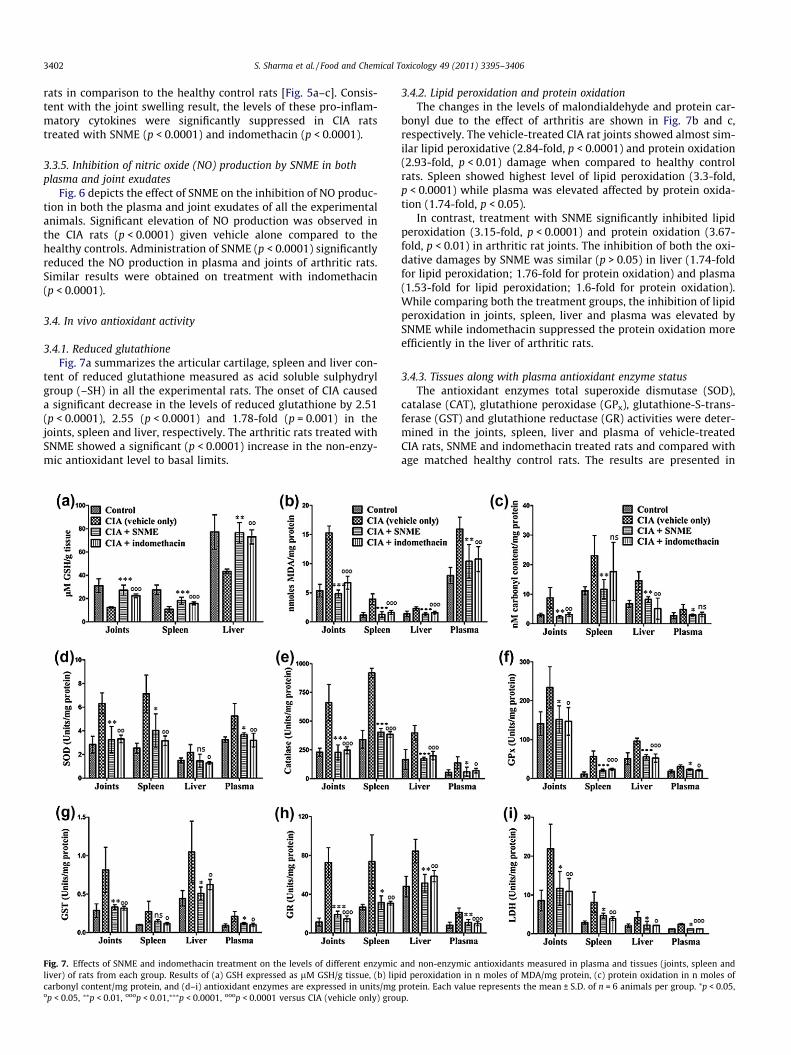
3.4.1. Reduced glutathioneFig. 7a summarizes the articular cartilage, spleen and liver con-
tent of reduced glutathione measured as acid soluble sulphydrylgroup (–SH) in all the experimental rats. The onset of CIA causeda significant decrease in the levels of reduced glutathione by 2.51(p < 0.0001), 2.55 (p < 0.0001) and 1.78-fold (p = 0.001) in thejoints, spleen and liver, respectively. The arthritic rats treated withSNME showed a significant (p < 0.0001) increase in the non-enzy-mic antioxidant level to basal limits.
Fig. 7. Effects of SNME and indomethacin treatment on the levels of different enzymicliver) of rats from each group. Results of (a) GSH expressed as lM GSH/g tissue, (b) lipcarbonyl content/mg protein, and (d–i) antioxidant enzymes are expressed in units/mgop < 0.05, ⁄⁄p < 0.01, ooop < 0.01,⁄⁄⁄p < 0.0001, ooop < 0.0001 versus CIA (vehicle only) grou
3.4.2. Lipid peroxidation and protein oxidationThe changes in the levels of malondialdehyde and protein car-
bonyl due to the effect of arthritis are shown in Fig. 7b and c,respectively. The vehicle-treated CIA rat joints showed almost sim-ilar lipid peroxidative (2.84-fold, p < 0.0001) and protein oxidation(2.93-fold, p < 0.01) damage when compared to healthy controlrats. Spleen showed highest level of lipid peroxidation (3.3-fold,p < 0.0001) while plasma was elevated affected by protein oxida-tion (1.74-fold, p < 0.05).
