ALTITUDINAL VARIATION OF PHENOLIC CONTENTS IN FLOWERING HEADS OF Arnica montana cv. ARBO: A THREE YEAR COMPARISON RENATE SPITALER • ANDREA WINKLER • ISABELLA LINS • SEMA YANAR • HERMANN STUPPNER • CHRISTIAN ZIDORN* Institut für Pharmazie, Abteilung Pharmakognosie, Universität Innsbruck, Innrain 52, Josef- Moeller-Haus, A-6020 Innsbruck, Austria

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ALTITUDINAL VARIATION OF PHENOLIC CONTENTS IN
FLOWERING HEADS OF Arnica montana cv. ARBO:
A THREE YEAR COMPARISON
RENATE SPITALER • ANDREA WINKLER • ISABELLA LINS • SEMA YANAR •
HERMANN STUPPNER • CHRISTIAN ZIDORN*
Institut für Pharmazie, Abteilung Pharmakognosie, Universität Innsbruck, Innrain 52, Josef-
Moeller-Haus, A-6020 Innsbruck, Austria

2
Abstract – In continuation of our studies of altitudinal effects on the secondary metabolite
profile of flowering heads from taxa of the Asteraceae (R. Spitaler et al., 2006
Phytochemistry 67: 409-417) we investigated altitudinal variation of phenolic contents and
radical scavenging potential from cultivated plants of Arnica montana cv. ARBO during the
growing seasons 2003, 2004, and 2005. Moreover, by conducting experiments on potted
plants, we excluded that differences in phenolic contents from plants grown at different
altitudes were primarily related to differences in soil composition at these sites. To assess
altitudinal and interseasonal variation, plants of A. montana cultivar ARBO were grown in
nine experimental plots at altitudes between 590 and 2230 m at Mount Patscherkofel near
Innsbruck/Austria. In all growing seasons and regardless of the soil the plants were grown in,
the proportion of flavonoids with vicinal free hydroxy groups in ring B to flavonoids lacking
this feature as well as the total amount of caffeic acid derivatives significantly increased with
elevation. These increases of anti-oxidant phenolics corresponded to an increase of the radical
scavenging potential of extracts from plants grown in different altitudes. These results are
discussed with regards to previous studies suggesting that enhanced UV-B radiation as well as
decreased temperatures might trigger augmented biosynthesis of UV-absorbing and anti-
oxidant phenolics in higher plants.
Key Words – Arnica montana, Altitudinal effects, Chemical ecology, Phenolics, Caffeic acid
esters, Flavonoids, Natural products, UV radiation, Radical scavenging activity, Temperature
regime.

3
INTRODUCTION
Previous studies on altitudinal variation of phenolic compounds in different wild and
cultivated taxa from the Asteraceae suggested that factors related to the altitude of the
growing site are causing significant shifts in the quantity of secondary metabolites in
flowering heads (Zidorn and Stuppner, 2001; Zidorn et al., 2005; Spitaler et al., 2006). Such
altitudinal effects were first observed in wild Asteraceae populations growing in their natural
habitats, e.g. in populations from Leontodon helveticus Mérat emend. Widder growing in the
Eastern Alps (Zidorn and Stuppner, 2001). Subsequent studies confirmed that altitudinal
effects also occur in populations from neophytic taxa in their newly acquired distribution
areas (Zidorn et al., 2005). Genetical differences as the only source of altitudinal variation
were finally ruled out by cultivation experiments with Arnica montana L. cv. ARBO (Spitaler
et al., 2006) in the Tyrolean Alps.
A multitude of environmental factors changes with the altitude of the growing site. These
factors include precipitation, mean temperature, soil, wind speed, low and high temperature
extremes, duration of snow cover, length of the vegetation period, and the intensity of
radiation under clear sky conditions. Enhanced UV-B radiation and lower temperatures in
high altitudes have been most intensely discussed to have an impact on plant secondary
metabolism (Körner, 1999; Bilger et al., 2007). The increase of irradiance and especially of
UV-B radiation is well documented for the Alps and the Innsbruck area in particular
(Blumthaler et al., 1997). Irradiance under clear sky conditions increases by 8 % for total
irradiance, 9 % for UV-A radiation, and 18 % for erythemal effective radiation (mainly UV-
B) per 1000 m of altitude.
As a protective response against damage from excessive UV-B radiation plants enhance
the biosynthesis of UV-B absorbing and antioxidant phenolic compounds (Markham et al.,
1998b; Körner, 1999). The induction of enzymes involved in the biosynthesis of flavonoids
with experimentally supplemented UV radiation is well known (Wellmann, 1975; Jaakola and
Määttä-Riihinen, 2004). The role of phenolics in protecting plants from peak UV-B exposure
is not limited to UV-B screening (Markham et al., 1998b) but observed changes in ratios
between different types of flavonoids (ortho-dihydroxylated versus other flavonoids) indicate

4
that the enhanced radical scavenging potentials of ortho-dihydroxylated flavonoids like
luteolin and quercetin are also contributing to their UV-B protective activity (Markham et al.,
1998a; Spitaler et al., 2006).
Field experiments to assess the actual impact of the altitude of the growing site on plant
secondary metabolism have been performed only for a limited number of species. The present
study aimed to clarify three questions on the altitudinal effects on secondary metabolite
spectra of A. montana. These questions are based on our first Arnica field trial (Spitaler et al.,
2006): (i) Do (random) natural climate fluctuations cause the variation observed in the
previous study,. (ii) are the previously observed altitudinal variations due to differences in soil
characteristics, and (iii) do differences in profiles of phenolics correlate with antioxidant
capacity?
