Vol. 37, no. 1 Journal of Vector Ecology 197 Altitudinal genetic and morphometric variation among populations of Culex theileri eobald (Diptera: Culicidae) from northeastern Turkey Berna Demirci 1 , Yoosook Lee 2 , Gregory C. Lanzaro 2 , and Bulent Alten 1 1 Hacettepe University, Science Faculty, Department of Biology, 06800 Beytepe, Ankara, Turkey, [email protected] 2 Department of Pathology, Microbiology and Immunology, School of Veterinary Medicine, University of California, Davis, CA 95616, U.S.A. Received 6 October 2011; Accepted 29 December 2011 ABSTRACT: Enviromental conditions, including such important climatic variables as temperature and precipitation, change with altitude; thus, elevation plays a significant role in determining population and community structure in a variety of organisms. Using single nucleotide polymorphisms (SNPs) and geometric morphometrics, nine populations of Culex theileri eobald occurring in different ecological subregions at altitudes between 808-2,130 m in northeastern Turkey were compared. e wing size and shape data indicate that there are significant phenotypic differences among them, while Cx theileri populations are not genetically differentiated in the northeast part of Turkey. e size and shape variation analysis of wings showed that there is a positive correlation between wing (body) size/shape and altitude. Journal of Vector Ecology 37 (1): 197-209. 2012. Keyword Index: Culex theileri, SNPs, altitudinal variation, geometric morphometrics. INTRODUCTION Culex theileri eobald has a wide Afrotropical, southern Palaearctic, and northern Oriental distribution. In Europe, it has been reported from Portugal, Spain, France, Italy, Yugoslovia, Greece, Hungary, Bulgaria, and Ukrania (Becker et al. 2003), and it is one of the most common mosquito species in northeastern Turkey. e larvae occur in spring in flooded meadows, stagnant or slowly moving streams, ditches, rock pools, drains, swamps, and rice fields but also frequently in artificial containers. Larvae can be found in highly polluted water (Aitken 1954, Ramos et al. 1977) and can tolerate a range of salinity, from fresh or slightly saline water (2 g NaCl/l) up to 16.6 g NaCl/l and pH 5.5-9.5 (Ramos et al. 1977). Cx. theileri has a very wide distribution in permanent breeding sites in the eastern part of Turkey and it prefers large natural larval habitats that are generaly present in rural areas (Aldemir et al. 2010, Simsek 2004). Cx. theileri is naturally infected with West Nile, Riſt Valley fever, and Sindbis arboviruses in South Africa (Jupp et al.1966, Mclntosh et al. 1967, Harbach 1988). In Iran, females were found to be naturally infected with third-stage larvae of Dirofilaria immitis (Azari-Hamidian et al. 2009). Field and laboratory studies verified that the species is a natural vector of D. immitis on Maderia Island, Portugal (Santa-Ana et al. 2006). In Iraq, Cx. theileri has been implicated in West Nile virus transmission (Abul-Hab 1967). Enviromental conditions, including such important climatic variables as temperature and precipitation, change sharply with altitude; thus, elevation plays a significant role in determining population and community structure in a variety of organisms. Important life history variables, such as growth rate, development time, longevity, and fecundity depend on temperature, especially in terrestial insects; these traits should be strongly affected by elevation gradients. Clearly these differences, especially the phenological ones between populations at different altitudes, may reduce or even eliminate the potential for gene flow, in particular in species which have a narrow time for reproductive activity. Knowledge of genetic variation and gene flow among vector populations is useful in following and predicting spatial and temporal dispersion of important genetic traits such as vector competence and insecticide resistance. Vector population structure and movement patterns also have clear implications for the ecology and evolution of vector-borne disease systems as well as for disease epidemiology and control (May and Anderson 1983, Gauthier and Tibayrenic 2005). Despite its broad distribution and evidence indicating that Cx. theileri is a competent vector of human and domestic animal pathogens, very little genetic data have been collected for this species. One study successfully amplified the cytochrome oxidase I gene (Azari- Hamidian et al. 2009), but there are no other genetic data available on the species. Here, we used nuclear single nucleotide polymorphisms (SNPs) to elucidate how Culex theileri eobald populations are structured genetically along elevational gradients in northeastern Turkey. e SNPs that we used for this study are the first molecular markers that were identified for Cx. theileri (Unpublished data). SNPs are the most abundant source of genetic variation among individual organisms. Previous studies have reported that the average SNP density in humans is one SNP per kb and about one SNP every 125 bp in the genome of the mosquito

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vol. 37, no. 1 Journal of Vector Ecology 197
Altitudinal genetic and morphometric variation among populations of Culex theileri Theobald (Diptera: Culicidae) from northeastern Turkey
Berna Demirci1, Yoosook Lee2, Gregory C. Lanzaro2, and Bulent Alten1
1Hacettepe University, Science Faculty, Department of Biology, 06800 Beytepe, Ankara, Turkey, [email protected] of Pathology, Microbiology and Immunology, School of Veterinary Medicine, University of California, Davis, CA
95616, U.S.A.
Received 6 October 2011; Accepted 29 December 2011
ABSTRACT: Enviromental conditions, including such important climatic variables as temperature and precipitation, change with altitude; thus, elevation plays a significant role in determining population and community structure in a variety of organisms. Using single nucleotide polymorphisms (SNPs) and geometric morphometrics, nine populations of Culex theileri Theobald occurring in different ecological subregions at altitudes between 808-2,130 m in northeastern Turkey were compared. The wing size and shape data indicate that there are significant phenotypic differences among them, while Cx theileri populations are not genetically differentiated in the northeast part of Turkey. The size and shape variation analysis of wings showed that there is a positive correlation between wing (body) size/shape and altitude. Journal of Vector Ecology 37 (1): 197-209. 2012.
Keyword Index: Culex theileri, SNPs, altitudinal variation, geometric morphometrics.
INTRODUCTION
Culex theileri Theobald has a wide Afrotropical, southern Palaearctic, and northern Oriental distribution. In Europe, it has been reported from Portugal, Spain, France, Italy, Yugoslovia, Greece, Hungary, Bulgaria, and Ukrania (Becker et al. 2003), and it is one of the most common mosquito species in northeastern Turkey. The larvae occur in spring in flooded meadows, stagnant or slowly moving streams, ditches, rock pools, drains, swamps, and rice fields but also frequently in artificial containers. Larvae can be found in highly polluted water (Aitken 1954, Ramos et al. 1977) and can tolerate a range of salinity, from fresh or slightly saline water (2 g NaCl/l) up to 16.6 g NaCl/l and pH 5.5-9.5 (Ramos et al. 1977). Cx. theileri has a very wide distribution in permanent breeding sites in the eastern part of Turkey and it prefers large natural larval habitats that are generaly present in rural areas (Aldemir et al. 2010, Simsek 2004).
Cx. theileri is naturally infected with West Nile, Rift Valley fever, and Sindbis arboviruses in South Africa (Jupp et al.1966, Mclntosh et al. 1967, Harbach 1988). In Iran, females were found to be naturally infected with third-stage larvae of Dirofilaria immitis (Azari-Hamidian et al. 2009). Field and laboratory studies verified that the species is a natural vector of D. immitis on Maderia Island, Portugal (Santa-Ana et al. 2006). In Iraq, Cx. theileri has been implicated in West Nile virus transmission (Abul-Hab 1967).
Enviromental conditions, including such important climatic variables as temperature and precipitation, change sharply with altitude; thus, elevation plays a significant role in determining population and community structure in a
variety of organisms. Important life history variables, such as growth rate, development time, longevity, and fecundity depend on temperature, especially in terrestial insects; these traits should be strongly affected by elevation gradients. Clearly these differences, especially the phenological ones between populations at different altitudes, may reduce or even eliminate the potential for gene flow, in particular in species which have a narrow time for reproductive activity. Knowledge of genetic variation and gene flow among vector populations is useful in following and predicting spatial and temporal dispersion of important genetic traits such as vector competence and insecticide resistance. Vector population structure and movement patterns also have clear implications for the ecology and evolution of vector-borne disease systems as well as for disease epidemiology and control (May and Anderson 1983, Gauthier and Tibayrenic 2005).
