Age-associated changes in CD90 expression on thymocytes and in TCR-dependent stages of thymocyte maturation in male rats Gordana Leposavic ´ a,b, * , Vesna Pes ˇic ´ b , Dus ˇko Kosec a , Katarina Radojevic ´ a , Nevena Arsenovic ´-Ranin c , Ivan Pilipovic ´ a , Milica Peris ˇic ´ a , Bosiljka Plec ´as ˇ-Solarovic ´ b a Institute of Immunology and Virology ‘Torlak’, Immunology Research Center ‘Branislav Jankovic ´’, Belgrade, Serbia and Montenegro b Department of Physiology, Faculty of Pharmacy, Belgrade, Serbia and Montenegro c Department of Immunology and Microbiology, Faculty of Pharmacy, Belgrade, Serbia and Montenegro Received 18 October 2005; received in revised form 1 March 2006; accepted 7 March 2006 Available online 2 May 2006 Abstract To elucidate the effects of ageing on T-cell-maturation, in 3- and 18-month-old rats, we analysed the expression of: (i) CD4/CD8/TCRab and (ii) Thy-1, which is supposed to be a regulator of TCRab signalling, and thereby the thymocyte selection thresholds. Since an essential role for TCRab signalling in the development of CD4C25CT reg -cells was suggested, the frequency of these cells was also quantified. We demonstrated that, as for mice, early thymocyte differentiational steps within the CD4-8- double negative (DN) developmental stage are age-sensitive. Furthermore, we revealed that TCRab-dependent stages of T-cell development are affected by ageing, most likely due to an impaired expression of Thy-1 on TCRab low thymocytes entering selection processes. The diminished frequency of the post-selection CD4C8C double positive (DP) cells in aged rats, together with an overrepresentation of mature single positive (SP) cells, most probably suggests more efficient differentiational transition from the DP TCRab high to the SP TCRab high developmental stage, which is followed by an increase in pre-migration proliferation of the mature SP cells. Moreover, the study indicated impaired intrathymic generation of CD4C25CT reg -cells in aged rats, thus providing a possible explanation for the increased frequency of autoimmune diseases in ageing. q 2006 Elsevier Inc. All rights reserved. Keywords: Ageing; T-cell differentiation; CD90 expression; Thymocyte apoptosis; ConA; CD4C25C thymocytes 1. Introduction Empirical data on a substantial increase of morbidity and mortality due to infectious diseases in old age (Grubeck- Loebenstein, 1997; Aspinall, 2000) suggest a decline in the efficiency of immune system function with age. In the same line are findings of diminished protective immunity following prophylactic vaccination against influenza, and re-emergence of such latent infections as Varicella zoster in aged individuals (Globerson and Effros, 2000). Moreover, an increase in the frequency of cancer and autoimmune diseases was also found to accompany ageing (Wick et al., 2000). However, contrary to the classical view of ‘immunosenescence’ as the age- associated decline in immune response, there is an emerging consensus that immunological ageing is more related to a complex reorganization than to a simple decline in immune system functions (Globerson and Effros, 2000; Romanyukha et al., 2003). The involution of the thymic lymphoepithelial component is one of the most prominent features of ageing in the immune system. Based on data available in animals and humans, it is generally accepted that the volume of true thymic tissue (excluding the perivascular space, and the adipose and fibrous tissue) attains maximum size at puberty, after, which it gradually decreases (Fabris et al., 1988; George and Ritter, 1996; Bodey et al., 1997; Shanker, 2004). Therefore, with ageing, the thymic tissue weakens as a source of naive T lymphocytes (Brill et al., 1990; Consolini et al., 2000; Romanyukha et al., 2003). The reduced T-cell output, together with an increase in apoptosis of naive T-cells limits the ability of aged individuals to respond to newly encountered antigens. The markedly reduced size of the naive T-cell subpopulation, together with an increased number of memory cells in the periphery, is a clear-cut characteristic of ageing in the immune system (Romanyukha et al., 2003). Experimental Gerontology 41 (2006) 574–589 www.elsevier.com/locate/expgero 0531-5565/$ - see front matter q 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.exger.2006.03.006 * Corresponding author. Address: Department of Physiology, Faculty of Pharmacy, 450 Vojvode Stepe, 11151 Belgrade (KumodraZ ˇ ), Serbia and Montenegro. Tel.: C381 11 39 70 379; fax: C381 11 46 74 65. E-mail address: [email protected] (G. Leposavic ´).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Age-associated changes in CD90 expression on thymocytes and in
TCR-dependent stages of thymocyte maturation in male rats
Gordana Leposavic a,b,*, Vesna Pesic b, Dusko Kosec a, Katarina Radojevic a,
Nevena Arsenovic-Ranin c, Ivan Pilipovic a, Milica Perisic a, Bosiljka Plecas-Solarovic b
a Institute of Immunology and Virology ‘Torlak’, Immunology Research Center ‘Branislav Jankovic’, Belgrade, Serbia and Montenegrob Department of Physiology, Faculty of Pharmacy, Belgrade, Serbia and Montenegro
c Department of Immunology and Microbiology, Faculty of Pharmacy, Belgrade, Serbia and Montenegro
Received 18 October 2005; received in revised form 1 March 2006; accepted 7 March 2006
Available online 2 May 2006
Abstract
To elucidate the effects of ageing on T-cell-maturation, in 3- and 18-month-old rats, we analysed the expression of: (i) CD4/CD8/TCRab and
(ii) Thy-1, which is supposed to be a regulator of TCRab signalling, and thereby the thymocyte selection thresholds. Since an essential role for
TCRab signalling in the development of CD4C25CTreg-cells was suggested, the frequency of these cells was also quantified. We demonstrated
that, as for mice, early thymocyte differentiational steps within the CD4-8- double negative (DN) developmental stage are age-sensitive.
Furthermore, we revealed that TCRab-dependent stages of T-cell development are affected by ageing, most likely due to an impaired expression
of Thy-1 on TCRablow thymocytes entering selection processes. The diminished frequency of the post-selection CD4C8C double positive (DP)
cells in aged rats, together with an overrepresentation of mature single positive (SP) cells, most probably suggests more efficient differentiational
transition from the DP TCRabhigh to the SP TCRabhigh developmental stage, which is followed by an increase in pre-migration proliferation of the
mature SP cells. Moreover, the study indicated impaired intrathymic generation of CD4C25CTreg-cells in aged rats, thus providing a possible
explanation for the increased frequency of autoimmune diseases in ageing.
q 2006 Elsevier Inc. All rights reserved.
Keywords: Ageing; T-cell differentiation; CD90 expression; Thymocyte apoptosis; ConA; CD4C25C thymocytes
1. Introduction
Empirical data on a substantial increase of morbidity and
mortality due to infectious diseases in old age (Grubeck-
Loebenstein, 1997; Aspinall, 2000) suggest a decline in the
efficiency of immune system function with age. In the same
line are findings of diminished protective immunity following
prophylactic vaccination against influenza, and re-emergence
of such latent infections as Varicella zoster in aged individuals
(Globerson and Effros, 2000). Moreover, an increase in the
frequency of cancer and autoimmune diseases was also found
to accompany ageing (Wick et al., 2000). However, contrary
to the classical view of ‘immunosenescence’ as the age-
associated decline in immune response, there is an emerging
0531-5565/$ - see front matter q 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.exger.2006.03.006
* Corresponding author. Address: Department of Physiology, Faculty of
Pharmacy, 450 Vojvode Stepe, 11151 Belgrade (KumodraZ), Serbia and
Montenegro. Tel.: C381 11 39 70 379; fax: C381 11 46 74 65.
E-mail address: [email protected] (G. Leposavic).
consensus that immunological ageing is more related to a
complex reorganization than to a simple decline in immune
system functions (Globerson and Effros, 2000; Romanyukha
et al., 2003).
The involution of the thymic lymphoepithelial component is
one of the most prominent features of ageing in the immune
system. Based on data available in animals and humans, it is
generally accepted that the volume of true thymic tissue
(excluding the perivascular space, and the adipose and fibrous
tissue) attains maximum size at puberty, after, which it
gradually decreases (Fabris et al., 1988; George and Ritter,
1996; Bodey et al., 1997; Shanker, 2004). Therefore, with
ageing, the thymic tissue weakens as a source of naive T
lymphocytes (Brill et al., 1990; Consolini et al., 2000;
Romanyukha et al., 2003). The reduced T-cell output, together
with an increase in apoptosis of naive T-cells limits the ability
of aged individuals to respond to newly encountered antigens.
The markedly reduced size of the naive T-cell subpopulation,
together with an increased number of memory cells in the
periphery, is a clear-cut characteristic of ageing in the immune
system (Romanyukha et al., 2003).
Experimental Gerontology 41 (2006) 574–589
www.elsevier.com/locate/expgero

G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589 575
It has been supposed that the decline in thymic function with
ageing is a consequence of changes at both the prethymic
thymocyte precursor level and the intrathymic stromal cell
level (Hirokawa et al., 1986; Yu et al., 1997; Mackall et al.,
1998). The underlying mechanisms of age-related thymic
involution have not been fully depicted yet, although clear
experimental support for both non-genetic, i.e. hormonal
(Kelley et al., 1986; Fabris et al., 1988, 1997; Li et al., 1992;
Hirokawa et al., 1997; Leposavic et al., 2002), neural (Madden
et al., 1997) and genetic mechanisms (Lau and Spain, 2000;
Hsu et al., 2005) exists. Moreover, in accordance with the well-
documented symbiotic relation between the developing T
lymphocytes and the thymic nonlymphoid component (Ritter
and Boyd, 1993), it has been suggested that the age-related
alterations in thymocyte development contribute to thymic
involution (Lau and Spain, 2000).
Multifaceted age-related changes in both the multiple-step
process of intrathymic T-cell maturation and its intrathymic
regulation have been assumed. The earliest thymocytes
lack functional TCR, as well as CD4 and CD8 (Michie and
Zuniga-Pflucker, 2002; Zamoyska and Lovatt, 2004). Within
this stage thymocytes express an invariant pre-Ta protein and
rearrange the TCRb locus. Successful rearrangement of the
TCRb locus precedes its covalent pairing with the pre-Tachain. This is known as b selection, while subsequent
association with CD3 produces the functional pre-TCRacomplex (Michie and Zuniga-Pflucker, 2002; Zamoyska and
Lovatt, 2004). These cells are signalled to proliferate and turn
on expression of the CD4 and CD8 genes, becoming double
positive (DP) cells (Jameson et al., 1995; Guidos, 1996; Michie
and Zuniga-Pflucker, 2002; Zamoyska and Lovatt, 2004).
Thymocytes unable to rearrange their TCRb locus successfully
are deleted by apoptosis. Following TCRa locus rearrange-
ment, the thymocytes interact with epithelial cells expressing
an optimised set of self-peptides within the MHC complex.
From this point, the thymocyte will survive if signals generated
by TCR engagement by MHC/peptide are interpreted as
appropriate (positive selection), but will be deleted by
apoptosis if the generated signals are interpreted as either
inappropriately weak (death by neglect) or inappropriately
strong (negative selection) (Jameson et al., 1995; Guidos,
1996; Michie and Zuniga-Pflucker, 2002; Zamoyska and
Lovatt, 2004). The positively selected thymocytes upregulate
TCRab expression and downregulate one of the coreceptors,
either CD4 or CD8, to give the mature MHC-restricted self-
tolerant CD4C or CD8C single positive (SP) mature T-cells
(Jameson et al., 1995; Guidos, 1996; Zamoyska and Lovatt,
2004). Since, thymocytes progress through phenotypically
well-defined steps during maturation, T-cell maturation can be
monitored by analysis of the distribution of thymocyte subsets.
