DOI: 10.1126/scitranslmed.3009539 , 251ra118 (2014); 6 Sci Transl Med et al. Vanessa O. Zambelli acute inflammatory pain Aldehyde dehydrogenase-2 regulates nociception in rodent models of Editor's Summary enzyme. sensitivity to pain reported in East Asian people, many of whom carry a mutation in this same aldehyde-degrading possibly without the addiction that plagues opiate pain killers. In addition, these results may explain the greater the magnitude of the pain response. New drugs that modulate aldehyde levels could prove beneficial for patients, effect. Indeed, the authors found a tight correlation between the local concentration of aldehydes at the injury site and rodents' susceptibility to a painful stimulus. Conversely, activating the enzyme with the drug Alda-1 reverses this markedly increases -- a known pain-causing molecule -- inactivating mutation of the enzyme that degrades aldehydes find that an et al. Pain is a seriously undertreated condition, and new drugs are sorely needed. Now, Zambelli New Path to Pain Control http://stm.sciencemag.org/content/6/251/251ra118.full.html can be found at: and other services, including high-resolution figures, A complete electronic version of this article http://stm.sciencemag.org/content/suppl/2014/08/25/6.251.251ra118.DC1.html can be found in the online version of this article at: Supplementary Material http://stm.sciencemag.org/content/scitransmed/4/159/159ec200.full.html http://stm.sciencemag.org/content/scitransmed/3/90/90ra60.full.html http://stm.sciencemag.org/content/scitransmed/3/101/101ra91.full.html http://stm.sciencemag.org/content/scitransmed/4/143/143ra100.full.html http://stm.sciencemag.org/content/scitransmed/5/183/183ra56.full.html can be found online at: Related Resources for this article http://www.sciencemag.org/about/permissions.dtl in whole or in part can be found at: article permission to reproduce this of this article or about obtaining reprints Information about obtaining is a registered trademark of AAAS. Science Translational Medicine rights reserved. The title NW, Washington, DC 20005. Copyright 2014 by the American Association for the Advancement of Science; all last week in December, by the American Association for the Advancement of Science, 1200 New York Avenue (print ISSN 1946-6234; online ISSN 1946-6242) is published weekly, except the Science Translational Medicine on August 29, 2014 stm.sciencemag.org Downloaded from on August 29, 2014 stm.sciencemag.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DOI: 10.1126/scitranslmed.3009539, 251ra118 (2014);6 Sci Transl Med
et al.Vanessa O. Zambelliacute inflammatory painAldehyde dehydrogenase-2 regulates nociception in rodent models of
Editor's Summary
enzyme.sensitivity to pain reported in East Asian people, many of whom carry a mutation in this same aldehyde-degrading possibly without the addiction that plagues opiate pain killers. In addition, these results may explain the greaterthe magnitude of the pain response. New drugs that modulate aldehyde levels could prove beneficial for patients, effect. Indeed, the authors found a tight correlation between the local concentration of aldehydes at the injury site androdents' susceptibility to a painful stimulus. Conversely, activating the enzyme with the drug Alda-1 reverses this
markedly increases−−a known pain-causing molecule−−inactivating mutation of the enzyme that degrades aldehydes find that an et al.Pain is a seriously undertreated condition, and new drugs are sorely needed. Now, Zambelli
New Path to Pain Control
http://stm.sciencemag.org/content/6/251/251ra118.full.htmlcan be found at:
and other services, including high-resolution figures,A complete electronic version of this article
http://stm.sciencemag.org/content/suppl/2014/08/25/6.251.251ra118.DC1.html can be found in the online version of this article at: Supplementary Material
http://stm.sciencemag.org/content/scitransmed/4/159/159ec200.full.html http://stm.sciencemag.org/content/scitransmed/3/90/90ra60.full.html
http://stm.sciencemag.org/content/scitransmed/3/101/101ra91.full.html http://stm.sciencemag.org/content/scitransmed/4/143/143ra100.full.html
http://stm.sciencemag.org/content/scitransmed/5/183/183ra56.full.html can be found online at:Related Resources for this article
http://www.sciencemag.org/about/permissions.dtl in whole or in part can be found at: article
permission to reproduce this of this article or about obtaining reprintsInformation about obtaining
is a registered trademark of AAAS. Science Translational Medicinerights reserved. The title NW, Washington, DC 20005. Copyright 2014 by the American Association for the Advancement of Science; alllast week in December, by the American Association for the Advancement of Science, 1200 New York Avenue
(print ISSN 1946-6234; online ISSN 1946-6242) is published weekly, except theScience Translational Medicine
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
o
n A
ugus
t 29,
201
4st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om

R E S EARCH ART I C L E
PA IN
Aldehyde dehydrogenase-2 regulates nociception inrodent models of acute inflammatory painVanessa O. Zambelli,1,2* Eric R. Gross,1,3* Che-Hong Chen,1 Vanessa P. Gutierrez,2
Yara Cury,2 Daria Mochly-Rosen1†
ugus
t 29,
201
4
Exogenous aldehydes can cause pain in animal models, suggesting that aldehyde dehydrogenase-2 (ALDH2),which metabolizes many aldehydes, may regulate nociception. To test this hypothesis, we generated a knock-inmouse with an inactivating point mutation in ALDH2 (ALDH2*2), which is also present in human ALDH2 of ~540million East Asians. The ALDH2*1/*2 heterozygotic mice exhibited a larger response to painful stimuli thantheir wild-type littermates, and this heightened nociception was inhibited by an ALDH2-selective activator(Alda-1). No effect on inflammation per se was observed. Using a rat model, we then showed that nociceptiontightly correlated with ALDH activity (R2 = 0.90) and that reduced nociception was associated with less earlygrowth response protein 1 (EGR1) in the spinal cord and less reactive aldehyde accumulation at the insult site(including acetaldehyde and 4-hydroxynonenal). Further, acetaldehyde- and formalin-induced nociceptive be-havior was greater in the ALDH2*1/*2 mice than in the wild-type mice. Finally, Alda-1 treatment was evenbeneficial when given after the inflammatory agent was administered. Our data in rodent models suggest thatthe mitochondrial enzyme ALDH2 regulates nociception and could serve as a molecular target for pain control,with ALDH2 activators, such as Alda-1, as potential non-narcotic, cardiac-safe analgesics. Furthermore, ourresults suggest a possible genetic basis for East Asians’ apparent lower pain tolerance.
A
onst
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
INTRODUCTION
Pain is an international health problem, affecting about one in everyfive individuals (1). About 200 million opioid prescriptions are writtenannually in the United States, and in 2013, vicodin was the overallnumber one prescribed medication (2). According to the NationalCenter for Health Statistics data recently released for 2010, opioiddrugs caused 75% of all drug-induced deaths and were responsiblefor 16,651 fatalities (3). Secondary health complications, includingopioid abuse and dependence, afflict 2.1 million people (4). In addi-tion, nonsteroidal anti-inflammatory pain medications (NSAIDs) areused by over 30 million people each day in the United States foranalgesia (5). Yet, NSAIDs and cyclooxygenase-2 (COX-2) inhibitorsalso increase the risk of gastrointestinal bleeding and cardiac events,and their safety has been questioned for certain patient populations,such as those with cardiac disease (6, 7). Thus, new molecular targetsthat regulate pain are needed to develop therapeutics for pain controlwith fewer deleterious addictive and cardiovascular effects.
Reactive aldehydes, including 4-hydroxynonenal (4-HNE), formal-dehyde, and acetaldehyde, cause pain when directly applied in rodents(8–10). We therefore determined whether altering the enzymatic ac-tivity of the mitochondrial aldehyde dehydrogenase-2 (ALDH2), whichcatalyzes removal of these reactive aldehydes, alters pain responses. Wewere also interested in this question because a common inactivatingpoint mutation in mitochondrial ALDH2 (Glu487 to Lys487), found in36% of Han Chinese, affects about 8% of the world population (11).The ALDH2*2 in the Han Chinese codes for a dominant-negative var-iant reducing ALDH2 enzymatic activity by ~60 to 80% in heterozy-gotes (ALDH2*1/*2) and by ~95% in homozygotes (ALDH2*2/*2)
1Department of Chemical and Systems Biology, Stanford University School of Medicine,Stanford, CA 94305, USA. 2Laboratory of Pain and Signaling, Butantan Institute, São Paulo05503-900, Brazil. 3Department of Anesthesiology, Perioperative, and Pain Medicine,Stanford University School of Medicine, Stanford, CA 94305, USA.*These authors contributed equally to this study.†Corresponding author. E-mail: [email protected]
www.Scienc
compared to wild-type ALDH2*1/*1 (11). The ALDH2*2 variantcauses flushing after ethanol consumption, a result of acetaldehyde ac-cumulation (11). The ALDH2*2 inactivating mutation also causes re-duced metabolism of other reactive aldehydes, including malondialdehyde(MDA) and 4-HNE (12), and the rate of formaldehyde metabolism inhuman mitochondrial liver fractions from ALDH2*1/*2 subjects isabout three times slower than in those from ALDH2*1/*1 subjects (13).
Our laboratory has developed a small molecule that selectively en-hances the activity of ALDH2, Alda-1 [N-(1,3-benzodioxol-5-ylmethyl)-2,6-dichlorobenzamide] (14). Alda-1 corrects the structural defect inthe mutant ALDH2*2, thereby increasing ALDH2*2 activity (15). Here,we determined whether ALDH2 enzymatic activity modulates acuteinflammatory-induced hyperalgesia and whether the ALDH2 activatorAlda-1 could be a potential drug to reduce pain.