METHODS AND MATERIALS
Plant Material Plantlets from A. montana cultivar ARBO were purchased from Saatzucht
Steinach GmbH (Steinach/Germany) in May 2002 and planted out at Mount Patscherkofel as
described previously (Spitaler et al., 2006). The experimental plots were located at the
following altitudes above mean sea level: site 1, 590 m; site 2, 870 m; site 3, 1020 m; site 4,
1260 m; site 5, 1430 m; site 6, 1620 m; site 7, 1890 m; site 8, 1960 m; site 9, 2230 m. In
2004, a second batch of plantlets was planted in black plastic pots (23.5 cm diam., depth: 18.5
cm) filled with a mixture (1/1, v/v) of peat and sand. These pots were transferred to the
experimental sites during the summer of 2004 and dug into the surrounding soil. To exclude
ontogenetic differences, only flowering heads with at least two rows of flowering ray florets
were collected (Douglas et al., 2004). Terminal flowering heads of each stem were collected
separately from lateral capitula. Only terminal flowering heads were analyzed (Spitaler et al.,
2006). Flowering heads were collected in batches of twelve, air-dried, and afterwards kept at -
20 °C until analysis. For each site and harvest year three batches of flowering heads were
collected and analyzed separately. Flowering heads were pooled in batches of twelve to
minimize the effects of chance factors due to variations between individual plants, without the

5
necessity of an unreasonably high number of HPLC analyses. Three batches per altitude and
season (and soil treatment) were analyzed to gain some idea about the variation within the
populations during each season at each altitude.
Plants for phytochemical analyses were collected at the following dates (numbers in
brackets refer to the nine experimental sites from 1, lowest altitude, to 9, highest altitude):
2003 growing season: see Spitaler et al. (2006); 2004 growing season: 07.06. (1); 17.06. (2,
3); 30.06. (4, 5, 6); 08.07. (6, 7); 21.07. (4, 6, 7); and 04.08. (7, 8); 2005 growing season
natural soil: 31.05. (1); 06.06. (2); 10.06. (1); 22.06. (4, 5, 6); 30.06. (4, 5, 6, 7); 09.07. (7, 8);
14.07, (7, 8); 21.07. (8); 04.08. (9); and 12.08. (9); and 2005 growing season potted plants:
30.05. (2); 31.05. (1); 10.06. (3); 17.06. (2, 3): 22.06. (3, 4, 5, 6); 30.06. (4, 5); 09.07. (7, 8);
14.07. (8); 21.07. (7, 8); 28.07. (9); 04.08. (7, 9); and 12.08. (9).
Extract preparation Each batch of twelve air-dried flowering heads was ground and phenolics
were analyzed by HPLC/DAD as described previously (Spitaler et al., 2006). In detail, after
adding 2.50 mg of the internal standard compound cynarin as a stock solution in MeOH,
(CH3)2CO, and H2O (3/1/1, v/v/v), ground flowering heads (500.0 mg) were sonicated twice
for 30 min with a mixture of MeOH, (CH3)2CO, and H2O (3/1/1, v/v/v) and once for 30 min
with a mixture of MeOH and H2O (1/1, v/v) (total extraction volume 25 ml for each cycle).
The extracts were filtered, the remaining plant material rinsed with 20 ml of a mixture of
MeOH, (CH3)2CO, and H2O (3/1/1, v/v/v) and the combined extracts were filled up to 100.0
ml with a mixture of MeOH, (CH3)2CO, and H2O (3/1/1, v/v/v); 10.0 ml of this solution were
brought to dryness in vacuo and redissolved in 2.00 ml of a mixture of MeOH, (CH3)2CO, and
H2O (3/1/1, v/v/v). After filtration, this solution was used for HPLC analysis. Comparative
investigations using different extraction media and longer times of sonication, and a larger
number of sonication cycles proved that the chosen procedure led to an exhaustive extraction.
All quantitative analyses were run in triplicate. The extraction procedure for the DPPH assays
was identical to the one described above with the exception that no cynarin (a potent radical
scavenger itself) was added before the first step of extraction.

6
HPLC analyses HPLC analyses of phenolics were performed as described previously
(Spitaler et al., 2006). In detail, HP-1090 and HP-1100 ChemStations equipped with DAD
detectors were employed and the following parameters applied: column, Phenomenex Synergi
Hydro-Rp 80A 150 x 4.6 mm (4 m material); guard column, Phenomenex Security Guard
C18 (ODS, Octadecyl) 4.0 mm x 3.0 mm; mobile phase A, H2O/HCOOH/CH3COOH
(99/0.9/0.1, v/v/v); phase B, MeCN/MeOH/HCOOH/CH3COOH (89/10/0.9/0.1, v/v/v/v);
flow rate, 1.00 ml/min; injection volume 10 l; detection wavelength 350 nm; oven
temperature, ambient; linear gradient, 0 min 5% B, 5 min 15% B, 20 min 16% B, 35 min
18% B, 45 min 19% B, 55 min 27.5% B, 60 min 65% B, 65 min 98% B, 70 min stop; post
time, 12 min. The amounts of phenolics were estimated by comparing the peak areas obtained
for the particular flavonoids F1-F6 and caffeic acid derivatives P1-P9 with the peak area
obtained for the internal standard cynarin (CYN).
Compound identification Phenolics were grouped into flavonoids (F) and caffeic acid
derivatives (phenolic acids, P) based on their characteristic UV spectra (F: a broad maximum
at 350 nm, P: a maximum at 330 nm with a shoulder at 295 nm). Peaks assignable to
flavonoids (F1-F6) and phenolic acids (P1-P9) were numbered consecutively with increasing
HPLC retention times. Peak characterization and compound identification was performed as
previously described (Spitaler et al., 2006). Peak identities are assigned in Table 1.
