RESEARCH ARTICLE A. Penna E. Garce´s M. Vila M. G. Giacobbe S. Fraga A. Luglie` I. Bravo E. Bertozzini C. Vernesi Alexandrium catenella (Dinophyceae), a toxic ribotype expanding in the NW Mediterranean Sea Received: 15 November 2004 / Accepted: 10 June 2005 / Published online: 11 August 2005 Ó Springer-Verlag 2005 Abstract The presence of the paralytic shellfish poisoning (PSP) dinoflagellate Alexandrium catenella in the north western (NW) Mediterranean Sea has been known since 1983. From this date on, the species has spread along the Spanish and Italian coastlines. Information concerning A. catenella isolates in the NW Mediterranean Sea was gained through phylogenetic studies. Twenty established toxic cultures of A. catenella taken from various NW Mediterranean Sea locations were analysed by nucleotide sequencing of the 5.8S rDNA and internal transcribed spacer regions. These rDNA ribosomal markers resulted useful in delineating the phylogenetic position of this species in the genus Alexandrium as well as in determin- ing relationships between A. catenella isolates from dif- ferent geographic areas. The phylogenetic position of the Mediterranean A. catenella ribotype, when compared to the ‘‘Alexandrium tamarense/catenella/fundyense species complex’’, fits this species complex well. All the Medi- terranean A. catenella isolates were constituted by only one genetic ribotype. By comparing the isolate sequences with those of other geographic areas, it revealed that the Mediterranean A. catenella ribotype was closely related to the A. catenella from Japan, Western Pacific Ocean. It was also evident that in temperate Japanese waters, a genetic variability was detected within A. catenella iso- lates; in fact, all strains resulted divergent showing as many as 15 mutational steps. The possibility that A. catenella has been recently introduced into the Med- iterranean basin from temperate Asian areas is discussed. Introduction A Harmful Algal Bloom (HAB) is defined by a rapid and intense increase in the cell density of a noxious species in seawater (Smayda 1997). Although only a 2% of the world’s marine microalgae, represented by approximately 60–80 species, are harmful (Sournia 1995), they are an important threat affecting marine fauna directly or through the food chain and producing severe economic losses and negative impact on the environment and human health. Since the 1980s, there has been an increase in the frequency and intensity of HABs in the Mediterranean Sea. These events are characterized by blooms of various dinoflagellates, especially the genus Alexandrium Halim, comprising various toxic and non-toxic species (Garce´s et al. 2000; Vila et al. 2001a). A. catenella is, among others, a pro- ducer of neurotoxic PSP compounds. Although this species is commonly reported as a cold-water micro- organism in several areas of the world, such as the Western Pacific Ocean of North America (Scholin and Anderson 1994), in the upwelling coastal waters of Chile Communicated by R. Cattaneo-Vietti, Genova A. Penna (&) E. Bertozzini Centro Biologia Ambientale, University of Urbino, V.le Trieste 296, 61100 Pesaro, Italy E-mail: [email protected] Tel.: +39-0721-423526 Fax: +39-0721-423522 E. Garce´s M. Vila Institut de Cie`ncies del Mar, Passeig Marı´tim de la Barceloneta, 37-49, 08003 Barcelona, Spain M. G. Giacobbe Istituto per l’Ambiente Marino Costiero, CNR, Sp. S. Raineri 86, 98122 Messina, Italy S. Fraga I. Bravo Instituto Espan˜ol de Oceanografı´a, Apdo. 1552, 36200 Vigo, Spain A. Luglie` Dip. Botanica ed Ecologia Vegetale, University of Sassari, 07100 Sassari, Italy C. Vernesi Centro di Ecologia Alpina, Viote del Monte Bondone, 38040 Trento, Italy Marine Biology (2005) 148: 13–23 DOI 10.1007/s00227-005-0067-5

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH ARTICLE
A. Penna Æ E. Garces Æ M. Vila Æ M. G. Giacobbe
S. Fraga Æ A. Luglie Æ I. Bravo Æ E. BertozziniC. Vernesi
Alexandrium catenella (Dinophyceae), a toxic ribotype expandingin the NW Mediterranean Sea
Received: 15 November 2004 / Accepted: 10 June 2005 / Published online: 11 August 2005� Springer-Verlag 2005
Abstract The presence of the paralytic shellfish poisoning(PSP) dinoflagellate Alexandrium catenella in the northwestern (NW) Mediterranean Sea has been known since1983. From this date on, the species has spread along theSpanish and Italian coastlines. Information concerningA. catenella isolates in the NW Mediterranean Sea wasgained through phylogenetic studies. Twenty establishedtoxic cultures of A. catenella taken from various NWMediterranean Sea locations were analysed by nucleotidesequencing of the 5.8S rDNA and internal transcribedspacer regions. These rDNA ribosomal markers resulteduseful in delineating the phylogenetic position of thisspecies in the genus Alexandrium as well as in determin-ing relationships between A. catenella isolates from dif-ferent geographic areas. The phylogenetic position of the
Mediterranean A. catenella ribotype, when compared tothe ‘‘Alexandrium tamarense/catenella/fundyense speciescomplex’’, fits this species complex well. All the Medi-terranean A. catenella isolates were constituted by onlyone genetic ribotype. By comparing the isolate sequenceswith those of other geographic areas, it revealed that theMediterranean A. catenella ribotype was closely relatedto the A. catenella from Japan, Western Pacific Ocean. Itwas also evident that in temperate Japanese waters, agenetic variability was detected within A. catenella iso-lates; in fact, all strains resulted divergent showing asmany as 15 mutational steps. The possibility thatA. catenella has been recently introduced into the Med-iterranean basin from temperate Asian areas is discussed.
Introduction
A Harmful Algal Bloom (HAB) is defined by a rapidand intense increase in the cell density of a noxiousspecies in seawater (Smayda 1997). Although only a 2%of the world’s marine microalgae, represented byapproximately 60–80 species, are harmful (Sournia1995), they are an important threat affecting marinefauna directly or through the food chain and producingsevere economic losses and negative impact on theenvironment and human health. Since the 1980s, therehas been an increase in the frequency and intensity ofHABs in the Mediterranean Sea. These events arecharacterized by blooms of various dinoflagellates,especially the genus Alexandrium Halim, comprisingvarious toxic and non-toxic species (Garces et al. 2000;Vila et al. 2001a). A. catenella is, among others, a pro-ducer of neurotoxic PSP compounds. Although thisspecies is commonly reported as a cold-water micro-organism in several areas of the world, such as theWestern Pacific Ocean of North America (Scholin andAnderson 1994), in the upwelling coastal waters of Chile
Communicated by R. Cattaneo-Vietti, Genova
A. Penna (&) Æ E. BertozziniCentro Biologia Ambientale, University of Urbino,V.le Trieste 296, 61100 Pesaro, ItalyE-mail: [email protected].: +39-0721-423526Fax: +39-0721-423522
E. Garces Æ M. VilaInstitut de Ciencies del Mar,Passeig Marıtim de la Barceloneta,37-49, 08003 Barcelona, Spain
M. G. GiacobbeIstituto per l’Ambiente Marino Costiero, CNR,Sp. S. Raineri 86, 98122 Messina, Italy
S. Fraga Æ I. BravoInstituto Espanol de Oceanografıa, Apdo. 1552,36200 Vigo, Spain
A. LuglieDip. Botanica ed Ecologia Vegetale, University of Sassari,07100 Sassari, Italy
C. VernesiCentro di Ecologia Alpina, Viote del Monte Bondone,38040 Trento, Italy
Marine Biology (2005) 148: 13–23DOI 10.1007/s00227-005-0067-5

(Cordova and Muller 2002), Southern Argentina(Benavides et al. 1995), Western South Africa (RuizSebastian et al. 2005) and New Zealand (MacKenzieet al. 2004), it has also been observed in the temperatewaters of Japan (Adachi et al. 1994), South-easternAustralia (Hallegraeff et al. 1991), China (Yeung et al.2002) and the Western Mediterranean Sea (Vila et al.2001b; Luglie et al. 2003a). Before the first widespreadPSP toxic event, occurring along 100 km of the Catalancoastline of Spain in 1998 and 1999 (Vila et al. 2001b),the presence of A. catenella in the Mediterranean hadonly been reported as a sporadic occurrence in theBalearic-Catalan Sea in 1983 (Margalef and Estrada1987). In 1994, a bloom of Alexandrium sp. (later iden-tified as A. catenella), characterized by a cell density upto 1.0 · 104 cells l�1 was reported in Valencia Harbour(Spain) (Gomis et al. 1996). Furthermore, A. catenellaalso appeared in the Mediterranean for the first time inthe waters of Tunisia, during the summer of 1997(S. Turki, personal communication), and in the OlbiaHarbour, on the Tyrrhenian Sea (Italy) in the summersof 1999 to 2002 (Luglie et al. 2003b).
