www.landesbioscience.com Human Vaccines & Immunotherapeutics 1 Human Vaccines & Immunotherapeutics 8:6, 1-12; June 2012; © 2012 Landes Bioscience RESEARCH PAPER RESEARCH PAPER *Correspondence to: Jordan L. Plieskatt; Email: [email protected] Submitted: 12/01/11; Revised: 02/10/12; Accepted: 02/16/12 http://dx.doi.org/10.4161/hv.19726 Introduction The neglected tropical diseases (NTDs) are a group of 17 major chronic and disabling infections affecting more than 1 billion people worldwide, primarily those living in rural areas, urban slums or conflict zones. 1,2 Human hookworm infection is one of the world’s most common and debilitating NTDs responsible for an estimated 600 million cases in the world’s poorest countries; most of these infections are caused by Necator americanus. 3-5 The clinical signature of hookworm infection is anemia and protein malnutrition, 6 which results from hookworm associated blood loss, especially in children and pregnant women. 7,8 Blood loss from hookworm is responsible for a disease burden equivalent to up to one-half of the disability-adjusted life years lost that result from better known devastating infections such as malaria, 9 in addition to losses in childhood intelligence, memory and cogni- tion that are reflected in reductions in school performance and future wage-earning. 10 In many regions of Africa hookworm infection is syndemic with malaria and schistosomiasis. 11-14 A bivalent recombinant vaccine for human hookworm disease is under development. One of the lead candidate antigens in the vaccine is a glutathione S-transferase cloned from the hookworm Necator americanus (Na-GST-1) which is expressed in the yeast Pichia pastoris. Based on preliminary studies demonstrating that the recombinant protein was not stable in an acetate buffer at pH 6, we undertook an extensive stability analysis of the molecule. To improve and optimize stability we complemented traditional methods employed for macromolecule and vaccine stabilization with biophysical techniques that were incorporated into a systematic process based on an eigenvector approach. Large data sets, obtained from a variety of experimental methods were used to establish a color map (“empirical phase diagram”) of the physical stability of the vaccine antigen over a wide range of temperature and pH. The resulting map defined “apparent phase boundaries” that were used to develop high throughput screening assays. These assays were then employed to identify excipients that stabilized the antigen against physical degradation that could otherwise result in losses of physicochemical integrity, immunogenicity and potency of the vaccine. Based on these evaluations, the recombinant Na-GST-1 antigen was reformulated and ultimately produced under Good Manufacturing Practices and with an acceptable stability profile. Advances in vaccines against neglected tropical diseases Enhancing physical stability of a recombinant hookworm vaccine through biophysical and formulation studies Jordan L. Plieskatt, 1, * Wanderson C. Rezende, 1,† Chris M. Olsen, 2 Jared M. Trefethen, 2 Sangeeta B. Joshi, 2 C. Russell Middaugh, 2 Peter J. Hotez 1,† and Maria Elena Bottazzi 1,† 1 Department of Microbiology, Immunology and Tropical Medicine; The George Washington University; Washington, DC USA; 2 Department of Pharmaceutical Chemistry; University of Kansas; Kansas USA; † Current Affiliation: Sabin Vaccine Institute and Texas Children’s Hospital Center for Vaccine Development; National School of Tropical Medicine; Baylor College of Medicine; Houston, TX USA. Key words: neglected tropical diseases, recombinant vaccines, hookworm, Necator americanus, product stability, biophysical studies, formulation This manuscript has been published online, prior to printing. Once the issue is complete and page numbers have been assigned, the citation will change accordingly. Public health control of hookworm in endemic countries relies on periodic deworming with a benzimidazole anthelminthic drug. 15 However, drug failure with the benzimidazole meben- dazole is now widespread when treating hookworm, 16 possibly because of resistance, 17 while administration of another benz- imidazole, albendazole, can fail within a few months. 18 Thus, the development of a hookworm vaccine could help to affect the global control or elimination of hookworm. 14 A bivalent human hookworm vaccine is under development and will soon enter phase 1 clinical testing. 14 This vaccine will be comprised of two recombinant N. americanus antigens formu- lated on an aluminum salt. One of the antigens is a 45 kDa recom- binant aspartic protease known as Na-APR-1 while the other is a 24 kDa glutathione S-transferase designated Na-GST-1. 14 These enzymes are required for adult parasite hemoglobin digestion and heme detoxification, respectively. These candidate antigens were selected based on their ability to protect against N. ameri- canus and other hookworm challenge infections, as well as other criteria. 14 Both antigens have undergone extensive preclinical

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Human Vaccines & Immunotherapeutics 1
Human Vaccines & Immunotherapeutics 8:6, 1-12; June 2012; © 2012 Landes Bioscience
ReseaRcH papeR ReseaRcH papeR
*Correspondence to: Jordan L. Plieskatt; Email: [email protected]: 12/01/11; Revised: 02/10/12; Accepted: 02/16/12http://dx.doi.org/10.4161/hv.19726
Introduction
The neglected tropical diseases (NTDs) are a group of 17 major chronic and disabling infections affecting more than 1 billion people worldwide, primarily those living in rural areas, urban slums or conflict zones.1,2 Human hookworm infection is one of the world’s most common and debilitating NTDs responsible for an estimated 600 million cases in the world’s poorest countries; most of these infections are caused by Necator americanus.3-5 The clinical signature of hookworm infection is anemia and protein malnutrition,6 which results from hookworm associated blood loss, especially in children and pregnant women.7,8 Blood loss from hookworm is responsible for a disease burden equivalent to up to one-half of the disability-adjusted life years lost that result from better known devastating infections such as malaria,9 in addition to losses in childhood intelligence, memory and cogni-tion that are reflected in reductions in school performance and future wage-earning.10 In many regions of Africa hookworm infection is syndemic with malaria and schistosomiasis.11-14
a bivalent recombinant vaccine for human hookworm disease is under development. One of the lead candidate antigens in the vaccine is a glutathione s-transferase cloned from the hookworm Necator americanus (Na-GsT-1) which is expressed in the yeast Pichia pastoris. Based on preliminary studies demonstrating that the recombinant protein was not stable in an acetate buffer at pH 6, we undertook an extensive stability analysis of the molecule. To improve and optimize stability we complemented traditional methods employed for macromolecule and vaccine stabilization with biophysical techniques that were incorporated into a systematic process based on an eigenvector approach. Large data sets, obtained from a variety of experimental methods were used to establish a color map (“empirical phase diagram”) of the physical stability of the vaccine antigen over a wide range of temperature and pH. The resulting map defined “apparent phase boundaries” that were used to develop high throughput screening assays. These assays were then employed to identify excipients that stabilized the antigen against physical degradation that could otherwise result in losses of physicochemical integrity, immunogenicity and potency of the vaccine. Based on these evaluations, the recombinant Na-GsT-1 antigen was reformulated and ultimately produced under Good Manufacturing practices and with an acceptable stability profile.
Advances in vaccines against neglected tropical diseases
Enhancing physical stability of a recombinant hookworm vaccine through biophysical and formulation studies
Jordan L. plieskatt,1,* Wanderson c. Rezende,1,† chris M. Olsen,2 Jared M. Trefethen,2 sangeeta B. Joshi,2 c. Russell Middaugh,2 peter J. Hotez1,† and Maria elena Bottazzi1,†
1Department of Microbiology, Immunology and Tropical Medicine; The George Washington University; Washington, Dc Usa; 2Department of pharmaceutical chemistry; University of Kansas; Kansas Usa; †current affiliation: sabin Vaccine Institute and Texas children’s Hospital center for Vaccine Development; National school of Tropical
Medicine; Baylor college of Medicine; Houston, TX Usa.
Key words: neglected tropical diseases, recombinant vaccines, hookworm, Necator americanus, product stability, biophysical studies, formulation
Thi
s m
anus
crip
t ha
s be
en p
ublis
hed
onlin
e, p
rior
to
prin
ting
. Onc
e th
e is
sue
is c
ompl
ete
and
page
num
bers
hav
e be
en a
ssig
ned,
the
cit
atio
n w
ill c
hang
e ac
cord
ingl
y.