Some age-sensitive maturational steps during early thymocyte
maturation have been clearly identified in mice (Brill et al.,
1990; Yehuda et al., 1998). Moreover, it has been suggested
that later, TCR-dependent, checkpoints in T-cell development
may be also altered in the course of ageing in mice (Lau and
Spain, 2000). However, data related to the age-associated
changes in T-cell maturation in rat thymus are rather limited.
Therefore, this study was designed in order to identify subtle
thymocyte differentiational steps, which are affected by ageing,
with special focus on the late TCRab-dependent steps. To
reach this goal (i) the surface density of TCRab on thymocytes
in respect to CD4/CD8 expression and (ii) expression of Thy-1,
which is supposed to be a regulator of TCRab signalling, and
thereby of the thymocyte selection thresholds (Hueber et al.,
1997; Killeen, 1997), were examined. The rationale for the
analysis of Thy-1 is provided by data indicating that the age-
associated decline in thymus weight and thymic cellularity
leads to an increase in the density of noradrenergic nerve fibres
in the parenchyma and noradrenaline concentration in the
thymus (Madden et al., 1997), on the one hand, and by the
observation that exogenous cAMP and noradrenaline can
induce decreases in steady state Thy-1 mRNA levels in
T-lineage cells through posttranscriptional destabilization of
Thy-1 mRNA (Wajeman-Chao et al., 1998), on the other hand.
In addition, thymocyte apoptosis and proliferation, events
associated with thymocyte maturation, were also analysed.
Having in mind: (i) the age-related increase in the frequency
of self-reactive lymphocytes and autoimmune diseases (Wick
et al., 2000; Shanker, 2004) and (ii) that under normal
circumstances, some self-reactive T-cells fail to be eliminated
in the thymus and escape into the periphery where they are
exposed to continuous control by a dominant T-cell-mediated
protective mechanism produced by intrathymically generated
regulatory T-cells (Treg-cells) that express CD4 and CD25
(Seddon and Mason, 2000; Shevach, 2002; Godfrey et al.,
2000), one may assume that in aged individuals the
autoreactive T/Treg-cell balance is affected due either to an
absolute or a relative decline in the thymic production of Treg-
cells. To explore this hypothesis the relative number of CD4C25C cells in the thymus of both young adult and aged rats was
quantified.
2. Materials and methods
2.1. Animals
Male Wistar rats were purchased from the Military Medical
Academy Animal House, Belgrade, housed in polyethylene
cages containing sterilized wood shavings and given rodent
chow and tap water ad libitum. They were housed in a room
maintained at a temperature of 21G2 8C, with a 12 h light/dark
cycle. Rats were euthanized under ether anaesthesia by
exsanguination. The experimental protocol was approved by
the Institutional Animal Care and Use Committee.
Rats were euthanized at the age of either 3 (6 rats) or 18
months (5 rats). Animals that showed overt signs of illness,
including low body weight, tumours or splenomegaly were
excluded from the study. Each thymus was aseptically isolated,
trimmed of all excess body fat and gently blotted on sterile
gauze to remove excess blood. The two lobi were divided and
individually weighed. The left lobus was fixed for stereological
analysis, while the right lobus was dissociated by gently
grinding the tissue on a sterile 60-mm sieve screen in complete
RPMI-1640 medium. The single-cell suspension thus obtained

G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589576
was washed three times in ice-cold complete RPMI-1640
medium. The cells were then counted in an improved Neubauer
hemocytometer and the number of cells per milligram thymic
tissue, and consequently the total number of cells per thymus,
were estimated. The viability of such cell preparations,
determined by Trypan blue exclusion, was routinely greater
than 95%.
2.2. Chemicals, antibodies and immunoconjugates
Sodium azide, Merocyanine (MC540) and Concanavalin A
(Con A) were purchased from Sigma–Aldrich Chemie GmbH,
Taufkirchen, Germany. Dexamethasone (Dexason, Dex) was
obtained from Galenika A.D., Zemun, Serbia and Montenegro.
RPMI 1640 powdered medium (Sigma–Aldrich Chemie
GmbH) was dissolved in redistilled water according to the
manufacturer’s instructions. To prepare complete RPMI
medium, 2 mM L-glutamine (Serva, Heidelberg, Germany),
1 mM sodium pyruvate (Serva), 100 units/ml penicillin (ICN,
Costa Mesa, CA, USA), 100 mg/ml streptomycin (ICN) and
10% fetal calf serum (FCS) (Gibco, Grand Island, NY, USA)
were added. FCS was inactivated by heating the serum at 56 8C
for 30 min.
For immunolabeling the following monoclonal antibodies
(mAbs): phycoerythrin (PE)-conjugated anti-CD4 (clone
W3/25, Serotec, Oxford, UK), fluorescein-isothiocyanate
(FITC)-conjugated anti-CD8 (clone MRC OX-8, Serotec),
peridinin chlorophyll protein (PerCP)-conjugated anti-TCRab(clone R73, BD Biosciences Pharmingen, Mountain View, CA,
USA), biotin-conjugated anti-CD25 (clone MRC OX-39,
Serotec), FITC-conjugated anti-CD44 (Pgp-1) (clone OX-49,
Serotec), FITC-conjugated anti-CD161 (clone 10/78, Serotec),
FITC-conjugated anti-CD90 (Thy-1.1) (clone HIS 51, BD
Biosciences Pharmingen), anti-CD3 (clone G4.18, BD
Biosciences Pharmingen) and-RT6.1 (clone P4/16, Serotec)
were used as the first-step reagents. As the second-step
reagents, streptavidin-PerCP, streptavidin-PE, FITC-conju-
gated F(ab 0)2 rabbit anti-rat IgG purchased from Serotec,
were applied, respectively. The appropriate IgG isotype
controls were obtained from BD Biosciences Pharmingen.
2.3. Tissue processing and stereological analysis
After fixation in Bouin’s solution and dehydration in a
graded series of ethyl alcohol, the thymic specimens were
embedded in paraffin, serially cut into 5 mm thick sections,
which were then stained with haematoxylin and eosin. Every
40th section (approximately 20 sections per organ) was
subjected to analysis of the volume of lymphoepithelial thymic
tissue, denoted as true thymic tissue (George and Ritter, 1996;
Shanker, 2004), and thymic cellularity.
Stereological measurements were performed by a point
counting method (Weibel, 1979) using image analysis software
(Micro Image, Version 4.0, OLYMPUS). The test areas were
randomly chosen, and each image, acquired using a digital
camera, was saved, overlaid with the corresponding grid and
analysed.
Absolute volumes of the cortical and medullary lymphoe-
pithelial thymic compartments, and interlobular, subcapsular
and perivascular connective and adipose tissues, were
estimated from the volume of the processed and embedded
organ and the volume density (Vv) of the corresponding
compartment (the percentage of whole organ volume occupied
by the analysed structure). During processing and embedding
the thymic tissue shrunk by approximately 34% in both groups,
and this was determined stereologically, as previously
described (Plecas-Solarovic et al., 2004). Vv of each thymic
compartment was determined under !40 magnification, using
an orthogonal test grid with 130 points, according to the
equation: VvZPf/Pt (Pf represents the number of test points
hitting the analysed structure, while Pt is the total number of
test points falling on the organ). The total number of analysed
test-areas was 100 per animal.
The total number of thymocytes/thymic lobus was
calculated from the numerical density (Nv) of thymocytes, as
the number of cells per unit volume, and the absolute volume of
the lobus (Plecas-Solarovic et al., 2004). From the total number
of thymocytes/lobus the number of thymocytes/mg thymic
tissue, and consecutively the total number of thymocytes/
thymus, were calculated. The Nv of thymocytes was estimated
under immersion magnification, using a grid corresponding to
the multipurpose M42 test-system (Weibel, 1979). For each
thymic compartment 60 test areas per animal were measured.
Nv of thymocytes was determined according to Weibel and
Gomez (Weibel, 1979).
2.4. Phenotyping of lymphocytes by flow cytometry
analysis (FCA)
Immunolabeling of the cell suspensions was performed as
previously described (Leposavic et al., 2005). Briefly, aliquots
of 1!106 lymphoid cells in 100 ml RPMI 1640 medium were
centrifuged at 350g for 5 min, at 4 8C, to yield a pellet. The
cells were incubated for 30 min on ice in the dark with
fluorochrome-conjugated mAbs (direct labelling) or with
fluorochrome-conjugated and biotin-conjugated mAbs
(indirect labelling), and then washed three times in the
medium. When biotin-conjugated mAbs were applied, the
cells were incubated again for 30 min on ice with appropriate
fluorochrome-conjugated streptavidin. After labelling the cells
were washed twice in the medium, and then in ice-cold
phosphate buffered saline (PBS, pH 7.4) containing 0.01%
sodium azide. The cells were then fixed in 0.5 ml of 1%
paraformaldehyde in PBS and kept at 4 8C in the dark until
analysis. Twenty-thousand cells per sample were analysed
using a FACScan flow cytometer (Becton Dickinson, Mountain
View, CA, USA). Non-specific IgG isotype-matched controls
were used for each fluorochrome type to define background
staining, while dead cells and debris were excluded from
analysis by selective gating based on anterior and right-angle
scatter. The percentage of positive cells for each labelling was
determined using the CellQuest software (Becton Dickinson).
In addition, CD90 expression was analysed by estimation of the
mean channel number, which represents density of the surface

ged rats
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589 577
marker expression. The mean channel number was determined
for the thymocytes from both young adult and aged rats, and
the relative change in the mean intensity of fluorescence (MIF)
was calculated as follows:
MIF for histogram of young adult ratsKMIF for histogram of a
MIF for histogram of aged rats
(Kamath et al., 1998).
2.5. Detection of apoptotic thymocytes
Since, thymocytes are normally rapidly eliminated by
phagocytes in vivo, the relative number of apoptotic cells
was quantified after 18 h of cultivation, as previously suggested
(Kamath et al., 1998). One hundred microliters of freshly
isolated thymocytes at a concentration of 5!106 cells/ml were
plated in 96-well flat-bottom plates (Nunc A/S, Roskilde,
Denmark) in RPMI 1640 complete media. To these cells,
100 ml of complete medium containing Dex was added to attain
a final concentration of 1 mM Dex, a dose known to induce
apoptosis of rat thymocytes in vitro (Brown et al., 1993). In
control cultures 100 ml of complete medium was added. Cells
were harvested after 18 h of culture for analysis of the relative
numbers of apoptotic cells. After the estimation of cell viability
by Trypan blue dye exclusion, MC 540 and FITC-conjugated
Annexin V/propidium iodide (PI) assays were performed.
2.5.1. Detection of apoptotic thymocytes using lypophilic
dye MC 540
The staining was performed according to the procedure
described by Mower et al. (1994). For labelling, just before
FACS analysis, 5 ml of 1 mg/ml stock MC540 solution in
double-distilled H2O was added to 1 ml of thymocyte
suspension containing 106–107 cells. The stock solution of
MC540 was filtered through a 0.22 nm filter and stored in
the dark at 4 8C no longer than 1 month. All samples were
analysed on a FACScan flow cytometer by CellQuest
Software (Becton Dickinson). According to the intensity of
MC540 fluorescence, on the one hand, and forward scatter,
on the other hand, (Cohen, 1991), three subsets of
apoptotic cells can be distinguished: (1) cells in early
apoptosis, (2) cells in advanced apoptosis and (3) cells in
late apoptosis/necrosis. It has been reported that apoptotic
cells remain MC540 bright until their DNA content reaches
a certain point and then display decreasing levels of
MC540 staining (Reid et al., 1996). Thus, generally, the
cells in late apoptosis/necrosis are characterized by low
Table 1
Thymus weight, thymus weight/body weight ratio, volume of true thymic tissue, num
3- and 18-month-old Wistar rats
Absolute thymus
weight (mg)
Thymus weight/
body weight (%)
Volume o
thymic tis
3-Month-old 570.2G31.1 1.67G0.07 336.6G1
18-Month-old 482.5G104.2 0.7G0.1** 123.6G3
The data are expressed as meanGSEM (nZ5–7). *p!0.05, **p!0.01.
forward scatter and an undetectable level of MC540
binding.
