Adolescents Exhibit Behavioral Differences From Adults During Instrumental Learning and Extinction David A. Sturman, Daniel R. Mandell, and Bita Moghaddam University of Pittsburgh Adolescence is associated with the development of brain regions linked to cognition and emotion. Such changes are thought to contribute to the behavioral and neuropsychiatric vulnerabilities of this period. We compared adolescent (Postnatal Days 28 – 42) and adult (Postnatal Day 60) rats as they performed a simple instrumental task and extinction. Rats were trained to poke into a hole for a food-pellet reinforcer. After six days of training, rats underwent extinction sessions in which the previously rewarded behavior was no longer reinforced. During extinction, we examined the effects of continued presentation of a cue light and food restriction. Adults and adolescents exhibited similar performance during training, although adolescents made more task-irrelevant pokes, consistent with increased exploration. Adults made more premature pokes, which could indicate a more exclusive focus on the task. During extinction, adolescents made more perseverative (previously reinforced) pokes than adults. This behavior was strongly modu- lated by the combination of motivational factors present (food restriction and cue light), indicating that adolescents were differentially sensitive to them. Furthermore, food restriction induced greater open-field activity in adolescents but not in adults. Thus, as the neural circuitry of motivated behavior develops substantially during adolescence, so too does the behavioral sensitivity to motivational factors. Under- standing how such factors differently affect adolescents may shed light on mechanisms that lead to the development of disorders that are manifested during this period. Keywords: addiction, instrumental learning, extinction, schizophrenia, motivation Adolescence is a major transitional period between childhood and adulthood. It encompasses puberty, a time of reproductive development, and is characterized in humans and rodents by nu- merous nonreproductive sociobehavioral changes (Spear, 2000). It is during adolescence that the symptoms of several psychiatric disorders typically arise, including depression, eating disorders, and schizophrenia (Pine, 2002; Sisk & Zehr, 2005; Volkmar, 1996). Characteristic adolescent behavioral changes include ele- vated social interaction (Csikszentmihalyi, Larson, & Prescott, 1977) and increased novelty-seeking and risk-taking behavior (Adriani, Chiarotti, & Laviola, 1998; Macrı `, Adriani, Chiarotti, & Laviola, 2002; Spear, 2000; Stansfield & Kirstein, 2006; Stein- berg, 2008). These latter behaviors correlate with drug and alcohol use (Andrucci, Archer, Pancoast, & Gordon, 1989), and adoles- cence is often considered a period of increased addiction vulner- ability (Adriani & Laviola, 2004; Chambers, Taylor, & Potenza, 2003). Furthermore, as in human adolescents, adolescent mice exhibit greater impulsivity than adults, as measured by reduced preference for larger delayed food reinforcers over more immedi- ate smaller ones (Adriani & Laviola, 2003). Along with these behavioral changes, the adolescent brain un- dergoes extensive remodeling (McCutcheon & Marinelli, 2009), with neurogenesis (Pinos et al., 2001), axonal growth (Benes, Taylor, & Cunningham, 2000; Cunningham, Bhattacharyya, & Benes, 2002), myelination (Benes, Turtle, Khan, & Farol, 1994), apoptosis (Nunez, Sodhi, & Juraska, 2002), and synaptic and receptor pruning (Andersen, Thompson, Rutstein, Hostetter, & Teicher, 2000; Meyer, Ferres-Torres, & Mas, 1978; Teicher, Andersen, & Hostetter, 1995) accompanying shifts in white-matter density and cortical gray-matter volume (Benes et al., 1994; Giedd et al., 1999; Juraska & Markham, 2004; Paus, 2005; Paus et al., 2001). The mesocortical dopaminergic circuitry, considered broadly relevant to motivated behavior (for reviews, see Cools [2008], Salamone & Correa [2002], Berridge [2007], and Floresco and Magyar [2006]) also undergoes considerable development during adolescence (Chambers et al., 2003; Ernst & Fudge, 2009; Lewis, 1997; Spear, 2000). Much of the literature on adolescence has focused on drug- related behavioral differences such as ethanol or psychostimulant sensitivity differences (Badanich, Maldonado, & Kirstein, 2008; Bolanos, Glatt, & Jackson, 