Feature Review AINTEGUMENTA-LIKE proteins: hubs in a plethora of networks Anneke Horstman 1 , Viola Willemsen 2 , Kim Boutilier 1 , and Renze Heidstra 2 1 Plant Research International, Wageningen University and Research Centre, Droevendaalsesteeg 1, 6708 PB Wageningen, The Netherlands 2 Plant Developmental Biology, Wageningen University and Research Centre, Droevendaalsesteeg 1, 6708 PB Wageningen, The Netherlands Members of the AINTEGUMENTA-LIKE (AIL) family of APETALA 2/ETHYLENE RESPONSE FACTOR (AP2/ERF) domain transcription factors are expressed in all dividing tissues in the plant, where they have central roles in developmental processes such as embryogenesis, stem cell niche specification, meristem maintenance, organ positioning, and growth. When overexpressed, AIL pro- teins induce adventitious growth, including somatic embryogenesis and ectopic organ formation. The Arabi- dopsis (Arabidopsis thaliana) genome contains eight AIL genes, including AINTEGUMENTA, BABY BOOM, and the PLETHORA genes. Studies on these transcription factors have revealed their intricate relationship with auxin as well as their involvement in an increasing number of gene regulatory networks, in which extensive crosstalk and feedback loops have a major role. The AIL transcription factor family in Arabidopsis The eight AINTEGUMENTA-LIKE (AIL) transcription factor genes within the Arabidopsis (Arabidopsis thaliana) genome [1] include AINTEGUMENTA (ANT) [2,3], BABY BOOM (BBM) [4], and the PLETHORA (PLT) genes [5,6] (Box 1; Figure 1), which are all expressed in young, divid- ing tissues in the plant. They have overlapping roles in the establishment and maintenance of meristems, as well as organ initiation and growth (Table 1). A wealth of genetic studies have shown that AIL proteins are master regula- tors of these developmental processes. Loss-of-function combinations and gain-of-function mutants of this gene family show spectacular phenotypes in which meristems or complete organs are missing or arise at ectopic positions. The central role of this family in meristem and organ development extends as far back as mosses [7–9]. In this review, we provide an overview of recent AIL research in the model plant Arabidopsis and point out future chal- lenges in AIL research. AIL function during embryogenesis PLT1, PLT2, AIL6/PLT3, and BBM (collectively called PLT/BBM) genes have a major role in basal patterning of the embryo (Figure 2A). PLT1 and PLT2 gene expression has been described from the octant stage onwards, in the lower tier of the embryo proper [5]. Early embryonic AIL6/ PLT3 and BBM expression has not been reported. Later in embryogenesis, PLT1 expression becomes restricted to the quiescent centre (QC) and surrounding stem cells, where- as the PLT2, AIL6/PLT3, and BBM expression domains are slightly expanded to include the ground tissue and provascular cells. Post-embryonically, the AIL6/PLT3 ex- pression maximum is in the columella stem cells, in con- trast to the QC peak expression observed for PLT1, PLT2, and BBM [5,6]. Only combinations of plt1, plt2, ail6/plt3, and bbm mutants show embryonic abnormalities [5,6]. Subtle defects in plt1;plt2 embryos include enlarged mis-specified QC progenitor cells, and plt1;plt2 seedlings show defective root development, confirming that PLT1 and PLT2 are required for stem cell niche specification [5]. plt1;plt2;ail6/plt3 triple mutant embryos show aberrant organization of the embryonic root pole and seedlings are rootless. plt2;bbm double mutants fail to develop past the early embryo stage, indicating the importance of PLT2 and BBM for embryogenesis [6]. BBM or AIL5/PLT5 overexpression induces the ectopic formation of embryos on the meristem, cotyledons, and first leaves of seedlings [4,10]. Overexpression of PLT1 or PLT2 during embryogenesis ectopically induces root stem cell niches and, in the most extreme cases, can lead to a complete transformation of the embryo towards root iden- tity [5]. In line with this, induced overexpression of PLT2 in seedlings can also produce roots from the shoot apex [6]. Together with the mutant phenotypes, these results sug- gest that BBM, AIL5/PLT5, PLT1, and PLT2 genes act as master regulators for early embryo and root development. This might reflect a specific function for these genes in embryo initiation or the maintenance of cell potency. In most plant species, the initial phase of embryo cell proliferation and morphogenesis is followed by the matu- ration phase, in which cell division stops, storage reserves accumulate, and the seed becomes desiccation tolerant and dormant. In Arabidopsis, the onset of seed maturation is characterized by an increase in the level of the plant hormone abscisic acid (ABA) and by increased expression Review 1360-1385/$ – see front matter ß 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tplants.2013.10.010 Corresponding author: Heidstra, R. ([email protected]). Keywords: AINTEGUMENTA-LIKE; development; embryogenesis; meristem maintenance. 146 Trends in Plant Science, March 2014, Vol. 19, No. 3

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Feature Review
AINTEGUMENTA-LIKE proteins: hubsin a plethora of networksAnneke Horstman1, Viola Willemsen2, Kim Boutilier1, and Renze Heidstra2
1 Plant Research International, Wageningen University and Research Centre, Droevendaalsesteeg 1, 6708 PB Wageningen,
The Netherlands2 Plant Developmental Biology, Wageningen University and Research Centre, Droevendaalsesteeg 1, 6708 PB Wageningen,
The Netherlands
Review
Members of the AINTEGUMENTA-LIKE (AIL) family ofAPETALA 2/ETHYLENE RESPONSE FACTOR (AP2/ERF)domain transcription factors are expressed in all dividingtissues in the plant, where they have central roles indevelopmental processes such as embryogenesis, stemcell niche specification, meristem maintenance, organpositioning, and growth. When overexpressed, AIL pro-teins induce adventitious growth, including somaticembryogenesis and ectopic organ formation. The Arabi-dopsis (Arabidopsis thaliana) genome contains eight AILgenes, including AINTEGUMENTA, BABY BOOM, andthe PLETHORA genes. Studies on these transcriptionfactors have revealed their intricate relationship withauxin as well as their involvement in an increasingnumber of gene regulatory networks, in which extensivecrosstalk and feedback loops have a major role.
The AIL transcription factor family in Arabidopsis
The eight AINTEGUMENTA-LIKE (AIL) transcriptionfactor genes within the Arabidopsis (Arabidopsis thaliana)genome [1] include AINTEGUMENTA (ANT) [2,3], BABYBOOM (BBM) [4], and the PLETHORA (PLT) genes [5,6](Box 1; Figure 1), which are all expressed in young, divid-ing tissues in the plant. They have overlapping roles in theestablishment and maintenance of meristems, as well asorgan initiation and growth (Table 1). A wealth of geneticstudies have shown that AIL proteins are master regula-tors of these developmental processes. Loss-of-functioncombinations and gain-of-function mutants of this genefamily show spectacular phenotypes in which meristems orcomplete organs are missing or arise at ectopic positions.The central role of this family in meristem and organdevelopment extends as far back as mosses [7–9]. In thisreview, we provide an overview of recent AIL research inthe model plant Arabidopsis and point out future chal-lenges in AIL research.
1360-1385/$ – see front matter
� 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tplants.2013.10.010
Corresponding author: Heidstra, R. ([email protected]).Keywords: AINTEGUMENTA-LIKE; development; embryogenesis; meristemmaintenance.
146 Trends in Plant Science, March 2014, Vol. 19, No. 3
AIL function during embryogenesisPLT1, PLT2, AIL6/PLT3, and BBM (collectively calledPLT/BBM) genes have a major role in basal patterning ofthe embryo (Figure 2A). PLT1 and PLT2 gene expressionhas been described from the octant stage onwards, in thelower tier of the embryo proper [5]. Early embryonic AIL6/PLT3 and BBM expression has not been reported. Later inembryogenesis, PLT1 expression becomes restricted to thequiescent centre (QC) and surrounding stem cells, where-as the PLT2, AIL6/PLT3, and BBM expression domainsare slightly expanded to include the ground tissue andprovascular cells. Post-embryonically, the AIL6/PLT3 ex-pression maximum is in the columella stem cells, in con-trast to the QC peak expression observed for PLT1, PLT2,and BBM [5,6]. Only combinations of plt1, plt2, ail6/plt3,and bbm mutants show embryonic abnormalities [5,6].Subtle defects in plt1;plt2 embryos include enlargedmis-specified QC progenitor cells, and plt1;plt2 seedlingsshow defective root development, confirming that PLT1and PLT2 are required for stem cell niche specification [5].plt1;plt2;ail6/plt3 triple mutant embryos show aberrantorganization of the embryonic root pole and seedlings arerootless. plt2;bbm double mutants fail to develop past theearly embryo stage, indicating the importance of PLT2and BBM for embryogenesis [6].
BBM or AIL5/PLT5 overexpression induces the ectopicformation of embryos on the meristem, cotyledons, andfirst leaves of seedlings [4,10]. Overexpression of PLT1 orPLT2 during embryogenesis ectopically induces root stemcell niches and, in the most extreme cases, can lead to acomplete transformation of the embryo towards root iden-tity [5]. In line with this, induced overexpression of PLT2 inseedlings can also produce roots from the shoot apex [6].Together with the mutant phenotypes, these results sug-gest that BBM, AIL5/PLT5, PLT1, and PLT2 genes act asmaster regulators for early embryo and root development.This might reflect a specific function for these genes inembryo initiation or the maintenance of cell potency.
In most plant species, the initial phase of embryo cellproliferation and morphogenesis is followed by the matu-ration phase, in which cell division stops, storage reservesaccumulate, and the seed becomes desiccation tolerant anddormant. In Arabidopsis, the onset of seed maturation ischaracterized by an increase in the level of the planthormone abscisic acid (ABA) and by increased expression

Box 1. AILs form a subgroup within the large AP2/ERF family
The AINTEGUMENTA-LIKE (AIL) proteins are members of the
APETALA 2/ETHYLENE RESPONSE FACTOR (AP2/ERF) domain family
of transcription factors that are found in a phylogenetically wide
group of plants, including moss, algae, gymnosperms, and angios-
perms, and comprise the second largest group of transcription factors
in plants, with up to 200 members in a single genome [1,121,126–133].
The AP2/ERF domain, a DNA binding domain of approximately 60 to
70 amino acids [134], was identified initially in the Arabidopsis AP2
protein [135], and shortly thereafter in four tobacco (Nicotiana
tabacum) ERFs [136]. The AP2/ERF domain was thought to be plant-
specific until the discovery that the same domain exists in HNH (His-
Asp-His) homing endonucleases from viruses, cyanobacteria, ciliates,
and parasitic protists [55,137,138]. Homing endonucleases are
invasive DNA sequences that are usually found in self-splicing introns
or inteins, which can transpose and duplicate themselves within and
between species [139].