A relevant tool in taxonomic studies is provided bythe use of molecular markers to assess the phyloge-netic position of eukaryotic unicellular organisms(Manhart and McCourt 1992; Saunders et al. 1997).Genetic analyses based on the LSU rRNA gene, wereused to investigate the phylogenetic relationships be-tween the different species of genus Alexandrium(Scholin et al. 1994; Adachi et al. 1996; Medlin et al.1998; Higman et al. 2001; Guillou et al. 2002; Johnet al. 2003; Ruiz Sebastian et al. 2005) and it wasfound that the three morphospecies A. tamarense,A. catenella and A. fundyense should be considered asa ‘‘species complex’’. It was demonstrated that differ-ent ribotypes could contain specimens assigned to eachof the three morphospecies of the ‘‘species complex’’and that their isolates usually correlate more closelywith their geographic origin than with their morpho-type. Subsequently, the ‘‘A. tamarense/catenella/fundy-ense species complex’’ was subdivided into fiveribotype/geographic clades according to the isolate’sgeographical origin as follows: toxic North American(NA), Temperate Asian (TA), Tropical Asian (TROP)as well as non - toxic Western European (WE), Tas-manian (TASM), and the new Mediterranean (ME)clade (Scholin et al. 1995; John et al. 2003; RuizSebastian et al. 2005).
Lilly et al. (2002) found that two clones of A. cate-nella isolated from Thau Lagoon, which is located onthe Mediterranean coast of France, corresponded to theTA clade based on the LSU rRNA and, as they did notfind any other isolates in Europe belonging to this clade,they concluded that most probably A. catenella wasintroduced into Thau Lagoon through ballast waters.
In this study, in order to delineate the geographicorigin of the toxic species A. catenella in the NW Medi-terranean, phylogenetic analyses were carried out. OurA. catenella isolates from the Catalan-Balearic and Tyr-
rhenian Seas of the NW Mediterranean were comparedwith isolates from Japan and Chile based on the 5.8SrDNA and ITS region sequences. Furthermore, thephylogenetic relationship between the different A. cate-nella isolates within, as well as outside the Mediterraneanand other Alexandrium species were delineated by usingthe ribosomal markers 5.8S rDNA gene and ITS regions.
Materials and methods
Sample collection and culture
Several strains of A. catenella isolated for this study arelisted in Table 1. Water and sediment samples werecollected from a number of different sites along theCatalan (Catalan/Balear Sea, Spain) and Sardinian(Tyrrhenian Sea, Italy) coasts (Fig. 1). Each culturestrain was established from vegetative cells or restingcysts of a single water or sediment sample and main-tained in F/2, K, or L1 media (Guillard 1975; Kelleret al. 1987; Guillard and Hargraves 1993) at 17 –21±1�C and a 14:10 h (light:dark) photoperiod. Illu-mination was provided by a photon irradiance of about100 lmol m�2 s�1. All isolates were identified by mor-phological characterization following the criteria ofBalech (1995). Thecal plates of vegetative cells wereexamined under a UV epifluorescence microscope usingFluorescent brightener 28 (SIGMA-ALDRICH), stain-ing and phase contrast by light microscopy (LM). Allthe A. catenella strains listed in Table 1 resulted PSP-toxic (J. M. Franco, personal communication) andcurrently maintained in the CNR (Consiglio Nazionaledelle Ricerche, Messina, Italy), CSIC (Institut de Cien-cies del Mar, Barcelona, Spain) or IEO (Instituto Es-panol de Oceanografia, Vigo, Spain) culture collections.
DNA extraction, PCR amplification and sequencing
Genomic DNA was extracted from approximately 10 mlculture in logarithmic growth phase using the Dynabe-ads DNA DirectTM Kit (Dynal, A.S., Oslo, Norway),the Puregene DNA Isolation Kit (Gentra Systems,Minneapolis, MN, USA) or DNeasy Plant Kit (Qiagen,Valencia, CA, USA), according to the manufacturer’sinstructions.
The PCR amplification conditions of the 5.8S rDNAgene and ITS regions were obtained as described byPenna and Magnani (1999). Three PCR-amplifiedproducts of the 5.8S rDNA gene and ITS regions werepooled, purified and then directly sequenced or clonedfor sequence analyses. The amplified PCR fragmentswere cloned in the vector pDrive Cloning Vector (Qia-gen) and sequenced. Nucleotide sequencing was per-formed using the ABI PRISM 310 Genetic Analyzer(Perkin Elmer, Applied Biosystems, Foster City, CA,USA) and the dye terminator method was used
14

Table 1 Sample locations in the Mediterranean Sea, date, origin, source and EMBL accession numbers of the different A. catenella strainsisolated from this study
Strain Location Date Origin cellularphase
Source EMBLAccession no.
CNR-ACATA4 Tyrrhenian Sea, Olbia St. A, Italy 27-May-2002 Vegetative M.G. Giacobbe/A. Luglie AJ532910CNR-ACATC2 Tyrrhenian Sea, Olbia St. C, Italy 27-May-2002 Vegetative M.G. Giacobbe/A. Luglie AJ580317CNR-ACATS1 Tyrrhenian Sea, Olbia St. S, Italy 02-Jun-2002 Vegetative M.G. Giacobbe/A. Luglie AJ580319CNR-ACATS2 Tyrrhenian Sea, Olbia St. S, Italy 02-Jun-2002 Vegetative M.G. Giacobbe/A. Luglie AJ580318CNR-ACATS3 Tyrrhenian Sea, Olbia St. S, Italy 02-Jun-2002 Vegetative M.G. Giacobbe/A. Luglie AJ580320CSIC-D4 Catalan Sea, Barcelona, Spain 30-May-1998 Vegetative E. Garces AJ298900IEO-715 Catalan Sea, Barcelona, Spain 12-Jun-2003 Vegetative S. Fraga AJ968681CSIC-C5 Catalan Sea, Tarragona, Spain 30-May-1998 Vegetative M. Delgado AJ580321CSIC-C7 Catalan Sea, Tarragona, Spain 30-May-1998 Vegetative M. Delgado AJ580322CSIC-5T Catalan Sea, Tarragona, Spain 10-Jun-2001 Cyst I. Bravo/E. Garces AJ580325CSIC-6T Catalan Sea, Tarragona, Spain 10-Jun-2001 Cyst I. Bravo/E. Garces AJ580323CSIC-8T Catalan Sea, Tarragona, Spain 10-Jun-2001 Cyst I. Bravo/E. Garces AJ608265IEO-AC1C Catalan Sea, Tarragona, Spain 15-Mar-2002 Cyst I. Bravo/E. Garces AJ532911IEO-AC2C Catalan Sea, Barcelona, Spain 15-Mar-2002 Cyst I. Bravo/E. Garces AJ532912IEO-AC9T Catalan Sea, Tarragona, Spain 28-Oct-2002 Cyst I. Bravo/E. Garces AJ630645IEO-594 Catalan Sea, Tarragona, Spain 01-Oct-2002 Vegetative S. Fraga AJ968682IEO-709 Catalan Sea, Tarragona, Spain 29-Apr-2003 Vegetative S. Fraga AJ968683ATTL01 Catalan Sea,Thau Lagoon, France 01-Nov-1998 Vegetative D. Kulis AJ608263ATTL02 Catalan Sea,Thau Lagoon, France 01-Nov-1998 Vegetative D. Kulis AJ608264IEO-816 Catalan Sea,Thau Lagoon, France 26-Oct-2004 Vegetative S. Fraga AJ968680
CNR Consiglio Nazionale delle Ricerche, Messina, Italy; CSIC Institut de Ciencies del Mar, Barcelona, Spain; IEO Instituto Espanol deOceanografia, Vigo, Spain
Fig. 1 Location of the samplingareas in the WesternMediterranean Sea. Site 1Olbia, Tyrrhenian Sea, Italy.Site 2 Barcelona, Catalan Sea,Spain. Site 3 Tarragona,Catalan Sea, Spain. Site 4 ThauLagoon, Catalan Sea, France
15
according to the manufacturer’s instructions (ABIPRISM Big Dye Terminator Cycle Sequencing Readyreaction Kit, Perkin Elmer Corp.). Each clone was se-quenced in both directions three times, in order to re-move ambiguities. The sequences were deposited atEMBL Bank and are listed in Table 1.
