Allosteric Regulation and Temperature Dependence of Oxygen Binding in Human Neuroglobin and Cytoglobin MOLECULAR MECHANISMS AND PHYSIOLOGICAL SIGNIFICANCE* Received for publication, June 25, 2004, and in revised form, July 28, 2004 Published, JBC Papers in Press, August 6, 2004, DOI 10.1074/jbc.M407126200 Angela Fago‡§¶, Christian Hundahl‡§, Sylvia Dewilde**, Kambiz Gilany, Luc Moens, and Roy E. Weber‡ From the ‡Department of Zoophysiology, Institute of Biological Sciences, C. F. Møllers Alle 131, Aarhus University, DK-8000 Aarhus C, Denmark, and the Department of Biomedical Sciences, University of Antwerp, B-2610 Antwerp, Belgium Two new globin proteins have recently been discovered in vertebrates, neuroglobin in neurons and cytoglobin in all tissues, both showing heme hexacoordination by the distal His(E7) in the absence of gaseous ligands. In anal- ogy to hemoglobin and myoglobin, neuroglobin and cyto- globin are supposedly involved in O 2 storage and deliv- ery, although their physiological role remains to be solved. Here we report O 2 equilibria of recombinant hu- man neuroglobin (NGB) and cytoglobin (CYGB) measured under close to physiological conditions and at varying temperature and pH ranges. NGB shows both alkaline and acid Bohr effects (pH-dependent O 2 affinity) and tem- perature-dependent enthalpy of oxygenation. O 2 and CO binding equilibrium studies on neuroglobin mutants strongly suggest that the bound O 2 is stabilized by inter- actions with His(E7) and that this residue functions as a major Bohr group in the presence of Lys(E10). As shown by the titration of free thiols with 4,4-dithiodipyridine and by mass spectrometry, this mechanism of modulating O 2 affinity is independent of formation of an internal di- sulfide bond under the experimental conditions used, which stabilize thiols in the reduced form. In CYGB, O 2 binding is cooperative, consistent with its proposed di- meric structure. Similar to myoglobin but in contrast to NGB, O 2 binding to CYGB is pH-independent and exother- mic throughout the temperature range investigated. Our data support the hypothesis that CYGB may be involved in O 2 -requiring metabolic processes. In contrast, the lower O 2 affinity in NGB does not appear compatible with a physiological role involving mitochondrial O 2 supply at the low O 2 tensions found within neurons. Neuroglobin (Ngb) 1 and cytoglobin (Cygb) are recently dis- covered members of the vertebrate globin family, which in- cludes the probably most extensively studied proteins, hemo- globin (Hb), and myoglobin (Mb). Both are expressed at low levels (M range), Ngb in various regions of the brain and in the retinal cells (1– 4) and Cygb in all of the tissues so far investi- gated (5, 6). An important characteristic is their heme hexaco- ordination in the absence of exogenous ligands where the distal histidine, His(E7), occupies the sixth coordination position of the iron atom in both the ferrous and the ferric valency states (7–12). Before the discovery of Ngb and Cygb, globin heme hexacoordination was regarded as a feature of certain plant hemoglobins and invertebrate or bacterial globins but not of vertebrate globins. In contrast to Ngb and Cygb, Hb and Mb are present at high levels (mM range) in the red blood cells and in the striated muscle of vertebrates and show pentacoordinate heme geometry when deoxygenated. Despite their low degree of sequence similarity with other vertebrate globins, human NGB and CYGB show the same characteristic -helix globin fold as vertebrate Hb and Mb (7, 10). A phylogenetic tree for human globin proteins indicates that NGB diverged early in evolution and may represent the most ancestral human globin protein, whereas CYGB ap- pears to be more closely related to Mb (13). Despite their ancient origin, neither protein has changed significantly dur- ing evolution (13), which may reflect conservation of a critical biological function. Although monomeric, Ngb is capable of conformational changes that affect ligand affinity. A number of studies have reported the existence of at least two identifiable conformations on the basis of heterogeneity of reaction rates (8, 14) and infrared (15), NMR (16), and resonance Raman spectra (9) that are indicative of an open and closed conformation of the heme pocket. Two distinguishable conformations resulting in a dif- ferent heme pocket accessibility have been identified in a CYGB mutant by x-ray crystallography (10). Also, the forma- tion of an internal disulfide bond increases the rate of dissoci- ation of the distal His from the heme and thereby O 2 affinity in NGB and to a lesser extent in CYGB (17). Although our knowledge of the structural properties of Ngb and Cygb is rapidly increasing, the physiological role of Ngb and Cygb is still largely unknown. A major hint to the role of Ngb came from the work of Sun et al. (18, 19) who found that Ngb may protect neurons during episodes of O 2 lack. This important finding has been often referred to in the context of the general view that Ngb is involved in O 2 binding and deliv- ery to sustain mitochondrial respiration similar to Hb and Mb. Remarkably, the function of certain nerve globins from inver- * This work was supported by the Danish Natural Science Research Council and the EU (Grant QLG3-CT-2002-01548). The costs of publi- cation of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. § Both authors contributed equally to this work. ¶ To whom correspondence should be addressed: Dept. of Zoophysi- ology, Institute of Biological Sciences, C. F. Møllers Alle 131, Aarhus University, DK-8000 Aarhus C, Denmark. Tel.: 45-89422591; Fax: 45- 86194186; E-mail: [email protected]. ** Postdoctoral fellow of the Fund for Scientific Research (Flanders University). 1 The abbreviations used are: Ngb, neuroglobin; NGB, human neuro- globin; Cygb, cytoglobin; CYGB, human cytoglobin; FPLC, fast protein liquid chromatography; Mb, myoglobin; Hb, hemoglobin; HbA 0 , FPLC- purified adult human hemoglobin; 4-PDS, 4,4-dithiodipyridine; H(E7)V, His(E7)3Val mouse Ngb mutant; H(E7)Q, His(E7)3Gln hu- man NGB mutant; K(E10)L, Lys(E10)3Leu human NGB mutant; H(E7)L/K(E10)L, double mutant His(E7)3Leu, Lys(E10)3Leu of hu- man NGB; GSS, triple mutant Cys(CD7)3Gly, Cys(D5)3Ser, Cys(G19)3Ser of human NGB; DTT, 1,4-dithioerythritol; MALDI-TOF, matrix-assisted laser desorption ionization time-of-flight. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 43, Issue of October 22, pp. 44417–44426, 2004 © 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. This paper is available on line at http://www.jbc.org 44417 at TEXAS TECH UNIVERSITY - Trial Subscription on March 8, 2009 www.jbc.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Allosteric Regulation and Temperature Dependence ofOxygen Binding in Human Neuroglobin and CytoglobinMOLECULAR MECHANISMS AND PHYSIOLOGICAL SIGNIFICANCE*
Received for publication, June 25, 2004, and in revised form, July 28, 2004Published, JBC Papers in Press, August 6, 2004, DOI 10.1074/jbc.M407126200
Angela Fago‡§¶, Christian Hundahl‡§, Sylvia Dewilde�**, Kambiz Gilany�, Luc Moens�,and Roy E. Weber‡From the ‡Department of Zoophysiology, Institute of Biological Sciences, C. F. Møllers Alle 131, Aarhus University,DK-8000 Aarhus C, Denmark, and the �Department of Biomedical Sciences, University of Antwerp, B-2610Antwerp, Belgium
Two new globin proteins have recently been discoveredin vertebrates, neuroglobin in neurons and cytoglobin inall tissues, both showing heme hexacoordination by thedistal His(E7) in the absence of gaseous ligands. In anal-ogy to hemoglobin and myoglobin, neuroglobin and cyto-globin are supposedly involved in O2 storage and deliv-ery, although their physiological role remains to besolved. Here we report O2 equilibria of recombinant hu-man neuroglobin (NGB) and cytoglobin (CYGB) measuredunder close to physiological conditions and at varyingtemperature and pH ranges. NGB shows both alkalineand acid Bohr effects (pH-dependent O2 affinity) and tem-perature-dependent enthalpy of oxygenation. O2 and CObinding equilibrium studies on neuroglobin mutantsstrongly suggest that the bound O2 is stabilized by inter-actions with His(E7) and that this residue functions as amajor Bohr group in the presence of Lys(E10). As shownby the titration of free thiols with 4,4�-dithiodipyridineand by mass spectrometry, this mechanism of modulatingO2 affinity is independent of formation of an internal di-sulfide bond under the experimental conditions used,which stabilize thiols in the reduced form. In CYGB, O2binding is cooperative, consistent with its proposed di-meric structure. Similar to myoglobin but in contrast toNGB, O2 binding to CYGB is pH-independent and exother-mic throughout the temperature range investigated. Ourdata support the hypothesis that CYGB may be involvedin O2-requiring metabolic processes. In contrast, thelower O2 affinity in NGB does not appear compatible witha physiological role involving mitochondrial O2 supply atthe low O2 tensions found within neurons.
Neuroglobin (Ngb)1 and cytoglobin (Cygb) are recently dis-covered members of the vertebrate globin family, which in-
cludes the probably most extensively studied proteins, hemo-globin (Hb), and myoglobin (Mb). Both are expressed at lowlevels (�M range), Ngb in various regions of the brain and in theretinal cells (1–4) and Cygb in all of the tissues so far investi-gated (5, 6). An important characteristic is their heme hexaco-ordination in the absence of exogenous ligands where the distalhistidine, His(E7), occupies the sixth coordination position ofthe iron atom in both the ferrous and the ferric valency states(7–12). Before the discovery of Ngb and Cygb, globin hemehexacoordination was regarded as a feature of certain planthemoglobins and invertebrate or bacterial globins but not ofvertebrate globins. In contrast to Ngb and Cygb, Hb and Mb arepresent at high levels (mM range) in the red blood cells and inthe striated muscle of vertebrates and show pentacoordinateheme geometry when deoxygenated.
