Amblyomma triste (Koch, 1844) (Acari: Ixodidae) Ovaries: An ultrastructural analysis Patrı ´cia Rosa de Oliveira a , Gerva ´sio Henrique Bechara b , Maria Izabel Camargo Mathias a, * a Departamento de Biologia, I.B., UNESP, Av. 24 A, No. 1515, Cx. Postal 199, CEP: 13506-900, Rio Claro, SP, Brazil b Departamento de Patologia Veterina ´ ria, FCAV, UNESP, Via de Acesso Prof. Paulo Castellane, s/n, CEP: 14884-900, Jaboticabal, SP, Brazil Received 19 April 2006; received in revised form 11 February 2007; accepted 14 February 2007 Available online 28 February 2007 Abstract This study presents an ultrastructural analysis of the ovary of the tick, Amblyomma triste. The ovary of this species is of the panoistic type that is, without nursing and follicular cells. It is composed of a layer of epithelial cells forming a wall and of germinative cells that generate the oocytes which remain attached to the external margin of this wall by a multicellular pedicel. The different developmental stages in the oocytes had been described by Oliveira et al. [Oliveira, P.R., Bechara, G.H., Camargo-Mathias, M.I., 2006. Amblyomma triste (Koch, 1844) (Acari: Ixodidae): Morphological description of the ovary and of vitellogenesis. Experimental Parasitology 113, 179–185]. The results of the investigation suggest that besides exogenous production of vitellogenic elements, endogenous production can take place simultaneously, contributing to the development and growth of the oocytes. Ó 2007 Elsevier Inc. All rights reserved. Index Descriptors and Abbreviations: Amblyomma triste; Tick; Ovary; Vitellogenesis; Ultrastructure 1. Introduction The economic importance of ticks is widely acknowl- edged and is related to their feeding habits. During feeding, many species of ticks can transmit diseases caused by pro- tozoa, viruses, Rickettsias and spirochetes (Rey, 1973) to man and other animals. The tick species Amblyomma triste is distributed in the Neotropics and has been reported in Equador (Keirans, 1984), Argentina (Ivancovich, 1980), Uruguay and Brazil (Sinkoc et al., 1997) infesting tapir (Kohls, 1956), dogs (Correa, 1954), capybara (Hydrochaeris hydrochaeris) (Sinkoc et al., 1997), marsh deer Blastocerus dichotomus (Szabo ´ et al., 2003) and humans (Venzal et al., 2003). It has also been reported also from opossum (Didelphis mar- supialis) in an endemic area for Brazilian spotted fever in Pedreiras, State of Sa ˜o Paulo (Lemos et al., 1997). According to Till (1961), Sonenshine (1991) and Said (1992), the female reproductive system of ticks generally consists of a large U-shaped ovary located at the poster- ior region of the body, with a pair of oviducts, an uterus, a muscular connection tube, a vagina and a gen- ital opening. In this report, we describe the ultrastructure of the ovary, thereby contributing to our understanding of vitel- logenesis in A. triste. 2. Materials and methods Semi-feeding A. triste females from the tick colony were maintained under controlled conditions (28 °C, 80% humidity and 12 h light/dark cycle) at the Department of Animal Pathology, Veterinary College, UNESP – Jabotica- bal, SP, Brazil. Twenty-five specimens maintained in the refrigerator for thermal shock anesthesia were dissected in a saline solution (NaCl 7.5 g/L, Na 2 HPO 4 2.38 g/L and KH 2 PO 4 2.72 g/L). 0014-4894/$ - see front matter Ó 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.exppara.2007.02.006 * Corresponding author. Fax: +55 19 3526 4135. E-mail address: [email protected] (M.I. Camargo Mathias). www.elsevier.com/locate/yexpr Experimental Parasitology 116 (2007) 407–413

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/yexpr
Experimental Parasitology 116 (2007) 407–413
Amblyomma triste (Koch, 1844) (Acari: Ixodidae)Ovaries: An ultrastructural analysis
Patrıcia Rosa de Oliveira a, Gervasio Henrique Bechara b, Maria Izabel Camargo Mathias a,*
a Departamento de Biologia, I.B., UNESP, Av. 24 A, No. 1515, Cx. Postal 199, CEP: 13506-900, Rio Claro, SP, Brazilb Departamento de Patologia Veterinaria, FCAV, UNESP, Via de Acesso Prof. Paulo Castellane, s/n, CEP: 14884-900, Jaboticabal, SP, Brazil
Received 19 April 2006; received in revised form 11 February 2007; accepted 14 February 2007Available online 28 February 2007
Abstract
This study presents an ultrastructural analysis of the ovary of the tick, Amblyomma triste. The ovary of this species is of the panoistictype that is, without nursing and follicular cells. It is composed of a layer of epithelial cells forming a wall and of germinative cells thatgenerate the oocytes which remain attached to the external margin of this wall by a multicellular pedicel. The different developmentalstages in the oocytes had been described by Oliveira et al. [Oliveira, P.R., Bechara, G.H., Camargo-Mathias, M.I., 2006. Amblyomma
triste (Koch, 1844) (Acari: Ixodidae): Morphological description of the ovary and of vitellogenesis. Experimental Parasitology 113,179–185]. The results of the investigation suggest that besides exogenous production of vitellogenic elements, endogenous productioncan take place simultaneously, contributing to the development and growth of the oocytes.� 2007 Elsevier Inc. All rights reserved.
