AhpC (alkyl hydroperoxide reductase) from Anabaena sp. PCC 7120 protects Escherichia coli from multiple abiotic stresses Yogesh Mishra, Neha Chaurasia, Lal Chand Rai * Molecular Biology Section, Laboratory of Algal Biology, Center of Advanced Study in Botany, Banaras Hindu University, Varanasi-221005, India article info Article history: Received 9 February 2009 Available online 25 February 2009 Keywords: Anabaena sp. PCC 7120 Alkyl hydroperoxide reductase (AhpC) Abiotic stress Cloning RT-PCR abstract Alkyl hydroperoxide reductase (AhpC) is known to detoxify peroxides and reactive sulfur species (RSS). However, the relationship between its expression and combating of abiotic stresses is still not clear. To investigate this relationship, the genes encoding the alkyl hydroperoxide reductase (ahpC) from Anabaena sp. PCC 7120 were introduced into E. coli using pGEX-5X-2 vector and their possible func- tions against heat, salt, carbofuron, cadmium, copper and UV-B were analyzed. The transformed E. coli cells registered significantly increase in growth than the control cells under temperature (47 °C), NaCl (6% w/v), carbofuron (0.025 mg ml 1 ), CdCl 2 (4 mM), CuCl 2 (1 mM), and UV-B (10 min) exposure. Enhanced expression of ahpC gene as measured by semi-quantitative RT-PCR under afore- mentioned stresses at different time points demonstrated its role in offering tolerance against multiple abiotic stresses. Ó 2009 Elsevier Inc. All rights reserved. Introduction Alkyl hydroperoxide reductase (AhpC) is a key component of a large family of thiol-specific antioxidant (TSA) proteins distributed among prokaryotes and eukaryotes. Kim and Rhee [1] reported this protein from Saccharomyces cerevisiae and named as ‘‘Protector Protein”. Soon after, Jacobson et al. [2] reported the presence of AhpC in the bacterium Salmonella typhimurium. AhpC is known to scavenge a variety of peroxides, reactive oxygen, nitrogen and sulfur species [3,4]. A critical perusal of literature suggests that information on AhpC have remained confined to yeast and bacteria as for the molecular mechanism of its function against abiotic stresses is concerned. Bioinformatics analysis of the AhpC/TSA family revealed the presence of two highly conserved cysteine residues corresponding to Cys-47 and Cys-170 in yeast TSA. The N-terminal cysteine is conserved in all family members while the C-terminal cysteine is conserved in all except six members. Both Cys-47 and Cys-170 have been reported to be indispensable for the maintenance of the dimeric structure of the oxidized TSA and Cys-47. However, Cys-170 is not essential for the antioxidant activ- ity as measured in vitro [3]. During the last few years the AhpC has been studied from a variety of aerobic and anaerobic bacteria including Clostridium pasteurianum [5] and Bacillus subtilis [6] with a major focus to understand the role of AhpC in (i) scavenging ROS and peroxides, (ii) virulence, (iii) interaction with other pathways, (iv) coloniza- tion, and (v) switching from a peroxide reductase to a molecular chaperone function. However, information on cyanobacterial AhpC has remained confined to Synechocystis [7]. Keeping in mind the reports that AhpC affects gene expression, protects DNA from oxidative damage, behaves as molecular chap- erone under peroxide stress and binds metals, it was hypothesized that enhanced expression of this protein in Escherichia coli might offer protection from abiotic stresses. This study has been designed to clone ahpC gene from the filamentous diazotrophic cyanobacte- rium Anabaena sp. PCC 7120 into E. coli and examine its tolerance to abiotic stresses like heat, salt, pesticide, heavy metals and UV-B radiation. Materials and methods Cyanobacterial and bacterial strains and plasmids. Anabaena sp. PCC 7120 was grown photoautotrophically in BG-11 medium [8] buffered with Tris/HCl at 24 ± 2 °C under day light fluorescent tubes emitting 72 lmol photon m 2 s 1 PAR (photosynthetically active radiation) light intensity with a photoperiod of 14:10 h at pH 7.5. The cultures were shaken by hand 2–3 times daily. Esche- richia coli strain DH5a and E. coli BL21 (Novagen) were used as hosts for cloning and expression, respectively. Escherichia coli cul- tures were stored as 10% (v/v) glycerol stocks at 80 °C and main- tained on Luria–Bertani (LB) plates containing 1.4% (w/v) agar at 37 °C. Cells harboring recombinant plasmids were grown and maintained on LB media supplemented with 100 lg/ml ampicillin [9]. Plasmid pGEX-5X-2 (GE Healthcare) was used as a vector for cloning. 0006-291X/$ - see front matter Ó 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2009.02.100 * Corresponding author. Fax: +91 542 2368174. E-mail addresses: [email protected], [email protected] (L.C. Rai). Biochemical and Biophysical Research Communications 381 (2009) 606–611 Contents lists available at ScienceDirect Biochemical and Biophysical Research Communications journal homepage: www.elsevier.com/locate/ybbrc

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biochemical and Biophysical Research Communications 381 (2009) 606–611
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
AhpC (alkyl hydroperoxide reductase) from Anabaena sp. PCC 7120 protectsEscherichia coli from multiple abiotic stresses
Yogesh Mishra, Neha Chaurasia, Lal Chand Rai *
Molecular Biology Section, Laboratory of Algal Biology, Center of Advanced Study in Botany, Banaras Hindu University, Varanasi-221005, India
a r t i c l e i n f o
Article history:Received 9 February 2009Available online 25 February 2009
Keywords:Anabaena sp. PCC 7120Alkyl hydroperoxide reductase (AhpC)Abiotic stressCloningRT-PCR
0006-291X/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.bbrc.2009.02.100
* Corresponding author. Fax: +91 542 2368174.E-mail addresses: [email protected], [email protected]
a b s t r a c t
Alkyl hydroperoxide reductase (AhpC) is known to detoxify peroxides and reactive sulfur species (RSS).However, the relationship between its expression and combating of abiotic stresses is still not clear. Toinvestigate this relationship, the genes encoding the alkyl hydroperoxide reductase (ahpC) fromAnabaena sp. PCC 7120 were introduced into E. coli using pGEX-5X-2 vector and their possible func-tions against heat, salt, carbofuron, cadmium, copper and UV-B were analyzed. The transformedE. coli cells registered significantly increase in growth than the control cells under temperature(47 �C), NaCl (6% w/v), carbofuron (0.025 mg ml�1), CdCl2 (4 mM), CuCl2 (1 mM), and UV-B (10 min)exposure. Enhanced expression of ahpC gene as measured by semi-quantitative RT-PCR under afore-mentioned stresses at different time points demonstrated its role in offering tolerance against multipleabiotic stresses.