1998; Little, Kuhn, Wilson, & Swartzwelder, 1996; Moy, Duncan, Knapp, & Breese, 1998; Pau- tassi, Myers, Spear, Molina, & Spear, 2008; Spear & Brake, 1983; Varlinskaya & Spear, 2006). For example, adolescents tend to exhibit less amphetamine- and cocaine-induced locomotor stimu- lation and stereotypy (Bolanos et al., 1998; Laviola, Adriani, Terranova, & Gerra, 1999; Mathews & McCormick, 2007; Spear & Brake, 1983). Conversely, adolescent rats show greater sensi- tivity to the cataleptic effects of neuroleptics such as haloperidol (Shalaby & Spear, 1980; Spear & Brake, 1983). Less is known David A. Sturman, Daniel R. Mandell, and Bita Moghaddam, Depart- ment of Neuroscience, University of Pittsburgh. This work was supported by National Institute of Mental Health Grants MH48404 and MH065468 and the Andrew Mellon Foundation for a predoctoral fellowship (David A. Sturman). Correspondence concerning this article should be addressed to Bita Moghaddam, Department of Neuroscience, University of Pittsburgh, A210 Langley Hall, Pittsburgh, PA 15260. E-mail: [email protected] Behavioral Neuroscience © 2010 American Psychological Association 2010, Vol. 124, No. 1, 16 –25 0735-7044/10/$12.00 DOI: 10.1037/a0018463 16

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Adolescents Exhibit Behavioral Differences From Adults DuringInstrumental Learning and Extinction
David A. Sturman, Daniel R. Mandell, and Bita MoghaddamUniversity of Pittsburgh
Adolescence is associated with the development of brain regions linked to cognition and emotion. Suchchanges are thought to contribute to the behavioral and neuropsychiatric vulnerabilities of this period. Wecompared adolescent (Postnatal Days 28–42) and adult (Postnatal Day 60�) rats as they performed asimple instrumental task and extinction. Rats were trained to poke into a hole for a food-pellet reinforcer.After six days of training, rats underwent extinction sessions in which the previously rewarded behaviorwas no longer reinforced. During extinction, we examined the effects of continued presentation of a cuelight and food restriction. Adults and adolescents exhibited similar performance during training, althoughadolescents made more task-irrelevant pokes, consistent with increased exploration. Adults made morepremature pokes, which could indicate a more exclusive focus on the task. During extinction, adolescentsmade more perseverative (previously reinforced) pokes than adults. This behavior was strongly modu-lated by the combination of motivational factors present (food restriction and cue light), indicating thatadolescents were differentially sensitive to them. Furthermore, food restriction induced greater open-fieldactivity in adolescents but not in adults. Thus, as the neural circuitry of motivated behavior developssubstantially during adolescence, so too does the behavioral sensitivity to motivational factors. Under-standing how such factors differently affect adolescents may shed light on mechanisms that lead to thedevelopment of disorders that are manifested during this period.
Keywords: addiction, instrumental learning, extinction, schizophrenia, motivation
Adolescence is a major transitional period between childhoodand adulthood. It encompasses puberty, a time of reproductivedevelopment, and is characterized in humans and rodents by nu-merous nonreproductive sociobehavioral changes (Spear, 2000). Itis during adolescence that the symptoms of several psychiatricdisorders typically arise, including depression, eating disorders,and schizophrenia (Pine, 2002; Sisk & Zehr, 2005; Volkmar,1996). Characteristic adolescent behavioral changes include ele-vated social interaction (Csikszentmihalyi, Larson, & Prescott,1977) and increased novelty-seeking and risk-taking behavior(Adriani, Chiarotti, & Laviola, 1998; Macrı̀, Adriani, Chiarotti, &Laviola, 2002; Spear, 2000; Stansfield & Kirstein, 2006; Stein-berg, 2008). These latter behaviors correlate with drug and alcoholuse (Andrucci, Archer, Pancoast, & Gordon, 1989), and adoles-cence is often considered a period of increased addiction vulner-ability (Adriani & Laviola, 2004; Chambers, Taylor, & Potenza,2003). Furthermore, as in human adolescents, adolescent miceexhibit greater impulsivity than adults, as measured by reducedpreference for larger delayed food reinforcers over more immedi-ate smaller ones (Adriani & Laviola, 2003).