Despite its broad distribution and evidence indicating that Cx. theileri is a competent vector of human and domestic animal pathogens, very little genetic data have been collected for this species. One study successfully amplified the cytochrome oxidase I gene (Azari- Hamidian et al. 2009), but there are no other genetic data available on the species. Here, we used nuclear single nucleotide polymorphisms (SNPs) to elucidate how Culex theileri Theobald populations are structured genetically along elevational gradients in northeastern Turkey. The SNPs that we used for this study are the first molecular markers that were identified for Cx. theileri (Unpublished data). SNPs are the most abundant source of genetic variation among individual organisms. Previous studies have reported that the average SNP density in humans is one SNP per kb and about one SNP every 125 bp in the genome of the mosquito

198 Journal of Vector Ecology June 2012
Anopheles gambiae (Morlais and Severson 2003). Neutral SNPs may provide useful markers for population genetics studies. Non-synonymous SNPs, which alter the amino acid sequence and potentially the function of encoded proteins, are useful markers for association studies to detect genetic variation and to link these with phenotypic traits of interest.
Altitude also plays an important role in the size and shape of many species. Body size is one of the most significant features of organisms and is correlated with a large number of ecological and physiological variables. In mosquitoes, body size has been shown to affect factors such as longevity, fecundity, and size of the blood meal (Silver and Renshaw 1999). The size of adult mosquitoes may determine their effectiveness as vectors, and their longevity is an essential parameter in disease transmission as it increases the potential for infective bites to hosts. Similar to size, biological shape is one of the most conspicuous aspects of an organism’s phenotype and provides a link between the genotype and the environment (Ricklefs and Miles 1994). In insects, phenotypic variation among members of species complexes has been intensely investigated, especially concerning the size and shape of adult wings (James and Partridge 1998, Morin et al. 1999). In Drosophila, significant morphometric wing shape variations were found in populations from different geographic regions, forming clusters or latitudinal clines (Van’T Land et al. 1999), and laboratory studies of Drosophila populations showed that temperature change results in change in wing shape along with change in size (Cavicchi et al. 1985). Some studies report that variations in wing shape may be related to fitness or possible targets of natural selection (Buggs 2007, Gay et al. 2008), intrapopulation genetic variation, life history such as conditions during larval development (Swindell and Bouzat 2006), and the direction of selection throughout the population sources (Hansen and Houle 2008).
The effect of altitude on wing size (body size) and wing shape was measured using geometric morphometrics methods. Geometric morphometrics is a powerful tool for capturing the shape characteristics in several morphological aspects, particularly head, wings, and genitalia (Zelditch et al. 2004). This method involves examining the structures from which Cartesian coordinates can be taken. Wing size is an index of body size (Tantawy and Vetukhiv 1960, Cowley
and Atchley 1990) and the wings are very appropriate structures for biological shape studies because their 2-D flattened shape bears several useful landmarks (Grodnitsky 1999, Zelditch et al. 2004). Several authors have employed insect wings, particularly in 2-D morphometrical studies in systematics and phylogeny (Rohlf 1993, Debat et al. 2003, Klingenberg 2003, Gumiel et al. 2003).
We focused our collection efforts on the northeasern part of Turkey, because elevation can change dramatically over short distances, providing a convenient surrogate for many variables, such as temperature and land cover, that affect mosquito ecology.
MATERIALS AND METHODS
The study areaMosquito trapping was conducted from June to
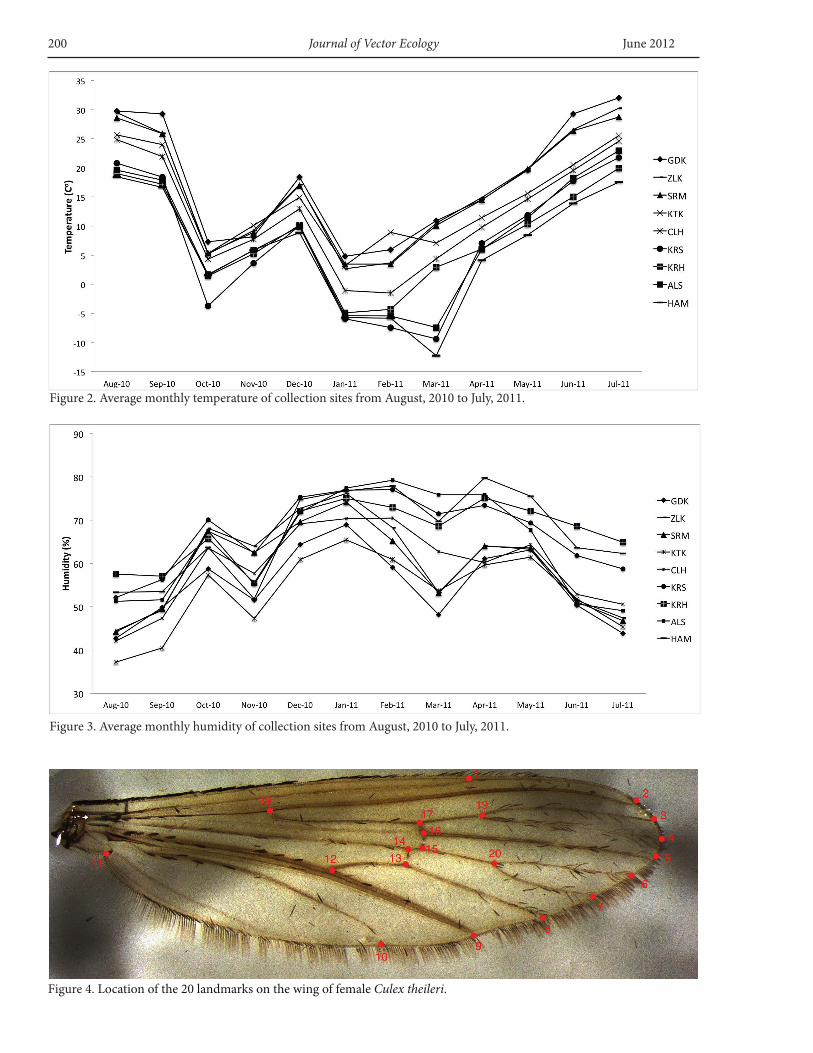
September, 2009 in northeastern Turkey, along habitat-climate-elevation gradients ranging from plain habitats below 1,000 m to foothills low montane areas (1,000-1,700 m), mid-range montane areas (1,700-2,000 m) and high montane areas above 2,000 m. Sampling included sites located along the Aras River and in the Kars Plateau (Figure 1). Aerial distance between the most remote locations (Godekli and Hamamli) is ≈200 km and the distance between the closest sites (Alisofu and Hamamli) is ≈5 km. Selected environmental site characteristics are given in Table 1. Data loggers were placed at sampling stations to record temperature and humidity three times per day. Monthly temperature and humidity obtained from data loggers are given in Figures 2 and 3.
Kars plateau is located in the coldest region in Turkey and continental climate conditions prevail in the area. It is covered with snow for four to five months each year and the thickness of the snow can reach 80 cm in some areas. According to the average climatic data of the 70 years between 1935 and 2005 (T.S.M.S), the annual average temperature was 4.4° C; the highest temperature average was in July (17.3° C) and August (17.4° C) and the lowest temperature average was in January (-11.2° C). The annual rainfall is 489.5 mm.
The Aras River emerges in the vicinity of Erzurum Province in eastern Anatolia and flows into the Caspian
City Abbr. Altitude (m) Latitude Longitude Habitat typeGodekli GDK 808 39.820188° 44.596330° The Igdır Plain, with many mosquito breeding
habitats, intensive agricultural activities.Zulfikarkoy ZLK 848 39.994245° 44.145304°Surmeli SRM 944 40.065487° 43.787455° Transition zone between the Aras Valley and the
Igdır Plain, limited breeding habitats, intensive agricultural activities.
Kotek KTK 1,350 40.253812° 42.943545°Cilehane CLH 1,620 40.220713° 43.015578°Kars KA 1,768 40.592670° 43.077831°
Kars Plateau, limited agricultural activities and limited breeding habitats.
Karahamza KR 1,876 40.431349° 42.754236°Alisofu ALS 2,070 40.342701° 42.682848°Hamamli HAM 2,130 40.306989° 42.700422°
Table 1. Some ecological features of sampling stations.

Vol. 37, no. 1 Journal of Vector Ecology 199
Figure 1. Collection sites of Culex theileri from the northeast part of Turkey. A). Elevation map of collection sites. B). Habitat type map of collection sites.
