Advanced glycation end products inhibitors from Alpinia zerumbet rhizomes Jamnian Chompoo a , Atul Upadhyay a , Wataru Kishimoto b , Tadahiro Makise c , Shinkichi Tawata b,⇑ a Department of Bioscience and Biotechnology, The United Graduate School of Agricultural Science, Kagoshima University, Kagoshima 890-0065, Japan b Department of Bioscience and Biotechnology, Faculty of Agriculture, University of the Ryukyus, Senbaru 1, Nishihara-cho, Okinawa 903-0219, Japan c Life Up Clinic, 750 Minamiuehara, Nakagusuku, Okinawa 901-2424, Japan article info Article history: Received 10 December 2010 Received in revised form 7 April 2011 Accepted 12 April 2011 Available online 20 April 2011 Keywords: Alpinia zerumbet Diabetes Advanced glycation end products (AGEs) 5,6-Dehydrokawain Dihydro-5,6-dehydrokawain 8(17),12-Labdadiene-15,16-dial abstract Advanced glycation end products (AGEs) are major factors responsible for the complication of diabetes. The present study was carried out to investigate the inhibitory activities on fructosamine adduct and a- dicarbonyl formations by hexane extracts of various parts of Alpinia zerumbet. Furthermore, we isolated two previously known compounds, namely 5,6-dehydrokawain (DK) and dihydro-5,6-dehydrokawain (DDK). 8(17),12-Labdadiene-15,16-dial (labdadiene) was isolated for the first time from the rhizome of A. zerumbet. The results showed that labdadiene (IC 50 = 51.06 lg/mL) had similar activity to rutin and quercetin against fructosamine adduct. The inhibition of a-dicarbonyl compounds formation by labdadi- ene was significantly higher than that of DK and DDK. Our results indicate that labdadiene is a potent antiglycation agent which was found to inhibit AGEs formation in three different steps in the pathway. These data indicate that labdadiene could be used to prevent glycation-associated complications in diabetes. Ó 2011 Published by Elsevier Ltd. 1. Introduction Advanced glycation end products (AGEs) are a complex and het- erogeneous group of compounds that have been implicated in dia- betes-related complications. The AGEs are closely associated with hyperglycaemia and their patho-biochemistry could explain many of the changes observed in diabetes-related complications (Singh, Barden, Mori, & Beilin, 2001). AGEs are the products of non- enzymatic glycation and oxidation of proteins and lipids. Glycation adducts of proteins are formed when proteins react with glucose- reactive a-oxoaldehydes or dicarbonyls, such as glyoxal, methyl- glyoxal and 3-deoxyglucosone (3-DG) (Brownlee, 2001). These adducts have recently been proposed to be formed from all stages of the glycation process, by degradation of glucose or Schiff’s bases in early glycation, or from Amadori products such as fructosamine in the intermediate stages of glycation. Thus, a-dicarbonyl could be considered an important focal point of how glucose can go on to form AGEs by the classical Maillard reaction, using AGEs formation pathway (Singh et al., 2001), as well as from in vivo factors, such as the catabolism of ketone bodies and threonine, and lipid peroxida- tion (Thornalley, Langborg, & Minhas, 1999). Therefore, inhibition of fructosamine adducts and a-dicarbonyls could reduce AGEs formation and hence lessen the chance of diabetic complications. Alpinia zerumbet (family: Zingiberaceae) is a perennial plant growing widely in the subtropical and tropical regions. It is used in traditional medicine for its anti-inflammatory, bacteriostatic and fungistatic properties (Zoghbi, Andrade, & Maia, 1999). Dihy- dro-5,6-dehydrokawain (DDK) and 5,6-dehydrokawain (DK) have been detected in the rhizomes of Alpinia speciosa (Itokawa, Morita, & Mihashi, 1981). Furthermore, DK and DDK are reported to inhibit the aggregation of ATP release from rabbit platelets (Teng et al., 1990). DK and DDK are described to have anti-ulcerogenic and antithrombotic activities (Mpalantinos, Moura, Parente, & Kuster, 1998). The inhibitory effects of DK on human platelet aggregation, anti-inflammatory and cancer chemopreventive therapeutic prop- erties are reported (Jantan et al., 2008). Labdadiene, isolated from the Zingiberaceae (Sirat, Masri, & Rahman, 1994), was traditionally used as a medicine against inflammatory diseases (Kunnumakkara et al., 2008). The cardiovascular effects induced by labdadiene were evaluated in male Wistar rats (Oliveira et al., 2006). Moreover, labdadiene has also been reported to inhibit lipid peroxidation, cyclooxygenase enzymes and human tumour cell proliferation (Liu & Nair, 2011). Our laboratory has reported DDK and phenolic compounds and their antioxidant activities in leaves and rhizomes of A. zerumbet (Elzaawely, Xuan, & Tawata, 2007). Recently we have reported the HIV-1 integrase and neuraminidase inhibitory proper- ties of A. zerumbet and have found that DK and DDK have signifi- cant bioactivities (Upadhyay, Chompoo, Kishimoto, Makise, & Tawata, 2011). In this study, we primarily investigated the inhibitory proper- ties on AGEs formation of hexane extracts from six different parts of A. zerumbet. Since we had previously found high bioactivities in 0308-8146/$ - see front matter Ó 2011 Published by Elsevier Ltd. doi:10.1016/j.foodchem.2011.04.034 ⇑ Corresponding author. Tel.: +81 98 895 8803; fax: +81 98 895 8734. E-mail addresses: [email protected] (J. Chompoo), [email protected] (A. Upadhyay), [email protected] (W. Kishimoto), [email protected] (T. Makise), [email protected] (S. Tawata). Food Chemistry 129 (2011) 709–715 Contents lists available at ScienceDirect Food Chemistry journal homepage: www.elsevier.com/locate/foodchem

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Food Chemistry 129 (2011) 709–715
Contents lists available at ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Advanced glycation end products inhibitors from Alpinia zerumbet rhizomes
Jamnian Chompoo a, Atul Upadhyay a, Wataru Kishimoto b, Tadahiro Makise c, Shinkichi Tawata b,⇑a Department of Bioscience and Biotechnology, The United Graduate School of Agricultural Science, Kagoshima University, Kagoshima 890-0065, Japanb Department of Bioscience and Biotechnology, Faculty of Agriculture, University of the Ryukyus, Senbaru 1, Nishihara-cho, Okinawa 903-0219, Japanc Life Up Clinic, 750 Minamiuehara, Nakagusuku, Okinawa 901-2424, Japan
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 December 2010Received in revised form 7 April 2011Accepted 12 April 2011Available online 20 April 2011
Keywords:Alpinia zerumbetDiabetesAdvanced glycation end products (AGEs)5,6-DehydrokawainDihydro-5,6-dehydrokawain8(17),12-Labdadiene-15,16-dial
0308-8146/$ - see front matter � 2011 Published bydoi:10.1016/j.foodchem.2011.04.034
⇑ Corresponding author. Tel.: +81 98 895 8803; faxE-mail addresses: [email protected] (J. Chom
Upadhyay), [email protected] (W. KishimMakise), [email protected] (S. Tawata).