Phylogenetic analyses
Sequence alignment was carried out with SAM (Karpluset al. 1998) software, which implements a Hidden
Markov Chain algorithm, and subsequently checked byeye. The 5.8S rDNA-coding region and flanking se-quences of ITS were aligned with other ITS sequences ofAlexandrium species listed in GenBank. The sequencealignment analyses were also carried out including othersequence data of A. catenella and Alexandrium speciesfrom the GenBank (Table 2). Phylogenetic relation-ships, based on the ITS-5.8S rDNA, were inferred usingthe neighbor-joining (NJ) maximum likelihood (ML)and Bayesian (BI) methods. The sequence of Lingulodi-nium polyedrum (GenBank AF377944) was used asoutgroup. To choose the best-fit model of nucleotide
Table 2 List of Alexandrium species 5.8S rDNA gene and ITS region sequences used in this study
Species Strain Samplinglocation
Geographicclade
Sourcea Accessionnumber
Toxicity
A. affine H1 Harima Nada, Japan Temperate Asia Adachi et al. 1996 AB006995 No-PSPA. affine IEO-PA4V Vigo, Spain Europe Atlantic This study AJ632094 No-PSPA. affine IEO-PA8V Andalucia, Spain Mediterranean This study AJ632095 No-PSPA. catenella M17 Harima Nada, Japan Temperate Asia Adachi et al. 1996 AB006990 Yes-PSPA. catenella OFX102 Ofunato Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. catenella OFY101 Ofunato Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. catenella TNY11 Tanabe Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. catenella TNX22 Tanabe Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. catenella Y-2 Yamakawa, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. catenella K0-3 Uranouchi Inlet, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. catenella ACC01 Chile North America Marin et al. 2001 AJ272120 Yes-PSPA. fraterculus AF-1 Pusan, Korea Temperate Asia Cho et al. 1999 AF208242 No-PSPA. fundyense GtCA29 Cape Ann, MA, USA North America Adachi et al. 1996 NR Yes-PSPA. insuetum S1 Shoudoshima, Japan Temperate Asia Adachi et al. 1996 AB006996 No-PSPA. margalefi CNR-AM1 Oliveri, Italy Mediterranean Penna et al. 1999 AJ251208 No-PSPA. minutum LAC 27 Trieste, Italy Mediterranean Penna et al. 2000 AJ005050 Yes-PSPA. minutum CNR-AMIA1 Siracusa, Italy Mediterranean Penna et al. 2003 AJ621734 Yes-PSPA. minutum CNR-AMIA4 Siracusa, Italy Mediterranean Penna et al. 2003 AJ318460 Yes-PSPA. minutum CNR-AMIA5 Siracusa, Italy Mediterranean Penna et al. 2003 AJ532913 Yes-PSPA. minutum IEO-AL8C Arenys, Spain Mediterranean Penna et al. 2003 AJ532914 Yes-PSPA. minutum IEO-AL9C Arenys, Spain Mediterranean Penna et al. 2003 AJ621733 Yes-PSPA. minutum CSIC-D1 Arenys, Spain Mediterranean Penna et al. 2003 AJ312945 Yes-PSPA. pseudogonyaulax H1 Harima Nada, Japan Temperate Asia Adachi et al. 1996 AB006997 No-PSPA. tamarense CU-15 Gulf of Thailand, Thailand Tropical Asia Adachi et al. 1996 AB006992 Yes-PSPA. tamarense FK-788 Funka Bay, Japan Temperate Asia Adachi et al. 1996 AB006993 Yes-PSPA. tamarense PW06 Port Benny, AK, USA North America Adachi et al. 1996 NR Yes-PSPA. tamarense AT-A Chinhae Bay, Korea Temperate Asia Cho E.S. 2001 AF374224 PSP?A. tamarense AT-B Chinhae Bay, Korea Temperate Asia Cho E.S. 2001 AF374225 PSP?A. tamarense AT-2 Chinhae Bay, Korea Temperate Asia Cho E.S. 2001 AF374227 PSP?A. tamarense AT-6 Chinhae Bay, Korea Temperate Asia Cho E.S. 2001 AF374228 PSP?A. tamarense AT-10 Chinhae Bay, Korea Temperate Asia Cho E.S. 2001 AF374226 PSP?A. tamarense At503A-A Mikawa Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. tamarense At304A Mikawa Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. tamarense OK875-1 Okirai Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. tamarense OFX151 Ofunato Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. tamarense OFX191 Ofunato Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. tamarense HIAI Hiroshima Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. tamarense HI38 Hiroshima Bay, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. tamarense AT4 Harima Nada, Japan Temperate Asia Adachi et al. 1996 NR Yes-PSPA. tamarense PE1V Vigo, Spain Western Europe Penna et al. 1999 AJ005047 Yes-PSPA. tamarense WKS-1 Kushimoto, Japan Western Europe Adachi et al. 1996 AB006991 Yes-PSPA. tamiyavanichii MS-01 Sebatu, Malaysia Tropical Asia Usup et al. 2002 AF145224 No-PSPA. taylori CSIC-AV8 La Fosca, Spain Mediterranean Penna et al. 1999 AJ251654 No-PSPA. taylori CNR-AT4 Siracusa, Italy Mediterranean Penna et al. 1999 AJ251653 No-PSPA. taylori CNR-ATAYB2 Vulcano, Italy Mediterranean Penna et al. 2000 AJ300451 No-PSPA. taylori Temporary-cyst Vulcano, Italy Mediterranean Penna et al. 2000 AJ291785 No-PSPA. taylori CBA-1 Kavala, Greece Mediterranean Penna et al. 2001 AJ416856 No-PSP
aWho sequenced the strain; IEO Instituto Espanol de Oceanogra-fia, Vigo, Spain; CNR Consiglio Nazionale delle Ricerche, Messina,Italy; CSIC Institut de Ciencies del Mar, Barcelona, Spain; CCMP
Provasoli-Guillard National Center for Culture Marine Phyto-plankton, Bigelow Laboratory, USA; CBA Centro Biologia Am-bientale, University of Urbino, Urbino, Italy; NR not reported
16
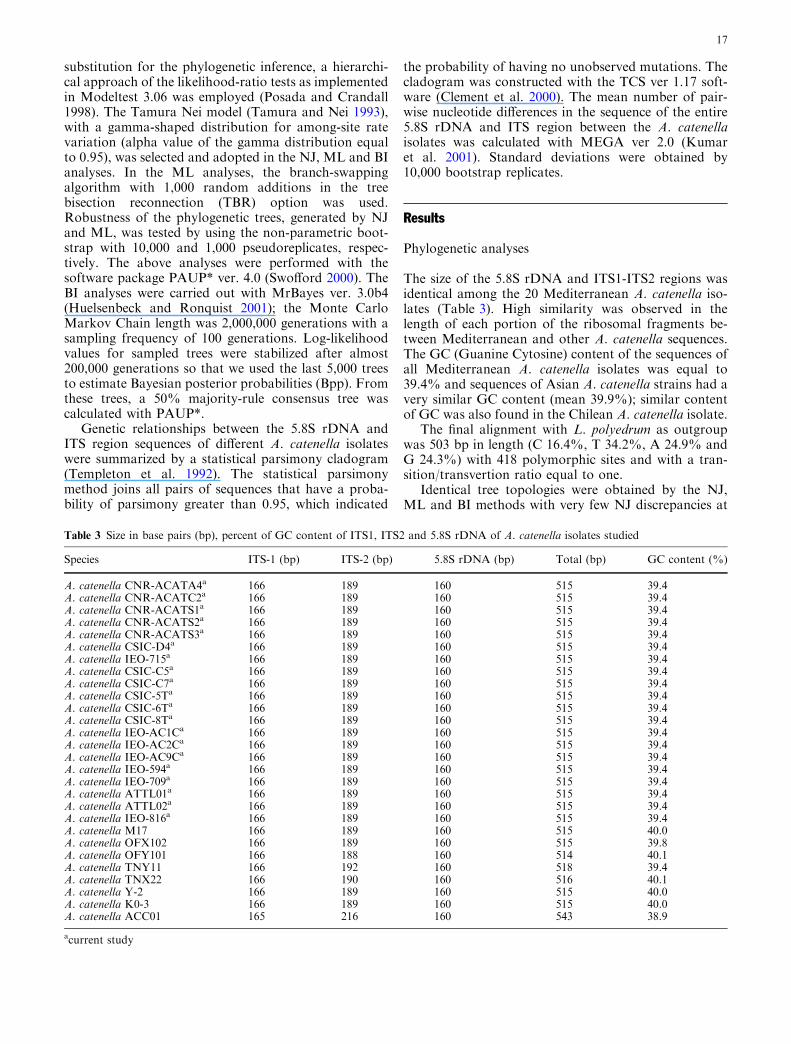