Public health control of hookworm in endemic countries relies on periodic deworming with a benzimidazole anthelminthic drug.15 However, drug failure with the benzimidazole meben-dazole is now widespread when treating hookworm,16 possibly because of resistance,17 while administration of another benz-imidazole, albendazole, can fail within a few months.18 Thus, the development of a hookworm vaccine could help to affect the global control or elimination of hookworm.14
A bivalent human hookworm vaccine is under development and will soon enter phase 1 clinical testing.14 This vaccine will be comprised of two recombinant N. americanus antigens formu-lated on an aluminum salt. One of the antigens is a 45 kDa recom-binant aspartic protease known as Na-APR-1 while the other is a 24 kDa glutathione S-transferase designated Na-GST-1.14 These enzymes are required for adult parasite hemoglobin digestion and heme detoxification, respectively. These candidate antigens were selected based on their ability to protect against N. ameri-canus and other hookworm challenge infections, as well as other criteria.14 Both antigens have undergone extensive preclinical

© 2012 Landes Bioscience.
Do not distribute.
2 Human Vaccines & Immunotherapeutics Volume 8 Issue 6
Early testing of the purified recombinant protein revealed that the molecule was not stable in the acetate buffer at pH 6. While performing traditional empirical stability testing stressing the molecule at 2–8°C for various time periods, stability failure was observed within three months due to protein degradation (Fig. 1A). Analysis using SDS-PAGE revealed that the recombi-nant protein had reduced relative purity. As seen in Figure 1A, when the recombinant protein was run under reducing condi-tions, the protein stored at 2–8°C for 3 mo had a relative purity that decreased from 93–95% as maintained when stored under frozen conditions (below -50°C) to 87–88%, which was below the allowable passing specification of purity (>90%). The relative purity also decreased with a similar banding pattern under non-reducing conditions (data not shown). Peak optical density (OD) also decreased from reference (0.57) to a peak OD of (0.39–0.41) for replicate samples stored at 2–8°C.
Subsequent analysis of the recombinant protein stored below -50°C at the same testing points also resulted in stability failure due to precipitation/aggregation seen upon freeze/thaw (Fig. 1B). Multiple samples upon freeze thaw showed a visibly cloudy solution with particulates (data not shown). These samples were centrifuged to isolate the precipitate. Supernatant from these samples were run on SDS-PAGE to confirm identity (Fig. 1B). The visible pellet was then resuspended in sample buffer and run on SDS-PAGE to con-firm the presence of Na-GST-1 protein in the precipitate (Fig. 1B).
Recombinant protein samples were also analyzed by a second orthogonal method, size exclusion HPLC (SE-HPLC). Briefly, 50 μg of a reference protein sample stored under frozen conditions (below -50°C) was analyzed alongside samples stored at 2–8°C. Figure 1C shows the expected profile of Na-GST-1 recombinant protein with an area count of approximately 5.2 million. Figure 1D show the recombinant protein stored at 2–8°C for 3 mo with a decreased main peak (approximately 4 million area counts) in addition to several minor peaks at later retention time. These results preliminarily can be correlated with the minor bands observed with SDS-PAGE analysis leading to decreased purity (82% by SE-HPLC).
The minor bands detected with SDS-PAGE were sequenced to determine whether these bands are products of the Na-GST-1 protein (data not shown). Results obtained from N-terminal sequencing identified a full-length band and two minor bands (approximately 10 kDa and 7 kDA). The 10 kDA fragment rep-resents a potential N-terminal cleavage at approximately the 21st amino acid of the Na-GST-1 full length sequence, while the 7 kDa fragment maintained an intact N-terminus with probable cleavage at the C-terminus of the protein. In summary, both frag-ments are product derived with peptide hydroloysis (proteolytic cleavage) as the potential cause of decreased purity.
A functional assay was used to determine the enzymatic activity of the Na-GST-1 recombinant protein. This assay deter-mines if the changes in protein stability had an impact on pro-tein functionality. Recombinant Na-GST-1 protein samples in a non-stabilizing buffer (50 mM sodium acetate pH 6.0) were subjected to heat-induced degradation (100°C or 55°C for 5 min) and also reducing conditions. The induced protein aggregation caused by ionic interactions was confirmed by non-reducing
testing in laboratory animal models of hookworm infection and reviewed in reference 14.
The Na-GST-1 Vaccine Candidate Antigen
The Na-GST-1 recombinant protein was expressed in the yeast strain Pichia pastoris.19,20 Amino acid sequence align-ment with GSTs from other organisms show that Na-GST-1 belongs to a nematode-specific nu-class of GST family unique in its high heme-binding capacities.20 Na-GST-1 may function in parasite heme- and hematin-detoxification. Therefore, host antibody specific to Na-GST-1 might neutralize function and facilitate oxidative damage to the parasite.14,20 X-ray crystal-lographic studies of Na-GST-1 revealed that the enzyme can form homodimers in solution to create large binding cavities that are accessible to heme and hematin.19 To date, process development of the Na-GST-1 recombinant protein included suitability studies to ensure the production of consistent, stable and reproducible recombinant protein. This paper provides a summary of the stability studies performed during process development to understand and evaluate the physical state of the vaccine target as a function of stress (i.e., temperature and pH). In addition, we present data for the screening and iden-tification of buffering agents and stabilizers (sugars and other excipients) to prevent the degradation and enhance the stabil-ity of formulated Na-GST-1. Finally, we show ongoing stability data of a GMP manufactured Na-GST-1 recombinant protein in the selected buffer and a vaccine formulation for up to 18 mo post manufacture.
Results
Stability profile of the Na-GST-1 recombinant protein. Process development and manufacture rely on developing assays for the product’s characterization (appearance, identity, confor-mation, integrity and purity), identification of contaminants and evaluation of stability. For the Na-GST-1 vaccine antigen, process development was performed at the 10-L scale and manu-facturing was performed at the 60-L scale (unpublished data). The purified recombinant protein was initially produced in a final buffer consisting of 50 mM sodium acetate pH 6.0 and stored under freezing conditions (below -50°C). The purified protein during manufacture was analyzed according to a quality control program including multiple assays for identity (western blot, N-terminal sequencing and mass spectrometry) and integ-rity (SDS-PAGE and size-exclusion liquid chromatography) as well as purity assays including those to detect residual host cell proteins such as a slot-blot immunoassay developed for P. pasto-ris proteins (unpublished data). Na-GST-1 manufactured under this developed process contained no detectable host cell proteins (<0.16% w/w) and an overall purity of the intact main species of >90% (data not shown). A summary of the purified protein analysis and specifications can be reviewed in Tables S1 and S2 respectively. Enzymatic activity was confirmed with an in vitro enzymatic assay used to monitor integrity, proper folding and functionality.20

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Human Vaccines & Immunotherapeutics 3
In summary, our investigations demonstrated that the initial buffer selected was not adequate to provide suitable stability for this protein. Specifically the pH of the final buffer was close to the theoretical pI of 6.3 of Na-GST-1, as confirmed by isoelec-tric focusing gels (data not shown), which may contribute to the generation of hydrophobic and covalent interactions. Therefore, we initiated extensive biophysical screening studies to better
SDS-PAGE analysis (data not shown). The disruption of protein structure was then confirmed by the loss of enzymatic activity of Na-GST-1 (Fig. 2). Upon induced heat stress, as compared with a control protein stored at <-50°C and not exposed to elevated temperatures prior to analysis, aggregated Na-GST-1 was found to have lost its enzymatic activity indicating thermal stability to be an important factor in maintaining a native active protein.
Figure 1. (a) sDs-paGe under reducing conditions containing 2 μg of Na-GsT-1 in 50 mM sodium acetate pH 6.0 buffer. Lane a represents reference protein stored at <-50°c and Lanes B–e are replicate samples of the recombinant protein stored at 2–8°c. (B) sDs-paGe under non-reducing conditions of Na-GsT-1 samples. Lanes (a and B) correspond to supernatant from two protein samples where particulates were observed after freeze thaw. Lanes (c and D) represent protein samples from lanes (a and B) after they were centrifuged to isolate particulates and resuspended in sample buffer. (c and D) se-HpLc of two recombinant Na-GsT-1 protein samples. (c) represents recombinant protein stored at freezing temperature (below -50°c). (D) rep-resents recombinant protein stored at 2–8°c for 3 mo with a decrease in main peak area, purity and detection of lower molecular weight breakdown products.

© 2012 Landes Bioscience.
Do not distribute.
4 Human Vaccines & Immunotherapeutics Volume 8 Issue 6
Figure 3B displays the effect of temperature on Na-GST-1 sec-ondary structure by following the signal at 222 nm (primarily α helix) as the temperature was increased from 10 to 87.5°C. At pH 3, there was little to no change in the signal at 222 nm as a func-tion of temperature due to the already conformationally altered nature of the protein. At pH 4, the onset of the transition began at ~27°C, with a Tm of ~32°C. There was an initial decrease in the signal at ~20°C for pH 5 and a second larger unfolding of the α-helical structure at ~41°C with an observed T
M at 45°C.