Advanced glycation end products (AGEs) are major factors responsible for the complication of diabetes.The present study was carried out to investigate the inhibitory activities on fructosamine adduct and a-dicarbonyl formations by hexane extracts of various parts of Alpinia zerumbet. Furthermore, we isolatedtwo previously known compounds, namely 5,6-dehydrokawain (DK) and dihydro-5,6-dehydrokawain(DDK). 8(17),12-Labdadiene-15,16-dial (labdadiene) was isolated for the first time from the rhizome ofA. zerumbet. The results showed that labdadiene (IC50 = 51.06 lg/mL) had similar activity to rutin andquercetin against fructosamine adduct. The inhibition of a-dicarbonyl compounds formation by labdadi-ene was significantly higher than that of DK and DDK. Our results indicate that labdadiene is a potentantiglycation agent which was found to inhibit AGEs formation in three different steps in the pathway.These data indicate that labdadiene could be used to prevent glycation-associated complications indiabetes.
� 2011 Published by Elsevier Ltd.
1. Introduction Alpinia zerumbet (family: Zingiberaceae) is a perennial plant
Advanced glycation end products (AGEs) are a complex and het-erogeneous group of compounds that have been implicated in dia-betes-related complications. The AGEs are closely associated withhyperglycaemia and their patho-biochemistry could explain manyof the changes observed in diabetes-related complications (Singh,Barden, Mori, & Beilin, 2001). AGEs are the products of non-enzymatic glycation and oxidation of proteins and lipids. Glycationadducts of proteins are formed when proteins react with glucose-reactive a-oxoaldehydes or dicarbonyls, such as glyoxal, methyl-glyoxal and 3-deoxyglucosone (3-DG) (Brownlee, 2001). Theseadducts have recently been proposed to be formed from all stagesof the glycation process, by degradation of glucose or Schiff’s basesin early glycation, or from Amadori products such as fructosaminein the intermediate stages of glycation. Thus, a-dicarbonyl could beconsidered an important focal point of how glucose can go on toform AGEs by the classical Maillard reaction, using AGEs formationpathway (Singh et al., 2001), as well as from in vivo factors, such asthe catabolism of ketone bodies and threonine, and lipid peroxida-tion (Thornalley, Langborg, & Minhas, 1999). Therefore, inhibitionof fructosamine adducts and a-dicarbonyls could reduce AGEsformation and hence lessen the chance of diabetic complications.
Elsevier Ltd.
: +81 98 895 8734.poo), [email protected] (A.
oto), [email protected] (T.
growing widely in the subtropical and tropical regions. It is usedin traditional medicine for its anti-inflammatory, bacteriostaticand fungistatic properties (Zoghbi, Andrade, & Maia, 1999). Dihy-dro-5,6-dehydrokawain (DDK) and 5,6-dehydrokawain (DK) havebeen detected in the rhizomes of Alpinia speciosa (Itokawa, Morita,& Mihashi, 1981). Furthermore, DK and DDK are reported to inhibitthe aggregation of ATP release from rabbit platelets (Teng et al.,1990). DK and DDK are described to have anti-ulcerogenic andantithrombotic activities (Mpalantinos, Moura, Parente, & Kuster,1998). The inhibitory effects of DK on human platelet aggregation,anti-inflammatory and cancer chemopreventive therapeutic prop-erties are reported (Jantan et al., 2008). Labdadiene, isolated fromthe Zingiberaceae (Sirat, Masri, & Rahman, 1994), was traditionallyused as a medicine against inflammatory diseases (Kunnumakkaraet al., 2008). The cardiovascular effects induced by labdadiene wereevaluated in male Wistar rats (Oliveira et al., 2006). Moreover,labdadiene has also been reported to inhibit lipid peroxidation,cyclooxygenase enzymes and human tumour cell proliferation(Liu & Nair, 2011). Our laboratory has reported DDK and phenoliccompounds and their antioxidant activities in leaves and rhizomesof A. zerumbet (Elzaawely, Xuan, & Tawata, 2007). Recently we havereported the HIV-1 integrase and neuraminidase inhibitory proper-ties of A. zerumbet and have found that DK and DDK have signifi-cant bioactivities (Upadhyay, Chompoo, Kishimoto, Makise, &Tawata, 2011).
In this study, we primarily investigated the inhibitory proper-ties on AGEs formation of hexane extracts from six different partsof A. zerumbet. Since we had previously found high bioactivities in

710 J. Chompoo et al. / Food Chemistry 129 (2011) 709–715
DK and DDK (Upadhyay et al., 2011), we focused on thesecompounds for AGEs inhibitory properties. Furthermore, we iso-lated labdadiene for the first time from A. zerumbet and investi-gated its activities. We examined the inhibition of fructosamineadducts and a-dicarbonyl formation in order to check its efficacyagainst AGEs. To the best of our knowledge, this is the maidenreport on AGEs inhibition from A. zerumbet and isolatedcompounds.
2. Materials and methods
2.1. Chemicals and reagents
Bovine serum albumin (BSA), copper(II) sulphate, D(+)-glucose,2,4-dinitrophenylhydrazine (DNPH), heptafluorobutyric acid(HFBA), 40% glyoxal solution, nitro blue tetrazolium chloride(NBT), sodium azide, and guanidine were obtained from Wako PureChemical Industries Ltd. (Osaka, Japan). Trichloroacetic acid (TCA)was purchased from Sigma–Aldrich (St Louis, MO). Trim-ethylaminoacetohydrazide chloride (Girard’s reagent T) wasbought from Tokyo Chemical Industry Co. Ltd. (Tokyo, Japan). Alka-line PBS (137 mM NaCl, 8.1 mM Na2HPO4, 2.68 mM KCl, 1.47 mMKH2PO4) was adjusted to pH 10 with NaOH.
2.2. Preparation of crude extracts and isolated compounds
The parts of A. zerumbet (including rhizomes, stems, leaves,flowers, pericarps and seeds) were collected at University of theRyukyus, Okinawa, Japan. The samples were oven dried and groundto fine powder. Fifteen grams each dried samples were soaked in400 mL of hexane for 24 h. The extracts were filtered through filterpaper and evaporated under vacuum to remove the solvent. Thecrude extracts were kept at 4 �C for experiments. DK and DDK wereisolated from the rhizomes as reported previously (Upadhyay et al.,2011). Labdadiene was isolated from the hexane extract of the rhi-zomes. Briefly, the dried rhizomes were extracted with hexane for48 h and the crude hexane extract was filtered and dried undervacuum at 40 �C. The crude extract was further separated using sil-ica gel column chromatography using hexane:acetone (9:1). Thealiquots from column chromatography were separated using Dia-ion HP-20 resin column (Mitsubishi Chemical Co., Tokyo, Japan)with increasing methanol concentration from 50% to 100%, and fur-ther purified by HPLC to get labdadiene. Labdadiene was collectedat 235 nm using a COSMOSIL HPLC column (Vision WorkstationBiocad Family, USA) (10 mm i.d. � 250 mm; 5 lm particle size).The mobile phase was water (solvent A) and methanol (solventB) at flow rate of 2 mL/min. The gradient elution was performedas follows: 0–10 min, 80% B isocratic; 10–20 min, linear gradient80–100% B, 20–40 min, 100% B isocratic. The chemical structuresof isolated compounds were identified using GC–MS and NMR(Fig. 1).