Sea in Azerbaijan. The river forms part of the international boundaries of Turkey-Armenia, Iran-Nakhichevan (Azerbaijan), and Iran-Armenia. The Aras Valley is an important ecological corridor where desert fauna come into Anatolia. Suitable climate, poor drainage, high water table, and salinity combine to produce large populations of a number of mosquito species in the valley (Gunduz et al. 2009, Aldemir et al. 2010). Between 1990 and 2004, the annual average temperature, annual total rainfall, and annual average relative humidity in the area were 9.9° C, 250 mm, and 51.8%, respectively.
Mosquito collectionsCx. theileri adults were collected from cattle and sheep
barns using mouth aspirators and New Jersey light traps (NJLTs) containing 40-watt light bulbs. On each trapping night, five to seven light traps were placed in each of the sampling localities. NJLTs were placed 1.5 m above ground, and they were operated from 18:00 to 06:00. Specimens were stored in individual microtubes and kept in dry ice. The specimens were transported to the laboratory the following morning for identification using the keys and descriptions provided by Schaffner et al. (2001).
DNA extraction and SNP discoveryWhole mosquito tissue, except wings, was homogenized
using a Qiagen Tissuelyser and genomic DNA extracted
using a BioSprint96 DNA Blood Kit (Qiagen, CA). 12 genes; cytochrome b (CYTB), cytochrome c oxidase subunit 1 (COI), esterase B1 (ESTB1), fatty acid-binding protein (FABP), forkhead transcription factor (FOXO), heat shock protein 70 (HSP70), myosin light chain 2 (MyoLC-2), odorant receptor 10 (ODR10), v-type ATP synthase (VATPS), vitellogenin (VIT), and 5.8s rRNA were selected for this study and DNA sequences of specimens from different altitudes were used for SNPs discovery. Amplification and/or sequencing of all 12 DNA fragments was not possible in all the individuals we studied; therefore the number of individuals varied for different fragments (from a minimum number of 14 to maximum of 20).
Primers, except for the cytochrome B gene, were obtained from Dr. Anthony J. Cornel, University of California. Two primers (N1N-PDR and C3B-PDR) published in a previous study were used for amplifying the cytb gene (Esseghir et al. 1997) and the sequence data of the COI gene region used for this study were obtained from GenBank (Accession number: FJ210900). The full list of primer sequences are summarized in Table 2.
For amplification of the cytb gene fragment, the 50ul PCR reaction contained 0.4 µM of each forward and reverse primer, 1X PCR reaction buffer (Applied Biosystems), 2 µM MgCl2, 200 µM dNTP mix, 0.5U Taq polymerase, and 3µl of DNA template. Thermal cycling was performed using the following program: 2 min at 94° C; 2 cycles for 30 s at
200 Journal of Vector Ecology June 2012
Figure 2. Average monthly temperature of collection sites from August, 2010 to July, 2011.
Figure 3. Average monthly humidity of collection sites from August, 2010 to July, 2011.
Figure 4. Location of the 20 landmarks on the wing of female Culex theileri.

Vol. 37, no. 1 Journal of Vector Ecology 201
94° C, 30 s at 40° C, 1 min at 72° C; 30 cycles of 30 s at 94° C, 30 s at 45° C, 1 min at 72° C; followed by a final 3 min extension at 72˚C. For amplification of the other gene fragments, each 50 µl PCR reaction contained 0.5 µM of each forward and reverse primer, 1X PCR reaction buffer (Applied Biosystems), 1.5 mM MgCl2, 200 µM dNTP mix, 1.25U Taq polymerase, and 2 µl of DNA template. Thermal cycling was performed using the following program: 5 min at 95° C; 35 cycles of 30 s at 95° C, 30 s at between 48 and 54° C, 30 s at 72° C; followed by a final 5-min extension at 72° C. For each gene fragment, we optimized the PCR reaction by adjusting either the PCR mix and/or thermal cycling conditions for optimal amplification.
Amplicons were sequenced at the UC DNA Sequencing Facility (College of Biological Sciences, UC Davis) using an ABI 3730 Genetic Analyzer (AppliedBiosystems, Carlsbad, CA). ChromasLite ver. 2.01 (Technelysium, Australia) was used to view chromatograms and convert chromatogram to text sequence. Sequences were aligned using BioEdit version 7.0.5.3 (http://www.mbio.ncsu.edu/RNaseP/info/programs/BIOEDIT/bioedit.html). DnaSP version 5.10.01 was used for sequence analysis. Non-coding and coding regions were based on the gene annotation reported in the mosquito Cx. pipiens quinquefasciatus genome on Ensembl (http://metazoa.ensembl.org). The true reading frame in Cx. theileri may differ from these gene annotations, however the Cx. pipiens quinquefasciatus genome is the best available approximation.
SNP genotyping and validationGenotyping was performed using IPLEX reagents
and protocols for multiplex PCR, single base primer extension (SBE), and generation of mass spectra, as per the manufacturer’s description in the iPLEX assay protocol (http://www.sequenom.com/Assets/pdfs/appnotes/ 8876–006.pdf). The SNPs were validated using matrix-assisted laser desorption/ionization time of-flight mass spectroscopy (MALDI-TOF MS) assays. The IPLEX PCR primer pairs are described in Table 2.
Genetic data analysis Expected heterozygosity (He), observed heterozygosity
(Ho), and tests for differences of He and Ho among groups were computed using Arlequin version 3.1. We also used Arlequin to test for departures from Hardy-Weinberg proportions, to test for linkage-disequilibrium for each pair of loci within each site, and to determine variation in haplotype frequences among and within regions by analysis of molecular variance (AMOVA). Arlequin software also estimated pairwise FST values and Slatkin linearized FST [FST/(1- FST )] among collections and computed the significance of the variance components associated with each level of genetic structure by a nonparametric permutation test with 100,000 pseudoreplicates.
STRUCTURE ver. 2.2 software was used for the genetic identification of clustered populations, allogenous introductions, and potential admixed colonies with a 100,000 burn-in period and 100,000 Markov chain Monte
Carlo (MCMC) repetitions. This software uses a model-based clustering method to infer population structure using multiloci genotypes. Each individual in the colony was probabilistically assigned to K populations. Runs of various lengths were performed with different numbers of genetic clusters (K), testing all values of K from 1 to 9 (the number of sampling sites). Generally, the number of clusters (K) in the data is inferred from the posterior probability distrubution Pr (K/X) calculated from the posterior probability of the data Pr (X/K) (X being the genotypes of the sampled individuals).
We examined the correlations between genetic distance and geographical distance matrices using Mantel tests as executed by the IBD program using 10,000 matrix randomizations. Geographic distances between sample sites were determined from their latitude and longitude coordinates using the XY to line option of the ARCGIS version 10. To determine if elevation plays a role in structuring populations of Cx. theileri, we calculated the absolute difference in elevation between each pair of sites and constructed a matrix of pairwise elevation differences between sites. We then assessed the effects of elevation on genetic distance using Mantel tests as executed by IBD.
Geometric morphometrics analysisAll specimens were screened under a light microscope
for the presence of known endoparasites to prevent possible variations affecting the morphometric data (Mayr and Ashlock 1991). The body parts were then removed from each specimen with forceps and mounted in Entellane (Merck KgaA, Darmstadt, Germany) on labelled slides. Before mounting, for proper visibility of veins, the wings were kept in 5% KOH solution for 20 min at room temperature to clear hairs. The wings were then placed in 95% alcohol for <10 s, after which they were transferred for washing to cups containing distilled water. Slides were photographed, digitized, and archived. All specimens were scored by a single experimenter (B.D). Photographs were first entered into tps-UTIL1.28 (Rohlf 2007).