The observed TM
values for the Na-GST-1 protein at pH 6–8 are approximately 52°C, 56°C and 59°C, respectively, while onset temperatures for the main transition ranged from 50–53°C. Overall, the onset temperature increased with increasing pH, as did the T
M. Thus, the secondary structure stability of Na-GST-1
appears to be higher at higher (near neutral) pH values.Fluorescence studies. Changes in the tertiary structure of
Na-GST-1 were studied by monitoring changes in the trypto-phan (Trp) emission peak position as a function of both pH and temperature (Fig. 3C). In general, a red shift in the Trp emission maximum as a function of temperature was observed, suggesting that the indole side chain was exposed to a more polar (aque-ous) environment upon thermal unfolding. At pH 3, however, a slight red shift in the starting wavelength to 332 nm and an overall lack of change in position as a function of temperature indicated that Na-GST-1 was already structurally perturbed at this pH (consistent with the observations from the CD studies). The initial value of ~328 nm at all other pH values suggests that the indole side chains of the Trp residues were relatively relatively
understand the origin of the observed degradation and to provide solution candidates that would improve stability.
Biophysical characterization and construction of a mul-tiple technique-based empirical phase diagram (EPD) for the Na-GST-1 recombinant protein. To improve and optimize pro-tein stability, a systematic approach to the characterization and stabilization of vaccine antigens was undertaken. This approach used a combination of biophysical techniques over a range of tem-perature and pH. The large data sets, obtained from techniques such as circular dichroism, intrinsic and extrinsic fluorescence and light scattering, have been used previously to evaluate vac-cine stability and were used to establish a stability map (“EPD”) of Na-GST-1.21-28
Circular dichroism studies. Changes in the secondary struc-ture of Na-GST-1 were studied by monitoring the change in the far UV-CD signal as a function of both pH and temperature. Figure 3A shows the pH dependence of the secondary structure of Na-GST-1. In general, a broad peak from 240 nm to 200 nm was observed at all pH values examined suggesting a mixture of secondary structure types. A minimum at 222 nm and a shoulder at 208 nm indicate a predominance of α-helical structure while the broad nature of the peak between the two minima suggests contribution from β sheet as well. The CD spectra of Na-GST-1 from pH 6–8 are similar, while a decrease in pH caused an over-all reduction in the signal intensity from pH 5 to 3. At pH 4, the CD signal begins to significantly decrease in intensity indicating a major loss of secondary structure; and at pH 3, the CD spec-trum indicates that Na-GST-1 was severely structurally altered.
Figure 2. enzymatic activity of 40 μg Na-GsT-1 protein measured by fluorescence assay and reported in relative fluorescence units (RFU) under dif-ferent treatment conditions: control = untreated (frozen protein and thawed for use); heat treated at two temperatures (55°c and 100°c) and under reducing conditions with DTT. Results shown are average of duplicate measurements.

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Human Vaccines & Immunotherapeutics 5
to unfold, an increase in fluorescence intensity was observed (excluding Na-GST-1 at pH 3, which has been shown to already be in an altered state), followed by a decrease in the signal inten-sity (~60°C) possibly due to protein aggregation (see below) and/or the intrinsic temperature effect. The following order in pro-tein stability was observed based on onset temperatures: pH 4 (30°C)<pH 5 (42°C)<pH 6 (50°C)~pH 7 (50°C)~pH 8 (50°C). In agreement with the CD results, the fluorescence data shows that GST-1 is thermally more stable at near neutral pH values.
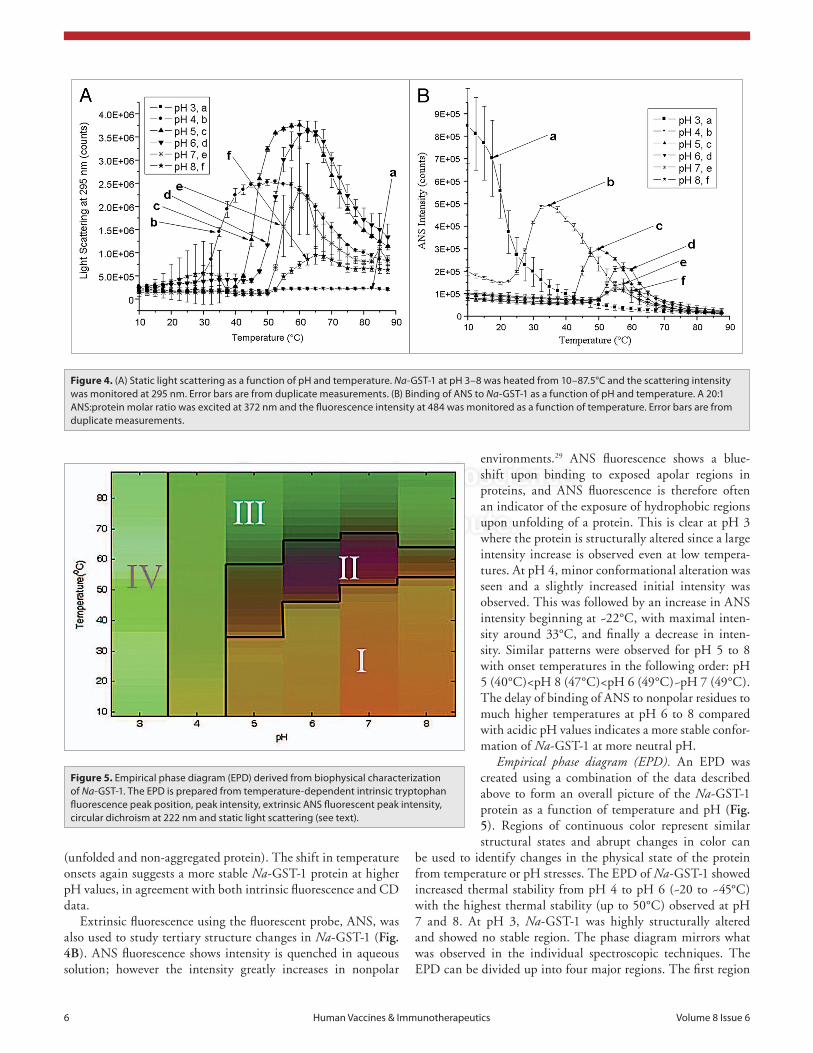
Static light scattering at the excitation wavelength was also examined (Fig. 4A). Generally, an increase in light scattering indicates an increase in the size of the particles in solution. In the case of Na-GST-1, light scattering at 295 increased from pH 4 to 7 and then decreased again at pH 8. The onset temperatures for scattering increased with increasing pH in the following order: pH 4 (28°C)<pH 5 (38°C)<pH 6 (45°C)<pH 7 (50°C)~pH 8 (50°C), with no changes in light scattering observed for pH 3
buried in an apolar environment and upon heating the peak posi-tions shifted to longer wavelengths (~334 nm) due to exposure to a more polar environment. The magnitude of the red shift was slightly decreased at pH 4, while similar shifts were observed at pH 5–8. Following a similar trend to that observed in CD, the onset temperatures for the transitions were in the following order: pH 4 (~28°C)<pH 5 (~38°C)<pH 6 (~45°C)~pH 7 (~45°C)<pH 8 (~48°C). Tm values for these transitions were calculated using a first derivative method to yield the following order: pH 4 (32.2°C)<pH 5 (45.5°C<pH 6 (52.0°C)~pH 7 (53.0°C)~pH 8 (52.3°C). The overall noise at higher temperatures was presum-ably due to sample aggregation.
Fluorescence intensity values as a function of temperature are also used to provide information about the unfolding of a protein’s tertiary structure (Fig. 3D). A general decrease in fluo-rescence intensity was observed as temperature was increased, due to the intrinsic effect of temperature. As the protein began
Figure 3. (a) The effect of ph on the secondary structure of Na-GsT-1 at 10°c. each data point represents individual measurements of cD spectra from a single representative sample. (B) ellipticity at 222 nm as a function of temperature was measured over the pH range of 3–8. Duplicate measurements were taken and the average value displayed. (c) Tryptophan emission fluorescence peak position of Na-GsT-1 as a function of pH and temperature. Na-GsT-1 at pH 3–8 was heated from 10–87.5°c, and the fluorescence emission maximum was determined after excitation at 295 nm. (D) Tryptophan emission fluorescence intensity of Na-GsT-1 as a function of pH and temperature. Na-GsT-1 at pH 3–8 was heated from 10–87.5°c. error bars are from duplicate measurements.