The AIL proteins fall under the AP2 subfamily of proteins, which
contain two AP2/ERF domains separated by a linker region [126]. This
AP2 subfamily is divided into the euAP2 lineage, which is characterized
by a miR172 binding motif that lies C-terminal to the AP2/ERF domains,
and the AINTEGUMENTA (ANT) lineage, which is characterized by
signature amino acid insertions in the first and second AP2/ERF
domain. The ANT lineage is further divided into the basalANT and
euANT/AIL lineages (see Figure 1 in main text) on the basis of a number
of conserved amino acid insertions in the first AP2/ERF domain
(euANT1) and the N-terminal region (euANT2–4) of the euANT/AIL
proteins [127,140]. These motifs are generally well conserved among
AILs, both within and between species [127,140,141].
The 3D structure of AP2/ERF proteins has only been resolved for the
single AP2 domain-containing protein ERF1. The 3D solution structure
of ERF1 showed that its AP2 domain forms a three-stranded anti-
parallel b-sheet that lies mostly parallel to a conserved a-helix [142].
Modelling and DNA-binding experiments suggest that the anti-
parallel b-sheet is responsible for the DNA-binding properties of
ERF1 [142], which has since been confirmed for other single AP2
domain proteins in the large ERF1 subfamily [113,143]. Homology
modelling of Arabidopsis ANT using the ERF1 3D structure as a
template suggests that each AP2 repeat of ANT forms an a-helix,
similarly to ERF1. However, unlike ERF1, the first AP2 repeat of ANT is
predicted to contain two b-sheets, at different positions from those in
ERF1, whereas the second AP2/ERF repeat does not seem to form b-
sheets [109]. Both AP2 domains of ANT are required for DNA binding,
and each domain is thought to use different amino acids to contact
the DNA [109,110]. It was proposed that the first AP2 repeat binds to
the 50 part of the target sequence and the second AP2 repeat to the 30
part of the target sequence, with the ANT linker region serving as a
bridge [110]. The importance of the linker region between the AP2/
ERF domains is illustrated by the high conservation of this region
within the AIL gene family and by the observation that mutations in
the linker abolish DNA binding in vitro. These observations led to the
suggestion that the linker region directly binds DNA or serves to
position the AP2 repeats on the DNA [109].
AP2/ERF domain proteins regulate two major processes in plants:
response to stress and control of growth and development. Notably,
AIL proteins have been shown to function exclusively in pathways
related to development.
Review Trends in Plant Science March 2014, Vol. 19, No. 3
of an interwoven network of transcription factors, includ-ing ABSCISIC ACID-INSENSITIVE 3 (ABI3), that togeth-er regulate expression of maturation phase genes.(reviewed in [11,12]). Recently, a heterologous systembased on activation of the Phaseolus vulgaris ABI3 tran-scription factor PvALF and expression of its target genePHASEOLIN (PHAS) identified AIL5/PLT5 as being co-expressed with PHAS [13]. AIL5/PLT5 is bound by PvALFin the presence of ABA and is required for expression ofendogenous seed storage genes in Arabidopsis [13]. It is notknown whether AIL5/PLT5 directly binds to seed storagegenes to activate their expression or whether it acts up-stream of other maturation phase transcription factors,thereby regulating the final phase of embryo development.
AIL–auxin feedback loop
The phytohormone auxin has been shown to play an im-portant part in the formation of the apical–basal axis of theembryo. Auxin binding by its receptors TRANSPORT IN-HIBITOR RESISTANT 1 (TIR1) and TIR1-related proteinspromotes degradation of the AUXIN/INDOLE ACETICACID (Aux/IAA) family proteins that bind to AUXIN RE-SPONSE FACTOR (ARF) family transcription factors,inhibiting ARF transcriptional activity on auxin targetgenes (reviewed in [14]).
The expression of PLT1 and PLT2 is dependent on theredundant action of ARF5/MONOPTEROS (MP) and that ofits close homologue NONPHOTOTROPIC HYPOCOTYL 4(NPH4/ARF7) [5,15]. Although the dynamics of auxin-in-duced PLT1 and PLT2 transcription implies that they are(late) auxin response genes, exogenous application of auxinfails to rescue plt1;plt2 mutants, indicating that auxincannot bypass the requirement for PLT1 and PLT2 [5].Expression of the auxin efflux facilitator PIN-FORMED 4(PIN4) is largely absent in plt1;plt2 embryos, showing that
PIN4 is downstream of PLT1 and PLT2 in the transcrip-tional network. Reduced transcription of PIN1 and PIN3 isalso observed in plt1;plt2;ail6/plt3 triple mutant embryos[6,16]. By contrast, the PLT1 expression domain is expandedto the whole embryo in pin2;pin3;pin4;pin7 mutants.Explanted pin2;pin3;pin4;pin7 embryos show impaired cot-yledon development, and root hairs emerge at apical posi-tions on the seedling. These findings suggest a feedback loopin which PIN proteins, by directing auxin transport andaccumulation, restrict PLT1 and PLT2 expression to thebasal embryo domain to initiate embryonic root specifica-tion. In turn, PLT activity regulates PIN transcription tostabilize the position of the root primordium [16] (Figure 3).
Defects in QC patterning are observed in RHO guanyl-nucleotide exchange factor 7 (RopGEF7) RNA interferencemutants during embryogenesis that correlate with thereduced expression of PIN1, PLT1, and PLT2 [17]. Rop-GEF7, which encodes a RAC/ROP GTPase activator, isexpressed in the same domain as PLT1 and PLT2, and itsexpression is unaffected in plt1;plt2 double mutants [17]. Ithas been suggested that RopGEF7, via RAC/ROP GTPases[18], regulates correct PIN endocytosis, thereby directlyaffecting local auxin concentrations and indirectly affect-ing PLT1 and PLT2 expression to mediate root meristempatterning [17].
Reduced expression of PLT1, PLT2, BBM, and severalPIN genes is observed in JAGGED LATERAL ORGAN(JLO) mutant embryos [19]. Strong jlo mutants are embryolethal, but plants carrying the hypomorphic jlo2 allele pro-duce viable embryos in addition to those resembling mp andbdl mutant embryos [19,20]. JLO might control embryopatterning, either alone or together with its interactionpartner ASYMMETRIC LEAVES 2 (AS2) [21], throughthe auxin-dependent MP–BDL pathway, or by directly reg-ulating PLT/BBM gene expression (Figure 3).
147
98
100
100
100
98
88
41
3151
75
95
975834
630.05
ADAP
AIL1
PLT1
euANT
basalANT
PLT2
PLT7
AIL6/PLT3
ANT
AIL5/PLT5
BBM
Wrinkled
Other
Selaginella
Selaginella
Selaginella
Physcomitrella
Physcomitrella
98
81
TRENDS in Plant Science
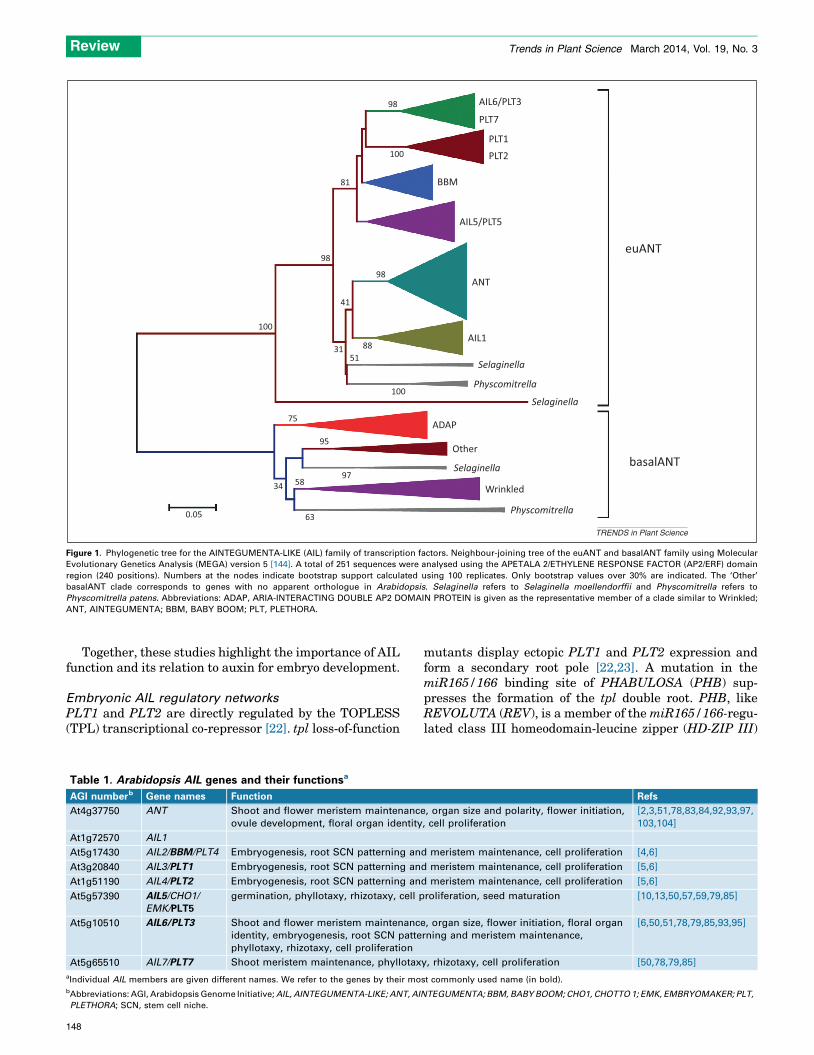
Figure 1. Phylogenetic tree for the AINTEGUMENTA-LIKE (AIL) family of transcription factors. Neighbour-joining tree of the euANT and basalANT family using Molecular
Evolutionary Genetics Analysis (MEGA) version 5 [144]. A total of 251 sequences were analysed using the APETALA 2/ETHYLENE RESPONSE FACTOR (AP2/ERF) domain
region (240 positions). Numbers at the nodes indicate bootstrap support calculated using 100 replicates. Only bootstrap values over 30% are indicated. The ‘Other’
basalANT clade corresponds to genes with no apparent orthologue in Arabidopsis. Selaginella refers to Selaginella moellendorffii and Physcomitrella refers to
Physcomitrella patens. Abbreviations: ADAP, ARIA-INTERACTING DOUBLE AP2 DOMAIN PROTEIN is given as the representative member of a clade similar to Wrinkled;
ANT, AINTEGUMENTA; BBM, BABY BOOM; PLT, PLETHORA.
Review Trends in Plant Science March 2014, Vol. 19, No. 3
Together, these studies highlight the importance of AILfunction and its relation to auxin for embryo development.