substitution for the phylogenetic inference, a hierarchi-cal approach of the likelihood-ratio tests as implementedin Modeltest 3.06 was employed (Posada and Crandall1998). The Tamura Nei model (Tamura and Nei 1993),with a gamma-shaped distribution for among-site ratevariation (alpha value of the gamma distribution equalto 0.95), was selected and adopted in the NJ, ML and BIanalyses. In the ML analyses, the branch-swappingalgorithm with 1,000 random additions in the treebisection reconnection (TBR) option was used.Robustness of the phylogenetic trees, generated by NJand ML, was tested by using the non-parametric boot-strap with 10,000 and 1,000 pseudoreplicates, respec-tively. The above analyses were performed with thesoftware package PAUP* ver. 4.0 (Swofford 2000). TheBI analyses were carried out with MrBayes ver. 3.0b4(Huelsenbeck and Ronquist 2001); the Monte CarloMarkov Chain length was 2,000,000 generations with asampling frequency of 100 generations. Log-likelihoodvalues for sampled trees were stabilized after almost200,000 generations so that we used the last 5,000 treesto estimate Bayesian posterior probabilities (Bpp). Fromthese trees, a 50% majority-rule consensus tree wascalculated with PAUP*.
Genetic relationships between the 5.8S rDNA andITS region sequences of different A. catenella isolateswere summarized by a statistical parsimony cladogram(Templeton et al. 1992). The statistical parsimonymethod joins all pairs of sequences that have a proba-bility of parsimony greater than 0.95, which indicated
the probability of having no unobserved mutations. Thecladogram was constructed with the TCS ver 1.17 soft-ware (Clement et al. 2000). The mean number of pair-wise nucleotide differences in the sequence of the entire5.8S rDNA and ITS region between the A. catenellaisolates was calculated with MEGA ver 2.0 (Kumaret al. 2001). Standard deviations were obtained by10,000 bootstrap replicates.
Results
Phylogenetic analyses
The size of the 5.8S rDNA and ITS1-ITS2 regions wasidentical among the 20 Mediterranean A. catenella iso-lates (Table 3). High similarity was observed in thelength of each portion of the ribosomal fragments be-tween Mediterranean and other A. catenella sequences.The GC (Guanine Cytosine) content of the sequences ofall Mediterranean A. catenella isolates was equal to39.4% and sequences of Asian A. catenella strains had avery similar GC content (mean 39.9%); similar contentof GC was also found in the Chilean A. catenella isolate.
The final alignment with L. polyedrum as outgroupwas 503 bp in length (C 16.4%, T 34.2%, A 24.9% andG 24.3%) with 418 polymorphic sites and with a tran-sition/transvertion ratio equal to one.
Identical tree topologies were obtained by the NJ,ML and BI methods with very few NJ discrepancies at
Table 3 Size in base pairs (bp), percent of GC content of ITS1, ITS2 and 5.8S rDNA of A. catenella isolates studied
Species ITS-1 (bp) ITS-2 (bp) 5.8S rDNA (bp) Total (bp) GC content (%)
A. catenella CNR-ACATA4a 166 189 160 515 39.4A. catenella CNR-ACATC2a 166 189 160 515 39.4A. catenella CNR-ACATS1a 166 189 160 515 39.4A. catenella CNR-ACATS2a 166 189 160 515 39.4A. catenella CNR-ACATS3a 166 189 160 515 39.4A. catenella CSIC-D4a 166 189 160 515 39.4A. catenella IEO-715a 166 189 160 515 39.4A. catenella CSIC-C5a 166 189 160 515 39.4A. catenella CSIC-C7a 166 189 160 515 39.4A. catenella CSIC-5Ta 166 189 160 515 39.4A. catenella CSIC-6Ta 166 189 160 515 39.4A. catenella CSIC-8Ta 166 189 160 515 39.4A. catenella IEO-AC1Ca 166 189 160 515 39.4A. catenella IEO-AC2Ca 166 189 160 515 39.4A. catenella IEO-AC9Ca 166 189 160 515 39.4A. catenella IEO-594a 166 189 160 515 39.4A. catenella IEO-709a 166 189 160 515 39.4A. catenella ATTL01a 166 189 160 515 39.4A. catenella ATTL02a 166 189 160 515 39.4A. catenella IEO-816a 166 189 160 515 39.4A. catenella M17 166 189 160 515 40.0A. catenella OFX102 166 189 160 515 39.8A. catenella OFY101 166 188 160 514 40.1A. catenella TNY11 166 192 160 518 39.4A. catenella TNX22 166 190 160 516 40.1A. catenella Y-2 166 189 160 515 40.0A. catenella K0-3 166 189 160 515 40.0A. catenella ACC01 165 216 160 543 38.9
acurrent study
17

some branch nodes in the NJ tree; only the ML phylo-genetic tree is shown (Fig. 2). Within the genus Alex-andrium, the toxic Mediterranean A. catenella isolateshad identical sequence and grouped together with thetoxic A. catenella isolates from Temperate Asia (Japan);this grouping was well supported by high bootstrap(87% and 91% for NJ and ML, respectively) values and
a 100% of posterior probability of the BI inference.Furthermore, the Mediterranean A. catenella isolatesalong with the Japanese isolates were placed in the ‘A.tamarense/catenella/fundyense species complex’. Here,the ‘A. tamarense/catenella/fundyense species complex’comprised the different geographic ribotypes or cladesand not the three morphospecies (A. fundyense, A.catenella and A. tamarense). Further, the A. catenellaACC01 isolate from Chile clustered together with theNorth American clade sequences. It was also evidentthat the two Western European clade A. tamarense iso-lates grouped together as a sister clade of the A. catenellaribotype. Also, these latter clades and their branchingorder within the ‘A. tamarense/catenella/fundyense spe-cies complex’ were well supported by bootstrap (NJ andML) and posterior probability (BI) values.