� 2009 Elsevier Inc. All rights reserved.
Introduction
Alkyl hydroperoxide reductase (AhpC) is a key component of alarge family of thiol-specific antioxidant (TSA) proteins distributedamong prokaryotes and eukaryotes. Kim and Rhee [1] reported thisprotein from Saccharomyces cerevisiae and named as ‘‘ProtectorProtein”. Soon after, Jacobson et al. [2] reported the presence ofAhpC in the bacterium Salmonella typhimurium. AhpC is known toscavenge a variety of peroxides, reactive oxygen, nitrogen andsulfur species [3,4]. A critical perusal of literature suggests thatinformation on AhpC have remained confined to yeast and bacteriaas for the molecular mechanism of its function against abioticstresses is concerned. Bioinformatics analysis of the AhpC/TSAfamily revealed the presence of two highly conserved cysteineresidues corresponding to Cys-47 and Cys-170 in yeast TSA. TheN-terminal cysteine is conserved in all family members while theC-terminal cysteine is conserved in all except six members. BothCys-47 and Cys-170 have been reported to be indispensable forthe maintenance of the dimeric structure of the oxidized TSA andCys-47. However, Cys-170 is not essential for the antioxidant activ-ity as measured in vitro [3].
During the last few years the AhpC has been studied from avariety of aerobic and anaerobic bacteria including Clostridiumpasteurianum [5] and Bacillus subtilis [6] with a major focus tounderstand the role of AhpC in (i) scavenging ROS and peroxides,(ii) virulence, (iii) interaction with other pathways, (iv) coloniza-
ll rights reserved.
c.in (L.C. Rai).
tion, and (v) switching from a peroxide reductase to a molecularchaperone function. However, information on cyanobacterial AhpChas remained confined to Synechocystis [7].
Keeping in mind the reports that AhpC affects gene expression,protects DNA from oxidative damage, behaves as molecular chap-erone under peroxide stress and binds metals, it was hypothesizedthat enhanced expression of this protein in Escherichia coli mightoffer protection from abiotic stresses. This study has been designedto clone ahpC gene from the filamentous diazotrophic cyanobacte-rium Anabaena sp. PCC 7120 into E. coli and examine its toleranceto abiotic stresses like heat, salt, pesticide, heavy metals and UV-Bradiation.
Materials and methods
Cyanobacterial and bacterial strains and plasmids. Anabaena sp.PCC 7120 was grown photoautotrophically in BG-11 medium [8]buffered with Tris/HCl at 24 ± 2 �C under day light fluorescenttubes emitting 72 lmol photon m�2 s�1 PAR (photosyntheticallyactive radiation) light intensity with a photoperiod of 14:10 h atpH 7.5. The cultures were shaken by hand 2–3 times daily. Esche-richia coli strain DH5a and E. coli BL21 (Novagen) were used ashosts for cloning and expression, respectively. Escherichia coli cul-tures were stored as 10% (v/v) glycerol stocks at �80 �C and main-tained on Luria–Bertani (LB) plates containing 1.4% (w/v) agar at37 �C. Cells harboring recombinant plasmids were grown andmaintained on LB media supplemented with 100 lg/ml ampicillin[9]. Plasmid pGEX-5X-2 (GE Healthcare) was used as a vector forcloning.

Y. Mishra et al. / Biochemical and Biophysical Research Communications 381 (2009) 606–611 607
Cloning of the ahpC gene from Anabaena sp. PCC 7120. GenomicDNA from Anabaena sp. PCC 7120 was isolated as previously de-scribed [10]. An open reading frame alr4404, putatively encodingalkyl hydroperoxide reductase was amplified by polymerase chainreaction using genomic DNA as template with a pair of primers, Pf(50CGGAATTCCCATGGCTCTCCGTCTTGGT30) and Pr (50ATTTGCGGCCGCTTACTTGTTAGGTTGAGGAGT30). The underlined bases repre-sent EcoRI and NotI recognition sites, respectively. The PCR wasdone in a reaction mixture of 25 ll for 30 cycles at 94 �C for1.5 min, 62 �C for 1 min, and 72 �C for 2 min using standard PCRconditions (100 ng DNA, 2.5 ll of 10� PCR buffer with 15 mMMgCl2, 200 lM dNTPs, 10 pmol of each primer and 0.2 U TaqDNA polymerase in an Icycler (Bio-Rad, USA). The amplified prod-uct was purified using standard freeze–thaw method for cloning.