Along with these behavioral changes, the adolescent brain un-dergoes extensive remodeling (McCutcheon & Marinelli, 2009),with neurogenesis (Pinos et al., 2001), axonal growth (Benes,Taylor, & Cunningham, 2000; Cunningham, Bhattacharyya, &Benes, 2002), myelination (Benes, Turtle, Khan, & Farol, 1994),apoptosis (Nunez, Sodhi, & Juraska, 2002), and synaptic andreceptor pruning (Andersen, Thompson, Rutstein, Hostetter, &Teicher, 2000; Meyer, Ferres-Torres, & Mas, 1978; Teicher,Andersen, & Hostetter, 1995) accompanying shifts in white-matterdensity and cortical gray-matter volume (Benes et al., 1994; Gieddet al., 1999; Juraska & Markham, 2004; Paus, 2005; Paus et al.,2001). The mesocortical dopaminergic circuitry, consideredbroadly relevant to motivated behavior (for reviews, see Cools[2008], Salamone & Correa [2002], Berridge [2007], and Florescoand Magyar [2006]) also undergoes considerable developmentduring adolescence (Chambers et al., 2003; Ernst & Fudge, 2009;Lewis, 1997; Spear, 2000).
Much of the literature on adolescence has focused on drug-related behavioral differences such as ethanol or psychostimulantsensitivity differences (Badanich, Maldonado, & Kirstein, 2008;Bolanos, Glatt, & Jackson, 1998; Little, Kuhn, Wilson, &Swartzwelder, 1996; Moy, Duncan, Knapp, & Breese, 1998; Pau-tassi, Myers, Spear, Molina, & Spear, 2008; Spear & Brake, 1983;Varlinskaya & Spear, 2006). For example, adolescents tend toexhibit less amphetamine- and cocaine-induced locomotor stimu-lation and stereotypy (Bolanos et al., 1998; Laviola, Adriani,Terranova, & Gerra, 1999; Mathews & McCormick, 2007; Spear& Brake, 1983). Conversely, adolescent rats show greater sensi-tivity to the cataleptic effects of neuroleptics such as haloperidol(Shalaby & Spear, 1980; Spear & Brake, 1983). Less is known
David A. Sturman, Daniel R. Mandell, and Bita Moghaddam, Depart-ment of Neuroscience, University of Pittsburgh.
This work was supported by National Institute of Mental Health GrantsMH48404 and MH065468 and the Andrew Mellon Foundation for apredoctoral fellowship (David A. Sturman).
Correspondence concerning this article should be addressed to BitaMoghaddam, Department of Neuroscience, University of Pittsburgh, A210Langley Hall, Pittsburgh, PA 15260. E-mail: [email protected]
Behavioral Neuroscience © 2010 American Psychological Association2010, Vol. 124, No. 1, 16–25 0735-7044/10/$12.00 DOI: 10.1037/a0018463
16

about developmental differences in motivated behavior more gen-erally, such as how various nonpharmacological factors mightaffect adolescents differently than adults. The aim of this studywas to characterize several similarities and differences in adoles-cent motivated behavior during an instrumental learning task andextinction. Although motivation can be a somewhat elusive con-cept, others have defined it as “the set of processes through whichorganisms regulate the probability, proximity and availability ofstimuli” (Salamone & Correa, 2002, p. 5). Here we use the termmotivational factor to identify elements that increase behavioralmanifestations of such processes in an organism. We tested ado-lescent and adult rats in a simple paradigm in which they learnedto pair a particular action (nose poke) with a desired outcome (foodpellet) and followed the training period with extinction, at whichpoint the action–outcome association was no longer reinforced.During extinction, we examined how continued food restrictionand presentation of a task cue may differently affect adolescents.We further compared the impact of one of these motivationalfactors, food restriction, on adolescent and adult open-field activ-ity. Studying such differences may inform our understanding ofhow adolescent neurodevelopment leads to typical age-specificbehavioral propensities and disease processes.