DPPH Assays DPPH assays were performed in 96-well-plates using a Hidex Chameleon plate
reader in the absorption mode at 515 nm. 2,2-Diphenyl-1-picrylhydraziyl was purchased from
Sigma-Aldrich, Steinheim, Germany. Ascorbic acid, which was used as a reference
compound was purchased from Merck, Darmstadt, Germany. The final concentration of
DPPH in the test mixture was set to 40.0 mg/l. Arnica extracts were tested in final
concentrations of 1.00 mg/ml, 0.50 mg/ml, 0.25 mg/ml, 0.125 mg/ml, and 0.0625 mg/ml. The
reference compounds ascorbic acid (Merck, Darmstadt, Germany) was measured in final
concentrations of 20.0 µg/ml, 10.0 µg/ml, 6.00 µg/ml, 2.00 µg/ml, and 1.00 µg/ml. Each
concentration was applied six times on one plate and for each extracts three plates were

7
prepared. After the solutions were mixed, plates were slightly shaken and kept in the dark for
exactly 30 min. Then plates were measured three times at 515 nm.
IC50 values for each replicate of the extracts and the ascorbic acid solutions were calculated
using the following formula: IC50 = [(50 - LP)/(HP - LP) * (HC - LC)] + LC. LP = low
percentage, i.e. highest percent inhibition less than 50%; HP = high percentage, i.e. lowest
percent inhibition greater than 50%; HC = high concentration, i.e. concentration of test
substance (or extract) at the high percentage, LC = low concentration, i.e. concentration of
test substance (or extract) at the low percentage. To get standardized results for the radical
scavenging activity of the extracts, which can be compared to results from other studies,
inhibitory concentrations were expressed as ascorbic acid equivalents per gram dried plant
material using the following formula: AAE (mg/g) = 1000 * IC50AA / IC50AMO. AAE: ascorbic
acid equivalents, IC50AA: 50% inhibitory concentration of ascorbic acid (AA), IC50AMO: 50%
inhibitory concentration of A. montana (AMO) flowering heads.
Data analysis Pearson correlation coefficients and linear regression equations were calculated
using the MINITAB 13.31 software package (State College, PA, USA).
RESULTS
Variation of phenolic compounds in relation to the altitude of the growing site Analogous to
the previous investigation, six flavonoids F1-F6 and nine phenolic acids P1-P9 were
quantified by HPLC: quercetin 3-O--D-glucoside F1, patuletin 3-O--D-glucoside F2,
kaempferol 3-O--D-glucoside F3, kaempferol 3-O--D-glucuronide F4, 6-
methoxykaempferol 3-O--D-glucoside F5, hispidulin F6, chlorogenic acid P1, 3,5-
dicaffeoylquinic acid P4, 1-methoxyoxaloyl-3,5-dicaffeoylquinic acid P5, 4,5-
dicaffeoylquinic acid P6 as well as unknown hydroxycinnamate esters P2, P3, P7, P8, and P9
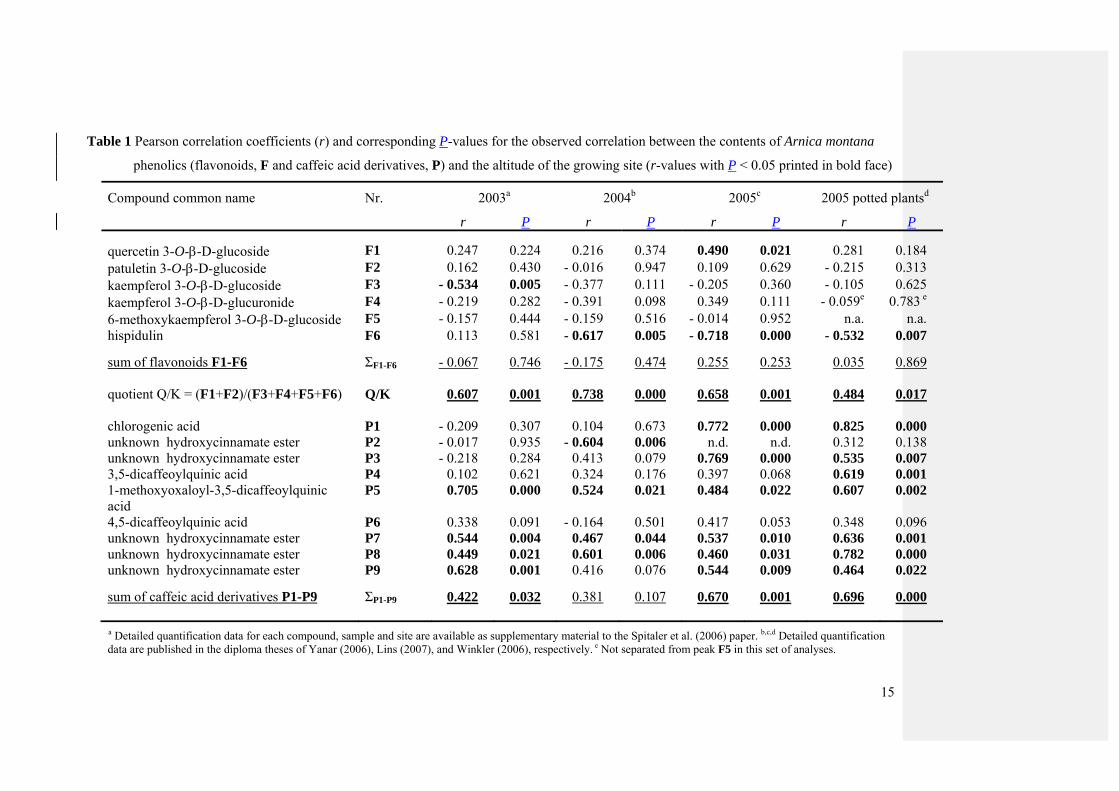
(Spitaler et al., 2006 and Table 1). In Table 1 Pearson correlation coefficients for individual

8
compounds and for the sums of compound groups (flavonoids and caffeic acid derivatives as
well as the ratio of ortho-dihydroxy versus other flavonoids) are summarized.