In contrast, treatment with SNME significantly inhibited lipidperoxidation (3.15-fold, p < 0.0001) and protein oxidation (3.67-fold, p < 0.01) in arthritic rat joints. The inhibition of both the oxi-dative damages by SNME was similar (p > 0.05) in liver (1.74-foldfor lipid peroxidation; 1.76-fold for protein oxidation) and plasma(1.53-fold for lipid peroxidation; 1.6-fold for protein oxidation).While comparing both the treatment groups, the inhibition of lipidperoxidation in joints, spleen, liver and plasma was elevated bySNME while indomethacin suppressed the protein oxidation moreefficiently in the liver of arthritic rats.
3.4.3. Tissues along with plasma antioxidant enzyme statusThe antioxidant enzymes total superoxide dismutase (SOD),
catalase (CAT), glutathione peroxidase (GPx), glutathione-S-trans-ferase (GST) and glutathione reductase (GR) activities were deter-mined in the joints, spleen, liver and plasma of vehicle-treatedCIA rats, SNME and indomethacin treated rats and compared withage matched healthy control rats. The results are presented in
and non-enzymic antioxidants measured in plasma and tissues (joints, spleen andid peroxidation in n moles of MDA/mg protein, (c) protein oxidation in n moles ofprotein. Each value represents the mean ± S.D. of n = 6 animals per group. ⁄p < 0.05,p.

S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406 3403
Fig. 7(d–h). The levels of GST, GR and CAT were increased by 2.86(p < 0.01), 6.39 (p < 0.0001) and 2.87-fold (p < 0.0001) in the jointsof vehicle-treated CIA rats. The spleen from the vehicle-treated ar-thritic rats showed an increase in SOD by 2.81-fold and the GPx le-vel was increased by 5-fold (p < 0.0001). The activity of CAT andGST was increased by almost 2.4-fold (p < 0.05) in arthritic rat liverwith a 1.44-fold increase in the activity of SOD. The antioxidant en-zyme levels of GR and GST were increased by 2.6 (p < 0.0001) and2.32 (p < 0.01), respectively in plasma of vehicle-treated CIA rats incomparison to the healthy control rats. Generally, the onset ofarthritis caused a significant increase in the activities of SOD,CAT, GPx, GST and GR antioxidant enzymes especially in arthriticrat joints and spleen. Treatment with SNME and indomethacinwere able to normalize the altered antioxidant enzyme levels ofall tissues occurring due to arthritis.
3.4.4. Lactate dehydrogenaseFig. 7i depicts the levels of lactate dehydrogenase in tissues
along with plasma of all the experimental rats. The levels of LDHwere found to be dramatically increased in spleen, joints, plasmaand liver of vehicle-treated arthritic rats. Administration of SNMEto arthritic rats significantly inhibited the activity of LDH by1.96-fold (p < 0.0001) in plasma, 1.87-fold (p < 0.05) in joints andliver and 1.73-fold (p < 0.05) in spleen. Although the inhibitionwas more pronounced by indomethacin treatment in joints(2.01-fold, p < 0.01) and spleen (2.06-fold, p < 0.01) of arthritic rats.
4. Discussion
Plant kingdom is a rich source of active components that havebeen shown to promote the specific cellular and humoral immuneresponse in different ways. This property has increased the interestof the scientific community to discover and develop various thera-peutic interventions to be used as medicine against several ail-ments. This study was conducted to investigate the effect ofSNME in comparison with indomethacin, on the progression ofCIA, and the mechanisms underlying this effect.