Among the other species of Alexandrium examinedto date, A. taylori appeared to diverge earlier. Suc-cessively, species diverged into two clades. One cladeconsisted of A. pseudogonyaulaux and Alexandrium sp.,while the other comprised the remaining species of the
A. catenella ACC01
A. tamarense 788A
A. tamarense AT-B
A. tamarense At304A
A. tamarense HIAI
A. tamarense HI38
A. tamarense OFX151
A. tamarense At503A-A
A. tamarense PW06
A. fundyense GtCA29
A.catenella M17A.catenella TNY11A.catenella OFX102A.catenella Y-2
A.catenella TNX22
A.catenella K0-3
A.catenella OFY101
A. tamarense WKS1
A. tamarense PE1V
A. fraterculus AF1
A. tamayavanichi MS01
A. tamarense CU-15
A. affine IEO-PA4V
A. affine H1
A. margalefi CNR-AM1
A. insuetum S1
A.minutum LAC27
Alexandrium sp. AS1
A. pseudogonyaulax H1A. taylori CNR-AT4
A. taylori AV-8
0.1 substitutions/site
/100100
/79
100
505052
96100100
100100100
100100100
/53
5193
100
/6070
8791
100
647199
8494
100
9999
100
A. catenella (20 Mediterranean isolates)
A. affine IEO-PA8V
N
70
TROPICALASIA clade(TROP)
L. polyedrumAF377944
NORTHAMERICA clade(NA)
TEMPERATEASIA clade(TA)
WESTERNEUROPE clade (WE)
Alexan
driu
mtam
arense/caten
e lla/fun
dyen
sesp
eciesco
mp
lex
Fig. 2 Consensus maximum likelihood phylogenetic tree of thegenus Alexandrium based on the ITS-5.8S rDNA sequences.Numbers on the major nodes represent, from the top to thebottom, NJ (10,000 pseudo-replicates) ML (1,000 pseudo-repli-cates) bootstrap values and Bayesian posterior probability. Onlybootstrap and posterior probability values >50% are shown. Thesymbol/shows the absence of the bootstrap value due to thetopological discrepancy between NJ and ML-BI inferences. Thetree is rooted using L. polyedrum AF377944 as outgroup. The 5.8SrDNA-ITS sequences from this study are in zbold. A. tamarenseAT-B is a representative of the following isolates of A. tamarense,as AT-A, AT-2, AT-6, AT-10, OK875-1, OFX191, AT-4, since theyshare the same sequence. A. minutum and A. taylori isolates hadidentical sequences within each species and only A. minutumLAC27, A. taylori CNR-AT4 and A. taylori AV-8 were shown
18

genus Alexandrium. The bootstrap and posteriorprobability values were usually high, thus confirmingthe utility of the 5.8S rDNA-ITS regions as phyloge-netic markers.
The entire sequence of the 5.8S rDNA and ITS regionsof the Mediterranean, Temperate Asian and Chileanisolates were aligned (523 bp). A total of 9 haplotypes(Fig. 3) and 122 polymorphic sites were found, with 110
substitutions (56 transitions and 54 transversions) and 14indels. A unique isolate from Chile can be resulted alsocompletely divergent from the Mediterranean and Japa-nese haplotypes (112 substitutions), characterized as aseparate clade by the statistical parsimony method. AllMediterranean isolates shared the same haplotype(Mediterranean haplotype). On the contrary, in theJapanese temperate area all the isolates evinced diver-gent: in fact, they are separated by as many as 15 muta-tional steps, having 14 polymorphic sites with 13substitutions (4 transitions, 9 transversions) and 1 indel.
Sequence divergence within A. catenella isolates
No genetic diversity was observed in the Mediterraneanisolates (Table 4); indeed, the 20 isolates of A. catenellasequenced in this study were identical. At the same time,within Japanese isolates, a certain degree of nucleotidediversity was observed. Considering the sequencedivergence among the three different geographicalgroups, even though only one Chilean isolate sequencewas available, it was evident that the A. catenella isolatesof both Mediterranean and Japanese origin revealedmuch less differentiations; in contrast, Mediterraneanand Japanese A. catenella isolates were separated fromthe Chilean isolate by a higher net number of nucleotidedifferences. The higher value of the molecular diversityindex observed when comparing Japanese isolates thanthat found for Mediterranean isolates is due to highervariability in the Japanese population compared to thatof the Mediterranean population, the Japanese popula-tion having an isolate sequence designated M17, whichresults almost identical to the Mediterranean isolate se-quences (only three mutational steps).
Discussion and conclusions
The 5.8S gene rDNA and ITS regions were useful inexamining the relationships within the A. catenella spe-
Mediterranean
n=20
haplotype
Fig. 3 Statistical parsimony network of haplotypes of A. catenellaisolates based on the 5.8S rDNA and ITS region sequences. Thesize of the circles is proportional to the number of isolates foundhaving that haplotype. Small open circles indicate missing haplo-types
Table 4 Molecular diversity indexes in the 5.8S rDNA and ITSregions among and within isolates of A. catenella
Species MediterraneanSea
W Pacific/Japan
E Pacific/Chile
A. catenella(Mediterranean Sea)(n=20)
0.00
A. catenella(W Pacific, Japan)(n=7)
0.03% 0.05%
A. catenella(E Pacific, Chile)(n=1)
1.16% 1.23% nd
Data are expressed as mean l± SD (Standard Deviation (calcu-lated on 10.000 bootstrap replicates); n, number of isolates; nd, notdetermined) pair-wise nucleotide sequence differences within iso-lates (on the diagonal), net number of differences between isolates(above the diagonal)
19

cies and in tracing phylogenetic differentiations at orbelow the species level. These spacers are the most rap-idly changing sequences in the cistron because of theirrelative lack of functional constraints (Kooistra et al.2001). Different levels of polymorphism among speciesor individuals can result from sexual reproduction, re-distribution of the rDNA cistrons in the genome andploidy levels of the vegetative life stage (Wendel et al.1995); further, the variability in the sequence traits canresult from hybridization between genetically distinctpopulations, either by mixing of isolated populations orby gene flow.
In this study, the phylogenetic analysis within thegenus Alexandrium, based on 5.8S rDNA and ITS re-gion variability, generally agrees with the findings ofprevious studies in which different ribosomal geneticmarkers of SSU and LSU rRNA were used as well(Scholin et al. 1995; Adachi et al. 1996; John et al. 2003;MacKenzie et al. 2004), and clearly confirms A. catenellaas a sister group of A. tamarense. This result is sup-ported both by the topological agreement among thedifferent phylogenetic methods used and the high boot-strap and posterior probability values at the branchinternal node that separate the two clades. Moreover, itis interesting to note that in the A. catenella group, allMediterranean isolates share the same ITS-5.8S rDNAsequence (100% homology) and cluster together withthe different isolates of the Japanese temperate areas.This grouping is very well supported by both bootstrapand posterior probability values to 100%. The totalalignment of the ITS1 and ITS2 regions among differentmicroalgal isolates is usually neglected, since greatervariability can be found in the ITS2 region (Orsini et al.2004; Rodriguez-Lanetty et al. 2004).