The 1H NMR (400 MHz) and 13C NMR (100 MHz) spectra wererecorded on a Bruker Biospin GmbH (Rheinstetten, Germany) inCDCl3. Chemical shifts are expressed in parts per million (d) rela-tive to TMS. 2D NMR experiments (H, C-COSY, HMQC, HMBC) wereobtained using standard pulse sequences on TopSpin 2.1 programVersion 2.1.0. Electron ionization mass spectrometry.
2.3. Spectral details for labdadiene
Electron ionisation mass spectrometry (EIMS): m/z (rel. int.);302 (20), 137 (100), 123 (50), 109 (35), 95 (73), 81 (70), 69 (55),55 (48), 41 (50). 1H NMR (CDCl3): d 0.73, 0.82 and 0.89 (s, each3H, CH3, 18, 19, 20), 1.04–2.52 (m, 14H, CH2, CH, 1, 2, 3, 5, 6, 7, 9,10, 11), 3.44 (s, 2H, CH2, 14), 4.37 (s, 1H, CH2, 17), 4.86 (s, 1H,
CH2, 17), 6.76 (t, 1H, CH, 12), 9.40 (s, 1H, CHO, 15) and 9.63(s, 1H, CHO, 16); 13C NMR (CDCl3): d 14.4 (C-20), 19.3 (C-2), 21.7(C-19), 24.1 (C-6), 33.6 (C-4), 37.9 (C-14), 39.3 (C-1), 39.4 (C-7),39.6 (C-10), 42.0 (C-3), 55.4 (C-5), 56.5 (C-9), 107.8 (C-17), 134.9(C-13), 148.0 (C-8), 159.9 (C-12), 193.5 (C-16) and 197.3 (C-15).
2.4. Quantification of DK, DDK and labdadiene using HPLC
The isolated compounds were measured using a ShimadzuHPLC (SCL-10A vp, Shimadzu Co., Kyoto, Japan) coupled with aUV–Vis detector (SPD-20A, Shimadzu). Separations were achievedon a TSKgel ODS-100Z column (15 cm � 4.6 mm i.d.; 5 lm particlesize; Agilent Technologies, Santa Clara, CA). The mobile phase waswater with 1% acetic acid (v/v) (solvent A) and methanol with 1%acetic acid (solvent B) at a flow rate of 0.8 mL/min. DK and DDKwere measured at 280 nm. The gradient elution was performedas follows: 0–10 min, 50% B isocratic; 10–20 min, linear gradient50–100% B and 20–26 min, 100–50% B. The gradient elution forlabdadiene was performed as follows: 0–10 min, 80% B isocratic;10–20 min, linear gradient 80–100% B and 20–35 min, 100% Band monitored at 235 nm. A 5-lL methanolic solution of crude ex-tracts at 1 mg/mL was used for the identification of the compounds(Fig. 1). The quantification of each compound was determinedbased on peak area measurements.
2.5. Preparation of AGEs–BSA
Each sample was dissolved in DMSO. The reaction mixturescontaining 400 lg bovine serum albumin, 200 mM glucose, 10 lLof test sample and 50 mM phosphate buffer (pH 7.4) to a total vol-ume of 500 lL, were incubated at 60 �C for 30 h. The blank, whichcontained the protein and glucose but no test sample, was kept at4 �C until measurement (Matsuura et al., 2002).
2.6. Measurement of glycation of BSA
AGEs–BSA aliquots or glycated materials of 100 lL were trans-ferred to 1.5-mL plastic tubes, and 10 lL of 100% (w/v) TCA wereadded to each tube. The supernatant containing glucose, inhibitorand the interfering substances was removed after centrifugation(15,000 rpm, 4 �C, 4 min) (Wu, Hsieh, Wang, & Chen, 2009). Then,the precipitate of AGEs–BSA was dissolved with 100 lL of PBS toserve for screening. The comparison of fluorescence spectrum(excitation 360 nm) and the change in fluorescence intensity (exci-tation 360 nm, and emission 450 nm) based on AGEs was moni-tored using a spectrofluorometer (GENios, Tecan Group Ltd.,Männedorf, Switzerland). Percent inhibition of AGEs formation byeach extract or compound was calculated using the followingequation:
Inhibitory activities ð%Þ ¼ ½1� ðF i=F0Þ� � 100;
where Fi is fluorescence of solution with inhibitors and F0 is fluores-cence of the solution without inhibitors. The concentration requiredfor 50% inhibition (IC50) of the fluorescence intensity was deter-mined graphically.
2.7. Determination of fructosamine adduct
The fructosamine adduct was determined by using NBT assay asdescribed previously (Baker, Zyzak, Thorpe, & Baynes, 1994). Ali-quots of glycated materials (40 lL) were added to the reactionmixture which contained 160 lL of NBT reagent (300 lM) insodium carbonate buffer (100 mM, pH 10.35), incubated at roomtemperature for 30 min, and absorbance measured at 530 nmusing a microplate reader (Benchmark microplate reader;
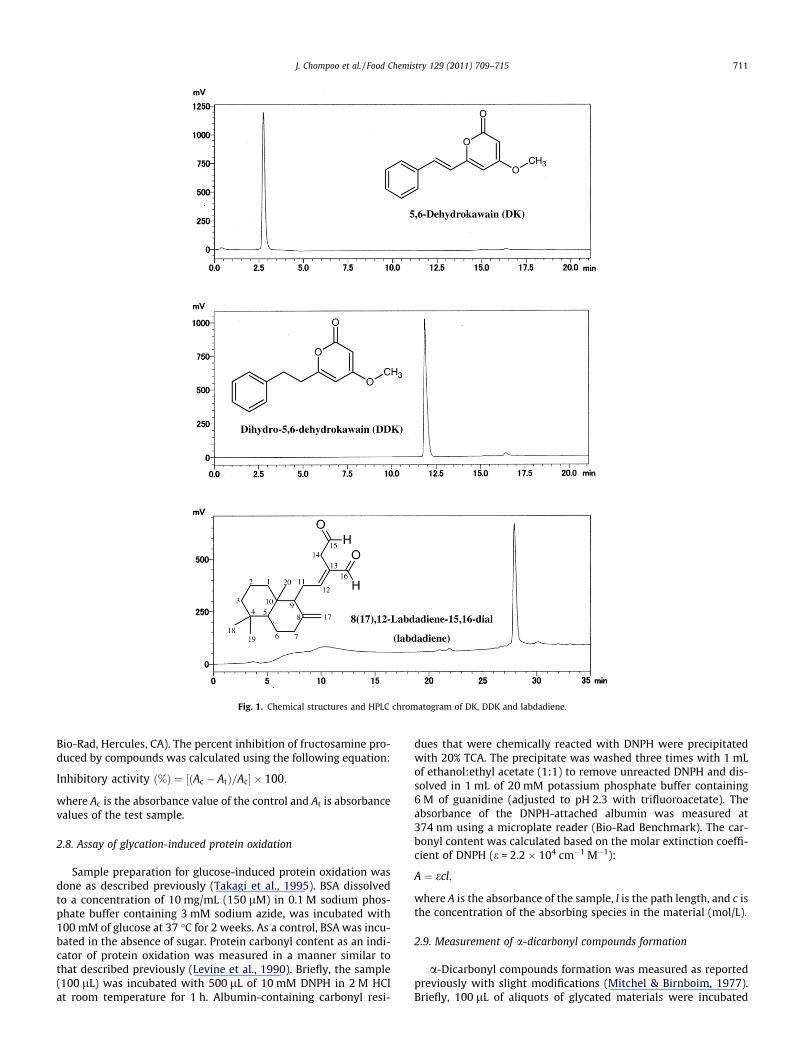
Fig. 1. Chemical structures and HPLC chromatogram of DK, DDK and labdadiene.