The results from in vitro studies showed that SNME is an effec-tive scavenger of both superoxide and hydrogen peroxide radicalsat different magnitudes of potency. This may be due to the pheno-lic content, particularly flavonoids present in SNME, which are be-lieved to be potent superoxide radical scavengers (Visavadiya et al.,2009). Moreover, presence of such compounds in the extract mayhelp neutralize hydrogen peroxide into water through electrondonation. The antioxidative properties of SNME on scavenginghydrogen peroxide radicals were found to be superior (IC50 valuewas 175–180 lg/ml) while the ability to inhibit superoxide radi-cals was lower than that reported for Salix caprea flower extract(Alam et al., 2006). SNME was also checked for its inhibitory effectson another harmful ROS, hypochlorous acid and showed equal po-tency as standard antioxidant compounds. The antioxidant effectsof SNME on the suppression of iron-ascorbate induced lipid perox-idation in rat brain cytosolic fraction were studied by measuringthe amount of malondialdehyde (MDA). SNME inhibited MDA for-mation more efficiently than rutin and this inhibition could becaused by direct scavenging of superoxide radicals or by reducingthe ferricyanide complex to ferrous form.
Furthermore, SNME noticeably decreased the amount of nitritegenerated by the decomposition of sodium nitroprusside and wasfound to be equally efficient with rutin. The observed reductionof nitric oxide may be due to the antioxidant properties of SNME,which compete with oxygen to react with nitric oxide therebyinhibiting the generation of peroxynitrite. The reduction ability(Fe3+ ? Fe2+) of SNME was relatively more pronounced than curcu-min and rutin at almost all the concentrations. These values were
also higher than that obtained for S. caprea flower extract (Alamet al., 2006). This activity of SNME is attributed to the high antiox-idant contents present in the bark of S. nigra, which could reactwith free radicals to stabilize and terminate radical chain reactions(Roginsky and Lissi, 2005). The phenolic content of SNME washigher than that reported for leaves and bark of S. caprea and S. alba(Kähkönen et al., 1999). However, values obtained in this workshowed less phenolic and flavonoid content compared to the ex-tracts of S. aegyptiaca reported by Enayat and Banerjee (2009).
Studies for the anti-arthritic properties of SNME were carriedout using CIA rats. SNME effectively improved the arthritic symp-toms throughout the experiment, leaving the arthritic score and in-dex closer to that of the healthy control group. Paw swelling(expressed as arthritic index) is an important factor in determiningthe degree of inflammation and therapeutic efficacy of the plantextract in question. A significant reduction in paw swelling (evi-dent by Fig. 2a) was observed in both SNME and indomethacin-treated groups, which indicates the anti-inflammatory potentialof SNME. Surprisingly, the CIA inhibition by SNME (93.7%) wasfound to be more pronounced than indomethacin (78.68%). It canbe suggested that the salicin content present in SNME might beresponsible for the observed anti-inflammatory effects. However,the phenols and flavonoids present in the extract might also pre-dict its therapeutic effectiveness. Also, the significant weight gainof the SNME-treated rats, showed the low toxicity of SNME atthe given dosage. Through both histologic and radiographic evalu-ations, we found that both SNME and indomethacin treatmentbrought about significant reduction in the severity of clinical indi-ces like soft tissue swelling, bone erosion, cartilage damage, mas-sive cell influx and joint space narrowing. These results thusconfirm that SNME had significant protective effects on the carti-lage and bone in the joints of rats immunized with CII.
In RA as well as CIA, the primary site of inflammation is thesynovial tissue, which can release cytokines into the systemic cir-culation causing measurable plasma levels of TNF-a, IL-1b and IL-6to shoot up. Compelling evidence suggests that the secretion ofcytokine IL1 is increased on exposure of mononuclear cells to typeII collagen. These cytokines including TNF-a are also released bysuperoxide radicals (via mechanisms not yet fully defined), macro-phages/monocytes and synovial lining cells (synoviocytes) (Sipeet al., 1994). Interestingly, the administration of SNME dramati-cally reduced the levels of these three pro-inflammatory cytokinesto almost basal levels. It is, therefore, proposed that SNME, beingan important negative regulator of these pro-inflammatory cyto-kines related to RA might deactivate the inflammatory responseof infiltrating and proliferating synovial cells. Hence, it is plausibleto suggest that part of the beneficial anti-inflammatory and carti-lage/bone protective effects of SNME may be mediated throughthe inhibition of TNF-a and IL-b. This, in turn, would lead to re-duced production of free radicals and subsequent damage.