Despite their low degree of sequence similarity with othervertebrate globins, human NGB and CYGB show the samecharacteristic �-helix globin fold as vertebrate Hb and Mb (7,10). A phylogenetic tree for human globin proteins indicatesthat NGB diverged early in evolution and may represent themost ancestral human globin protein, whereas CYGB ap-pears to be more closely related to Mb (13). Despite theirancient origin, neither protein has changed significantly dur-ing evolution (13), which may reflect conservation of a criticalbiological function.
Although monomeric, Ngb is capable of conformationalchanges that affect ligand affinity. A number of studies havereported the existence of at least two identifiable conformationson the basis of heterogeneity of reaction rates (8, 14) andinfrared (15), NMR (16), and resonance Raman spectra (9) thatare indicative of an open and closed conformation of the hemepocket. Two distinguishable conformations resulting in a dif-ferent heme pocket accessibility have been identified in aCYGB mutant by x-ray crystallography (10). Also, the forma-tion of an internal disulfide bond increases the rate of dissoci-ation of the distal His from the heme and thereby O2 affinity inNGB and to a lesser extent in CYGB (17).
Although our knowledge of the structural properties of Ngband Cygb is rapidly increasing, the physiological role of Ngband Cygb is still largely unknown. A major hint to the role ofNgb came from the work of Sun et al. (18, 19) who found thatNgb may protect neurons during episodes of O2 lack. Thisimportant finding has been often referred to in the context ofthe general view that Ngb is involved in O2 binding and deliv-ery to sustain mitochondrial respiration similar to Hb and Mb.Remarkably, the function of certain nerve globins from inver-
* This work was supported by the Danish Natural Science ResearchCouncil and the EU (Grant QLG3-CT-2002-01548). The costs of publi-cation of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
§ Both authors contributed equally to this work.¶ To whom correspondence should be addressed: Dept. of Zoophysi-
ology, Institute of Biological Sciences, C. F. Møllers Alle 131, AarhusUniversity, DK-8000 Aarhus C, Denmark. Tel.: 45-89422591; Fax: 45-86194186; E-mail: [email protected].
** Postdoctoral fellow of the Fund for Scientific Research (FlandersUniversity).
1 The abbreviations used are: Ngb, neuroglobin; NGB, human neuro-globin; Cygb, cytoglobin; CYGB, human cytoglobin; FPLC, fast proteinliquid chromatography; Mb, myoglobin; Hb, hemoglobin; HbA0, FPLC-purified adult human hemoglobin; 4-PDS, 4,4�-dithiodipyridine;H(E7)V, His(E7)3Val mouse Ngb mutant; H(E7)Q, His(E7)3Gln hu-man NGB mutant; K(E10)L, Lys(E10)3Leu human NGB mutant;H(E7)L/K(E10)L, double mutant His(E7)3Leu, Lys(E10)3Leu of hu-
man NGB; GSS, triple mutant Cys(CD7)3Gly, Cys(D5)3Ser,Cys(G19)3Ser of human NGB; DTT, 1,4-dithioerythritol; MALDI-TOF,matrix-assisted laser desorption ionization time-of-flight.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 43, Issue of October 22, pp. 44417–44426, 2004© 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 44417
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from

tebrates, whose sequence similarity with mammalian Ngbs isrelatively high (1), is to supply O2 to the nerves (20). Anotherpiece of evidence is that Ngb appears located in the vicinity ofmitochondria as clearly shown in histochemical studies per-formed on retina sections (2). In contrast, Cygb has recentlybeen proposed to be involved in O2-requiring steps in the syn-thesis of collagen based on its expression pattern in fibroticcells (21). Still, practically no direct measurements have beenundertaken yet on the O2 affinity of either Ngb or Cygb tovalidate these hypotheses, as the frequently reported P50 valueof 1 torr for NGB has been mainly derived on the basis ofkinetic rate constants (8). Moreover, discrepancies on thisvalue exist because another kinetic study reported a �1000-fold higher O2 affinity of NGB (22). For these reasons, we haveused equilibrium methods to directly measure O2 bindingcurves for NGB and CYGB, which are reported in this study.
Alternative hypotheses on the functional role of Ngb exist. Inaccordance with its protective effect during hypoxia (18, 19), wehave recently shown (14) that NGB in its Fe(II)NO (nitricoxide) form is an efficient scavenger of peroxynitrite, which isgenerated at high rate following brain ischemia (23). Moreover,in contrast to Hb and Mb, NGB apparently does not generatethe cytotoxic ferryl Fe(IV) form of the protein in the reactionwith peroxides (14). However, these observations do not pre-clude a possible role of NGB in O2 metabolism, given that Hband Mb may also scavenge peroxynitrite and other reactive O2
species (24).To assess whether NGB and CYGB may contribute to phys-
iological functions related to O2 transport and storage, we havestudied their O2 binding equilibria over a range of pH valuesand temperatures and correlated these functional properties tothe protein molecular structure and to existing data on O2
levels present in intact cells.
EXPERIMENTAL PROCEDURES
Reagents—The reagent 4,4�-dithiodipyridine (4-PDS) was from Al-drich (AldrithiolTM-4). All of the other reagents were from Sigma and ofhighest purity available. Water was of Milli Q grade.
Protein Expression and Purification—Recombinant wild-type NGBand CYGB and mouse Ngb were expressed in Escherichia coli andpurified as described (8, 10). The mutants of human NGB and mouseNgb were constructed by site-directed mutagenesis using theQuikChange protocol (Stratagene). The mutants of human NGB in-cluded H(E7)Q with Gln replacing His(E7), K(E10)L with Leu replacingLys(10), the double mutant H(E7)L/K(E10)L with Leu replacing bothHis(E7) and Lys(E10), and the triple mutant here for simplicity calledGSS where all of the cysteine residues in positions CD7, D5, and G19are replaced by Gly, Ser, and Ser, respectively. The mouse Ngb mutantH(E7)V with Val replacing His(E7) was also used. The purity of sampleswas routinely checked using a PhastSystem (Amersham Biosciences) byisoelectrofocusing on polyacrylamide gels at the pH 3–9 range and bySDS 15% polyacrylamide gels. Whenever necessary, contaminants wereremoved by FPLC anion-exchange chromatography using a Hi-Trap QFF 5-ml column (Amersham Biosciences) by a linear gradient of 0.2 M
NaCl in 0.02 M Hepes, pH 7.0, containing 0.5 mM EDTA. The divalentmetal chelator EDTA was routinely added to all buffers to a finalconcentration of 0.5 mM. Human HbA0 was obtained by FPLC anion-exchange chromatography on a HiLoad 26/10 Q-Sepharose column (Am-ersham Biosciences) using a linear gradient of 0.15 M NaCl in 0.05 M
Tris buffer, pH 8.3, containing 0.5 mM EDTA. Aliquots were stored inliquid N2. Purified NGB was stored in liquid N2 as either a Met or COderivative (see below) in small aliquots (�1 mM heme).
Determination of Extinction Coefficients—Absorption molar extinc-tion coefficients of Met NGB were determined after complete oxidationof the protein by potassium ferricyanide and elimination of excessferricyanide by gel filtration (25). The millimolar heme concentration ofMet NGB was obtained after adding solid potassium cyanide in molarexcess over heme and using the extinction coefficient of 11 mM�1 cm�1
at 540 nm for Mb (25). Extinction coefficients of deoxy and CO forms ofNGB were obtained under anaerobic conditions by adding sodium di-thionite (1 mg/ml) and CO gas in molar excess to a solution of Met NGBof a known concentration. The spectra of pure oxy NGB were measured
after exposing to air the samples previously reduced to the Fe(II) formin anaerobic vials by the enzymatic reducing system described by Ha-yashi et al. (26). The molar extinction coefficients of oxy NGB were thencalculated from the known concentration of the Met NGB sample used.
Protein Reduction—To reduce Fe(III) to Fe(II) and possible S–Sbonds before O2 binding equilibrium experiments, samples (�1 mM
heme) were dialyzed against 1–2 mg/ml sodium dithionite and 1–2mg/ml DTT dissolved in ice-cold 50 mM Bistris buffer, pH 7.6, contain-ing 0.5 mM EDTA under strictly anaerobic conditions. The buffer wasextensively bubbled with N2 and CO before the addition of solid dithio-nite and DTT. After a 1-h reduction, three exchanges of degassedCO-equilibrated buffer were made to eliminate dithionite and DTT. Allsteps were made on ice and in the dark. The reduced protein then waskept on ice in closed anaerobic vials containing CO until use for O2
equilibrium experiments (within the same day) or stored in liquidnitrogen in CO-flushed cryovials. Reduction under the air of Met Ngb bythe Met enzymatic reducing system (26) as previously reported (8) wasnot successful in our hands.