Index Descriptors and Abbreviations: Amblyomma triste; Tick; Ovary; Vitellogenesis; Ultrastructure
1. Introduction
The economic importance of ticks is widely acknowl-edged and is related to their feeding habits. During feeding,many species of ticks can transmit diseases caused by pro-tozoa, viruses, Rickettsias and spirochetes (Rey, 1973) toman and other animals.
The tick species Amblyomma triste is distributed in theNeotropics and has been reported in Equador (Keirans,1984), Argentina (Ivancovich, 1980), Uruguay and Brazil(Sinkoc et al., 1997) infesting tapir (Kohls, 1956), dogs(Correa, 1954), capybara (Hydrochaeris hydrochaeris)(Sinkoc et al., 1997), marsh deer Blastocerus dichotomus
(Szabo et al., 2003) and humans (Venzal et al., 2003). Ithas also been reported also from opossum (Didelphis mar-
supialis) in an endemic area for Brazilian spotted fever inPedreiras, State of Sao Paulo (Lemos et al., 1997).
0014-4894/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.exppara.2007.02.006
* Corresponding author. Fax: +55 19 3526 4135.E-mail address: [email protected] (M.I. Camargo Mathias).
According to Till (1961), Sonenshine (1991) and Said(1992), the female reproductive system of ticks generallyconsists of a large U-shaped ovary located at the poster-ior region of the body, with a pair of oviducts, anuterus, a muscular connection tube, a vagina and a gen-ital opening.
In this report, we describe the ultrastructure of theovary, thereby contributing to our understanding of vitel-logenesis in A. triste.
2. Materials and methods
Semi-feeding A. triste females from the tick colony weremaintained under controlled conditions (28 �C, 80%humidity and 12 h light/dark cycle) at the Department ofAnimal Pathology, Veterinary College, UNESP – Jabotica-bal, SP, Brazil.
Twenty-five specimens maintained in the refrigeratorfor thermal shock anesthesia were dissected in a salinesolution (NaCl 7.5 g/L, Na2HPO4 2.38 g/L and KH2PO4
2.72 g/L).
408 P.R. de Oliveira et al. / Experimental Parasitology 116 (2007) 407–413
2.1. Transmission electron microscopy (TEM)
The material was fixed in 2.5% glutaraldehyde, postfixedin 1% OsO4, and embedded in Epon Araldite. The materialwas then embedded in pure Epon resin and polymerized at60 �C for 72 h. Ultrathin sections were contrasted with ura-nyl acetate and lead citrate. Afterwards, screens containingultrathin sections of the material were examined and pho-tographed in a Phillips 100 Transmission Electron Micro-scope (TEM).