© 2012 Landes Bioscience.
Do not distribute.
6 Human Vaccines & Immunotherapeutics Volume 8 Issue 6
environments.29 ANS fluorescence shows a blue-shift upon binding to exposed apolar regions in proteins, and ANS fluorescence is therefore often an indicator of the exposure of hydrophobic regions upon unfolding of a protein. This is clear at pH 3 where the protein is structurally altered since a large intensity increase is observed even at low tempera-tures. At pH 4, minor conformational alteration was seen and a slightly increased initial intensity was observed. This was followed by an increase in ANS intensity beginning at ~22°C, with maximal inten-sity around 33°C, and finally a decrease in inten-sity. Similar patterns were observed for pH 5 to 8 with onset temperatures in the following order: pH 5 (40°C)<pH 8 (47°C)<pH 6 (49°C)~pH 7 (49°C). The delay of binding of ANS to nonpolar residues to much higher temperatures at pH 6 to 8 compared with acidic pH values indicates a more stable confor-mation of Na-GST-1 at more neutral pH.
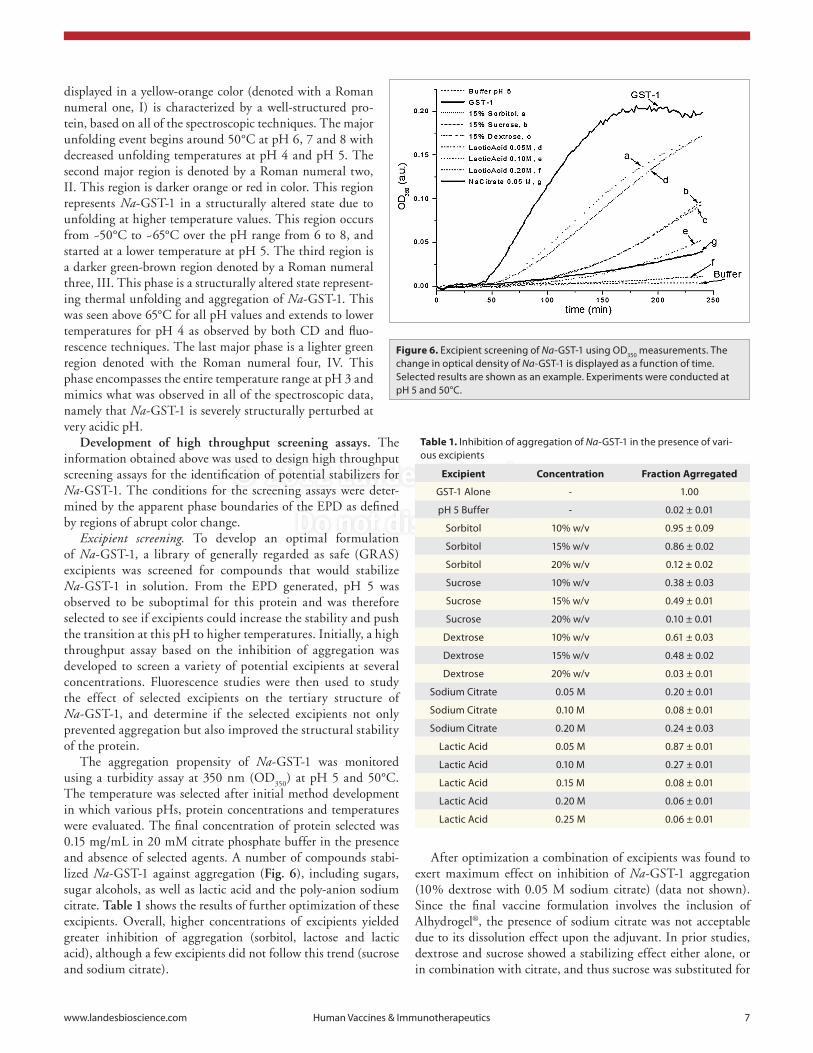
Empirical phase diagram (EPD). An EPD was created using a combination of the data described above to form an overall picture of the Na-GST-1 protein as a function of temperature and pH (Fig. 5). Regions of continuous color represent similar structural states and abrupt changes in color can
be used to identify changes in the physical state of the protein from temperature or pH stresses. The EPD of Na-GST-1 showed increased thermal stability from pH 4 to pH 6 (~20 to ~45°C) with the highest thermal stability (up to 50°C) observed at pH 7 and 8. At pH 3, Na-GST-1 was highly structurally altered and showed no stable region. The phase diagram mirrors what was observed in the individual spectroscopic techniques. The EPD can be divided up into four major regions. The first region
(unfolded and non-aggregated protein). The shift in temperature onsets again suggests a more stable Na-GST-1 protein at higher pH values, in agreement with both intrinsic fluorescence and CD data.
Extrinsic fluorescence using the fluorescent probe, ANS, was also used to study tertiary structure changes in Na-GST-1 (Fig. 4B). ANS fluorescence shows intensity is quenched in aqueous solution; however the intensity greatly increases in nonpolar
Figure 4. (a) static light scattering as a function of pH and temperature. Na-GsT-1 at pH 3–8 was heated from 10–87.5°c and the scattering intensity was monitored at 295 nm. error bars are from duplicate measurements. (B) Binding of aNs to Na-GsT-1 as a function of pH and temperature. a 20:1 aNs:protein molar ratio was excited at 372 nm and the fluorescence intensity at 484 was monitored as a function of temperature. error bars are from duplicate measurements.
Figure 5. empirical phase diagram (epD) derived from biophysical characterization of Na-GsT-1. The epD is prepared from temperature-dependent intrinsic tryptophan fluorescence peak position, peak intensity, extrinsic aNs fluorescent peak intensity, circular dichroism at 222 nm and static light scattering (see text).
© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Human Vaccines & Immunotherapeutics 7
After optimization a combination of excipients was found to exert maximum effect on inhibition of Na-GST-1 aggregation (10% dextrose with 0.05 M sodium citrate) (data not shown). Since the final vaccine formulation involves the inclusion of Alhydrogel®, the presence of sodium citrate was not acceptable due to its dissolution effect upon the adjuvant. In prior studies, dextrose and sucrose showed a stabilizing effect either alone, or in combination with citrate, and thus sucrose was substituted for
displayed in a yellow-orange color (denoted with a Roman numeral one, I) is characterized by a well-structured pro-tein, based on all of the spectroscopic techniques. The major unfolding event begins around 50°C at pH 6, 7 and 8 with decreased unfolding temperatures at pH 4 and pH 5. The second major region is denoted by a Roman numeral two, II. This region is darker orange or red in color. This region represents Na-GST-1 in a structurally altered state due to unfolding at higher temperature values. This region occurs from ~50°C to ~65°C over the pH range from 6 to 8, and started at a lower temperature at pH 5. The third region is a darker green-brown region denoted by a Roman numeral three, III. This phase is a structurally altered state represent-ing thermal unfolding and aggregation of Na-GST-1. This was seen above 65°C for all pH values and extends to lower temperatures for pH 4 as observed by both CD and fluo-rescence techniques. The last major phase is a lighter green region denoted with the Roman numeral four, IV. This phase encompasses the entire temperature range at pH 3 and mimics what was observed in all of the spectroscopic data, namely that Na-GST-1 is severely structurally perturbed at very acidic pH.
Development of high throughput screening assays. The information obtained above was used to design high throughput screening assays for the identification of potential stabilizers for Na-GST-1. The conditions for the screening assays were deter-mined by the apparent phase boundaries of the EPD as defined by regions of abrupt color change.
Excipient screening. To develop an optimal formulation of Na-GST-1, a library of generally regarded as safe (GRAS) excipients was screened for compounds that would stabilize Na-GST-1 in solution. From the EPD generated, pH 5 was observed to be suboptimal for this protein and was therefore selected to see if excipients could increase the stability and push the transition at this pH to higher temperatures. Initially, a high throughput assay based on the inhibition of aggregation was developed to screen a variety of potential excipients at several concentrations. Fluorescence studies were then used to study the effect of selected excipients on the tertiary structure of Na-GST-1, and determine if the selected excipients not only prevented aggregation but also improved the structural stability of the protein.
The aggregation propensity of Na-GST-1 was monitored using a turbidity assay at 350 nm (OD
350) at pH 5 and 50°C.