Additionally, the phylogenetic analysis of the ITS -5.8S rDNA gene of the genus Alexandrium is in agree-ment with the distinct geographic clades (NA, TA, WEand ME clades) of the ‘A. tamarense/catenella/fundyensespecies complex’ already described using other molecu-lar markers in previous reports (Scholin et al. 1994;Medlin et al. 1998; John et al. 2003; Ruiz Sebastian et al.2005). In fact, in our analyses, the ACC01 isolate fromChile, morphologically attributed to the A. catenellaspecies, falls into the A. tamarense group. This is sup-ported by high bootstrap and posterior probability val-ues. Also, the position of the A. tamarense CU15, an ITSrDNA Thai type (Adachi et al. 1996), diverged from the‘A. tamarense/catenella/fundyense species complex’ andgrouped together with A. tamiyavanichii MS-01. This isin agreement with several phylogenetic studies on thegenus Alexandrium, such as those carried out by Usupet al. (2002), John et al. (2003) and Ruiz Sebastian et al.(2005) and further morphological studies should becarried out on the A. tamarense strain CU15.
In this study, a phylogenetic network among differenthaplotypes of A. catenella from Mediterranean andother world areas has been inferred. This phylogeneticnetwork is based on the statistical parsimony methodthat joins all pairs of haplotypes having a probability of
parsimony greater that 0.95 and is widely used in thecontext of phylogeography (Templeton et al. 1992;Vernesi et al. 2002). All the Mediterranean isolatesshared the same sequence, which is very similar to thatof a Japanese isolate (isolate M17) being separated byonly three substitutions. The most relevant aspect is thatin temperate Japanese isolates, seven different sequencesbelonging to seven isolates respectively, are present, witha mean pair-wise difference of 0.05%; whereas, in theNW Mediterranean Sea isolates, only one nucleotidesequence of ITS-5.8S rDNA is found. In the temperateJapanese area, it is possible to find that isolates sampledin the same locality (Tanabe Bay), such as TNX22 andTNY11, display sequences that are separated by 11mutation steps (Fig. 3). On the contrary, all the NWMediterranean A. catenella isolates, although from dis-tant areas, such as Tarragona and Olbia, share the sameITS-5.8S rDNA sequence.
The monomorphism of the A. catenella isolates in theNW Mediterranean Sea does not seem to be due to thesampling strategy, since the NW Mediterranean group ismade up of a much higher number of strains (more thandouble) when compared to the Japanese group. Therecovery of identical Mediterranean A. catenella se-quences embedded within the Japanese genetic vari-ability could suggest that the Mediterranean isolatesresulted from a founder originating from TemperateAsia.
The statistical parsimony analysis seems to indicate aclose relationship between the single Mediterraneanhaplotype and Asian haplotypes. Thus, it is possible thatthe founder of the Mediterranean A. catenella group islocated in the temperate Asian areas. This finding can beconsidered a quite recent phenomenon. It seems that themutation process has not had sufficient time to introducenew variants despite the fact that the ITS-5.8S rDNAmutational rate is rather fast (Mandal 1984), generatingnew haplotypes in a relatively short period of time.Moreover, from the time the Mediterranean group wasestablished, no hybridization between local and im-ported genotypes has occurred.
Although the nucleotide ITS-5.8S rDNA sequencesof A. catenella found in the Mediterranean are identical,this does not imply that these strains have homogenousphysiological characteristics. Recently, Figueroa et al.(2005) examined some clonal strains used in this study(IEO-AC1C1, IEO-AC5T, IEO-AC6T, IEO-AC9T),investigating the vegetative growth rate, mating type andseveral aspects of sexual reproduction. Notable differ-ences in vegetative growth rate, cyst production, ex-cystment success and viability were found.
Our findings on the phylogenetic relationships be-tween several Mediterranean A. catenella isolates andthe Japanese isolates are in agreement with the study byLilly et al. (2002), in which only two A. catenella strainswere analysed with the ML method based on the D1–D2domains of LSU rDNA. To reinforce the hypothesis ofthe introduction of this species into the Mediterranean,the retrieving of historical data regarding the occurrence
20

of A. catenella is discussed in this study (Table 5). Al-though many phytoplankton studies have been carriedout in the Mediterranean Sea, there is a lack of infor-mation on the presence of A. catenella before 1983. Thenthere was a lack of information until 1994, when anexpansion of this species, affecting several localities inthe NW Mediterranean basin, such as the ValenciaHarbour (since 1994), several Catalan harbours (since1996), Thau Lagoon (1998) and Olbia Bay (1999) hasbeen documented. At the same time, there is only onereference to this species in the Eastern Mediterraneanbasin: the eutrophic Alexandria Harbour in Egypt(Mikhail 2001). The Mediterranean is a confined seacommunicating with the Atlantic Ocean by the GibraltarStrait, and more recently, to the Red Sea by the artificialSuez Canal (1869). The introduction of A. catenella fromAsia by natural ways is unlikely because of the greatgeographic boundaries, while man-assisted activitiesappear to present a more realistic cause. Ship ballastwaters are known to transport viable cysts of phyto-plankton species (Hallegraeff 1998) and the harboursaffected by this species (Barcelona, Tarragona, Valenciaand Olbia) are characterized by high commercial ship-ping traffic.
In addition, aquaculture activities are present inValencia Harbour, Olbia Bay and Thau Lagoon. Thetransfer of mollusc stocks is known to be another vectorof alien phytoplankton species introduction (Laabir andGentien 1999). Once the A. catenella species has beenintroduced, especially in closed areas such as harbours,bays or lagoons, sediments become full of cysts. Thiscould be the case in Tarragona harbour, where numbersup to 4,000 cysts cm�3 of wet sediment were observed (I.Bravo, personal communication). Thus, when the spe-cies has been successfully established, natural oranthropogenic ways of dispersion could facilitate theexpansion to other areas (Maso et al. 2003).
In conclusion, it seems that A. catenella was probablyintroduced into the Western Mediterranean from tem-perate Asia, and based on the data available until now,from Japanese waters via anthropogenic pathways. Thiscan be supported by either the high genetic similaritybetween the Mediterranean isolates and a Japanesestrain, as shown by the phylogenetic analyses based onthe ITS-5.8S rDNA gene and the absence of A. catenellabefore 1983 in historical data followed by its detection indifferent sites in the Western basin of the Mediterranean.Some possible mechanisms may explain this new geo-
Table 5 Historical references of A. catenella occurrences in the Mediterranean Sea [win, winter; spr, spring; sum, summer; aut, autumn]
Year State and sites Occurence period References
1983 Spain: between Catalan coast and Balearic Islands Sum Margalef and Estrada (1987)1994 Spain: Valencia harbour Spr Gomis et al. (1996);
M. Delgado (personal communication)1995 France: Thau lagoon Aut – win Abadie et al. (1999): as A. tamarense1996 Spain: Barcelona harbour (Catalan coast) Sum Vila et al. (2001a)1997 Spain: Catalan coast (different harbours)
Tunis: Canal de TunisSumSum
Vila et al. (2001b)Souad Turki (personal communication)
1998 Egypt: Alexandria (eastern harbour) Mikhail (2001)France: Thau lagoon Aut Abadie et al. (1999): as A. tamarense
Lilly et al. (2002)Spain: Catalan coast (different harbours) Spr – sum – aut Vila et al. (2001b)Catalan coast (open coastal waters) Sum
1999 Egypt: Alexandria (eastern harbour) Mikhail (2001)Italy: Olbia Habour (Sardinia) Sum Luglie et al. (2003b)Spain: Catalan coast (harbours) Spr – sum – aut Vila et al. (2001b)Catalan coast (open coastal waters) Sum Vila et al. (2001c)
2000 Egypt: Alexandria (eastern harbour) Mikhail (2001)Italy: Olbia harbour (Sardinia) Sum A. Luglie and M. G. Giacobbe
(personal communication)Spain: Catalan coast (different harbours) Spr – sum – aut Sampedro et al. (2003)
2001 Italy: Olbia harbour (Sardinia) Sum Luglie et al. (2003a)Spain: Catalan coast (harbours) Spr – sum – aut Sampedro et al. (2003)Andaluzian coast Aut Fernandez et al. (2004)
2002 Italy: Olbia harbour (Sardinia)Spain: Catalan coast (different harbours)
Spr – sumSpr – sum – aut
Luglie et al. (2003a)Sampedro et al. (2003)