Oxygen Binding Equilibria—O2 equilibrium curves were measuredon ultrathin (�0.05 mm) layers of 4-�l protein samples using a modifiedgas diffusion chamber fed by cascaded W�sthoff gas-mixing pumps,which generate precise mixtures of O2 or air and ultrapure (�99.998%)N2 (27, 28). The absorbance change following the stepwise increases/decreases in O2 tension was continuously monitored at a fixed wave-length in the Soret region, typically 428 nm for Ngb and 426 nm forCYGB. Equilibrium at each oxygenation step was achieved within 2–4min, consistently with the rapid (millisecond to second) kinetics ofligand binding (17). In some experiments, the absorbance spectra (400–600 nm) were measured using a UV-visible Cary 50 Probe spectropho-tometer equipped with optic fibers to verify the reversible change in thespectrum from the deoxygenated to oxygenated form. A BMS 2 MK 2thermostated microelectrode (Radiometer) was used to measure the pHvalue in 100-�l subsamples. Samples were typically in 0.1 M Bistris orHepes buffers containing 0.5 mM EDTA at a protein concentration of0.03–0.05 mM heme. Identical binding curves were obtained in eitherBistris or Hepes buffers. The effect of protein concentration on the O2
affinity was checked at pH 7.55 at 0.036 and 0.09 mM heme, which isclose to the highest local concentration estimated for Ngb in retinal cells(�0.1 mM) (2). To avoid buildup of oxidized heme during O2 equilibriumexperiments, samples contained the Met-reducing enzymatic system(26) in the following amounts: glucose 6-phosphate (15 mM); glucose6-phosphate dehydrogenase (0.0073 mg/ml); NADP (1 mM); ferredoxin-NADP reductase (0.0017 mM); ferredoxin (0.0038 mM); and catalase(0.0015 mM). Rapid (2–3 saturation points) control experiments per-formed in the absence of reducing system using the H(E7)V and H(E7)Qmutants confirmed that O2 affinity was not altered by the cofactors andenzymes added. O2 binding equilibria were also measured in bufferedsuspensions of intact E. coli cells expressing NGB. In these experi-ments, the reducing enzymatic system was omitted.
Values for O2 affinity (P50, O2 tension at half-saturation) and coop-erativity (n50) were interpolated from the zero-intercept and the slope,respectively, of Hill plots, log[Y/(1 � Y)] versus logPO2, where Y is theO2 saturation.
Autoxidation Rates—Oxygenated samples were prepared by the re-duction of the Met form with sodium dithionite followed by immediatepassage through a small (5 ml) PD-10 gel filtration column (AmershamBiosciences) kept at 4 °C in order to minimize heme oxidation duringdesalting (�2–3 min). Rates of spontaneous oxidation were measured inair at 25 °C from the decrease in absorbance at 576 nm over time usinga HP 8543 UV-visible diode array spectrophotometer. Absorbancetraces were fitted to monoexponential decays. Protein concentrationwas within the range of 0.006–0.010 mM. The buffer used was 0.05 M
Hepes, 0.5 mM EDTA.CO Saturation Experiments—The effect of pH on CO saturation levels
in NGB was investigated at 10 and 30 °C in anaerobic cells at a proteinconcentration of 1.5 �M heme by adding 2 �l of CO-saturated water (1 mM,Ref. 25) in 0.1 M Bistris buffers containing 0.5 mM EDTA in the presenceof 2 mg/ml sodium dithionite and 1 mg/ml DTT. CO saturation wascalculated by multi-component spectral analysis on a HP 8543 UV-visiblediode array spectrophotometer using reference spectra for pure deoxy andCO-NGB obtained as described above.
Sulfhydryl Reactivity—The reaction of 4-PDS with the thiol groups ofglobin proteins to form 4-thiopyridone (29, 30) was employed to meas-ure free cysteines in NGB and CYGB samples at pH 7.0. To verify thepropensity to form disulfide bridges under various conditions, sampleswere incubated for 3 h at 20 °C either in 0.1 M Bistris buffer containing0.5 mM EDTA in the presence and absence of the enzymatic Met-reducing system or in 0.1 M phosphate buffer with and without EDTA
O2 Equilibria in Neuroglobin and Cytoglobin44418
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from

added. We used a 4-molar excess of 4-PDS over heme at a proteinconcentration of 0.010 mM heme. The increase in absorbance with timewas monitored in quartz cuvettes at 324 nm corresponding to themaximum absorbance for 4-thiopyridone (extinction coefficient 19.8cm�1 mM�1) using as reference a protein sample at identical hemeconcentration not containing 4-PDS.
Reaction with 4-PDS was also employed to create the internal S–Sbond between Cys(CD7) and Cys(D5) in NGB. In the reaction, deriva-tization of Cys(G19) with 4-PDS also occurs. According to Grassetti andMurray (29), free thiols are quantitatively oxidized to the S–S formwhen reacting with dithiopyridine, provided that they are located suf-ficiently close to each other. After reaction with 4-PDS, the protein wasdialyzed to remove excess reagent and used in equilibrium determina-tion of O2 binding.
Mass Spectrometry—NGB dissolved in 5 mM Tris-HCl, pH 8.5, andnot previously exposed to reducing agents was analyzed by mass spec-troscopy to detect the formation of internal disulfide bridge. Samplepreparation was similar to that reported by Hamdane et al. (17). Thenative sample at 5 mM concentration was dissolved in 50:50 methanol:water in 5% formic acid and injected into home-made nanoelectrosprayneedles. Nanoelectrospray mass spectrometry spectra were acquired ona Q-TOFII (Micromass, Wythwnshave, United Kingdom) over the mass-to-charge ratio (m/z) range of 700–2000. The multiple-charged electro-spray spectra were deconvoluted by the MaxEnt1 software to producethe molecular mass of the protein. Before each measurement, the massscale was calibrated using myoglobin. NGB was alkylated by addingdifferent concentrations of iodoacetamide in 20 mM ammonium bicar-bonate and incubated for 20 min at room temperature. The sample wasdried in a 10-kDa cut-off centrifugal filter device (Centricon YM10,Millipore, Bedford, MA). The retained proteins were then dissolved in50:50 methanol:water in 5% formic acid, and the mass was measured asdescribe above.
NGB was digested with trypsin overnight. The digested mixture wasanalyzed by Applied Biosystems 4700 MALDI-TOF/TOF. For sampleloading, 5 mg ml�1 �-cyano-4-hydroxycinnamic acid (Sigma) was dis-solved in acetonitrile, 0.1% trifluoroacetic acid (70:30, v/v). A thin layerof homogeneous matrix crystals was prepared on the target by adding0.5 ml of �-cyano-4-hydroxycinnamic acid and allowed to dry. 0.5 ml of0.1% trifluoroacetic acid was placed on the top of the matrix layerfollowed by 1 �l of trypsin-digested solution (31).
RESULTS
The molar extinction coefficients calculated for NGB in theMet, oxygenated, deoxygenated, and CO forms and used in thisstudy are shown in Table I.
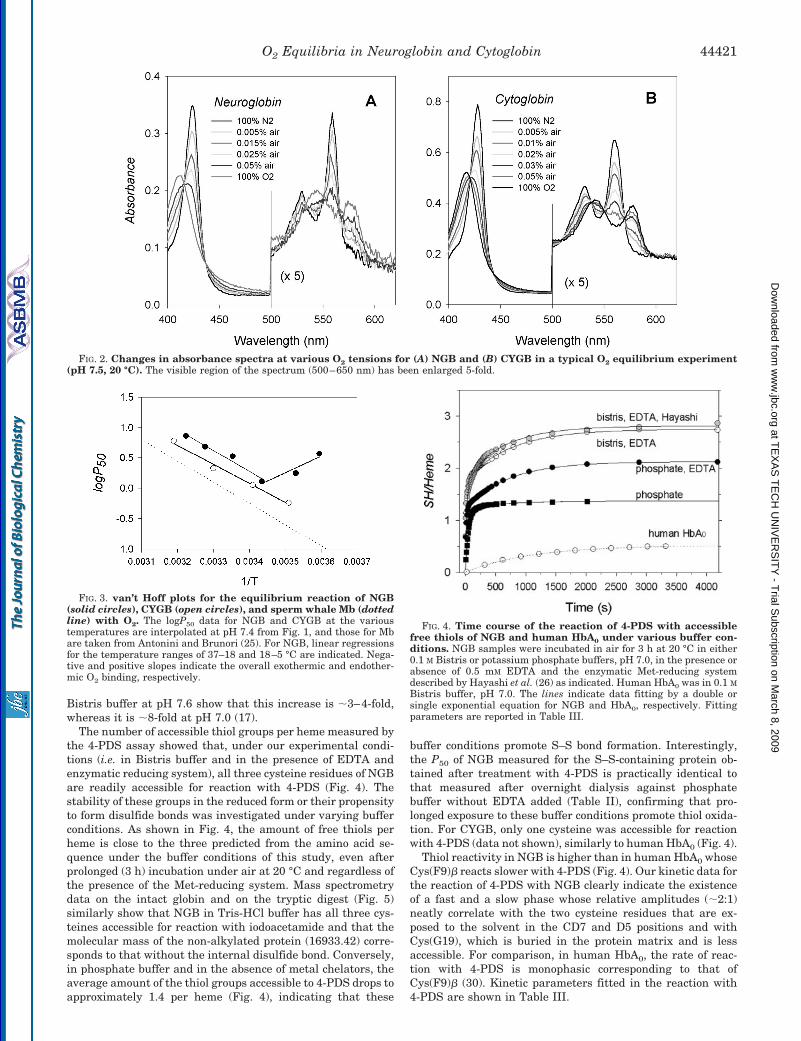
O2 affinity and cooperativity for NGB and CYGB as a func-tion of pH and at various temperatures are reported in Fig. 1,A, B, and D, with selected P50 values reported in Table II.Other samples studied were the human NGB mutants H(E7)Q,K(E10)L, H(E7)L/K(E10)L, and GSS and mouse Ngb wild-typeand mutant H(E7)V as well as NGB expressed in intact E. colicells (Fig. 1C). Hill coefficients for NGB are close to unity, asexpected for a monomeric protein (Fig. 1, A–C). In CYGB, Hillcoefficients vary with temperature and are in the range of0.63–1.63 (Fig. 1D), indicating the existence of both positiveand negative heme-heme interactions. Reversible changes fromdeoxy to oxy absorbance spectra of NGB and CYGB are shownin Fig. 2 for a typical O2 equilibrium experiment. For bothproteins, the presence of a single set of isosbestic points and ofthe characteristic oxy spectrum with peaks at 543 and 576 nm
(Fig. 2) shows that the protein remains in the ferrous stateduring measurements of O2-binding equilibria. The recovery ofthe fully deoxygenated spectrum or absorbance value aftercomplete oxygenation at the end of each experiment (data notshown) confirmed the absence of a significant amount of ferricprotein.