2.5.2. Detection of apoptotic thymocytes using Annexin
V/propidium iodide
Thymocyte apoptosis was also examined using an Annexin
V/PI apoptosis detection kit (BD Biosciences Pharmingen).
Apoptotic cell staining was performed according to the
producer’s manual. Twenty thousand cells per sample were
analysed using a FACScan flow cytometer (Becton Dickinson)
and CellQuest software (Becton Dickinson). Since, it has been
shown that: (i) normal thymocytes undergoing spontaneous
apoptosis in vitro initially appear as Annexin VC/PI- cells and
(ii) the same cells, with increasing culture duration, become
Annexin VC/PIC cells (Okasha et al., 2001), Annexin VC/
PI- single positive cells are considered early apoptotic, whereas
Annexin VC/PIC double positive cells are believed to be in a
late stage of apoptosis rather than in necrosis (Okasha et al.,
2001;Vermes et al., 1995). Annexin VK/PIC single positive
cells are reckoned to be mainly dead cells (Okasha et al., 2001;
Vermes et al., 1995).
2.6. Detection of BrdUC cells
Bromodeoxyuridine (BrdU) incorporation was utilized to
identify DNA-synthesizing cells in vitro. A total of 2!105
cells/well (100 ml) of thymocytes from young adult or aged rats
were dispersed into plastic 96-well plates (Nunc A/S) and
cultured for 48 h at 37 8C in a 5% CO2 humified air atmosphere
without or with 2.5 mg/ml ConA in a total volume of 200 ml
complete RPMI 1640 culture medium. All cultures were run in
triplicate. During the last 18 h of culture the cells were pulsed
by 1 mM BrdU/well. For the detection of BrdU incorporation
into cells, the BrdU/7-AAD Flow kit (BD Biosciences
Pharmingen) was used. 7-AAD labelling enabled detection of
the late apoptotic/necrotic hypodiploid cells and subsequently
their exclusion from the analysis. To determine the phenotypic
characteristics of the BrdUC cells, the staining with FITC-
conjugated anti-BrdU Abs and 7-AAD was combined with
immunolabelling with biotin-conjugated anti-CD3 mAbs as the
first step-reagent, and streptavidin-PE as the second-step
reagent. The staining was performed according to the
BrdU/7-AAD flow kit producer’s manual. All samples were
ber ot thymocytes/mg thymic tissue and total number of thymocytes/thymus in
f true
sue (mm3)
Thymocyte number
(!106)/mg thymic tissue
Total thymocyte
number/thymus (!108)
9.2 1.47G0.25 8.41G0.53
9.4* 0.24G0.06** 1.21G0.45*
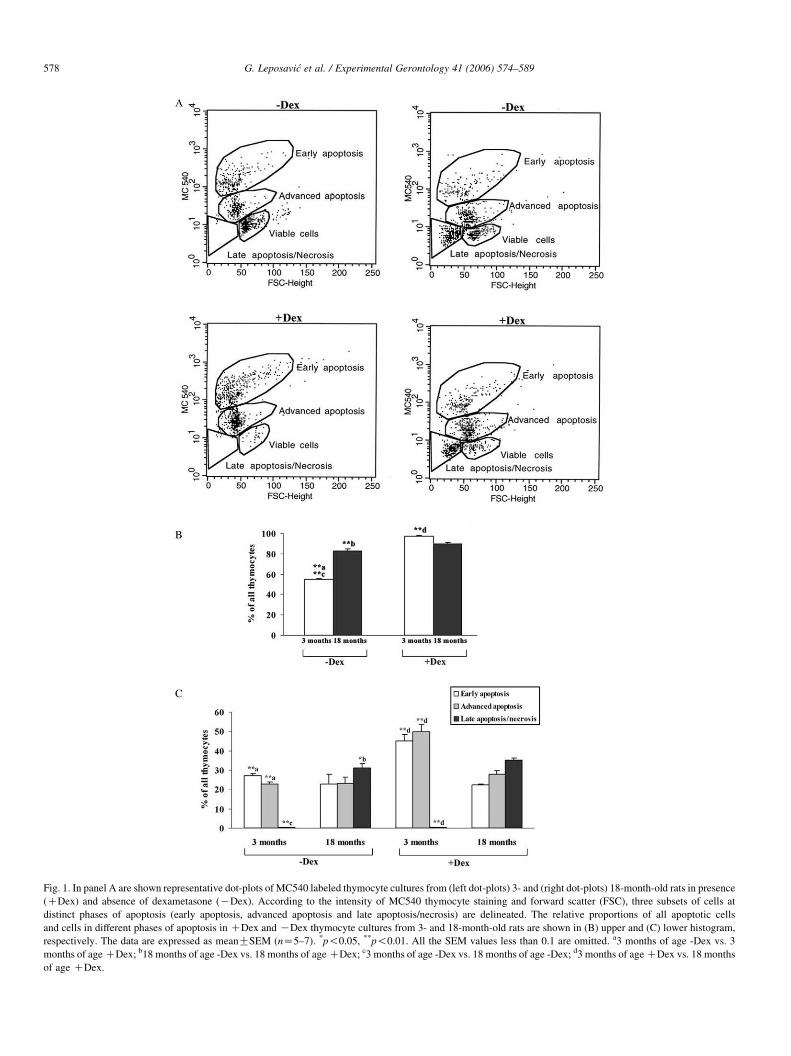
Fig. 1. In panel A are shown representative dot-plots of MC540 labeled thymocyte cultures from (left dot-plots) 3- and (right dot-plots) 18-month-old rats in presence
(CDex) and absence of dexametasone (KDex). According to the intensity of MC540 thymocyte staining and forward scatter (FSC), three subsets of cells at
distinct phases of apoptosis (early apoptosis, advanced apoptosis and late apoptosis/necrosis) are delineated. The relative proportions of all apoptotic cells
and cells in different phases of apoptosis in CDex and KDex thymocyte cultures from 3- and 18-month-old rats are shown in (B) upper and (C) lower histogram,
respectively. The data are expressed as meanGSEM (nZ5–7). *p!0.05, **p!0.01. All the SEM values less than 0.1 are omitted. a3 months of age -Dex vs. 3
months of age CDex; b18 months of age -Dex vs. 18 months of age CDex; c3 months of age -Dex vs. 18 months of age -Dex; d3 months of age CDex vs. 18 months
of age CDex.
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589578
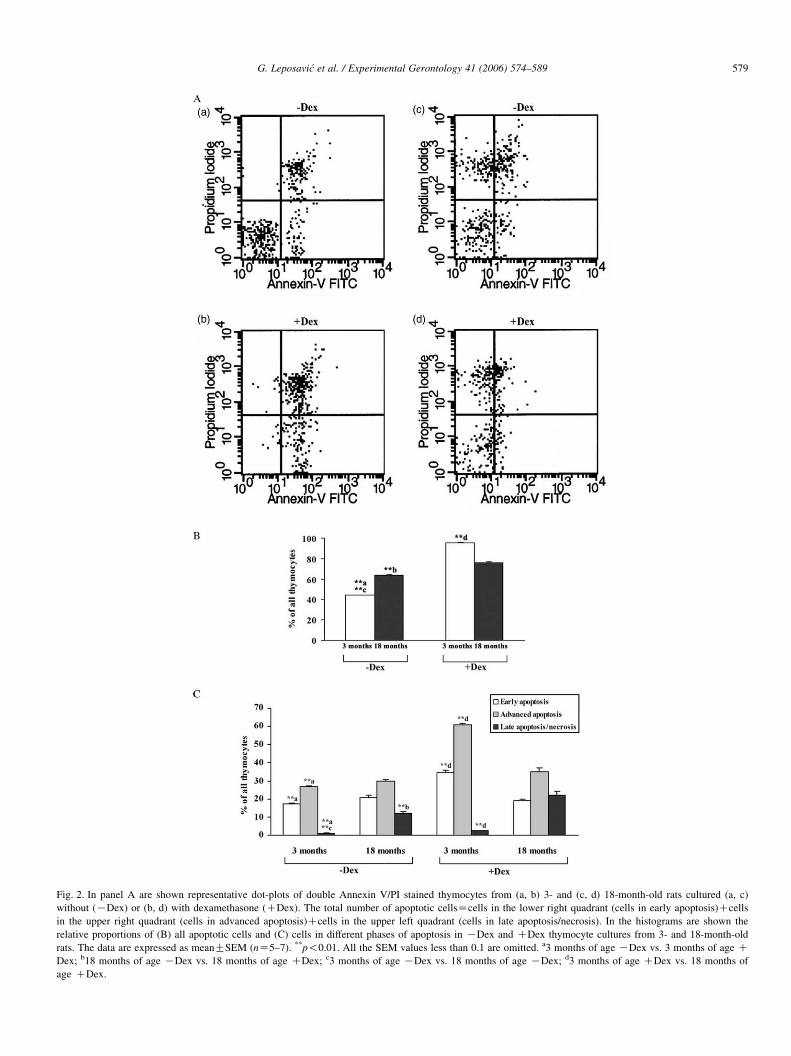
Fig. 2. In panel A are shown representative dot-plots of double Annexin V/PI stained thymocytes from (a, b) 3- and (c, d) 18-month-old rats cultured (a, c)
without (KDex) or (b, d) with dexamethasone (CDex). The total number of apoptotic cellsZcells in the lower right quadrant (cells in early apoptosis)Ccells
in the upper right quadrant (cells in advanced apoptosis)Ccells in the upper left quadrant (cells in late apoptosis/necrosis). In the histograms are shown the
relative proportions of (B) all apoptotic cells and (C) cells in different phases of apoptosis in KDex and CDex thymocyte cultures from 3- and 18-month-old
rats. The data are expressed as meanGSEM (nZ5–7). **p!0.01. All the SEM values less than 0.1 are omitted. a3 months of age KDex vs. 3 months of age CDex; b18 months of age KDex vs. 18 months of age CDex; c3 months of age KDex vs. 18 months of age KDex; d3 months of age CDex vs. 18 months of
age CDex.
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589 579

Fig. 3. Relative proportions of BrdUC cells in the thymocyte cultures from 3-
and 18-month-old rats, as determined by flow cytometric analysis of BrdU
incorporation. The thymocytes were cultured for 48h in medium without ConA
(KConA) or with ConA (CConA). The data are expressed as meanGSEM
(nZ5–7). **p!0.01. All the SEM values less than 0.03 are omitted. a3 months
of age KConA vs. 3 months of age CConA; b18 months of age KConA vs. 18
months of age CConA; c3 months of age KConA vs. 18 months of age
KConA; d3 months of age CConA vs. 18 months of age CConA.
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589580
analysed on the FACScan flow cytometer by CellQuest
software (Becton Dickinson).
2.7. Statistical analysis
Data are expressed as mean (X)Gstandard error of the mean
(SEM). To assess the significance of differences between
means, the Mann–Whitney U-test was applied using the
program, SPSS 10.0 for Windows. PR0.05 was considered
significant.