Embryonic AIL regulatory networks
PLT1 and PLT2 are directly regulated by the TOPLESS(TPL) transcriptional co-repressor [22]. tpl loss-of-function
Table 1. Arabidopsis AIL genes and their functionsa
AGI numberb Gene names Function
At4g37750 ANT Shoot and flower meristem maintenance
ovule development, floral organ identity
At1g72570 AIL1
At5g17430 AIL2/BBM/PLT4 Embryogenesis, root SCN patterning an
At3g20840 AIL3/PLT1 Embryogenesis, root SCN patterning an
At1g51190 AIL4/PLT2 Embryogenesis, root SCN patterning an
At5g57390 AIL5/CHO1/
EMK/PLT5
germination, phyllotaxy, rhizotaxy, cell p
At5g10510 AIL6/PLT3 Shoot and flower meristem maintenance
identity, embryogenesis, root SCN patte
phyllotaxy, rhizotaxy, cell proliferation
At5g65510 AIL7/PLT7 Shoot meristem maintenance, phyllotax
aIndividual AIL members are given different names. We refer to the genes by their mo
bAbbreviations: AGI, Arabidopsis Genome Initiative; AIL, AINTEGUMENTA-LIKE; ANT, AIN
PLETHORA; SCN, stem cell niche.
148
mutants display ectopic PLT1 and PLT2 expression andform a secondary root pole [22,23]. A mutation in themiR165/166 binding site of PHABULOSA (PHB) sup-presses the formation of the tpl double root. PHB, likeREVOLUTA (REV), is a member of the miR165/166-regu-lated class III homeodomain-leucine zipper (HD-ZIP III)
Refs
, organ size and polarity, flower initiation,
, cell proliferation
[2,3,51,78,83,84,92,93,97,
103,104]
d meristem maintenance, cell proliferation [4,6]
d meristem maintenance, cell proliferation [5,6]
d meristem maintenance, cell proliferation [5,6]
roliferation, seed maturation [10,13,50,57,59,79,85]
, organ size, flower initiation, floral organ
rning and meristem maintenance,
[6,50,51,78,79,85,93,95]
y, rhizotaxy, cell proliferation [50,78,79,85]
st commonly used name (in bold).
TEGUMENTA; BBM, BABY BOOM; CHO1, CHOTTO 1; EMK, EMBRYOMAKER; PLT,
(A)
(F)(E)
(D)
(B)
(C) Peri
Stemcells
Col
Stele EnCor
EpiLrc
Vasc Gt
Stage I Stage II
AIL6/PLT3,AIL5/PLT5,PLT7
AIL6/PLT3,AIL5/PLT5,PLT7
ANT
ANT
ANT
AIL6/PLT3
AIL6/PLT3
ANTAIL6/PLT3
PLT7
PLT1,PLT2,AIL6/PLT3,BBM
PLT1,PLT2,AIL6/PLT3,BBM
TRENDS in Plant Science
137°
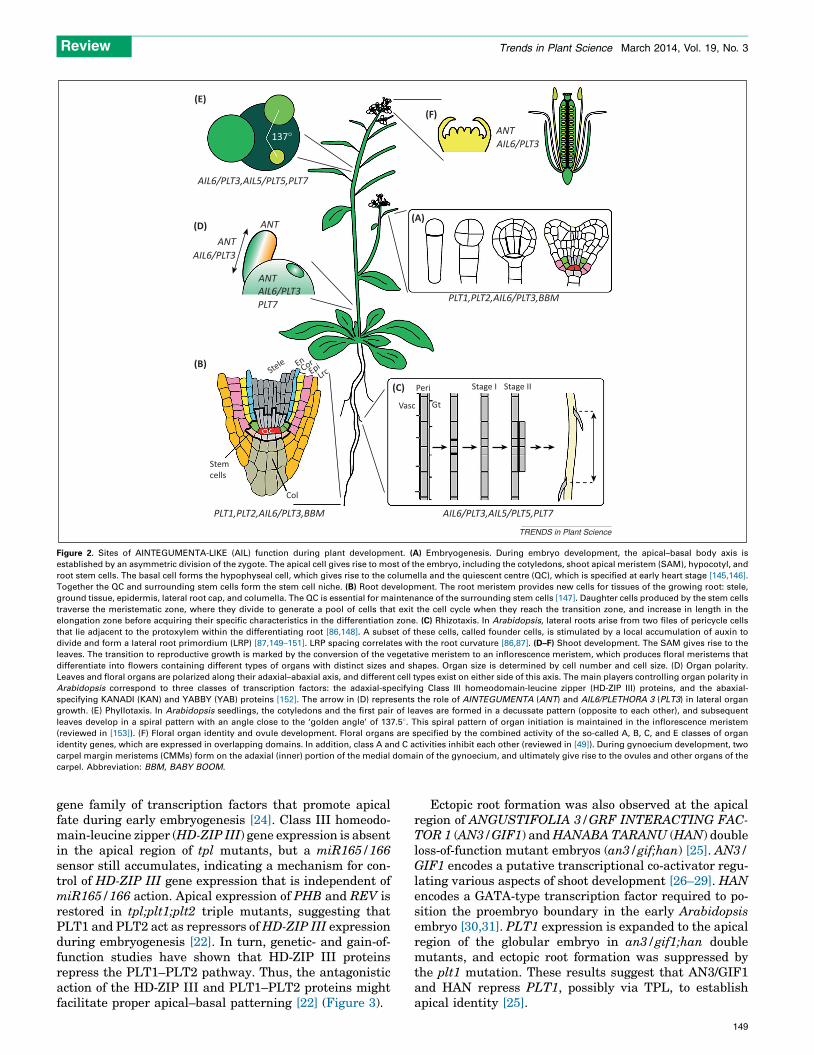
Figure 2. Sites of AINTEGUMENTA-LIKE (AIL) function during plant development. (A) Embryogenesis. During embryo development, the apical–basal body axis is
established by an asymmetric division of the zygote. The apical cell gives rise to most of the embryo, including the cotyledons, shoot apical meristem (SAM), hypocotyl, and
root stem cells. The basal cell forms the hypophyseal cell, which gives rise to the columella and the quiescent centre (QC), which is specified at early heart stage [145,146].
Together the QC and surrounding stem cells form the stem cell niche. (B) Root development. The root meristem provides new cells for tissues of the growing root: stele,
ground tissue, epidermis, lateral root cap, and columella. The QC is essential for maintenance of the surrounding stem cells [147]. Daughter cells produced by the stem cells
traverse the meristematic zone, where they divide to generate a pool of cells that exit the cell cycle when they reach the transition zone, and increase in length in the
elongation zone before acquiring their specific characteristics in the differentiation zone. (C) Rhizotaxis. In Arabidopsis, lateral roots arise from two files of pericycle cells
that lie adjacent to the protoxylem within the differentiating root [86,148]. A subset of these cells, called founder cells, is stimulated by a local accumulation of auxin to
divide and form a lateral root primordium (LRP) [87,149–151]. LRP spacing correlates with the root curvature [86,87]. (D–F) Shoot development. The SAM gives rise to the
leaves. The transition to reproductive growth is marked by the conversion of the vegetative meristem to an inflorescence meristem, which produces floral meristems that
differentiate into flowers containing different types of organs with distinct sizes and shapes. Organ size is determined by cell number and cell size. (D) Organ polarity.
Leaves and floral organs are polarized along their adaxial–abaxial axis, and different cell types exist on either side of this axis. The main players controlling organ polarity in
Arabidopsis correspond to three classes of transcription factors: the adaxial-specifying Class III homeodomain-leucine zipper (HD-ZIP III) proteins, and the abaxial-
specifying KANADI (KAN) and YABBY (YAB) proteins [152]. The arrow in (D) represents the role of AINTEGUMENTA (ANT) and AIL6/PLETHORA 3 (PLT3) in lateral organ
growth. (E) Phyllotaxis. In Arabidopsis seedlings, the cotyledons and the first pair of leaves are formed in a decussate pattern (opposite to each other), and subsequent
leaves develop in a spiral pattern with an angle close to the ‘golden angle’ of 137.58. This spiral pattern of organ initiation is maintained in the inflorescence meristem
(reviewed in [153]). (F) Floral organ identity and ovule development. Floral organs are specified by the combined activity of the so-called A, B, C, and E classes of organ
identity genes, which are expressed in overlapping domains. In addition, class A and C activities inhibit each other (reviewed in [49]). During gynoecium development, two
carpel margin meristems (CMMs) form on the adaxial (inner) portion of the medial domain of the gynoecium, and ultimately give rise to the ovules and other organs of the
carpel. Abbreviation: BBM, BABY BOOM.
Review Trends in Plant Science March 2014, Vol. 19, No. 3
gene family of transcription factors that promote apicalfate during early embryogenesis [24]. Class III homeodo-main-leucine zipper (HD-ZIP III) gene expression is absentin the apical region of tpl mutants, but a miR165/166sensor still accumulates, indicating a mechanism for con-trol of HD-ZIP III gene expression that is independent ofmiR165/166 action. Apical expression of PHB and REV isrestored in tpl;plt1;plt2 triple mutants, suggesting thatPLT1 and PLT2 act as repressors of HD-ZIP III expressionduring embryogenesis [22]. In turn, genetic- and gain-of-function studies have shown that HD-ZIP III proteinsrepress the PLT1–PLT2 pathway. Thus, the antagonisticaction of the HD-ZIP III and PLT1–PLT2 proteins mightfacilitate proper apical–basal patterning [22] (Figure 3).
Ectopic root formation was also observed at the apicalregion of ANGUSTIFOLIA 3/GRF INTERACTING FAC-TOR 1 (AN3/GIF1) and HANABA TARANU (HAN) doubleloss-of-function mutant embryos (an3/gif;han) [25]. AN3/GIF1 encodes a putative transcriptional co-activator regu-lating various aspects of shoot development [26–29]. HANencodes a GATA-type transcription factor required to po-sition the proembryo boundary in the early Arabidopsisembryo [30,31]. PLT1 expression is expanded to the apicalregion of the globular embryo in an3/gif1;han doublemutants, and ectopic root formation was suppressed bythe plt1 mutation. These results suggest that AN3/GIF1and HAN repress PLT1, possibly via TPL, to establishapical identity [25].