A total of 277 specimens was used for the geometric morphometrics anaysis (GDK-38, ZLK-35, PRL-37, CLH-21, KTK-27, KRS-46, KRH-43, ALS-21, HAM-9). Landmarks (20) of the right wings of 277 Cx. theileri females were used for the analysis following the method of Rohlf and Slice (1990) (Figure 4). The landmarks are the intersections of wing veins with the wing margin and intersections of cross veins with major veins. The landmark configurations were scaled, translated, and rotated against the consensus configuration by the GLS Procrustes superimposition method (Bookstein 1991, Rohlf 1993, Alibert et al. 2001). The coordinates were analyzed using tps-RELW1.34 (Rohlf 2007) to calculate the eigen values for each principal warp. Consensus configurations and relative warps for each specimen were conducted. The variability in wing shape was assessed using the scores obtained for each individual on the first two relative warps (technically a principal component analysis). The relative warps correspond to the principal components and define

202 Journal of Vector Ecology June 2012
a shape space in which individuals are replaced (Alibert et al. 2001). As well as examining the effects of location and line on shape, we also tested for clinal changes in shape using a multivariate approach. The TpsReg program was used for this analysis. Mean Procrustes residuals of the individuals were computed and this was followed by a multivariate regression of shape on altitude. Altitude was considered as a linear variable in this analysis. A Canonical Variates Analysis (CANOVAR) was also conducted on landmark data by IMP CVAGEN6n to compare populations collected from different altitudes, using group membership information and Barlett’s test examined differences among groups. Also, squared Mahalanobis distances were derived from CVA in order to quantify the intraspecific phenetic relationships and their statistical significance was computed by nonparametric permutation tests (1,000 runs each) after Bonferroni correction. These distances were used in an unweighted pair-group method with arithmetic average (UPGMA) cluster analysis to produce a dendrogram.
Size morphometry of Cx. theileri was investigated using centroid sizes of wings as an estimator with nonparametric Kruskal-Wallis test (Sokal and Rohlf 1995). Centroid size is the square root of the sum of squared distances of a set of landmarks from their centroid, i.e., the square root of the sum of variances of landmarks about that centroid in x- and y- directions (Bookstein 1991). Altitutinal trend in centroid size were tested by linear regression of line means for each sampling station. Finally, the allometric relationship was tested between wing-shape measures and centroid size. The allometric content of shape was estimated by a simple linear regression of the first relative warp (RW1) on CS. The allometry was also explored by a multivariate regression analysis (shape variables on CS) and its significance estimated according to non-parametric methods based on permutation (1,000 runs) (Good 2000).
RESULTS
Genetic variability and differentiationA total of 2,416 (GDK-94, ZLK-120, PRL-241, CLH-
140, KTK-146, KA-1180, KR-394, ALS-90, HAM-11) adult mosquitoes was collected from nine different sampling stations and 278 of these mosquitoes (GDK-46, ZLK-22, PRL-28, CLH-29, KTK-28, KA-35, KR-35, ALS-44, HAM-11) were analyzed in this study. Cx. theileri’s abundance was not linearly changed by elevation, but while it was more abundant in Kars and Karahamza, its abundance decreased toward the lowest/highest elevation. Also, we could only sample 11 individuals from Hamamli.
Within each population, deviations from the Hardy-Weinberg predictions were found in four of the 22 loci, showing a strong heterozygote excess (ODOR-507, ODOR-48, HSP70-117, HSP70-492) (Table 2). Consequently, all analyses were done with and without these loci. Linkage disequilibrium was observed in three pairs of loci ( FOXO-084 and FOXO-228; MLC2-066#1 and MLC2-066#2; VIT-026#1 and VIT-026#1) and all analyses were performed by choosing one locus of each pair that were in linkage
disequilibrium.Structure analysis and pair-wise Fst values indicated
that there is no genetic differentiation among populations. FST values are very small and not significantly different from zero for all pairwise comparisons. The pairwise Fst values and p values between populations are reported in Table 3. When considering all individuals, the posterior probability of the data for a given value of K is highest for K= 1. Because of this lack of geographic population structure, further investigation by means of Mantel tests was not carried out.
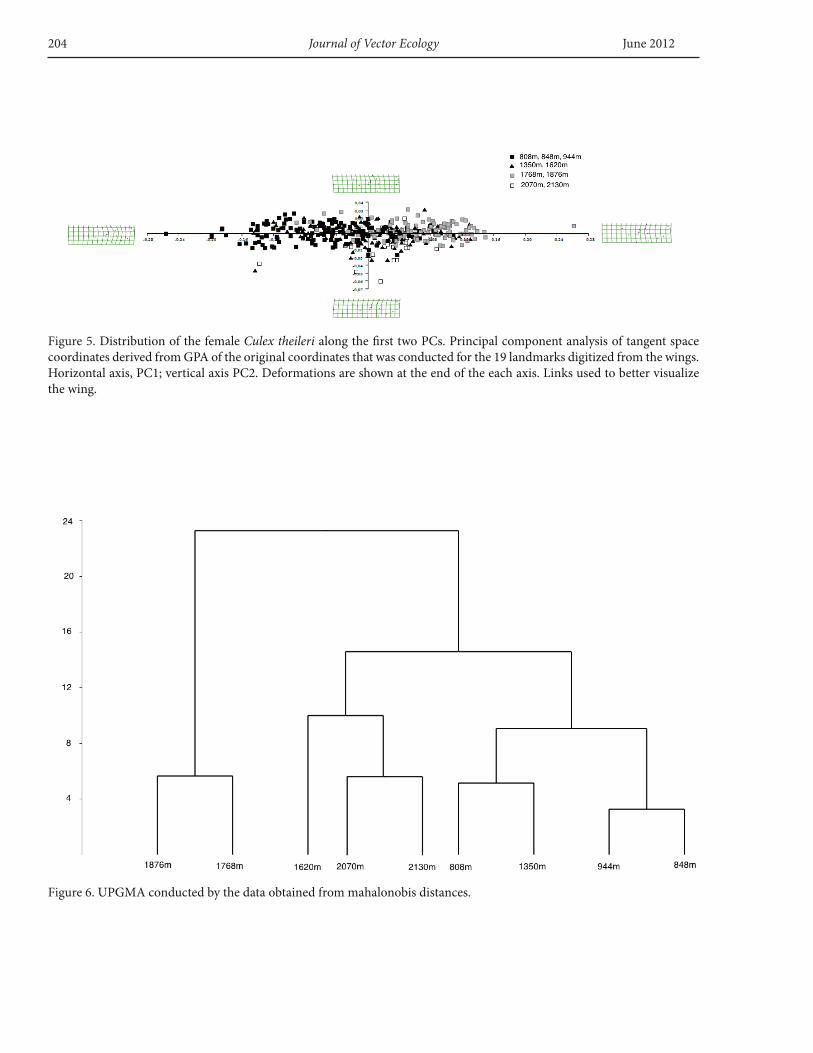
Geometric morphometricsWhen a PCA was conducted on the 20 wing landmarks,
the two first PCs summarized 19.85% and 14.79% of the total variance, respectively (Figure 5). Main deformations centered on the medial of the wing on landmarks 14-15 and 16 (Figure 4). To examine clinal patterns, a multivariate regression of shape on altitude was undertaken. When partial warps and uniform components were considered, the regression of shape variables on altitude was significant (r2 =0.014, F=3.346, p<0.05).
However, CVA (Canonical Variates Analyses-CANOVAR) show some overlap in local areas (figure not shown). There were three statistically significant groups: MANOVA Axis 1 Lambda=0.1477, χ2=484.8621 df=288, p=3.19744e-012, Axis 2 Lambda=0.2561, χ2=345.2950 df=245, p=2.48224e-005, and Axis 3 Lambda=0.3667, χ2=254.3003 df=204, p=0.00956143. When the shape differences of populations were analyzed by UPGM derived from square Mahalonobis distances, KRH (1,786) and KRS (1,876 m) comprised the first group, GDK (808 m), ZLK (848 m), SRM (944 m), KTK (1350 m) comprised the second group, and CLH (1,620 m), ALS (2,070 m), HAM (2,130 m) populations comprised the third group (Figure 6), with a pattern showing KRH (1,786 m) and KRS (1,876 m) populations were further from the other populations.