J. Chompoo et al. / Food Chemistry 129 (2011) 709–715 711
Bio-Rad, Hercules, CA). The percent inhibition of fructosamine pro-duced by compounds was calculated using the following equation:
Inhibitory activity ð%Þ ¼ ½ðAc � AtÞ=Ac� � 100;
where Ac is the absorbance value of the control and At is absorbancevalues of the test sample.
2.8. Assay of glycation-induced protein oxidation
Sample preparation for glucose-induced protein oxidation wasdone as described previously (Takagi et al., 1995). BSA dissolvedto a concentration of 10 mg/mL (150 lM) in 0.1 M sodium phos-phate buffer containing 3 mM sodium azide, was incubated with100 mM of glucose at 37 �C for 2 weeks. As a control, BSA was incu-bated in the absence of sugar. Protein carbonyl content as an indi-cator of protein oxidation was measured in a manner similar tothat described previously (Levine et al., 1990). Briefly, the sample(100 lL) was incubated with 500 lL of 10 mM DNPH in 2 M HClat room temperature for 1 h. Albumin-containing carbonyl resi-
dues that were chemically reacted with DNPH were precipitatedwith 20% TCA. The precipitate was washed three times with 1 mLof ethanol:ethyl acetate (1:1) to remove unreacted DNPH and dis-solved in 1 mL of 20 mM potassium phosphate buffer containing6 M of guanidine (adjusted to pH 2.3 with trifluoroacetate). Theabsorbance of the DNPH-attached albumin was measured at374 nm using a microplate reader (Bio-Rad Benchmark). The car-bonyl content was calculated based on the molar extinction coeffi-cient of DNPH (e = 2.2 � 104 cm�1 M�1):
A ¼ ecl;
where A is the absorbance of the sample, l is the path length, and c isthe concentration of the absorbing species in the material (mol/L).
2.9. Measurement of a-dicarbonyl compounds formation
a-Dicarbonyl compounds formation was measured as reportedpreviously with slight modifications (Mitchel & Birnboim, 1977).Briefly, 100 lL of aliquots of glycated materials were incubated

712 J. Chompoo et al. / Food Chemistry 129 (2011) 709–715
at room temperature for 1 h with a reaction mixture containing50 lL of Girard-T stock solution (500 mM) and 850 lL of sodiumformate (500 mM, pH 2.9). Absorbance was measured at 290 nmusing a spectrophotometer (UV Mini 1240, Shimadzu, Kyoto, Ja-pan), and glyoxal contents were calculated using a standard curvefor glyoxal. A calibration curve was prepared using 40% glyoxalsolution in a similar way.
2.10. HPLC analyses of glyoxal content
Aliquots of glucose reaction (1 mL) were initially prepared forHPLC analysis by adding 300 lL of 2 M HCl (pH 3.0) and 50 lL ofGirard-T stock solution and then incubating at room temperaturefor 24 h to allow sufficient time for product formation from slowreacting dicarbonyl compounds, such as glyoxal. Aliquots, 2 lL,were analysed by reversed-phase HPLC, using TSKgel-NH2-100 col-umn (Tosoh Bioscience LLC, USA), 5 cm � 2.0 mm i.d., 3 lm particlesize, with absorbance detection at 280 nm. An isocratic solution of14% acetonitrile in water containing 0.25% HFBA for 10 min wasused as eluting solvent (Well-Knecht, Zyzak, Litchfield, Thorpe, &Baynes, 1995). The amounts of glyoxal content in different samplesof varying concentrations were calculated from the standard curve(y = 2,525,707x + 125,777; r2 = 0.993). The inhibition percent is cal-culated as
Inhibition activity ð%Þ ¼ ½ðAc � AsÞ=Gc� � 100;
where Ac is absorbance value of control and As is absorbance valueof test samples. The IC50 is the concentration of sample at whichglyoxal formation was reduced by 50%.
2.11. Statistical analysis
The data were analysed by one-way ANOVA. Upon significantdifference, means were separated using Tukey HSD range test atp = 0.01 with three replications. In some cases, only means andstandard error of the sample means are presented. All statisticalanalyses were performed using SPSS Version 16.0 for WindowsVista (SPSS Inc., Chicago, IL).
Table 1Amount of DK, DDK and labdadiene in hexane extracts from different parts of A.zerumbet.
Sample Amount of compound (mg/g hexane extracts)
DK DDK Labdadiene
Rhizomes 3.13 ± 0.02 aA 5.41 ± 0.02 b 3.97 ± 0.02 aStems 2.08 ± 0.07 b 3.70 ± 0.01 c 0.00 ± 0.00 dLeaves 1.67 ± 0.06 c 3.38 ± 0.03 d 0.00 ± 0.00 dFlowers 2.22 ± 0.01 b 6.08 ± 0.06 a 0.00 ± 0.00 dPericarps 1.58 ± 0.07 c 0.13 ± 0.01 e 0.35 ± 0.01 cSeeds 0.11 ± 0.02 d 0.22 ± 0.02 e 2.91 ± 0.01 b
A The data represent the means ± SD of three determinations. Values with thesame letter in one column are not significantly different (p = 0.01) from each other.