RESULTS
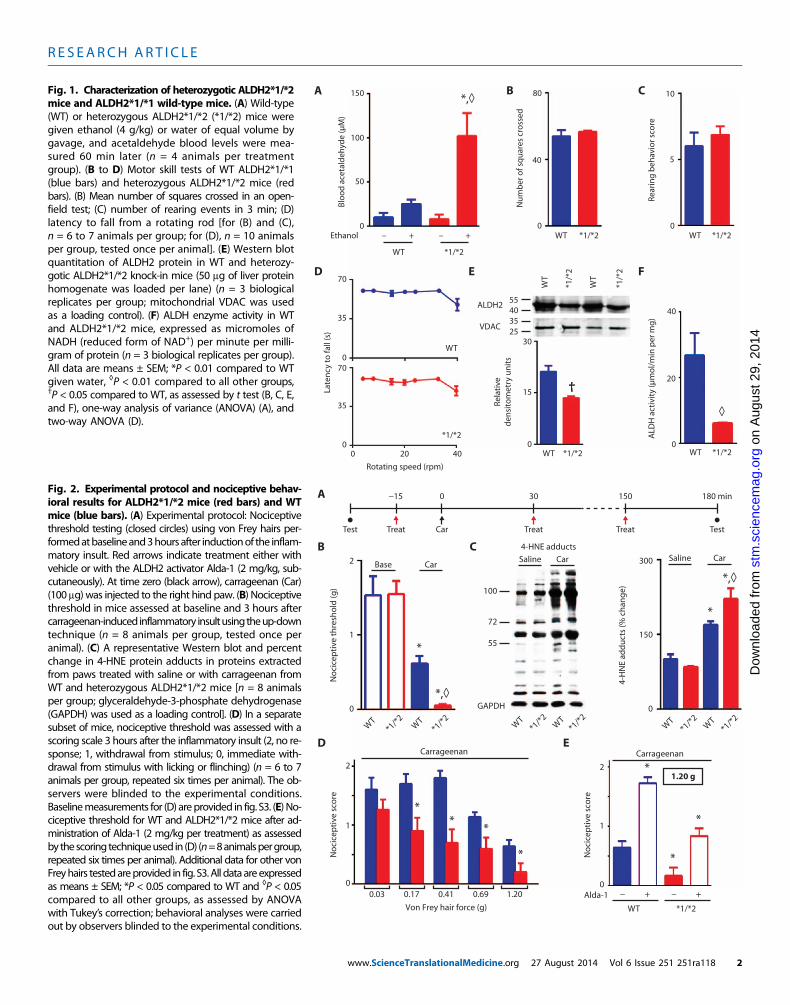
We generated knock-in mice carrying the inactivating Lys487 pointmutation in ALDH2, identical to the mutation found in Han Chinese[(11); denoted ALDH2*2] (fig. S1). To confirm that the mutant micemimic the human phenotype, we challenged them with ethanol anddetermined blood acetaldehyde levels. Similar to human heterozygotes(16), heterozygote mice accumulated five times higher blood acetaldehydeconcentrations than did wild-type ALDH2 (ALDH2*1/*1) mice (Fig. 1A).Before nociceptive testing, we performed behavioral tests to confirmthat the ALDH2 inactivating mutation did not affect mouse gross mo-tor skills. No differences were detected in the mean number of squarescrossed in an open field (Fig. 1B) or in rearing behavior (Fig. 1C) be-tween wild-type and ALDH2*1/*2 mice. Similarly, in a rotarod test, nodifferences were seen between wild-type and ALDH2*1/*2 mice (Fig.1D and fig. S2). When assessed in liver tissue homogenate, the abun-dance of ALDH2*1/*2 protein was about 44% lower than that in wild-type mice (Fig. 1E), and ALDH activity in the heterozygotic mice was
eTranslationalMedicine.org 27 August 2014 Vol 6 Issue 251 251ra118 1
R E S EARCH ART I C L E
on
Aug
ust 2
9, 2
014
ag.o
rg
A B C
D E F
Blo
od
ace
tald
ehyd
e (µ
M)
Nu
mb
er o
f sq
uar
es c
ross
ed
– + – +
WT *1/*2
Rear
ing
beh
avio
r sco
re
Late
ncy
to fa
ll (s
)
Ethanol WT *1/*2 *1/*2
*1/*2 *1/*2
WT
WT WT
WT
*1/*2
150
100
50
0
55403525
ALDH2
VDAC
30
15
0
WT
WT
*1/*
2
*1/*
2
80
40
0
10
5
0
70
35
070
35
00 20 40
Rela
tive
den
sito
met
ry u
nit
s
40
20
0
ALD
H a
ctiv
ity
(µm
ol/
min
per
mg
)
†
◊
*,◊
Rotating speed (rpm)
Fig. 1. Characterization of heterozygotic ALDH2*1/*2
mice and ALDH2*1/*1 wild-type mice. (A) Wild-type(WT) or heterozygous ALDH2*1/*2 (*1/*2) mice weregiven ethanol (4 g/kg) or water of equal volume bygavage, and acetaldehyde blood levels were mea-sured 60 min later (n = 4 animals per treatmentgroup). (B to D) Motor skill tests of WT ALDH2*1/*1(blue bars) and heterozygous ALDH2*1/*2 mice (redbars). (B) Mean number of squares crossed in an open-field test; (C) number of rearing events in 3 min; (D)latency to fall from a rotating rod [for (B) and (C),n = 6 to 7 animals per group; for (D), n = 10 animalsper group, tested once per animal]. (E) Western blotquantitation of ALDH2 protein in WT and heterozy-gotic ALDH2*1/*2 knock-in mice (50 mg of liver proteinhomogenate was loaded per lane) (n = 3 biologicalreplicates per group; mitochondrial VDAC was usedas a loading control). (F) ALDH enzyme activity in WTand ALDH2*1/*2 mice, expressed as micromoles ofNADH (reduced form of NAD+) per minute per milli-gram of protein (n = 3 biological replicates per group).All data are means ± SEM; *P < 0.01 compared to WTgiven water, ◊P < 0.01 compared to all other groups,†P < 0.05 compared to WT, as assessed by t test (B, C, E,and F), one-way analysis of variance (ANOVA) (A), andtwo-way ANOVA (D).stm
.sci
ence
mD
ownl
oade
d fr
om
A
B C
D E
−15 0 30 150 180 min
Test TestTreatTreat TreatCar
WT
*1/*2
*1/*2W
T
100
72
55
Saline Car
4-HNE adducts
1
2
0
No
cice
pti
ve t
hre
sho
ld (g
)
150
300
0
WT
*1/*2
*1/*2W
T
4-H
NE
add
uct
s (%
ch
ang
e)
0.03 0.17 0.41 0.69 1.20
1
2
0
CarBase
*,◊
*
*,◊
*
**
*
*
Carrageenan
No
cice
pti
ve s
core
1
2
0
No
cice
pti
ve s
core
*
*
*
Carrageenan
1.20 g
− + − +
WT *1/*2
Alda-1
Saline Car
Von Frey hair force (g)
WT
*1/*2
*1/*2W
T
GAPDH
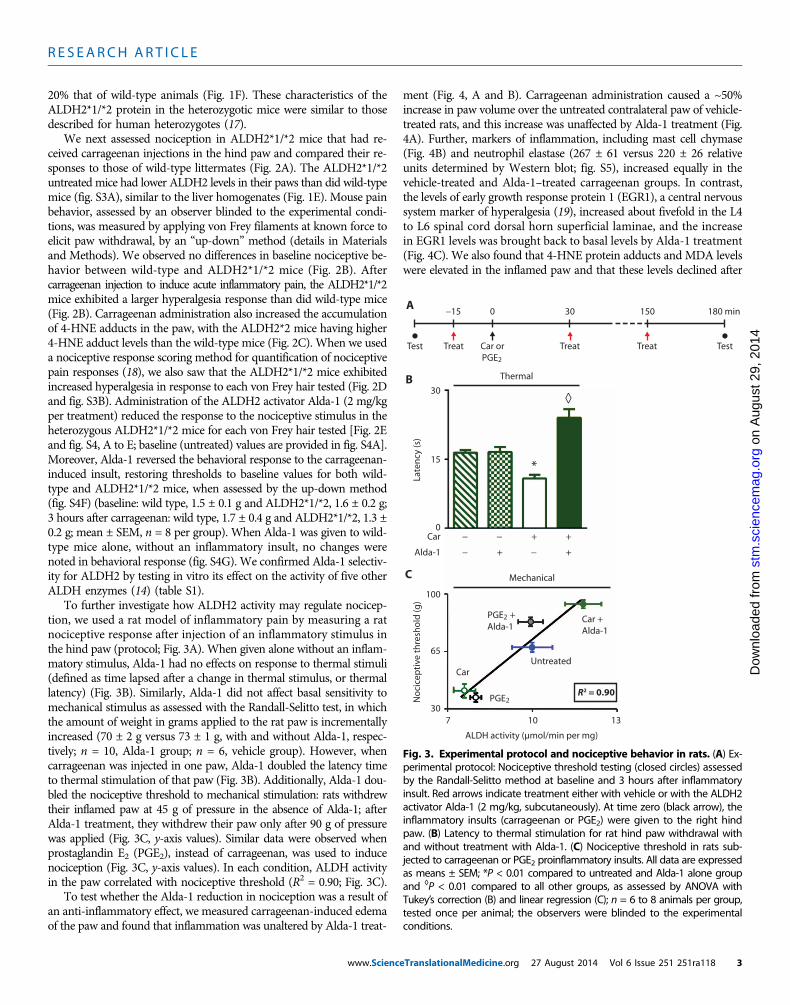
Fig. 2. Experimental protocol and nociceptive behav-ioral results for ALDH2*1/*2 mice (red bars) and WT
mice (blue bars). (A) Experimental protocol: Nociceptivethreshold testing (closed circles) using von Frey hairs per-formedatbaselineand3hoursafter inductionof the inflam-matory insult. Red arrows indicate treatment either withvehicle or with the ALDH2 activator Alda-1 (2 mg/kg, sub-cutaneously). At time zero (black arrow), carrageenan (Car)(100 mg) was injected to the right hind paw. (B) Nociceptivethreshold in mice assessed at baseline and 3 hours aftercarrageenan-inducedinflammatory insultusingtheup-downtechnique (n = 8 animals per group, tested once peranimal). (C) A representative Western blot and percentchange in 4-HNE protein adducts in proteins extractedfrom paws treated with saline or with carrageenan fromWT and heterozygous ALDH2*1/*2 mice [n = 8 animalsper group; glyceraldehyde-3-phosphate dehydrogenase(GAPDH) was used as a loading control]. (D) In a separatesubset of mice, nociceptive threshold was assessed with ascoring scale 3 hours after the inflammatory insult (2, no re-sponse; 1, withdrawal from stimulus; 0, immediate with-drawal from stimulus with licking or flinching) (n = 6 to 7animals per group, repeated six times per animal). The ob-servers were blinded to the experimental conditions.Baselinemeasurements for (D) areprovided in fig. S3. (E) No-ciceptive threshold for WT and ALDH2*1/*2 mice after ad-ministration of Alda-1 (2 mg/kg per treatment) as assessedby thescoringtechniqueused in (D) (n=8animalspergroup,repeated six times per animal). Additional data for other vonFreyhairs testedareprovided in fig. S3.All dataareexpressedas means ± SEM; *P < 0.05 compared to WT and ◊P < 0.05compared to all other groups, as assessed by ANOVAwith Tukey’s correction; behavioral analyses were carriedout by observers blinded to the experimental conditions.www.ScienceTranslationalMedicine.org 27 August 2014 Vol 6 Issue 251 251ra118 2
R E S EARCH ART I C L E
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
20% that of wild-type animals (Fig. 1F). These characteristics of theALDH2*1/*2 protein in the heterozygotic mice were similar to thosedescribed for human heterozygotes (17).