A major finding of this study is that NGB displays bothalkaline and acid Bohr effects (increases and decreases in P50
with a pH decrease, respectively), depending on the tempera-ture and on the pH range (Fig. 1, A and B). In contrast, the O2
affinity of CYGB is virtually independent of changes in pH anddecreases linearly when temperature increases (Fig. 1D). Accord-ingly, the van’t Hoff plot calculated at pH 7.4 shows an unusualbiphasic behavior for NGB and a linear one for CYGB (Fig. 3),showing temperature-dependent or temperature-independentoverall enthalpies of oxygenation (�H, slope of the plot � 2.303R,where R is the gas constant), respectively. Specifically, at pH 7.4and in the range of 5–37 °C, O2 binding in NGB is exothermic attemperatures above �18 °C (�H � �15.7 kcal mol�1) and endo-thermic at temperatures below �18 °C (�H � 12.7 kcal mol�1).In contrast, in CYGB as in Mb or human HbA, the overall heatliberated upon O2 binding (�H � �14.3 kcal mol�1) does notdepend on the temperature of the reaction within the tempera-ture range investigated and is very similar to that of sperm whaleMb (�H � �14.9 kcal mol�1) (Fig. 3).
The Bohr effect found in purified recombinant NGB (Fig. 1, Aand B) is also present in intact E. coli cells overexpressing NGBand in purified recombinant mouse Ngb (Fig. 1C), indicatingthat this is a distinct property of mammalian Ngbs within cellsor in vitro buffer solutions. Although bacterial cells do notnaturally express NGB, they may provide a reducing environ-ment similar to that of neurons and retinal cells of vertebratesexpressing Ngb and to that provided by the Met-reducing sys-tem present in our in vitro experiments. Attempts to measureO2 binding equilibria in situ using rabbit and mouse retinasections in which the local concentration of Ngb should be closeto 0.1 mM (2) were unsuccessful, suggesting that in vivo Ngbconcentration may in fact be lower than that estimated fromimmunohistochemical staining. An increase in protein concen-tration from 0.036 to 0.090 mM did not alter O2 affinity (P50 �2.9 mm Hg, pH 7.55 0.01, 25 °C), suggesting that O2-linkedassociation, as known from hagfish (32) and lamprey Hbs (33)and several invertebrate Hbs, does not occur in NGB.
The stereochemical origin of the Bohr effect has been inves-tigated using the mutants of either human NGB or mouse Ngbwhere the distal His(E7) or Lys(E10) has been replaced byneutral residues. Previous studies have indicated His(E7) to becrucial in the control of the affinity for external ligands (8, 17)and that protonation at these residues alters heme reactivity(34). As shown in Fig. 1C, the His(E7) mutants, H(E7)V andH(E7)Q, show a markedly increased O2 affinity (low P50 values)and a loss of pH sensitivity compared with the wild-type pro-tein. This finding indicates the essential role of heme hexaco-ordination by His(E7) in the control of ligand affinity and of itsdependence on pH. In the K(E10)L mutant, the Bohr effect inthe pH range of 6.5–7.5 is reversed compared with that in thewild-type protein so that O2 affinity increases with a pH de-crease. It was not possible to measure O2 equilibria for thedouble mutant H(E7)L/K(E10)L of human NGB because of itsextremely high O2 affinity.
We also investigated the functional role of cysteine residuesin position CD7, D5, and G19 by using the triple mutant GSSof human NGB in which all three cysteines are replaced. Thismutant protein is the same as that characterized by x-raycrystallography (7). As shown in Fig. 1C, the Bohr effect curveof the GSS mutant is superimposable to that of the wild-type
TABLE IAbsorbance maxima (�, nm) and corresponding extinction
coefficients (�, mM�1 cm�1) for NGB
Derivative � �
nm mM�1 cm�1
Met 532 10.7Oxy 543 14.7
576 11.6Deoxy 528 16.2
558 28.9CO 538 11.5
561 11.2
O2 Equilibria in Neuroglobin and Cytoglobin 44419
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from
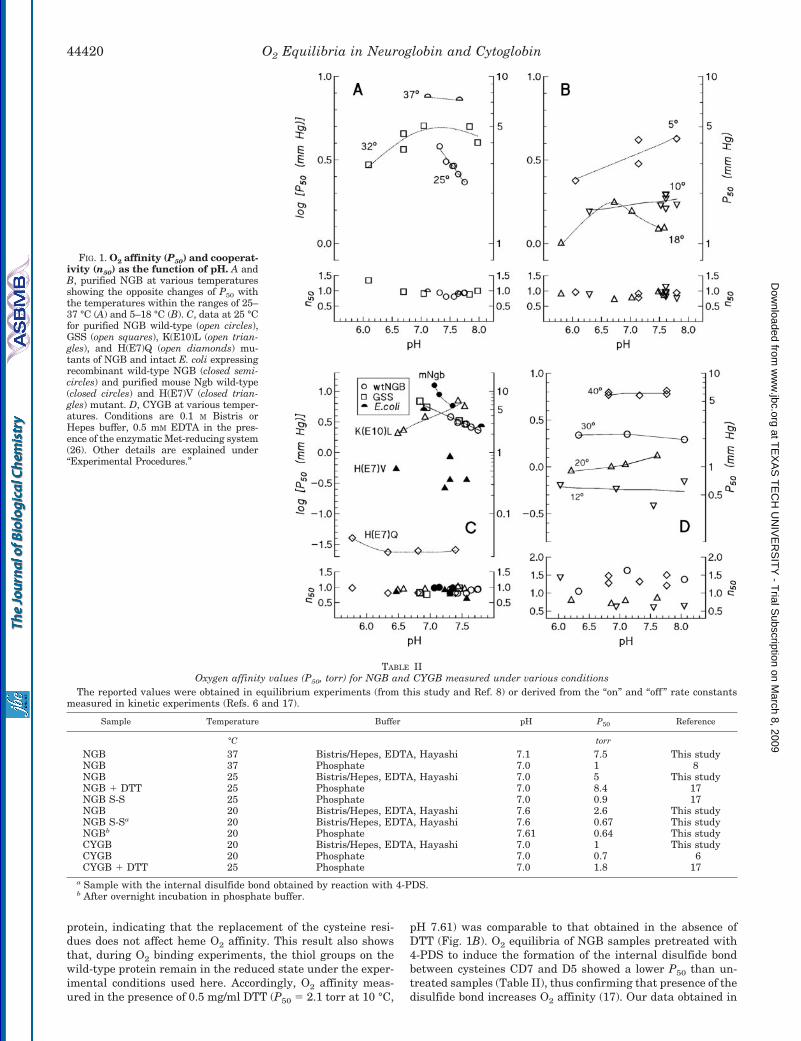
protein, indicating that the replacement of the cysteine resi-dues does not affect heme O2 affinity. This result also showsthat, during O2 binding experiments, the thiol groups on thewild-type protein remain in the reduced state under the exper-imental conditions used here. Accordingly, O2 affinity meas-ured in the presence of 0.5 mg/ml DTT (P50 � 2.1 torr at 10 °C,
pH 7.61) was comparable to that obtained in the absence ofDTT (Fig. 1B). O2 equilibria of NGB samples pretreated with4-PDS to induce the formation of the internal disulfide bondbetween cysteines CD7 and D5 showed a lower P50 than un-treated samples (Table II), thus confirming that presence of thedisulfide bond increases O2 affinity (17). Our data obtained in
FIG. 1. O2 affinity (P50) and cooperat-ivity (n50) as the function of pH. A andB, purified NGB at various temperaturesshowing the opposite changes of P50 withthe temperatures within the ranges of 25–37 °C (A) and 5–18 °C (B). C, data at 25 °Cfor purified NGB wild-type (open circles),GSS (open squares), K(E10)L (open trian-gles), and H(E7)Q (open diamonds) mu-tants of NGB and intact E. coli expressingrecombinant wild-type NGB (closed semi-circles) and purified mouse Ngb wild-type(closed circles) and H(E7)V (closed trian-gles) mutant. D, CYGB at various temper-atures. Conditions are 0.1 M Bistris orHepes buffer, 0.5 mM EDTA in the pres-ence of the enzymatic Met-reducing system(26). Other details are explained under“Experimental Procedures.”
TABLE IIOxygen affinity values (P50, torr) for NGB and CYGB measured under various conditions
The reported values were obtained in equilibrium experiments (from this study and Ref. 8) or derived from the “on” and “off ” rate constantsmeasured in kinetic experiments (Refs. 6 and 17).