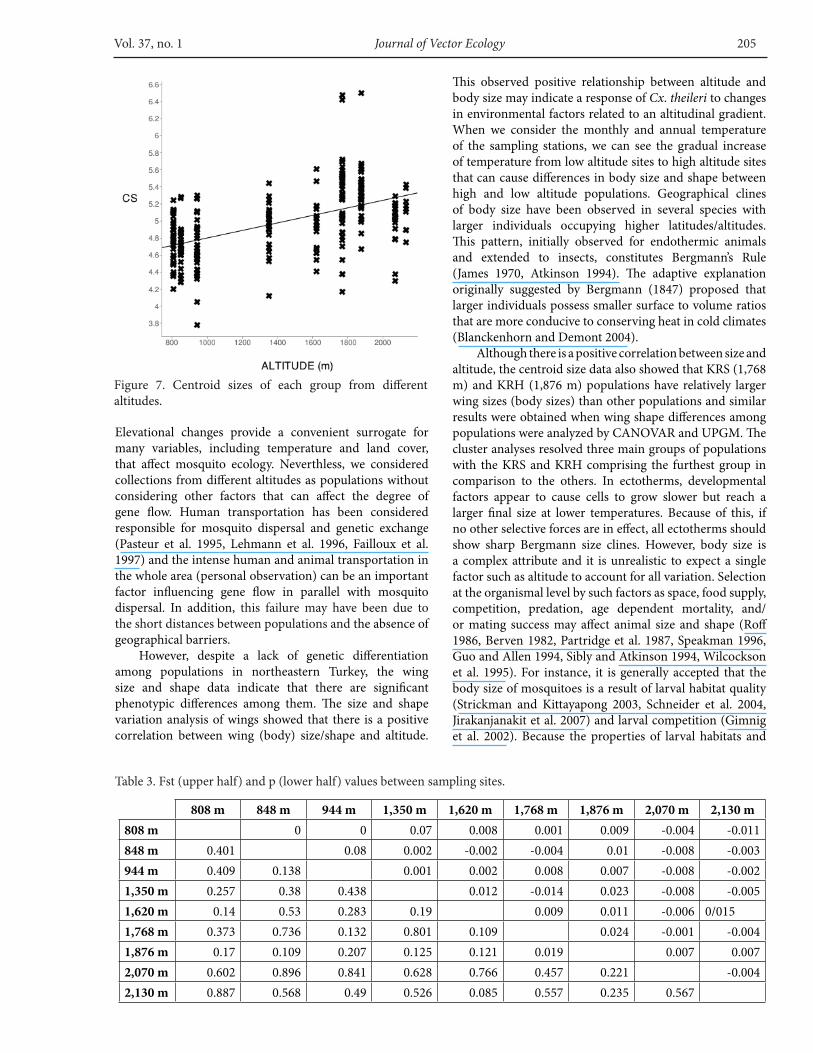
Centroid sizes were used as measures of overall wing size differences among different regions for each individual. The size differences were significant between most populations with samples from populations at an elevation of 1,768 m and 1,876 m having relatively larger wings (Figure 7). Centroid size showed a linear association with altitude (r2 =0.287, F=110.8, p< 0.0000), indicating that individuals with larger body sizes occurred at higher altitudes (Figure 7). However, the test for allometry between centroid size and principal component scores indicated there is no significant relationship between size and shape (r2 =0.003, F=0.925, p=0.2).
DISCUSSION
Based on our data, we conclude that populations of Cx. theileri are not genetically differentiated in northeast Turkey. Failure to find genetic differentiation may to some extent relate to our sampling scheme, which was conservative with regard to our hypothesis. As mentioned above, we focused collection efforts on the northeastern part of Turkey because of the dramatic elevation changes over short distances.
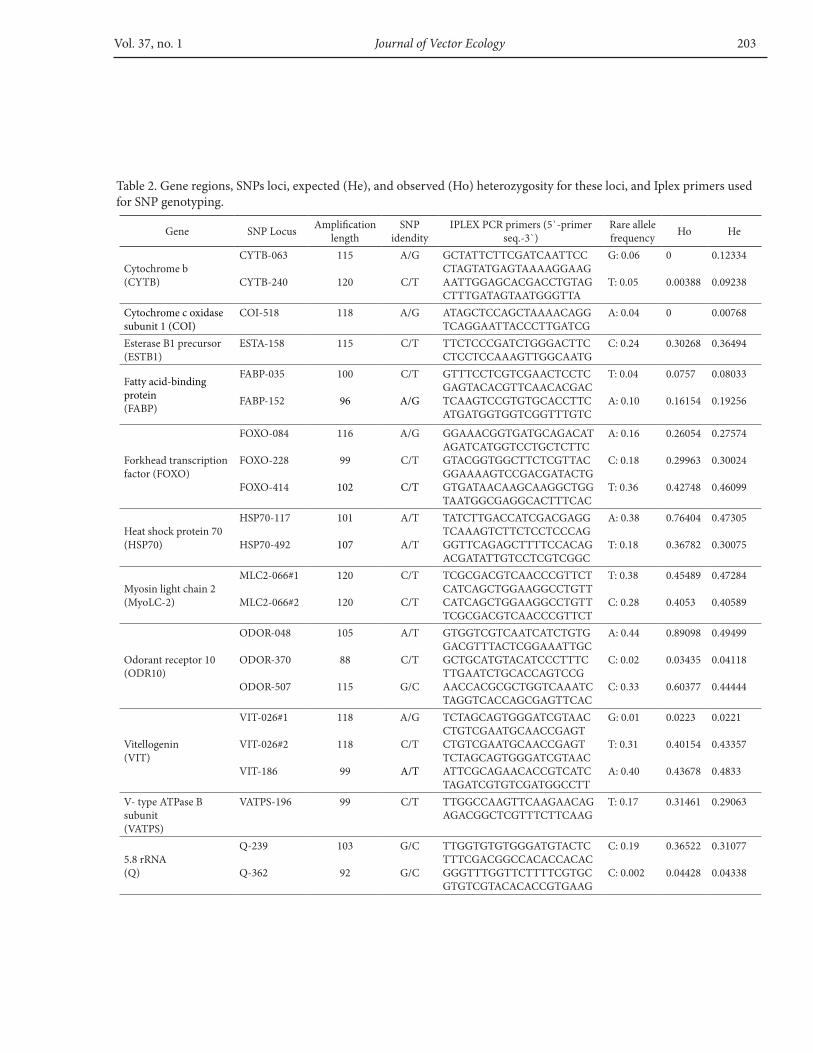
Vol. 37, no. 1 Journal of Vector Ecology 203
Gene SNP Locus Amplification length
SNP idendity
IPLEX PCR primers (5`-primer seq.-3`)
Rare allele frequency Ho He
Cytochrome b(CYTB)
CYTB-063
CYTB-240
115
120
A/G
C/T
GCTATTCTTCGATCAATTCCCTAGTATGAGTAAAAGGAAGAATTGGAGCACGACCTGTAGCTTTGATAGTAATGGGTTA
G: 0.06
T: 0.05
0
0.00388
0.12334
0.09238
Cytochrome c oxidase subunit 1 (COI)
COI-518 118 A/G ATAGCTCCAGCTAAAACAGGTCAGGAATTACCCTTGATCG
A: 0.04 0 0.00768
Esterase B1 precursor(ESTB1)
ESTA-158 115 C/T TTCTCCCGATCTGGGACTTCCTCCTCCAAAGTTGGCAATG
C: 0.24 0.30268 0.36494
Fatty acid-binding protein(FABP)
FABP-035
FABP-152
100
96
C/T
A/G
GTTTCCTCGTCGAACTCCTCGAGTACACGTTCAACACGACTCAAGTCCGTGTGCACCTTCATGATGGTGGTCGGTTTGTC
T: 0.04
A: 0.10
0.0757
0.16154
0.08033
0.19256
Forkhead transcription factor (FOXO)
FOXO-084
FOXO-228
FOXO-414
116
99
102
A/G
C/T
C/T
GGAAACGGTGATGCAGACATAGATCATGGTCCTGCTCTTCGTACGGTGGCTTCTCGTTACGGAAAAGTCCGACGATACTGGTGATAACAAGCAAGGCTGGTAATGGCGAGGCACTTTCAC
A: 0.16
C: 0.18
T: 0.36
0.26054
0.29963
0.42748
0.27574
0.30024
0.46099
Heat shock protein 70(HSP70)
HSP70-117
HSP70-492
101
107
A/T
A/T
TATCTTGACCATCGACGAGGTCAAAGTCTTCTCCTCCCAGGGTTCAGAGCTTTTCCACAG ACGATATTGTCCTCGTCGGC
A: 0.38
T: 0.18
0.76404
0.36782
0.47305
0.30075
Myosin light chain 2(MyoLC-2)
MLC2-066#1
MLC2-066#2
120
120
C/T
C/T
TCGCGACGTCAACCCGTTCTCATCAGCTGGAAGGCCTGTTCATCAGCTGGAAGGCCTGTTTCGCGACGTCAACCCGTTCT
T: 0.38
C: 0.28
0.45489
0.4053
0.47284
0.40589
Odorant receptor 10(ODR10)
ODOR-048
ODOR-370
ODOR-507
105
88
115
A/T
C/T
G/C
GTGGTCGTCAATCATCTGTGGACGTTTACTCGGAAATTGCGCTGCATGTACATCCCTTTCTTGAATCTGCACCAGTCCGAACCACGCGCTGGTCAAATCTAGGTCACCAGCGAGTTCAC
A: 0.44
C: 0.02
C: 0.33
0.89098
0.03435
0.60377
0.49499
0.04118
0.44444
Vitellogenin(VIT)
VIT-026#1
VIT-026#2
VIT-186
118
118
99
A/G
C/T
A/T
TCTAGCAGTGGGATCGTAACCTGTCGAATGCAACCGAGTCTGTCGAATGCAACCGAGTTCTAGCAGTGGGATCGTAACATTCGCAGAACACCGTCATCTAGATCGTGTCGATGGCCTT
G: 0.01
T: 0.31
A: 0.40
0.0223
0.40154
0.43678
0.0221
0.43357
0.4833
V- type ATPase B subunit(VATPS)
VATPS-196 99 C/T TTGGCCAAGTTCAAGAACAGAGACGGCTCGTTTCTTCAAG
T: 0.17 0.31461 0.29063
5.8 rRNA(Q)
Q-239
Q-362
103
92
G/C
G/C
TTGGTGTGTGGGATGTACTCTTTCGACGGCCACACCACACGGGTTTGGTTCTTTTCGTGCGTGTCGTACACACCGTGAAG
C: 0.19
C: 0.002
0.36522
0.04428
0.31077
0.04338
Table 2. Gene regions, SNPs loci, expected (He), and observed (Ho) heterozygosity for these loci, and Iplex primers used for SNP genotyping.