Table 2Effect of hexane extracts from various parts of A. zerumbet on advanced glycation end pro
Sample Inhibition 50% (lg/mL)
Glycation of BSA Fructosamine adducts a-Dicarbonyl form
Rhizomes 164 ± 8.11 aA 278 ± 2.86 a 176 ± 0.51 bStems 275 ± 7.25 d 353 ± 4.86 c 434 ± 1.86 cLeaves 266 ± 1.38 cd 294 ± 5.06 ab 858 ± 4.22 dFlowers 231 ± 1.25 b 305 ± 3.05 b 90 ± 5.79 aPericarps 791 ± 7.80 e 848 ± 0.83 e 459 ± 0.08 cSeeds 245 ± 5.92 bc 475 ± 6.45 d 431 ± 3.19 c
A The data represent the means ± SD of three determinations. Values with the same le
3. Results
3.1. Amounts of DK, DDK and labdadiene
The amounts of DK, DDK and labdadiene in the crude extractsfrom different parts of A. zerumbet are shown in Table 1. DK andDDK were found in all parts of the plant. The rhizomes extract con-tained higher amount of DK (3.13 mg/g hexane extract), however,flowers extract contained a higher quantity of DDK (6.08 mg/g hex-ane extract). The hexane extract of rhizomes contained higheramount of labdadiene (3.97 mg/g hexane extracts) than seedsand pericarp (0.35 and 2.91 mg/g hexane extracts, respectively).However, the hexane extracts of leaves and flowers did not containlabdadiene.
3.2. Inhibitory effect of crude extracts and isolated compounds onglycation of BSA
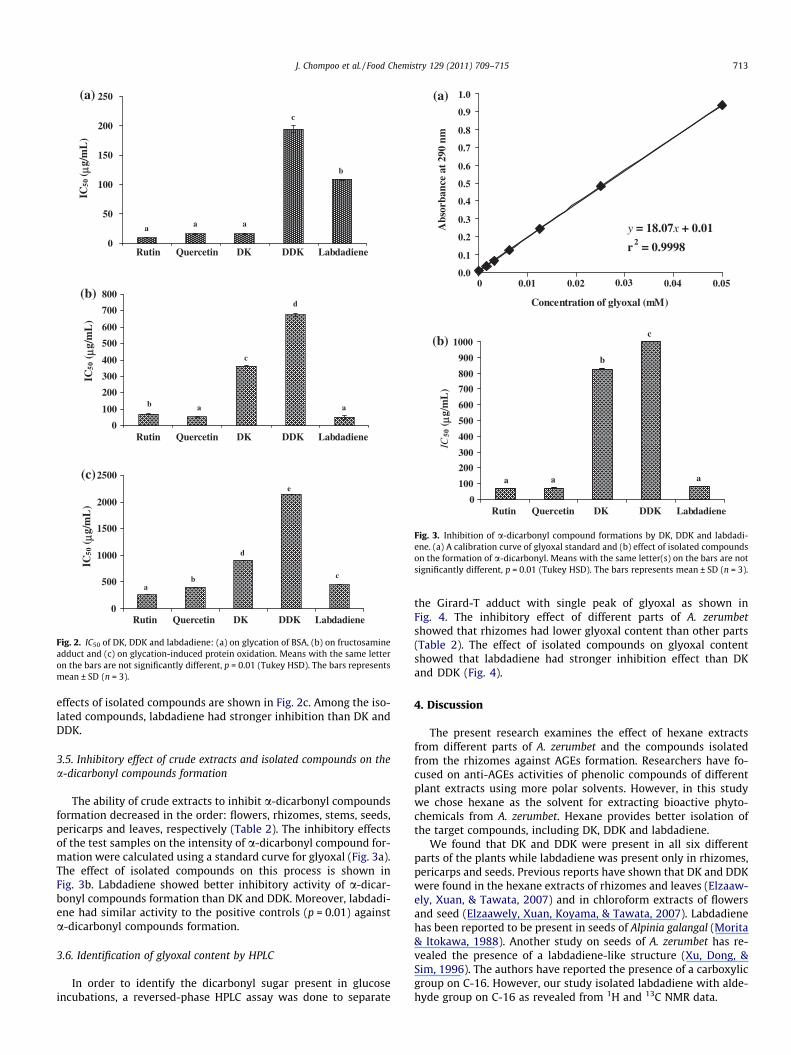
The ability of hexane extracts from various parts of A. zerumbetto inhibit AGEs formation was evaluated using the BSA–glucose as-say, in which bovine serum albumin served as the model proteinand glucose as the glycating agent. The results showed that rhi-zomes had higher inhibitory activity than the other parts (Table 2).On carrying out inhibition with isolated compounds, DK(IC50 = 15.9 lg/mL) had stronger inhibitory activity than labdadi-ene and DDK (IC50 = 108 and 194 lg/mL, respectively). On statisti-cal analysis of the results shown in Fig. 2a, it was found that theactivity of DK was not significantly different from that of the twopositive controls rutin and quercetin (IC50 = 9.65 and 15.9 lg/mL,respectively) (p = 0.01).
3.3. Inhibitory effect of crude extracts and isolated compounds on thefructosamine adduct
The results of inhibitory effect of hexane extracts on fructos-amine adduct showed that the IC50 of crude extracts from rhizomesand leaves on fructosamine adduct were stronger than stems, flow-ers, pericarps and seeds (Table 2). Labdadiene (IC50 = 51.1 lg/mL)had higher inhibitory effects than DK and DDK (IC50 = 360 and673 lg/mL, respectively) (Fig. 2b). Moreover, labdadiene had simi-lar activity against fructosamine adduct formation as positivecontrols.
3.4. Inhibitory effect of crude extracts and isolated compounds onglycation-induced protein oxidation
Oxidative modifications of BSA preparations were demon-strated using a combination of carbonyl assay; the extent of pro-tein carbonyl formation was determined by DNPH-reagent. Theinhibitory effects of crude extracts are shown in Table 2. The hex-ane extract of flower showed the strongest activity. The inhibitory
ducts.
ation Glycation-induced protein oxidation Glyoxal content by HPLC
1150 ± 6.45 b 110 ± 0.66 a3224 ± 1.83 d 419 ± 0.99 d1392 ± 0.73 c 784 ± 1.25 f1027 ± 1.14 a 285 ± 3.98 b4213 ± 1.87 f 588 ± 8.80 e3721 ± 0.59 e 400 ± 1.59 c
tter in one column are not significantly different (p = 0.01) from each other.