We next assessed nociception in ALDH2*1/*2 mice that had re-ceived carrageenan injections in the hind paw and compared their re-sponses to those of wild-type littermates (Fig. 2A). The ALDH2*1/*2untreated mice had lower ALDH2 levels in their paws than did wild-typemice (fig. S3A), similar to the liver homogenates (Fig. 1E). Mouse painbehavior, assessed by an observer blinded to the experimental condi-tions, was measured by applying von Frey filaments at known force toelicit paw withdrawal, by an “up-down” method (details in Materialsand Methods). We observed no differences in baseline nociceptive be-havior between wild-type and ALDH2*1/*2 mice (Fig. 2B). Aftercarrageenan injection to induce acute inflammatory pain, the ALDH2*1/*2mice exhibited a larger hyperalgesia response than did wild-type mice(Fig. 2B). Carrageenan administration also increased the accumulationof 4-HNE adducts in the paw, with the ALDH2*2 mice having higher4-HNE adduct levels than the wild-type mice (Fig. 2C). When we useda nociceptive response scoring method for quantification of nociceptivepain responses (18), we also saw that the ALDH2*1/*2 mice exhibitedincreased hyperalgesia in response to each von Frey hair tested (Fig. 2Dand fig. S3B). Administration of the ALDH2 activator Alda-1 (2 mg/kgper treatment) reduced the response to the nociceptive stimulus in theheterozygous ALDH2*1/*2 mice for each von Frey hair tested [Fig. 2Eand fig. S4, A to E; baseline (untreated) values are provided in fig. S4A].Moreover, Alda-1 reversed the behavioral response to the carrageenan-induced insult, restoring thresholds to baseline values for both wild-type and ALDH2*1/*2 mice, when assessed by the up-down method(fig. S4F) (baseline: wild type, 1.5 ± 0.1 g and ALDH2*1/*2, 1.6 ± 0.2 g;3 hours after carrageenan: wild type, 1.7 ± 0.4 g and ALDH2*1/*2, 1.3 ±0.2 g; mean ± SEM, n = 8 per group). When Alda-1 was given to wild-type mice alone, without an inflammatory insult, no changes werenoted in behavioral response (fig. S4G). We confirmed Alda-1 selectiv-ity for ALDH2 by testing in vitro its effect on the activity of five otherALDH enzymes (14) (table S1).
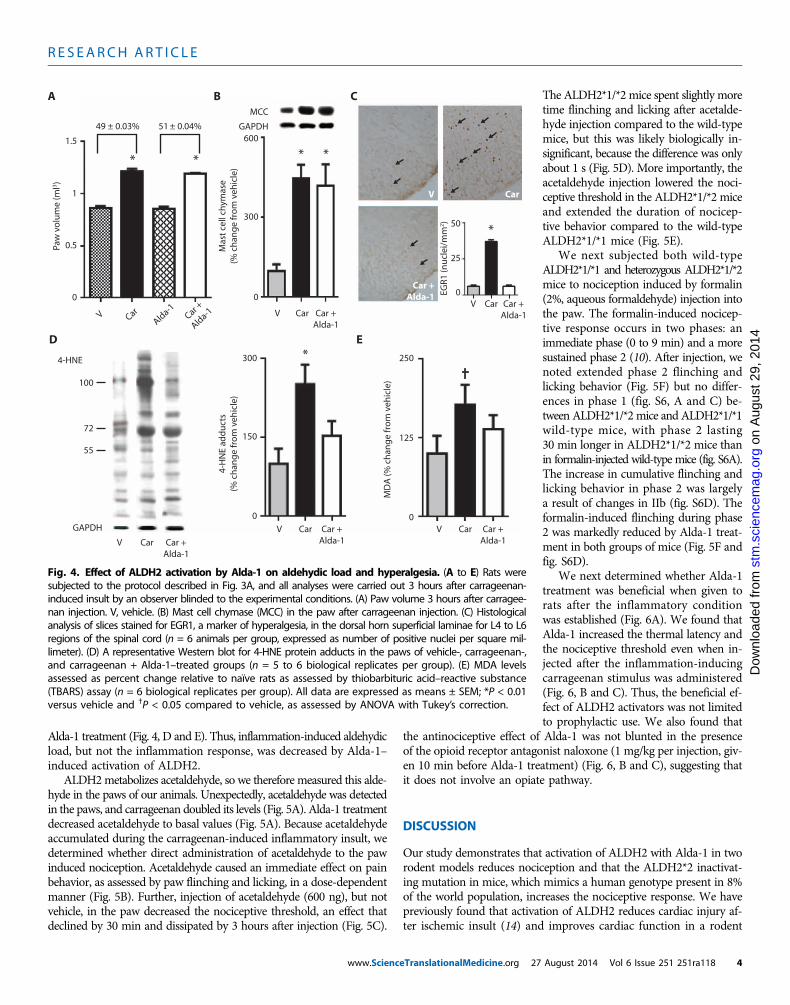
To further investigate how ALDH2 activity may regulate nocicep-tion, we used a rat model of inflammatory pain by measuring a ratnociceptive response after injection of an inflammatory stimulus inthe hind paw (protocol; Fig. 3A). When given alone without an inflam-matory stimulus, Alda-1 had no effects on response to thermal stimuli(defined as time lapsed after a change in thermal stimulus, or thermallatency) (Fig. 3B). Similarly, Alda-1 did not affect basal sensitivity tomechanical stimulus as assessed with the Randall-Selitto test, in whichthe amount of weight in grams applied to the rat paw is incrementallyincreased (70 ± 2 g versus 73 ± 1 g, with and without Alda-1, respec-tively; n = 10, Alda-1 group; n = 6, vehicle group). However, whencarrageenan was injected in one paw, Alda-1 doubled the latency timeto thermal stimulation of that paw (Fig. 3B). Additionally, Alda-1 dou-bled the nociceptive threshold to mechanical stimulation: rats withdrewtheir inflamed paw at 45 g of pressure in the absence of Alda-1; afterAlda-1 treatment, they withdrew their paw only after 90 g of pressurewas applied (Fig. 3C, y-axis values). Similar data were observed whenprostaglandin E2 (PGE2), instead of carrageenan, was used to inducenociception (Fig. 3C, y-axis values). In each condition, ALDH activityin the paw correlated with nociceptive threshold (R2 = 0.90; Fig. 3C).