Sample Temperature Buffer pH P50 Reference
°C torr
NGB 37 Bistris/Hepes, EDTA, Hayashi 7.1 7.5 This studyNGB 37 Phosphate 7.0 1 8NGB 25 Bistris/Hepes, EDTA, Hayashi 7.0 5 This studyNGB DTT 25 Phosphate 7.0 8.4 17NGB S-S 25 Phosphate 7.0 0.9 17NGB 20 Bistris/Hepes, EDTA, Hayashi 7.6 2.6 This studyNGB S-Sa 20 Bistris/Hepes, EDTA, Hayashi 7.6 0.67 This studyNGBb 20 Phosphate 7.61 0.64 This studyCYGB 20 Bistris/Hepes, EDTA, Hayashi 7.0 1 This studyCYGB 20 Phosphate 7.0 0.7 6CYGB DTT 25 Phosphate 7.0 1.8 17
a Sample with the internal disulfide bond obtained by reaction with 4-PDS.b After overnight incubation in phosphate buffer.
O2 Equilibria in Neuroglobin and Cytoglobin44420
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from
Bistris buffer at pH 7.6 show that this increase is �3–4-fold,whereas it is �8-fold at pH 7.0 (17).
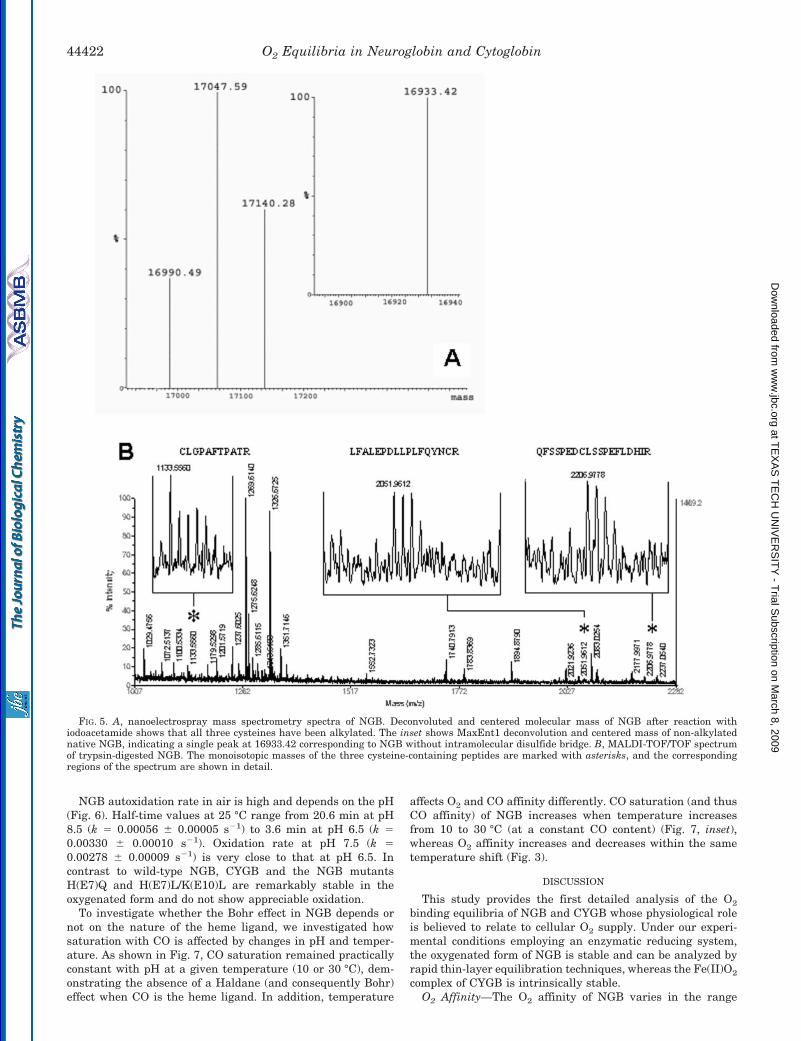
The number of accessible thiol groups per heme measured bythe 4-PDS assay showed that, under our experimental condi-tions (i.e. in Bistris buffer and in the presence of EDTA andenzymatic reducing system), all three cysteine residues of NGBare readily accessible for reaction with 4-PDS (Fig. 4). Thestability of these groups in the reduced form or their propensityto form disulfide bonds was investigated under varying bufferconditions. As shown in Fig. 4, the amount of free thiols perheme is close to the three predicted from the amino acid se-quence under the buffer conditions of this study, even afterprolonged (3 h) incubation under air at 20 °C and regardless ofthe presence of the Met-reducing system. Mass spectrometrydata on the intact globin and on the tryptic digest (Fig. 5)similarly show that NGB in Tris-HCl buffer has all three cys-teines accessible for reaction with iodoacetamide and that themolecular mass of the non-alkylated protein (16933.42) corre-sponds to that without the internal disulfide bond. Conversely,in phosphate buffer and in the absence of metal chelators, theaverage amount of the thiol groups accessible to 4-PDS drops toapproximately 1.4 per heme (Fig. 4), indicating that these
buffer conditions promote S–S bond formation. Interestingly,the P50 of NGB measured for the S–S-containing protein ob-tained after treatment with 4-PDS is practically identical tothat measured after overnight dialysis against phosphatebuffer without EDTA added (Table II), confirming that pro-longed exposure to these buffer conditions promote thiol oxida-tion. For CYGB, only one cysteine was accessible for reactionwith 4-PDS (data not shown), similarly to human HbA0 (Fig. 4).
Thiol reactivity in NGB is higher than in human HbA0 whoseCys(F9)� reacts slower with 4-PDS (Fig. 4). Our kinetic data forthe reaction of 4-PDS with NGB clearly indicate the existenceof a fast and a slow phase whose relative amplitudes (�2:1)neatly correlate with the two cysteine residues that are ex-posed to the solvent in the CD7 and D5 positions and withCys(G19), which is buried in the protein matrix and is lessaccessible. For comparison, in human HbA0, the rate of reac-tion with 4-PDS is monophasic corresponding to that ofCys(F9)� (30). Kinetic parameters fitted in the reaction with4-PDS are shown in Table III.
FIG. 2. Changes in absorbance spectra at various O2 tensions for (A) NGB and (B) CYGB in a typical O2 equilibrium experiment(pH 7.5, 20 °C). The visible region of the spectrum (500–650 nm) has been enlarged 5-fold.
FIG. 3. van’t Hoff plots for the equilibrium reaction of NGB(solid circles), CYGB (open circles), and sperm whale Mb (dottedline) with O2. The logP50 data for NGB and CYGB at the varioustemperatures are interpolated at pH 7.4 from Fig. 1, and those for Mbare taken from Antonini and Brunori (25). For NGB, linear regressionsfor the temperature ranges of 37–18 and 18–5 °C are indicated. Nega-tive and positive slopes indicate the overall exothermic and endother-mic O2 binding, respectively.
FIG. 4. Time course of the reaction of 4-PDS with accessiblefree thiols of NGB and human HbA0 under various buffer con-ditions. NGB samples were incubated in air for 3 h at 20 °C in either0.1 M Bistris or potassium phosphate buffers, pH 7.0, in the presence orabsence of 0.5 mM EDTA and the enzymatic Met-reducing systemdescribed by Hayashi et al. (26) as indicated. Human HbA0 was in 0.1 M
Bistris buffer, pH 7.0. The lines indicate data fitting by a double orsingle exponential equation for NGB and HbA0, respectively. Fittingparameters are reported in Table III.
O2 Equilibria in Neuroglobin and Cytoglobin 44421
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from
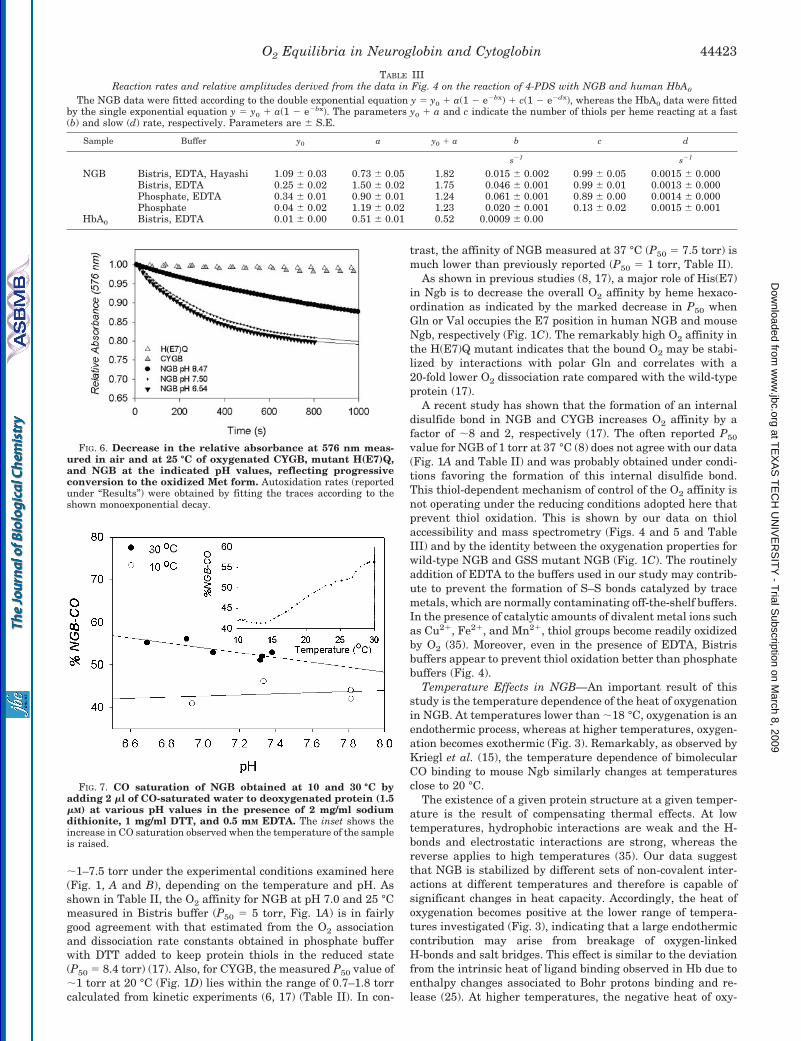
NGB autoxidation rate in air is high and depends on the pH(Fig. 6). Half-time values at 25 °C range from 20.6 min at pH8.5 (k � 0.00056 0.00005 s�1) to 3.6 min at pH 6.5 (k �0.00330 0.00010 s�1). Oxidation rate at pH 7.5 (k �0.00278 0.00009 s�1) is very close to that at pH 6.5. Incontrast to wild-type NGB, CYGB and the NGB mutantsH(E7)Q and H(E7)L/K(E10)L are remarkably stable in theoxygenated form and do not show appreciable oxidation.