2002 Greece: Greek coast Spr – sum Gotsis-Skretas (personal communication):as Alexandrium sp.
2003 Algery: Argel harbour (Argelia)Italy: Olbia harbour (Sardinia)Spain: Catalan coast (different harbours)
SumSpr – sum – autWin – spr – sum – aut
Illoul (personal communication)A. Luglie (personal communication)HAEDAT database
2004 Spain: Catalan coast (different harbours) Win – spr – sum – ?? HAEDAT database
The regular monitoring of toxic species in the Catalan coast began in 1989, in Sardinia in 1992 and Sicily in 2001. During the STRATEGYEU Project (2001–2004) extensive monitoring was done in the Mediterranean: Catalan-Balear Sea, Sardinia, Sicily and Greece [N, 46stations] sampled weekly from May to October. [HAEDAT Centro Cientifico y de Comunicacion sobre Algas Nocivas, COI-IEO, Spain]
21

graphic expansion of A. catenella on a regional scaleobserved in Mediterranean coastal areas. These mecha-nisms may be due to the natural population dispersal orviable cyst dissemination in new areas and/or theincreasing number of coastal and harbour sites, char-acterized by high level of nutrients and low turbulence,which may favour dinoflagellate presence (Garces et al.2000; Vila and Maso 2005).
In any case, whether these phenomena are the resultof the introduction of new species or due to emergenthidden flora, remains an open debate in studies onharmful algal species.
Acknowledgements This study was supported by the EU-FP5Strategy Project EVK3-CT- 00046-2001 and CCVIEO. Specialthanks to Dr. Mercedes Maso, Prof. Mauro Magnani and Dr.Francesca Andreoni for technical assistance in sequence analysesand to Jose M. Franco and CRM (Centro Ricerche Marine, Ce-senatico, Italy) for providing toxin information and also to Ko-stantinos Koukaras for supplying a culture strain.
References
Abadie E, Amzil Z, Belin C, Comps MA, Elziere-Papayanni P,Lassus P, LeBec C, Marcaillou-Le Baut C, Nezan E, Poggi R(1999) Contamination de l’etang de Thau par Alexandriumtamarense. Episode de novembre a decembre 1998. ReportObservation et surveillance de la mer cotiere. Ifremer, Pluzane,p 44
Adachi M, Sako Y, Ishida Y (1994) Restriction fragment lengthpolymorphism of ribosomal DNA internal transcribed spacerand 5.8S regions in Japanese Alexandrium species (Dinophy-ceae). J Phycol 30:857–865
Adachi M, Sako Y, Ishida Y (1996) Analyses of Alexandrium(Dinophyceae) species using sequences of the 5.8S ribosomalDNA and internal transcribed spacer regions. J Phycol 32:424–432
Balech E (1995) The Genus Alexandrium Halim (Dinoflagellata).Sherkin Island Marine Station, Sherkin Island Co., Cork, Ire-land
Benavides H, Prado L, Diaz S, Carreto Jj (1995) An exceptionalbloom of Alexandrium catenella in the Beagle Channel,Argentina. In: Lassus P, Arzul G, Erard-Le Denn E, Gentien P,Marcaillou-Le Baut C (eds) Harmful marine algal blooms.Proliferation D’algues Marines Nuisibles. Lavoisier, Paris, pp113–119
Clement M, Posada D, Crandall KA (2000) TCS: a computerprogram to estimate gene genealogies. Mol Ecol 9:1657–1659
Cordova JL, Muller I (2002) Use of PCR and partial sequencing ofthe large-subunit rRNA gene to identify Alexandrium catenella(Dinophyceae) from the South of Chile. Harmful Algae 1:343–350
Fernandez R, Maman L, Jaen D, Marquez I (2004) Control yseguimiento del fitoplancton toxico en las costas anadaluzasdurante los anos 2001 y 2002. In: Norte M, Fernandez R (eds)VII Reunion Iberica sobre Fitoplancton Toxico y biotoxinas.Instituto Universitario de Biorganica de la Universidad de laLaguna. La Laguna, pp 99–107
Figueroa RI, Bravo I, Garces E (2005) Effects of nutritional factorsand different parental crosses on the encystment and excyst-ment of Alexandrium catenella in culture. Phycologia (in press)
Garces E, Maso M, Vila M, Camp J (2000) HABs events in theMediterranean Sea: are they increasing? A case study of the lastdecade in the NW Mediterranean and the genus Alexandrium.Harmful Algal News 20:1–11
Gomis C, Alcober J, Bernabeu A (1996) Seguimiento de las pob-laciones fitoplanctonicas en las bateas mrjilloneras del Puerto
de Valencia 1991–1994. In: Matamoros E, Delgado M (eds) IVReunion Iberica de fitoplancton toxico y biotoxinas St. Carlesde la Rapita (Tarragona): Generalitat de Catalunya. Depart-ment d’Agricultura, Ramaderia i Pesca, pp 29–38
Guillard RRL (1975) Culture of phytoplankton for feeding marineinvertebrates. In: Smith WL, Chanley MH (eds) Culture ofmarine invertebrate animals. Plenum Press, NewYork, pp 26–60
Guillard RRL, Hargraves PE (1993) Stichochrysis inmobilis is adiatom, not a chrysophyte. Phycologia 32:234–236
Guillou L, Nezan E, Cueff V, Erard-Le Denn E, Cambon-BonavitaMA, Gentien P, Barbier G (2002) Genetic diversity andmolecular detection of three toxic dinoflagellate genera (Alex-andrium, Dinopysis and Karenia) from the French Coasts.Protist 153:223–238
Hallegraeff GM, Bolch CJ, Blackburn SI, Oshima Y (1991) Speciesof toxigenic dinoflagellate genus Alexandrium in south-easternAustralian waters. Bot Mar 34:575–587
Hallegraeff GM (1998) Transport of toxic dinoflagellates via ships’ballast water: bioeconomic risk assessment and efficacy ofpossible ballast water management strategies. Mar Ecol ProgrSer 168:297–309
Higman WA, Stone DM, Lewis JM (2001) Sequence comparisonsof toxic and non-toxic Alexandrium tamarense (Dinophyceae)isolates from UK waters. Phycologia 40:256–262
Huelsenbeck JP, Ronquist F (2001) MrBAYES: Bayesian inferenceof phylogenetic trees. Bioinformatics 17:754–755
John U, Fensome R, Medlin LK (2003) The application of amolecular clock based on molecular sequences and the fossilrecord to explain biogeographic distribution within the Alex-andrium tamarense ‘‘Species Complex’’ (Dinophyceae). MolBiol Evol 20:1015–1027
Karplus K, Barrett C, Hughey R (1998) Hidden markov models fordetecting remote protein homologies. Bioinformatics 14:846–856
Keller MD, Selvin RC, Claus W, Guillard RRL (1987) Media forthe culture of oceanic ultraphytoplankton. J Phycol 23:633–638
Kooistra WHCF, de Boer MK, Vrieling EG, Connell LB, GieskesWC (2001) Variation along the ITS markers across strains ofFibrocapsa japonica (Raphidophyceae) suggests hybridizationevents and recent range expansion. J Sea Res 46:213–222
Kumar S, Tamura K, Jacobsen IB, Nei M (2001) MEGA 2:Molecular Evolutionary Genetics Analysis Software. ArizonaState University, Tempe, AZ, USA
Laabir M, Gentien P (1999) Survival of toxic dinoflagellates aftergut passage in the Pacific oyster Crassostrea gigas Thunburg.J Shell Res 18:217–222
Lilly EL, Kulis DM, Gentien P, Anderson DM (2002) Paralyticshellfish poisoning toxins in France linked to a human-intro-duced strain of Alexandrium catenella from western Pacific:evidence from DNA and toxin analysis. J Plank Res 24:443–452