Construction of expression vector. The purified PCR product wasdigested with EcoRI and NotI (NEB) and the resultant DNAfragment was cloned into the expression vector pGEX-5X-2, di-gested with the same restriction enzymes. To construct the recom-binant, plasmid pGEX-5X-2-ahpC was introduced into E. coli BL21,the latter was then grown in LB medium. The plasmid was thenisolated and the DNA sequence of alr4404 was confirmed bysequencing.
Expression analysis of ahpC gene using RT-PCR. For expressionanalysis of ahpC, E. coli. BL21 harboring the pGEX-5X-2-ahpC plas-mid was grown in LB medium supplemented with 100 lg ml�1
ampicillin in an orbital shaker (200 rpm) at 37 �C. When A600 ofthe culture reached a value of 0.5, isopropyl b-D-1-thiogalactopyra-noside (IPTG) was added at a final concentration of 0.5 mM and theculture was grown for another 6 h at 37 �C. RNA was isolated frommid-exponential phase cells harboring the pGEX-5X-2-ahpC beforeand after IPTG induction using the TRIzol reagent as per theinstructions given in the manufacturer’s protocol. All RNA sampleswere diluted to a concentration of 3 lg ml�1 prior to RT. In a clearnuclease free 0.2 mL microcentrifuge tube 1 lL of 10 mM dNTPmix, 1 lL RNA template and 6 lL of nuclease free water was mixedgently and incubated at 70 �C for 10 min to remove any secondarystructure of RNA. This was now placed on ice. About 15 U of M-MLV reverse transcriptase (SIGMA, USA) was added along withthe RT buffer. The final volume was adjusted to 20 ll with nucleasefree water. The reaction mixture was incubated at 25 �C for 5 minfollowed by 37 �C for 50 min. To test the purity of the cDNA controlreaction mixtures were prepared in the same way as mentionedabove except that M-MLV RT was not added. To ascertain the equalconcentration of RNA in the different samples RT-PCR of 16S rDNAwas also performed and used as an internal control. The primerpair 8F 50AGAGTTTGATCCTGGCTCAG30 and 518R 50ATTA-CCGCGGCTGCTGG30 was used to amplify 16S rDNA by using thePCR cycle as follows: initial denaturation at 95 �C for 5 min, fol-lowed by 30 cycles of incubation each consisting of 1 min denatur-ation at 94 �C, 1.5 min annealing at 58 �C, 2 min extension at 72 �Cand a final extension of 10 min at 72 �C. To allow relative quantifi-cation of the ahpC gene, preliminary experiments were carried outwith stepwise reduction of the number of PCR cycles to determinethe maximum cycle number where samples do not reach amplifi-cation plateau. For amplification of the ahpC genes, 1 lL of the RTreaction product was used in subsequent PCR. PCR was performedin 25 ll final volume of reaction mixture containing 100 ng of DNA,2.5 ll of 10X PCR buffer with 15 mM MgCl2, 200 lM dNTPs,10 pmol of each primer and 0.2 U Taq DNA polymerase (BangaloreGenei, India) in an Icycler (Bio-Rad, USA). The Icycler profile was asfollows: initial denaturation for 5 min at 94 �C followed by 40 incu-bation cycles each consisting of 1.5 min denaturation at 94 �C,1 min annealing at 62 �C, 2 min extension at 72 �C and a final10 min extension at 72 �C. The intensities of the RT-PCR productson agarose gels were quantified with the Gel Doc 2000 systemusing the Quantity one software (BioRad, USA).
Enzyme activity assay. For enzymatic assay of alkyl hydroperox-ide reductase, E. coli BL21 harboring the pGEX-5X-2-ahpC andpGEX-5X-2 plasmids was grown as described previously in expres-sion analysis of ahpC gene using RT-PCR. Cells harvested by centri-fugation were suspended in 20 mM sodium phosphate buffer (pH7.4) and disrupted by ultrasonication. The crude extract was cen-trifuged at 15,000 rpm for 1 h and the clear supernatant used forenzymatic assay. The rate of DTT oxidation catalyzed by AhpC inthe presence of the peroxide substrate was measured by monitor-ing the change in absorbance at 310 nm due to the formation of theDTT disulfide. Typical conditions for the assays include enzyme(5.6 lM) incubated for 10 min with 10 mM DTT and 2 mM t-butylhydroperoxide in 50 mM potassium phosphate buffer, pH 7.5, con-taining 300 mM NaCl and 1 mM EDTA in a 1-ml quartz cuvette. Therate of DTT oxidation was measured spectrophotometrically at310 nm at room temperature [11].