3. Results
3.1. Age-associated changes in thymus weight and total
number of thymocytes
There was no significant difference in the absolute weight of
the thymuses from 3- and 18-month-old rats. However, the
relative thymic weight (ratio to body weight) was significantly
(p!0.01) diminished in aged rats and its average value was
2.4-fold less than that in young adults (Table 1). Furthermore,
stereological analysis revealed that the average volume of
processed true thymic tissue was 2.7-fold (p!0.05) lower in
aged compared with young adult animals (Table 1). This
decrease in the volume of true thymic tissue reflected, at least
partly, a marked reduction in thymus cellularity. The number
of thymocytes per milligram of thymic tissue, and conse-
quently the total number of thymocytes per thymus, were
significantly reduced in aged rats compared with young adults
as determined by both direct counting of the cells in the
thymocyte suspension (0.16G0.06!106/mg thymic tissue in
aged vs. 1.81G0.23!106/mg thymic tissue in young adult rats,
p!0.05 and 0.79G0.17!108/thymus in aged vs. 10.74G1.26!108/thymus in young adult rats, p! 0.01) and
sterological analysis (Table 1).
3.2. Influence of ageing on thymocyte apoptosis
The reduced number of thymocytes may reflect: (i)
decreased ingress of T-cell precursors into the thymus; (ii)
increased elimination of thymocytes by apoptosis and (iii)
decreased proliferation of developing T-cells. To assess the
putative contribution of increased thymocyte apoptotic
elimination to the observed reduction in thymus cellularity,
not only spontaneous thymocyte apoptosis, but also the
sensitivity of thymocytes to apoptotic signalling induced by
glucocorticoids, as well known inducers of thymocyte
apoptosis, was examined. Using both MC540 and Annexin
V/PI assays it was demonstrated that the frequency of apoptotic
cells, in the absence of known inducers of apoptosis, was
significantly (p!0.01) greater in the cultures of thymocytes
from aged than in those from young adult rats (Figs. 1 and 2).
Furthermore, it was demonstrated that this increase in apoptotic
cells from aged rats reflected a drastic (p!0.01) rise in the
relative frequency of cells in late apoptosis/necrosis, while the
relative numbers of cells in earlier phases of apoptosis did not
significantly differ between cultures of thymocytes from rats of
different age (Figs. 1 and 2). This finding may indicate that
thymocytes from aged rats in culture underwent spontaneous
apoptosis earlier than those from young adult rats.
As we expected, both the assays for detection of apoptotic
cells demonstrated that Dex in the cultures of thymocytes
caused a significant (p!0.01) rise in the relative number of
apoptotic cells irrespective of the age of animals from, which
they were isolated (Figs. 1 and 2). However, to our surprise, in
the presence of Dex, the frequency of apoptotic cells in the
cultures of thymocytes from aged rats was significantly (p!0.01) lower than that in cultures of thymocytes from young
adults. Furthermore, the Dex-induced increase in the percen-
tage of apoptotic cells in cultures of thymocytes from young
adult rats reflected a significantly (p!0.01) increased
frequency of cells in early and advanced apoptosis, while
that in aged rats was associated with a significantly (p!0.05
and 0.01 by MC540 and Annexin V/PI staining, respectively)
increased frequency of cells in late apoptosis/necrosis (Figs. 1
and 2). Although, the overall number of apoptotic cells in
DexC cultures of thymocytes from aged rats was significantly
(p!0.01) lower than that in cultures of thymocytes from young
adult rats, it seems that these cells underwent apoptosis earlier
in the presence of Dex.
3.3. Influence of ageing on BrdU incorporation in thymocytes
In order to clarify, the mechanisms underlying the reduced
cellularity of thymus from aged animals, we attempted not only
to identify cells within thymocyte subpopulations delineated by
the surface density of TCR, which were actually involved in
DNA replication, but also to assess the proliferative potential
of these cell subpopulations. For this purpose BrdU incorpor-
ation in DNA was tested in cultures of thymocytes from young
adult and aged rats in the absence and presence of ConA. In the
absence of ConA the frequency of BrdUC was significantly
(p!0.01) lower in the thymocyte cultures from aged rats than
in those from young adults (Fig. 3). ConA evoked significant
increases in the frequency of BrdUC cells in the thymocyte
cultures from rats of both ages. However, ConA was
more effective in increasing the relative number of cells
incorporating BrdUC in cultures of thymocytes from young
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589 581
rats (the average number of BrdUC cells increased by 38%)
than in those from aged rats (the average number of BrdUCcells increased by 28%).
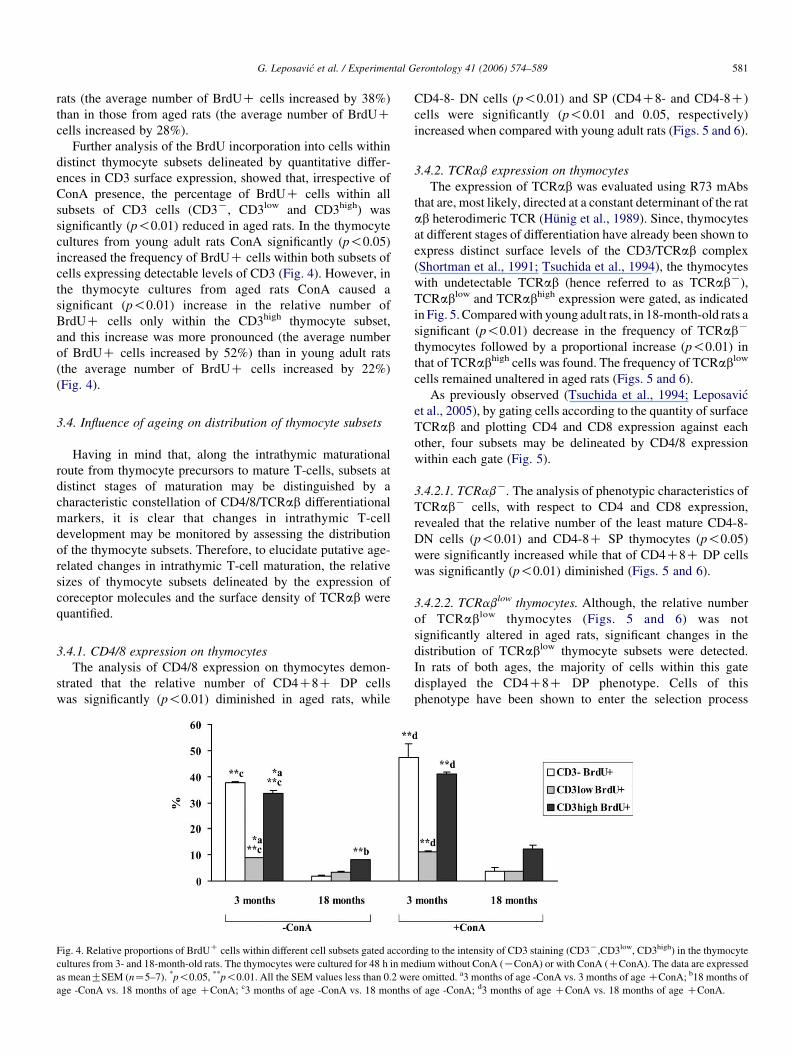
Further analysis of the BrdU incorporation into cells within
distinct thymocyte subsets delineated by quantitative differ-
ences in CD3 surface expression, showed that, irrespective of
ConA presence, the percentage of BrdUC cells within all
subsets of CD3 cells (CD3K, CD3low and CD3high) was
significantly (p!0.01) reduced in aged rats. In the thymocyte
cultures from young adult rats ConA significantly (p!0.05)
increased the frequency of BrdUC cells within both subsets of
cells expressing detectable levels of CD3 (Fig. 4). However, in
the thymocyte cultures from aged rats ConA caused a
significant (p!0.01) increase in the relative number of
BrdUC cells only within the CD3high thymocyte subset,
and this increase was more pronounced (the average number
of BrdUC cells increased by 52%) than in young adult rats
(the average number of BrdUC cells increased by 22%)
(Fig. 4).
3.4. Influence of ageing on distribution of thymocyte subsets
Having in mind that, along the intrathymic maturational
route from thymocyte precursors to mature T-cells, subsets at
distinct stages of maturation may be distinguished by a
characteristic constellation of CD4/8/TCRab differentiational
markers, it is clear that changes in intrathymic T-cell
development may be monitored by assessing the distribution
of the thymocyte subsets. Therefore, to elucidate putative age-
related changes in intrathymic T-cell maturation, the relative
sizes of thymocyte subsets delineated by the expression of
coreceptor molecules and the surface density of TCRab were
quantified.
3.4.1. CD4/8 expression on thymocytes
The analysis of CD4/8 expression on thymocytes demon-
strated that the relative number of CD4C8C DP cells
was significantly (p!0.01) diminished in aged rats, while
Fig. 4. Relative proportions of BrdUC cells within different cell subsets gated accord
cultures from 3- and 18-month-old rats. The thymocytes were cultured for 48 h in me
as meanGSEM (nZ5–7). *p!0.05, **p!0.01. All the SEM values less than 0.2 we
age -ConA vs. 18 months of age CConA; c3 months of age -ConA vs. 18 months
CD4-8- DN cells (p!0.01) and SP (CD4C8- and CD4-8C)
cells were significantly (p!0.01 and 0.05, respectively)
increased when compared with young adult rats (Figs. 5 and 6).
3.4.2. TCRab expression on thymocytes
The expression of TCRab was evaluated using R73 mAbs
that are, most likely, directed at a constant determinant of the rat
ab heterodimeric TCR (Hunig et al., 1989). Since, thymocytes
at different stages of differentiation have already been shown to
express distinct surface levels of the CD3/TCRab complex
(Shortman et al., 1991; Tsuchida et al., 1994), the thymocytes
with undetectable TCRab (hence referred to as TCRabK),
TCRablow and TCRabhigh expression were gated, as indicated
in Fig. 5. Compared with young adult rats, in 18-month-old rats a
significant (p!0.01) decrease in the frequency of TCRabK
thymocytes followed by a proportional increase (p!0.01) in
that of TCRabhigh cells was found. The frequency of TCRablow
cells remained unaltered in aged rats (Figs. 5 and 6).
As previously observed (Tsuchida et al., 1994; Leposavic
et al., 2005), by gating cells according to the quantity of surface
TCRab and plotting CD4 and CD8 expression against each
other, four subsets may be delineated by CD4/8 expression
within each gate (Fig. 5).
3.4.2.1. TCRabK. The analysis of phenotypic characteristics of
TCRabK cells, with respect to CD4 and CD8 expression,
revealed that the relative number of the least mature CD4-8-
DN cells (p!0.01) and CD4-8C SP thymocytes (p!0.05)
were significantly increased while that of CD4C8C DP cells
was significantly (p!0.01) diminished (Figs. 5 and 6).
3.4.2.2. TCRablow thymocytes. Although, the relative number
of TCRablow thymocytes (Figs. 5 and 6) was not
significantly altered in aged rats, significant changes in the
distribution of TCRablow thymocyte subsets were detected.
In rats of both ages, the majority of cells within this gate
displayed the CD4C8C DP phenotype. Cells of this
phenotype have been shown to enter the selection process
ing to the intensity of CD3 staining (CD3K,CD3low, CD3high) in the thymocyte
dium without ConA (KConA) or with ConA (CConA). The data are expressed
re omitted. a3 months of age -ConA vs. 3 months of age CConA; b18 months of
of age -ConA; d3 months of age CConA vs. 18 months of age CConA.