0
500
1000
1500
2000
2500
Rutin Quercetin DK DDK Labdadiene
Rutin Quercetin DK DDK Labdadiene
Rutin Quercetin DK DDK Labdadiene
a b c
d
e
IC50
( µg/
mL
)
0
100
200
300
400
500
600
700
800
b a
c
d
a
IC50
( µg/
mL
)
(b)
(c)
0
50
100
150
200
250
a a a
c
b
IC50
( µg/
mL
)(a)
Fig. 2. IC50 of DK, DDK and labdadiene: (a) on glycation of BSA, (b) on fructosamineadduct and (c) on glycation-induced protein oxidation. Means with the same letteron the bars are not significantly different, p = 0.01 (Tukey HSD). The bars representsmean ± SD (n = 3).
y = 18.07x + 0.01
r2 = 0.9998
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
0 0.01 0.02 0.03 0.04 0.05
Abs
orba
nce
at 2
90 n
m
Concentration of glyoxal (mM)
(a)
0
100
200
300
400
500
600
700
800
900
1000
Rutin Quercetin DK DDK Labdadiene
a
b
a a
IC50
( µg/
mL
)
c(b)
Fig. 3. Inhibition of a-dicarbonyl compound formations by DK, DDK and labdadi-ene. (a) A calibration curve of glyoxal standard and (b) effect of isolated compoundson the formation of a-dicarbonyl. Means with the same letter(s) on the bars are notsignificantly different, p = 0.01 (Tukey HSD). The bars represents mean ± SD (n = 3).
J. Chompoo et al. / Food Chemistry 129 (2011) 709–715 713
effects of isolated compounds are shown in Fig. 2c. Among the iso-lated compounds, labdadiene had stronger inhibition than DK andDDK.
3.5. Inhibitory effect of crude extracts and isolated compounds on thea-dicarbonyl compounds formation
The ability of crude extracts to inhibit a-dicarbonyl compoundsformation decreased in the order: flowers, rhizomes, stems, seeds,pericarps and leaves, respectively (Table 2). The inhibitory effectsof the test samples on the intensity of a-dicarbonyl compound for-mation were calculated using a standard curve for glyoxal (Fig. 3a).The effect of isolated compounds on this process is shown inFig. 3b. Labdadiene showed better inhibitory activity of a-dicar-bonyl compounds formation than DK and DDK. Moreover, labdadi-ene had similar activity to the positive controls (p = 0.01) againsta-dicarbonyl compounds formation.
3.6. Identification of glyoxal content by HPLC
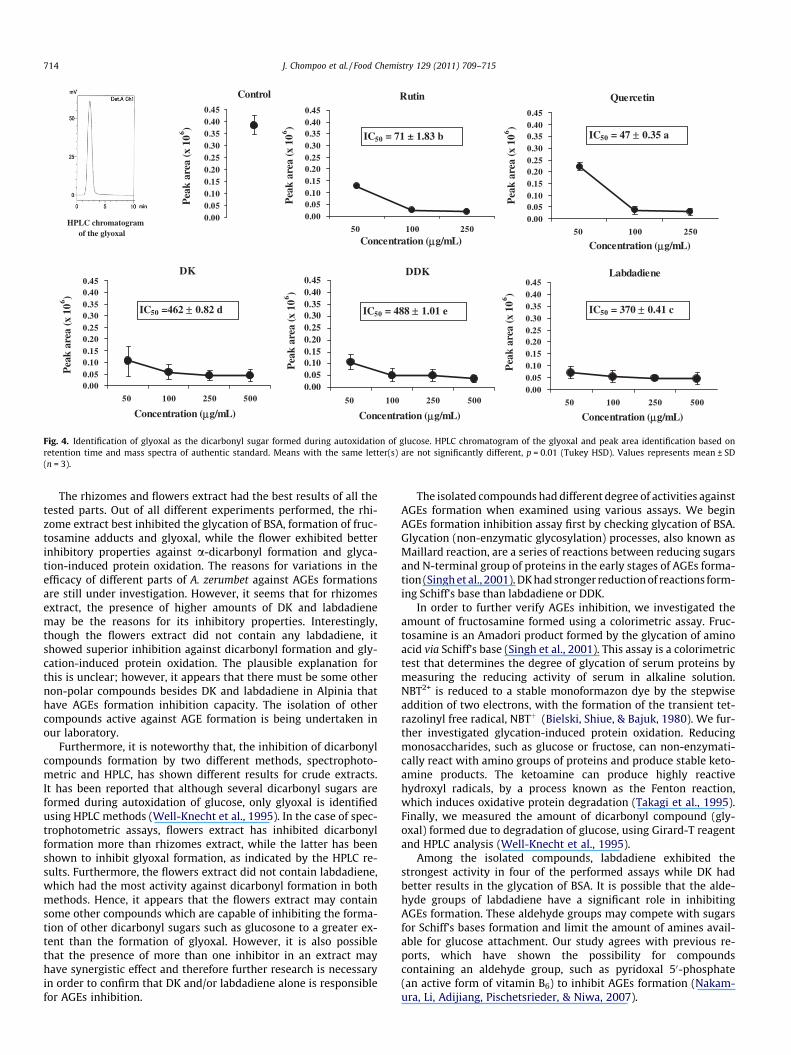
In order to identify the dicarbonyl sugar present in glucoseincubations, a reversed-phase HPLC assay was done to separate
the Girard-T adduct with single peak of glyoxal as shown inFig. 4. The inhibitory effect of different parts of A. zerumbetshowed that rhizomes had lower glyoxal content than other parts(Table 2). The effect of isolated compounds on glyoxal contentshowed that labdadiene had stronger inhibition effect than DKand DDK (Fig. 4).
4. Discussion
The present research examines the effect of hexane extractsfrom different parts of A. zerumbet and the compounds isolatedfrom the rhizomes against AGEs formation. Researchers have fo-cused on anti-AGEs activities of phenolic compounds of differentplant extracts using more polar solvents. However, in this studywe chose hexane as the solvent for extracting bioactive phyto-chemicals from A. zerumbet. Hexane provides better isolation ofthe target compounds, including DK, DDK and labdadiene.
We found that DK and DDK were present in all six differentparts of the plants while labdadiene was present only in rhizomes,pericarps and seeds. Previous reports have shown that DK and DDKwere found in the hexane extracts of rhizomes and leaves (Elzaaw-ely, Xuan, & Tawata, 2007) and in chloroform extracts of flowersand seed (Elzaawely, Xuan, Koyama, & Tawata, 2007). Labdadienehas been reported to be present in seeds of Alpinia galangal (Morita& Itokawa, 1988). Another study on seeds of A. zerumbet has re-vealed the presence of a labdadiene-like structure (Xu, Dong, &Sim, 1996). The authors have reported the presence of a carboxylicgroup on C-16. However, our study isolated labdadiene with alde-hyde group on C-16 as revealed from 1H and 13C NMR data.