SDS–PAGE analysis. For preparation of cell-free extracts (forSDS–PAGE) bacteria were harvested by centrifugation at5000 rpm for 10 min in SIGMA 3 K-30 laboratory centrifuge, Ger-many and washed twice with extraction buffer (50 mM Tris–HCl,10 mM MgCl2, 20 mM KCl, pH 7.5). The pellet was re-suspendedin 5 ml extraction buffer and subjected to grinding under liquidnitrogen to break the cells. The extract was centrifuged at10,000 rpm for 60 min. The supernatant was subjected to ace-tone/TCA precipitation overnight at �20 �C followed by centrifuga-tion at 10,000 rpm for 15 min. The pellet was re-suspended in500 ll Tris buffer (10 mM Tris–HCl, 50 mM NaCl, 1 mM EDTA, pH7.5). The sample was stored in small aliquots at �20 �C until fur-ther characterization.
Western blot analysis. In order to confirm the expression of ahpCgene in E. coli cells, SDS–PAGE was carried out as per the method ofSambrook and Russell [9]. The gels were transferred to a PVDFmembrane (Millipore Immobilon-P), using a dual mini-electroblotsystem (Precision Instruments, Varanasi, India). The gel cassettewas kept in transfer buffer (3.03 g l�1 Tris base, 14.4 g l�1 glycine,and 200 ml methanol (99% v/v pure) for 12 h or overnight at 15 Vat 4 �C. Membrane was blocked for 4 h in TTBS (Tris buffer salinecontaining 0.1% Tween 20) and 5% (w/v) non-fat dried milk. Theprimary antibody was diluted as per the instructions of the donors.The membrane incubated overnight was washed five times for5 min each in TTBS. This was then incubated in a Goat anti RabbitIgG HRP (horseradish peroxidase) conjugated secondary antibody(Genei, India) for 4 h. Following four consecutive 5 min wash inTTBS the membrane was developed with DAB/NiCl2 visualizationsolution. The reaction was terminated by washing the PVDF mem-brane with Milli Q water. The immunoblots were dried betweenfilter paper to reduce the background staining and photographedusing a gel documentation system (Bio-Rad, USA). Polyclonal anti-bodies used for the detection of AhpC were obtained as generousgift from Prof. Dr. Karl-Josef Dietz, Germany.
Assay for abiotic stress tolerance in transformed E. coli cells. In or-der to assess the role of alkyl hydroperoxide reductase in multiplestress tolerance, effects of copper, UV-B, salt, carbofuron, cadmiumand temperature on the growth of transformed E. coli BL21 cellswith pGEX-5X-2 (empty vector) and pGEX-5X-2-ahpC (recombi-nant plasmid) were examined.
Mode of stress application: NaCl autoclaved separately wasadded directly into the sterilized medium to achieve an appropri-ate concentration. Stock solution of CdCl2�2H2O (100 mM),CuCl2�2H2O (1 M) and carbofuron (120 mg ml�1) was prepared inglass-distilled water and sterilized by passing through a Milliporemembrane filter (0.22 lm). Temperature treatment to culture sus-pensions was given in a temperature controlled incubator. About2 mL of bacterial culture suspension (OD 0.5) was transferred intoquartz cuvette (Pye Unicam B538751 A, thickness 1 mm, capacity4 ml) exposed to artificial UV-B radiation (from UV-B lamp, CAT
0.6 pGEX-5X-2-ahpC (-IPTG)
608 Y. Mishra et al. / Biochemical and Biophysical Research Communications 381 (2009) 606–611
No. 34408, fotodyne, Inc. USA giving its maximum output at310 nm).
Measurement of survival and growth. Survival of the E. coli cells(transformed with empty vector) against salt, carbofuron, cad-mium and temperature was measured by treating them with dif-ferent concentrations of NaCl (2%, 4%, 6%, 8% and 10%),carbofuron (0.01, 0.02, 0.03, 0.04 and 0.05 mg ml�1), CdCl2 (2, 4,6 and 8 mM), temperature (42, 47, 52 and 57 �C), CuCl2 (0.5, 1, 2,4 and 6 mM) and UV-B (5, 10, 20, 30, 40, 50 and 60 min), respec-tively. The LC50 for NaCl, carbofuron, cadmium (CdCl2), tempera-ture, copper (CuCl2) and UV-B were determined by the platecolony count method. Approximately 50% survival of E. coli was ob-served at NaCl (6%), carbofuron (0.025 mg ml�1), CdCl2 (4 mM),temperature (47 �C), CuCl2 (1 mM) and UV-B (10 min.), respec-tively. The E. coli cells transformed with empty vector failed toshow any perceptible change in their growth when subjected todoses lower than LC50 of the selected stresses (data not shown).All growth experiments were conducted on a rotary shaker(200 rpm) at 37 �C.