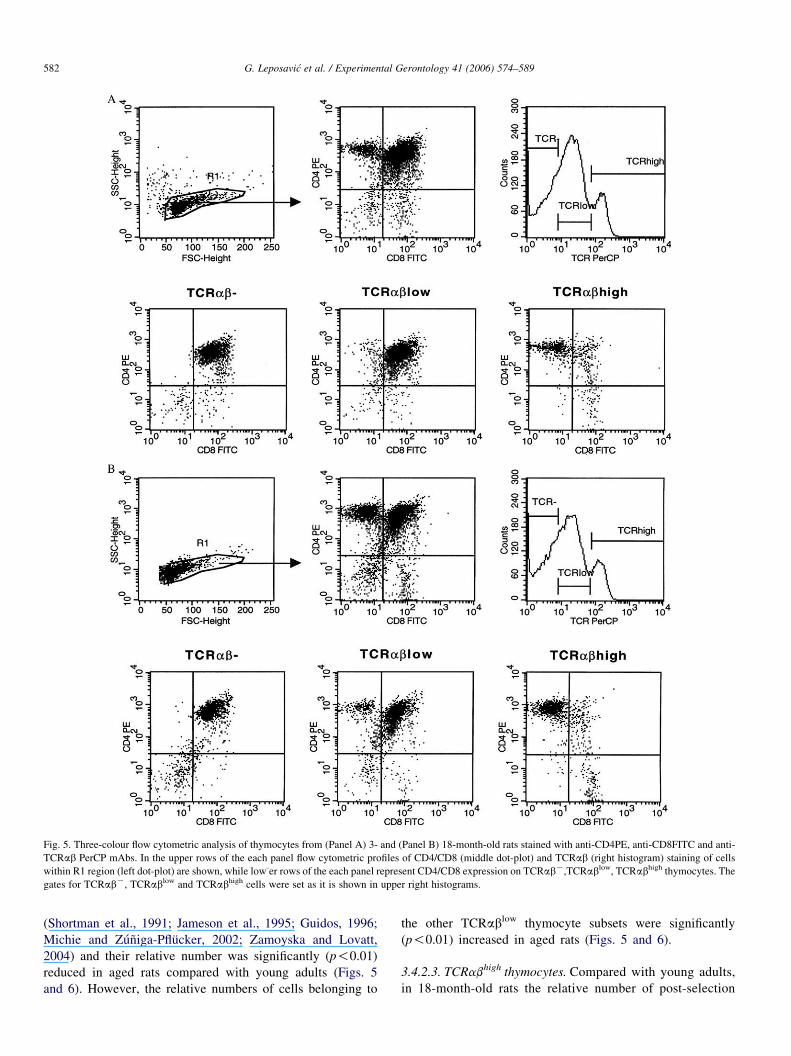
Fig. 5. Three-colour flow cytometric analysis of thymocytes from (Panel A) 3- and (Panel B) 18-month-old rats stained with anti-CD4PE, anti-CD8FITC and anti-
TCRab PerCP mAbs. In the upper rows of the each panel flow cytometric profiles of CD4/CD8 (middle dot-plot) and TCRab (right histogram) staining of cells
within R1 region (left dot-plot) are shown, while low-er rows of the each panel represent CD4/CD8 expression on TCRabK,TCRablow, TCRabhigh thymocytes. The
gates for TCRabK, TCRablow and TCRabhigh cells were set as it is shown in upper right histograms.
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589582
(Shortman et al., 1991; Jameson et al., 1995; Guidos, 1996;
Michie and Zuniga-Pflucker, 2002; Zamoyska and Lovatt,
2004) and their relative number was significantly (p!0.01)
reduced in aged rats compared with young adults (Figs. 5
and 6). However, the relative numbers of cells belonging to
the other TCRablow thymocyte subsets were significantly
(p!0.01) increased in aged rats (Figs. 5 and 6).
3.4.2.3. TCRabhigh thymocytes. Compared with young adults,
in 18-month-old rats the relative number of post-selection
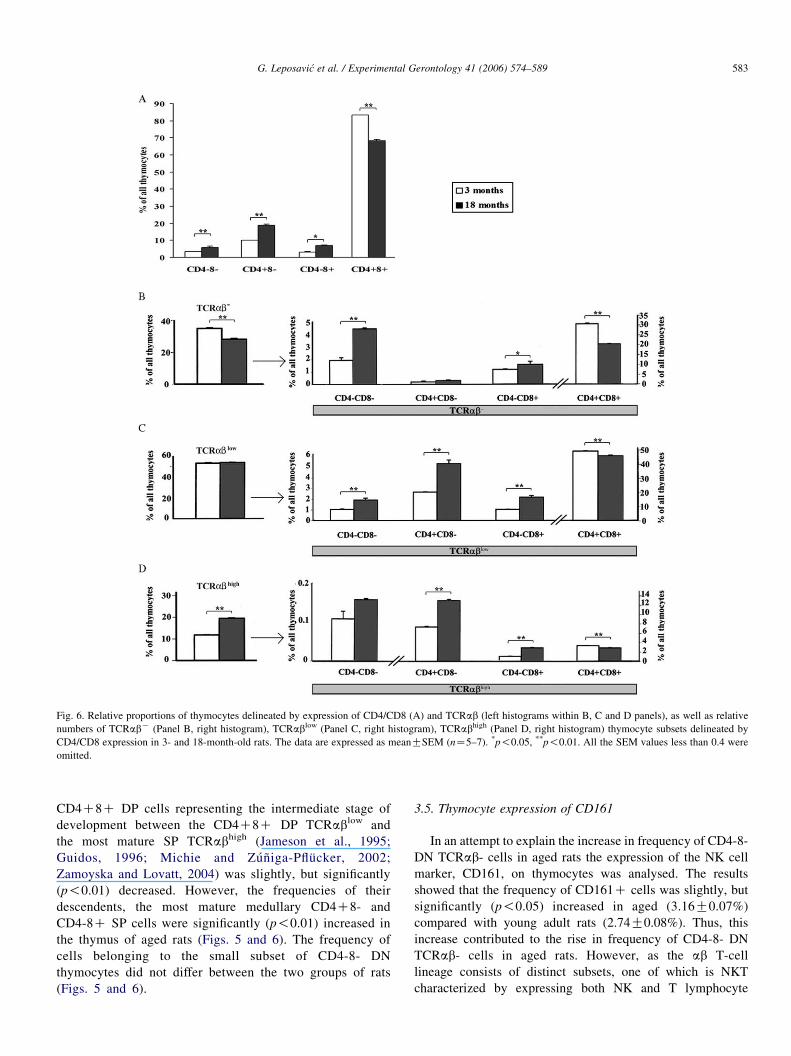
Fig. 6. Relative proportions of thymocytes delineated by expression of CD4/CD8 (A) and TCRab (left histograms within B, C and D panels), as well as relative
numbers of TCRabK (Panel B, right histogram), TCRablow (Panel C, right histogram), TCRabhigh (Panel D, right histogram) thymocyte subsets delineated by
CD4/CD8 expression in 3- and 18-month-old rats. The data are expressed as meanGSEM (nZ5–7). *p!0.05, **p!0.01. All the SEM values less than 0.4 were
omitted.
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589 583
CD4C8C DP cells representing the intermediate stage of
development between the CD4C8C DP TCRablow and
the most mature SP TCRabhigh (Jameson et al., 1995;
Guidos, 1996; Michie and Zuniga-Pflucker, 2002;
Zamoyska and Lovatt, 2004) was slightly, but significantly
(p!0.01) decreased. However, the frequencies of their
descendents, the most mature medullary CD4C8- and
CD4-8C SP cells were significantly (p!0.01) increased in
the thymus of aged rats (Figs. 5 and 6). The frequency of
cells belonging to the small subset of CD4-8- DN
thymocytes did not differ between the two groups of rats
(Figs. 5 and 6).
3.5. Thymocyte expression of CD161
In an attempt to explain the increase in frequency of CD4-8-
DN TCRab- cells in aged rats the expression of the NK cell
marker, CD161, on thymocytes was analysed. The results
showed that the frequency of CD161C cells was slightly, but
significantly (p!0.05) increased in aged (3.16G0.07%)
compared with young adult rats (2.74G0.08%). Thus, this
increase contributed to the rise in frequency of CD4-8- DN
TCRab- cells in aged rats. However, as the ab T-cell
lineage consists of distinct subsets, one of which is NKT
characterized by expressing both NK and T lymphocyte
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589584
markers (Godfrey et al., 2000; Pear et al., 2004), to answer
whether an increase in the relative number of NKT (CD161CTCRabC) cells contributed to the rise in CD161C cells in
aged rats, the relative number of these cells was quantified, as
well. However, there was no significant difference in the
relative proportion of NKT-cells between aged (2.08G0.29%)
and young adult (1.60G0.07%) rats.
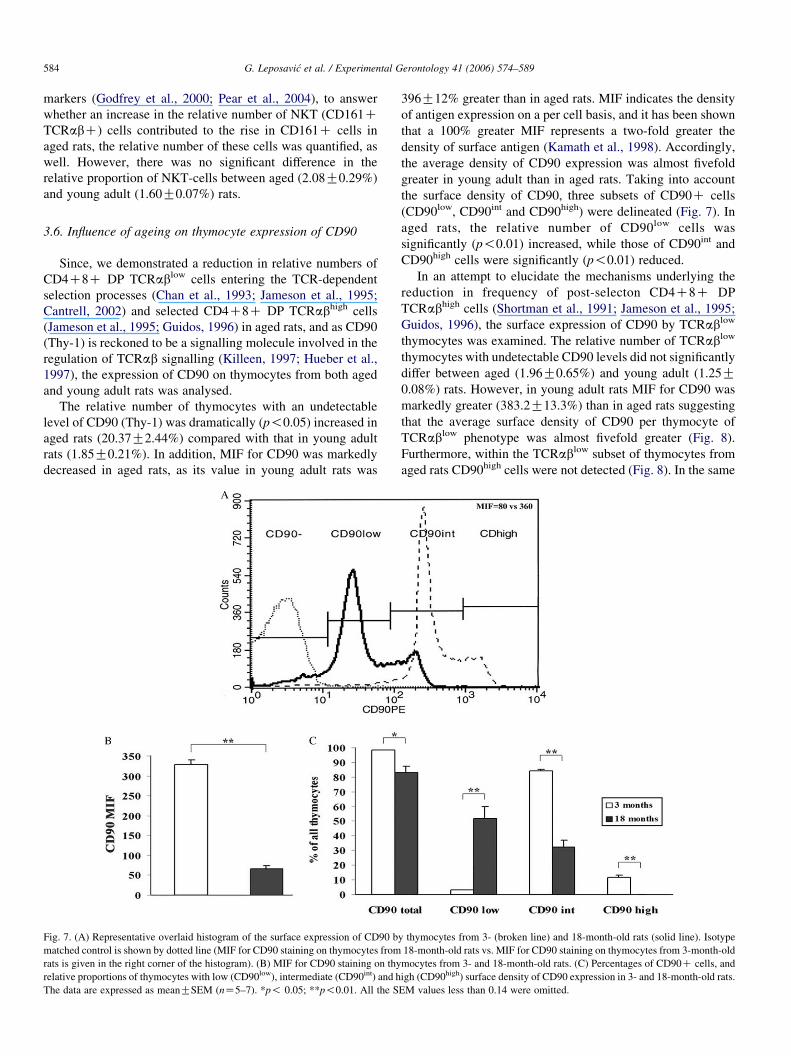
3.6. Influence of ageing on thymocyte expression of CD90
Since, we demonstrated a reduction in relative numbers of
CD4C8C DP TCRablow cells entering the TCR-dependent
selection processes (Chan et al., 1993; Jameson et al., 1995;
Cantrell, 2002) and selected CD4C8C DP TCRabhigh cells
(Jameson et al., 1995; Guidos, 1996) in aged rats, and as CD90
(Thy-1) is reckoned to be a signalling molecule involved in the
regulation of TCRab signalling (Killeen, 1997; Hueber et al.,
1997), the expression of CD90 on thymocytes from both aged
and young adult rats was analysed.