To test whether the Alda-1 reduction in nociception was a result ofan anti-inflammatory effect, we measured carrageenan-induced edemaof the paw and found that inflammation was unaltered by Alda-1 treat-
www.Scienc
ment (Fig. 4, A and B). Carrageenan administration caused a ~50%increase in paw volume over the untreated contralateral paw of vehicle-treated rats, and this increase was unaffected by Alda-1 treatment (Fig.4A). Further, markers of inflammation, including mast cell chymase(Fig. 4B) and neutrophil elastase (267 ± 61 versus 220 ± 26 relativeunits determined by Western blot; fig. S5), increased equally in thevehicle-treated and Alda-1–treated carrageenan groups. In contrast,the levels of early growth response protein 1 (EGR1), a central nervoussystem marker of hyperalgesia (19), increased about fivefold in the L4to L6 spinal cord dorsal horn superficial laminae, and the increasein EGR1 levels was brought back to basal levels by Alda-1 treatment(Fig. 4C). We also found that 4-HNE protein adducts and MDA levelswere elevated in the inflamed paw and that these levels declined after
A
B
C
−15 0 30 150 180 min
Test TestTreatTreat TreatCar orPGE2
Late
ncy
(s)
30
15
0− +− +
+ +−−
Car
Alda-1
100
65
30
No
cice
pti
ve t
hre
sho
ld (g
)
PGE2
Alda-1
Car
Car +Alda-1
Untreated
PGE2
7 10 13
ALDH activity (µmol/min per mg)
*
◊
Mechanical
Thermal
R2 = 0.90
+
Fig. 3. Experimental protocol and nociceptive behavior in rats. (A) Ex-perimental protocol: Nociceptive threshold testing (closed circles) assessed
by the Randall-Selitto method at baseline and 3 hours after inflammatoryinsult. Red arrows indicate treatment either with vehicle or with the ALDH2activator Alda-1 (2 mg/kg, subcutaneously). At time zero (black arrow), theinflammatory insults (carrageenan or PGE2) were given to the right hindpaw. (B) Latency to thermal stimulation for rat hind paw withdrawal withand without treatment with Alda-1. (C) Nociceptive threshold in rats sub-jected to carrageenan or PGE2 proinflammatory insults. All data are expressedas means ± SEM; *P < 0.01 compared to untreated and Alda-1 alone groupand ◊P < 0.01 compared to all other groups, as assessed by ANOVA withTukey’s correction (B) and linear regression (C); n = 6 to 8 animals per group,tested once per animal; the observers were blinded to the experimentalconditions.eTranslationalMedicine.org 27 August 2014 Vol 6 Issue 251 251ra118 3
R E S EARCH ART I C L E
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
Alda-1 treatment (Fig. 4, D and E). Thus, inflammation-induced aldehydicload, but not the inflammation response, was decreased by Alda-1–induced activation of ALDH2.ALDH2metabolizes acetaldehyde, so we therefore measured this alde-hyde in the paws of our animals. Unexpectedly, acetaldehyde was detectedin the paws, and carrageenan doubled its levels (Fig. 5A). Alda-1 treatmentdecreased acetaldehyde to basal values (Fig. 5A). Because acetaldehydeaccumulated during the carrageenan-induced inflammatory insult, wedetermined whether direct administration of acetaldehyde to the pawinduced nociception. Acetaldehyde caused an immediate effect on painbehavior, as assessed by paw flinching and licking, in a dose-dependentmanner (Fig. 5B). Further, injection of acetaldehyde (600 ng), but notvehicle, in the paw decreased the nociceptive threshold, an effect thatdeclined by 30 min and dissipated by 3 hours after injection (Fig. 5C).
www.ScienceTranslationalMedicine.org 27
The ALDH2*1/*2 mice spent slightly moretime flinching and licking after acetalde-hyde injection compared to the wild-typemice, but this was likely biologically in-significant, because the difference was onlyabout 1 s (Fig. 5D). More importantly, theacetaldehyde injection lowered the noci-ceptive threshold in the ALDH2*1/*2 miceand extended the duration of nocicep-tive behavior compared to the wild-typeALDH2*1/*1 mice (Fig. 5E).
We next subjected both wild-typeALDH2*1/*1 and heterozygous ALDH2*1/*2mice to nociception induced by formalin(2%, aqueous formaldehyde) injection intothe paw. The formalin-induced nocicep-tive response occurs in two phases: animmediate phase (0 to 9 min) and a moresustained phase 2 (10). After injection, wenoted extended phase 2 flinching andlicking behavior (Fig. 5F) but no differ-ences in phase 1 (fig. S6, A and C) be-tween ALDH2*1/*2 mice and ALDH2*1/*1wild-type mice, with phase 2 lasting30 min longer in ALDH2*1/*2 mice thanin formalin-injected wild-type mice (fig. S6A).The increase in cumulative flinching andlicking behavior in phase 2 was largelya result of changes in IIb (fig. S6D). Theformalin-induced flinching during phase2 was markedly reduced by Alda-1 treat-ment in both groups of mice (Fig. 5F andfig. S6D).
We next determined whether Alda-1treatment was beneficial when given torats after the inflammatory conditionwas established (Fig. 6A). We found thatAlda-1 increased the thermal latency andthe nociceptive threshold even when in-jected after the inflammation-inducingcarrageenan stimulus was administered(Fig. 6, B and C). Thus, the beneficial ef-fect of ALDH2 activators was not limitedto prophylactic use. We also found that
the antinociceptive effect of Alda-1 was not blunted in the presenceof the opioid receptor antagonist naloxone (1 mg/kg per injection, giv-en 10 min before Alda-1 treatment) (Fig. 6, B and C), suggesting thatit does not involve an opiate pathway.
DISCUSSION
Our study demonstrates that activation of ALDH2 with Alda-1 in tworodent models reduces nociception and that the ALDH2*2 inactivat-ing mutation in mice, which mimics a human genotype present in 8%of the world population, increases the nociceptive response. We havepreviously found that activation of ALDH2 reduces cardiac injury af-ter ischemic insult (14) and improves cardiac function in a rodent
A B C
D E
Paw
vo
lum
e (m
l3 )
1.5
1
0.5
0
V CarAld
a-1Car +
Alda-1
49 ± 0.03% 51 ± 0.04%
* *
Mas
t ce
ll ch
ymas
e(%
ch
ang
e fr
om
veh
icle
)
MCC
GAPDH600
300
0
* *
V Car Car +Alda-1
EGR1
(nu
clei
/mm
2 ) 50
25
0V Car Car +
Alda-1
55
100
72
4-HNE
GAPDH
V Car Car +Alda-1
4-H
NE
add
uct
s(%
ch
ang
e fr
om
veh
icle
)
300
150
0
V Car Car +Alda-1
V Car Car +Alda-1
MD
A (%
ch
ang
e fr
om
veh
icle
)
250
125
0
*
*
V Car
Car +Alda-1
†
Fig. 4. Effect of ALDH2 activation by Alda-1 on aldehydic load and hyperalgesia. (A to E) Rats weresubjected to the protocol described in Fig. 3A, and all analyses were carried out 3 hours after carrageenan-
induced insult by an observer blinded to the experimental conditions. (A) Paw volume 3 hours after carragee-nan injection. V, vehicle. (B) Mast cell chymase (MCC) in the paw after carrageenan injection. (C) Histologicalanalysis of slices stained for EGR1, a marker of hyperalgesia, in the dorsal horn superficial laminae for L4 to L6regions of the spinal cord (n = 6 animals per group, expressed as number of positive nuclei per square mil-limeter). (D) A representative Western blot for 4-HNE protein adducts in the paws of vehicle-, carrageenan-,and carrageenan + Alda-1–treated groups (n = 5 to 6 biological replicates per group). (E) MDA levelsassessed as percent change relative to naïve rats as assessed by thiobarbituric acid–reactive substance(TBARS) assay (n = 6 biological replicates per group). All data are expressed as means ± SEM; *P < 0.01versus vehicle and †P < 0.05 compared to vehicle, as assessed by ANOVA with Tukey’s correction.August 2014 Vol 6 Issue 251 251ra118 4

R E S EARCH ART I C L E
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
model of heart failure (20). Here, we also show that the reduced noci-ception by Alda-1 treatment is insensitive to opiate receptor antagonists.Together, our data suggest that ALDH2 activators, such as Alda-1, mayrepresent a non-narcotic, cardiac-safe class of therapeutic agents foracute inflammatory-induced pain.
Pain is a subjective feeling that can only be measured in animalmodels as a behavioral response to nociceptive stimulus. We used twodifferent techniques to assess nociception in mice: a scoring system usedby some East Asian laboratories (18) and the up-down technique usedby the Western world (21) to study pain behavior in rodents. Further,because of the small paw volume in a mouse and the time required
www.ScienceTranslationalMedicine.org 27
to breed a number of mice for our be-havioral studies, we used a second modelof pain in rats and performed more ex-tensive studies in that model.
Reactive aldehydes, includingMDA and4-HNE, induce pain responses (22, 23).These aldehydes are produced by reactiveoxygen species–induced peroxidation ofexogenous and endogenous polyunsatu-rated fatty acids. Blood levels of 4-HNEin healthy individuals (~0.5 mM) can in-crease 10 to 100 times under oxidativestress (24, 25). Similar increases in MDAlevels after oxidative stress have also beenreported (26). Although its main sourceis ethanol metabolism (27, 28), acetaldehydemay be formed by pyruvate dehydrogenasein the mitochondria, and the accumula-tion of aldehydes impairs mitochondrialALDH2 activity (14); these sources mayhave contributed to the acetaldehyde ac-cumulation that we observed. Regardless,our data demonstrate the importance ofALDH2 in controlling the levels of reactivealdehydes, including 4-HNE, MDA, andacetaldehyde, and reducing pain sensation.
Although additional transgenic micewith alterations in ALDH2 exist, includinga knockout ALDH2 mouse (29, 30) andan overexpressing ALDH2*2 mouse (31),the ALDH2*2 mice that we have generatedare particularly suitable for preclinical andtranslational studies because they mimicthe human ALDH2*2 point mutation.Like humans carrying the ALDH2*2 mu-tation, no noticeable gross motor differenceswere identified in mice with this mutation.In addition, ALDH2 protein levels and en-zymatic activity in the heterozygotic micewere decreased similarly to these valuesin heterozygotic humans (17).
A number of limitations to our studyshould be noted:We only studied ALDH2*1/*2 heterozygotic, not homozygotic *2/*2,mice. We also did not determine whetherthe beneficial effect of Alda-1 treatmentresults from central or peripheral action,
nor did we determine Alda-1 efficacy in chronic pain or arthritis.Nevertheless, because mice with this mutation exhibited reduced re-sponse to nociception after Alda-1 treatment, subjects with the ALDH2*2variant may experience more severe inflammatory pain and benefitfrom drugs that activate ALDH2.