Single colony of E. coli BL21(DE3) cells transformed either withempty vector or recombinant plasmid was inoculated in tubes con-taining fresh LB medium and 100 lg ml�1 ampicillin and grownovernight. On the following day the cultures were diluted to 0.05optical density (OD) with fresh LB medium (25 ml in 100 ml coni-cal flask) and incubated until OD value reached approximately 0.5.For abiotic stress treatment, 50 ll of E. coli culture (OD 0.5) wasinoculated into flasks containing 50 ml culture medium and LC50
doses of NaCl, carbofuron, CdCl2 and CuCl2. For UV-B treatment2 ml of E. coli culture (OD 0.5) was exposed to 10 min. of UV-Band 50 ll of such UV-B treated cells were inoculated into tubescontaining 50 ml of liquid medium and grown in dark at 37 �C.For heat treatment, 50 ml of liquid medium inoculated by 50 llof E. coli cells (OD 0.5) was incubated at 47 �C. The bacterial sus-pension was harvested at every 30 min and optical density mea-sured by spectrophotometer (GE healthcare, USA). Specificgrowth rate was calculated by using the equation: l = [ln (n2/n1)]/[t2-t1] where l stands for specific growth rate and n1, n2 areabsorbance of culture suspension at the beginning (t1) and end
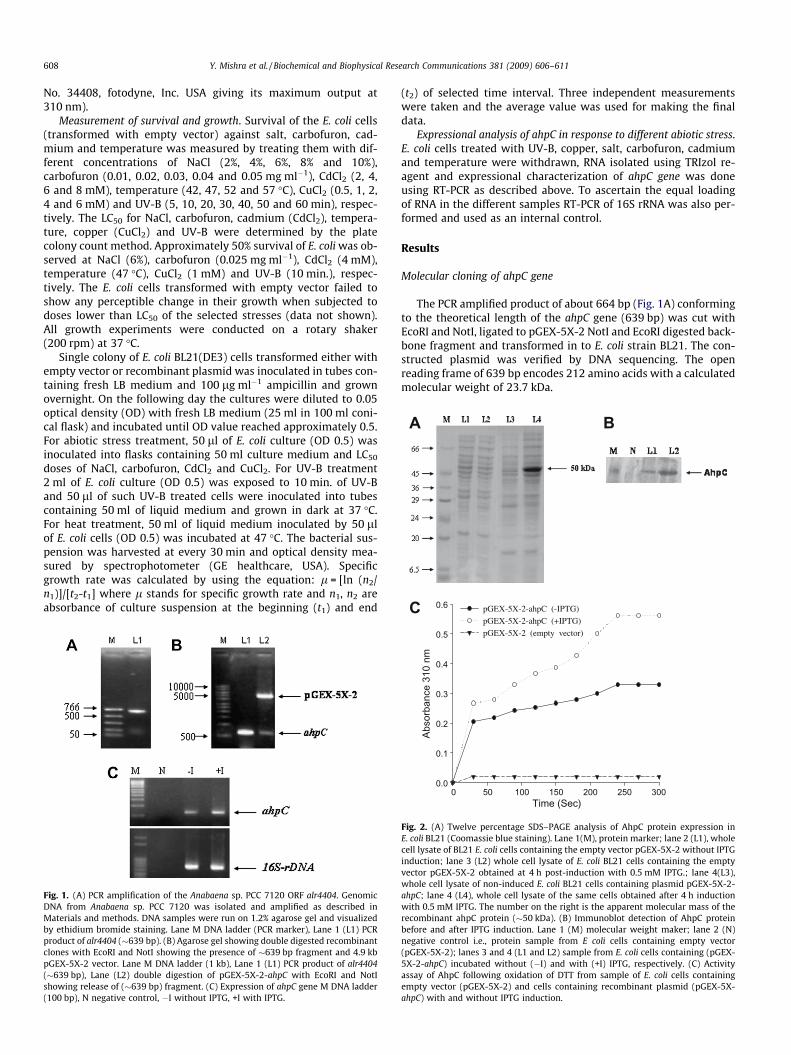
Fig. 1. (A) PCR amplification of the Anabaena sp. PCC 7120 ORF alr4404. GenomicDNA from Anabaena sp. PCC 7120 was isolated and amplified as described inMaterials and methods. DNA samples were run on 1.2% agarose gel and visualizedby ethidium bromide staining. Lane M DNA ladder (PCR marker), Lane 1 (L1) PCRproduct of alr4404 (�639 bp). (B) Agarose gel showing double digested recombinantclones with EcoRI and NotI showing the presence of �639 bp fragment and 4.9 kbpGEX-5X-2 vector. Lane M DNA ladder (1 kb), Lane 1 (L1) PCR product of alr4404(�639 bp), Lane (L2) double digestion of pGEX-5X-2-ahpC with EcoRI and NotIshowing release of (�639 bp) fragment. (C) Expression of ahpC gene M DNA ladder(100 bp), N negative control, �I without IPTG, +I with IPTG.
(t2) of selected time interval. Three independent measurementswere taken and the average value was used for making the finaldata.
Expressional analysis of ahpC in response to different abiotic stress.E. coli cells treated with UV-B, copper, salt, carbofuron, cadmiumand temperature were withdrawn, RNA isolated using TRIzol re-agent and expressional characterization of ahpC gene was doneusing RT-PCR as described above. To ascertain the equal loadingof RNA in the different samples RT-PCR of 16S rRNA was also per-formed and used as an internal control.
Results
Molecular cloning of ahpC gene
The PCR amplified product of about 664 bp (Fig. 1A) conformingto the theoretical length of the ahpC gene (639 bp) was cut withEcoRI and NotI, ligated to pGEX-5X-2 NotI and EcoRI digested back-bone fragment and transformed in to E. coli strain BL21. The con-structed plasmid was verified by DNA sequencing. The openreading frame of 639 bp encodes 212 amino acids with a calculatedmolecular weight of 23.7 kDa.