The reactive free radical NO is a major macrophage-derivedinflammatory mediator and has been reported to be involved in di-verse physiologic and pathologic processes, including arthritis(Wallace, 2005). The increased production of NO in arthritic ani-mals may be owed to activation of inducible nitric oxide synthase(iNOS) in response to inflammatory cytokines such as IL-1b andTNF-a, since CIA model is characterized in terms of iNOS-drivenhigh output of NO production (Ialenti et al., 1993). We demonstratehere that the formation of nitrite (metabolite of NO) both in plasmaand joints caused by CIA is reduced in SNME-treated rats. The de-creased level of NO might be secondary effect to the reduced for-mation of endogenous TNF-a and IL-1.
There is wealth of evidence, both direct and indirect, thatthe production of free radicals at inflammatory sites damage cellu-lar components in cartilage directly (Henrotin et al., 2003).Reaction of NO with superoxide anion can modify the free and

3404 S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406
or/protein-bound amino acid residues and inhibit the enzymeactivities. Their reaction also forms peroxynitrite, a potent oxidiz-ing molecule capable of eliciting lipid peroxidation (Hitchon andEl-Gabalawy, 2004). In this study, we observed a significant in-crease in the lipid peroxide and protein carbonyl levels with theonset of arthritis. Peroxidation of lipids disturb the assembly ofthe membrane causing changes in fluidity and permeability, alter-ations of ion transport and probably contributes to accelerated ath-erosclerosis in RA (Hitchon and El-Gabalawy, 2004). Oxidativemodification of proteins leads to diminished specific protein func-tions and affect a variety of cellular functional proteins like recep-tors, signal transduction mechanisms, transport systems, andenzymes (Surh and Packer, 2005). Treatment with SNME decreasedmalondialdehyde and protein carbonyl concentrations in thejoints, spleen, liver tissue and plasma. It can be thought that thebeneficial effects of SNME against accumulation of lipid and pro-tein damage may be partly related to its direct free radical scaveng-ing and antioxidative potential as discussed above. No significantdifference (p > 0.05) was observed with respect to protein oxida-tion in spleen and plasma of arthritic rats. This in vivo lipid perox-idation inhibitory potential of SNME is well supported by ourin vitro tests in which the concentration-dependent decrease inthe MDA level signifies the potential of SNME in protecting againstdamage to membrane functions.
The concentration of GSH reflects the endogenous defensesagainst hydrogen peroxide formation and organic peroxides. Its re-duced level in vehicle treated CIA rats might be due to its excessiveconsumption during oxidative stress and cellular lysis associatedwith the development of arthritis (Hassan et al., 2001). The reduc-tion in the level of this non-enzymic antioxidant also contributesto increased lipid peroxidation in CIA rats as compared to healthycontrol rats (Kim et al., 2010). In group of rats treated with SNMEand indomethacin, levels of GSH rescued to reach normal levelsprobably either by competing in scavenging for free radicals orby the direct detoxification of reactive oxygen species and/or
Fig. 8. Possible mechanism underlying the anti-arthritic potential of S
neutralization of reactive intermediate species generated from oxi-dative stress.
A growing body of evidence indicates that superoxide radicalsalso result in collagen degradation that may accelerate otherinflammatory reactions and tissue destruction through the recruit-ment and activation of neutrophils (Salvemini et al., 2001). SOD is akey antioxidant enzyme that protects cell from superoxide toxicity.Besides, SOD has a double-barreled impact because its degradationproduct that is, hydrogen peroxide interacts with glutathione inthe presence of GPx and ultimately forms water and oxygen. To thisend, CAT also protects the cells by dismutating hydrogen peroxideinto water and oxygen or by using it as an oxidant. The activities ofthese three enzymes were found to be increased in the tissues andplasma of vehicle treated CIA rats. The highest activities of SOD andCAT in arthritic joints and spleen and that of GPx in spleen and livermake these tissues more susceptible to oxidative damage by theaction of oxygen free radicals. Indeed increased ROS/superoxideradicals have been documented at inflammation sites such assynovial joints, which in turn contribute to lipid peroxidation byincreasing hydrogen peroxide levels. Thus, to respond to an in-crease in superoxide radicals, the cells must not only elevate SODbut CAT and GPx as well, the latter to counter the effect of hydro-gen peroxide. Our literature survey revealed a very high discrep-ancy in the antioxidant system findings. As to SOD activity,significant decreases (Karatas et al., 2003; Kiziltunc et al., 1998)and increases (Geetha et al., 1998; Kasama et al., 1988) were re-ported. Related to CAT and GPx defense system; similar to our datasignificantly high activities of enzymes were observed (Geethaet al., 1998; Kasama et al., 1988; Braven et al., 1989) in RA patients.Another important enzyme of the glutathione defense system, i.e.,GR regenerates reduced glutathione from oxidized glutathione atthe expense of NADPH. As expected, the GR activity was also foundto be increased in the tissues examined along with plasma (Changet al., 1978). The highest augment of the activities was seen injoints and spleen of arthritic rats, which could be due to its
NME. The inhibitory actions of SNME treatment are represented.