DDK
0.000.050.100.150.200.250.300.350.400.45
50 100 250 500
Pea
k ar
ea (x
106 )
Concentration (µg/mL)
DK
0.000.050.100.150.200.250.300.350.400.45
50 100 250 500
Pea
k ar
ea (x
106 )
Concentration (µg/mL)
Labdadiene
0.000.050.100.150.200.250.300.350.400.45
50 100 250 500
Pea
k ar
ea (x
106 )
Concentration (µg/mL)
Quercetin
0.000.050.100.150.200.250.300.350.400.45
50 100 250
Pea
k ar
ea (x
106 )
Concentration (µg/mL)
Rutin
0.000.050.100.150.200.250.300.350.400.45
50 100 250
Pea
k ar
ea (x
106 )
Concentration (µg/mL)
IC50 = 71 ± 1.83 b
IC50 =462 ± 0.82 d IC50 = 488 ± 1.01 e IC50 = 370 ± 0.41 c
IC50 = 47 ± 0.35 a
Control
0.000.050.100.150.200.250.300.350.400.45
Pea
k ar
ea (x
106 )
HPLC chromatogram of the glyoxal
Fig. 4. Identification of glyoxal as the dicarbonyl sugar formed during autoxidation of glucose. HPLC chromatogram of the glyoxal and peak area identification based onretention time and mass spectra of authentic standard. Means with the same letter(s) are not significantly different, p = 0.01 (Tukey HSD). Values represents mean ± SD(n = 3).
714 J. Chompoo et al. / Food Chemistry 129 (2011) 709–715
The rhizomes and flowers extract had the best results of all thetested parts. Out of all different experiments performed, the rhi-zome extract best inhibited the glycation of BSA, formation of fruc-tosamine adducts and glyoxal, while the flower exhibited betterinhibitory properties against a-dicarbonyl formation and glyca-tion-induced protein oxidation. The reasons for variations in theefficacy of different parts of A. zerumbet against AGEs formationsare still under investigation. However, it seems that for rhizomesextract, the presence of higher amounts of DK and labdadienemay be the reasons for its inhibitory properties. Interestingly,though the flowers extract did not contain any labdadiene, itshowed superior inhibition against dicarbonyl formation and gly-cation-induced protein oxidation. The plausible explanation forthis is unclear; however, it appears that there must be some othernon-polar compounds besides DK and labdadiene in Alpinia thathave AGEs formation inhibition capacity. The isolation of othercompounds active against AGE formation is being undertaken inour laboratory.
Furthermore, it is noteworthy that, the inhibition of dicarbonylcompounds formation by two different methods, spectrophoto-metric and HPLC, has shown different results for crude extracts.It has been reported that although several dicarbonyl sugars areformed during autoxidation of glucose, only glyoxal is identifiedusing HPLC methods (Well-Knecht et al., 1995). In the case of spec-trophotometric assays, flowers extract has inhibited dicarbonylformation more than rhizomes extract, while the latter has beenshown to inhibit glyoxal formation, as indicated by the HPLC re-sults. Furthermore, the flowers extract did not contain labdadiene,which had the most activity against dicarbonyl formation in bothmethods. Hence, it appears that the flowers extract may containsome other compounds which are capable of inhibiting the forma-tion of other dicarbonyl sugars such as glucosone to a greater ex-tent than the formation of glyoxal. However, it is also possiblethat the presence of more than one inhibitor in an extract mayhave synergistic effect and therefore further research is necessaryin order to confirm that DK and/or labdadiene alone is responsiblefor AGEs inhibition.
The isolated compounds had different degree of activities againstAGEs formation when examined using various assays. We beginAGEs formation inhibition assay first by checking glycation of BSA.Glycation (non-enzymatic glycosylation) processes, also known asMaillard reaction, are a series of reactions between reducing sugarsand N-terminal group of proteins in the early stages of AGEs forma-tion (Singh et al., 2001). DK had stronger reduction of reactions form-ing Schiff’s base than labdadiene or DDK.
In order to further verify AGEs inhibition, we investigated theamount of fructosamine formed using a colorimetric assay. Fruc-tosamine is an Amadori product formed by the glycation of aminoacid via Schiff’s base (Singh et al., 2001). This assay is a colorimetrictest that determines the degree of glycation of serum proteins bymeasuring the reducing activity of serum in alkaline solution.NBT2+ is reduced to a stable monoformazon dye by the stepwiseaddition of two electrons, with the formation of the transient tet-razolinyl free radical, NBTþ� (Bielski, Shiue, & Bajuk, 1980). We fur-ther investigated glycation-induced protein oxidation. Reducingmonosaccharides, such as glucose or fructose, can non-enzymati-cally react with amino groups of proteins and produce stable keto-amine products. The ketoamine can produce highly reactivehydroxyl radicals, by a process known as the Fenton reaction,which induces oxidative protein degradation (Takagi et al., 1995).Finally, we measured the amount of dicarbonyl compound (gly-oxal) formed due to degradation of glucose, using Girard-T reagentand HPLC analysis (Well-Knecht et al., 1995).
Among the isolated compounds, labdadiene exhibited thestrongest activity in four of the performed assays while DK hadbetter results in the glycation of BSA. It is possible that the alde-hyde groups of labdadiene have a significant role in inhibitingAGEs formation. These aldehyde groups may compete with sugarsfor Schiff’s bases formation and limit the amount of amines avail-able for glucose attachment. Our study agrees with previous re-ports, which have shown the possibility for compoundscontaining an aldehyde group, such as pyridoxal 50-phosphate(an active form of vitamin B6) to inhibit AGEs formation (Nakam-ura, Li, Adijiang, Pischetsrieder, & Niwa, 2007).

J. Chompoo et al. / Food Chemistry 129 (2011) 709–715 715
The fructosamine adduct assay revealed that labdadiene hasstrong activity when compared with the reported positive controlsrutin and quercetin. However, the mechanism of inhibition is likelyto be different. The metal chelation by the vicinal dihydroxylgroups of these flavonoids and/or entrapment of reactive aminogroup supposedly have inhibiting roles (Cervantes-Laurean et al.,2006; Morel et al., 1993).
In all different forms of inhibition of AGEs by the isolated com-pounds, we showed that these compounds have inhibitory activityat different steps of AGEs formation pathways. The isolated com-pounds inhibited glycation of amino group, the first step in thispathway. Furthermore, these compounds suppressed the forma-tion of Amadori products, thereby reducing AGEs formation. Final-ly, these compounds also reduced dicarbonyl compoundsformation, which is the result of glucose degradation or glyca-tion-induced protein oxidation. Therefore, our study revealed thatthe isolated compounds from A. zerumbet had three-fold activitiesin inhibiting AGEs formation.
5. Conclusion
The rhizomes of A. zerumbet have several metabolites that haveAGEs inhibitory properties. We isolated two kawains, DK and DDK,and for the first time isolated labdadiene from the rhizomes of A.zerumbet. Our results showed three-fold inhibitory properties ofthese compounds against AGEs formation. We found that labdadi-ene showed the strongest activity and had significant results whencompared with rutin and quercetin. Thus, labdadiene could beused as a preventive measure against glycation-associated compli-cations in diabetes.
Acknowledgement
The authors would like to thank Tatsunori Higa for his assis-tance with the experiments.
References
Baker, J. R., Zyzak, D. V., Thorpe, S. R., & Baynes, J. W. (1994). Chemistry of thefructosamine assay: D-glucosone is the product of oxidation of Amadoricompounds. Clinical Chemistry, 40, 1950–1955.