Known genetic mutations regulating pain sensation are limited tosmall familial subsets of the human population (32, 33) and to rarepoint mutations in ion channels (34, 35). Our findings in rodents sug-gest that the ALDH2 point mutation ALDH2*2, present in 8% of theworld population, might also contribute to lower pain thresholds inhumans. The prevalence of the ALDH2*2 point mutation, which is
A B C
D F
Ace
tald
ehyd
e(n
g/g
tis
sue)
V Car Car +Alda-1
1000
500
0100 300 600–
Acetaldehyde (ng)
10
5
0
Lick
ing
or f
linch
ing
(s)
15
No
cice
pti
ve t
hre
sho
ld (g
)
100
50
00 10 15 20 30 180
Time aftertreatment (min)
WT
*1/*2
*1/*2W
T
(−) (+)
8
4
0
Lick
ing
or f
linch
ing
(s)
2
1
0
0 10 30 60 120
Time aftertreatment (min)
Phas
e II
cum
ula
tive
lick
ing
an
dfli
nch
ing
beh
avio
r (s)
No
cice
pti
ve t
hre
sho
ld (g
)
800
400
0
WT
*1/*2
*1/*2W
T
Alda-1 (−) (+)(−) (+)
AcetaldehydeAcetaldehyde
Acetaldehyde
Formalin
*
*
* **
*
*
◊◊
◊
◊
∆
∆
†
†
†
Acetaldehyde
E
Fig. 5. ALDH2 involvement in acetaldehyde- and formaldehyde-induced acute nociception. (A)Quantification of acetaldehyde in carrageenan-induced inflamed paw (n = 3 biological replicates per
group, repeated three times for each replicate). (B) Acetaldehyde at the amounts measured in the paw[in (A)] was injected into the hind paw, and licking and flinching were measured. (C) Nociceptive thresholdafter administration of 600 ng of acetaldehyde (n = 4 to 7 per animals group, tested once for each animal).(D) A single acetaldehyde (100 ng) injection was administered to the paw, and licking and flinching weremeasured (n = 8 per group, tested once per animal). (E) Using the up-down technique, the intensity andduration of the acetaldehyde-induced nociceptive behavior were measured in ALDH2*1/*2 mice and WTALDH2*1/*1 mice (n = 8 per group, tested once per animal). (F) Phase 2 cumulative flinching and lickingfor WT ALDH2*1/*1 mice and ALDH2*1/*2 mice (n = 8 per group, tested once per animal). All data areexpressed as means ± SEM; *P < 0.001 versus vehicle, †P < 0.05 versus all other groups, ◊P < 0.001 com-pared to all other groups, and DP < 0.001 compared to respective vehicle-treated groups, as assessed byANOVA with Tukey’s correction; blue data points and bars, WT ALDH2*1/*1 mice; red data points and bars,ALDH2*1/*2 mice.August 2014 Vol 6 Issue 251 251ra118 5

R E S EARCH ART I C L E
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
unique to East Asian ethnicity, is ~36% (11) and in some regions ashigh as 45% (36). Asians are reported to have a lower pain tolerance thanother ethnicities (37–40) and are more responsive to painful stimuli suchas skin irritation (39), capsaicin (40), heat (41), cold (42), or mechanicalstimulus (37) compared with other ethnic populations. Although thesedifferences are ascribed primarily to social or cultural factors (38), it ispossible that increased pain sensitivity among Asians can be attri-buted, at least in part, to the ALDH2*2 mutation. Case reports providefurther evidence for a role for ALDH2 in pain sensation, suggestingthat the ALDH2 inhibitor disulfirammay cause painful peripheral neu-ropathy in a dose-dependent and reversible manner (43). Nevertheless,
www.Scienc
our data in rodents indicating that ALDH2 activity attenuates nocicep-tion remain to be confirmed in humans.
Our findings that Alda-1 corrects ALDH2 inactivity due to the mu-tation in ALDH2*2 (15) suggest that compounds like Alda-1 may ben-efit East Asian and other populations in reducing inflammatory pain.The apparent non–opioid-dependent mechanism for Alda-1 would cir-cumvent the potential for addiction and abuse seen with opioids usedfor analgesia (44). Moreover, Alda-1 is beneficial when given duringischemic injury (14) and reduces ischemia-induced cardiac arrhythmias(45). Although more studies are required, this non-opioid, cardiac-safeprofile for Alda-1 may be particularly useful for patients with cardiacrisk factors and known cardiac disease because the use of some tradi-tional NSAIDs or COX-2 inhibitors was confirmed in a large meta-analysis to increase cardiovascular risk (7).
In summary, our study identified ALDH2 as a regulator of acutenociception in rodents. Although confirmatory studies are needed, ourrodent studies suggest that the ALDH2*2 mutation may underlie dif-ferences in pain behavior observed in humans. We showed that het-erozygous mice with an ALDH2 inactivating mutation, ALDH2*1/*2,have increased reactive aldehyde-induced adducts and increased noci-ception when compared to wild-type mice; these effects were correctedby Alda-1 treatment. Increasing the activity of mitochondrial ALDH2with Alda-1 reduced aldehydic load, limited changes in the marker ofhyperalgesia EGR1, and provided analgesia. Our data suggest that ac-tivators of ALDH2, such as Alda-1, may be a promising therapeuticdrug class to reduce pain that likely acts independently of opioidreceptor signaling and without increasing cardiovascular risk.
MATERIALS AND METHODS
Study designAll procedures were in accordance with the guidelines of the InternationalAssociation for the Study of Pain and approved by Stanford University(#25505) and the Butantan Institute, São Paulo, Brazil (CEUAIB #838).
The study objective tested whether ALDH2 regulated inflammatorypain in rodent models. We tested our hypothesis by using (i) ALDH2*1/*2knock-in mice litter-matched to C57BL/6 wild-type ALDH2 mice; (ii)Wistar rats; (iii) an ALDH2 activator, Alda-1, which also corrects thedecreased enzymatic activity of the ALDH2*1/*2 mutation; and (iv) in-flammatory painmodels by administering carrageenan, PGE2, acetalde-hyde, or formalin to the rodent paw.Weused bothmechanical and thermalmethods of nociception detection, in addition to biochemical analysis.
On the basis of our previous experience, a minimum of six rodentsper group is required to obtain statistically meaningful behavioral data.An experimental group size of six or more animals is necessary toachieve at least a 20% minimal difference in behavior for a power of95% with a < 0.05 and b < 20%. All behavioral tests were performedbetween 9:00 a.m. and 4:00 p.m. by observers blinded to the treatmentgroups. MaleWistar rats (170 to 190 g) andmalemice (18 to 21 g) werehoused in a temperature- and light-controlled room for at least 3 daysbefore use. All animals were randomized and assigned to testing groupsto generate biological replicates for each group, and the experimenterwas blinded as to genotype and drug treatment given.
DrugsSaline (0.1 ml; control), carrageenan (200 mg for rats and 100 mg formice; Marine Colloids), PGE2 (100 ng, Sigma), or formalin (2%,
A0 30 150 180 min
Test TestTreatTreatCarNal Nal
B
C
30
15
0
0
100
50
Car Car +Alda-1
Car +Nal
Car +Nal +
Alda-1
Car Car +Alda-1
Car +Nal
Car +Nal +
Alda-1
**
**
Late
ncy
(s)
No
cice
pti
ve t
hre
sho
ld (g
)
Fig. 6. Effect of Alda-1 after the inflammatory insult on nociceptivethreshold. (A) Nociceptive threshold testing (closed circles) assessed by
either the Hargreaves or the Randall-Selitto method at baseline and 3hours after inflammatory insult. Red arrows, treatment with either vehicleor the ALDH2 activator Alda-1 (2 mg/kg, subcutaneously). Naloxone (Nal,1 mg/kg) or vehicle was given subcutaneously (green arrows) 10 minbefore each Alda-1 treatment. At time zero (black arrow), the inflammatoryinsult (carrageenan) was given to the right hind paw. (B and C) Alda-1 wasgiven 30 and 150 min after carrageenan (n = 9 per group, tested once peranimal; *P < 0.0001 as assessed by ANOVA followed by Tukey’s correction).Error bars represent means ± SEM. The observers were blinded to the ex-perimental conditions.eTranslationalMedicine.org 27 August 2014 Vol 6 Issue 251 251ra118 6

R E S EARCH ART I C L E
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
Sigma) was administered by intraplantar injection into the right hindpaw to induce nociception. These doses were chosen on the basis of pre-vious studies by our laboratory groups (46) and published literature (47).
Alda-1 dose (2 mg/kg per injection, dissolved in 50% polyethyleneglycol/50% dimethyl sulfoxide) was established in preliminary studiesand given in multiple doses because of its estimated short half-life.Alda-1 was injected subcutaneously to the dorsal side of the neck15 min before carrageenan or saline and again at 30 and 150 min aftercarrageenan, PGE2, or saline injection. One subset of animals receivedAlda-1 only 30 and 150 min after carrageenan injection. Where indicated,naloxone (1 mg/kg per injection, Sigma) or vehicle (saline) was given10 min before each Alda-1 treatment to the dorsal side of the neck.
Knock-in mice generation (see also fig. S1)ALDH2*2 knock-in mice with a C57BL/6 background were generatedby homologous recombination. The 8.0-kb genomic fragment encom-passing the mouse ALDH2 locus was cloned to the gene-targetingvector pPNT to introduce a single amino acid substitution. Site-directedmutagenesis introduced a single nucleotide substitution (G to C) withinexon 12 of the ALDH2 genomic fragment corresponding to the positionof human E487K mutation. Plasmid DNA from the constructed pPNTvector was electroporated to embryonic stem (ES) cells. Positive ESclones were selected on the basis of neomycin and thymine kinasemarkers and confirmed by polymerase chain reaction (PCR), restric-tion fragment length polymorphism, and DNA sequencing. PositiveES cells were microinjected into C57BL/6 blastocytes and implantedinto pseudopregnant females.