204 Journal of Vector Ecology June 2012
Figure 6. UPGMA conducted by the data obtained from mahalonobis distances.
Figure 5. Distribution of the female Culex theileri along the first two PCs. Principal component analysis of tangent space coordinates derived from GPA of the original coordinates that was conducted for the 19 landmarks digitized from the wings. Horizontal axis, PC1; vertical axis PC2. Deformations are shown at the end of the each axis. Links used to better visualize the wing.
Vol. 37, no. 1 Journal of Vector Ecology 205
Elevational changes provide a convenient surrogate for many variables, including temperature and land cover, that affect mosquito ecology. Neverthless, we considered collections from different altitudes as populations without considering other factors that can affect the degree of gene flow. Human transportation has been considered responsible for mosquito dispersal and genetic exchange (Pasteur et al. 1995, Lehmann et al. 1996, Failloux et al. 1997) and the intense human and animal transportation in the whole area (personal observation) can be an important factor influencing gene flow in parallel with mosquito dispersal. In addition, this failure may have been due to the short distances between populations and the absence of geographical barriers.
However, despite a lack of genetic differentiation among populations in northeastern Turkey, the wing size and shape data indicate that there are significant phenotypic differences among them. The size and shape variation analysis of wings showed that there is a positive correlation between wing (body) size/shape and altitude.
This observed positive relationship between altitude and body size may indicate a response of Cx. theileri to changes in environmental factors related to an altitudinal gradient. When we consider the monthly and annual temperature of the sampling stations, we can see the gradual increase of temperature from low altitude sites to high altitude sites that can cause differences in body size and shape between high and low altitude populations. Geographical clines of body size have been observed in several species with larger individuals occupying higher latitudes/altitudes. This pattern, initially observed for endothermic animals and extended to insects, constitutes Bergmann’s Rule (James 1970, Atkinson 1994). The adaptive explanation originally suggested by Bergmann (1847) proposed that larger individuals possess smaller surface to volume ratios that are more conducive to conserving heat in cold climates (Blanckenhorn and Demont 2004).
Although there is a positive correlation between size and altitude, the centroid size data also showed that KRS (1,768 m) and KRH (1,876 m) populations have relatively larger wing sizes (body sizes) than other populations and similar results were obtained when wing shape differences among populations were analyzed by CANOVAR and UPGM. The cluster analyses resolved three main groups of populations with the KRS and KRH comprising the furthest group in comparison to the others. In ectotherms, developmental factors appear to cause cells to grow slower but reach a larger final size at lower temperatures. Because of this, if no other selective forces are in effect, all ectotherms should show sharp Bergmann size clines. However, body size is a complex attribute and it is unrealistic to expect a single factor such as altitude to account for all variation. Selection at the organismal level by such factors as space, food supply, competition, predation, age dependent mortality, and/or mating success may affect animal size and shape (Roff 1986, Berven 1982, Partridge et al. 1987, Speakman 1996, Guo and Allen 1994, Sibly and Atkinson 1994, Wilcockson et al. 1995). For instance, it is generally accepted that the body size of mosquitoes is a result of larval habitat quality (Strickman and Kittayapong 2003, Schneider et al. 2004, Jirakanjanakit et al. 2007) and larval competition (Gimnig et al. 2002). Because the properties of larval habitats and
Figure 7. Centroid sizes of each group from different altitudes.
Table 3. Fst (upper half) and p (lower half) values between sampling sites.
808 m 848 m 944 m 1,350 m 1,620 m 1,768 m 1,876 m 2,070 m 2,130 m808 m 0 0 0.07 0.008 0.001 0.009 -0.004 -0.011848 m 0.401 0.08 0.002 -0.002 -0.004 0.01 -0.008 -0.003944 m 0.409 0.138 0.001 0.002 0.008 0.007 -0.008 -0.0021,350 m 0.257 0.38 0.438 0.012 -0.014 0.023 -0.008 -0.0051,620 m 0.14 0.53 0.283 0.19 0.009 0.011 -0.006 0/0151,768 m 0.373 0.736 0.132 0.801 0.109 0.024 -0.001 -0.0041,876 m 0.17 0.109 0.207 0.125 0.121 0.019 0.007 0.0072,070 m 0.602 0.896 0.841 0.628 0.766 0.457 0.221 -0.0042,130 m 0.887 0.568 0.49 0.526 0.085 0.557 0.235 0.567

206 Journal of Vector Ecology June 2012
crowding were not recorded in this study, we cannot exclude their influence on variation in the observed size differences. Similar to size, wing shape might be influenced by many factors in the microenvironment. Variation in wing shape can be related to flight performance and therefore selection against dispersal at higher altitudes can be driving variation. Nonlinear patterns for shape might also arise from selection as environmental variables such as relative humidity show nonlinear changes with latitude (Hoffmann and Shirriffs 2002).
Several factors such as gene flow between selective environments (Garant et al. 2007), divergent natural selection (Schluter 2000), and phenotypic plasticity (Pigliucci 2001) may influence adaptive phenotypic divergence among populations. In the presence of gene flow between populations, the rate of local adaptation of non-plastic traits can be constrained, favoring the evolution of phenotypic plasticity (Antonovics 1976, Bradshaw 1965). Plasticity could allow individuals to phenotypically adapt to new conditions within one or two generations, and thus would allow them to persist in novel environments. Single genotypes can change an organism’s chemistry, physiology, development, morphology, or behavior in response to environmental cues (Agrawal 2001). The ability of an organism to produce alternative phenotypes as a response to environmental change is often an adaptive strategy to minimize loss of fitness in a harsher environment or to maximize fitness in a more favorable environment. At the same time, high gene flow may result in the selection for increased plasticity (Crispo 2008). Conceivably, if individuals are highly plastic with respect to traits that are adaptive in alternate forms under different environmental conditions, increased gene flow between selective environments may be permitted (Crispo 2008). Some studies have shown that newly colonized populations and invasive species are more plastic than their native counterparts, suggesting that plasticity may have facilitated colonization of the newly occupied areas (reviewed by Daehler 2003, Richards et al. 2006). The presence of Culex larvae and adults throughout the year in the in the western Orange Free State in southern Africa, where in summer ambient temperature may reach a maximum of 37° C, whereas the minimum temperature in winter may be as low as -10° C, may indicate high plasticity for temperature tolerance (Van Der Linde et al. 1990) and the broad temperature tolerance of this species probably makes an important contribution to its wide temporal and geographic distribution (Hewitt et al. 1982).