Bielski, B. H. J., Shiue, G. G., & Bajuk, S. (1980). Reduction of nitro blue terazolium byCO�2 and O�2 radicals. The Journal of Physical Chemistry, 84, 830–833.
Brownlee, M. (2001). Biochemistry and molecular cell biology of diabeticcomplications. Nature, 414, 813–820.
Cervantes-Laurean, D., Schramm, D. D., Jacobson, E. L., Halaweish, I., Bruckner, G. G.,& Boisonneault, G. A. (2006). Inhibition of advanced glycation end productformation on collagen by rutin and its metabolites. Journal of NutritionalBiochemistry, 17, 531–540.
Elzaawely, A. A., Xuan, T. D., Koyama, H., & Tawata, S. (2007). Antioxidant activityand contents of essential oil and phenolic compounds in flowers and seeds ofAlpinia zerumbet (Pers.) B.L. Burtt & R.M. Sm. Food Chemistry, 104, 1648–1653.
Elzaawely, A. A., Xuan, T. D., & Tawata, S. (2007). Essential oils, kava pyrones andphenolic compounds from leaves and rhizomes of Alpinia zerumbet (Pers.) B.L.Burtt & R.M.Sm. and their antioxidant activity. Food Chemistry, 103, 486–494.
Itokawa, H., Morita, M., & Mihashi, S. (1981). Phenolic compounds from therhizomes of Alpinia speciosa. Phytochemistry, 20, 2503–2506.
Jantan, I., Raweh, S. M., Sirat, H. M., Jamil, S., Yasin, Y. H. M., Jalil, J., et al. (2008).Inhibitory effect of compounds from Zingiberaceae species on human plateletaggregation. Phytomedicine, 15, 306–309.
Kunnumakkara, A. B., Ichikawa, H., Anand, P., Mohankumar, C. J., Hema, P. S., Nair,M. S., et al. (2008). Coronarin D, a labdane diterpene, inhibits both constitutiveand inducible nuclear factor-jB pathway activation, leading to potentiation ofapoptosis, inhibition of invasion, and suppression of osteoclastogenesis.Molecular Cancer Therapeutics, 7, 3306–3317.
Levine, R. L., Garland, D., Oliver, C. N., Amici, A., Climent, I., Lenz, A. G., et al. (1990).Determination of carbonyl content in oxidatively modified proteins. Methods inEnzymology, 186, 464–478.
Liu, Y., & Nair, M. G. (2011). Ladane diterpenes in Curcuma mangga rhizomes inhibitlipid peroxidation, cyclooxygenase enzymes and human tumour cellproliferation. Food Chemistry, 124, 527–532.
Matsuura, N., Aradate, T., Sasaki, C., Kojima, H., Ohara, M., Hasegawa, J., et al. (2002).Screening system for the Maillard reaction inhibitor from natural productextracts. Journal of Health Science, 48, 520–526.
Mitchel, R. E. J., & Birnboim, H. C. (1977). The use of Girard-T reagent in a rapid andsensitive method for measuring glyoxal and certain other a-dicarbonylcompounds. Analytical Biochemistry, 81, 47–56.
Morel, I., Lescoat, G., Cogrel, P., Sergent, O., Pasdeloup, N., Cillard, P., et al. (1993).Antioxidant and iron-chelating activities of the flavonoids catechin, quercetinand diosmetin on iron-loaded rat hepatocyte cultures. BiochemicalPharmacology, 45, 13–19.
Morita, H., & Itokawa, H. (1988). Cytotoxic and antifungal diterpenes from the seedsof Alpinia galanga. Planta Medica, 54, 117–120.
Mpalantinos, M. A., Moura, R. S., Parente, J. P., & Kuster, R. M. (1998). Biologicallyactive flavonoids and kava pyrones from the aqueous extract of Alpiniazerumbet. Phytotherapy Research, 12, 442–444.
Nakamura, S., Li, H., Adijiang, A., Pischetsrieder, M., & Niwa, T. (2007). Pyridoxalphosphate prevents progression of diabetic nephropathy. Nephrology DialysisTransplantation, 22, 2165–2174.
Oliveira, A. P., Furtado, F. F., Silva, M. S., Tavares, J. F., Mafra, R. A., Araújo, D. A. M.,et al. (2006). Calcium channel blockade as a target for the cardiovascular effectsinduced by the 8(17),12E,14-labdatrien-18-oic acid (labdane-302). VascularPharmacology, 44, 338–344.
Singh, R., Barden, A., Mori, T., & Beilin, L. (2001). Advanced glycation end-products:A review. Diabetologia, 44, 129–146.
Sirat, H. M., Masri, D., & Rahman, A. A. (1994). The distribution of labdane diterpenesin the Zingiberaceae of Malaysia. Phytochemistry, 36, 699–701.
Takagi, Y., Kashiwagi, A., Tanaka, Y., Asahina, T., Kikkawa, R., & Shigeta, Y. (1995).Significance of fructose-induced protein oxidation and formation of advancedglycation end product. Journal of Diabetes and Its Complications, 9, 87–91.
Teng, C. M., Hsu, S. Y., Lin, C. H., Yu, S. M., Wang, K. J., Lin, M. H., et al. (1990).Antiplatelet action of dehydrokawain derivatives isolated from Alpinia speciosarhizoma. The Chinese Journal of Physiology, 33, 41–48.
Thornalley, P. J., Langborg, A., & Minhas, H. S. (1999). Formation of glyoxal,methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose.Biochemical Journal, 344, 109–116.
Upadhyay, A., Chompoo, J., Kishimoto, W., Makise, T., & Tawata, S. (2011). HIV-1integrase and neuraminidase inhibitors from Alpinia zerumbet. Journal ofAgricultural and Food Chemistry. <http://www.dx.doi.org/10.1021/jf10481k>.
Well-Knecht, K. J., Zyzak, D. V., Litchfield, J. E., Thorpe, S. R., & Baynes, J. W. (1995).Mechanism of autoxidative glycosylation: Identification of glyoxal andarabinose as intermediates in the autoxidative modification of proteins byglucose. Biochemistry, 34, 3702–3709.
Wu, J. W., Hsieh, C. L., Wang, H. Y., & Chen, H. Y. (2009). Inhibitory effects of guava(Psidium guajava L.) leaf extracts and its active compounds on glycation processof protein. Food Chemistry, 113, 78–84.
Xu, H. X., Dong, H., & Sim, K. Y. (1996). Labdane diterpenes from Alpinia zerumbet.Phytochemistry, 42, 149–151.
Zoghbi, M. G. B., Andrade, E. H. A., & Maia, J. G. S. (1999). Volatile constituents fromleaves and flowers of Alpinia speciosa K. Schum. and A. purpurata (Viell.) Schum.Flavour and Fragrance Journal, 14, 411–414.
Related Documents