The temperature was selected after initial method development in which various pHs, protein concentrations and temperatures were evaluated. The final concentration of protein selected was 0.15 mg/mL in 20 mM citrate phosphate buffer in the presence and absence of selected agents. A number of compounds stabi-lized Na-GST-1 against aggregation (Fig. 6), including sugars, sugar alcohols, as well as lactic acid and the poly-anion sodium citrate. Table 1 shows the results of further optimization of these excipients. Overall, higher concentrations of excipients yielded greater inhibition of aggregation (sorbitol, lactose and lactic acid), although a few excipients did not follow this trend (sucrose and sodium citrate).
Figure 6. excipient screening of Na-GsT-1 using OD350 measurements. The change in optical density of Na-GsT-1 is displayed as a function of time. selected results are shown as an example. experiments were conducted at pH 5 and 50°c.
Table 1. Inhibition of aggregation of Na-GsT-1 in the presence of vari-ous excipients
Excipient Concentration Fraction Agrregated
GsT-1 alone - 1.00
pH 5 Buffer - 0.02 ± 0.01
sorbitol 10% w/v 0.95 ± 0.09
sorbitol 15% w/v 0.86 ± 0.02
sorbitol 20% w/v 0.12 ± 0.02
sucrose 10% w/v 0.38 ± 0.03
sucrose 15% w/v 0.49 ± 0.01
sucrose 20% w/v 0.10 ± 0.01
Dextrose 10% w/v 0.61 ± 0.03
Dextrose 15% w/v 0.48 ± 0.02
Dextrose 20% w/v 0.03 ± 0.01
sodium citrate 0.05 M 0.20 ± 0.01
sodium citrate 0.10 M 0.08 ± 0.01
sodium citrate 0.20 M 0.24 ± 0.03
Lactic acid 0.05 M 0.87 ± 0.01
Lactic acid 0.10 M 0.27 ± 0.01
Lactic acid 0.15 M 0.08 ± 0.01
Lactic acid 0.20 M 0.06 ± 0.01
Lactic acid 0.25 M 0.06 ± 0.01

© 2012 Landes Bioscience.
Do not distribute.
8 Human Vaccines & Immunotherapeutics Volume 8 Issue 6
is also consistent with previous lots of Na-GST-1 manufactured in 50 mM sodium acetate pH 6 (data not shown).
Additional accelerated stability studies were also completed on the recombinant protein and formulated vaccine includ-ing studies using elevated temperatures (25°C and 37°C) and multiple freeze thaw cycles according to International Code for Harmonization (ICH) and included in regulatory filings to support the overall stability program of the clinical product. Multiple assays including isoelectric focusing gels were used during these studies to monitor deamidation and stability (data not shown) and showed the recombinant protein was stable at 25°C for up to 6 mo and formulated vaccine at 37°C for 6 mo (data not shown). The only change to note was the adsorption to Alhydrogel® at elevated temperature and extended time became stronger and resulted in an increase in potency (unpublished data). A representative set of tests to show the overall stability of the purified Na-GST-1 manufactured protein stored at 2–8°C up to 18 mo is shown in Table 2. Multiple reference lots were also monitored under storage at -50°C with no significant change (data not shown).
Discussion
The initial buffer for Na-GST-1 was selected based on preliminary orthogonal data and based on buffers used for a prior hookworm vaccine candidate, Na-ASP-2.30 Because some recombinant vac-cine antigens are inherently unstable, extensive formulation and stabilization studies are often necessary. This activity, which was not explored in the early phases of the Na-GST-1 program lead to initial stability failures of the Na-GST-1 recombinant protein. This underscores the importance of monitoring and evaluation of the protein at different temperatures, pH values, and under different buffer conditions using traditional approaches as well as a more systematic process based on a novel eigenvector approach. While most stability indicating assays showed a loss of physical stability (i.e., aggregation or color and appearance failure with particulates) an enzymatic assay was used in an attempt to cor-relate these thermal stability results to the functional activity of Na-GST-1. The functional assay demonstrated that samples in the original (not stabilized) buffer when exposed to elevated temperatures and resulting in subsequent biochemical stability failure also showed a significant loss of native enzymatic activity. This information provided further evidence that thermal stabil-ity would be important in maintaining a suitable vaccine candi-date and advanced biophysical screening techniques were thus explored.
Multiple techniques that monitored a variety of protein char-acteristics (secondary and tertiary structure, as well as aggrega-tion propensity) were used to study the effect of temperature and pH on the structural stability of Na-GST-1. CD, intrinsic Trp fluorescence, extrinsic ANS fluorescence and static light scatter-ing all indicated that the Na-GST-1 was unstable at low pH, but showed a marked increase in thermal stability to ~50°C near bio-logical pH conditions (pH 7 to 8). The data from these various techniques were combined to create an empirical phase diagram, which was then used to select conditions to screen a variety of
sodium citrate. After the completion of initial stabilizer screen-ing under stressed conditions (pH 5.0 and 50°C), final studies were performed at optimal pH 7.4 in a 10 mM imidazole buffer to evaluate the effect of the potential stabilizers on the confor-mational stability of Na-GST-1. Thus, intrinsic Trp fluorescence was used to monitor Trp peak position shifts, peak intensity values and static light scattering of Na-GST-1 in 10% dextrose alone, 15% sucrose alone and a mixture of 10% dextrose and 15% sucrose. Individually, the excipients stabilized Na-GST-1, but combined excipients produced a significant enhancement in thermal stability of ~5°C in Tm compared with Na-GST-1 alone (data not shown).
Formulation development of Na-GST-1. Na-GST-1 formu-lated with Alhydrogel®. Using the information generated from the biophysical profiling and excipient screening, the indicated buffers were tested for their compatibility with Alhydrogel®. Alhydrogel® is the selected human adjuvant of choice for the vaccine development program given its regulatory history, access and compatibility with other program candidates. Preliminary hamster challenge trials demonstrated efficicacy of Na-GST-1 absorbed to Alhydrogel®20 in addition to potency assay develop-ment in a mouse model (data now shown) and preclinical toxicol-ogy studies (data not shown). The intended Na-GST-1 vaccine consists of a formulation of 0.1 mg/mL protein and 0.8 mg/mL Alhydrogel® stored at 2–8°C. The buffers containing 10% dex-trose, 50 mM sodium citrate, 10 mM imidazole at pH 7.4 and 10% dextrose, 10 mM imidazole at pH 7.4 were used to prepare formulated Na-GST-1. After adsorption, the supernatants were sampled and analyzed for the presence of unabsorbed protein in the supernatant both by SDS-PAGE and silver staining. A series of standards on the gel were also used to determine the percentage free protein. In the absence of citrate treatment, 100% of the pro-tein was absorbed to the Alhydrogel®. When the formulation was treated with citrate at a concentration of 50 mM the Na-GST-1 protein desorbed from the Alhydrogel® and could be detected in the supernatant (Fig. S1). The complete binding of Na-GST-1 to Alhydrogel® and in the 10% dextrose, 10 mM imidazole buffer pH 7.4 buffer, has been monitored and confirmed, for up to 18 mo (Data not shown).
Ongoing stability assessment of recombinant protein Na-GST-1 in 10% dextrose, 10 mM imidazole pH 7.4. Na-GST-1 was manu-factured under GMP and formulated into a final buffer of 10% dextrose, 10 mM imidazole pH 7.4. In addition to the character-ization assays at the time of manufacture (shown in Tables S1 and S2), ongoing stability analysis up to 18 mo has been completed. Figure S2 shows a representative mass spectrometry analysis for Na-GST-1 at manufacture and 18 mo post manufacture when stored at 2–8°C. The predicted mass of Na-GST-1 is 23,679 Da without an intra-molecular disulfide bond. In Figure S2, two main peaks are seen at 23, 676 and 23,692 Da. The second peak and other subsequent peaks are +16 Daltons, which would corre-spond to possibly one single oxidation event. Na-GST-1 contains four methionine residues, which are possible sites for oxidation events. When stored in the 10% dextrose, 10 mM Imidazole pH 7.4 buffer, there is no increase in minor peaks with respect to the parent peak over time, however. This mass spectrometry profile

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Human Vaccines & Immunotherapeutics 9
suggested formulation would have included a combination of 10% dextrose and 15% sucrose in 10 mM imidazole buffer at pH 7.4. After further optimization and to decrease the osmolal-ity of the solution 10% dextrose in 10 mM imidazole pH 7.4 was selected. It should also be noted that sucrose can hydrolyze during storage at high temperatures or low pH to form reducing monosaccharides such as dextrose that may react with primary amino groups of proteins and produce glycation products which may alter a protein’s physicochemical stability. Furthermore, since dextrose is a reducing sugar, its effect on the chemical stability of Na-GST-1 protein needs to be more thoroughly investigated. While advance biophysical characterization and construction of a multiple technique-based empirical phase diagram (EPD) for Na-GST-1 provides immediate screening and feedback of buffer composition, selected assays should be continued as part of the overall stability program of the molecule to monitor changes over time and the overall impact on the clinical products efficacy and stability. Additionally, orthogonal techniques such as color and appearance and size exclusion HPLC should not be overlooked as these often provide real time feedback of the molecule and its behavior and stability in the selected buffer. Finally, the chemi-cal degradation pathway was not thoroughly explored with these methods and this candidate and should be explored as the candi-date moves forward through clinical trials.