S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406 3405
induction to counter the effect of chronic oxidative stress thatarises due to arthritis. Also, the increased GR activity is importantfor the detoxification pathway of oxygen free radicals. After SNMEtreatment, the alterations produced in the arthritic rats with re-spect to antioxidants concentrations were modulated to near nor-mal levels. Therefore, reversal of free radical induced oxidativedamage by SNME and indomethacin treatment indicates its protec-tive nature against protein oxidation through glutathione redox cy-cle and ability to directly detoxify the ROS generated fromexposure to CII.
GST is the detoxicant enzyme that metabolize toxic electro-philes, detoxifies endogenous compounds such as peroxidised lip-ids and is considered to be secondary antioxidant enzyme (Hayesand Pulford, 1995). In the present study, we have obtained markedincrease in the activity of GST with the onset of CIA, in line with thefindings of Surapneni et al. (Surapneni and Gopan, 2008) and Osta-lowska et al. (Ostalowska et al., 2006) for RA. The increase in theactivity of antioxidant enzyme can be attributed to response ofthe tissues to the increased oxidative stress due to CIA thereby sug-gesting an increased defense of the body against the effects of toxicelectrophilic chemicals and hydrophobic compounds. This bio-chemical alteration was significantly ameliorated nearly to controlvalues after administration of SNME and indomethacin to CIA rats.
LDH, a cytoplasmic cellular enzyme was also measured in thetissues and plasma of arthritic rats due to its importance as an indi-cator of the damage to cellular integrity induced by pathologicalconditions. The activity of LDH was found to be most pronouncedin the spleen, thereby indicating the natural killer activity of spleencells. SNME treatment normalized the altered enzymic values,comparable to indomethacin.
In conclusion, our results have demonstrated that SNME hashighly potent antioxidant activity under in vitro conditions. Be-sides, it has a great potential to ameliorate the progression of CIAby controlling inflammatory proteins, nitric oxide and antioxidantenzymes (Fig. 8). Finally, we propose that SNME being anti-inflam-matory and antioxidant in nature might have a preventive effectfor the inflammatory arthritis like RA. In further experiments, stud-ies with purified fractions of the extract will be conducted for fur-ther pharmacological characterization.
Conflict of Interest
The authors declare that there are no conflict of interest.
Acknowledgement
Authors wish to thank the University Grants Commission, Indiafor the financial assistance.
References
Afolayan, A.J., Aboyade, O.M., Sofidiya, M.O., 2008. Total phenolic content and freeradical scavenging activity of Malva parviflora L.(Malvaceae). J. Biol. Sci. 8, 945–949.
Alam, M.S., Kaur, G., Jabbar, Z., Javed, K., Athar, M., 2006. Evaluation of antioxidantactivity of Salix caprea flowers. Phytother. Res. 20, 479–483.
Ayoola, G.A., Coker, H.A.B., Adesegun, S.A., Adepoju-Bello, A.A., Obaweya, K.,Ezennia, E.C., Atangbayila, T.O., 2008. Phytochemical screening andantioxidant activities of some selected medicinal plants used for malariatherapy in southwestern Nigeria. Trop. J. Pharm. Res. 7, 1019–1024.
Biswas, S., Sahu, D., Saroha, A., Das, H.R., 2009. Glycoproteomic analysis of collageninduced arthritis in rats. Trends Carbohydr. Res. 1, 12–20.
Braven, J., Ansari, N., Figgitt, D.P., Fisher, A., Luders, C., Hickling, P., Whittaker, M.,1989. A comparison of glutathione reductase and glutathione peroxidaseactivities in patients with rheumatoid arthritis and healthy adults. Br. J.Rheumatol. 28, 212–215.