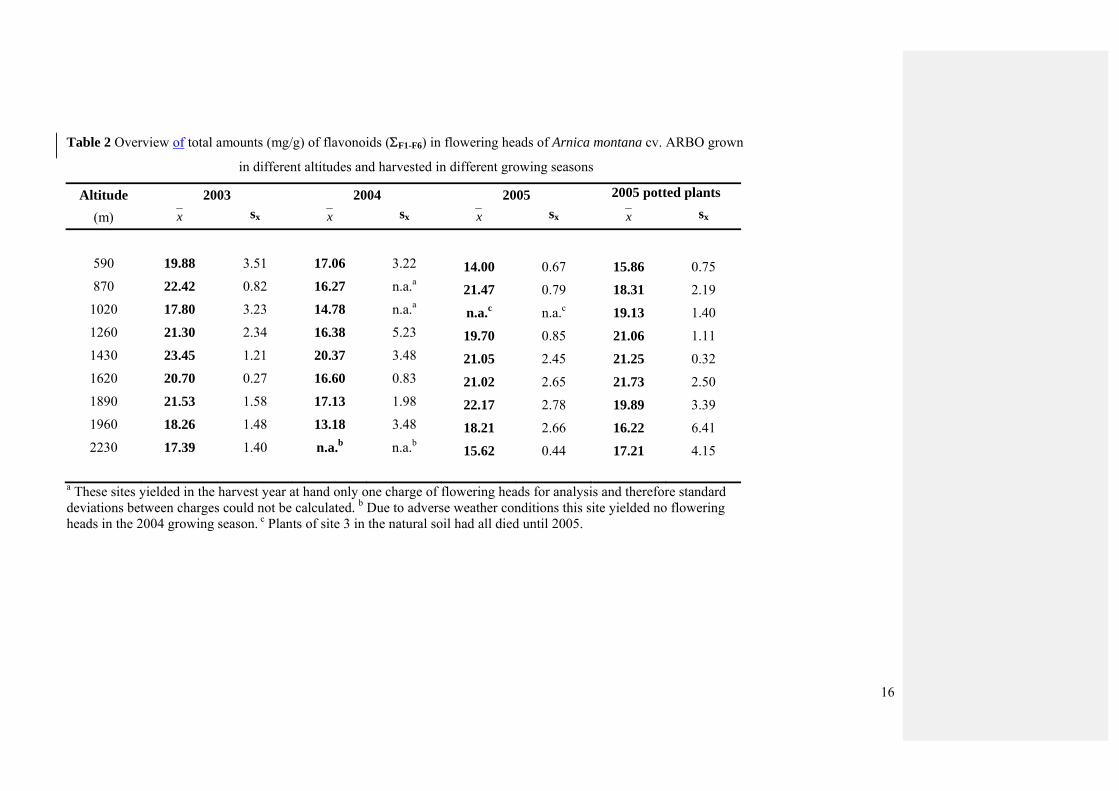
Quantification data for the total contents of flavonoids are summarized in Table 2. The
total content of flavonoids showed no significant positive correlation with the altitude of the
growing site in any of the growing seasons (Table 1). Moreover, neither did most of the
individual compounds show statistically significant correlation with the altitude of the
growing site. In contrast, the ratio of flavonoids with a 3’,4’-dihydroxylation pattern in ring B
to flavonoids lacking this feature [(F1+F2)/(F3+F4+F5+F6)] showed highly significant
increases with the altitude of the growing site (ranging from r = 0.484, P = 0.017 to r = 0.738,
p < 0.001). This correlation was remarkably stable over all growing seasons including potted
plants and plants grown in the natural soil. When all results for the quotient of Q/K from all
growing seasons and soil treatments were combined (which might be a bit problematic from a
purely statistical point of view), the Pearson correlation coefficient was r = 0.528 (P < 0.001)
(Fig. 1).