149

OBE1;OBE2
PLT1;PLT2 PLT3/AIL6ANT
MP;NPH4
Auxin
YUCCAPIN
AN3/GIF1
JLO(-AS2)
BBM
TPL HD-ZIP III
PLT5/AIL5PLT7
ARF7;ARF19
TPST
RGF/GLV
AB14
GA
JA
MYC2
GCN5ADA2b
PKLAG
AP3
SLR/IAA14
ARGOS
ADF9
HAN
ABA
RopGEF7
TRENDS in Plant Science
Figure 3. AINTEGUMENTA-LIKE (AIL) gene regulatory networks. Genes in purple, green or orange are directly associated with the jasmonate (JA), auxin, or abscisic acid
(ABA) pathways, respectively. Unbroken black lines represent direct interactions, whereas broken lines show possible interactions. Abbreviations: ABI4, ABA INSENSITIVE
4; ADF9, ACTIN DEPOLYMERIZING FACTOR 9; AG, AGAMOUS; AN3/GIF1, ANGUSTIFOLIA 3/GRF INTERACTING FACTOR 1; ANT, AINTEGUMENTA; AP3, APETALA 3; ARF,
AUXIN RESPONSE FACTOR; ARGOS, AUXIN-REGULATED GENE INVOLVED IN ORGAN SIZE; BBM, BABY BOOM; GA, gibberellic acid; GCN5, GENERAL CONTROL NON-
REPRESSIBLE 5; GLV, GOLVEN; HAN, HANABA TARANU; JLO, JAGGED LATERAL ORGAN; MP, MONOPTEROS; NPH4, NONPHOTOTROPIC HYPOCOTYL 4; OBE, OBERON;
PIN, PIN-FORMED; PKL, PICKLE; PLT, PLETHORA; RGF, ROOT GROWTH FACTOR; RopGEF7, RHO guanyl-nucleotide exchange factor 7; SLR, SOLITARY ROOT: TPL,
TOPLESS; TPST, TYROSYLPROTEIN SULFOTRANSFERASE.
Review Trends in Plant Science March 2014, Vol. 19, No. 3
Root development and meristem maintenancePLT/BBM expression patterns set up during embryogene-sis are maintained during post-embryonic root develop-ment (Figure 2B), where they show partly overlappinggradients of protein accumulation, with the highest pro-teins levels in the stem cell niche [5,6]. Concomitantly, theSHORT ROOT (SHR) transcription factor is expressed inthe stele and moves to the surrounding cells, including theQC, where it activates SCARECROW (SCR) [32,33].plt1;plt2, shr and scr mutants display loss of (different)QC identity markers and premature termination of rootgrowth [5,34]. PLT1 transcription is not affected in shr andscr mutants, and PLT1–PLT2 activity is not required forSHR or SCR expression. Thus, QC specification requiresthe parallel action of PLT1–PLT2 and SHR–SCR path-ways [5].
Additive activities of AIL genes
The QC is the site of the root auxin maximum [35–37],which is generated by local biosynthesis [38–42] andthrough the activity of directional auxin transporters suchas PIN proteins [16,43–45]. The PIN-mediated, root-direct-ed auxin transport in the vasculature and the shoot-direct-ed transport in the lateral root cap and epidermis arethought to generate an auxin gradient in the root that iscrucial for root meristem size and maintenance[16,43,46,47]. The proposed root auxin gradient correlateswith an expression gradient of PLT/BBM proteins that actin a largely additive fashion [5,6]. First, stepwise loweringof PLT/BBM gene dosage results in increasingly enhancedroot phenotypes. Second, complementing plt1;plt2 mutantswith a shortened gradient of PLT2 expression results inseverely decreased root and meristem sizes, but rescuesthe stem cell niche. Third, expression of PLT2 in thetransition zone of the plt1;plt2 mutant meristem prolongs
150
meristem activity but fails to maintain the stem cell niche.Finally, inducible PLT2 overexpression results in continu-ous growth of the meristematic zone while inhibiting cellexpansion at the elongation zone [6]. Simultaneously low-ering RETINOBLASTOMA-RELATED (RBR) levels, as anindependent stem cell input [48], and inducing overexpres-sion of PLT2 increases stem cell numbers and activity [6].Thus, high PLT/BBM levels in the QC are required tospecify and maintain the root stem cell niche, intermediatelevels are required for the proliferation of root meristemcells, and exit from the meristem to differentiation requiresPLT/BBM levels to drop below a certain threshold [6].Together, the auxin–PLT/BBM gradient can provide aconcentration-based mechanism for specification andmaintenance of root stem cells, as well as for regulatingthe proliferation, elongation, and differentiation of cells.
Translating auxin to AIL expression
Several studies implicate ARFs as general upstream med-iators of the auxin–AIL pathway during embryogenesis(MP and NPH4), flower primordia initiation and ovuledevelopment (MP), and lateral root formation (ARF7 andARF19) [5,49–51] (see below).
Auxin also positively regulates TYROSYLPROTEINSULFOTRANSFERASE (TPST) and several tyrosine-sul-fated peptide encoding ROOT GROWTH FACTOR (RGF)/GOLVEN (GLV) genes that are highly expressed in the rootstem cell niche [52–54]. tpst mutants have pleiotropicphenotypes, including short roots, additional QC cells,and differentiation of columella stem cells. The observedroot defects correlate with impaired expression of auxinbiosynthesis genes and PLT1 and PLT2 transcript andprotein levels. tpst root defects were partially restored inthe presence of RGF1 peptide. Induced overexpression ofPLT2 also partially rescued tpst mutant root defects,

Review Trends in Plant Science March 2014, Vol. 19, No. 3
whereas plt1;plt2 mutant roots could not be complementedby addition of RGF1 peptide [52,53]. In addition, RGF1-treated roots show an expanded PLT2 protein expressiondomain without associated changes in transcription [52].Together these results suggest TPST-sulfated RGF pep-tides link the auxin pathway to transcriptional and post-transcriptional regulation of PLT/BBM in root stem cellmaintenance (Figure 3).
AIL genes integrate multiple hormonal inputs
Germination results in emergence of the radicle (hypocotyl +embryonic root) from the seed through cell enlargement andactivation of the embryonic root meristem [55]. This processis regulated by environmental factors, such as nutrients,light, and temperature, as well as by the endogenous planthormones ABA and gibberellic acid (GA) [56]. The ail5/plt5mutant was first identified (and named chotto1; Table 1) as amutant showing reduced sensitivity to ABA during germi-nation [57]. Increased AIL5/PLT5 expression in the radicleof imbibed seeds requires the transcription factor ABAINSENSITIVE 4 (ABI4), a key component of ABA-mediatedsignalling in seeds [58], and genetic analyses confirms thatAIL5/PLT5 acts downstream of ABI4 [59] (Figure 3). Fresh-ly harvested ail5/plt5 mutant seeds show reduced primaryseed dormancy, which was recently also shown for abi4 [59–61]. However, neither ABA nor GA seems to regulate AIL5/PLT5 expression [59,61]. Genetic analysis further showsthat the ail5/plt5 mutant background partially restores GAbiosynthesis and the associated delayed germination phe-notype in an ABA over-accumulating mutant. However,ail5/plt5 is not able to restore germination in a GA-deficientmutant [61]. Finally, ail5/plt5 seedlings display partialinsensitivity to excess nitrate independent of ABI4. Theseresults indicate that AIL5/PLT5 integrates multiple andindependent inputs towards regulation of germination andseedling growth.
Jasmonate (JA) negatively affects root growth in a dose-dependent manner resulting in reduced root cell sizes, areduced meristem size and activity, and QC divisionsfollowed by columella stem cell differentiation [62]. TheJA growth inhibitory effect is mediated via its receptorCOI1 [63,64]. The downstream MYC2 transcription factor[65,66] acts, at least in part, through direct transcriptionaldownregulation of the PLT1 and PLT2 genes, independentof the auxin pathway [62] (Figure 3). The effects of JAapplication are only partly reduced in plt1;plt2 mutant andPLT2 overexpression roots, suggesting that other PLT/BBM members also play a part in mediating JA effects [62].
Regulation of PLT expression at the chromatin level
Chromatin factors affect key regulators of cell proliferationand stem cell maintenance, such as WUS, WOX5, PLT1,and PLT2 genes [67–72]. The histone acetyltransferaseGENERAL CONTROL NON-REPRESSIBLE 5 (GCN5)and its associated factor ADA2b were found to regulatePLT1 and PLT2 gene expression levels and genetic analy-sis confirmed that GCN5 and ADA2b act in the PLT1–PLT2 pathway [71] (Figure 3). In addition, the root meri-stem of gcn5 mutants, which is gradually lost owing tocompromised stem cell niche maintenance, is partiallyrescued by induced overexpression of PLT2 [71].
Mutation of the chromatin remodelling factor PICKLE(PKL) causes short primary roots with a reduced meristemsize due to impaired stem cell niche activity. By contrast,mutation of the Polycomb-group (PcG) gene CURLY LEAF(CLF) results in longer roots with an increased meristemsize, which is associated with increased stem cell activity[67]. These mutants do not show altered auxin accumula-tion. Rather, their phenotypes correlated with, and wereshown genetically to be caused by, decreased PLT1–PLT2expression or increased WOX5 expression, respectively.Decreased expression levels of root stem cell and meristemmarker genes in pkl correlated with increased levels of(PcG-mediated) trimethylation of histone H3 on lysine 27(H3K27me3) at their promoter regions, indicating that rootstem cell niche activity is regulated by the antagonisticactivities of PcG proteins and PKL [67]. The similar phe-notypes of gcn5 and pkl suggest linked gene activities.PKL-mediated remodelling might allow the recruitmentof a GCN5–ADA2b-containing histone acetyltransferasecomplex, thereby generating a suitable chromatin statefor the accurate interpretation of auxin-dependent PLTsignals and reinforcing stem cell fate [67]. Such a stem cell-specific chromatin state would be analogous to that ob-served in animal systems [73,74].
The OBERON 1 (OBE1) and OBE2 genes encode planthomeodomain (PHD)-finger proteins, so-called epigeneticreaders that recognize and bind to both methylated andunmodified histone H3 tails [75,76]. The OBE proteins actredundantly in MP-dependent embryonic root initiation[76,77]. obe1;obe2 double mutants resemble mp mutantsand lack PLT1 and PLT2 expression, whereas MP expres-sion is unaffected [76]. Because PLT1 and PLT2 expressionis dependent on MP [5], OBE1 and OBE2 might act tocontrol embryonic root meristem formation downstream orat the level of MP, and in this way control PLT1 and PLT2expression [76] (Figure 3).
Shoot and flower meristem maintenanceANT, AIL6/PLT3, and PLT7 are expressed in distinct butoverlapping domains in the inflorescence and vegetativeshoot meristems. AIL6/PLT3 and PLT7 are expressedthroughout the meristem, but their expression is increasedin the peripheral zones and in the central zone, respective-ly [78,79]. By contrast, ANT is expressed exclusively in theperipheral zones and marks the cryptic bract region of thefloral meristem [80,81]. The ant;ail6/plt3;plt7 triple mu-tant shoot stops growing after the production of a fewleaves owing to reduced cell divisions in the meristemand differentiation of the meristematic cells, showing thatANT, AIL6/PLT3, and PLT7 genes are required for shootapical meristem (SAM) maintenance [78] (Figure 2D). Theant and ail6/plt3 mutations enhance wus and shoot mer-istemless (stm) phenotypes, whereas plt7 partially rescuesthese phenotypes, indicating that ANT, AIL6/PLT3, andPLT7 do not function in a strictly redundant fashion [78].