Germ-line transmission of the ALDH2 E487K mutation by homol-ogous recombination was derived from selected ES cell lines from foundermice. The genotype of the E487K mutation was confirmed by direct ge-nomic DNA sequencing of PCR fragments from amplified genomic DNA.The specific primers EG475 (TACTGTCAAAGTGCCACAGAAGAA-CTCGTAA), EG460 (AACCTGCGTGCAATCCATCTTGTTCAATGG),and EG399 (TTGGCCTTCCACTGGGA GTGGGTCCCTCTGTC)were used for the amplification of a 1.3-kb fragment from exon 13 tothe 3′ untranslated region of neomycin marker and a 3.0-kb fragmentfrom exon 13 to downstream of the neomycin marker, respectively, forthe presence of the mutated allele. In contrast, for the wild-type allele,a 1.4-kb fragment devoid of the neomycin marker was amplified withEG475 and EG399 primers. We confirmed the expression of the ALDH2E487K mutant protein, which exhibited a change in charge as a resultof the glutamate-to-lysine substitution, by isoelectric focusing gel elec-trophoresis and Western blot with an ALDH2-specific antibody (SantaCruz Biotechnology, 1:500). The founder mice were backcrossed to theC57BL/6 background for at least seven generations to achieve a homog-eneous genetic background, and the E487K mutation was transmittedas a single Mendelian gene.
Nociception assessmentThe experimental performer was blinded to both genotype and treatments.Nociceptive response was assessed by testing mechanical and thermal hy-persensitivity. Mechanical sensitivity was measured by the von Freymethod for mice and the Randall-Selitto method for rats immediatelybefore and 180 min after carrageenan or PGE2 injection. Thermal sensi-tivity was measured by the Hargreaves method. Both mice and rats wereacclimated to the testing equipment 1 day before baseline testing.
To measure mechanical sensitivity, mice were placed individuallyin a plastic cage with a wire mesh bottom for testing. After acclimation
www.Scienc
to the plastic cage for at least 15 min, a von Frey hair was pressed per-pendicularly against the plantar skin surface and held for about 5 suntil the hair slightly buckled (36, 48). Mice were randomly separatedinto two testing groups, one scored with the up-down technique andthe other with a pain scoring system.
The up-down technique was performed as follows: Before and aftercarrageenan administration, von Frey hairs of increasing force wereapplied to the mice hind paw until the paw was withdrawn. Once aresponse was identified, the next lower von Frey hair was applied. Foreach positive response (paw withdrawal), the next lesser filament wastested. For each negative response, the next higher filament was tested.The sequence of positive and negative responses was recorded andincorporated in an established curve-fitting algorithm to determinemechanical sensitivity (49).
The scoring system technique was performed as follows: Hind pawnociception was assessed with six different von Frey hairs (0.03, 0.17,0.41, 0.69, 1.20, and 1.48 g). Using the same stimulation intensity asthe 0.03-g von Frey hair, each individual hair was applied six times toeach hind paw at intervals of several seconds. Because the response toeach von Frey stimulus is fairly subjective, we used a method of objec-tifying these nociceptive responses by assigning a specific number tothe response. The scoring system was as follows: 2, no response; 1, with-draws paw from hair; 0, immediate flinching or hind paw licking. Thismethod was previously shown by others to provide a reliable, objectiveassessment of pain response using von Frey hairs (18, 50).
Nociceptive threshold was also determined with the Randall-Selittopaw pressure test (Ugo Basile), as described (48). The baseline valuesvaried between 65 and 75 g of force. Animals with baseline out of thisrange (10%) were excluded before initiating the study to achieve uni-form testing.
Thermal sensitivity, as assessed by hind paw withdrawal threshold,was measured by the Hargreaves method (51). Rats were placed in theapparatus to acclimate for 10 min. A light source was then applied tothe hind paw, and the time until the rat withdrew the paw from thelight source was assessed. A 30-s exposure time was used to limitthermal damage to the paw region, with light source intensity adjustedin preliminary studies to cause a latency response of 15 s in control rats.
Edema assessment. Inflammation-induced edema of the injectedpaw was assessed with a plethysmometer (Ugo Basile) (52).
Evaluation of general motor activity (open field and rotarod).The general activity of ALDH2*1/*2 knock-in and wild-type mice wasassessed in an open-field arena (Ugo Basile). Each animal was individ-ually placed in the center zone, and behavioral parameters were re-corded for 3 min (48). Hand-operated counters were used to tallythe number of squares crossed and rearing behavior (number of timesstanding on hind legs). For rotarod testing, mice were trained at a fixedspeed of 10 rpm for three consecutive days. Mice were then tested atdifferent speeds, and latency to fall was measured. For the accelerationtask, the rotarod was accelerated from 5 to 40 rpm over 5 min. The falllatency was recorded.
Blood acetaldehyde determination. Blood samples (50 to 100 ml)were drawn from the retro-orbital sinus under anesthesia. Levels ofacetaldehyde were determined by a fluorescence-based high-performanceliquid chromatography method, as described previously (53).
Tissue acetaldehyde measurements. Paw samples were quicklysealed in microcentrifuge tubes and weighed, and 3 volumes (v/w) ofLiberase TL (0.75 mg/ml) (Roche) was added for overnight digestionat 37°C. Acetaldehyde standard solutions (100 to 5000 ng/ml) and
eTranslationalMedicine.org 27 August 2014 Vol 6 Issue 251 251ra118 7

R E S EARCH ART I C L E
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
tissue samples were reacted with saturated 2,4-dinitrophenylhydrazine(DNPH) solution.
To establish liquid chromatography–tandem mass spectrometry(LC-MS/MS) conditions, a standard of acetaldehyde DNPH was used.Chromatograms of standards established characteristic retention timesof acetaldehyde and verified that the MS signal was linear over therange of 0.1 to 5 mg/ml. The acetaldehyde peak area derivatives werecalculated and plotted against the concentration of the calibrationstandards with Analyst 1.5.1 software.
Aliquots (30 ml) were taken from each paw homogenate sample.Acetaldehyde was extracted from samples of paw homogenate by add-ing 150 ml of methanol containing DNPH (40 mg/ml). Both unknownsamples and standards were vortexed and centrifuged to separate the pre-cipitate. Ten microliters of sample extracts was injected into LC-MS/MSfor analysis. Automatic peak detection, integration, and data processingwere performed by the AB SCIEX Analyst 1.5.1 software package. Con-centrations of acetaldehyde were calculated by plotting peak area ofunknown samples against a standard curve. The data were normalizedto the paw volume obtained by plesthysmography.
Acetaldehyde experiments. Rats were acclimated to the test en-vironment 15 min before experiments. Acetaldehyde (100, 300, and600 ng/g of paw tissue) was diluted in saline containing 0.5% Tween80 and injected intraplantarly. Hind paw licking and flicking behaviorwas quantified for 10 min. Immediately after this test, the paw thresh-old was assessed at 10, 15, 20, 30, and 180 min after acetaldehydeinjection, using the Randall-Selitto test as described. For mice, acetal-dehyde (100 ng/g of paw tissue) was injected intraplantarly, andlicking and flicking behavior was quantified for 10 min. Immediatelyafterward, the paw threshold was assessed at 10, 30, 60, and 120 min,using the von Frey test as described.
Formalin experiments. Mice were placed individually in a plasticcage. After acclimation for ~15 min, formalin (2%, 20 ml, in saline) wasinjected, and flinches as well as time spent licking the injected hind pawwere recorded.Behaviorwasdivided intophase1 (0 to10min) andphase 2(10 to 90 min), with phase 2 being further divided into phase 2a (10 to40min) andphase2b (40 to90min) asdescribed (10). For a subset of experi-ments, Alda-1 was given once, 5min before formalin was administered.
Biochemical studies for rats and miceOne hundred eighty minutes after treatment, rat paw tissue washomogenized and centrifuged at 700g for 5 min at 4°C, and the pelletwas discarded. Protein concentration in the supernatant was deter-mined by Bradford assay.
ALDH enzymatic measurementALDH enzymatic activity was determined as described (14).
Western blot analysisWestern blot was performed as described (14). For 4-HNE protein ad-duct detection, an antibody against the reductively stabilized 4-HNEamino acid adduct (Alpha Diagnostic Intl., 1:1000) was used. For mastcell chymase and neutrophil elastase, detection antibodies from SantaCruz Biotechnology (1:500) were used. Quantification analysis of blotswas performed with ImageJ software, using GAPDH as a loading control.
TBARS assayTBARS, which measures MDA, was performed with an OXItek TBARSassay kit (ZeptoMetrix). The concentration of TBARS was expressed in
www.Scienc
picomoles per milligram of protein by using a standard curve of MDAat 0 to 200 pmol concentrations and expressed as percent control.
ImmunohistochemistryAnimals were anesthetized with ketamine (90 mg/kg) and xylazine(15 mg/kg) after 3 hours of the experimental protocol. Subsequently,animals were perfused with 4% paraformaldehyde (PFA) in 0.1 Mphosphate buffer (pH 7.4). Spinal cord sections (L4 to L6) were re-moved, post-fixed for 4 hours in PFA (4%), and transferred to a 30%sucrose solution in phosphate buffer to ensure cryoprotection. Spinalcords were sectioned (10 mm) on a cryostat and displayed on histo-logical slides. The sections were immunostained for EGR1 gene ex-pression, using a rabbit polyclonal antibody (Santa Cruz Biotechnology,1:500) in phosphate buffer containing 0.3% Triton X-100 and 5% nor-mal donkey serum overnight at 24°C. The sections were then washedin phosphate buffer and incubated with a biotinylated donkey anti-rabbit serum (Vector Labs, 1:200) in phosphate buffer for 2 hours at 24°C.The sections were washed again in phosphate buffer and incubated withthe avidin-biotin-peroxidase complex (Elite ABC, Vector Labs). Thesections were then reacted with 0.05% 3,3-diaminobenzidine and0.01% hydrogen peroxide in phosphate buffer. Intensification wasconducted with 0.05% osmium tetroxide in water. The sections weredehydrated, cleared, and mounted on a coverslip. Normal rat serumserved as control. Digital images of light microscopy were collected,and cells with positive EGR were counted.