Chang, J.C., van der Hoeven, L.H., Haddox, C.H., 1978. Glutathione reductase in thered blood cells. Ann. Clin. Lab. Sci. 8, 23–29.
Choi, E.M., 2007. Oxidative status of DBA/1J mice with type II collagen inducedarthritis. J. Appl. Toxicol. 27, 472–481.
Choi, E.M., Kim, Y.H., 2008. A preliminary study of the effects of an extract ofLigularia fischeri leaves on type II collagen-induced arthritis in DBA/1J mice.Food Chem. Toxicol. 46, 375–379.
Coelho, M.G., Reis, P.A., Gava, V.B., Marques, P.R., Gayer, C.R., Laranja, G.A.,Felzenswalb, L., Sabino, K.C., 2004. Anti-arthritic effect and subacutetoxicological evaluation of Baccharis genistelloides aqueous extract. Toxicol.Lett. 154, 69–80.
Cuzzocrea, S., McDonald, M.C., Mota-Filipe, H., Mazzon, E., Costantino, G., Britti, D.,Mazzullo, G., Caputi, A.P., Thiemermann, C., 2000. Beneficial effects of tempol, amembrane permeable radical scavenger, in a rodent model of collagen-inducedarthritis. Arthritis Rheum. 43, 320–328.
Dahiru, D., Onubiyi, J.A., Umaru, H.A., 2006. Phytochemical screening andantiulcerogenic effect of Moringa oleifera aqueous leaf extract. Afr. J. Tradit.Complement. Altern. Med. 3, 70–75.
Enayat, S., Banerjee, S., 2009. Comparative antioxidant activity of extracts leaves,bark and catkins of Salix aegyptiaca sp.. Food Chem. 116, 23–28.
Fay, J., Varoga, D., Wruck, C.J., Kurz, B., Goldring, M.B., Pufe, T., 2006. Reactive oxygenspecies induce expression of vascular endothelial growth factor in chondrocytesand human articular cartilage explants. Arthritis Res. Ther. 8, R189.
Geetha, T., Varalakshmi, P., Latha, R.M., 1998. Effect of triterpenes from CrataevaNurvala stem bark on lipid peroxidation in adjuvant-induced arthritis in rats.Pharmacol. Res. 37, 191–195.
Hassan, M.Q., Hadi, R.A., Al-Rawi, Z.S., Padron, V.A., Stohs, S.J., 2001. The glutathionedefense system in the pathogenesis of rheumatoid arthritis. J. Appl. Toxicol. 21,69–73.
Hayes, J.D., Pulford, D.J., 1995. The glutathione S-transferase supergene family:regulation of GST and the contribution of isoenzymes to cancerchemoprotection and drug resistance. Crit. Rev. Biochem. Mol. Biol. 30, 445–600.
Hazra, B., Biswas, S., Mandal, N., 2008. Antioxidant and free radical scavengingactivity of Spondias pinnata. BMC Complement. Altern. Med. 8, 63.
Henrotin, Y.E., Bruckner, P., Pujol, J.P., 2003. The role of reactive oxygen species inhomeostasis and degradation of cartilage. Osteoarthritis Cartilage 11, 747–755.
Hitchon, C.A., El-Gabalawy, H.S., 2004. Oxidation in rheumatoid arthritis. ArthritisRes. Ther. 6, 265–278.
Ialenti, A., Moncada, S., Di Rosa, M., 1993. Modulation of adjuvant arthritis byendogenous nitric oxide. Br. J. Pharmacol. 110, 701–706.
Jyoti, A., Sethi, P., Sharma, D., 2009. Curcumin protects against electrobehavioralprogression of seizures in the iron-induced experimental model ofepileptogenesis. Epilepsy Behav. 14, 300–308.
Kähkönen, M.P., Hopia, A.I., Vuorela, H.J., Rauha, J.P., Pihlaja, K., Kujala, T.S.,Heinonen, M., 1999. Antioxidant activity of plant extracts containing phenoliccompounds. J. Agric. Food Chem. 47, 3954–3962.