Ongoing product development. The Na-GST-1 molecule has since been manufactured under cGMP conditions in 10% dextrose, 10 mM Imidazole pH 7.4, and in addition to multiple reference lots, is continually monitored as part of wider stability studies according to ICH guidelines. These studies, not discussed within this manuscript, are used to monitor the impact of the selected buffer components over time and the overall stability of Na-GST-1. The selected buffer has demonstrated physical stabil-ity (including no detectable deamidation) of the molecule through 18 mo at 2–8°C and short-term stability (6 mo) studies involving elevated temperatures (25°C and 37°C) that may be encountered
compounds for their ability to stabilize Na-GST-1 against both aggregation and thermal unfolding.
A wide variety of excipients that are generally recognized as safe (GRAS) were used in the excipient screening studies. These excipients belonged to different classes of compounds such as sugars, sugar alcohols, amino acids, salts, polymers and deter-gents among others. Each excipient was individually tested for its ability to stabilize protein structure and prevent aggrega-tion against thermal stress. A number of compounds stabilized Na-GST-1 against aggregation, including sugars, sugar alcohols, as well as lactic acid, and the poly-anion sodium citrate. Protein stabilization by sugars and polyols has been studied extensively and is thought in most cases to be due to non-specific effects such as preferential hydration.31,32 Kendrick et al. suggested that the degree of preferential exclusion and the protein’s chemical poten-tial is related to the degree of solvent accessible surface area.33 Thus, the system will favor the least extent of solvent accessible surface area and stabilize highly folded states of the proteins. Excipients such as detergents, salts and ionic compounds on the other hand are thought generally to exert their effects by directly interfering with protein-protein interactions and perturbation of solubility.34 Anionic compounds have been shown to selectively accumulate on the surface of proteins and directly impact protein stability and conformational integrity by shifting the conforma-tional equilibrium to native forms.32
While these advanced studies provide a vast array of infor-mation on both pH and buffer excipients that might result in the most physically stable candidate, they may not be a feasible buffer composition when taken into the overall clinical develop-ment program. Due to the addition of adjuvant (Alhydrogel®) to the formulations, the sugars dextrose and sucrose were selected while the poly-anionic sodium citrate was excluded. Using intrinsic tryptophan fluorescence in conjunction with static light scattering, the effects of the sugars on the tertiary structure and aggregation of the Na-GST-1 protein were studied. Overall the
Table 2. Representative stability testing of Na-GsT-1 recombinant protein manufactured in 10% D-glucose, 10 mM Imidazole pH 7.4 buffer and stored at 2–8°c up to 18 mo
Months Post Manufacture (stored at 2–8°C)
Test Expected Result 0 3 6 9 12 18
size exclusion-HpLc
One predominant peak eluting between 36–40
min. comparable to reference standard. peak
area >90% total area.
apex: 38.5 area: 99.32%
apex: 38.5 area: 99.52%
apex: 38.7 area: 99.24%
apex: 37.6 area: 98.98%
apex: 38.3 area: 98.18%
apex: 36.7 area: 98.64%
sDs-paGe/coomassie Blue
staining and Densitometry
>90% in main band by densitometry (non-reduced and reduced)
Non-reduced: 95.1%
Reduced: 97.4%
Non-reduced: 94.8%
Reduced: 95.8%
Non-reduced: 98.5%
Reduced: 98.2%
Non-reduced: 93.9%
Reduced: 93.7%
Non-reduced: 95.3%
Reduced: 94.1
Non-reduced: 95.1%
Reduced:
94.1%
pH pH 6.5–8 7.35 7.32 7.32 7.32 7.37 7.30
protein content by absorbance at
280 nm1.8–2.2 mg/mL 2.01 mg/mL 2.05 mg/mL 2.1 mg/mL 2.04 mg/mL 2.1 mg/mL 2.05 mg/mL

© 2012 Landes Bioscience.
Do not distribute.
10 Human Vaccines & Immunotherapeutics Volume 8 Issue 6
integration of the main bands as well as any other visible bands (if present).
Size exclusion-high performance liquid chromatography (SE-HPLC). A Waters Alliance 2,695 system with an in-line photo-diode array detector (2,996 PDA) and Empower 2 soft-ware was used with a G2000SWXL TSK-GEL-HPLC column (TOSOH Bioscience, 08540) fitted with a guard column and mobile phase of 50 mM sodium acetate pH 4.8 and flow rate of 0.25 mL/min. Na-GST-1 samples were diluted to a concentration of 1 mg/mL in 50 mM sodium acetate, pH 4.8. Following equili-bration and system suitability of the column with BioRad gel fil-tration standards [Fifty μg of molecular weight standards were run prior to Na-GST-1 and were used to calibrate the elution time of Na-GST-1] samples (50 μg) were injected. The retention time of Na-GST-1 was determined, and the % purity of the main peak was determined from a single main peak, with the apex elut-ing between 36–40 min.
Mass spectrometry (MS). A Waters MicroMass Q-Tof Micro mass spectrometer was used to determine the molecular weight of rNa-GST-1. The recombinant protein was desalted and diluted into formic acid for analysis. Buffer solution was removed by ZipTip desalting prior to analysis. The mobile phase for this analysis was 50% acetonitrile/0.1% formic acid or various ratios of methanol/water with 0.1% formic acid. For mass spectrometry analysis, 2–3 μL of the eluted sample was introduced into the mass spectrometer. The mass spectrometer was controlled and the data processed using the MassLynx 4.0 operating system. About a 100 scans are combined (to average out fluctuations in the elctrospray), the baseline subtracted, the data smoothed and the centroid of each observed peak determined. The software cal-culated the mass of the protein from two consecutive multiply charged ions in the spectrum which were manually defined.
Enzymatic activity. Na-GST-1 in 50 mM sodium acetate pH 6.0 was thawed from storage at <-50°C and either immediately used in the enzymatic assay or exposed to elevated temperatures (55°C and 100°C) for 5 min prior to analysis. Dilutions of the Na-GST-1 recombinant protein were prepared to a final concen-tration of 0.5 mg/mL and serially diluted 1:1 to 0.002 mg/mL. The diluted protein in a black microplate were then used with a fluorometric kit (Biovision, K260-100) that utilizes the reac-tion of monochlorobimane (MCB) with glutathione, which is catalyzed by Na-GST-1. Plates were measured in a Perkin Elmer Victor 3 at emission/excitation of 380/460 for 0.1 sec/well with a small emission aperture and CW-lamp energy of 2,496. Relative fluorescence units (RFU) at 10 min intervals were compared between Na-GST-1 and a quantitative control provide (GST mU/μL), which was also serially diluted. Duplicate samples typi-cally had a SD <500 RFU which is equivalent to approximately <2% variability within sample sets.
Biophysical profiling. Sample preparation. The recombinant Na-GST-1 protein (~1.3 mg/mL) was stored at -80°C in 50 mM sodium acetate buffer. Samples were thawed at room temperature and dialyzed against a constant buffer concentration of 20 mM citrate-phosphate in which the ratio of citric acid and sodium phosphate dibasic was varied to adjust the pH range from 3 to 8. Additionally, the ionic strength of the solutions (buffers and
in the supply train. Additionally, a pre-clinical toxicology study has been completed to demonstrate the safety of the molecule, its adjuvant and buffer components and used to support regulatory filings. Finally, the buffer components of Na-GST-1 are being evaluated for compatibility with the second recombinant protein (Na-APR-1) of the bivalent human hookworm vaccine. This pro-tein, which is currently under development, is also undergoing biophysical characterization and stabilization through similar methodology as presented in this manuscript.