Statistical analysisData are expressed as means ± SEM. ANOVA with Tukey’s post hocanalysis was used for behavioral pain studies and biochemical analysis.When only two groups were analyzed and compared, an unpaired ttest was performed. For Fig. 3C, linear regression analysis and Pearsoncorrelation testing were used to assess concordance. A value of P <0.05 was considered significant. GraphPad Prism software was usedfor all statistical analysis.
SUPPLEMENTARY MATERIALS
www.sciencetranslationalmedicine.org/cgi/content/full/6/251/251ra118/DC1Fig. S1. Scheme for generation of ALDH2*2 (E487K) knock-in mice.Fig. S2. Rotarod data.Fig. S3. ALDH2 protein level in mouse paw and additional nociceptive data.Fig. S4. Nociceptive testing.Fig. S5. Neutrophil elastase protein levels.Fig. S6. Formalin-induced pain model.Table S1. ALDH activation by Alda-1.
REFERENCES AND NOTES
1. D. S. Goldberg, S. J. McGee, Pain as a global public health priority. BMC Public Health 11,770 (2011).
2. N. D. Volkow, T. A. McLellan, J. H. Cotto, M. Karithanom, S. R. Weiss, Characteristics of opioidprescriptions in 2009. JAMA 305, 1299–1301 (2011).
3. C. M. Jones, K. A. Mack, L. J. Paulozzi, Pharmaceutical overdose deaths, United States, 2010.JAMA 309, 657–659 (2013).
4. Substance Abuse and Mental Health Services Administration, Results from the 2012 NationalSurvey on Drug Use and Health, NSDUH Series H-46, HHS Publication No. (SMA) 13-4795.Rockville, MD (2013).
5. American Gastroenterological Association, Study shows long-term use of NSAIDs causessevere intestinal damage (ScienceDaily); http://www.sciencedaily.com/releases/2005/01/050111123706.htm [retrieved 16 January 2014].
eTranslationalMedicine.org 27 August 2014 Vol 6 Issue 251 251ra118 8

R E S EARCH ART I C L E
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
6. D. Mukherjee, S. E. Nissen, E. J. Topol, Risk of cardiovascular events associated with selectiveCOX-2 inhibitors. JAMA 286, 954–959 (2001).
7. Coxib and traditional NSAID Trialists’ (CNT) Collaboration, N. Bhala, J. Emberson, A. Merhi,S. Abramson, N. Arber, J. A. Baron, C. Bombardier, C. Cannon, M. E. Farkouh, G. A. FitzGerald,P. Goss, H. Halls, E. Hawk, C. Hawkey, C. Hennekens, M. Hochberg, L. E. Holland, P. M. Kearney,L. Laine, A. Lanas, P. Lance, A. Laupacis, J. Oates, C. Patrono, T. J. Schnitzer, S. Solomon,P. Tugwell, K. Wilson, J. Wittes, C. Baigent, Vascular and upper gastrointestinal effects ofnon-steroidal anti-inflammatory drugs: Meta-analyses of individual participant datafrom randomised trials. Lancet 382, 769–779 (2013).
8. S. Bang, K. Y. Kim, S. Yoo, Y. G. Kim, S. W. Hwang, Transient receptor potential A1 mediatesacetaldehyde-evoked pain sensation. Eur. J. Neurosci. 26, 2516–2523 (2007).
9. M. Trevisani, J. Siemens, S. Materazzi, D. M. Bautista, R. Nassini, B. Campi, N. Imamachi, E. Andrè,R. Patacchini, G. S. Cottrell, R. Gatti, A. I. Basbaum, N. W. Bunnett, D. Julius, P. Geppetti,4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammationthrough activation of the irritant receptor TRPA1. Proc. Natl. Acad. Sci. U.S.A. 104, 13519–13524(2007).
10. C. R. McNamara, J. Mandel-Brehm, D. M. Bautista, J. Siemens, K. L. Deranian, M. Zhao, N. J. Hayward,J. A. Chong, D. Julius, M. M. Moran, C. M. Fanger, TRPA1 mediates formalin-induced pain.Proc. Natl. Acad. Sci. U.S.A. 104, 13525–13530 (2007).
11. P. J. Brooks, M. A. Enoch, D. Goldman, T. K. Li, A. Yokoyama, The alcohol flushing response:An unrecognized risk factor for esophageal cancer from alcohol consumption. PLOS Med.6, e50 (2009).
12. C. H. Chen, J. C. Ferreira, E. R. Gross, D. Mochly-Rosen, Targeting aldehyde dehydrogenase2: New therapeutic opportunities. Physiol. Rev. 94, 1–34 (2014).
13. R. S. Wang, T. Nakajima, T. Kawamoto, T. Honma, Effects of aldehyde dehydrogenase-2genetic polymorphisms on metabolism of structurally different aldehydes in human liver.Drug Metab. Dispos. 30, 69–73 (2002).
14. C. H. Chen, G. R. Budas, E. N. Churchill, M. H. Disatnik, T. D. Hurley, D. Mochly-Rosen, Ac-tivation of aldehyde dehydrogenase-2 reduces ischemic damage to the heart. Science 321,1493–1495 (2008).
15. S. Perez-Miller, H. Younus, R. Vanam, C. H. Chen, D. Mochly-Rosen, T. D. Hurley, Alda-1 is anagonist and chemical chaperone for the common human aldehyde dehydrogenase 2 variant.Nat. Struct. Mol. Biol. 17, 159–164 (2010).
16. A. Yokoyama, E. Tsutsumi, H. Imazeki, Y. Suwa, C. Nakamura, T. Mizukami, T. Yokoyama,Salivary acetaldehyde concentration according to alcoholic beverage consumed and aldehydedehydrogenase-2 genotype. Alcohol. Clin. Exp. Res. 32, 1607–1614 (2008).
17. C. L. Lai, C. T. Yao, G. Y. Chau, L. F. Yang, T. Y. Kuo, C. P. Chiang, S. J. Yin, Dominance of theinactive Asian variant over activity and protein contents of mitochondrial aldehyde de-hydrogenase 2 in human liver. Alcohol. Clin. Exp. Res. 38, 44–50 (2014).
18. I. Takasaki, T. Andoh, K. Shiraki, Y. Kuraishi, Allodynia and hyperalgesia induced by herpessimplex virus type-1 infection in mice. Pain 86, 95–101 (2000).
19. O. I. Rahman, R. Terayama, T. Ikeda, M. Koganemaru, T. Nakamura, R. Shiba, T. Nishimori,Differential effects of NMDA and AMPA/KA receptor antagonists on c-Fos or Zif/268 expres-sion in the rat spinal dorsal horn induced by noxious thermal or mechanical stimulation, orformalin injection. Neurosci. Res. 43, 389–399 (2002).
20. K. M. Gomes, J. C. Campos, L. R. Bechara, B. Queliconi, V. M. Lima, M. H. Disatnik, P. Magno,C. H. Chen, P. C. Brum, A. J. Kowaltowski, D. Mochly-Rosen, J. C. Ferreira, Aldehyde de-hydrogenase 2 activation in heart failure restores mitochondrial function and improvesventricular function and remodelling. Cardiovasc. Res. cvu125 (2014).
21. Y. Sun, P. Sahbaie, D. Y. Liang, W. W. Li, X. Q. Li, X. Y. Shi, J. D. Clark, Epigenetic regulation of spinalCXCR2 signaling in incisional hypersensitivity in mice. Anesthesiology 119, 1198–1208 (2013).
22. Y. Riahi, G. Cohen, O. Shamni, S. Sasson, Signaling and cytotoxic functions of 4-hydroxyalkenals.Am. J. Physiol. Endocrinol. Metab. 299, E879–E886 (2010).
23. A. M. Patwardhan, A. N. Akopian, N. B. Ruparel, A. Diogenes, S. T. Weintraub, C. Uhlson, R. C. Murphy,K. M. Hargreaves, Heat generates oxidized linoleic acid metabolites that activate TRPV1 andproduce pain in rodents. J. Clin. Invest. 120, 1617–1626 (2010).
24. H. Esterbauer, R. J. Schaur, H. Zollner, Chemistry and biochemistry of 4-hydroxynonenal,malonaldehyde and related aldehydes. Free Radic. Biol. Med. 11, 81–128 (1991).
25. H. Strohmaier, H. Hinghofer-Szalkay, R. J. Schaur, Detection of 4-hydroxynonenal (HNE) asa physiological component in human plasma. J. Lipid Mediat. Cell Signal. 11, 51–61 (1995).
26. F. Nielsen, B. B. Mikkelsen, J. B. Nielsen, H. R. Andersen, P. Grandjean, Plasma malondialdehydeas biomarker for oxidative stress: Reference interval and effects of life-style factors. Clin. Chem.43, 1209–1214 (1997).
27. R. Parrilla, K. Okawa, K. O. Lindros, U. J. Zimmerman, K. Kobayashi, J. R. Williamson, Functionalcompartmentation of acetaldehyde oxidation in rat liver. J. Biol. Chem. 249, 4926–4933 (1974).
28. C. J. Eriksson, M. Marselos, T. Koivula, Role of cytosolic rat liver aldehyde dehydrogenase inthe oxidation of acetaldehyde during ethanol metabolism in vivo. Biochem. J. 152, 709–712(1975).
29. E. Fernandez, W. Koek, Q. Ran, G. A. Gerhardt, C. P. France, R. Strong, Monoamine metabolismand behavioral responses to ethanol in mitochondrial aldehyde dehydrogenase knockoutmice. Alcohol. Clin. Exp. Res. 30, 1650–1658 (2006).
www.Scienc
30. K. Kitagawa, T. Kawamoto, N. Kunugita, T. Tsukiyama, K. Okamoto, A. Yoshida, K. Nakayama,K. Nakayama, Aldehyde dehydrogenase (ALDH) 2 associates with oxidation of methoxyacetaldehyde;in vitro analysis with liver subcellular fraction derived from human and Aldh2 gene targetingmouse. FEBS Lett. 476, 306–311 (2000).