The relative number of thymocytes with an undetectable
level of CD90 (Thy-1) was dramatically (p!0.05) increased in
aged rats (20.37G2.44%) compared with that in young adult
rats (1.85G0.21%). In addition, MIF for CD90 was markedly
decreased in aged rats, as its value in young adult rats was
Fig. 7. (A) Representative overlaid histogram of the surface expression of CD90 b
matched control is shown by dotted line (MIF for CD90 staining on thymocytes from
rats is given in the right corner of the histogram). (B) MIF for CD90 staining on thy
relative proportions of thymocytes with low (CD90low), intermediate (CD90int) and h
The data are expressed as meanGSEM (nZ5–7). *p! 0.05; **p!0.01. All the S
396G12% greater than in aged rats. MIF indicates the density
of antigen expression on a per cell basis, and it has been shown
that a 100% greater MIF represents a two-fold greater the
density of surface antigen (Kamath et al., 1998). Accordingly,
the average density of CD90 expression was almost fivefold
greater in young adult than in aged rats. Taking into account
the surface density of CD90, three subsets of CD90C cells
(CD90low, CD90int and CD90high) were delineated (Fig. 7). In
aged rats, the relative number of CD90low cells was
significantly (p!0.01) increased, while those of CD90int and
CD90high cells were significantly (p!0.01) reduced.
In an attempt to elucidate the mechanisms underlying the
reduction in frequency of post-selecton CD4C8C DP
TCRabhigh cells (Shortman et al., 1991; Jameson et al., 1995;
Guidos, 1996), the surface expression of CD90 by TCRablow
thymocytes was examined. The relative number of TCRablow
thymocytes with undetectable CD90 levels did not significantly
differ between aged (1.96G0.65%) and young adult (1.25G0.08%) rats. However, in young adult rats MIF for CD90 was
markedly greater (383.2G13.3%) than in aged rats suggesting
that the average surface density of CD90 per thymocyte of
TCRablow phenotype was almost fivefold greater (Fig. 8).
Furthermore, within the TCRablow subset of thymocytes from
aged rats CD90high cells were not detected (Fig. 8). In the same
y thymocytes from 3- (broken line) and 18-month-old rats (solid line). Isotype
18-month-old rats vs. MIF for CD90 staining on thymocytes from 3-month-old
mocytes from 3- and 18-month-old rats. (C) Percentages of CD90C cells, and
igh (CD90high) surface density of CD90 expression in 3- and 18-month-old rats.
EM values less than 0.14 were omitted.
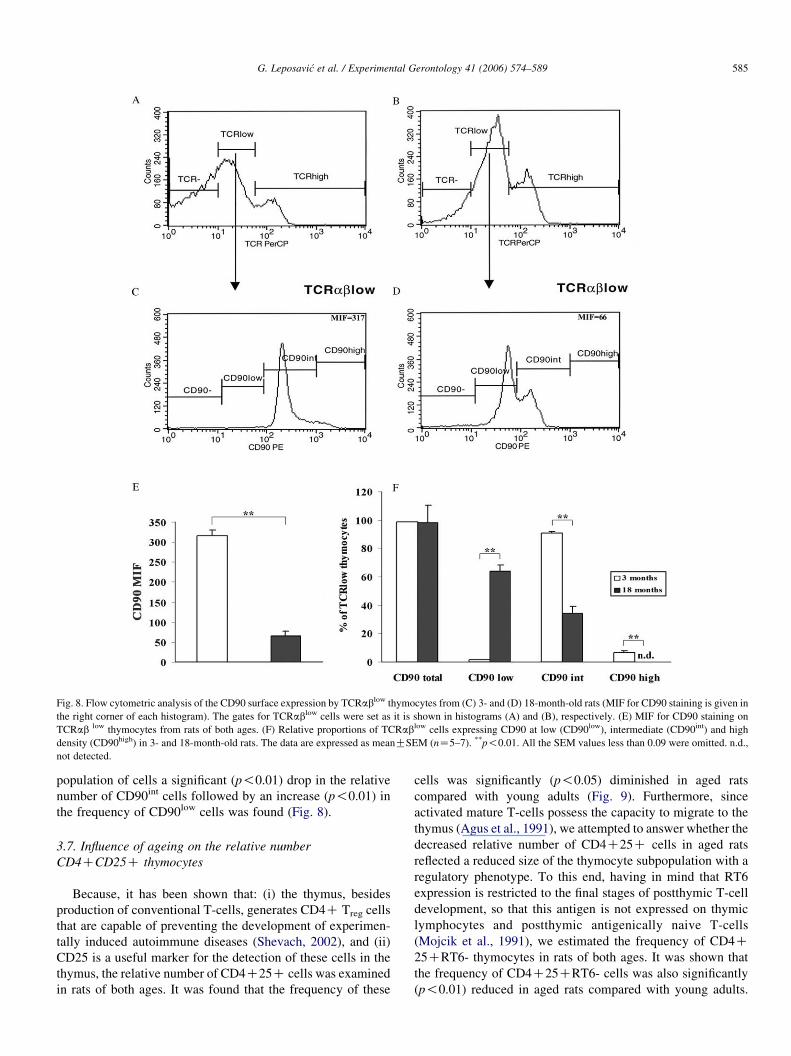
Fig. 8. Flow cytometric analysis of the CD90 surface expression by TCRablow thymocytes from (C) 3- and (D) 18-month-old rats (MIF for CD90 staining is given in
the right corner of each histogram). The gates for TCRablow cells were set as it is shown in histograms (A) and (B), respectively. (E) MIF for CD90 staining on
TCRab low thymocytes from rats of both ages. (F) Relative proportions of TCRablow cells expressing CD90 at low (CD90low), intermediate (CD90int) and high
density (CD90high) in 3- and 18-month-old rats. The data are expressed as meanGSEM (nZ5–7). **p!0.01. All the SEM values less than 0.09 were omitted. n.d.,
not detected.
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589 585
population of cells a significant (p!0.01) drop in the relative
number of CD90int cells followed by an increase (p!0.01) in
the frequency of CD90low cells was found (Fig. 8).
3.7. Influence of ageing on the relative number
CD4CCD25C thymocytes
Because, it has been shown that: (i) the thymus, besides
production of conventional T-cells, generates CD4C Treg cells
that are capable of preventing the development of experimen-
tally induced autoimmune diseases (Shevach, 2002), and (ii)
CD25 is a useful marker for the detection of these cells in the
thymus, the relative number of CD4C25C cells was examined
in rats of both ages. It was found that the frequency of these
cells was significantly (p!0.05) diminished in aged rats
compared with young adults (Fig. 9). Furthermore, since
activated mature T-cells possess the capacity to migrate to the
thymus (Agus et al., 1991), we attempted to answer whether the
decreased relative number of CD4C25C cells in aged rats
reflected a reduced size of the thymocyte subpopulation with a
regulatory phenotype. To this end, having in mind that RT6
expression is restricted to the final stages of postthymic T-cell
development, so that this antigen is not expressed on thymic
lymphocytes and postthymic antigenically naive T-cells
(Mojcik et al., 1991), we estimated the frequency of CD4C25CRT6- thymocytes in rats of both ages. It was shown that
the frequency of CD4C25CRT6- cells was also significantly
(p!0.01) reduced in aged rats compared with young adults.

Fig. 9. Flow cytometric analysis of CD4/CD25 expression by (panel A) all and (panel B) RT6K thymocytes from (left dot-plots) 3- and (right dot-plots) 18-month-
old rats. Upper single parameter histograms in panel B represent RT6 expression in (left) 3- and in (right) 18-month-old rats. Single-parameter overlaid histograms
represent CD4 expression by (Panel A) all and (Panel B) CD25CRT6K thymocytes from 3- (white histogram) and 18-month-old rats (gray histogram). Relative
proportions of the all CD4CCD25C and CD4CCD25CRT6K thymocytes in rats of both ages are shown in right histograms in Panel A and Panel B, respectively.
The data are expressed as meanGSEM (nZ5–7). *p!0.05; **p!0.01.
G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589586
This finding speaks in favour of a reduction in the relative
contribution of the cells expressing regulatory CD4C25Cphenotype in aged rats.
4. Discussion
This study has demonstrated a sharp drop in the cellularity
and total volume of lymphoepithelial thymic tissue in 18-
month-old Wistar rats compared with their 3-month-old
counterparts. However, in spite of this the thymic weight did
not significantly differ between these two groups of rats due to
an age-related increase in the volume of nonlymphoid, i.e.
adipose and connective tissue. Contrary to this finding, an age-
related reduction in the overall thymus weight was reported in
many strains of rats and mice (Fabris et al., 1988; Li et al.,
1992; Bodey et al., 1997; Leposavic et al., 2002). This
discrepancy may be related to a strong genetic contribution
influencing age-related thymic changes, for example, Buffalo
strain rats do not experience thymic involution (Hirokawa
et al., 1994). It has been suggested that: (i) reduction in the
volume of thymic lymphoepithelial tissue produces a radical
decrease in the number of recent thymic emigrants and (ii) the
functional capacity of the residual true thymic tissue correlates
with its anatomical measurements (Pawelec et al., 1999). The
drop in thymus cellularity with age is fully consistent with the
findings in the cultures of thymocytes from aged rats showing:
(i) the accelerated apoptotic elimination of thymocytes and (ii)
the reduced frequency of cells incorporating BrdU spon-
taneously or in a response to ConA stimulation. The reduced
frequency of apoptotic cells in response to Dex in thymocyte
cultures from aged rats compared with that in thymocyte
cultures from young adults may be explained by a significant
reduction in the relative numbers of DP cells, which are
believed to be the most sensitive to glucocorticoid-induced
apoptosis (Nieto et al., 1992). The present data on apoptotic
and proliferative characteristics of thymocytes from aged rats
in culture are in accordance with those showing that
thymocytes from aged mice and rats exhibit an increase in
the expression of cell death genes, i.e. p53, bax and caspase 3
(Kapasi and Singhal, 1999) and a significant decrease in
spontaneous and mitogen-induced proliferation (Kaplan and
Garvey, 1981; Yu et al., 1997; Globerson and Effros, 2000; Hsu
et al., 2002, 2005; Tian et al., 2003), respectively.
The study has also shown substantial changes in the
distribution of thymocyte subsets delineated by expression of
CD4/8/TCRab in aged rats compared with young adults
suggesting age-associated alterations in thymocyte develop-
ment. In accordance with previous work (Capri et al., 2000) an
increased frequency of the least mature CD4-8- DN TCRabK
cells was found in aged rats. The relative number of these cells
has been also reported to increase in the thymus of aged mice
(Aspinall, 1997; Thoman, 1997; Yehuda et al., 1998; Hsu et al.,
2002). The present findings, similarly to those for mice
(Ales-Martinez et al., 1988), indicate that the increase in the
frequency of CD4-8- DN TCRabK cells in aged rats partly
reflected an overrepresentaion of NK cells in the thymus of
aged animals compared with young adults. Therefore, a
possible contribution of some other mechanisms such as:

G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589 587
(i) an increased entry of precursor cells; (ii) reduced apoptotic
loss of these cells and (iii) their increased proliferation, to the
increase in frequency of CD4-8- DN TCRabK cells in aged
rats should be also taken into consideration. There are no data
to support the first option. On the contrary, it appears that there
is no significant age-dependent loss in the number of bone-
marrow-resident pre-T precursor cells (Sharp et al., 1990), and
it has been suggested that ‘seeding’ of the aged thymic
reticulum by blood–borne precursors is similar to that of the
young thymus (Donskoy and Goldschneider, 1992; Thoman,
1995, 1997; Foss et al., 2001; Shanker, 2004). There are also no
data to support the contribution of either reduced apoptotic loss
or increased cell proliferation to the age-associated increase in
the frequency of CD4-8- DN TCRabK cells. The increase in
frequency of these cells in the thymus of aged mice has been
ascribed to developmental blocks that occur with differentia-
tive transition within the CD4-8- DN TCRabK developmental
stage itself and to decreased progression from this to the next
CD4C8C DP stage (Aspinall, 1997; Thoman, 1997; Yehuda
et al., 1998; Capri et al., 2000; Hsu et al., 2002). The hereby
reported reduction in the frequency of BrdUC cells within
CD3K subsets of thymocytes from aged rats in culture seems to
be consistent with the previous assumption indicating that the
transition of thymocytes through differentiation steps within
the DN stage becomes limited leading to a restriction in the
number of cells capable of proliferation and further transition
to the DP stage. In keeping with this assumption is the
reduction in the relative numbers of CD4-8- DN TCRabK
descendent CD4C8C DP TCRabK cells in aged rats.