Kane, C.W., 2006. Herbal Medicine of the American Southwest: A Guide to theIdentification, Collection, Preparation and Use of the Medicinal and EdiblePlants of the Southwestern United States. Lincoln Town Press, Tucson, USA.
Karatas, F., Ozates, I., Canatan, H., Halifeoglu, I., Karatepe, M., Colakt, R., 2003.Antioxidant status and lipid peroxidation in patients with rheumatoid arthritis.Indian J. Med. Res. 118, 178–181.
Kasama, T., Kobayashi, K., Sekine, F., Negishi, M., Ide, H., Takahashi, T., Niwa, Y.,1988. Follow-up study of lipid peroxides, superoxide dismutase and glutathioneperoxidase in the synovial membrane, serum and liver of young and old micewith collagen-induced arthritis. Life Sci. 43, 1887–1896.
Kaur, P., Arora, S., 2011. Superoxide anion radical scavenging activity of Cassiasiamea and Cassia javanica. Med. Chem. Res. 20, 9–15.
Khayyal, M.T., El-Ghazaly, M.A., Abdallah, D.M., Okpanyi, S.N., Kelber, O., Weiser, D.,2005. Mechanisms involved in the anti-inflammatory effect of a standardizedwillow bark extract. Arzneimittelforschung 55, 677–687.
Kim, K.R., Chung, T.Y., Shin, H., Son, S.H., Park, K.K., Choi, J.H., Chung, W.Y., 2010. Redginseng saponin extract attenuates murine collagen-induced arthritis byreducing pro-inflammatory responses and matrix metalloproteinase-3expression. Biol. Pharm. Bull. 33, 604–610.
Kizil, G., Kizil, M., Yavuz, M., Emen, S., Hakimoglu, F., 2008. Antioxidant activities ofethanol extracts of Hypericum triquetrifolium and Hypericum scabroides. Pharm.Biol. 46, 231–242.
Kiziltunc, A., Cogalgil, S., Cerrahoglu, L., 1998. Carnitine and antioxidants levels inpatients with rheumatoid arthritis. Scand. J. Rheumatol. 27, 441–445.
Lin, J.Y., Tang, C.Y., 2007. Determination of total phenolic and flavonoid contents inselected fruits and vegetables, as well as their stimulatory effects on mousesplenocyte proliferation. Food Chem. 101, 140–147.
Mahdi, J.G., Mahdi, A.J., Mahdi, A.J., Bowen, I.D., 2006. The historical analysis ofaspirin discovery, its relation to the willow tree and antiproliferative andanticancer potential. Cell Prolif. 39, 147–155.
Mathew, S., Abraham, T.E., 2006. Studies on the antioxidant activities of cinnamon(Cinnamomum verum) bark extracts, through various in vitro models. FoodChem. 94, 520–528.
Meier, B., Sticher, O., Julkunen-Tiitto, R., 1988. Pharmaceutical aspects of the use ofwillows in herbal remedies. Planta Med. 54, 559–560.
Minirth, F., Krusz, J.C., Horewell, A., Neal, V., 2005. The Christian’s Guide to NaturalProducts & Remedies: 1100 Herbs, Vitamins, Supplements And More! B&HPublishing Group, Canada.
Nahrstedt, A., Schmidt, M., Jaggi, R., Metz, J., Khayyal, M.T., 2007. Willow barkextract: the contribution of polyphenols to the overall effect. Wien MedWochenschr 157, 348–351.
Ogunlana, O.E., Ogunlana, O.O., 2008. In vitro assessment of the free radicalscavenging activity of Psidium guajava. Res. J. Agric. Biol. Sci. 4, 666–671.

3406 S. Sharma et al. / Food and Chemical Toxicology 49 (2011) 3395–3406
Ostalowska, A., Birkner, E., Wiecha, M., Kasperczyk, S., Kasperczyk, A., Kapolka, D.,Zon-Giebel, A., 2006. Lipid peroxidation and antioxidant enzymes in synovialfluid of patients with primary and secondary rheumatoid arthritis of the kneejoint. Osteoarthritis Cartilage 14, 139–145.
Prieto, P., Pineda, M., Aguilar, M., 1999. Spectrophotometric quantitation ofantioxidant capacity through the formation of a phosphomolybdenumcomplex: specific application to the determination of vitamin E. Anal.Biochem. 269, 337–341.