3. Results
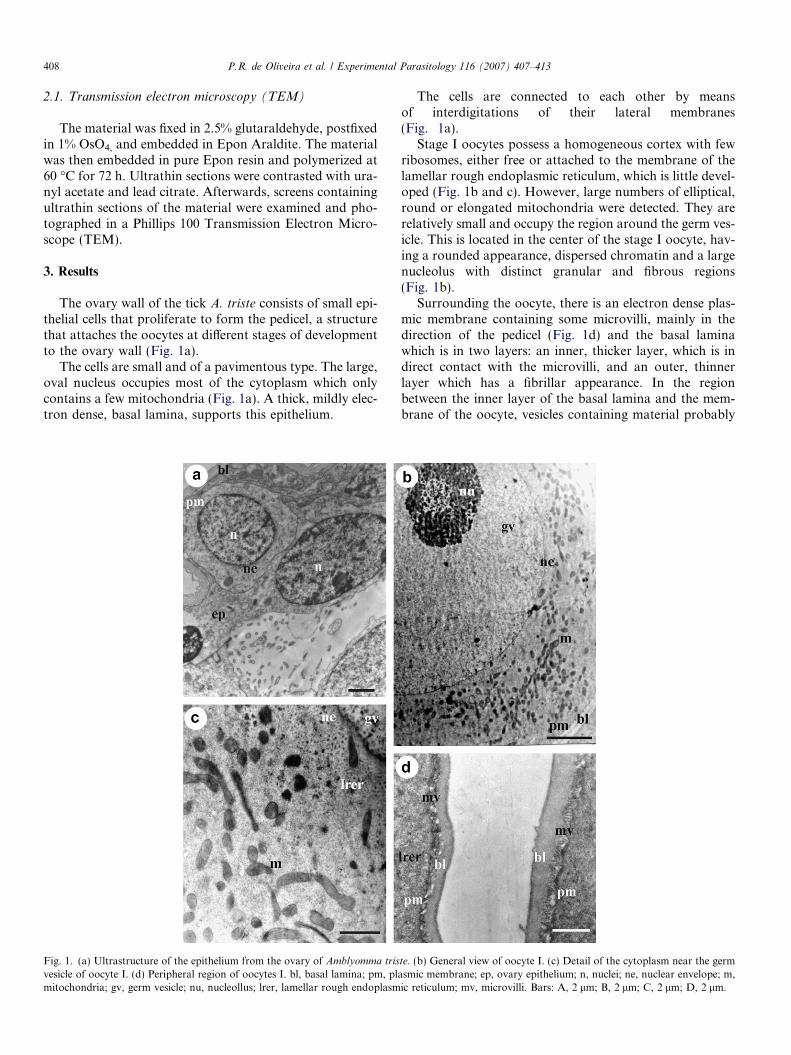
The ovary wall of the tick A. triste consists of small epi-thelial cells that proliferate to form the pedicel, a structurethat attaches the oocytes at different stages of developmentto the ovary wall (Fig. 1a).
The cells are small and of a pavimentous type. The large,oval nucleus occupies most of the cytoplasm which onlycontains a few mitochondria (Fig. 1a). A thick, mildly elec-tron dense, basal lamina, supports this epithelium.
Fig. 1. (a) Ultrastructure of the epithelium from the ovary of Amblyomma tris
vesicle of oocyte I. (d) Peripheral region of oocytes I. bl, basal lamina; pm, plmitochondria; gv, germ vesicle; nu, nucleollus; lrer, lamellar rough endoplasm
The cells are connected to each other by meansof interdigitations of their lateral membranes(Fig. 1a).
Stage I oocytes possess a homogeneous cortex with fewribosomes, either free or attached to the membrane of thelamellar rough endoplasmic reticulum, which is little devel-oped (Fig. 1b and c). However, large numbers of elliptical,round or elongated mitochondria were detected. They arerelatively small and occupy the region around the germ ves-icle. This is located in the center of the stage I oocyte, hav-ing a rounded appearance, dispersed chromatin and a largenucleolus with distinct granular and fibrous regions(Fig. 1b).
Surrounding the oocyte, there is an electron dense plas-mic membrane containing some microvilli, mainly in thedirection of the pedicel (Fig. 1d) and the basal laminawhich is in two layers: an inner, thicker layer, which is indirect contact with the microvilli, and an outer, thinnerlayer which has a fibrillar appearance. In the regionbetween the inner layer of the basal lamina and the mem-brane of the oocyte, vesicles containing material probably
te. (b) General view of oocyte I. (c) Detail of the cytoplasm near the germasmic membrane; ep, ovary epithelium; n, nuclei; ne, nuclear envelope; m,ic reticulum; mv, microvilli. Bars: A, 2 lm; B, 2 lm; C, 2 lm; D, 2 lm.
P.R. de Oliveira et al. / Experimental Parasitology 116 (2007) 407–413 409
being transported from the pedicel cells to the oocyte, canbe observed (Fig. 1d).