To investigate whether the Bohr effect in NGB depends ornot on the nature of the heme ligand, we investigated howsaturation with CO is affected by changes in pH and temper-ature. As shown in Fig. 7, CO saturation remained practicallyconstant with pH at a given temperature (10 or 30 °C), dem-onstrating the absence of a Haldane (and consequently Bohr)effect when CO is the heme ligand. In addition, temperature
affects O2 and CO affinity differently. CO saturation (and thusCO affinity) of NGB increases when temperature increasesfrom 10 to 30 °C (at a constant CO content) (Fig. 7, inset),whereas O2 affinity increases and decreases within the sametemperature shift (Fig. 3).
DISCUSSION
This study provides the first detailed analysis of the O2
binding equilibria of NGB and CYGB whose physiological roleis believed to relate to cellular O2 supply. Under our experi-mental conditions employing an enzymatic reducing system,the oxygenated form of NGB is stable and can be analyzed byrapid thin-layer equilibration techniques, whereas the Fe(II)O2
complex of CYGB is intrinsically stable.O2 Affinity—The O2 affinity of NGB varies in the range
FIG. 5. A, nanoelectrospray mass spectrometry spectra of NGB. Deconvoluted and centered molecular mass of NGB after reaction withiodoacetamide shows that all three cysteines have been alkylated. The inset shows MaxEnt1 deconvolution and centered mass of non-alkylatednative NGB, indicating a single peak at 16933.42 corresponding to NGB without intramolecular disulfide bridge. B, MALDI-TOF/TOF spectrumof trypsin-digested NGB. The monoisotopic masses of the three cysteine-containing peptides are marked with asterisks, and the correspondingregions of the spectrum are shown in detail.
O2 Equilibria in Neuroglobin and Cytoglobin44422
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from
�1–7.5 torr under the experimental conditions examined here(Fig. 1, A and B), depending on the temperature and pH. Asshown in Table II, the O2 affinity for NGB at pH 7.0 and 25 °Cmeasured in Bistris buffer (P50 � 5 torr, Fig. 1A) is in fairlygood agreement with that estimated from the O2 associationand dissociation rate constants obtained in phosphate bufferwith DTT added to keep protein thiols in the reduced state(P50 � 8.4 torr) (17). Also, for CYGB, the measured P50 value of�1 torr at 20 °C (Fig. 1D) lies within the range of 0.7–1.8 torrcalculated from kinetic experiments (6, 17) (Table II). In con-
trast, the affinity of NGB measured at 37 °C (P50 � 7.5 torr) ismuch lower than previously reported (P50 � 1 torr, Table II).
As shown in previous studies (8, 17), a major role of His(E7)in Ngb is to decrease the overall O2 affinity by heme hexaco-ordination as indicated by the marked decrease in P50 whenGln or Val occupies the E7 position in human NGB and mouseNgb, respectively (Fig. 1C). The remarkably high O2 affinity inthe H(E7)Q mutant indicates that the bound O2 may be stabi-lized by interactions with polar Gln and correlates with a20-fold lower O2 dissociation rate compared with the wild-typeprotein (17).
A recent study has shown that the formation of an internaldisulfide bond in NGB and CYGB increases O2 affinity by afactor of �8 and 2, respectively (17). The often reported P50
value for NGB of 1 torr at 37 °C (8) does not agree with our data(Fig. 1A and Table II) and was probably obtained under condi-tions favoring the formation of this internal disulfide bond.This thiol-dependent mechanism of control of the O2 affinity isnot operating under the reducing conditions adopted here thatprevent thiol oxidation. This is shown by our data on thiolaccessibility and mass spectrometry (Figs. 4 and 5 and TableIII) and by the identity between the oxygenation properties forwild-type NGB and GSS mutant NGB (Fig. 1C). The routinelyaddition of EDTA to the buffers used in our study may contrib-ute to prevent the formation of S–S bonds catalyzed by tracemetals, which are normally contaminating off-the-shelf buffers.In the presence of catalytic amounts of divalent metal ions suchas Cu2, Fe2, and Mn2, thiol groups become readily oxidizedby O2 (35). Moreover, even in the presence of EDTA, Bistrisbuffers appear to prevent thiol oxidation better than phosphatebuffers (Fig. 4).
Temperature Effects in NGB—An important result of thisstudy is the temperature dependence of the heat of oxygenationin NGB. At temperatures lower than �18 °C, oxygenation is anendothermic process, whereas at higher temperatures, oxygen-ation becomes exothermic (Fig. 3). Remarkably, as observed byKriegl et al. (15), the temperature dependence of bimolecularCO binding to mouse Ngb similarly changes at temperaturesclose to 20 °C.
The existence of a given protein structure at a given temper-ature is the result of compensating thermal effects. At lowtemperatures, hydrophobic interactions are weak and the H-bonds and electrostatic interactions are strong, whereas thereverse applies to high temperatures (35). Our data suggestthat NGB is stabilized by different sets of non-covalent inter-actions at different temperatures and therefore is capable ofsignificant changes in heat capacity. Accordingly, the heat ofoxygenation becomes positive at the lower range of tempera-tures investigated (Fig. 3), indicating that a large endothermiccontribution may arise from breakage of oxygen-linkedH-bonds and salt bridges. This effect is similar to the deviationfrom the intrinsic heat of ligand binding observed in Hb due toenthalpy changes associated to Bohr protons binding and re-lease (25). At higher temperatures, the negative heat of oxy-
FIG. 6. Decrease in the relative absorbance at 576 nm meas-ured in air and at 25 °C of oxygenated CYGB, mutant H(E7)Q,and NGB at the indicated pH values, reflecting progressiveconversion to the oxidized Met form. Autoxidation rates (reportedunder “Results”) were obtained by fitting the traces according to theshown monoexponential decay.
FIG. 7. CO saturation of NGB obtained at 10 and 30 °C byadding 2 �l of CO-saturated water to deoxygenated protein (1.5�M) at various pH values in the presence of 2 mg/ml sodiumdithionite, 1 mg/ml DTT, and 0.5 mM EDTA. The inset shows theincrease in CO saturation observed when the temperature of the sampleis raised.
TABLE IIIReaction rates and relative amplitudes derived from the data in Fig. 4 on the reaction of 4-PDS with NGB and human HbA0
The NGB data were fitted according to the double exponential equation y � y0 a(1 � e�bx) c(1 � e�dx), whereas the HbA0 data were fittedby the single exponential equation y � y0 a(1 � e�bx). The parameters y0 a and c indicate the number of thiols per heme reacting at a fast(b) and slow (d) rate, respectively. Parameters are S.E.
Sample Buffer y0 a y0 a b c d
s�1 s�1
NGB Bistris, EDTA, Hayashi 1.09 0.03 0.73 0.05 1.82 0.015 0.002 0.99 0.05 0.0015 0.000Bistris, EDTA 0.25 0.02 1.50 0.02 1.75 0.046 0.001 0.99 0.01 0.0013 0.000Phosphate, EDTA 0.34 0.01 0.90 0.01 1.24 0.061 0.001 0.89 0.00 0.0014 0.000Phosphate 0.04 0.02 1.19 0.02 1.23 0.020 0.001 0.13 0.02 0.0015 0.001
HbA0 Bistris, EDTA 0.01 0.00 0.51 0.01 0.52 0.0009 0.00
O2 Equilibria in Neuroglobin and Cytoglobin 44423
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from

genation of NGB (Fig. 3) is consistent with the loss of thesepolar interactions. Under these conditions, the negative �Hvalues found for NGB and CYGB are those typical for verte-brate globins (25). A recent kinetic study has reported lessnegative �H values due to a large positive contribution deriv-ing from heme dissociation of His(E7) upon O2 binding (36).Thermodynamic parameters, such as �H of Hb oxygenation,are highly dependent on the experimental conditions chosenincluding buffers and pH and temperature ranges (37, 38). AtpH 7.0 and at high temperatures (�20 °C), the experimentalconditions used by Uzan et al. (36), O2 affinity of NGB showsminimal temperature dependence showing that endothermicand exothermic reactions under these specific conditions mayoffset each other.