Time (Sec)0 50 100 150 200 250 300
Abso
rban
ce 3
10 n
m
0.0
0.1
0.2
0.3
0.4
0.5pGEX-5X-2-ahpC (+IPTG)
pGEX-5X-2 (empty vector)
Fig. 2. (A) Twelve percentage SDS–PAGE analysis of AhpC protein expression inE. coli BL21 (Coomassie blue staining). Lane 1(M), protein marker; lane 2 (L1), wholecell lysate of BL21 E. coli cells containing the empty vector pGEX-5X-2 without IPTGinduction; lane 3 (L2) whole cell lysate of E. coli BL21 cells containing the emptyvector pGEX-5X-2 obtained at 4 h post-induction with 0.5 mM IPTG.; lane 4(L3),whole cell lysate of non-induced E. coli BL21 cells containing plasmid pGEX-5X-2-ahpC; lane 4 (L4), whole cell lysate of the same cells obtained after 4 h inductionwith 0.5 mM IPTG. The number on the right is the apparent molecular mass of therecombinant ahpC protein (�50 kDa). (B) Immunoblot detection of AhpC proteinbefore and after IPTG induction. Lane 1 (M) molecular weight maker; lane 2 (N)negative control i.e., protein sample from E coli cells containing empty vector(pGEX-5X-2); lanes 3 and 4 (L1 and L2) sample from E. coli cells containing (pGEX-5X-2-ahpC) incubated without (�I) and with (+I) IPTG, respectively. (C) Activityassay of AhpC following oxidation of DTT from sample of E. coli cells containingempty vector (pGEX-5X-2) and cells containing recombinant plasmid (pGEX-5X-ahpC) with and without IPTG induction.
Y. Mishra et al. / Biochemical and Biophysical Research Communications 381 (2009) 606–611 609
Expressional analysis of ahpC gene in E. coli using RT-PCR and SDS–PAGE
A 3–4 h exposure of cells transformed with recombinant plas-mid to 0.5 mM IPTG was found to produce 4.5-fold increase inthe transcript level compared to non-induced cells (Fig. 1C). Like-wise, the fusion protein (on the SDS–PAGE) also showed inductionafter IPTG treatment (Fig. 2A).The molecular weight of GST-AhpCwas found to be �50 kDa.
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
pGEX-5X-2 (empty vector)
pGEX-5X-2 (empty vector) + 47ºCpGEX-5X-2-ahpC (recombinant plasmid) + 47ºC
Time (h)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
pGEX -5X-2( empty vector)
pGEX -5X-2 (empty vector) 2.5 mg carbofuron/100ml
pGEX-5X-2-ahpC (recombinant plasmid ) 2.5 mg carbofuron /100ml
0.0 2.5 5.0 7.5 10.0 12.5
Time (h)0.0 2.5 5.0 7.5 10.0 12.5
Time (h)0.0 2.5 5.0 7.5 10.0 12.5
Ab
sorb
an
ce (
60
0n
m)
Ab
sorb
an
ce (
60
0n
m)
Ab
sorb
an
ce (
60
0n
m)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
pGEX - 5X-2(empty vector)
pGEX-5X-2(empty vector) +1mM CuCl2pGEX-5X-2-ahpC(recombinant plasmid) + 1mMCuCl2
Fig. 3. Effect of (A) heat, (B) salt, (C) carbofuron (pesticide), (D) cadmium, (E) copper and(pGEX-5X-2-ahpC) and empty vector (pGEX-5X-2).The mean of three independent replicgrowth curves of E. coli cells in liquid medium exposed to heat, salt, carbofuron, cadmiu
Western blotting and enzymatic activity assay
Western blot analysis with anti-AhpC produced a single band of�50 kDa showing good agreement with the molecular weight(49.7) of ahpC gene deduced from the nucleotide sequence. Thisconfirmed that E. coli cells containing pGEX-5X-2-ahpC producedan AhpC protein (Fig. 2B). The activity of alkyl hydroperoxidereductase was assayed by DTT oxidation in the presence of theperoxide as substrate and measured by monitoring the change in
pGEX-5X-2 (empty vector)pGEX-5X-2 (empty vector) + 6% NaCl pGEX-5X-2-ahpC (recombinant plasmid) + 6% NaCl
pGEX-5X-2 (empty vector)pGEX-5X-2 (empty vector) + 4mM CdCl2
pGEX-5X-2-ahpC (recombinant plasmid) + 4mM CdCl
Time (h)0.0 2.5 5.0 7.5 10.0 12.5
Time (h)0.0 2.5 5.0 7.5 10.0 12.5
pGEX-5X-2(empty vector)
pGEX-5X-2(empty vector) 10min UV-B
pGEX-5X-2-ahpC(recombinant vector)+10min UV-B
Time (h)0.0 2.5 5.0 7.5 10.0 12.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Ab
sorb
an
ce (
60
0n
m)
Ab
sorb
an
ce (
60
0n
m)
Ab
sorb
an
ce (
60
0n
m)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
(F) UV-B on the growth of transformed E. coli cells containing recombinant plasmidates are plotted with error bars indicating standard deviations. A–F represents them, copper and UV-B, respectively.

610 Y. Mishra et al. / Biochemical and Biophysical Research Communications 381 (2009) 606–611
OD at 310 nm due to formation of the DTT disulfide. The cells con-taining (pGEX-5X-2-ahpC) plasmid showed increased activity overthose containing empty vector. This level was further increasedafter IPTG treatment. Control cells containing empty plasmidpGEX-5X-2 did not depict any enzymatic activity (Fig. 2C).