Method
Subjects
Adolescent (Postnatal Days 28–42; n � 42) and adult (PostnatalDay 60�; n � 42) male Sprague–Dawley rats (Harlan, Frederick,MD) were used. Preadolescent juvenile rats (Postnatal Day 21) andadults were received 4 days before beginning handling and operantbox habituation. Training on the instrumental task began immedi-ately after habituation, corresponding to 1 week after arrival (Post-natal Day 28 for adolescents). All subjects were housed in pairsunder 12-hr light–dark cycle conditions (lights on at 7 p.m.), andtesting was performed during rats’ active phase. Food restrictionwas imposed during the habituation period, at which time pread-olescents received 5 g and 8 g chow on consecutive days and thenwere maintained at 10 g chow per day on the final day of habit-uation and throughout training. This level of food restriction waschosen after observing that not all food was consumed from theprevious day in some cages before testing when early adolescentswere fed 5 g and then 8 g and then sustained at 12 g chow per day.Adults were given 15 g chow per day during habituation andtraining. All rats had ad lib access to water except during testing.Experimental protocols were approved by the University of Pitts-burgh Animal Care and Use Committee.
Instrumental Task
Operant chambers (Coulbourn Instruments, Allentown, PA)were equipped with a house light that illuminated the chamberduring the task, three nose-poke holes, a food trough, and a foodpellet delivery system. Nose-poke holes were arrayed horizontallyon the wall opposite the food trough. Entries into the nose-pokeholes or the food trough were detected by infrared photosensors. APC-based controller and Graphic State software (Coulbourn In-struments) were used to run the task and record the rats’ behavior.
During the 1st day of habituation, rats were placed in the operantchamber for 20 min with the house light on. During the 2nd and3rd days of habituation, rats were placed in the operant box for 20min with the house light on and food pellets (fortified dextrosepellets, 45 mg; Bio-Serv, Frenchtown, NJ) were delivered into thefood trough every 30 s.
Rats were then trained on a reinforcement schedule in which asingle instrumental nose poke was reinforced with the delivery ofa single food pellet. The house light was continuously illuminatedfor the duration of each session. Trials began with the illuminationof a light cue in the center (and only the center) nose-poke hole.After a rat poked into that hole, the light turned off, and one pelletwas delivered to the food trough, along with the illumination of afood trough light. The trial-onset cue remained illuminated untilthe rat performed the nose poke (instrumental response). For thenext trial to begin, the rat was required to poke into the food troughto retrieve the pellet, which turned off the food trough light, andthen wait for a fixed 5-s intertrial interval (ITI). Rats received dailytraining sessions over 6 consecutive days. Each session was ter-minated on the delivery of 99 pellets or the passage of 30 min.Nose pokes into either of the nonilluminated (left and right)modules were not reinforced, although this behavior was recordedand categorized as task-irrelevant pokes. Video cameras allowedbehavior to be monitored by the experimenter during testing. Ratsthat did not learn the instrumental task after 3 days were hand-shaped to the center hole and performed the task for three fullsessions after this. These rats (n � 5) were excluded from allinstrumental training analyses. However, because their extinctionand open-field behavior was not statistically different from that oftheir peers, these data were combined with their correspondinggroups.
Extinction
The day after the completion of the last training session, ratsbegan one of four extinction paradigms, during which the instru-mental behavior was no longer reinforced. In the first group (adultn � 18; adolescent n � 18), rats remained food restricted. Addi-tionally, the trial-onset cue that was previously associated with thebeginning of each trial during training continued to be presentedduring extinction. A poke to the illuminated hole would turn offthe cue light as before, but no food pellet was delivered. If the ratthen poked in the food trough, after a 5-s ITI the cue light waspresented again. If it did not poke into the food trough, the cuelight would reappear after a 15-s delay. In a second group (adultn � 6, adolescent n � 6), rats underwent extinction exactly as inthe first group, except that these rats were given ad lib access tofood in their home cages each day (beginning immediately afterthe last training session). In a third extinction group (adult n � 12;adolescent n � 12), food restriction was maintained; however, nocue light was presented to the rats during extinction. In a finalgroup (adult n � 6; adolescent n � 6), rats were given ad libaccess to food in their home cages and no cue was presentedduring extinction sessions.