31. I. Ohsawa, K. Nishimaki, Y. Murakami, Y. Suzuki, M. Ishikawa, S. Ohta, Age-dependent neuro-degeneration accompanying memory loss in transgenic mice defective in mitochondrialaldehyde dehydrogenase 2 activity. J. Neurosci. 28, 6239–6249 (2008).
32. J. J. Cox, F. Reimann, A. K. Nicholas, G. Thornton, E. Roberts, K. Springell, G. Karbani, H. Jafri,J. Mannan, Y. Raashid, L. Al-Gazali, H. Hamamy, E. M. Valente, S. Gorman, R. Williams, D. P. McHale,J. N. Wood, F. M. Gribble, C. G. Woods, An SCN9A channelopathy causes congenital inability toexperience pain. Nature 444, 894–898 (2006).
33. B. Kremeyer, F. Lopera, J. J. Cox, A. Momin, F. Rugiero, S. Marsh, C. G. Woods, N. G. Jones,K. J. Paterson, F. R. Fricker, A. Villegas, N. Acosta, N. G. Pineda-Trujillo, J. D. Ramírez, J. Zea,M. W. Burley, G. Bedoya, D. L. Bennett, J. N. Wood, A. Ruiz-Linares, A gain-of-functionmutation in TRPA1 causes familial episodic pain syndrome. Neuron 66, 671–680 (2010).
34. R. E. Sorge, T. Trang, R. Dorfman, S. B. Smith, S. Beggs, J. Ritchie, J. S. Austin, D. V. Zaykin,H. Vander Meulen, M. Costigan, T. A. Herbert, M. Yarkoni-Abitbul, D. Tichauer, J. Livneh,E. Gershon, M. Zheng, K. Tan, S. L. John, G. D. Slade, J. Jordan, C. J. Woolf, G. Peltz, W. Maixner,L. Diatchenko, Z. Seltzer, M. W. Salter, J. S. Mogil, Genetically determined P2X7 receptor poreformation regulates variability in chronic pain sensitivity. Nat. Med. 18, 595–599 (2012).
35. E. C. Emery, G. T. Young, E. M. Berrocoso, L. Chen, P. A. McNaughton, HCN2 ion channelsplay a central role in inflammatory and neuropathic pain. Science 333, 1462–1466 (2011).
36. N. Enomoto, A. Takada, T. Date, Genotyping of the aldehyde dehydrogenase 2 (ALDH2)gene using the polymerase chain reaction: Evidence for single point mutation in theALDH2 gene of ALDH2-deficiency. Gastroenterol. Jpn. 26, 440–447 (1991).
37. K. M. Woodrow, G. D. Friedman, A. B. Siegelaub, M. F. Collen, Pain tolerance: Differencesaccording to age, sex and race. Psychosom. Med. 34, 548–556 (1972).
38. M. Y. Chan, T. Hamamura, K. Janschewitz, Ethnic differences in physical pain sensitivity:Role of acculturation. Pain 154, 119–123 (2013).
39. L. N. Rowell, B. Mechlin, E. Ji, M. Addamo, S. S. Girdler, Asians differ from non-HispanicWhites in experimental pain sensitivity. Eur. J. Pain 15, 764–771 (2011).
40. H. Wang, A. D. Papoiu, R. C. Coghill, T. Patel, N. Wang, G. Yosipovitch, Ethnic differences inpain, itch and thermal detection in response to topical capsaicin: African Americansdisplay a notably limited hyperalgesia and neurogenic inflammation. Br. J. Dermatol.162, 1023–1029 (2010).
41. P. J. Watson, R. K. Latif, D. J. Rowbotham, Ethnic differences in thermal pain responses: Acomparison of South Asian and White British healthy males. Pain 118, 194–200 (2005).
42. A. Y. Hsieh, D. A. Tripp, L. J. Ji, M. J. Sullivan, Comparisons of catastrophizing, pain attitudes,and cold-pressor pain experience between Chinese and European Canadian young adults.J. Pain 11, 1187–1194 (2010).
43. G. B. Frisoni, V. Di Monda, Disulfiram neuropathy: A review (1971–1988) and report of acase. Alcohol Alcohol. 24, 429–437 (1989).
44. S. Okie, A flood of opioids, a rising tide of deaths. N. Engl. J. Med. 363, 1981–1985 (2010).45. K. Koda, M. Salazar-Rodriguez, F. Corti, N. Y. Chan, R. Estephan, R. B. Silver, D. Mochly-Rosen,
R. Levi, Aldehyde dehydrogenase activation prevents reperfusion arrhythmias by inhibitinglocal renin release from cardiac mast cells. Circulation 122, 771–781 (2010).
46. V. O. Zambelli, A. C. Fernandes, V. P. Gutierrez, J. C. Ferreira, C. A. Parada, D. Mochly-Rosen,Y. Cury, Peripheral sensitization increases opioid receptor expression and activation bycrotalphine in rats. PLOS One 9, e90576 (2014).
47. S. D. Shields, D. J. Cavanaugh, H. Lee, D. J. Anderson, A. I. Basbaum, Pain behavior in theformalin test persists after ablation of the great majority of C-fiber nociceptors. Pain 151,422–429 (2010).
48. V. P. Gutierrez, K. Konno, M. Chacur, S. C. Sampaio, G. Picolo, P. Brigatte, V. O. Zambelli, Y. Cury,Crotalphine induces potent antinociception in neuropathic pain by acting at peripheral opioidreceptors. Eur. J. Pharmacol. 594, 84–92 (2008).
49. L. R. Poree, T. Z. Guo, W. S. Kingery, M. Maze, The analgesic potency of dexmedetomidine isenhanced after nerve injury: A possible role for peripheral a2-adrenoceptors. Anesth. Analg. 87,941–948 (1998).
50. C. S. Dale, C. Altier, N. Cenac, R. Giorgi, M. A. Juliano, L. Juliano, G. W. Zamponi, N. Vergnolle,Analgesic properties of S100A9 C-terminal domain: A mechanism dependent on calciumchannel inhibition. Fundam. Clin. Pharmacol. 23, 427–438 (2009).
51. K. Hargreaves, R. Dubner, F. Brown, C. Flores, J. Joris, A new and sensitive method formeasuring thermal nociception in cutaneous hyperalgesia. Pain 32, 77–88 (1988).
52. J. R. Clapper, G. Moreno-Sanz, R. Russo, A. Guijarro, F. Vacondio, A. Duranti, A. Tontini, S. Sanchini,N. R. Sciolino, J. M. Spradley, A. G. Hohmann, A. Calignano, M. Mor, G. Tarzia, D. Piomelli,Anandamide suppresses pain initiation through a peripheral endocannabinoid mechanism.Nat. Neurosci. 13, 1265–1270 (2010).
53. G. S. Peng, M. F. Wang, C. Y. Chen, S. U. Luu, H. C. Chou, T. K. Li, S. J. Yin, Involvement ofacetaldehyde for full protection against alcoholism by homozygosity of the variant alleleof mitochondrial aldehyde dehydrogenase gene in Asians. Pharmacogenetics 9, 463–476(1999).
eTranslationalMedicine.org 27 August 2014 Vol 6 Issue 251 251ra118 9

R E S EARCH ART I C L E
Acknowledgments:We thank H. Zeng and Y. Chen-Tsai for their help in creating the ALDH2*2knock-in mice at the Stanford University Transgenic Research Center, M. Fridlib with acetalde-hyde measurement help at the Stanford University Biomaterials and Advanced Drug DeliveryCenter, and B. Small for proofreading and artistic assistance. Funding: Fellowship support wasprovided for E.R.G. and V.O.Z. by HL-109212 and 2011/08873-8, respectively. Y.C. is supportedby Fundação de Amparo à Pesquisa do Estado de São Paulo grant 2012/05035-4 and 2013/07467-1. All work from this manuscript was supported by NIH MERIT Award AA11147 to D.M.-R.Author contributions: D.M.-R. and V.O.Z. conceived the project. V.O.Z., E.R.G., C.-H.C., and V.P.G.designed and performed the experiments and carried out data analysis. E.R.G. and D.M.-R. wrotethe manuscript with input from V.O.Z. and Y.C. V.O.Z. and E.R.G. contributed to this projectequally. Competing interests: D.M.-R. and C.-H.C. are the founders of ALDEA Pharmaceuticals,and C.-H.C. is a consultant for the company. However, none of the research in the academiclaboratory is supported by or is reported to the company. A Stanford University patent related tothis manuscript was filed on 23 August 2013 as PCT International application no. US2013/
www.ScienceT
056432. Title of patent is “Methods and Compositions for Treating Pain.” Inventors are V.O.Z.,C.-H.C., and D.M.-R. The other authors declare that they have no competing interests. Data andmaterials availability: All data for this study are provided in the manuscript or SupplementaryMaterials. Materials used for this manuscript are available upon request.
Submitted 15 May 2014Accepted 5 August 2014Published 27 August 201410.1126/scitranslmed.3009539
Citation: V. O. Zambelli, E. R. Gross, C.-H. Chen, V. P. Gutierrez, Y. Cury, D. Mochly-Rosen,Aldehyde dehydrogenase-2 regulates nociception in rodent models of acute inflammatorypain. Sci. Transl. Med. 6, 251ra118 (2014).
ranslationalMedicine.org 27 August 2014 Vol 6 Issue 251 251ra118 10
on
Aug
ust 2
9, 2
014
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
Related Documents