However, a reduced frequency of CD4C8C DP TCRablow
cells in aged rats, in the light of a decreased relative number of
their immediate CD4C8C DP TCRabK precursors, on the
one hand, and a reduced relative number of their CD4C8C DP
TCRabhigh descendants, on the other hand, may suggest,
not only reduced thymocyte transition from the DN to DP
differentiational stage, but also less efficient positive selection
and/or overexaggerated negative selection. The dramatically
reduced surface density of Thy-1 on TCRablow thymocytes
(encompassing mainly CD4C8C DP cells) from aged rats
found here favours this assumption, since it has been
demonstrated that thymocytes from ThyK/K mice exhibit
inappropriate negative selection due to a lack of Thy-1-
mediated negative regulation of TCRab signalling and
activation thresholds during thymocyte differentiation (Hueber
et al., 1997). Several other findings in the present study also
favour this hypothesis. Firstly, the decreased frequency of
BrdUC cells within the CD3low subset in thymocyte cultures
from aged rats in the presence of ConA, which is shown to
induce positive selection and thereby survival and further
maturation of DP thymocytes (Lovatt et al., 2000; Pongracz
et al., 2003), seems to be consistent with the previous
assumption. Secondly, the increased frequency of apoptotic
cells in thymocyte cultures from aged rats compared with those
from young adult rats is also in keeping with the same
hypothesis. Finally, it has been reported that the relative
number of Thy 1.1C thymocytes is diminished in 24-month-
old female Wistar–Furth rats but could be augmented after
implantation of GH3 pituitary adenoma cells (Kelley et al.,
1986).
The increased frequency of SP (CD4-8C and CD4C8-)
TCRabhigh thymocytes in aged rats, together with the reduced
frequency of their CD4C8C DP TCRabhigh predecessors,
may indicate more efficient differentiation of the positively
selected cells. In favour of this option are data indicating that
when there is a developmental block in T-cell development,
reparatory mechanisms in the thymus increase the efficiency of
generation of mature SP cells facilitating events beyond the
block (Almeida et al., 2001). Furthermore, it may also suggest
an increase in pre-migrational proliferation of these cells and/
or increased re-entry of T-cells from the periphery. Having in
mind that, differently from immature T-cells, ConA causes
activation and proliferation of mature T-cells, the first option is
supported by data showing that ConA evoked a more
pronounced increase in the frequency of BrdUC cells within
the mature CD3high subpopulation in thymocyte cultures from
aged rats than in those from young adults. The increased
frequency of cells showing the TCRabCCD90- mature
peripheral phenotype (Williams, 1989), in the thymuses from
aged rats (data not shown) is fully consistent with the second
option. Furthermore, it should be pointed out that the increased
frequency of mature SP TCRabhigh cells in the thymus may
reflect decreased export of these cells into the periphery.
However, other findings suggesting that the ‘efficiency’ of
thymus cell export increases with low numbers of thymus cells,
but insufficiently to compensate for the reduced production of
mature thymus cells, speak against the latter option (Almeida
et al., 2001).
Finally, providing that, in aged rats, there is an increased
frequency of CD4C re-entrants in the thymus as has been
suggested for mice (Mehr et al., 1996), collectively taken, the
present findings on alterations in thymocyte subsets in aged rats
(increased frequencies of the least mature CD4-8-TCR- cells
and the most mature CD4C8-TCRabhigh cells, but decreased
frequency of DP cells) seem to fit the framework of a
previously stated hypothesis that T-cell maturation is subject to
intrathymic feedback regulation. Namely, the studies of
T-lymphocyte development in mouse fetal thymic stroma
colonized with immature thymocytes and CD4C cells from
young and old donors have indicated that thymocyte
development is subject to regulation through two CD4C cell
mediated feedback loops. It has been hypothesized that mature
CD4C cells, whether newly generated in the thymus or re-
entrants from the periphery, exert a negative feedback on the
DN to DP transition and DP subset growth, and a positive
feedback on the DP to CD4 SP transition (Mehr et al., 1996,
1997).
Moreover, a more pronounced increase in the frequency of
mature CD4-8C SP than in the frequency of mature CD4C8-
SP seems to be in keeping with the observation indicating that
the CD8 T-cell subset is less sensitive than the CD4 T- cell
subset to age-induced alterations (Pawelec et al., 1999).
The present study also demonstrated that the relative
number of CD4C25C cells was significantly diminished in
the thymus of aged rats reflecting, at least partly, a reduced

G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589588
frequency of intrathymically developing Treg cells. (Fontenot
and Rudensky, 2005). Taking collectively the data suggesting
that Thy-1- mediated down-regulation of TCR signalling is
impaired in aged rats and those indicating that naturally arising
Treg cell lineage commitment is induced in response to self-
reactive TCRs with an avidity range for self peptide—MHC
somewhere between that required for positive and negative
selection (Fontenot and Rudensky, 2005), a diminished Treg
cell lineage commitment, and therefore a reduced thymic
production of Treg cells in aged rats may be expected. This
assumption is fully consistent with the increased incidence of
autoimmune disorders in old age (Wick et al., 2000; Shanker,
2004).
The present study confirmed that early differentiational
steps within the CD4-8- DN stage of thymocyte development
are affected in rats by ageing, and demonstrated alterations in
TCRab-dependent stages of T-cell development that, at least
partly, may be related to impaired expression of Thy-1
adhesion and signalling molecules on TCRablow thymocytes
entering selection processes. Furthermore, it suggests that
impaired positive selection and/or overexaggerated negative
selection leads to a reduced frequency of positively selected
CD4C8C DP TCRabhigh cells in aged rats, and that these
changes are followed by more efficient differentiation of
CD4C8C DP TCRabhigh cells into mature SP cells exhibiting
increased pre-migrational proliferation. Moreover, the study
indicates that generation of CD4C Treg cells is also impaired in
the thymus of aged rats, thus providing an explanation for the
increased frequency of autoimmune diseases in aged animals.
Acknowledgements
This work is support by the grants 1239 and 145049B from
Ministry of Science and Environmental Protection of the
Republic of Serbia.
References
Agus, B.D., Surh, C.D., Sprent, J., 1991. Reentry of T cells to the adult thymus
is restricted to activated T cells. J. Exp. Med. 173, 1039–1046.
Ales-Martinez, J.E., Alvarez-Mon, M., Merino, F., Bonilla, F., Martinez-A., C.,
Durantez, A., De la Hera, A., 1988. Decreased TCR-CD3CT cell numbers
in healthy aged humans. Evidence that T cell defects are masked by a
reciprocal increase of TCR-CD3-CD2C natural killer cells. Eur.
J. Immunol. 18, 1827–1830.
Almeida, A.R.M., Borghans, J.A.M., Freitas, A.A., 2001. T cell homeostasis:
thymus regeneration and peripheral T cell restoration in mice with a
reduced fraction of competent precursors. J. Exp. Med. 194, 591–599.
Aspinall, R., 1997. Age-associated thymic atrophy in the mouse is due to a
deficiency affecting rearrangement of the TCR during intrathymic T cell
development. J. Immunol. 158, 3037–3045.
Aspinall, R., 2000. Longevity and immune response. Biogerontology 1,
273–278.
Bodey, B., Bodey Jr., B., Siegel, S.E., Kaiser, H.E., 1997. Involution of the
mammalian thymus, one of the heading regulators of ageing. In vivo 11,
421–440.
Brill, S., Ben-Menahem, D., Kukulansky, T., Tal, E., Globerson, A., 1990. Do
peripheral T-lymphocytes reflect splenic T-lymphocyte functions in
individual young and old mice? Aging: Immunol. Infect. Dis. 2, 221–228.
Brown, D.G., Sun, X.-M., Cohen, G., 1993. Dexamethasone-induced apoptosis
involves cleavage of DNA to large fragments prior to internucleosomal
fragmentation. J. Biol. Chem. 268, 3037–3039.
Cantrell, D.A., 2002. Transgenic analysis of thymocyte signal transduction.
Nat. Rev. Immunol. 2, 20–27.
Capri, M., Quaglino, D., Verzella, G., Monti, D., Bonafe, M., Cossarizza, A.,
Troiano, L., Zecca, L., Pasquali-Ronchetti, I., Franceschi, C., 2000. A
cytofluorimetric study of T lymphocyte subsets in rat lymphoid tissues
(thymus, lymph nodes) and peripheral blood: a continuous remodelling
during the first year of life. Exp. Gerontol. 35, 613–625.
Chan, S.H., Cosgrove, D., Waltzinger, C., Benoist, C., Mathis, D., 1993.
Another view of the selective model of thymocyte selection. Cell 73,
225–236.
Cohen, J.J., 1991. Programmed cell death in murine myeloid leukemia cells.
Photochem. Photobiol. 72, 114–120.
Consolini, R., Legitimo, A., Calleri, A., 2000. Distribution of age-related
thymulin titres in normal subjects through the course of life. Clin. Exp.
Immunol. 121, 444–447.
Donskoy, E., Goldschneider, I., 1992. Thymocytopoiesis is maintained by
blood-borne precursors throughout postnatal life. A study in parabiotic
mice. J. Immunol. 148, 1604–1612.
Fabris, N., Mocchegiani, M., Muzzoli, M., Provinciali, M., 1988. Immune-
neuroendocrine interactions during aging. PNEI 1, 5–20.
Fabris, N., Mocchegiani, M., Provinciali, M., 1997. Plasticity of neuroendo-
crine-thymus interactions during aging. Exp. Gerontol. 32, 415–429.
Fontenot, J.D., Rudensky, A.Y., 2005. A well adapted regulatory contrivance:
regulatory T cell development and the forkhead family transcription factor
Foxp3. Nat. Immunol. 6, 331–337.
Foss, D.L., Donskoy, E., Goldschneider, I., 2001. The importation of
hematogenous precursors by the thymus is a gated phenomenon in normal
adult mice. J. Exp. Med. 193, 365–374.
George, A.J.T., Ritter, M.A., 1996. Thymic involution with ageing:
obsolescence or good housekeeping? Immunol. Today 17, 267–269.
Globerson, A., Effros, R.B., 2000. Ageing of lymphocytes and lymphocytes in
the aged. Immunol. Today 21, 515–521.
Godfrey, D.I., Hammond, K.J., Poulton, L.D., Smyth, M.J., Baxter, A.G.,
2000. NKT cells: facts, functions and fallacies. Immunol. Today 21,
573–583.
Grubeck-Loebenstein, B., 1997. Changes in the aging immune system.
Biologicals 25, 205–208.
Guidos, C.J., 1996. Positive selection of CD4C and CD8C cells. Curr. Opin.
Immunol. 8, 225–232.
Hirokawa, K., Kubo, S., Utsuyama, M., Kurashima, C., Sado, T., 1986. Age-
related changes in the potential of bone marrow cells to repopulate the
thymus and splenic T cells in mice. Cell. Immunol. 100, 443–451.
Hirokawa, K., Utsuyama, M., Kasai, M., Kurashima, C., Ishijima, S.,
Zeng, Y.X., 1994. Understanding the mechanism of age-changes of
thymic function to promote T cell differentiation. Immunol. Lett. 40,
267–277.