Open Field
After operant-box testing during the 2nd and 3rd days of ex-tinction, a subset of food-restricted and ad-lib-fed adult and ado-
17ADOLESCENT INSTRUMENTAL LEARNING AND EXTINCTION

lescent rats (adult n � 24; adolescent n � 24) was placed in thecenter of an open field arena (1 m � 1 m, divided into 25 squares)under normal white lighting and given 5 min to explore whilebeing videotaped. An experimenter, unaware of the rats’ food-restriction status, rated the number of total square entries andentries into the central grids. A square entry was counted when arat’s hind legs passed from one square into the next.
Statistical Analysis
To delineate age-related differences in task performance duringtraining, age (between) � session (within) repeated-measures anal-yses of variance (ANOVAs) were performed on total trials persession, task-irrelevant pokes (left- and right-hole pokes), latencyfrom cue onset to instrumental poke, and latency from instrumentalpoke to food-trough poke. During extinction, repeated-measuresANOVAs were used to examine the effects of age, food-restrictionstatus, cue-light presence, and their interactions on perseverativeand task-irrelevant pokes. These models were broken up to moreeasily interpret potential three-way between-factor interactions.The following analyses were performed separately on rats forwhich a cue was or was not presented during extinction: Age �Food-Restriction Status (between) � Session (within); on food-restricted and ad-lib-fed rats, Age � Cue-Light Presence (be-tween) � Session (within); and for adults and adolescents, Food-Restriction Status � Cue-Light Presence (between) � Session(within). To assess open-field behavior, Age � Food-RestrictionStatus ANOVAs were performed on total grid entries and centralgrid entries. When necessary, significant ANOVA results weresupplemented with Fisher’s least significant difference post hoctests. In all repeated-measures ANOVAs for which the assumptionof sphericity was violated, the lower-bound correction was usedfor a maximally conservative degrees-of-freedom adjustment.Pearson’s correlation coefficient and analysis of covariance (usedin models with fixed factors such as age or food-restriction status)were used to determine the relationship between continuous vari-ables (e.g., total trials performed during training, total persevera-tive pokes during extinction) and to test for potential relationshipswithin each age group between body weight and training trials,task-irrelevant pokes, and perseveration during extinction. We alsoused Pearson’s correlation coefficient to examine the relationshipbetween task-irrelevant pokes (during training and during extinc-tion) and open-field behaviors, training trials, and perseverationduring extinction. To avoid the potential problem of singularpoking events registering as multiple pokes (e.g., several pokeswithin 1 s as a rat retrieves a food pellet), a distinct poke wasdefined as one that was not immediately preceded by a previouspoke within 1 s. The exception to this was counting an instrumen-tal response as such even immediately after a premature poke.
Results
A significant interaction effect was observed for the total num-ber of trials performed by adults versus adolescents across trainingsessions, F(1, 1) � 7.05, p � .01. This was the result of adultsperforming a significantly greater number of trials in Sessions 3–6,but not Sessions 1–2, when most of the initial learning took place.Total trial performance was generally stable for adolescents andadults beginning in Sessions 3–4 (Figure 1A). Examination of the
Figure 1. Adolescent and adult trial performance across training sessions.A: Adolescents perform similar total trials during the first two trainingsessions. From Session 3 onward, adults perform more total trials thanadolescents. B: Within-session average cumulative trial performance overtime plus and minus standard error (dashed lines). Sessions 1 and 2 indicatesimilar or slightly faster performance (steeper slope) by adolescents. Therate of performance is nearly equal during the early portions of Sessions3–6. The drop in trial performance rate about 5–10 min into those sessionsby adolescents contributes to their lower total trials and may reflect earliersatiety. * � significant difference between adolescents and adults.