Both the ant and ap2 single mutants have fewer cells intheir floral meristems (Figure 2F), resulting in a reducednumber of floral organs [82,83]. When ap2 and ant muta-tions are combined, sepal, petal, and stamen formation iscompletely abolished [2]. This organ loss is likely to be aresult of reduced proliferation owing to ectopic expression
151

Review Trends in Plant Science March 2014, Vol. 19, No. 3
of the growth-repressing class C MADS-box gene AGA-MOUS (AG) [2,84].
Organ developmentPhyllotaxy and rhizotaxy
Cotyledons and the first pair of leaves are formed oppositeto each other in Arabidopsis, and subsequent leaves devel-op in a spiral pattern (Figure 2E). Unlike AIL6/PLT3 andPLT7, AIL5/PLT5 is expressed in a largely uniform man-ner throughout the entire SAM [78,79]. In the ail5/plt5;ail6/plt3;plt7 triple mutant, the switch to the spiralphyllotactic pattern is delayed by a few leaf pairs, withdouble mutant combinations showing lower penetrance[79]. The spiral pattern in the inflorescence meristem(Figure 2E) is also affected in this triple mutant, whichshows a tendency to develop successive siliques at angles of�908 or �1808 instead of at 137.58 in wild-type Arabidop-sis. Reducing PIN1 levels leads to the same defects in thephyllotactic pattern as reduced AIL/PLT expression [79].It was shown recently by ail5/plt5;ail6/plt3;plt7 comple-mentation experiments that AIL/PLT-induced auxin bio-synthesis genes, YUCCA 1 (YUC1) and YUC4, mediatehigh auxin levels in the central zone that are required forthe correct spiral phyllotactic pattern [85].
Lateral root primordia (LRPs) are distributed along theArabidopsis root, and their spacing correlates with the rootcurvature [86,87] (Figure 2C). ARF7 and ARF19, togetherwith the gene encoding SOLITARY ROOT (SLR)/IAA14,which represses these ARFs, are all expressed in the vascu-lar tissue and lateral root initiation sites [88–91]. arf7;arf19double mutant roots are defective in the first asymmetricdivision that leads to LRP formation and almost completelylack lateral roots. AIL6/PLT3, AIL5/PLT5, and PLT7 areexpressed prior to the first founder cell division, and theirexpression is absent in arf7;arf19 mutants [50]. Outgrowthof LRP is severely impaired in ail6/plt3;plt7 double andail6/plt3;ail5/plt5;plt7 triple mutants, and LRPs are fre-quently grouped in longitudinal or radial clusters. Comple-mentation experiments reveal that expression of AIL6/PLT3, AIL5/PLT5, or PLT7 from their endogenous promo-ters rescues LRP emergence more readily than the cluster-ing phenotype, suggesting distinct targets and/or dosagedependency reminiscent of PLT/BBM function in the pri-mary root [6,50]. Overexpression of AIL6/PLT3, AIL5/PLT5, or PLT7 in arf7;arf19 can overcome the block inLRP formation. These results indicate that the AIL6/PLT3, AIL5/PLT5, and PLT7 genes are downstream com-ponents of the ARF7- and ARF19-mediated auxin responsemodule [50] (Figure 3), although they do not seem to bedirect targets of ARF7 and ARF19 [89,91].
Despite the differences in the processes of shoot and rootprimordia initiation, the involvement of the same threeAIL proteins in both processes suggests that similar mech-anisms regulate plant architecture in different organs.
Organ size
Loss-of-function ant mutants have smaller leaves andflowers as a result of having fewer cells, whereas over-expression of ANT prolongs the cell proliferation period,leading to larger leaves and flowers with more and/orlarger cells [83,92] (Figure 2D). The reduced cell prolifera-
152
tion observed in ant is enhanced in the ant;ail6/plt3 doublemutant, which has even smaller leaves [93]. Overexpres-sion of the auxin-inducible gene AUXIN-REGULATEDGENE INVOLVED IN ORGAN SIZE (ARGOS) prolongsANT expression and leads to the formation of larger organs[94]. This effect was lost in the ant mutant, suggesting thatARGOS functions downstream of auxin and upstream ofANT in organ size control [94] (Figure 3).
An increase in floral organ size due to increased cellnumber was also observed in AIL5/PLT5 and AIL6/PLT3overexpression lines [1,95]. However, high expressionlevels of AIL6/PLT3 block cellular differentiation, result-ing in floral organs with altered morphology [95]. The verysmall and aberrantly shaped sepals of these AIL6/PLT3overexpression lines are unable to cover the entire flowerbud, a phenotype that was also reported in BBM over-expression lines [4,95]. Together with the sustained meri-stem cell proliferation observed on PLT2 overexpression[6], these results indicate that AIL transcription factorsregulate the balance between cell proliferation and differ-entiation.
Organ polarity
Double and triple mutant combinations of ant with loss-of-function alleles of the polarity genes FILAMENTOUSFLOWER (FIL) and YABBY 3 produce dwarfed seedlingsthat develop small, narrow leaves with loss of both adaxialand abaxial epidermal cell characteristics, which are notobserved in single mutants [96,97]. These defects are alsopresent in some floral organs. The expression of the adaxialspecifying HD-ZIP III gene PHB is reduced in fil;antflowers, suggesting that ANT and FIL act together topromote organ polarity (Figure 2D) by activating PHB[97] (Figure 3).
Flower and floral organ developmentFloral initiation
LEAFY (LFY) encodes a transcriptional regulator thatpromotes the transition to flowering and also specifiesfloral meristem identity [98–101]. Auxin treatmentinduces higher LFY gene expression and results in preco-cious flower formation. Auxin response elements in thepromoter of LFY are bound by the MP and BDL proteins[51]. Initiation of flower primordia from the inflorescencemeristem (Figure 2F) is known to rely on MP; mp mutantsform naked inflorescence pins lacking flowers, phenocopy-ing loss-of-function mutants in the PIN1 auxin effluxcarrier [45,102]. MP and LFY expression overlaps withthat of the ANT and AIL6/PLT3 genes, and MP was shownto bind to the promoters and activate expression of ANTand AIL6/PLT3 [51]. The pin-like inflorescences formed inthe ant;ail6/plt3;lfy triple mutant and the additive abilityof ANT and LFY overexpression to rescue flower primordi-um initiation in the hypomorphic mp-S319 mutant indi-cate that ANT and LFY function in parallel pathwaysdownstream of MP in the initiation of flower primordia[51] (Figure 3). As with AIL proteins, LFY feeds backpositively into the auxin pathway [51,79,85]. MP-depen-dent upregulation of LFY, ANT, and AIL6/PLT3, togetherwith their positive feedback to the auxin pathway, mightcommit cells to flower primordium formation [51].

Review Trends in Plant Science March 2014, Vol. 19, No. 3
Floral organ identity
The petals of the ant single mutant develop stomata, whichare absent in wild-type petals, indicating partial loss ofpetal identity in this mutant [84]. A more severe phenotypein which the petals become partially stamenoid was ob-served in the hypomorphic ap2-1;ant double mutant [84].Expression of AG was extended to the second whorl in theap2-1;ant mutant, where it caused the partial homeotictransformation [84]. Single ail6/plt3 mutants develop nor-mally, but flowers of the ant;ail6/plt3 double mutant havereduced petal, stamen, and carpel identities and consistprimarily of small sepals, filamentous organs, and unfusedcarpel valves, showing that ANT and AIL6/PLT3 functionredundantly in determining floral organ identity [93](Figure 2F). This phenotype correlates with altered expres-sion of the floral organ identity genes APETALA 3 (AP3, B-class) and AG (C-class) [93], and suggests their regulationby ANT and AIL6/PLT3 (Figure 3).
Ovule development
The number of ovule primordia in the ant mutant isreduced owing to carpel margin meristem (CMM) defects.Also, the integuments of ant ovules do not initiate properlyand female gametophyte development is disrupted, lead-ing to female sterility [2,3,103,104]. The CUP SHAPEDCOTYLEDON 1 (CUC1) and CUC2 genes act in parallel toANT in controlling the number of developing ovule primor-dia [49]. ANT, CUC1, and CUC2 are downregulated inhypomorphic mp-S319 mutant pistils, which do not devel-op ovules owing to the absence of carpel margin tissue.Binding of MP to the ANT, CUC1, and CUC2 genomicregions was confirmed in this tissue [49]. Thus, MP acts asan upstream regulator of ANT during ovule development(Figure 3), reminiscent of its role in regulating AIL geneexpression during embryogenesis and floral meristem ini-tiation [5,49,51].
HUELLENLOS (HLL) and ANT have overlapping rolesin ovule primordium outgrowth [105]. The transcriptionalco-regulators SEUSS (SEU) and LEUNIG (LUG) showpartial redundancy with ANT in the control of medialdomain development [106,107], and these defects are as-sociated with reduced expression levels of the HD-ZIP IIIgenes REV and PHB [106]. Double mutant ant;rev gynoeciaexhibit loss of CMM-derived tissue, which is correlatedwith lower expression of PHB [108]. These studies showthat ANT has a central role in ovule development(Figure 2F) through the activation of HD-ZIP III genes(Figure 3).
AIL target genesAlthough many genetic interactions have been described forAIL genes, our knowledge about target genes that aredirectly regulated by AIL binding remains limited. BothAPETALA 2/ETHYLENE RESPONSE FACTOR (AP2/ERF) domains of ANT are required for DNA binding, andeach domain is thought to use different amino acids tocontact the DNA [109,110] (Box 1). An ANT DNA bindingmotif was determined using SELEX, an in vitro oligonucle-otide selection method, which yielded the consensus se-quence gCAC(A/G)N(A/T)TcCC(a/g)ANG(c/t) [110]. AIL5/PLT5 also binds to the ANT consensus sequence in vitro
[61], suggesting that the observed overlapping functions ofAIL proteins is realized through a set of common targetgenes. The ANT DNA binding motif is different from that ofother single AP2/ERF proteins, which bind to short GC-richboxes [111–113].
Currently, only the targets of BBM have been identified.Using a microarray approach in which the BBM proteinwas inducibly activated in the presence of the translationalinhibitor cycloheximide to identify direct targets, it wasshown that the vast majority of target genes are upregu-lated on DNA binding, suggesting that BBM acts mainly asa transcriptional activator [114]. BBM was reported tointeract with TPL-related (TPR) corepressors in yeast[115], suggesting that BBM also acts as a transcriptionalrepressor. AP2 has also been shown to form complexes withTOPLESS (TPL) to repress the floral organ identity genesAG, AP3, PI, and SEP3 [116], suggesting that similarprotein–protein complexes are co-opted by different groupsof AP2-family proteins.