Luglie A, Giacobbe MG, Sannio A, Fiocca F, Sechi N (2003b)First record of the dinoflagellate Alexandrium catenella (Whe-don & Kofoid) Balech (Dinophyta), a potential producer ofparalytic shellfish poisoning, in Italian waters (Sardinia, Tyr-rhenian Sea). Bocconea 16:1045–1052
Luglie A, Giacobbe MG, Sannio A, Fiocca F, Sechi N (2003a) Thegeographical distribution of Alexandrium catenella is extendingto Italy! First evidences from the Tyrrhenian Sea. In: Pro-ceeding of Xth international conference on Harmful Algae (inpress)
MacKenzie L, de Salas M, Adamson J, Beuzenberg V (2004) Thedinoflagellate genus Alexandrium (Halim) in New Zealandcoastal waters: comparative morphology, toxicity and molecu-lar genetics. Harmful Algae 3:71–92
Mandal RK (1984) The organisation and transcription of eukary-otic ribosomal RNA genes. Progr Nucl Acid Res Mol Biol31:115–158
Manhart JR, McCourt RM (1992) Molecular data and speciesconcepts in the algae. J Phycol 28:730–737
Margalef R, Estrada M (1987) Synoptic distribution of summermicroplankton (Algae and Protozoa) across the principal frontin the Western Mediterranean. Investigaciones Pesqueria51:121–140
22

Maso M, Garces E, Pages F, Camp J (2003) Drifting plastic debrisas a potential vector for dispersing Harmful Algal Bloom(HAB) species. Sci Mar 67:107–111
Masselin P, Amzil Z, Abadie E, Nezan E, Le Bec C, Carreras A,Chiantella C, Truquet P (2001) Paralytic shellfish poisoning onthe French Mediterranean coast in autumn 1998: Alexandrium‘‘tamarense complex’’ (Dinophyceae) as causative agent. In:Hallegraeff GM, Blackburn SI, Bolch CJ, Lewis RJ (eds)Harmful Algal blooms 2000. Proceedings of IXth internationalconference Harmful Algal Blooms, UNESCO, Paris, pp 26–29
Medlin LK, Lange M, Wellbrock U, Donner G, Elbrachter M,Hummert C, Luckas B (1998) Sequence comparison link toxicEuropean isolates of Alexandrium tamarense from the OrkneyIslands to toxic North American stocks. Eur J Protistol 34:329–335
Mikhail SK (2001) Toxic red tide species are on rise in Alexandriawaters (Egypt). Harmful Algae News 22:5
Orsini L, Procaccini G, Sarno D, Montresor M (2004) MultiplerDNA ITS-type within the diatom Pseudo-nitzschia delicatissima(Bacillariophyceae) and their relative abundance across a springbloom in the Gulf of Naples. Mar Ecol Progr Ser 271:87–98
Penna A, Magnani M (1999) Identification of Alexandrium (Din-ophyceae) species using PCR and rDNA-targeted probes. JPhycol 35:615–621
Posada D, Crandall KA (1998) Modeltest: testing the model ofDNA substitution. Bioinformatics 14:817–818
Rodriguez-Lanetty M, Krupp DA, Weis VM (2004) Distinct ITStypes of Symbiodinium in Clade C correlate with cnidarian/dinoflagellate specificity during onset of symbiosis. Mar EcolProgr Ser 275:97–102
Ruiz Sebastian C, Etheridge SM, Cook PA, O’Ryan C, Pitcher GC(2005) Phylogenetic analysis of toxic Alexandrium (Dinophy-ceae) isolates from South Africa: implications for the globalphylogeography of the Alexandrium tamarense species complex.Phycologia (in press)
Sampedro N, Vila M, Arın L, Garces E, Camp J (2003) Seguimi-ento del fitoplancton toxico en la costa catalana: incidenciasdurante 2000–2002. In: VIII Reunion Iberica de fitoplanctonToxico y Biotoxinas. La Laguna, Tenerife, pp 29–38
Saunders GW, Hill DRA, Sexton JP, Andersen RA (1997) Small-subunit ribosomal RNA sequences from selected dinoflagel-lates: testing classical evolutionary hypotheses with molecularsystematic methods. Plant System Evol 11:237–259
Scholin CA, Anderson DM (1994) Identification of group andstrain-specific genetic markers for globally distributed Alex-andrium (Dinophyceae). I RFLP analysis of SSUrRNA genes. JPhycol 30:744–754
Scholin CA, Herzog M, Sogin M, Anderson DM (1994) Identifi-cation of group-and strain — specific genetic markers forglobally distributed Alexandrium (Dinophyceae). 2. Sequence
analysis of a fragment of the LSU rRNA gene. J Phycol 30:999–1011
Scholin CA, Hallegraeff GM, Anderson DM (1995) Molecularevolution of the Alexandrium tamarense ‘‘species complex’’(Dinophyceae): dispersal in the North American and WestPacific regions. Phycologia 34:472–485
Smayda TJ (1997) What is a bloom? A commentary. LimnolOceanogr 42:1132–1136
Sournia A (1995) Red-tide and toxic marine phytoplankton of theworld ocean: an inquiry into biodiversity Harmful Toxic AlgalBlooms. Lavosier, Paris, pp 239–243
Swofford DL (2000) PAUP*: phylogenetic analysis using parsi-mony (* and other methods), version. 4.0b. Sinauer Associates,Sunderland
Tamura K, Nei M (1993) Estimation of the number of nucleotidesubstitutions in the control region of mitochondrial DNA inhumans and chimpanzees. Mol Biol Evol 10:512–526
Templeton AR, Crandall KA, Sing CF (1992) A cladistic analysisof phenotypic associations with haplotypes inferred fromrestriction endonuclease mapping and DNA sequence data. III.Cladogram estimation. Genetics 132:619–633
Usup G, Pin LC, Ahmad A, Teen LP (2002) Phylogenetic rela-tionship of Alexandrium tamiyavanichii (Dinophyceae) to otherAlexandrium species based on ribosomal RNA gene sequences.Harmful Algae 1:59–68
Vernesi C, Pecchioli E, Caramelli D, Tiedemann R, Randi E,Bertorelle G (2002) The genetic structure of natural and rein-troduced roe deer (Capreolus capreolus) populations in the Alpsand central Italy, with reference to the mitochondrial DNAphylogeography of Europe. Mol Ecol 11:1285–1297
Yeung PKK, Wong FTW, Wong JTY (2002) Large subunit rDNAsequences from Alexandrium catenella strains isolated duringalgal blooms in Hong Kong. J Appl Phycol 14:147–150
Vila M, Camp J, Garces E, Maso M, Delgado M (2001a) Highresolution spatio-temporal detection of HABs in confined wa-ters of the NW Mediterranean. J Plank Res 23:497–514
Vila M, Garces E, Maso M, Camp J (2001b) Is the distribution ofthe toxic dinoflagellate Alexandrium catenella expanding alongthe NW Mediterranean coast? Mar Ecol Progr Ser 222:73–83
Vila M, Delgado M, Camp J (2001c) First detection of widespreadtoxic events caused by Alexandrium catenella in the Mediter-ranean Sea. In: Hallegraeff GM, Blackburn SI, Bolch CJ, LewisRJ (eds) Harmful Agal Blooms (2000) Proceedings of IXthInternational Harmful Algal Blooms. UNESCO, Paris, pp 8–11
Vila M, Maso M (2005) Phytoplankton functional groups andharmful algal species in antropogenically impacted waters of theNW Mediterranean Sea. Sci Mar 69:31–45
Wendel JF, Schnabel A, Seelan T (1995) Bidirectional interlocusconcerted evolution following allopolypoid speciation in cotton(Gossypium). Proc Natl Acad Sci USA 92:280–284
23
Related Documents