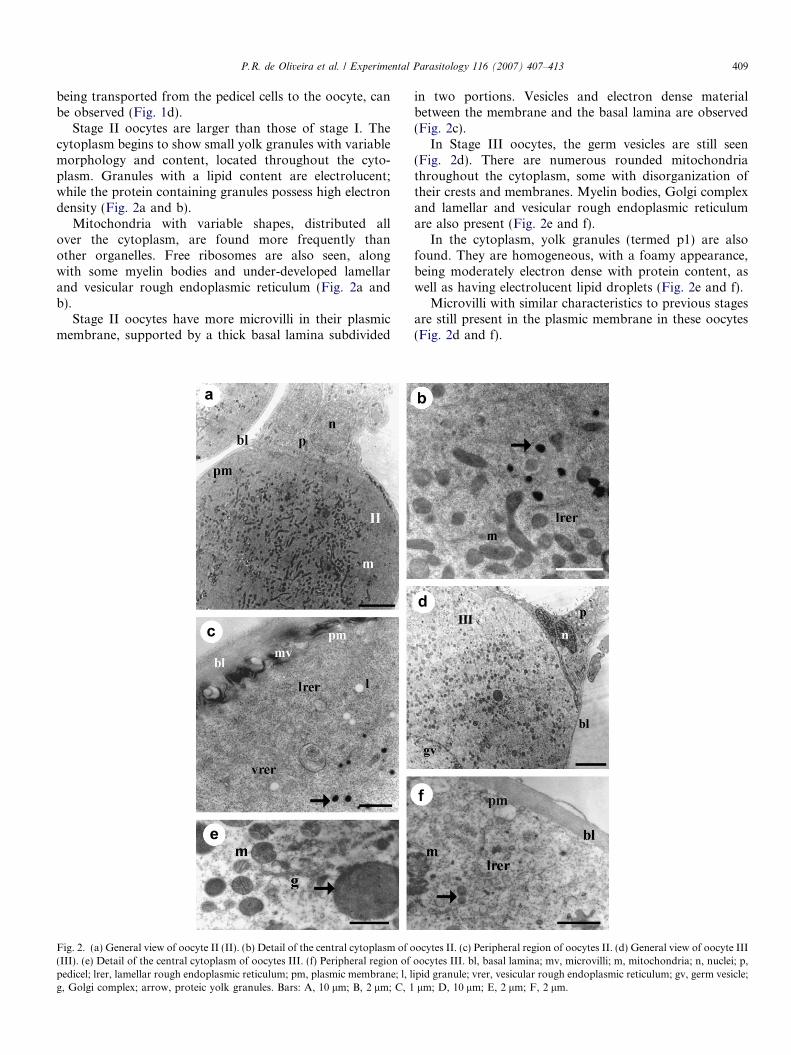
Stage II oocytes are larger than those of stage I. Thecytoplasm begins to show small yolk granules with variablemorphology and content, located throughout the cyto-plasm. Granules with a lipid content are electrolucent;while the protein containing granules possess high electrondensity (Fig. 2a and b).
Mitochondria with variable shapes, distributed allover the cytoplasm, are found more frequently thanother organelles. Free ribosomes are also seen, alongwith some myelin bodies and under-developed lamellarand vesicular rough endoplasmic reticulum (Fig. 2a andb).
Stage II oocytes have more microvilli in their plasmicmembrane, supported by a thick basal lamina subdivided
Fig. 2. (a) General view of oocyte II (II). (b) Detail of the central cytoplasm of o(III). (e) Detail of the central cytoplasm of oocytes III. (f) Peripheral region ofpedicel; lrer, lamellar rough endoplasmic reticulum; pm, plasmic membrane; l, lg, Golgi complex; arrow, proteic yolk granules. Bars: A, 10 lm; B, 2 lm; C, 1
in two portions. Vesicles and electron dense materialbetween the membrane and the basal lamina are observed(Fig. 2c).
In Stage III oocytes, the germ vesicles are still seen(Fig. 2d). There are numerous rounded mitochondriathroughout the cytoplasm, some with disorganization oftheir crests and membranes. Myelin bodies, Golgi complexand lamellar and vesicular rough endoplasmic reticulumare also present (Fig. 2e and f).
In the cytoplasm, yolk granules (termed p1) are alsofound. They are homogeneous, with a foamy appearance,being moderately electron dense with protein content, aswell as having electrolucent lipid droplets (Fig. 2e and f).
Microvilli with similar characteristics to previous stagesare still present in the plasmic membrane in these oocytes(Fig. 2d and f).
ocytes II. (c) Peripheral region of oocytes II. (d) General view of oocyte IIIoocytes III. bl, basal lamina; mv, microvilli; m, mitochondria; n, nuclei; p,ipid granule; vrer, vesicular rough endoplasmic reticulum; gv, germ vesicle;lm; D, 10 lm; E, 2 lm; F, 2 lm.