The functional diversity of BBM targets suggests thatthis protein functions in a wide range of developmentalpathways, which is consistent with the large number ofmutant studies in which BBM and other AIL genes wereshown to be regulated incorrectly. One of the few function-ally characterized BBM targets is RGF8/GLV6, whichencodes a homologue of the RGF1 signalling peptide thathas been shown to regulate PLT1 and PLT2 expression[52]. This suggests a positive feedback loop between RGF/GLV and PLT/BBM genes (Figure 3) that ensures suffi-ciently high PLT/BBM levels to preserve the meristem.Another gene that is directly upregulated by BBM isACTIN DEPOLYMERIZING FACTOR 9 (ADF9) [114],which encodes an ADF/cofilin cytoskeletal protein[117,118]. ADF9 is required for hormone-mediated cellproliferation in vitro [119]. Notably, only a few minutesof inducible BBM overexpression is sufficient to stimulatereorganization of the actin cytoskeleton. ADF9 thereforeprovides a potential link between the actin cytoskeletonand BBM-mediated meristematic growth. Finally, it wasshown that BBM also stimulates its own expression, sug-gesting it functions in a positive auto-regulatory feedbackloop to control its own activity [114]. BBM targets supporta role for AIL proteins in the regulation of cell proliferationand differentiation.
Concluding remarks and future prospectsAlmost 20 years of AIL research in Arabidopsis has shownthat these transcription factors are crucial for stem cellniche specification, meristem maintenance, organ initia-tion, and growth, and that they function as master reg-ulators of embryogenesis and root formation. At present,functions have been ascribed to all Arabidopsis AIL genesexcept AIL1. Research on AIL expression and function inother species is still at an early stage. Analysis of theexpression patterns of ANT-like genes from different spe-cies suggest that they are [120–122] or are not [120,123]conserved with the Arabidopsis ANT gene. Nevertheless,functional studies on AIL proteins from different speciesindicate that they also regulate stem cell specification andplant developmental processes, as in Arabidopsis. Forexample, in poplar (Populus spp.), ANT-like AILs have
153

Review Trends in Plant Science March 2014, Vol. 19, No. 3
been shown to control meristem activity during adventi-tious rooting [9] and to directly feed into the cell cyclemachinery to establish growth cessation on exposure toshort days [8]. Similarly, in the moss Physcomitrellapatens, four AIL genes are indispensable for the formationof one type of moss stem cell, the apical gametophore cell[7].
AIL proteins interact with auxin pathways throughoutplant development and at multiple levels, including viaARFs and through PIN feedback loops (Figure 3). Futurestudies will undoubtedly reveal additional auxin–AILrelationships, as well as new relationships with otherhormones, signalling pathways and chromatin-levelchanges. Factors involved in regulating AIL expressionduring the earliest stages of embryogenesis have not beenidentified, even though at least two AILs, BBM and PLT2,are required for progression through early embryogenesis.
Specific AIL family members seem to be expressed andfunction in either the root or shoot meristem. This divisioncould reflect protein subfunctionalization, which is alsosuggested by the different AIL–HD-ZIP III genetic inter-actions that appear as a general module in the regulationof several developmental processes (Figure 3). Alternative-ly, the regional expression and interaction of AIL proteinswith locally expressed transcriptional co-activators orrepressors might be responsible for this differential effect.Identification of the protein complexes in which AIL pro-teins act might shed light on this matter. In addition,promoter swapping to express root AILs in the shootand vice versa may reveal how much of this apparentsubfunctionalization is due to differences in protein char-acteristics versus gene expression patterns.
One aspect of AIL function is their dosage-dependentregulation of developmental processes, as observed duringroot development. This dosage-dependency could be me-diated by quantitative differences in the activation oftarget genes containing AIL-specific DNA binding motifs.Alternatively, AIL proteins could exert their dosage-de-pendent regulation via differences in binding affinity forDNA motifs that are present in common target genepromoters. The fact that both ANT and AIL5/PLT5 canbind the same DNA motif suggests a common set of down-stream targets. Elucidating the targets and DNA bindingsites of additional AIL proteins might clarify either of theabove scenarios.
A poorly understood aspect of AIL function is theirability to induce totipotency and pluripotency when over-expressed. This property offers new possibilities toimprove the efficiency of plant regeneration for a rangeof biotechnology applications. For example, the ability ofBBM to induce somatic embryogenesis was used toimprove Chinese white poplar and sweet pepper (Capsi-cum anuum) transformation [124,125]. More importantly,a better understanding of how AIL proteins function willprovide a framework for understanding how regenerationis regulated in vitro and during normal plant development.
AcknowledgementsWe thank Gabino Sanchez-Perez for constructing an updated phyloge-netic tree for the AIL family, and Ben Scheres, Gerco Angenent, andStephen Grigg for valuable comments on the manuscript. A.H. was
154
supported by a Technology Top Institute Green Genetics project. R.H.was supported by a Netherlands Genomics Initiative Horizon grant (no.93516054) from The Netherlands Organization for Scientific Research.
References1 Nole-Wilson, S. et al. (2005) AINTEGUMENTA-like (AIL) genes are
expressed in young tissues and may specify meristematic or division-competent states. Plant Mol. Biol. 57, 613–628
2 Elliott, R.C. et al. (1996) AINTEGUMENTA, an APETALA2-like geneof Arabidopsis with pleiotropic roles in ovule development and floralorgan growth. Plant Cell 8, 155–168
3 Klucher, K.M. et al. (1996) The AINTEGUMENTA gene ofArabidopsis required for ovule and female gametophytedevelopment is related to the floral homeotic gene APETALA2.Plant Cell 8, 137–153
4 Boutilier, K. et al. (2002) Ectopic expression of BABY BOOM triggers aconversion from vegetative to embryonic growth. Plant Cell 14,1737–1749
5 Aida, M. et al. (2004) The PLETHORA genes mediate patterning of theArabidopsis root stem cell niche. Cell 119, 109–120
6 Galinha, C. et al. (2007) PLETHORA proteins as dose-dependentmaster regulators of Arabidopsis root development. Nature 449,1053–1057
7 Aoyama, T. et al. (2012) AP2-type transcription factors determinestem cell identity in the moss Physcomitrella patens. Development139, 3120–3129
8 Karlberg, A. et al. (2011) Short day-mediated cessation of growthrequires the downregulation of AINTEGUMENTALIKE1transcription factor in hybrid aspen. PLoS Genet. 7, e1002361
9 Rigal, A. et al. (2012) The AINTEGUMENTA LIKE1 homeotictranscription factor PtAIL1 controls the formation of adventitiousroot primordia in poplar. Plant Physiol. 160, 1996–2006
10 Tsuwamoto, R. et al. (2010) Arabidopsis EMBRYOMAKER encodingan AP2 domain transcription factor plays a key role in developmentalchange from vegetative to embryonic phase. Plant Mol. Biol. 73,481–492
11 Braybrook, S.A. and Harada, J.J. (2008) LECs go crazy in embryodevelopment. Trends Plant Sci. 13, 624–630
12 Jia, H. et al. (2013) Distinct roles of LAFL network genes in promotingthe embryonic seedling fate in the absence of VAL repression. PlantPhysiol. 163, 1293–1305
13 Sundaram, S. et al. (2013) Gene networks and chromatin andtranscriptional regulation of the phaseolin promoter inArabidopsis. Plant Cell 25, 2601–2617
14 Mockaitis, K. and Estelle, M. (2008) Auxin receptors and plantdevelopment: a new signaling paradigm. Annu. Rev. Cell Dev. Biol.24, 55–80
15 Harper, R.M. et al. (2000) The NPH4 locus encodes the auxin responsefactor ARF7, a conditional regulator of differential growth in aerialArabidopsis tissue. Plant Cell 12, 757–770
16 Blilou, I. et al. (2005) The PIN auxin efflux facilitator network controlsgrowth and patterning in Arabidopsis roots. Nature 433, 39–44
17 Chen, M. et al. (2011) RopGEF7 regulates PLETHORA-dependentmaintenance of the root stem cell niche in Arabidopsis. Plant Cell 23,2880–2894
18 Hazak, O. and Yalovsky, S. (2010) An auxin regulated positivefeedback loop integrates Rho modulated cell polarity with patternformation. Plant Signal. Behav. 5, 709–711
19 Bureau, M. et al. (2010) JAGGED LATERAL ORGAN (JLO) controlsauxin dependent patterning during development of the Arabidopsisembryo and root. Plant Mol. Biol. 74, 479–491
20 Borghi, L. et al. (2007) Arabidopsis JAGGED LATERAL ORGANS isexpressed in boundaries and coordinates KNOX and PIN activity.Plant Cell 19, 1795–1808
21 Rast, M.I. and Simon, R. (2012) Arabidopsis JAGGED LATERALORGANS acts with ASYMMETRIC LEAVES2 to coordinate KNOXand PIN expression in shoot and root meristems. Plant Cell 24, 2917–2933
22 Smith, Z.R. and Long, J.A. (2010) Control of Arabidopsis apical-basalembryo polarity by antagonistic transcription factors. Nature 464,423–426
23 Long, J.A. et al. (2006) TOPLESS regulates apical embryonic fate inArabidopsis. Science 312, 1520–1523

Review Trends in Plant Science March 2014, Vol. 19, No. 3
24 Mallory, A.C. et al. (2004) MicroRNA control of PHABULOSA in leafdevelopment: importance of pairing to the microRNA 5’ region. EMBOJ. 23, 3356–3364
25 Kanei, M. et al. (2012) Stable establishment of cotyledon identityduring embryogenesis in Arabidopsis by ANGUSTIFOLIA3 andHANABA TARANU. Development 139, 2436–2446
26 Kim, J.H. and Kende, H. (2004) A transcriptional coactivator, AtGIF1,is involved in regulating leaf growth and morphology in Arabidopsis.Proc. Natl. Acad. Sci. U.S.A. 101, 13374–13379