Materials and Methods
Expression and purification of the Na-GST-1 recombinant protein. The Na-GST-1 gene was cloned into the pPICZαA (Invitrogen, V195-20) vector and expressed in P. pastoris X-33.20 Three chromatography steps were used to purify Na-GST-1 including a Q Sepharose XL (GE Healthcare, 17-5072-04) anion exchange capture step, followed by a Butyl Sepharose HP (GE Healthcare, 17-5432-04) chromatography step for additional polishing (unpublished data). A size exclusion chromatography step using Superdex 75 (GE Healthcare, 17-1044-04) removed additional high molecular weight host cell impurities in addition to serving as a buffer exchange step into a final buffer consist-ing of 50 mM sodium acetate pH 6 or 10% dextrose, 10 mM Imidazole pH 7.4.
Sodium dodecylsulfate-PAGE (SDS-PAGE) and staining. Sample preparation and electrophoresis. Sodium dodecylsulfate-PAGE (SDS-PAGE) was performed using pre-cast 4–20% Tris-glycine gels (Invitrogen) and an X-cell mini cell apparatus (Invitrogen. EC60252). Na-GST-1 protein samples were pre-pared by dilution to the desired concentration in their respective buffers (50 mM sodium acetate pH 6 or 10% dextrose, 10 mM imidazole pH 7.4), mixed 1:1 with 2x SDS-PAGE sample buffer (Invitrogen, LC2676) with or without a reducing agent (DTT). Samples are heated at 95–100°C for five minutes and gels run at a constant voltage of 135 ± 10 V per gel.
Coomassie staining. Gels were stained for 30 min (following an initial boiling) with a filtered solution of 40% methanol, 10% glacial acetic acid and two Phastgel Blue R tablets (GE Healthcare 17-0518-01) per liter. Destaining occurred with two changes (after boiling) of 5% methanol, 10% glacial acetic acid for 15 min each.
Silver staining. Gels were fixed in a 40% methanol, 10% ace-tic acid solution for at least 30 min, and then washed 3 times in 30% ethanol for 15 min each. Gels were then incubated in a 1x Dithionite solution for 5 min, washed with ultra-pure water and placed in a silver nitrate solution for 30 min. Following two washes with ultra-pure water the gel was developed in a sodium thiosulfate/carbonate solution containing formaldehyde for 10 min or until desired bands appeared. The reaction was stopped with multiple changes of 3.5% acetic acid solution and ultra-pure water.
Densitometry. A calibrated densitometer (Bio-Rad, GS-800) with Quantity One Software (Bio-Rad, 170-9600) was used to scan the gels. The purity of the sample on coomassie stained gels was determined after subtraction of background signal, and

© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com Human Vaccines & Immunotherapeutics 11
and light scattering), and the data were converted into a basis set for a multidimensional vector space. An N-dimensional vector was constructed for every combination of temperature and pH for which a measurement was taken (every 2.5°C, from 10 to 87.5°C and pH 3 to 8 at one unit intervals). The projectors of all vectors in the set were then summed to yield an N x N den-sity matrix with N eigenvectors. The three eigenvectors having the greatest eigenvalues were then used to transform the original N-dimensional vector set into three dimensions. Each vector is assigned to one of three different colors (red, green or blue), and the combination produces a unique color combination for each individual vector. All calculations were performed using Matlab software. Details of the EPD approach including the mathemati-cal theory and calculation process can be found elsewhere in ref-erences 21, 34 and 34.
Excipient screening. The aggregation of Na-GST-1 at pH 5 and 50°C was monitored by measuring the turbidity (optical den-sity at 350 nm) as a function of time. Duplicate samples in the presence or absence of various GRAS (generally regarded as safe) excipients were prepared at a final Na-GST-1 concentration of 0.15 mg/mL. Measurements were taken every two minutes over a period of four hours using a Molecular Devices Spectra Max M5 plate reader. The t = 0 value measured for each sample was sub-tracted from all subsequent measurements to calculate a ΔOD
350
across the time range. The percent inhibition of aggregation was then calculated as (1 - (ΔOD
350 sample/ΔOD
350 control)) x 100%,
using values corresponding to the time of maximum OD350
for the control sample (Na-GST-1 without excipient). After selec-tion of several promising excipients, secondary testing methods including intrinsic tryptophan fluorescence and light scattering were employed to further examine the effect of the additives on the conformational stability of Na-GST-1.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgements
This work supported by the Sabin Vaccine Institute through grants obtained from the Bill and Melinda Gates Foundation (Grant #32472 and #38988). The authors thank Mr. Aaron Miles for his supervisory role during the performance of the ana-lytical work.
Note
Supplemental material can be found at:www.landesbioscience.com/journals/hv/article/19726/
samples) was held constant at 0.15 using appropriate amounts of sodium chloride. The sample concentration was determined after dialysis using UV absorption spectroscopy at 280 nm and an extinction coefficient of 21,430 M-1cm-1, and then stored at 4°C.
Far-UV circular dichroism (CD) spectroscopy. CD spectra were obtained using a Jasco J-810 spectropolarimeter equipped with a peltier controlled 6-position cell holder. Spectra from 260 to 190 were collected at 2.5°C intervals as the temperature was increased from 10 to 87.5°C. A temperature ramp rate of 15°C/min and an incubation time of 120 sec were used. A scanning speed of 50 nm/min, resolution of 1 nm and a response time of 1 sec were used with a quartz cuvettes with pathlength of 0.1 cm. Buffer spectra were collected under the same set of conditions and sub-tracted from sample scans as a blank. Thermal melting curves were obtained by monitoring the CD signal at 222 nm as a func-tion of temperature.
Fluorescence spectroscopy. The intrinsic fluorescence of trypto-phan (Trp) was employed to detect tertiary structural changes of Na-GST-1 as a function of temperature. The spectra of Na-GST-1 were obtained using a Photon Technology International (PTI) spectrofluorometer equipped with a peltier controlled 4-position cell holder. The Trp in Na-GST-1 was excited at 295 and the emission spectra were recorded from 310 to 400 with a 1 nm step size and 1 sec integration time per point. Additionally, a second detector set at 180° from the emission signal was used to measure light scattering intensity at the excitation wavelength. Excitation, emission and static light scattering monochromator slits were set to 3 nm, 8 nm and 4 nm, respectively. Spectra were collected every 2.5°C with a 3 min equilibration time over the range of 10 to 87.5°C. Buffer spectra were collected under the same set of conditions and subtracted from sample scans as a blank.
Extrinsic fluoresce spectroscopy was conducted as a second-ary method to monitor the tertiary structure of Na-GST1 using the fluorescent dye, 1-anilino-8-naphthalene sulfonate (ANS). Stock solutions of ANS were prepared in 5% methanol w/v and the ANS concentration was determined using a molar extinction coefficient of 7,900 M-1cm-1 at 372 nm. The Na-GST-1 sample was mixed with a small aliquot of stock ANS solution to achieve an optimized ANS/protein molar ration of 20:1. The mixture was excited at 372 nm and the emission spectra were recorded from 400 to 600 nm.
Empirical phase diagrams (EPD). Empirical phase diagrams were constructed to create a visual representation of the data from the various techniques (CD, intrinsic Trp Fluorescence peak position and peak intensity, extrinsic ANS fluorescence intensity
References1. World Health Organization. Working to overcome the
global impact of neglected tropical diseases: First WHO report on neglected tropical diseases 2010; 172.
2. Hotez PJ. A Handful Of ‘Antipoverty’ Vaccines Exist For Neglected Diseases, But The World’s Poorest Billion People Need More. Health Aff(Millwood) 2011; 30:1080-7; http://dx.doi.org/10.1377/hlthaff.2011.0317.
3. Hotez PJ, Brindley PJ, Bethony JM, King CH, Pearce EJ, Jacobson J. Helminth infections: the great neglect-ed tropical diseases. J Clin Invest 2008; 118:1311-21; PMID:18382743; http://dx.doi.org/10.1172/JCI34261.
4. Hotez PJ, Fenwick A, Savioli L, Molyneux DH. Rescuing the bottom billion through control of neglected tropical diseases. Lancet 2009; 373:1570-5; PMID:19410718; http://dx.doi.org/10.1016/S0140-6736(09)60233-6.
5. Stoll NR. This wormy world. J Parasitol 1947; 33:1-18; PMID:20284977.
6. Hotez PJ, Brooker S, Bethony JM, Bottazzi ME, Loukas A, Xiao S. Hookworm infection. N Engl J Med 2004; 351:799-807; PMID:15317893; http://dx.doi.org/10.1056/NEJMra032492.
7. Smith JL, Brooker S. Impact of hookworm infec-tion and deworming on anaemia in non-pregnant populations: a systematic review. Trop Med Int Health 2010; 15:776-95; PMID:20500563; http://dx.doi.org/10.1111/j.1365-3156.2010.02542.x.