410 P.R. de Oliveira et al. / Experimental Parasitology 116 (2007) 407–413
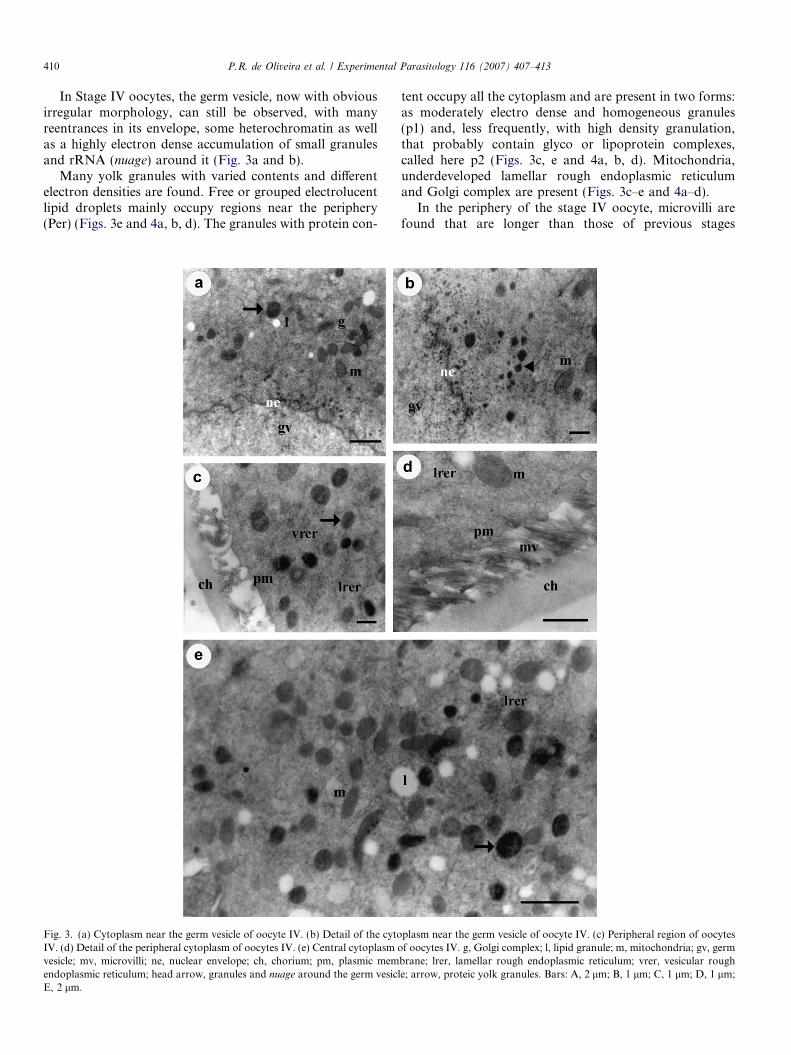
In Stage IV oocytes, the germ vesicle, now with obviousirregular morphology, can still be observed, with manyreentrances in its envelope, some heterochromatin as wellas a highly electron dense accumulation of small granulesand rRNA (nuage) around it (Fig. 3a and b).
Many yolk granules with varied contents and differentelectron densities are found. Free or grouped electrolucentlipid droplets mainly occupy regions near the periphery(Per) (Figs. 3e and 4a, b, d). The granules with protein con-
Fig. 3. (a) Cytoplasm near the germ vesicle of oocyte IV. (b) Detail of the cytoIV. (d) Detail of the peripheral cytoplasm of oocytes IV. (e) Central cytoplasm ovesicle; mv, microvilli; ne, nuclear envelope; ch, chorium; pm, plasmic memendoplasmic reticulum; head arrow, granules and nuage around the germ vesiclE, 2 lm.
tent occupy all the cytoplasm and are present in two forms:as moderately electro dense and homogeneous granules(p1) and, less frequently, with high density granulation,that probably contain glyco or lipoprotein complexes,called here p2 (Figs. 3c, e and 4a, b, d). Mitochondria,underdeveloped lamellar rough endoplasmic reticulumand Golgi complex are present (Figs. 3c–e and 4a–d).
In the periphery of the stage IV oocyte, microvilli arefound that are longer than those of previous stages
plasm near the germ vesicle of oocyte IV. (c) Peripheral region of oocytesf oocytes IV. g, Golgi complex; l, lipid granule; m, mitochondria; gv, germ
brane; lrer, lamellar rough endoplasmic reticulum; vrer, vesicular roughe; arrow, proteic yolk granules. Bars: A, 2 lm; B, 1 lm; C, 1 lm; D, 1 lm;
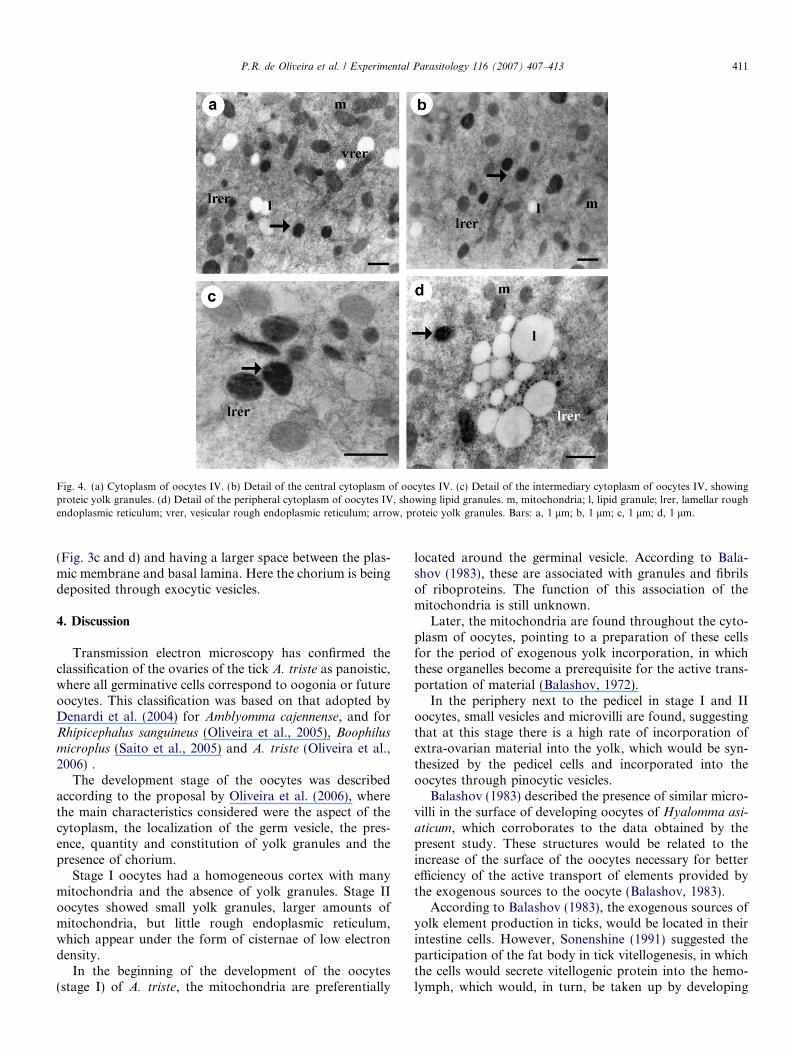
Fig. 4. (a) Cytoplasm of oocytes IV. (b) Detail of the central cytoplasm of oocytes IV. (c) Detail of the intermediary cytoplasm of oocytes IV, showingproteic yolk granules. (d) Detail of the peripheral cytoplasm of oocytes IV, showing lipid granules. m, mitochondria; l, lipid granule; lrer, lamellar roughendoplasmic reticulum; vrer, vesicular rough endoplasmic reticulum; arrow, proteic yolk granules. Bars: a, 1 lm; b, 1 lm; c, 1 lm; d, 1 lm.
P.R. de Oliveira et al. / Experimental Parasitology 116 (2007) 407–413 411
(Fig. 3c and d) and having a larger space between the plas-mic membrane and basal lamina. Here the chorium is beingdeposited through exocytic vesicles.
4. Discussion
Transmission electron microscopy has confirmed theclassification of the ovaries of the tick A. triste as panoistic,where all germinative cells correspond to oogonia or futureoocytes. This classification was based on that adopted byDenardi et al. (2004) for Amblyomma cajennense, and forRhipicephalus sanguineus (Oliveira et al., 2005), Boophilus