Molecular Basis for the Bohr Effect—The Bohr effect in hu-man HbA derives from a higher proton affinity in the deoxy-genated low affinity T state than in the fully liganded R state.A major Bohr group in human HbA is His(HC3)� whose pKa
rises in the R3T state transition due to the proximity ofAsp(FG1)�. Accordingly, the Bohr effect in human HbA is in-dependent of the nature of the ligand and has the same mag-nitude when the heme ligand is either O2 or CO (25). Analternative molecular mechanism must be operating in NGB,because only the affinity for O2 (Fig. 1, A–C) but not that for CO(Fig. 7) depends on pH at least within the pH range investi-gated here. This observation is consistent with the invarianceof resonance Raman and visible spectra of the CO derivativewithin the same pH range (9, 34, 39). Also, in the monomericglobin of the insect Chironomus thummi thummi, the Bohreffect is larger when O2 rather than CO is the heme ligand (40).These observations support the conclusion that in NGB theheme-bound O2, which in globins, including NGB (9), has aFe(III)O
2
. character (41), favors protonation of nearby distalresidues.
We propose that the Bohr effect in NGB originates fromstabilization of the bound O2 by the distal His(E7) in its un-protonated form and that protonation at this residue favors O2
dissociation. As shown, the Bohr effect is obliterated uponreplacement of His(E7) (Fig. 1C), indicating a key role of thisresidue in the pH-dependent control of O2 affinity. Our datashow that a pH decrease may have opposite effects, namely toeither decrease or increase O2 affinity, whereby both alkalineand acid Bohr effects are observed. When pH decreases, protonbinding at the His(E7) imidazole ring will progressively with-draw electrons from O2 and favor its heme dissociation,whereby a normal (alkaline) Bohr effect is observed. At thesame time, protonation at His(E7) will tend to open the hemepocket to solvate the positively charged imidazole (34). Thiswould then give free access to a high affinity pentacoordinateheme and therefore give rise to a reverse Bohr effect (increasein O2 affinity with a pH fall). As expected, each of these twoopposite effects will dominate depending on the temperatureand the pH range investigated, consistent with our results (Fig.1, A and B). In an elegant series of experiments, Nienhaus et al.(34) have recently shown that proton affinity of His(E7) isaffected by the presence of Lys(E10), which sequesters theheme pocket from the solvent by interacting with a heme pro-pionate chain (7). Replacement of Lys(E10) with a neutralresidue will no longer protect the heme pocket from the solvent,whereby the pH at which the His becomes exposed to thesolvent increases as shown in Fig. 1C and other data (34). Inaddition, electrostatic effects of Lys(E10) free �-amino groupand the paired heme propionate on the pKa of His(E7) may alsoplay an important role in modulating the Bohr effect of NGB.This model for the Bohr effect is supported by our data on thetemperature dependence of the O2 affinity. An increase in
temperature would render polar interactions, such as those in-volving His(E7) and Lys(E10), progressively less stable, and ac-cordingly, the enthalpy of oxygenation becomes negative (Fig. 3).Conversely, the endothermic oxygenation observed at the lowtemperature range investigated indicates that O2 binding is as-sociated with rupture, which requires heat, of polar interactions.
These conclusions nicely correlate with resonance Ramanspectroscopy of the CO derivative of mouse Ngb clearly showingclosed and open heme pocket conformations in which the ligandmay interact or not with a polar residue within the hemepocket (9). This model for the Bohr effect in NGB in whichheme-bound O2 directly interacts with a Bohr group predictsthat a decrease in pH should favor O2 dissociation as thesuperoxide ion and thus the autoxidation rate. However, be-tween pH 6.5 and 7.5, the autoxidation rate remains unaffected(Fig. 6), whereas O2 affinity changes (Fig. 1, A and B), suggest-ing a different control mechanism. Investigations on the struc-ture of the oxygenated form of Ngb have not been carried outyet because of its intrinsic instability and will represent animportant test to the hypothesis presented here.
The Bohr effect is not observed in CYGB, indicative of adifferent heme environment. In particular, the distal pocketin CYGB is more apolar than in NGB (10, 42), which resultsin a positive redox potential value compared with the nega-tive one seen in NGB (42) and in a higher stability of the oxyform toward autoxidation (Fig. 6) (42). Spectroscopic studieshave shown that, as in NGB, the heme-bound CO and possi-bly O2 may interact with the distal His(E7) of CYGB whoseheme pocket also adopts closed and open conformations (42).The recently solved crystal structure of CYGB (10, 12, 43) hasshown that Arg(E10) makes a salt bridge with one of theheme propionates, suggesting a role similar to that ofLys(E10) in NGB. The presence of Arg with a higher pKa thanLys in position E10 combined with a more hydrophobic envi-ronment of the distal heme pocket in CYGB probably de-creases the pKa of the distal His(E7), which may explain whythe O2 affinity does not change with pH in the range hereinvestigated (6.0–8.0).
Cooperative O2 Binding in CYGB—CYGB displays both pos-itive (cooperative) and negative (inhibitory) heme-heme inter-actions (Fig. 1D), indicating that ligation at one heme caneither increase or decrease the affinity at the other heme group.This finding is consistent with its dimeric structure proposedon the basis of x-ray crystallographic studies (10). Cooperativ-ity within a dimeric globin has been previously described in thehemoglobin from the mollusk Scapharca inaequivalvis (44). Inthis hemoglobin, the interface between the two subunits com-prises the helices E and F, an arrangement (the so-called EF-dimer) that brings the two hemes in close contact to each other(45). In Scapharca Hb, ligation at one heme is directly trans-mitted to the unligated heme through changes at the intersub-unit contact. The dimeric arrangement of human CYGB isradically different from that of the EF-dimer as shown by thethree-dimensional structure of the double cysteine mutant re-cently solved by x-ray crystallography (10, 43). The contactinterface consists mainly of the B and H helices and of the BChinge region, whereby the two heme groups face opposite sidesof the dimer surface. Thus, the mechanism of heme-heme co-operativity in CYGB must necessarily involve conformationaltransitions that are entirely different from those observed inScapharca Hb. Interestingly, in the crystal structure of thedouble cysteine mutant of CYGB, one heme of the dimer is inthe hexacoordinate form with His(E7) as the sixth heme ligandand the other heme is in the pentacoordinate form due to a shiftof the E1-E10 segment (10). These two structures of the hemepocket, corresponding to closed and open conformations, re-
O2 Equilibria in Neuroglobin and Cytoglobin44424
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from

spectively, suggest that the crystallization process may have“frozen” an intermediate species along the ligation pathway(10). No clear pathway of signal transmission can be identifiedat this stage, but it appears that the control of heme affinitydoes not depend on proximal effects as in human HbA since thebond between the heme and the proximal His(F8) remainsunaltered regardless of heme coordination on the distal side byHis(E7) (10, 43).
Physiological Consequences—Since their discovery as novelglobin proteins, NGB and CYGB have been assumed to partic-ipate to O2 transport and delivery similar to their homologousglobin proteins Mb and Hb. This hypothesis can be now verifiedbased on the measured values of O2 affinity (P50) for NGB andCYGB reported here. The reducing conditions used in thisstudy (without free metals and with the Hayashi enzymaticsystem) mimic those existing within living cells where oxidizedthiols in either the S–S (46, 47) or S–NO form (48) are intrin-sically unstable. This condition is largely due to in vivo highconcentrations of reduced GSH (�5 mM) and to the high (�500)ratio between GSH and GSSG (49). Oxidation of thiol groups tothe S–S form increases ligand affinity in NGB and CYGB underspecific in vitro conditions (17).
Under our experimental conditions, the O2 affinity of humanNGB falls in the lower end of the range of values for O2-bindingheme proteins with a P50 value at 37 °C of 7.5 torr at neutralpH (Fig. 1A). For comparison, Mb has typically a P50 of 2–3 torr(50). Within intact cells, partial O2 tension has been estimatedto vary between 1 and 10 torr (51) and to be �2–3 torr in musclecells (50). Unfortunately, no data are available on the O2 ten-sion existing inside neurons; however, given their high meta-bolic rate, which at rest can be compared with that of theheavily working muscle (52), the neurons are expected to ex-perience even lower O2 tensions than muscle cells. In the ret-ina, one of the tissues with the highest metabolic rate, cellularPO2 values close to zero, have been measured in dark-adaptedretina from cats (53). This means that, under in vivo conditions,the fractional O2 saturation of NGB would be too low to play asignificant role as O2 reserve. Assuming a P50 of 7.5 torr at37 °C and an intracellular PO2 of 1 torr, at approximatingnormal in vivo conditions, only �12% NGB molecules will besaturated with O2 given that, in monomeric proteins, Y �PO2/(PO2 P50), where Y is the fractional saturation. Al-though our data do not support a physiological role of NGBinvolving O2 binding and delivery in humans (and possibly inmammals), such a role may apply to invertebrate nerve hemo-globins that normally operate at lower temperatures andhigher internal PO2 and occur at higher concentration wherebythey can provide adequate O2 supply to sustain nerve electricalactivity as originally proposed (20). Thus, at present, the phys-iological function of NGB seems rather to relate to NO andperoxynitrite scavenging during hypoxia (14), which is consist-ent with the protective effect of this protein during lack of O2
supply (18, 19).The physiological function of CYGB conversely seems to be
consistent with O2 supply as evident from its O2 affinity andcooperativity and from the intrinsic high stability of theFe(II)O2 complex. However, due to the low in vivo concentra-tion estimated, CYGB may be involved in O2-requiring cellularreactions unrelated to mitochondrial respiration. These reac-tions may include Pro-hydroxylases involved in collagen syn-thesis, as recently proposed by Schmidt et al. (21), or Asn- andPro-hydroxylases involved in hydroxylation of hypoxia-induc-ible factor �. These two hydroxylases are localized in the nu-cleus and in the cytoplasm, respectively. Interestingly, CYGBhas been localized in the nucleus and in the cytoplasm (5, 54)and is up-regulated during hypoxia (21).