8. Brooker S, Hotez PJ, Bundy DA. Hookworm-related anaemia among pregnant women: a systematic review. PLoS Negl Trop Dis 2008; 2:291; PMID:18820740; http://dx.doi.org/10.1371/journal.pntd.0000291.

© 2012 Landes Bioscience.
Do not distribute.
12 Human Vaccines & Immunotherapeutics Volume 8 Issue 6
28. Kueltzo LA, Middaugh CR. Structural characterization of bovine granulocyte colony stimulating factor: effect of temperature and pH. J Pharm Sci 2003; 92:1793-804; PMID:12949998; http://dx.doi.org/10.1002/jps.10440.
29. Hawe A, Sutter M, Jiskoot W. Extrinsic fluorescent dyes as tools for protein characterization. Pharm Res 2008; 25:1487-99; PMID:18172579; http://dx.doi.org/10.1007/s11095-007-9516-9.
30. Goud GN, Bottazzi ME, Zhan B, Mendez S, Deumic V, Plieskatt J, et al. Expression of the Necator ameri-canus hookworm larval antigen Na-ASP-2 in Pichia pastoris and purification of the recombinant protein for use in human clinical trials. Vaccine 2005; 23:4754-64; PMID:16054275; http://dx.doi.org/10.1016/j.vac-cine.2005.04.040.
31. Lee JC, Timasheff SN. The stabilization of pro-teins by sucrose. J Biol Chem 1981; 256:7193-201; PMID:7251592.
32. Kamerzell TJ, Esfandiary R, Joshi SB, Middaugh CR, Volkin DB. Protein-excipient interactions: mecha-nisms and biophysical characterization applied to protein formulation development. Adv Drug Deliv Rev 2011; 63:1118-59; PMID:21855584; http://dx.doi.org/10.1016/j.addr.2011.07.006.
33. Kendrick BS, Chang BS, Arakawa T, Peterson B, Randolph TW, Manning MC, et al. Preferential exclu-sion of sucrose from recombinant interleukin-1 recep-tor antagonist: role in restricted conformational mobil-ity and compaction of native state. Proc Natl Acad Sci USA 1997; 94:11917-22; PMID:9342337; http://dx.doi.org/10.1073/pnas.94.22.11917.
34. Joshi S, Bhambhani A, Zeng Y, Middaugh CR. An Empirical Phase Diagram/High Throughput Screening Approach to the Characterization and Formulation of Biopharmaceuticals in “Formulation and Process Development Strategies for Manufacturing Biopharmaceuticals”, Ed. F.a.H. Jameel 2010: Wiley & Sons publication.
35. Maddux NR, Joshi SB, Volkin DB, Ralston JP, Middaugh CR. Multidimensional methods for the formulation of biopharmaceuticals and vac-cines. [published online]. J Pharm Sci 2011; In press; PMID:21647886; http://dx.doi.org/10.1002/jps.22618.
19. Asojo OA, Homma K, Sedlacek M, Ngamelue M, Goud GN, Zhan B, et al. X-ray structures of Na-GST-1 and Na-GST-2 two glutathione S-transferase from the human hookworm Necator americanus. BMC Struct Biol 2007; 7:42; PMID:17594497; http://dx.doi.org/10.1186/1472-6807-7-42.
20. Zhan B, Perally S, Brophy PM, Xue J, Goud G, Liu S, et al. Molecular cloning, biochemical characterization and partial protective immunity of the heme-binding glutathione S-transferases from the human hookworm Necator americanus. Infect Immun 2010; 78:1552-63; PMID:20145100; http://dx.doi.org/10.1128/IAI.00848-09.
21. Kueltzo LA, Ersoy B, Ralston JP, Middaugh CR. Derivative absorbance spectroscopy and protein phase diagrams as tools for comprehensive protein characterization: a bGCSF case study. J Pharm Sci 2003; 92:1805-20; PMID:12949999; http://dx.doi.org/10.1002/jps.10439.
22. Salnikova MS, Joshi SB, Rytting JH, Warny M, Middaugh CR. Physical characterization of clostridium difficile toxins and toxoids: effect of the formalde-hyde crosslinking on thermal stability. J Pharm Sci 2008; 97:3735-52; PMID:18257030; http://dx.doi.org/10.1002/jps.21261.
23. Peek LJ, Brandau DT, Jones LS, Joshi SB, Middaugh CR. A systematic approach to stabilizing EBA-175 RII-NG for use as a malaria vaccine. Vaccine 2006; 24:5839-51; PMID:16735084; http://dx.doi.org/10.1016/j.vaccine.2006.04.067.
24. Jiang G, Joshi SB, Peek LJ, Brandau DT, Huang J, Ferriter MS, et al. Anthrax vaccine powder for-mulations for nasal mucosal delivery. J Pharm Sci 2006; 95:80-96; PMID:16315230; http://dx.doi.org/10.1002/jps.20484.
25. Ausar SF, Foubert TR, Hudson MH, Vedvick TS, Middaugh CR. Conformational stability and disas-sembly of Norwalk virus-like particles. Effect of pH and temperature. J Biol Chem 2006; 281:19478-88; PMID:16675449; http://dx.doi.org/10.1074/jbc.M603313200.
26. Ausar SF, Rexroad J, Frolov VG, Look JL, Konar N, Middaugh CR. Analysis of the thermal and pH stability of human respiratory syncytial virus. Mol Pharm 2005; 2:491-9; PMID:16323956; http://dx.doi.org/10.1021/mp0500465.
27. Rexroad J, Evans RK, Middaugh CR. Effect of pH and ionic strength on the physical stability of adenovirus type 5. J Pharm Sci 2006; 95:237-47; PMID:16372304; http://dx.doi.org/10.1002/jps.20496.
9. Chan MS. The global burden of intestinal nema-tode infections—fifty years on. Parasitol Today 1997; 13:438-43; PMID:15275146; http://dx.doi.org/10.1016/S0169-4758(97)01144-7.
10. Bleakley H. Disease and development: evidence from hookworm eradication in the American South. Q J Econ 2007; 122:73-117; http://dx.doi.org/10.1162/qjec.121.1.73.
11. Brooker S, Akhwale W, Pullan R, Estambale B, Clarke SE, Snow RW, et al. Epidemiology of plasmodium-helminth co-infection in Africa: populations at risk, potential impact on anemia and prospects for combin-ing control. Am J Trop Med Hyg 2007; 77:88-98; PMID:18165479.
12. Hotez PJ, Mistry N, Rubinstein J, Sachs JD. Integrating neglected tropical diseases into AIDS, tuberculosis and malaria control. N Engl J Med 2011; 364:2086-9; PMID:21631320; http://dx.doi.org/10.1056/NEJMp1014637.
13. Hotez PJ, Bethony JM, Oliveira SC, Brindley PJ, Loukas A. Multivalent anthelminthic vaccine to prevent hookworm and schistosomiasis. Expert Rev Vaccines 2008; 7:745-52; PMID:18665774; http://dx.doi.org/10.1586/14760584.7.6.745.
14. Hotez PJ, Bethony JM, Diemert DJ, Pearson M, Loukas A. Developing vaccines to combat hook-worm infection and intestinal schistosomiasis. Nat Rev Microbiol 2010; 8:814-26; PMID:20948553; http://dx.doi.org/10.1038/nrmicro2438.
15. Hotez PJ. Mass drug administration and integrat-ed control for the world’s high-prevalence neglected tropical diseases. Clin Pharmacol Ther 2009; 85:659-64; PMID:19322166; http://dx.doi.org/10.1038/clpt.2009.16.
16. Keiser J, Utzinger J. Efficacy of current drugs against soil-transmitted helminth infections: systematic review and meta-analysis. JAMA 2008; 299:1937-48; PMID:18430913; http://dx.doi.org/10.1001/jama.299.16.1937.
17. Albonico M, Bickle Q, Ramsan M, Montresor A, Savioli L, Taylor M. Efficacy of mebendazole and levamisole alone or in combination against intestinal nematode infections after repeated targeted mebenda-zole treatment in Zanzibar. Bull World Health Organ 2003; 81:343-52; PMID:12856052.
18. Albonico M, Smith PG, Ercole E, Hall A, Chwaya HM, Alawi KS, et al. Rate of reinfection with intestinal nematodes after treatment of children with mebenda-zole or albendazole in a highly endemic area. Trans R Soc Trop Med Hyg 1995; 89:538-41; PMID:8560535; http://dx.doi.org/10.1016/0035-9203(95)90101-9.
Related Documents