The larger body size for KRS and KRH might indicate a gene x environment relationship with one genotype having a larger phenotypic value in one environment than in others. Cx. theileri experiences a broad range of environments in both temporal and spatial dimensions. Therefore, it might be more advantageous in terms of fitness to perform relatively well over a range of environmental conditions rather than to perform relatively well in one environment at a cost of performing well in others. Also, the greatest abundance of Cx. theileri in Kars and Karahamza may indicate that this altitude and these environmental conditions are more
suitable in these areas. The association between body size and fitness traits has been frequently investigated in insects. Investigations into the relative fitness of adult mosquitoes in the field have shown that relatively large-bodied females exhibit higher fitness than small-bodied females. Large body size has been related to increased survival and blood-feeding success in mosquito species from different genera (Haramis 1983, Reisen et al. 1984, Hawley 1985). Larger Aedes triseriatus (Say) females are more frequently parous (Haramis 1983), and larger Culex tarsalis Coquillett have higher fecundity (Bock and Milby 1981). However to fully understand the gene x environment relationship on fitness, a comprehensive life history perspective, including body size as one of several traits such as longevity, growth rate, fecundity, age at maturity that vary between different conditions as temperature, humidity, food availability, larval conditions, will be required because it is possible that selection on fitness is acting on a suite of life history traits and accordingly, these experiments should be considered for future studies.
REFERENCES CITED
Abul-Hab, J. 1967. Larvae of culicine mosquitoes in North Iraq. Bull. Entomol. Res. 57: 279-284.
Agrawal, A.A. 2001. Phenotypic plasticity in the interactions and evolution of species. Science 294: 321-326.
Aitken, Th. H.G. 1954. The Culicidae of Sardinia and Corsica (Diptera). Bull. Entomol. Res. 45: 437-494.
Aldemir, A., H. Bedir, B. Demirci, and B. Alten. 2010. Biting activity of mosquito species (Diptera: Culicidae) in the Turkey-Armenia border area, Ararat Valley, Turkey. J. Med. Entomol. 47: 22-27.
Alibert, P., B. Moureau, J.L. Dommergues, and B. David. 2001. Differentiation at a microgeographical scale within two species of ground beetle, Carabus auronitens and C. nemoralis (Coleoptera, Carabidae): A geometrical morphometric approach. Zool. Scripta. 30: 299-316.
Antonovics, J. 1976. The nature of limits to natural selection. Ann. Mo. Bot. Gard. 63: 224 -247.
Atkinson, D. 1994. Temperature and organism size: a biological law for ectotherms. Adv. Ecol. Res. 25: 1–58.
Aytekin, A.M. and N. Cagatay. 2002. A phenetic approach to the subgenera of bumblebees (Apidae: Hymenoptera). Mellifera 2: 60-64.
Azari–Hamidian, S., M.R. Yaghoobi–Ershadi, E. Javadian, M.R. Abai, I. Mobedi, Y.M. Linton, and R.E. Harbach. 2009. Distribution and ecology of mosquitoes in a focus of dirofilariasis in northwestern Iran, with the first finding of filarial larvae in naturally infected local mosquitoes. Med. Vet. Entomol. 23: 111-121.
Becker, N., D. Petri, M. Zgomba, C. Boase, C. Dahl , J. Lane, and A. Kaiser. 2003. Mosquitoes and Their Control. Kluwer Academic Press, NY. 302 pp.
Bergmann, C. 1847. Über die Verhältnisse der Wärmeökonomie der Thiere zu ihrer Grösse. Göttinger Studien pt. 1: 595-708.

Vol. 37, no. 1 Journal of Vector Ecology 207
Berven, K.A. 1982. The genetic basis of attitudinal variation in the wood frog, Rana sylvatica. An experimental analysis of life history traits. Evolution 36: 962-983.
Blanckenhorn, W.U. and M. Demont. 2004. Bergmann and converse Bergmann latitudinal clines in arthropods: Two ends of a continuum? Integr. Comp. Biol. 44: 413-424.
Bock, M.E and M.M Milby. 1981. Seasonal variation of wing length and egg raft size in Culex tarsalis. Proc. Calif. Mosq. Vector Contr. Assoc. 49: 64-66.
Bookstein, F.L. 1991. Morphometric Tools for Landmark Data. Cambridge University Press. 435 pp.
Bradshaw, A.D. 1965. Evolutionary significance of phenotypic plasticity in plants. Adv. Genet. 13: 115–155.
Buggs, R.J. 2007. Empirical study of hybrid zone movement. Heredity 99: 301-312.
Cavicchi, S., D. Guerra, G. Giorgi, and C. Pezzoli. 1985. Temperature-related divergence in experimental populations of Drosophila melanogaster. 1. Genetic and developmental basis of wing size shape variation. Genetics 109: 665-690.
Cowley, D.E. and W.R. Atchley. 1990. Development and quantitative genetics of correlation structure among body parts of Drosophila melanogaster. Am. Nat. 135: 242-268.
Crispo, E. 2008. Modifying effects of phenotypic plasticity on interactions among natural selection, adaptation and gene flow. J. Evolution. Biol. 21: 1460-1469.
Daehler, C.C. 2003. Performance comparisons of co-occurring native and alien invasive plants: implications for conservation and restoration. Annu. Rev. Ecol. Evol. Syst. 34: 183-211.
Debat, V., M. Begin, H. Legout, and J.R. David. 2003. Allometric and nonallometric components of Drosophila wing shape respond differently to developmental temperature. Evolution 57: 2773-2784.
Esseghir, S., P.D. Ready, R. Killick-Kendrick, and R. Ben-Ismail. 1997. Mitochondrial haplotypes and phylogeography of Phlebotomus vectors of Leishmania major. Insect Mol. Biol. 6: 211-225.
Failloux, A.B., M. Raymond, A. Ung, C. Chevillon, and N. Pasteur. 1997. Genetic differentiation associated with commercial traffic in the Polynesian mosquito., Aedes polynesiensis Marks 1951. Biol. J. Linn. Soc. 60: 107-118.
Garant, D., S.E. Forde, and A.P. Hendry. 2007. The multifarious effects of dispersal and gene flow on contemporary adaptation. Funct. Ecol. 21: 434-443.
Gauthier, C. and M. Tibayrenc. 2005. Population structure of malaria parasites: the driving epidemiological forces. Acta. Trop. 94: 241-250.
Gay, L., P.A. Crochet, D.A. Bell, and T. Lenormand. 2008. Comparing clines on molecular and phenotypic traits in hybrid zones: a window on tension zone models. Evolution 62: 2789-2806.
Gimnig, J.E., M. Ombok, S. Otieno, M.G. Kaufman, J.M. Vulule, and E.D. Walker. 2002. Density-dependent
development of Anopheles gambiae (Diptera: Culicidae) larvae in artificial habitats. J. Med. Entomol. 39: 162-172.
Good, P. 2000. Permutation Tests: A Practical Guide to Resampling Methods for Testing Hypotheses. Springer, New York.
Grodnitsky, D.L. 1999. Form and Function of Insect Wings. The Johns Hopkins University Press. Baltimore, MD. 280 pp.
Gumiel, M., S. Catala, F. Noireau, A. Rojas de Arias, A. Garcia, and J.P. Dujardin. 2003. Wing geometry in Triatoma infestans (Klug) and T. melanosoma Martinez, Olmedo and Carcavallo (Hemiptera: Reduviidae). Syst. Entomol. 28: 173-179.
Gunduz, Y.K., A. Aldemir, and B. Alten. 2009. Seasonal dynamics and nocturnal activities of mosquitoes (Diptera: Culicidae) in Aras Valley, Turkey. Turk. J. Zool. 33: 269-276.
Guo, X. and S.K. Allen Jr. 1994. Sex determination and polyploid gigantism in the dwarf-surf clam, Mulinia lateralis Say. Genetics 138: 1199–1206.
Hansen, T.F. and D. Houle. 2008. Measuring and comparing evolvability and constraint in multivariate characters. J. Evol. Biol. 21: 1201-1219.
Harbach, R.E. 1988. The mosquitoes of the subgenus Culex in southwestern Asia and Egypt (Diptera: Culicidae). Contrib. Am. Entomol. Inst. 24: 1-240.
Haramis, L.D. 1983. Increased adult size correlated with parity in Aedes triseriatus. Mosq. News 43: 77-79.
Hawley, W.A. 1985. The effect of larval density on adult longevity of a mosquito, Aedes sierrensis: epidemiological consequences. J. Anim. Ecol. 54: 955-964.
Hewitt, P.H., T.C.K. Van Der Linde, R. Van Pletzen, D.J. Kok, S. Fou- Rie, D.J. Mostert, and A. Nel. 1982. Temporal fluctuations of the numbers of female mosquitoes trapped at site in the western Orange Free State. J. Entomol. Soc. South. Afr. 45: 69-92.