microplus (Saito et al., 2005) and A. triste (Oliveira et al.,2006) .
The development stage of the oocytes was describedaccording to the proposal by Oliveira et al. (2006), wherethe main characteristics considered were the aspect of thecytoplasm, the localization of the germ vesicle, the pres-ence, quantity and constitution of yolk granules and thepresence of chorium.
Stage I oocytes had a homogeneous cortex with manymitochondria and the absence of yolk granules. Stage IIoocytes showed small yolk granules, larger amounts ofmitochondria, but little rough endoplasmic reticulum,which appear under the form of cisternae of low electrondensity.
In the beginning of the development of the oocytes(stage I) of A. triste, the mitochondria are preferentially
located around the germinal vesicle. According to Bala-shov (1983), these are associated with granules and fibrilsof riboproteins. The function of this association of themitochondria is still unknown.
Later, the mitochondria are found throughout the cyto-plasm of oocytes, pointing to a preparation of these cellsfor the period of exogenous yolk incorporation, in whichthese organelles become a prerequisite for the active trans-portation of material (Balashov, 1972).
In the periphery next to the pedicel in stage I and IIoocytes, small vesicles and microvilli are found, suggestingthat at this stage there is a high rate of incorporation ofextra-ovarian material into the yolk, which would be syn-thesized by the pedicel cells and incorporated into theoocytes through pinocytic vesicles.
Balashov (1983) described the presence of similar micro-villi in the surface of developing oocytes of Hyalomma asi-
aticum, which corroborates to the data obtained by thepresent study. These structures would be related to theincrease of the surface of the oocytes necessary for betterefficiency of the active transport of elements provided bythe exogenous sources to the oocyte (Balashov, 1983).
According to Balashov (1983), the exogenous sources ofyolk element production in ticks, would be located in theirintestine cells. However, Sonenshine (1991) suggested theparticipation of the fat body in tick vitellogenesis, in whichthe cells would secrete vitellogenic protein into the hemo-lymph, which would, in turn, be taken up by developing

412 P.R. de Oliveira et al. / Experimental Parasitology 116 (2007) 407–413
oocytes through endocytosis. Our results additionally sug-gest the participation of pedicel cells in the vitellogenesis,synthesizing and providing substances to the interior ofthe oocytes.