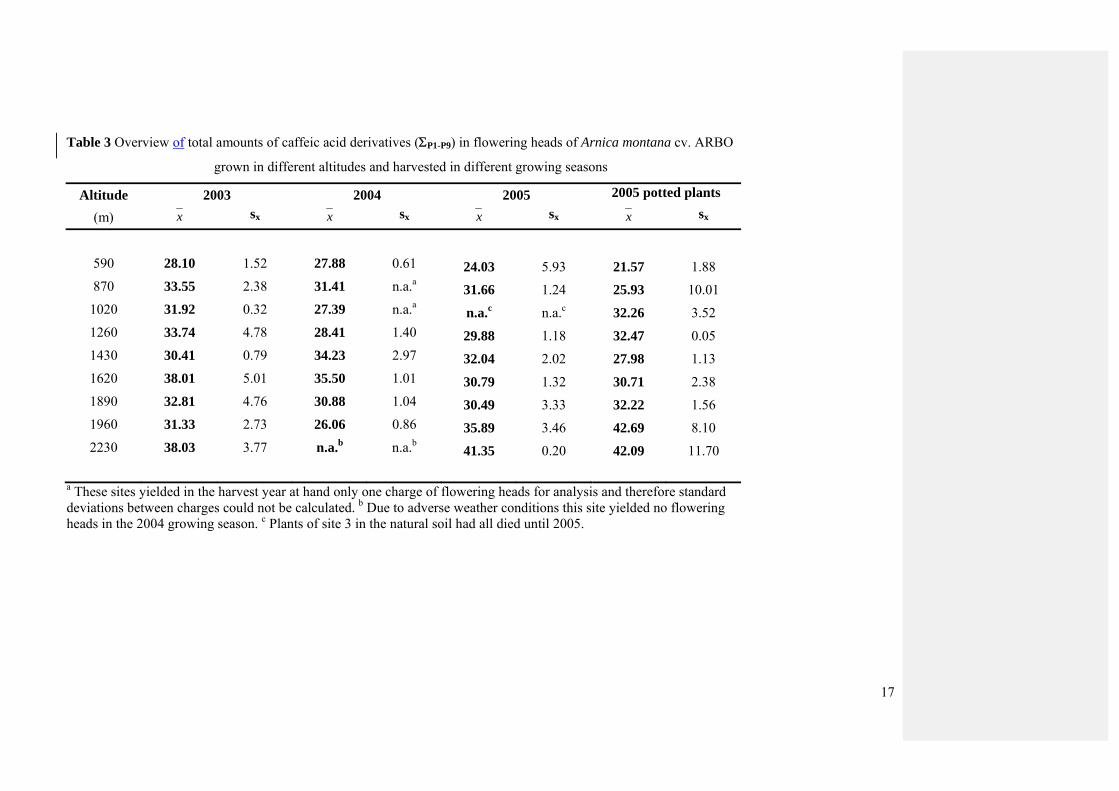
Quantification data of the total contents of caffeic acid derivatives (Table 3) revealed that
the total amounts of caffeic acid derivatives were positively correlated with the altitude of the
growing site in all investigated seasons, though the coefficients and their corresponding P-
values varied between r = 0.381, p = 0.107, and r = 0.696, P < 0.001 (Table 1).
The correlation of the quotient of the total amount of caffeic acid derivatives to the total
amount of flavonoids (ΣP1-P9/ΣF1-F6) with the altitude of the growing site also positively
correlated with the altitude of the growing site [r = 0.351, P = 0.001 for all investigated
samples]. However, it is hard to judge whether this result implies a significant shift in
biosynthetic pathways or whether it is merely a mathematical artefact resulting from the fact
that one variable (caffeic acid derivative contents) varies with altitude whereas the second
(total flavonoid contents) does not.
Variation of antioxidant activity in relation to the altitude of the growing site The radical
scavenging activity of extracts obtained from flowering heads harvested in different altitudes
was assessed for the samples harvested in 2004 and 2005, but not for the 2003 harvest,

9
because the DPPH assaying system for crude plant extracts had not yet been established in
our lab when the HPLC analyses of the 2003 samples were performed.
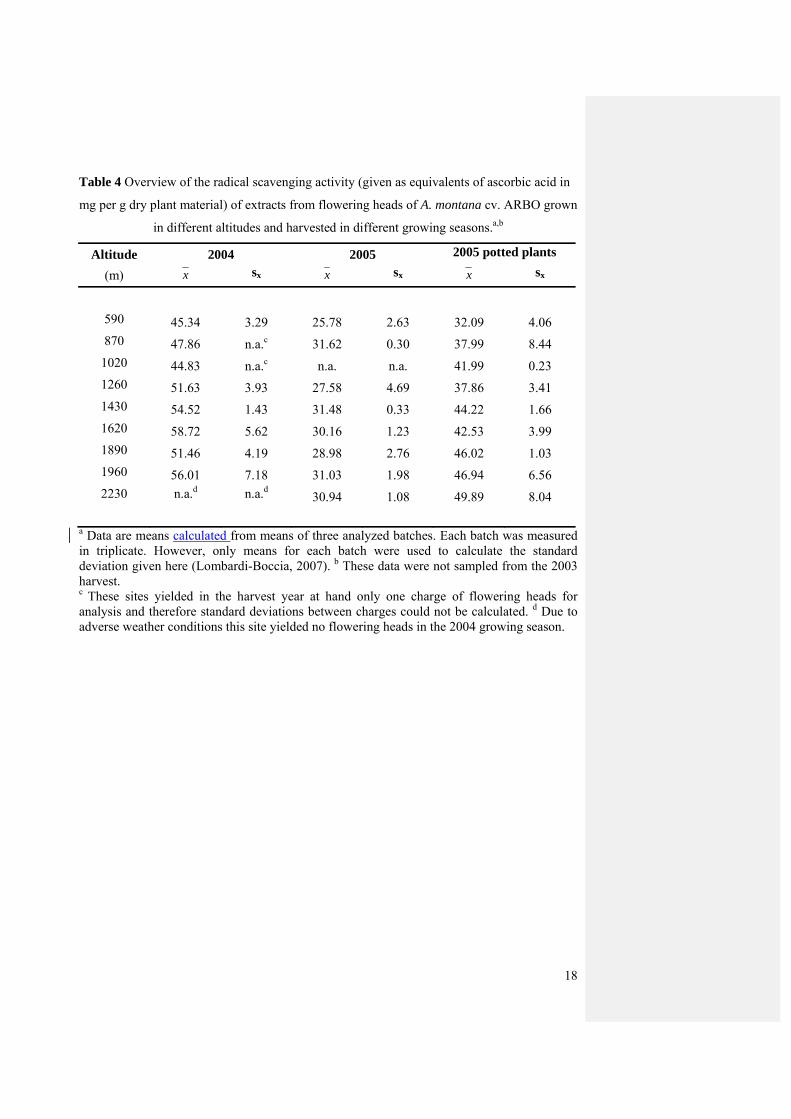
The results of the DPPH assays expressed as ascorbic acid equivalents (in mg) per gram
dried plant material are summarized in Table 4. The corresponding Pearson correlation
coefficients of altitude of the growing site and the DPPH radical scavenging potential were r
= 0.606 (P = 0.006) for 2004, r = 0.395 (P = 0.069) for 2005 for the plants in natural soil (n),
and r = 0.752 (p < 0.001) for 2005 for the potted plants (p). These correlation coefficients
were calculated from the mean values obtained for each batch, not from the mean values for
each site. The latter procedure would have resulted in even higher correlation coefficients [r =
0.787, P = 0.020 (2004), r = 0.491, P = 0.217 (2005n), r = 0.928, P < 0.001 (2005p)], though
due to smaller sample sizes also higher p-values.