Hoffmann A.A. and J. Shirriffs. 2002. Geographic variation for wing shape in Drosophila serrata. Evolution 56: 1068-1073.
James, F.C. 1970. Geographic size variation in birds and its relationship to climate. Ecology 51: 365-390.
James, A.C. and L. Partridge 1998. Geographic variation in competitive ability in Drosophila melanogaster. Am. Nat. 151: 530-537.
Jirakanjanakit, N., S. Leemingsawat, S. Thongrungkiat, C. Apiwathnasorn, S. Singhaniyom, C. Bellec, and J.P. Dujardin. 2007. Influence of larval density or food variation on the geometry of the wing of Aedes (Stegomyia) aegypti. Trop. Med. Intl. Hlth. 12: 1354- 1360.
Jupp, P.G., B.M. Mclntosh, and R.G. Brown. 1966. Laboratory transmission experiments with West Nile and Sindbis viruses and Culex (Culex) theileri Theobald. S. Afr. J. Med. Sci. 31: 95-97.
Klingenberg, C.P. 2003. Developmental instability as a research tool: Using patterns of fluctuating asymmetry

208 Journal of Vector Ecology June 2012
to infer the developmental origins of morphological integration. In: M. Polak (ed.) Developmental Instability, Causes, and Consequences. pp. 427-442. Oxford University Press.
Lehmann, T., W.A. Hawley, L. Kamau, D. Fontenille, F. Simard, and F.H. Collins. 1996. Genetic differentiation of Anopheles gambiae populations from East and West Africa: comparison of microsatellite and allozyme loci. Heredity 77 : 192-208.
May, R.M. and R.M. Anderson. 1983. Epidemiology and genetics in the coevolution of parasites and hosts. Proc. R. Soc. Lond. 219: 281-313.
Mayr, E. and P.D. Ashlock. 1991. Principles of Systematic Zoology. McGraw-Hill, New York. 416 pp.
Mclntosh, B.M., P.G. Jupp, D.B. Dickinson, G.M. McGillivary, and J. Sweetnam. 1967. Ecological studies on Sindbis and West Nile viruses in South Africa. I. Viral activity as revealed by infection of mosquitoes and sentinel fowls. S. Afr. J. Med. Sci. 32: 1-14.
Morin, J.P., B. Moreteau, G. Pétavy, R. Parkash, and J.R. David. 1999. Divergence of reaction norms of size characters between tropical and temperate populations of Drosophila melanogaster and D. simulans. J. Evol. Biol. 12: 329-339.
Morlais, I. and D.W. Severson. 2003. Intraspecific DNA variation in nuclear genes of the mosquito Aedes aegypti. Insect. Mol. Biol. 12: 631–639.
Partridge, L., E. Ewing, and A. Chandler. 1987. Male size and mating success in Drosophila melanogaster. The role of female and male behavior. Anim. Behav. 35: 555-562.
Pasteur, N., M. Marquine, F. Rousset, A.B. Failloux, C. Chevillon, and M. Raymond. 1995. The role of passive migration in the dispersal of resistance genes in Culex pipiens quinquefasciatus within French Polynesia. Genet. Res. 66: 139-146.
Pigliucci, M. 2001. Phenotypic Plasticity: Beyond Nature and Nuture. Johns Hopkins University Press, Baltimore, MD. 344 pp.
Ramos, H.C, H. Ribeiro, C.A. Pires, and R.A. Capela. 1977. Research on the mosquitoes of Portugal (Diptera, Culicidae). II - The mosquitoes of the Algarve. Ann. Inst. Hyg. Med. Trop. 5: 237-256.
Reisen, W.K, M.M. Milby, and M.E Bock. 1984.The effects of immature stress on selected events in the life history of Culex tarsalis. Mosq. News 44: 385-395.
Richards, C.L., O. Bossdorf, N.Z .Muth, J. Gurevitch, and M. Pigliucci. 2006. Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecol. Lett. 9: 981-993.
Ricklefs, R.E. and D.B. Miles. 1994. Ecological and evolutionary inferences from morphology: an ecological perspective. In: P.C. Wainwright and S.M. Reilly (eds.) Ecological Morphology. pp. 13-41. University of Chicago Press.
Roff, A. 1986. Predicting body size with life history models. BioScience 36: 316-323.
Rohlf, F.J. 1993. Relative warp analysis and an example of
its application to mosquito wings. In: L.F. Marcus, E. Bello, and A. Garcia-Valdecasas (eds.). Contributions to Morphometrics. Madrid, Museu Nacional de Ciencias Naturales. 264 pp.
Rohlf, F.J. 2003. TPSdig. Software. Version 1.38. Department of Ecology and Evolution, State University of New York at Stony Brook.
Rohlf, F.J. 2007. TpsUTIL. Department of Ecology and Evolution, State University of New York at Stony Brook.
Rohlf, F.J. and D. Slice. 1990. Extensions of the procrustes method for the optimal superimposition of landmarks. Syst. Zool. 39: 40-59.
Santa-Ana, M., M. Khadem and R. Capela. 2006. Natural infection of Culex theileri (Diptera: Culicidae) with Dirofilaria immitis (Nematoda: Filarioidea) on Madeira Island, Portugal. J. Med. Entomol. 43: 104-106.
Schaffner, F., G. Angel, B. Geoffroy, J.P. Hervy, A. Rhaiem, and J. Brunhes. 2001. Les moustiques dÕEurope (The mosquitoes of Europe), CD-ROM. Institut de Recherche pour Development/EID Mediterrane’ e, Montpellier, France.
Schluter, D. 2000. The Ecology of Adaptive Radiation. Oxford University Press, New York, NY.
Schneider, J.R., A.C. Morrison, H. Astete, T.W Scott, and M.L. Wilson. 2004. Adult size and distribution of Aedes aegypti (Diptera: Culicidae) associated with larval habitats in Iquitos, Peru. J. Med. Entomol. 41: 634-642.
Sibley, R.M. and D. Atkinson. 1994. How rearing temperature affects optimal adult size in ectotherms. Func. Ecol. 8: 486-493.
Silver, J.B. and M. Renshaw. 1999. Are mosquitoes from lower latitudes smaller? Med. Vet. Entomol. 13: 219-223.
Simsek, F.M. 2004. Seasonal larval and adult population dynamics and breeding habitat diversity of Culex theileri Theobald, 1903 (Diptera: Culicidae) in the Golbasi Dis- trict, Ankara, Turkey. Turk J. Zool. 28: 337-344.
Sokal, R.R. and F.J. Rohlf. 1995. Biometry. The Principles and Practice of Statistics in Biological Research. 3rd ed. W.H. Freeman and Company, NY. 887 pp.
Speakman, J.R. 1996. Energetics and the evolution of body size in small terrestrial mammals. Symp. Zool. Soc. Land. 69: 6-1.
Strickman, D. and P. Kittayapong. 2003. Dengue and its vectors in Thailand: calculated transmission risk from total pupal counts of Aedes aegypti and association of wing-length measurements with aspects of the larval habitat. Am. J. Trop. Med. Hyg. 68: 209-217.
Swindell, W. and J.L. Bouzat. 2006. Associations between environmental stress, selection history and quantitative genetic variation in Drosophila melanogaster. Genetica 127: 311-320.
Tantawy, A.O. and M.O. Vetukhiv. 1960. Effects of size on fecundity, longevity and viability in populations of Drosophila pseudoobscura. Am. Nat. 94: 395-403.
Van’T Land, J., P. Van Putten, B. Zwaan, A. Kamping, and W. Van Delden. 1999. Latitudinal variation in wild

Vol. 37, no. 1 Journal of Vector Ecology 209
populations of Drosophila melanogaster: Heritabilities and reaction norms. J. Evol. Biol. 12: 222-223.
Van Der Linde, T.C.K., P.H. Hewitt, A. Nel, and M.C. Van Der Westhuizen. 1990. Development rates and percentage hatching of Culex theileri Theobald (Diptera: Culicidae) eggs at various temperatures. J. Entomol. Soc. S. Afr. 53: 17-26.
Wilcockson, R.W., C.S. Crean, and T.H. Day. 1995. Heritability of a sexually selected character expressed only in males. Nature 374: 158-159.
Zelditch, M.L., H.D. Swiderski, H.D. Sheets, and W.L. Fink. 2004. Geometric Morphometrics for Biologists. Elsevier Academic Press. London. 443 pp.
Related Documents