In stage III oocytes many microvilli in the plasmic mem-brane were found. Large numbers of yolk granules of var-ied sizes were seen, while only a small amount of rough andsmooth endoplasmic reticulum, responsible for the endog-enous synthesis of lipids and proteins, respectively, werefound. This suggests that, in stage III oocytes, there is moreevidence that yolk elements come from extra-ovariansources.
According to Camargo-Mathias (1993) and Denardiet al. (2006), mitochondria are one of the sources for lipidsin the oocytes of the ant Neoponera villosa, confirming ear-lier observations (Bonhag, 1958; Wigglesworth, 1964).Ranade (1933) had suggested that mitochondria would betransformed into lipid yolk bodies and that the lipid mate-rial would come from the destruction of the mitochondrialcrests. Stage III oocytes of A. triste show such disorganiza-tion of the crests and membranes in the mitochondria, thepresence of lipids and little smooth endoplasmic reticulumas mentioned above. The present study therefore suggeststhat lipid components could be derived from mitochondriaas well as from an exogenous source.
In stage IV oocytes, numerous yolk granules of variedappearance and electron density, with many microvilli intheir plasmic membrane, and the deposition of choriumbetween the basal lamina and the plasmic membrane canbe seen. The presence of microvilli to this last stage ofdevelopment suggests that the incorporation of extra-ovar-ian yolk elements continues throughout vitellogenesis cor-roborating to the data obtained by Oliveira et al. (2005)for R. sanguineus.
The germ vesicle in stage IV oocytes has many reen-trances as well as nuclear pores that allow the passage ofmaterial (possibly rRNA) from the nucleus to the cyto-plasm. Consequently, around the germ vesicle, there is anincrease in the quantity of ribosomes, as well as formationof nuage and deposition of small electron dense granules.These could indicate endogenous synthesis of yolk in stageIV oocytes. The passage of material through the nuclearpores also was observed in the oocytes of camacuto scrimpAtya scabra (Cruz-Landim, 1997).
Diehl (1970) showed that vitellogenin proteins of ticksare immunologically identical to proteins from the hemo-lymph. This could be evidence for some extra-ovariansources of elements for the formation of yolk. Other tissuescould also synthesize these substances that would be trans-ported by hemolymph, taken up by microvilli andabsorbed through pinocytosis.
Balashov (1983) suggested that in the tick H. asiaticum,the yolk would be both endogenous and exogenous but thatendogenous production could occur before exogenous pro-duction. Our results contradict this, since oocytes in earlydevelopment (I and II) already possess evidence such as:microvilli in the plasmic membrane, and a large quantity of
mitochondria in the contact region oocyte/pedicel cells thatwould suggest exogenous transport of yolk elements.
Denardi et al. (2004), in oocytes of the A. cajennense andOliveira et al. (2005), in R. sanguineus described the chori-um as being subdivided into two layers: the endochorium(more internal and electro dense) and the exochorium(more external and less electron dense). These characteris-tics were not seen in the oocytes of A. triste.
Acknowledgments
We thank Miss Cristina Oishi Gridi Papp, Miss MonikaIamonte, Mr. Paulo Aruana Cezar, Mr. Antonio TeruyoshiYabuki and Mr. Ronaldo Del Vecchio for their technicalsupport and CAPES for financial support. Part of thiswork has been facilitated through the International Con-sortium of Ticks and Tick-borne Diseases (ICTTD-3)Coordination Action financed by the INCO program ofthe European Comission (Project No. 510561).
References
Balashov, Y.S., 1972. A translation of bloodsucking ticks (Ixodidae)vectors of diseases of man and animals. Miscellaneous Publications ofthe Entomological Society of America 8, 159–376.
Balashov, Y.S., 1983. The female reproductive system. In: Balashov, Y.S.(Ed.), An Atlas of Ixodid Tick Ultrastructure. Entomological Societyof America, Russian, pp. 98–128.
Bonhag, P.F., 1958. Ovarian structure and vitellogenesis in insects.Annual Review of Entomology 3, 137–160.
Correa, O., 1954. Carrapatos determinados no Rio Grande do Sul.Biologia, patologia e controle. Boletim da Diretoria de ProducaoAnimal 10 (18), 38–54.
Cruz-Landim, C., 1997. Cell reorganization and cell death during thesecretory cycle of the hypopharyngeal gland in Meliponinae beeworkers (Hymenoptera: Apidae). Acta Microscopica 6, 75–78.
Camargo-Mathias, M.I., 1993. Histoquımica e ultra-estrutura dos ovariosde operarias e rainhas de formigas Neoponera villosa (Hymenoptera:Ponerinae). Universidade Estadual Paulista, Rio Claro, 156 pp.