Pullaiah, T., 2006. Encyclopaedia of World Medicinal Plants. Regency, New Delhi,India.
Rioja, I., Bush, K.A., Buckton, J.B., Dickson, M.C., Life, P.F., 2004. Joint cytokinequantification in two rodent arthritis models: kinetics of expression, correlationof mRNA and protein levels and response to prednisolone treatment. Clin. Exp.Immunol. 137, 65–73.
Roginsky, V., Lissi, E.A., 2005. Review of methods to determine chain-breakingantioxidant activity in food. Food Chem. 92, 235–254.
Salvemini, D., Mazzon, E., Dugo, L., Serraino, I., De Sarro, A., Caputi, A.P., Cuzzocrea,S., 2001. Amelioration of Joint disease in a rat model of collogen inducedarthritis by M40403, a superoxide dismutase mimitic. Arthritis Rheum. 44,2909–2921.
Sanja, S.D., Sheth, N.R., Patel, N.K., Patel, D., Patel, B., 2009. Characterization andevaluation of antioxidant activity of Portulaca oleracea. Int. J. Pharm. Pharm. Sci.1, 74–84.
Sharma, R.K., Chatterji, S., Rai, D.K., Mehta, S., Rai, P.K., Singh, R.K., Watal, G., Sharma,B., 2009. Antioxidant activities and phenolic contents of the aqueous extracts ofsome Indian medicinal plants. J. Med. Plants Res. 3, 944–948.
Singh, R.P., Padmavathi, B., Rao, A.R., 2000. Modulatory influence of Adhatoda vesica(Justicia adhatoda) leaf extract on the enzymes of xenobiotic metabolism,
antioxidant status and lipid peroxidation in mice. Mol. Cell. Biochem. 213, 99–109.
Sipe, J.D., Martel-Pelletier, J., Otterness, I.G., Pelletier, J.P., 1994. Cytokine reductionin the treatment of joint conditions. Mediators Inflamm. 3, 243–256.
Soeken, K.L., Miller, S.A., Ernst, E., 2003. Herbal medicines for the treatment ofrheumatoid arthritis: a systematic review. Rheumatology (Oxford) 42, 652–659.
Surapneni, K.M., Gopan, V.S.C., 2008. Lipid peroxidation and antioxidant status inpatients with rheumatoid arthritis. Ind. J. Clin. Biochem. 23, 41–44.
Surh, Y.J., Packer, L., 2005. Oxidative stress, Inflammation and Health. Taylor &Francis Group.
Temraz, A., EL-tantawy, W.H., 2008. Characterization of antioxidant activity ofextract from Artemisia vulgaris. Pak. J. Pharm. Sci. 21, 321–326.
Verma, N., Tripathi, S.K., Sahu, D., Das, H.R., Das, R.H., 2009. Evaluation of inhibitoryactivities of plant extracts on production of LPS-stimulated pro-inflammatorymediators in J774 murine macrophages. Mol. Cell. Biochem. 336, 127–135.
Visavadiya, N.P., Soni, B., Dalwadi, N., 2009. Free radical scavenging andantiatherogenic activities of Sesamum indicum seed extracts in chemical andbiological model systems. Food Chem. Toxicol. 47, 2507–2515.
Wallace, J.L., 2005. Nitric oxide as a regulator of inflammatory processes. Mem. Inst.Oswaldo Cruz. 100, 5–9.
Wolfe, K., Wu, X., Liu, R.H., 2003. Antioxidant activity of apple peels. J. Agric. FoodChem. 51, 609–614.
Yeom, M.J., Lee, H.C., Kim, G.H., Lee, H.J., Shim, I., Oh, S.K., Kang, S.K., Hahm, D.H.,2006. Anti-arthritic effects of Ephedra sinica STAPF herb-acupuncture:inhibition of lipopolysaccharide-induced inflammation and adjuvant-Inducedpolyarthritis. J. Pharmacol. Sci. 100, 41–50.
Zaugg, S.E., Cefalo, D., Walker, E.B., 1997. Capillary electrophoretic analysis of salicinin Salix spp.. J. Chromatogr. 781, 487–490.
Related Documents