DISCUSSION
The presented data confirm that climatic factors related to the altitude of the growing site have
profound and reproducible effects on the quantitative composition of profiles of certain
secondary metabolites in flowering heads of genetically homogenous plants of A. montana.
The results also confirm that changes in the quantity of phenolic compounds duringin
flowering heads have an impact on the radical scavenging activity of extracts derived from
flowering these heads. Moreover, we were able to demonstrate that altitudinal effects on the
quantity and relative composition of phenolics are not primarily caused by changes in the soil
composition at the growing site.
The most pronounced effect and the one least varying within the years of observation was
the ratio of 3’,4’-dihydroxylated flavonoids to flavonoids without that substitution pattern
(quercetin versus kaempferol derivatives). The data revealed that the DPPH radical
scavenging potential of flowering heads from plants grown in higher altitudes was
significantly higher than that of lowland plants. This result was not surprising as caffeic acid
derivatives, whose amounts were also positively correlated with the altitude of the growing
site, are potent radical scavengers (Rice-Evans et al., 1996). However, the result is
nonetheless noteworthy as other compounds, which are present in the plant extracts but which
Formatiert: Hervorheben

10
are not detected by the employed HPLC system (like ascorbic acid) are also contributing to
the overall antioxidant potential of the analyzed extracts. Moreover, the positive correlation of
the ratio of quercetin derivatives to kaempferol derivatives to the altitude of the growing site
is also a factor contributing to the enhanced radical scavenging activity of flowering heads
from higher altitudes, because quercetin derivatives are three to four times more potent radical
scavengers than kaempferol derivatives (Rice-Evans et al., 1996).
The new results yield further proof to the assumption that factors related to the altitude
indeed have an effect on plant secondary metabolism. However, the data are not conclusive to
which factors are most decisive in causing these changes. In addition to the enhanced UV-B
radiation in higher altitudes, which was assumed to be the main factor for these differences in
previous studies (Spitaler et al., 2006; Alonso-Amelot et al., 2007), the decreased temperature
in higher altitudes might play a more important role than assumed previously. Bilger et al.
(2007) demonstrated that decreased temperatures during the growth period trigger increased
biosynthesis rates of phenolic compounds in a variety of plant species even in the absence of
UV-B radiation. Phenolic compounds play a vital role in the hydrogen peroxide scavenging
system of plants, which besides phenolics comprises peroxidase, ascorbic acid, and glutathion
(Takahama and Oniki, 1997). This system functions less efficiently at low temperatures, and
more phenolics have to be produced to prevent damage to the plants grown at lower
temperatures (Bilger et al., 2007). Even if the later harvest dates of the plants grown at higher
altitudes are taken into account, the mean temperature during the growing and flowering
phase of the high altitude plants was significantly lower than for the lowland plants. Thus, it
might be assumed that not only the increase of UV-B at higher altitude sites but also the lower
temperatures at these sites are contributing to the observed altitudinal variations of phenolic
compound contents.
As stated earlier (Spitaler et al., 2006) factors other than temperature and UV-B radiation,
which usually change with altitude in mountain ranges of Central Europe and which also
might have an impact on plant secondary metabolism, are nearly unaffected by altitude in the
Tyrolean Central Alps due to their continental climate. These other factors are therefore to be
excluded as potential factors contributing to the altitudinal variation observed in the
secondary metabolite profiles of A. montana cv. ARBO in the Tyrolean Alps.

11
Acknowledgements – The authors wish to thank the Thurn und Taxis family (Patsch) and the
Agrargemeinschaft Patsch for a permit to establish experimental plots on their land. CZ
thanks the Swarovski foundation for financial support.

12
REFERENCES
ALONSO-AMELOT, M. E., OLIVEROS-BASTIDAS, A., and CALCAGNO-PISARELLI, M. P. 2007.
Phenolics and condensed tannins of high altitude Pteridium arachnoideum in relation to
sunlight exposure, elevation, and rain regime. Biochem. Syst. Ecol. 2007, 35:1-10.
BILGER, W., ROLLAND, M., and NYBAKKEN, L. 2007. UV screening in higher plants induced
by low temperature in the absence of UV-B radiation. Photochem. Photobiol. Sci. 6:
190-195.
BLUMTHALER, M., AMBACH, W., and ELLINGER, R. 1997. Increase in solar UV radiation with
altitude. J. Photochem. Photobiol. B 39: 130-134.
DOUGLAS, J. A., SMALLFIELD, B. M., BURGESS, E. J., PERRY, N. B., ANDERSON, R. E.,
DOUGLAS, M. H., and GLENNIE, V. L. 2004. Sesquiterpene lactones in Arnica montana:
A rapid analytical method and the effects of flower maturity and simulated mechanical
harvesting on quality and yield. Planta Med. 70:166-170.