Denardi, S.E., Bechara, G.H., Oliveira, P.R., Nunes, E.T., Saito, K.C.,Camargo-Mathias, M.I., 2004. Morphological characterization of theovary and vitellogenesis dynamics in the Amblyomma cajennense
(Acari: Ixodidae). Veterinary Parasitology 125, 379–395.Denardi, S.E., Camargo-Mathias, M.I., Bechara, G.H., 2006. Ambly-
omma cajennense (Acari: Ixodidae): Salivary gland cells of partiallyengorged females ticks and the production of lipid by their mitochon-dria. Experimental Parasitology 113, 30–35.
Diehl, P.A., 1970. Zur Oogenese bei Ornithodoros moubata (Murray)(Ixodoidea: Argasidae) unter besonderer Berucksichtigung der Vitel-logenese. Acta Tropical 27, 301–355.
Ivancovich, J.C., 1980. Reclasificacion de algunas especies de garrapatasdel genero Amblyomma (Ixodoidea) en la Argentina. InvestigacionesAgropecuarias 15, 673–682.
Keirans, J.E., 1984. George Henry Faulkner Nuttall and the Nuttall tickcatalogue. United States Department of Agriculture Research SeriesMiscellaneous Publications 1438, 1610–1700.
Kohls, G.M., 1956. Concerning the identity of Amblyomma maculatum,Amblyomma tigrinum, Amblyomma triste and Amblyomma ovatum ofKoch, 1844. Proceedings of the Entomological Society of Washington58, 143–147.
Lemos, E.R.S., Machado, R.D., Coura, J.R., Guimaraes, M.A., Freire,N.M., Amorim, M., Gazeta, G.S., 1997. Epidemiological aspects of theBrazilian spotted fever: seasonal activity of ticks collected in an

P.R. de Oliveira et al. / Experimental Parasitology 116 (2007) 407–413 413
endemic area in Sao Paulo. Revista da Sociedade Brasileira deMedicina Tropical 30, 181–185.
Oliveira, P.R., Bechara, G.H., Denardi, S.E., Saito, K.C., Nunes, E.T.,Camargo-Mathias, M.I., 2005. Morphological characterization of theovary and oocytes vitellogenesis of the tick Rhipicephalus sanguineus
(Latreille, 1806) (Acari: Ixodidae). Experimental Parasitology 110,146–156.
Oliveira, P.R., Bechara, G.H., Camargo-Mathias, M.I., 2006. Ambly-
omma triste (Koch, 1844) (Acari: Ixodidae): morphological descrip-tion of the ovary and of vitellogenesis. Experimental Parasitology113, 179–185.
Ranade, V., 1933. On the cytoplasmic inclusions in the oogenesis ofPeriplaneta americana Linn. Allahad. University Science Bulletin 9,85–121.
Rey, L., 1973. Parasitologia. Guanabara Koogan, Sao Paulo, 212 pp.Said, E.A., 1992. A contribution to the anatomy and histology of the
female reproductive system of Amblyomma cajennense (Acarina:Ixodidae). Journal of the Egyptian Society of Parasitology 22, 385–393.
Saito, K.C., Bechara, G.H., Oliveira, P.R., Nunes, E.T., Denardi, S.E.,Camargo-Mathias, M.I., 2005. Morphological, histological, andultrastructural studies of the ovary of the tick Boophilus microplus
(Canestrini, 1887) (Acari: Ixodidae). Veterinary Parasitology 129, 299–311.
Sinkoc, A.L., Brum, J.G.W., Muller, G., Begrow, A., Paulsen, R.M.M.,1997. Ixodideos parasitos de capivaras (Hydrochaeris hydrochaeris L.,1766) da Estacao Ecologica do Taim, Rio Grande, RS. Ciencia Rural27, 119–122.
Sonenshine, D.E., 1991. Biology of Ticks. Oxford University Press, NewYork, pp. 280–303.
Szabo, M.P.J., Labruna, M.B., Pereira, M.C., Duarte, J.M.B., 2003. Ticks(Acari: Ixodidae) on Wild Marsh-Deer (Blastocerus dichotomus) fromSoutheast Brazil: Infestations before and after habitat loss. Journal ofMedical Entomology 40, 268–274.
Till, W.M., 1961. A Contribution to the Anatomy and Histology of theBrown Ear Tick Rhipicephalus appendiculatus. Swets & ZeitlingerPublishers, Amsterdam, 124 pp.
Venzal, J.M., Guglielmone, A.A., Estrada-Pena, A., Cabrera, P.A.,Castro, O., 2003. Ticks (Ixodida: Ixodidae) parasitising humans inUruguay. Annals of Tropical Medicine & Parasitology 97, 769–772.
Wigglesworth, V.B., 1964. The hormonal regulation of growth andreproduction in insects. In: Beament, J.W.L., Theherne, J.E., Wig-glesworth, V.B. (Eds.), Advances in Insect Physiology. AcademicPress, New York, pp. 247–336.
Related Documents