27 Horiguchi, G. et al. (2005) The transcription factor AtGRF5 and thetranscription coactivator AN3 regulate cell proliferation in leafprimordia of Arabidopsis thaliana. Plant J. 43, 68–78
28 Rodriguez, R.E. et al. (2010) Control of cell proliferation inArabidopsis thaliana by microRNA miR396. Development 137,103–112
29 Wang, L. et al. (2011) miR396-targeted AtGRF transcription factorsare required for coordination of cell division and differentiation duringleaf development in Arabidopsis. J. Exp. Bot. 62, 761–773
30 Zhao, Y. et al. (2004) HANABA TARANU is a GATA transcriptionfactor that regulates shoot apical meristem and flower development inArabidopsis. Plant Cell 16, 2586–2600
31 Nawy, T. et al. (2010) The GATA factor HANABA TARANU isrequired to position the proembryo boundary in the earlyArabidopsis embryo. Dev. Cell 19, 103–113
32 Helariutta, Y. et al. (2000) The SHORT-ROOT gene controls radialpatterning of the Arabidopsis root through radial signaling. Cell 101,555–567
33 Nakajima, K. et al. (2001) Intercellular movement of the putativetranscription factor SHR in root patterning. Nature 413, 307–311
34 Sabatini, S. et al. (2003) SCARECROW is involved in positioning thestem cell niche in the Arabidopsis root meristem. Genes Dev. 17,354–358
35 Petersson, S.V. et al. (2009) An auxin gradient and maximum in theArabidopsis root apex shown by high-resolution cell-specific analysisof IAA distribution and synthesis. Plant Cell 21, 1659–1668
36 Sabatini, S. et al. (1999) An auxin-dependent distal organizer ofpattern and polarity in the Arabidopsis root. Cell 99, 463–472
37 Brunoud, G. et al. (2012) A novel sensor to map auxin response anddistribution at high spatio-temporal resolution. Nature 482, 103–106
38 Brady, S.M. et al. (2007) A high-resolution root spatiotemporal mapreveals dominant expression patterns. Science 318, 801–806
39 Ikeda, Y. et al. (2009) Local auxin biosynthesis modulates gradient-directed planar polarity in Arabidopsis. Nat. Cell Biol. 11, 731–738
40 Ljung, K. et al. (2005) Sites and regulation of auxin biosynthesis inArabidopsis roots. Plant Cell 17, 1090–1104
41 Stepanova, A.N. et al. (2005) A Link between ethylene and auxinuncovered by the characterization of two root-specific ethylene-insensitive mutants in Arabidopsis. Plant Cell 17, 2230–2242
42 Stepanova, A.N. et al. (2008) TAA1-mediated auxin biosynthesis isessential for hormone crosstalk and plant development. Cell 133,177–191
43 Friml, J. et al. (2002) AtPIN4 mediates sink-driven auxin gradientsand root patterning in Arabidopsis. Cell 108, 661–673
44 Friml, J. et al. (2002) Lateral relocation of auxin efflux regulator PIN3mediates tropism in Arabidopsis. Nature 415, 806–809
45 Galweiler, L. et al. (1998) Regulation of polar auxin transport byAtPIN1 in Arabidopsis vascular tissue. Science 282, 2226–2230
46 Grieneisen, V.A. et al. (2007) Auxin transport is sufficient togenerate a maximum and gradient guiding root growth. Nature449, 1008–1013
47 Wisniewska, J. et al. (2006) Polar PIN localization directs auxin flowin plants. Science 312, 883
48 Wildwater, M. et al. (2005) The RETINOBLASTOMA-RELATEDgene regulates stem cell maintenance in Arabidopsis roots. Cell123, 1337–1349
49 Galbiati, F. et al. (2013) An integrative model of the control of ovuleprimordia formation. Plant J. 76, 446–455
50 Hofhuis, H. et al. (2013) Phyllotaxis and rhizotaxis in Arabidopsis aremodified by three PLETHORA transcription factors. Curr. Biol. 23,956–962
51 Yamaguchi, N. et al. (2013) A molecular framework for auxin-mediated initiation of flower primordia. Dev. Cell 24, 271–282
52 Matsuzaki, Y. et al. (2010) Secreted peptide signals required formaintenance of root stem cell niche in Arabidopsis. Science 329,1065–1067
53 Zhou, W. et al. (2010) Arabidopsis tyrosylprotein sulfotransferase actsin the auxin/PLETHORA pathway in regulating postembryonicmaintenance of the root stem cell niche. Plant Cell 22, 3692–3709
54 Fernandez, A. et al. (2013) Transcriptional and functionalclassification of the GOLVEN/ROOT GROWTH FACTOR/CLE-likesignaling peptides reveals their role in lateral root and hair formation.Plant Physiol. 161, 954–970
55 Balaji, S. et al. (2005) Discovery of the principal specific transcriptionfactors of Apicomplexa and their implication for the evolution ofthe AP2-integrase DNA binding domains. Nucleic Acids Res. 33,3994–4006
56 Bewley, J.D. (1997) Seed germination and dormancy. Plant Cell 9,1055–1066
57 Nambara, E. et al. (2002) A screen for genes that function in abscisicacid signaling in Arabidopsis thaliana. Genetics 161, 1247–1255
58 Finkelstein, R.R. et al. (1998) The Arabidopsis abscisic acid responselocus ABI4 encodes an APETALA2 domain protein. Plant Cell 10,1043–1054
59 Yamagishi, K. et al. (2009) CHOTTO1, a double AP2 domain protein ofArabidopsis thaliana, regulates germination and seedling growthunder excess supply of glucose and nitrate. Plant Cell Physiol. 50,330–340
60 Shu, K. et al. (2013) ABI4 regulates primary seed dormancy byregulating the biogenesis of abscisic acid and gibberellins inArabidopsis. PLoS Genet. 9, e1003577
61 Yano, R. et al. (2009) CHOTTO1, a putative double APETALA2 repeattranscription factor, is involved in abscisic acid-mediated repressionof gibberellin biosynthesis during seed germination in Arabidopsis.Plant Physiol. 151, 641–654
62 Chen, Q. et al. (2011) The basic helix-loop-helix transcription factorMYC2 directly represses PLETHORA expression during jasmonate-mediated modulation of the root stem cell niche in Arabidopsis. PlantCell 23, 3335–3352
63 Sheard, L.B. et al. (2010) Jasmonate perception by inositol-phosphate-potentiated COI1-JAZ co-receptor. Nature 468, 400–405
64 Yan, J. et al. (2009) The Arabidopsis CORONATINE INSENSITIVE1protein is a jasmonate receptor. Plant Cell 21, 2220–2236
65 Boter, M. et al. (2004) Conserved MYC transcription factors play a keyrole in jasmonate signaling both in tomato and Arabidopsis. GenesDev. 18, 1577–1591
66 Lorenzo, O. et al. (2004) JASMONATE-INSENSITIVE1 encodes aMYC transcription factor essential to discriminate between differentjasmonate-regulated defense responses in Arabidopsis. Plant Cell 16,1938–1950
67 Aichinger, E. et al. (2011) The CHD3 chromatin remodeler PICKLEand Polycomb group proteins antagonistically regulate meristemactivity in the Arabidopsis root. Plant Cell 23, 1047–1060
68 Anzola, J.M. et al. (2010) Putative Arabidopsis transcriptionaladaptor protein (PROPORZ1) is required to modulate histoneacetylation in response to auxin. Proc. Natl. Acad. Sci. U.S.A. 107,10308–10313
69 Bertrand, C. et al. (2003) Arabidopsis histone acetyltransferaseAtGCN5 regulates the floral meristem activity through theWUSCHEL/AGAMOUS pathway. J. Biol. Chem. 278, 28246–28251
70 Kaya, H. et al. (2001) FASCIATA genes for chromatin assembly factor-1 in Arabidopsis maintain the cellular organization of apicalmeristems. Cell 104, 131–142
71 Kornet, N. and Scheres, B. (2009) Members of the GCN5 histoneacetyltransferase complex regulate PLETHORA-mediated root stemcell niche maintenance and transit amplifying cell proliferation inArabidopsis. Plant Cell 21, 1070–1079
72 Kwon, C.S. et al. (2005) WUSCHEL is a primary target fortranscriptional regulation by SPLAYED in dynamic control of stemcell fate in Arabidopsis. Genes Dev. 19, 992–1003
73 Meshorer, E. and Misteli, T. (2006) Chromatin in pluripotentembryonic stem cells and differentiation. Nat. Rev. Mol. Cell Biol.7, 540–546
74 Serrano, L. et al. (2013) Chromatin structure, pluripotency anddifferentiation. Exp. Biol. Med. (Maywood) 238, 259–270
155

Review Trends in Plant Science March 2014, Vol. 19, No. 3
75 Musselman, C.A. and Kutateladze, T.G. (2011) Handpickingepigenetic marks with PHD fingers. Nucleic Acids Res. 39, 9061–9071
76 Saiga, S. et al. (2008) The Arabidopsis OBERON1 and OBERON2genes encode plant homeodomain finger proteins and are required forapical meristem maintenance. Development 135, 1751–1759
77 Thomas, C.L. et al. (2009) Arabidopsis plant homeodomain fingerproteins operate downstream of auxin accumulation in specifying thevasculature and primary root meristem. Plant J. 59, 426–436
78 Mudunkothge, J.S. and Krizek, B.A. (2012) Three Arabidopsis AIL/PLT genes act in combination to regulate shoot apical meristemfunction. Plant J. 71, 108–121
79 Prasad, K. et al. (2011) Arabidopsis PLETHORA transcription factorscontrol phyllotaxis. Curr. Biol. 21, 1123–1128
80 Long, J. and Barton, M.K. (2000) Initiation of axillary and floralmeristems in Arabidopsis. Dev. Biol. 218, 341–353
81 Long, J.A. and Barton, M.K. (1998) The development of apicalembryonic pattern in Arabidopsis. Development 125, 3027–3035
82 Bowman, J.L. et al. (1991) Expression of the Arabidopsis floralhomeotic gene AGAMOUS is restricted to specific cell types late inflower development. Plant Cell 3, 749–758
83 Krizek, B.A. (1999) Ectopic expression of AINTEGUMENTA inArabidopsis plants results in increased growth of floral organs.Dev. Genet. 25, 224–236
84 Krizek, B.A. et al. (2000) AINTEGUMENTA promotes petalidentity and acts as a negative regulator of AGAMOUS. Plant Cell12, 1357–1366
85 Pinon, V. et al. (2013) Local auxin biosynthesis regulation byPLETHORA transcription factors controls phyllotaxis inArabidopsis. Proc. Natl. Acad. Sci. U.S.A. 110, 1107–1112
86 Dubrovsky, J.G. et al. (2006) Lateral root initiation in Arabidopsis:developmental window, spatial patterning, density andpredictability. Ann. Bot. 97, 903–915
87 Laskowski, M. et al. (2008) Root system architecture from couplingcell shape to auxin transport. PLoS Biol. 6, e307
88 Fukaki, H. et al. (2002) Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene ofArabidopsis. Plant J. 29, 153–168
89 Okushima, Y. et al. (2005) Functional genomic analysis of the AUXINRESPONSE FACTOR gene family members in Arabidopsis thaliana:unique and overlapping functions of ARF7 and ARF19. Plant Cell 17,444–463