18 STURMAN, MANDELL, AND MOGHADDAM

average cumulative trials performed over time within sessionsdemonstrated that the difference between adolescents and adults intotal trials in Sessions 3–6 was associated with an early drop in therate of adolescent trial performance after initially being similar tothat of adults (Figure 1B). The rate of adolescent trial performancein the first session was slightly faster (a steeper line) than that ofadults, suggesting that on average, adolescents either learned thetask slightly earlier or were simply slightly more active during thisperiod (Figure 1B).
A significant Age � Session interaction was found for the meanlatency from cue onset to instrumental poke, F(1, 1) � 5.64, p �.05. Although the latency from trial-onset cue to center-hole pokewas initially lower for adolescents than for adults, in Sessions 4–6adults had a shorter average latency (Figure 2A). Because adoles-cents more readily reduced their response rate in the latter portionof most training sessions (Figure 1B), the age-related latencydifferences in Sessions 4–6 could be the result of this effect. Wetherefore examined this latency during the first 5 min of eachsession, when adolescents and adults performed trials at the high-est rate. A significant age difference was still present, F(1, 77) �9.03, p � .01; however, there was no significant difference inlatency for Sessions 3–6 (Figure 2B). There was also no signifi-cant main effect or interaction for the average latency from instru-mental poke to food-trough entry, F(1, 77) � 3.37, p � .05, andF(1, 1) � 2.49, p � .05, although adolescents appeared to exhibita shorter latency during the first training session (Figure 2C).Despite these latency similarities (and even slightly shorter cue-to-poke latencies for adolescents in early sessions), adults consis-tently performed more premature pokes across sessions, defined aspokes before the trial-onset cue during the ITI, F(1, 77) � 21.72,p � .001 (Figure 3A). Conversely, adolescents consistently per-formed more task-irrelevant (left- and right-hole) pokes than adultsduring training, F(1, 77) � 191.31, p � .001 (Figure 3B).
During extinction sessions when the trial-onset cue was present,food-restricted adolescents performed significantly more perse-verative (previously reinforced center hole) pokes than adults, F(1,33) � 33.16, p � .001. No Age � Session interaction was present,F(1, 1) � 1.28, p � .05 (Figure 4A). In food-restricted rats forwhich no cue light was presented during extinction, a main effectof age was still observed, F(1, 22) � 32.14, p � .001, and againno interaction was present, F(1, 1) � 0.535, p � .05 (Figure 4B).To test the hypothesis that the difference in perseverative pokesbetween food-restricted adolescents and adults was larger whenthe cue was present than when it was absent, we ran an Age �Cue-Light Presence (between) � Session (within) repeated-measures ANOVA. This model indicated via a significant Age �Cue-Light Presence interaction that food-restricted adolescentsperformed disproportionately more perseverative pokes than food-restricted adults when the cue light was present than when it wasabsent, F(1, 55) � 4.41, p � .05 (Figure 5A). Rats that were notfood restricted during extinction performed significantly fewerperseverative pokes than food-restricted rats when the cue waspresent, F(1, 43) � 35.07, p � .01. No differences were observedbetween ad-lib-fed adolescents and adults when the cue light waspresent, F(1, 10) � 1.73, p � .05 (Figure 4C), and no significantAge � Cue Light Presence interaction was present in ad-lib-fedrats, F(1, 20) � 1.95, p � .05 (Figure 5B). An Age � Food-Restriction Status interaction was observed, F(1, 43) � 6.15, p �.05, indicating that among rats to which the cue was presented
(Figures 4A and 4C), food restriction more strongly increasedperseverative pokes in adolescents than in adults (Figure 5C).When the cue light was absent (Figures 4B and 4D), food-restricted rats still performed more perseverative pokes than ad-
Figure 2. Evidence of slightly faster adolescent task acquisition followedby parity of performance. A: The mean latency from trial onset cue tocentral poke (instrumental response) was shorter in the first training ses-sion. During Sessions 4–6, this latency became shorter in adults than inadolescents. B: Taking only the first 5 min of each session, before thedrop-off in within-session cumulative trial performance seen in Figure 1B,we see that adolescent latencies are still shorter in early sessions, but therewere no significant latency differences in later sessions. C: There was alsono significant age difference in the mean latency from the central poke tothe retrieval of the food pellet. * � significant difference between agegroups.