Hirokawa, K., Utsuyama, M., Kobayashi, S., 1997. Hypothalamic control of
development and ageing of the thymus. Mech. Ageing Dev. 100,
177–185.
Hsu, H.-C., Mountz, J.D., Williams, R.W., Shelton, B.J., Yang, P.-A.,
Matsuki, Y., Xu, X., Dodd, C.H., Li, L., Geiger, H., Zhang, H.-G.,
Zant, G.V., 2002. Age-related change in thymic T-cell development is
associated with genetic loci on mouse chromosomes 1, 3, and 11. Mech.
Ageing Dev. 123, 1145–1158.
Hsu, H.-C., Li, L., Zhang, H.-G., Mountz, J.D., 2005. Genetic regulation of
thymic involution. Mech. Ageing Dev. 126, 87–97.
Hueber, A.O., Bernard, A.M., Battari, C.L., Marguet, D., Massol, P., Foa, C.,
Brun, N., Garsia, S., Steward, C., Pierres, M., He, H.T., 1997. Thymocytes
in Thy-1K/K mice show augmented TCR signaling and impared
differentiation. Curr. Biol. 7, 705–708.
Hunig, T., Wallny, H.J., Hartley, J.K., Lawetzky, A., Tiefenthaler, G., 1989. A
monoclonal antibody to a constant determinant of the rat T cell antigen
receptor that induces T cell activation. J. Exp. Med. 169, 73–86.
Jameson, S.C., Hogquist, K.A., Bevan, M.J., 1995. Positive selection of
thymocytes. Annu. Rev. Immunol. 13, 93–126.

G. Leposavic et al. / Experimental Gerontology 41 (2006) 574–589 589
Kamath, A.B., Nagarkatti, P.S., Nagarkatti, M., 1998. Characterization of
phenotypic alterations induced by 2,3,7,8-tetrachlorodibenzo-p-dioxin on
thymocytes in vivo and its effect on apoptosis. Toxicol. Appl. Pharmacol.
150, 117–124.
Kapasi, A.A., Singhal, P.C., 1999. Aging splenocyte and thymocyte apoptosis
is associated with enhanced expression of p53, Bax, and Caspase-3. Mol.
Cell. Biol. Res. Commun. 1, 78–81.
Kaplan, P.J., Garvey, J.S., 1981. Age-related changes in responsiveness of
various rat tissue lymphocytes to mitogens. Immunol. Lett. 3, 357–363.
Kelley, K.W., Brief, S., Westly, H.J., Novakofski, J., Bechtel, P.J., Simon, J.,
Walker, E.B., 1986. GH3 pituitary adenoma cells can reverse thymic aging
in rats. Proc. Natl Acad. Sci. USA 83, 5663–5667.
Killeen, N., 1997. Thy-1-hiding in full view. Curr. Biol. 7, R774–R777.
Lau, L., Spain, L.M., 2000. Altered ageing-related thymic involution in T cell
receptor transgenic, MHC-deficient, and CD4-deficient mice. Mech.
Ageing Dev. 114, 101–121.
Leposavic, G., Pejcic-Karapetrovic, B., Kosec, D., 2002. Alterations in
thymopoiesis in intact and peripubertally orchidectomized adult rats of
different age. Mech. Ageing Dev. 123, 401–411.
Leposavic, G., Pekic, S., Kosec, D., 2005. Gonadotropin-releasing hormone
agonist administration affects the thymopoiesis in adult female rats
independently on gonadal hormone production. Am. J. Reprod. Immunol.
53, 1–13.
Li, Y.M., Brunke, D.L., Dantzer, R., Kelley, K., 1992. Pituitary epithelial cell
implants reverse the accumulation of CD4–CD8-lymphocytes in thymus
glands of aged rats. Endocrinology 130, 2703–2709.
Lovatt, M., Yang, T.H., Stauss, H.J., Fisher, A.G., Merkenschlager, M., 2000.
Different doses of agonistic ligand drive the maturation of functional CD4
and CD8 T cells from immature precursors. Eur. J. Immunol. 30, 371–381.
Mackall, C.L., Punt, J.A., Morgan, P., Farr, A.G., Gress, R.E., 1998. Thymic
function in young/old chimeras: substantial thymic T cell regenerative
capacity despite irreversible age-associated thymic involution. Eur.
J. Immunol. 28, 1886–1893.
Madden, K.S., Rajan, S.T., Bellinger, D.B., Felten, S.Y., Felten, D.L., 1997.
Age-associated alterations in sympathetic neural interactions within the
immune system. Dev. Comp. Immunol. 21, 479–486.
Mehr, R., Perelson, A.S., Fridkis-Hareli, M., Globerson, A., 1996. Feedback
regulation of T cell development: manifestations in aging. Mech. Ageing
Dev. 91, 195–210.
Mehr, R., Perelson, A.S., Fridkis-Hareli, M., Globerson, A., 1997. Regulatory
feedback pathways in the thymus. Immunol. Today 18, 581–585.
Michie, A.M., Zuniga-Pflucker, J.C., 2002. Regulation of thymocyte
differentiation: pre-TCR signals and b-selection. Semin. Immunol. 14,
311–323.
Mojcik, C.F., Greiner, D.L., Goldschneider, I., 1991. Characterization of RT6-
bearing rat lymphocytes. Developmental relationships of RT6K and
RT6CT cell. Dev. Immunol. 1, 191–201.
Mower Jr., D.A., Peckham, D.W., Illera, V.A., Fishbaugh, J.K., Stunz, L.L.,
Ashman, R.F., 1994. Decreased membrane phospholipid packing and
decreased size precede DNA cleavage in mature mouse B cell apoptosis.
J. Immunol. 152, 4832–4842.
Nieto, M.A., Gonzalez, A., Gambon, F., Dıaz-Espada, F., Lopez-Rivas, A.,
1992. Apoptosis in human thymocytes after treatment with glucocorticoids.
Clin. Exp. Immunol. 88, 341–344.
Okasha, S.A., Ryu, S., Do, Y., McKallip, R.J., Nagarkatti, M., Nagarkatti, P.S.,
2001. Evidence for estradiol-induced apoptosis and dysregulated T cell
maturation in the thymus. Toxicology 163, 49–62.
Pawelec, G., Effros, R.B., Caruso, C., Remarque, E., Barnett, Y., Solana, R.,
1999. T cells and ageing. Front. Biosci. 4, d216–d269.
Pear, W.S., Tu, L., Stein, P.L., 2004. Lineage choices in the developing
thymus: choosing the T and NKT pathways. Curr. Opin. Immunol. 16,
167–173.
Plecas-Solarovic, B., Lalic, L.J., Leposavic, G., 2004. Age-dependent
morphometrical changes in the thymus of male propranol-treated rats.
Ann. Anat. 186, 141–147.
Pongracz, J., Parnell, S., Anderson, G., Jaffrezou, J.-P., Jenkinson, E., 2003.
Con A activates an Akt/PKB dependent survival mechanism to modulate
TCR induced cell death in double positive thymocytes. Mol. Immunol. 39,
1013–1023.
Reid, S., Cross, R., Snow, E.R., 1996. Combined Hoechst 33342 and
merocyanine 540 staining to examine murine B cell cycle stage, viability
and apoptosis. J. Immunol. Methods 192, 43–54.
Ritter, M.A., Boyd, R.L., 1993. Development in the thymus: it takes two to
tango. Immunol. Today 14, 462–469.
Romanyukha, A.A., Alexei, A., Yashin, A.I., 2003. Age related changes in
population of peripheral T cells: towards a model of immunoscenescence.
Mech. Ageing Dev. 124, 433–443.
Seddon, B., Mason, D., 2000. The third function of the thymus. Immunol.
Today 21, 95–99.
Shanker, A., 2004. Is thymus redundant after adulthood. Immunol. Lett. 91,
79–86.
Sharp, A., Kukulansky, T., Globerson, A., 1990. In vitro analysis of age-related
changes in the developmental potential of bone marrow thymocyte
progenitors. Eur. J. Immunol. 12, 2541–2546.
Shevach, E.M., 2002. CD4CCD25C suppressor T cells: more questions than
answers. Nat. Rev. Immunol. 2, 389–400.
Shortman, K., Vremec, D., Egerton, M., 1991. The kinetics of T cell antigen
receptor expression by subgroups of CD4C8C thymocytes: delineation of
CD4C8C32C thymocytes as post-selection intermediates leading to
mature T cells. J. Exp. Med. 173, 323–332.
Thoman, M.L., 1995. The pattern of T-lymphocyte differentiation is altered
during thymic involution. Mech. Ageing Dev. 58, 167–177.
Thoman, M.L., 1997. Early steps in T cell development are affected by ageing.
Cell. Immunol. 178, 117–123.
Tian, Y.-M., Tian, H.-J., Zhang, G.-Y., Dai, Y.-R., 2003. Effects of ginkgo
biloba extracts (EGb 761) on hydroxyl radical—induced thymocyte
apoptosis and on age-related thymic atrophy and peripheral immune
disfunctions in mice. Mech. Ageing Dev. 124, 977–983.
Tsuchida, M., Konishi, M., Jojima, K., Naito, K., Fujikura, Y., Fukumoto, T.,
1994. Analysis of cell surface antigens on glucocorticoid-treated rat
thymocytes with monoclonal antibodies. Immunol. Lett. 39, 209–217.
Vermes, I., Haanen, C., Steffens-Nakken, H., Reutelingsperger, C., 1995. A
novel assay for apoptosis. Flow cytometric detection of phosphatidylserine
expression on early apoptotic cells using fluorescein labelled Annexin V.
J. Immunol. Methods 184, 39–51.
Wajeman-Chao, S.A., Lancaster, S.A., Graf Jr., L.H., Chambers, D.A., 1998.
Mechanism of catecholamine-mediated destabilization of messenger RNA
encoding Thy-1 protein in T-lineage cells. J. Immunol. 161, 4825–4833.
Weibel, E.R., 1979. Stereological Methods: Practical Methods for Biological
Morphometry, vol. 1. Academic Press, London.
Wick, G., Jansen-Durr, P., Berger, P., Blasko, I., Grubeck-Loebenstein, B.,
2000. Disease of ageing. Vaccine 18, 1567–1583.
Williams, A.F., 1989. The structure of Thy-1 antigen. In: Reif, A.E.,
Schlesinger, M. (Eds.), Cell Surface Antigen Thy-1. Marcel Dekker, New
York, pp. 49–69.
Yehuda, A.B., Friedman, G., Wirtheim, E., Abel, L., Globerson, A., 1998.
Checkpoints in thymocytopoiesis in ageing: expression of recombination
activating genes RAG-1 and RAG-2. Mech. Ageing Dev. 192, 239–247.
Yu, S., Abel, L., Globerson, A., 1997. Thymocyte progenitors and T cell
development in ageing. Mech. Ageing Dev. 94, 103–111.
Zamoyska, R., Lovatt, M., 2004. Signalling in T-lymphocyte development:
integration of signalling pathways is the key. Curr. Opin. Immunol. 16,
191–196.
Related Documents





![Thymoglobulin (globuline anti-thymocytes [Lapin])](https://static.cupdf.com/doc/110x72/586dfe2c1a28ab114f8babb9/thymoglobulin-globuline-anti-thymocytes-lapin.jpg)


![Thymoglobulin (anti-thymocyte globulin [rabbit]) - Sanofiproducts.sanofi.ca/en/thymoglobulin.pdf · Thymoglobulin® (Anti-thymocyte Globulin [Rabbit]) ... (ATG) products, as protein](https://static.cupdf.com/doc/110x72/5aa587a17f8b9ab4788d4753/thymoglobulin-anti-thymocyte-globulin-rabbit-anti-thymocyte-globulin-rabbit.jpg)