90 Vanneste, S. et al. (2005) Cell cycle progression in the pericycle is notsufficient for SOLITARY ROOT/IAA14-mediated lateral rootinitiation in Arabidopsis thaliana. Plant Cell 17, 3035–3050
91 Wilmoth, J.C. et al. (2005) NPH4/ARF7 and ARF19 promote leafexpansion and auxin-induced lateral root formation. Plant J. 43,118–130
92 Mizukami, Y. and Fischer, R.L. (2000) Plant organ size control:AINTEGUMENTA regulates growth and cell numbers duringorganogenesis. Proc. Natl. Acad. Sci. U.S.A. 97, 942–947
93 Krizek, B. (2009) AINTEGUMENTA and AINTEGUMENTA-LIKE6act redundantly to regulate Arabidopsis floral growth and patterning.Plant Physiol. 150, 1916–1929
94 Hu, Y. et al. (2003) The Arabidopsis auxin-inducible gene ARGOScontrols lateral organ size. Plant Cell 15, 1951–1961
95 Krizek, B.A. and Eaddy, M. (2012) AINTEGUMENTA-LIKE6regulates cellular differentiation in flowers. Plant Mol. Biol. 78,199–209
96 Siegfried, K.R. et al. (1999) Members of the YABBY gene familyspecify abaxial cell fate in Arabidopsis. Development 126, 4117–4128
97 Nole-Wilson, S. and Krizek, B.A. (2006) AINTEGUMENTAcontributes to organ polarity and regulates growth of lateralorgans in combination with YABBY genes. Plant Physiol. 141,977–987
98 Schultz, E.A. and Haughn, G.W. (1991) LEAFY, a homeotic gene thatregulates inflorescence development in Arabidopsis. Plant Cell 3,771–781
99 Weigel, D. et al. (1992) LEAFY controls floral meristem identity inArabidopsis. Cell 69, 843–859
100 Weigel, D. and Meyerowitz, E.M. (1993) Activation of floral homeoticgenes in Arabidopsis. Science 261, 1723–1726
101 Weigel, D. and Nilsson, O. (1995) A developmental switch sufficientfor flower initiation in diverse plants. Nature 377, 495–500
156
102 Przemeck, G.K. et al. (1996) Studies on the role of the Arabidopsisgene MONOPTEROS in vascular development and plant cellaxialization. Planta 200, 229–237
103 Baker, S.C. et al. (1997) Interactions among genes regulating ovuledevelopment in Arabidopsis thaliana. Genetics 145, 1109–1124
104 Schneitz, K. et al. (1997) Dissection of sexual organ ontogenesis: agenetic analysis of ovule development in Arabidopsis thaliana.Development 124, 1367–1376
105 Schneitz, K. et al. (1998) Pattern formation and growth during floralorganogenesis: HUELLENLOS and AINTEGUMENTA are requiredfor the formation of the proximal region of the ovule primordium inArabidopsis thaliana. Development 125, 2555–2563
106 Azhakanandam, S. et al. (2008) SEUSS and AINTEGUMENTAmediate patterning and ovule initiation during gynoecium medialdomain development. Plant Physiol. 146, 1165–1181
107 Liu, Z.C. et al. (2000) Regulation of gynoecium marginal tissueformation by LEUNIG and AINTEGUMENTA. Plant Cell 12,1879–1891
108 Nole-Wilson, S. et al. (2010) Polar auxin transport together withAINTEGUMENTA and REVOLUTA coordinate early Arabidopsisgynoecium development. Dev. Biol. 346, 181–195
109 Krizek, B.A. (2003) AINTEGUMENTA utilizes a mode of DNArecognition distinct from that used by proteins containing a singleAP2 domain. Nucleic Acids Res. 31, 1859–1868
110 Nole-Wilson, S. and Krizek, B.A. (2000) DNA binding properties of theArabidopsis floral development protein AINTEGUMENTA. NucleicAcids Res. 28, 4076–4082
111 Fujimoto, S.Y. et al. (2000) Arabidopsis ethylene-responsive elementbinding factors act as transcriptional activators or repressors of GCCbox-mediated gene expression. Plant Cell 12, 393–404
112 Hao, D. et al. (1998) Unique mode of GCC box recognition by the DNA-binding domain of ethylene-responsive element-binding factor (ERFdomain) in plant. J. Biol. Chem. 273, 26857–26861
113 Shoji, T. et al. (2013) Divergent DNA-binding specificities of a group ofETHYLENE RESPONSE FACTOR transcription factors involved inplant defense. Plant Physiol. 162, 977–990
114 Passarinho, P. et al. (2008) BABY BOOM target genes provide diverseentry points into cell proliferation and cell growth pathways. PlantMol. Biol. 68, 225–237
115 Causier, B. et al. (2012) The TOPLESS interactome: a framework forgene repression in Arabidopsis. Plant Physiol. 158, 423–438
116 Krogan, N.T. et al. (2012) APETALA2 negatively regulates multiplefloral organ identity genes in Arabidopsis by recruiting the co-repressor TOPLESS and the histone deacetylase HDA19.Development 139, 4180–4190
117 Carlier, M.F. et al. (1997) Actin depolymerizing factor (ADF/cofilin)enhances the rate of filament turnover: implication in actin-basedmotility. J. Cell Biol. 136, 1307–1322
118 Gungabissoon, R.A. et al. (1998) Interaction of maize actin-depolymerising factor with actin and phosphoinositides and itsinhibition of plant phospholipase C. Plant J. 16, 689–696
119 Burgos-Rivera, B. et al. (2008) ACTIN DEPOLYMERIZINGFACTOR9 controls development and gene expression inArabidopsis. Plant Mol. Biol. 68, 619–632
120 Mizumoto, K. et al. (2009) Altered expression of wheatAINTEGUMENTA homolog, WANT-1, in pistil and pistil-liketransformed stamen of an alloplasmic line with Aegilops crassacytoplasm. Dev. Genes Evol. 219, 175–187
121 Shigyo, M. et al. (2006) Molecular evolution of the AP2 subfamily.Gene 366, 256–265
122 Yamada, T. et al. (2008) AINTEGUMENTA homolog expression inGnetum (gymnosperms) and implications for the evolution of ovulateaxes in seed plants. Evol. Dev. 10, 280–287
123 Rieu, I. et al. (2005) Isolation and expression analysis of a tobaccoAINTEGUMENTA ortholog (NtANTL). Plant Cell Physiol. 46,803–805
124 Deng, W. et al. (2009) A novel method for induction of plantregeneration via somatic embryogenesis. Plant Sci. 177, 43–48
125 Heidmann, I. et al. (2011) Efficient sweet pepper transformationmediated by the BABY BOOM transcription factor. Plant Cell Rep.30, 1107–1115
126 Riechmann, J.L. and Meyerowitz, E.M. (1998) The AP2/EREBPfamily of plant transcription factors. Biol. Chem. 379, 633–646

Review Trends in Plant Science March 2014, Vol. 19, No. 3
127 Kim, S. et al. (2006) Phylogeny and domain evolution in theAPETALA2-like gene family. Mol. Biol. Evol. 23, 107–120
128 Zhuang, J. et al. (2009) Genome-wide analysis of the putative AP2/ERF family genes in Vitis vinifera. Sci. Hort. 123, 73–81
129 Dietz, K.J. et al. (2010) AP2/EREBP transcription factors are part ofgene regulatory networks and integrate metabolic, hormonal andenvironmental signals in stress acclimation and retrogradesignalling. Protoplasma 245, 3–14
130 Licausi, F. et al. (2010) Genomic and transcriptomic analysis of theAP2/ERF superfamily in Vitis vinifera. BMC Genomics 11, 719
131 Sharoni, A.M. et al. (2011) Gene structures, classification andexpression models of the AP2/EREBP transcription factor family inrice. Plant Cell Physiol. 52, 344–360
132 Rashid, M. et al. (2012) AP2/ERF transcription factor in rice: genome-wide canvas and syntenic relationships between monocots andeudicots. Evol. Bioinform. Online 8, 321–355
133 Duan, C. et al. (2013) Identification of the Hevea brasiliensis AP2/ERFsuperfamily by RNA sequencing. BMC Genomics 14, 30
134 Okamuro, J.K. et al. (1997) The AP2 domain of APETALA2 defines alarge new family of DNA binding proteins in Arabidopsis. Proc. Natl.Acad. Sci. U.S.A. 94, 7076–7081
135 Jofuku, K.D. et al. (1994) Control of Arabidopsis flower and seeddevelopment by the homeotic gene APETALA2. Plant Cell 6, 1211–1225
136 Ohme-Takagi, M. and Shinshi, H. (1995) Ethylene-inducible DNAbinding proteins that interact with an ethylene-responsive element.Plant Cell 7, 173–182
137 Magnani, E. et al. (2004) From endonucleases to transcription factors:evolution of the AP2 DNA binding domain in plants. Plant Cell 16,2265–2277
138 Wuitschick, J.D. et al. (2004) Homing endonucleases encoded by germline-limited genes in Tetrahymena thermophila have APETELA2DNA binding domains. Eukaryot. Cell 3, 685–694
139 Taylor, G.K. and Stoddard, B.L. (2012) Structural, functional andevolutionary relationships between homing endonucleases andproteins from their host organisms. Nucleic Acids Res. 40, 5189–5200
140 El Ouakfaoui, S. et al. (2010) Control of somatic embryogenesis andembryo development by AP2 transcription factors. Plant Mol. Biol. 74,313–326
141 Bandupriya, H.D.D. et al. (2013) Isolation and characterization of anAINTEGUMENTA-like gene in different coconut (Cocos nucifera L.)varieties from Sri Lanka. Tree Genet. Genomes 9, 813–827
142 Allen, M.D. et al. (1998) A novel mode of DNA recognition by a beta-sheet revealed by the solution structure of the GCC-box bindingdomain in complex with DNA. EMBO J. 17, 5484–5496
143 Cao, Z.F. et al. (2001) Effect of two conserved amino acid residues onDREB1A function. Biochemistry (Mosc.) 66, 623–627
144 Tamura, K. et al. (2011) MEGA5: molecular evolutionary geneticsanalysis using maximum likelihood, evolutionary distance, andmaximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739
145 Ju rgens, G. and Mayer, U. (1994) EMBRYOS: Colour Atlas ofDevelopment, Wolfe Publishing
146 Scheres, B. et al. (1994) Embryonic origin of the Arabidopsisprimary root and root-meristem initials. Development 120,2475–2487
147 van den Berg, C. et al. (1997) Short-range control of cell differentiationin the Arabidopsis root meristem. Nature 390, 287–289
148 Parizot, B. et al. (2008) Diarch symmetry of the vascular bundle inArabidopsis root encompasses the pericycle and is reflected in distichlateral root initiation. Plant Physiol. 146, 140–148
149 Benkova, E. et al. (2003) Local, efflux-dependent auxin gradients as acommon module for plant organ formation. Cell 115, 591–602
150 De Smet, I. et al. (2007) Auxin-dependent regulation of lateral rootpositioning in the basal meristem of Arabidopsis. Development 134,681–690
151 Moreno-Risueno, M.A. et al. (2010) Oscillating gene expressiondetermines competence for periodic Arabidopsis root branching.Science 329, 1306–1311
152 Szakonyi, D. et al. (2010) Perspectives on leaf dorsoventral polarity. J.Plant Res. 123, 281–290
153 Kuhlemeier, C. (2007) Phyllotaxis. Trends Plant Sci. 12, 143–150
157
Related Documents