Effect of various abiotic stresses on growth of transformed E. coli cells
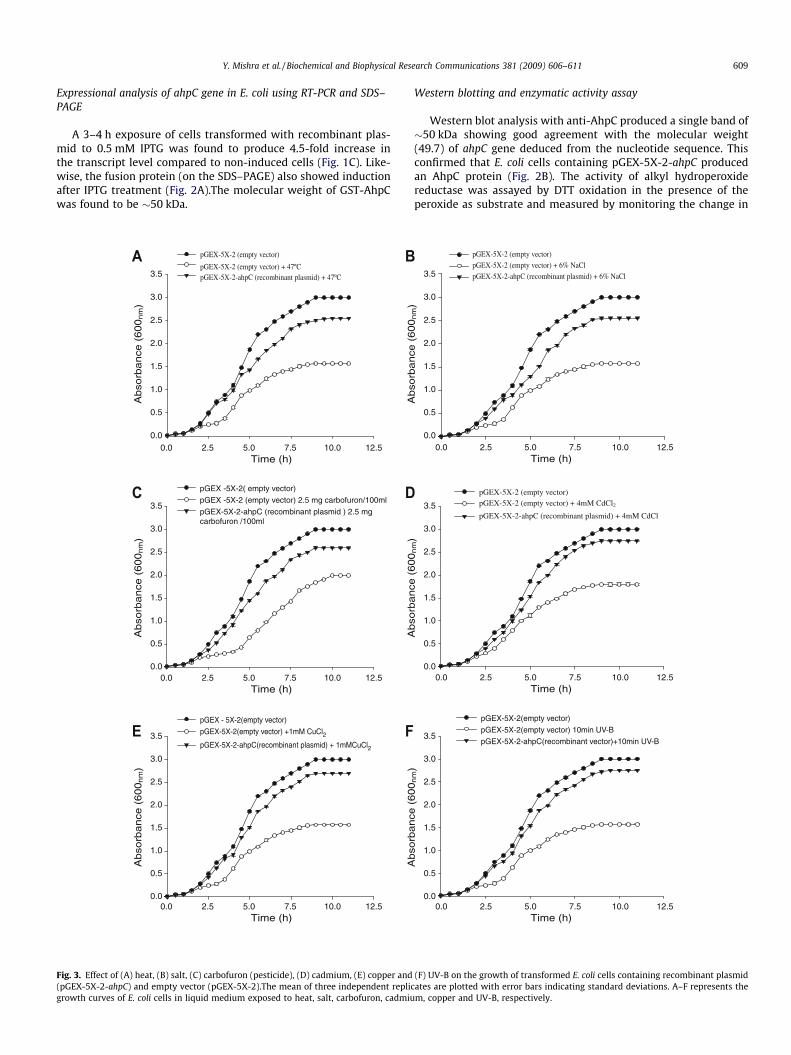
The transformed cells with recombinant plasmid (pGEX-5X-2-ahpC) showed better growth than those transformed with emptyvector (Fig. 3A–F). When LC50 dose for cells transformed withempty vector was applied on pGEX-5X-2-ahpC transformed cells,the decrease in specific growth rate was only 15%, 20%, 8%, 5%,25% and 23% under heat, salt, carbofuron, cadmium, copper andUV-B stress, respectively.
Expression of ahpC in response to various abiotic stresses
All the selected stresses triggered significant increase in thetranscript level of ahpC gene at various time points (Fig. 4A–F). Thisincrease being 1.10-, 1.23-, 2.51-fold in heat; 1.07-, 2.0-, 2.48-foldin salt; 1.98-, 2.34-, 2.42-fold in carbofuron; 1.72-, 2.42-, 2.54-foldin cadmium; 1.69-, 1.72-, 1.77-fold in copper and 1.58-, 1.91-,2.09-fold in UV-B over control after 3, 6 and 9 h, respectively.
Discussion
Molecular analysis of cyanobacterial ahpC revealed its involve-ment against heat, salt, carbofuron, cadmium, copper and UV-Bstresses. Specific growth rate of pGEX-5X-2-ahpC transformedE. coli cells was about 20–30% better than cells transformed with
Fig. 4. RT-PCR analysis of ahpC gene exposed to (A) heat, (B) salt, (C) carbofuro
pGEX-5X-2 under different stresses (Fig. 3A–F). The abiotic stressesused in the present study possessed a common mode of action e.g.disruption of redox homeostasis and generation of reactive oxygenspecies (ROS) resulting in impairment of key metabolic, regulatoryand dissipative pathways. The cyanobacterial ahpC gene in E. coliconceivably quenches the ROS and RSS thereby ameliorating thedestructive impacts caused by above stresses. It finds support fromNguyen-nhu and Knoops [4] where Ahp1p from S. cerevisiae pro-tected against metals. Similarly, Guimaraes et al. [12] reported thatAhpC is a key element protecting Mycobacterium tuberculosis fromoxidative stress. Notwithstanding above, the enhanced tolerance ofE. coli containing ahpC may be due to activation of the transcrip-tional activity of stress responsive genes. It is quite likely thatthe AhpC protein of transformed E. coli may act as general reduc-tant performing the general folding or chaperoning function asknown for heat shock proteins (HSPs). This speculation finds sup-port from the present result as well as the report of Antelmannet al. [6] where a new class of HSP III induced by heat or salt stressin strictly rB independent manner was identified as AhpC. Thus,we presume that increased accumulation of AhpC in E. coli cells of-fers protection against above stresses. To explore the relationshipbetween environmental stresses and ahpC induction, the expres-sion of ahpC gene was attested by RT-PCR (Fig. 4A–F). A significantup-regulation of ahpC transcript as observed under different stres-ses at different time points suggested stress dependent differentialregulation of ahpC gene. Our results are also supported by the re-port where up-regulation of ahpC gene was triggered by environ-mental stresses [13,14]. However, the spiking of IPTG in theculture medium failed to improve the stress tolerance of cellsexpressing cyanobacterial ahpC gene (data not shown). It is worthspeculating that a sufficient amount of AhpC protein for combatingstresses was already synthesized in the cells even without IPTG.
n (pesticide), (D) cadmium (E) copper and (F) UV-B at various time points.