Acknowledgments—We thank Kristina Litvin and Anna SofiaFagersten (Aarhus) for excellent technical assistance and Drs. MichaelC. Marden and Martino Bolognesi for critical reading of the paper.
REFERENCES
1. Burmester, T., Weich, B., Reinhardt, S., and Hankeln, T. (2000) Nature 407,520–523
2. Schmidt, M., Giessl, A., Laufs, T., Hankeln, T., Wolfrum, U., and Burmester,T. (2003) J. Biol. Chem. 278, 1932–1935
3. Reuss, S., Saaler-Reinhardt, S., Weich, B., Wystub, S., Reuss, M. H., Burm-ester, T., and Hankeln, T. (2002) Neuroscience 115, 645–656
4. Mammen, P. P. A., Shelton, J. M., Goetsch, S. C., Williams, S. C., Richardson,J. A., Garry, M. G., and Garry, D. J. (2002) J. Histochem. Cytochem. 50,1591–1598
5. Burmester, T., Ebner, B., Weich, B., and Hankeln, T. (2002) Mol. Biol. Evol.19, 416–421
6. Trent, J. T., III, and Hargrove, M. S. (2002) J. Biol. Chem. 277, 19538–195457. Pesce, A., Dewilde, S., Nardini, M., Moens, L., Ascenzi, P., Hankeln, T.,
Burmester, T., and Bolognesi, M. (2003) Structure 11, 1087–10958. Dewilde, S., Kiger, L., Burmester, T., Hankeln, T., Baudin-Creuza, V., Aerts,
T., Marden, M. C., Caubergs, R., and Moens, L. (2001) J. Biol. Chem. 276,38949–38955
9. Couture, M., Burmester, T., Hankeln, T., and Rousseau, D. L. (2001) J. Biol.Chem. 276, 36377–36382
10. de Sanctis, D., Dewilde, S., Pesce, A., Moens, L., Ascenzi, P., Hankeln, T.,Burmester, T., and Bolognesi, M. (2004) J. Mol. Biol. 336, 917–927
11. Vallone, B., Nienhaus, K., Brunori, M., and Nienhaus, G. U. (2004) Proteins56, 85–92
12. Sugimoto, H., Makino, M., Sawai, H., Kawada, N., Yoshizato, K., and Shiro, Y.J. Mol. Biol., in press
13. Pesce, A., Bolognesi, M., Bocedi, A., Ascenzi, P., Dewilde, S., Moens, L.,Hankeln, T., and Burmester, T. (2002) EMBO Rep. 3, 1146–1151
14. Herold, S., Fago, A., Weber, R. E., Dewilde, S., and Moens, L. (2004) J. Biol.Chem. 279, 22841–22847
15. Kriegl, J. M., Bhattacharyya, A. J., Nienhaus, K., Deng, P., Minkow, O., andNienhaus, G. U. (2002) Proc. Natl. Acad. Sci. U. S. A. 99, 7992–7997
16. Du, W., Syvitski, R., Dewilde, S., Moens, L., and La Mar, G. N. (2003) J. Am.Chem. Soc. 125, 8080–8081
17. Hamdane, D., Kiger, L., Dewilde, S., Green, B. N., Pesce, A., Uzan, J., Burm-ester, T., Hankeln, T., Bolognesi, M., Moens, L., and Marden, M. C. (2003)J. Biol. Chem. 278, 51713–51721
18. Sun, Y., Jin, K., Peel, A., Mao, X. O., Xie, L., and Greenberg, D. A. (2003) Proc.Natl. Acad. Sci. U. S. A. 100, 3497–3500
19. Sun, Y., Jin, K., Mao, X. O., Zhu, Y., and Greenberg, D. A. (2001) Proc. Natl.Acad. Sci. U. S. A. 98, 15306–15311
20. Kraus, D. W., and Colacino, J. M. (1986) Science 232, 90–9221. Schmidt, M., Gerlach, F., Avivi, A., Laufs, T., Wystub, S., Simpson, J. C., Nevo,
E., Saaler-Reinhardt, S., Reuss, S., Hankeln, T., and Burmester, T. (2004)J. Biol. Chem. 279, 8063–8069
22. Trent, J. T., III, Watts, R. A., and Hargrove, M. S. (2001) J. Biol. Chem. 276,30106–30110
23. Lipton, P. (1999) Physiol. Rev. 79, 1431–156824. Alayash, A. I., Patel, R. P., and Cashon, R. E. (2001) Antioxid. Redox Signal.
3, 313–32725. Antonini, E., and Brunori, M. (1971) Hemoglobin and Myoglobin in Their
Reactions with Ligands, North-Holland Publishing Company, Amsterdam26. Hayashi, A., Suzuki, T., and Shin, M. (1973) Biochim. Biophys. Acta 310,
309–31627. Weber, R. E. (1981) Nature 292, 386–38728. Weber, R. E. (1992) J. Appl. Physiol. 72, 1611–161529. Grassetti, D. R., and Murray, J. F. (1967) Arch. Biochem. Biophys. 119, 41–4930. Bonaventura, C., Tesh, S., Faulkner, K. M., Kraiter, D., and Crumbliss, A. L.
(1998) Biochemistry 37, 496–50631. Kussmann, M., Lassing, U., Sturmer, C. A., Przybylski, M., and Roepstorff, P.
(1997) J. Mass Spectrom. 32, 483–49332. Fago, A., Giangiacomo, L., D’Avino, R., Carratore, V., Romano, M., Boffi, A.,
and Chiancone, E. (2001) J. Biol. Chem. 276, 27415–2742333. Briehl, R. W. (1963) J. Biol. Chem. 238, 2361–236634. Nienhaus, K., Kriegl, J. M., and Nienhaus, G. U. (2004) J. Biol. Chem. 279,
22944–2295235. Creighton, T. E. (1993) Proteins. Structures and Molecular Properties, 2nd Ed.,
W. H. Freeman & Co., New York36. Uzan, J., Dewilde, S., Burmester, T., Hankeln, T., Moens, L., Hamdane, D.,
Marden, M. C., and Kiger, L. Biophys. J., in press37. Wyman, J. (1964) Adv. Protein Chem. 19, 223–28638. Fago, A., Wells, R. M. G., and Weber, R. E. (1997) Comp. Biochem. Physiol. B
118, 319–32639. Uno, T., Ryu, D., Tsutsumi, H., Tomisugi, Y., Ishikawa, Y., Wilkinson, A. J.,
Sato, H., and Hayashi, T. (2004) J. Biol. Chem. 279, 5886–589340. Gersonde, K., Sick, H., Overkamp, M., Smith, K. M., and Parish, D. W. (1986)
Eur. J. Biochem. 157, 393–40441. Perutz, M. F. (1979) Annu. Rev. Biochem. 48, 327–38642. Sawai, H., Kawada, N., Yoshizato, K., Nakajima, H., Aono, S., and Shiro, Y.
(2003) Biochemistry 42, 5133–514243. de Sanctis, D., Dewilde, S., Pesce, A., Moens, L., Ascenzi, P., Hankeln, T.,
Burmester, T., and Bolognesi, M. (2004) Biochem. Biophys. Res. Commun.316, 1217–1221
44. Chiancone, E., Vecchini, P., Verzili, D., Ascoli, F., and Antonini, E. (1981) J.Mol. Biol. 152, 577–592
45. Royer, W. E., Knapp, J. E., Strand, K., and Heaslet, H. A. (2001) TrendsBiochem. Sci. 26, 297–304
46. Schafer, F. Q., and Buettner, G. R. (2001) Free Radic. Biol. Med. 30, 1191–121247. Lodish, H., Berk, A., Zipursky, S. L., Matsudaira, P., Baltimore, D., and
O2 Equilibria in Neuroglobin and Cytoglobin 44425
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from

Darnell, J. E. (2000) Molecular Cell Biology, 4th Ed., W. H. Freeman & Co,New York
48. Gladwin, M. T., Wang, X., Reiter, C. D., Yang, B. K., Vivas, E. X., Bonaventura,C., and Schechter, A. N. (2002) J. Biol. Chem. 277, 27818–27828
49. Berg, J. M., Tymoczko, J. L., and Stryer, L. (2002) Biochemistry, p. 686, W. H.Freeman & Co., New York
50. Wittenberg, J. B., and Wittenberg, B. A. (2003) J. Exp. Biol. 206, 2011–202051. Halliwell, B., and Gutteridge, J. M. C. (1999) Free Radicals in Biology and
Medicine, 3rd Ed., Oxford University Press, New York52. Hochachka, P. W., and Somero, G. N. (2002) Biochemical Adaptation. Mecha-
nism, and Process in Physiological Evolution, Oxford University Press,Oxford
53. Wangsa-Wirawan, N. D., and Linsenmeier, R. A. (2003) Arch. Ophthalmol.121, 547–557
54. Geuens, E., Brouns, I., Flamez, D., Dewilde, S., Timmermans, J. P., andMoens, L. (2003) J. Biol. Chem. 278, 30417–30420
O2 Equilibria in Neuroglobin and Cytoglobin44426
at TE
XA
S T
EC
H U
NIV
ER
SIT
Y - T
rial Subscription on M
arch 8, 2009 w
ww
.jbc.orgD
ownloaded from
Related Documents