JAAKOLA, L. and MÄÄTTÄ-RIIHINEN, K. 2004. Activation of flavonoid biosynthesis by solar
radiation in bilberry (Vaccinium myrtillus L.) leaves. Planta 218:721-728.
KÖRNER, C. 1999. Alpine Plant Life. Functional Plant Ecology of High Mountain
Ecosystems. Springer, Berlin.
LINS, I. 2007. Quantitative Variationen im Inhaltsstoffspektrum von Arnica montana in
Abhängigkeit von der Höhenlage und chemosystematische Untersuchungen innerhalb
der Gattung Leontodon. Diploma Thesis University of Innsbruck, Innsbruck.
LOMBARDI-BOCCIA, G. 2007. Rebuttal to comment on nutrients and antioxidant molecules in
yellow plums (Prunus domestica L.) form conventional and organic productions: a
comparative study. J. Agric. Food Chem. 55:4279-4280.
MARKHAM, K. R., RYAN, K. G., BLOOR, S. J., and MITCHELL, K.A. 1998a. An increase in the
luteolin : apigenin ratio in Marchantia polymorpha on UV-B enhancement.
Phytochemistry 48:791-794.
MARKHAM, K. R., TANNER, G. J., CAASI-LIT, M., WHITECROSS, M. I., NAYUDU, M., and
MITCHELL, K. A. 1998b. Possible protective role for 3’,4’-dihydroxyflavones induced
by enhanced UV-B in a UV-tolerant rice cultivar. Phytochemistry 49:1913-1919.

13
RICE-EVANS, C. A., MILLER, N. J., and PAGANGA, G. 1996. Structure-antioxidant activity
relationships of flavonoids and phenolic acids. Free Rad. Biol. Med. 20:933-956.
SPITALER, R., SCHLORHAUFER, P. D., ELLMERER, E. P., MERFORT, I., BORTENSCHLAGER, S.,
STUPPNER, H., and ZIDORN, C. 2006. Altitudinal variation of secondary metabolite
profiles in flowering heads of Arnica montana cv. ARBO. Phytochemistry 67:409-417.
TAKAHAMA, U. and ONIKI, T. 1997. A peroxidase/phenolics/ascorbate system can scavenge
hydrogen peroxide in plant cells. Physiol. Plant. 101:845-852.
WELLMANN, E. 1975. UV Dose-dependent induction of enzymes related to flavonoid
biosynthesis in cell suspension cultures of parsley. FEBS Lett. 51:105-107.
WINKLER, A. 2006. Quantitative Variationen im Inhaltsstoffspektrum von Arnica montana in
Abhängigkeit von Höhenlage, Substrat und Herkunft. Diploma Thesis University of
Innsbruck, Innsbruck.
YANAR, S. 2006. Arnica montana - Quantifizierung der phenolischen Inhaltsstoffe und
Bestimmung der Radikalfängeraktivität der Blütenkopfextrakte. Diploma Thesis
University of Innsbruck, Innsbruck.
ZAMG 2002. Klimadaten von Österreich. CD-ROM. Zentralanstalt für Metereologie und
Geodynamik Wien.
ZIDORN, C., SCHUBERT, B., and STUPPNER, H. 2005. Altitudinal differences in the contents of
phenolics in flowering heads of three members of the tribe Lactuceae (Asteraceae)
occurring as introduced species in New Zealand. Biochem. Syst. Ecol. 33:855-872.
ZIDORN, C. and STUPPNER, H. 2001. Evaluation of chemosystematic characters in the genus
Leontodon. Taxon 50:115-133.

14
Figure legend and footnote
Fig. 1 Correlation of the altitude of the growing site and the quotient of ortho-dihydroxy-
substituted flavonoids to other flavonoids {Q/K = [(F1+F2)/(F3+F4+F5+F6)]} in
flowering heads of A. montana harvested in different years1-3
1 Each symbol represents one analyzed batch. 2 Plants grown in natural soil (n) and potted
plants (p), respectively. 3 Correlation coefficients and corresponding p-values for each
season are displayed in Table 1.
15
Table 1 Pearson correlation coefficients (r) and corresponding P-values for the observed correlation between the contents of Arnica montana
phenolics (flavonoids, F and caffeic acid derivatives, P) and the altitude of the growing site (r-values with P < 0.05 printed in bold face)
Compound common name Nr. 2003a 2004b 2005c 2005 potted plantsd
r P r P r P r P
quercetin 3-O--D-glucoside F1 0.247 0.224 0.216 0.374 0.490 0.021 0.281 0.184 patuletin 3-O--D-glucoside F2 0.162 0.430 - 0.016 0.947 0.109 0.629 - 0.215 0.313 kaempferol 3-O--D-glucoside F3 - 0.534 0.005 - 0.377 0.111 - 0.205 0.360 - 0.105 0.625 kaempferol 3-O--D-glucuronide F4 - 0.219 0.282 - 0.391 0.098 0.349 0.111 - 0.059e 0.783 e 6-methoxykaempferol 3-O--D-glucoside F5 - 0.157 0.444 - 0.159 0.516 - 0.014 0.952 n.a. n.a. hispidulin F6 0.113 0.581 - 0.617 0.005 - 0.718 0.000 - 0.532 0.007
sum of flavonoids F1-F6 ΣF1-F6 - 0.067 0.746 - 0.175 0.474 0.255 0.253 0.035 0.869 quotient Q/K = (F1+F2)/(F3+F4+F5+F6) Q/K 0.607 0.001 0.738 0.000 0.658 0.001 0.484 0.017 chlorogenic acid P1 - 0.209 0.307 0.104 0.673 0.772 0.000 0.825 0.000 unknown hydroxycinnamate ester P2 - 0.017 0.935 - 0.604 0.006 n.d. n.d. 0.312 0.138 unknown hydroxycinnamate ester P3 - 0.218 0.284 0.413 0.079 0.769 0.000 0.535 0.007 3,5-dicaffeoylquinic acid P4 0.102 0.621 0.324 0.176 0.397 0.068 0.619 0.001 1-methoxyoxaloyl-3,5-dicaffeoylquinic acid
P5 0.705 0.000 0.524 0.021 0.484 0.022 0.607 0.002
4,5-dicaffeoylquinic acid P6 0.338 0.091 - 0.164 0.501 0.417 0.053 0.348 0.096 unknown hydroxycinnamate ester P7 0.544 0.004 0.467 0.044 0.537 0.010 0.636 0.001 unknown hydroxycinnamate ester P8 0.449 0.021 0.601 0.006 0.460 0.031 0.782 0.000 unknown hydroxycinnamate ester P9 0.628 0.001 0.416 0.076 0.544 0.009 0.464 0.022
sum of caffeic acid derivatives P1-P9 ΣP1-P9 0.422 0.032 0.381 0.107 0.670 0.001 0.696 0.000 a Detailed quantification data for each compound, sample and site are available as supplementary material to the Spitaler et al. (2006) paper. b,c,d Detailed quantification data are published in the diploma theses of Yanar (2006), Lins (2007), and Winkler (2006), respectively. e Not separated from peak F5 in this set of analyses.