Y. Mishra et al. / Biochemical and Biophysical Research Communications 381 (2009) 606–611 611
These results confirmed the hypothesis, that enhanced expres-sion of ahpC gene from the Anabaena sp. PCC 7120, a heterologoussource, can confer tolerance in non-photosynthetic E. coli cellsagainst heat, salt, pesticide (carbofuron), cadmium, copper andUV-B. Therefore, engineering of this trait (ahpC) of Anabaena sp.PCC 7120 may open newer possibilities for development of agenetically modified cyanobacterium with greater agility toaccommodate environmental hardship and fix nitrogen under abi-otic stresses rampant in the paddy fields.
Acknowledgments
L.C. Rai is thankful to DST for financial support, and YogeshMishra and Neha Chaurasia to CSIR and UGC for SRF, respectively.We thank M. Gopinath, Department of Microbiology and Cell Biol-ogy, Indian Institute of Science, Bangalore, India for pGEX-5X-2,and BL21, and Dr. Shashi Pandey for help in cloning experimentsand the Program Coordinator, CAS in Botany for facilities.
References
[1] K.H. Kim, S.G. Rhee, Sequence of peptides from Saccharomyces cerevisiaeglutamine synthetase N-terminal peptide and ATP-binding domain, J. Biol.Chem. 263 (1988) 833–838.
[2] F.S. Jacobson, R.W. Morgan, M.F. Christman, B.N. Ames, An alkyl hydroperoxidereductase from Salmonella typhimurium involved in the defense of DNA againstoxidative damage: purification and properties, J. Biol. Chem. 264 (1989) 1488–1496.
[3] H.Z. Chae, T.B. Uhm, S.G. Rhee, Dimerization of thiol-specific antioxidant andthe essential role of cysteine 47, Proc. Natl. Acad. Sci. USA 91 (1994) 7022–7026.
[4] N.T. Nguyen-nhu, B. Knoops, Alkylhydroperoxide reductase I protectsSaccharomyces cerevisie against metal ion toxicity and glutathione depletion,Toxicol. Lett. 135 (2002) 219–228.
[5] C.M. Reynolds, J. Meyer, L.B. Poole, An NADH-dependent bacterial thioredoxinreductase-like protein in conjunction with a glutaredoxin homologue form aunique peroxiredoxin (AhpC) reducing system in Clostridium pasteurianum,Biochemistry 41 (2002) 1990–2001.
[6] H. Antelmann, S. Engelmann, R. Schmid, M. Hecker, General and oxidativestress responses in Bacillus subtilis: cloning, expression and mutation of thealkyl hydroperoxide reductase operon, J. Bacteriol. 178 (1996) 6571–6578.
[7] N.H. Matsuda, K. Motohashi, H. Yoshimura, A. Nozaki, K. Inoue, M. Ohmohri,Antioxidative stress system in cyanobacteria, J. Biol. Chem. 280 (2005) 840–846.
[8] R. Rippka, J. Deruelles, J.B. Waterbury, M. Herdman, R.Y. Stanier, Genericassignments, strain histories and properties of pure cultures of cyanobacteria,J. Gen. Microbiol. 111 (1979) 1–6.
[9] J. Sambrook, D.W. Russell, Molecular Cloning: A Laboratory Manual, ColdSpring Harbor Laboratory Press, Cold Spring Harbor, New York, 2001.
[10] A.K. Srivastava, A. Ara, P. Bhargava, Y. Mishra, S.P. Rai, L.C. Rai, A rapid and cost-effective method of genomic DNA isolation from cyanobacterial culture, matand soil suitable for genomic fingerprinting and community analysis, J. Appl.Phycol. 19 (2007) 373–382.
[11] K.S. Iyer, W.A. Klee, Direct spectrophotometer measurement of the rate ofreduction of disulfide bonds. The reactivity of the disulfide bonds of bovinelactalbumin, J. Biol. Chem. 248 (1973) 707–710.
[12] B.G. Guimaraes, H. Souchon, N. Honore, B.S. Joanis, R. Brosch, W. Shepard, S.T.Cole, P.M. Alzari, Structure and mechanism of the alkyl hydroperoxidase AhpC,a key element of the Mycobacterium tuberculosis defense system againstoxidative stress, J. Biol. Chem. 280 (2005) 25735–25742.
[13] S. Mongkolsuk, W. Whangsuk, M. Fuangthong, S. Loprasert, Mutation in OxyRresulting in peroxide resistance in Xanthomonas campestris, J. Bacteriol. 182(2000) 3846–3849.
[14] U.A. Ochsner, M.L. Vasil, E. Alsabbagh, K. Parvatiyar, D.J. Hassett, Role ofPseudomonas aeruginosa oxyR-recG-operon in oxidative stress defense andDNA repair: Oxy R-dependent regulation of katB-ankB and ahpC-ahpF, J.Bacteriol. 182 (2000) 4533–4544.
Related Documents