16
Table 2 Overview of total amounts (mg/g) of flavonoids (F1-F6) in flowering heads of Arnica montana cv. ARBO grown
in different altitudes and harvested in different growing seasons
Altitude 2003 2004 2005 2005 potted plants
(m) x sx x sx x sx x sx
590 19.88 3.51 17.06 3.22 14.00 0.67 15.86 0.75
870 22.42 0.82 16.27 n.a.a 21.47 0.79 18.31 2.19
1020 17.80 3.23 14.78 n.a.a n.a.c n.a.c 19.13 1.40
1260 21.30 2.34 16.38 5.23 19.70 0.85 21.06 1.11
1430 23.45 1.21 20.37 3.48 21.05 2.45 21.25 0.32
1620 20.70 0.27 16.60 0.83 21.02 2.65 21.73 2.50
1890 21.53 1.58 17.13 1.98 22.17 2.78 19.89 3.39
1960 18.26 1.48 13.18 3.48 18.21 2.66 16.22 6.41
2230 17.39 1.40 n.a.b n.a.b 15.62 0.44 17.21 4.15
a These sites yielded in the harvest year at hand only one charge of flowering heads for analysis and therefore standard deviations between charges could not be calculated. b Due to adverse weather conditions this site yielded no flowering heads in the 2004 growing season. c Plants of site 3 in the natural soil had all died until 2005.
17
Table 3 Overview of total amounts of caffeic acid derivatives (P1-P9) in flowering heads of Arnica montana cv. ARBO
grown in different altitudes and harvested in different growing seasons
Altitude 2003 2004 2005 2005 potted plants
(m) x sx x sx x sx x sx
590 28.10 1.52 27.88 0.61 24.03 5.93 21.57 1.88
870 33.55 2.38 31.41 n.a.a 31.66 1.24 25.93 10.01
1020 31.92 0.32 27.39 n.a.a n.a.c n.a.c 32.26 3.52
1260 33.74 4.78 28.41 1.40 29.88 1.18 32.47 0.05
1430 30.41 0.79 34.23 2.97 32.04 2.02 27.98 1.13
1620 38.01 5.01 35.50 1.01 30.79 1.32 30.71 2.38
1890 32.81 4.76 30.88 1.04 30.49 3.33 32.22 1.56
1960 31.33 2.73 26.06 0.86 35.89 3.46 42.69 8.10
2230 38.03 3.77 n.a.b n.a.b 41.35 0.20 42.09 11.70
a These sites yielded in the harvest year at hand only one charge of flowering heads for analysis and therefore standard deviations between charges could not be calculated. b Due to adverse weather conditions this site yielded no flowering heads in the 2004 growing season. c Plants of site 3 in the natural soil had all died until 2005.
18
Table 4 Overview of the radical scavenging activity (given as equivalents of ascorbic acid in
mg per g dry plant material) of extracts from flowering heads of A. montana cv. ARBO grown
in different altitudes and harvested in different growing seasons.a,b
Altitude 2004 2005 2005 potted plants
(m) x sx x sx x sx
590 45.34 3.29 25.78 2.63 32.09 4.06
870 47.86 n.a.c 31.62 0.30 37.99 8.44
1020 44.83 n.a.c n.a. n.a. 41.99 0.23
1260 51.63 3.93 27.58 4.69 37.86 3.41
1430 54.52 1.43 31.48 0.33 44.22 1.66
1620 58.72 5.62 30.16 1.23 42.53 3.99
1890 51.46 4.19 28.98 2.76 46.02 1.03
1960 56.01 7.18 31.03 1.98 46.94 6.56
2230 n.a.d n.a.d 30.94 1.08 49.89 8.04
a Data are means calculated from means of three analyzed batches. Each batch was measured in triplicate. However, only means for each batch were used to calculate the standard deviation given here (Lombardi-Boccia, 2007). b These data were not sampled from the 2003 harvest. c These sites yielded in the harvest year at hand only one charge of flowering heads for analysis and therefore standard deviations between charges could not be calculated. d Due to adverse weather conditions this site yielded no flowering heads in the 2004 growing season.
Related Documents