19ADOLESCENT INSTRUMENTAL LEARNING AND EXTINCTION

lib-fed rats, F(1, 32) � 11.57, p � .01. However, no Age �Food-Restriction Status interaction was observed when the cuelight was absent, F(1, 32) � 0.148, p � .05, indicating that foodrestriction did not have a stronger effect on adolescents than onadults when the cue was absent (Figure 5D). When rats were nolonger food restricted and the cue was omitted, adults performedfewer perseverative pokes than adolescents, F(1, 10) � 39.79, p �.001 (Figure 4D). We performed a Cue Presence � Food-Restriction Status (between) � Session (within) ANOVA sepa-rately on adolescents and adults. Adolescents exhibited a signifi-cant Food-Restriction Status � Cue Presence interaction, F(1,38) � 11.96, p � .001 (Figure 5E), indicating that cue presenceinteracted with food restriction to further increase perseveration inthese younger rats. In adults, however, no such interaction wasobserved, F(1, 37) � 2.43, p � .05 (Figure 5F). Total trialsperformed during training was a significant covariate for adultsand adolescents predictive of total perseverative pokes duringextinction, F(1, 75) � 11.49, p � .001. This indicates a positivelinear relationship between training trials and perseveration withinage groups, although adolescents tended to perform fewer totaltrials during training but more perseverative pokes during extinc-
tion than did adults. No statistically significant relationship wasobserved for trial performance, task-irrelevant poking, or perse-verative pokes during extinction as a function of the covariatebody weight when food-restriction status was included as a fixedfactor ( p � .05; data not shown).
Adolescents continued to perform significantly more task-irrelevant pokes than adults during extinction, F(1, 76) � 124.31,p � .001 (Figure 6A). A main effect of food restriction on totaltask-irrelevant pokes across extinction sessions was observed, F(1,80) � 7.25, p � .01. However, post hoc comparisons indicated thatalthough ad-lib-fed adults performed significantly fewer total task-irrelevant pokes than food-restricted adults, this effect was notsignificant for adolescents (Figure 6B). There was no statisticallysignificant correlation between task-irrelevant pokes and open-field grid entries, total training trials, or perseveration duringextinction for adolescents or adults ( p � .05; data not shown).
Food-restricted adolescents gained weight throughout the exper-iment. On the 1st day of testing, food-restricted adolescents (Post-natal Day 28) weighed (M � SD) 74.9 � 6.1 g. By the 1st day ofextinction, food-restricted adolescents (Postnatal Day 34) weighed110.4 � 7.2 g. On the 5th day of extinction, food-restrictedadolescents (Postnatal Day 38) weighed 116.9 � 8.2 g.
In the open field, adolescents performed more total grid entriesthan adults, F(1, 43) � 90.48, p � .001, and an Age � Food-Restriction Status interaction was significant, F(1, 43) � 5.55, p �
Figure 3. Adolescents and adults performed different behaviors betweentrials. A: Adults consistently performed more premature (precue) pokesduring the intertrial interval than adolescents. In both age groups, thenumber of premature pokes increased, but this increase was more pro-nounced in adults than in adolescents. B: Adolescents consistently per-formed more task-irrelevant (left- and right-hole) pokes than adults. * �significant age-related differences.
Figure 4. Age-related differences in perseverative (previously rein-forced) poking behavior were modulated by the presence of motivationalfactors. A: When rats remained food restricted and the cue light continuedto be presented during extinction, adolescents consistently performed moreperseverative pokes than adults. B: When the cue light was omitted but ratswere still food restricted, adolescents still perseverated more, but thisdifference was smaller than in A. C: When the rats had ad lib access to foodoutside of the task and the cue was still presented, there were no perse-veration differences between adolescents and adults. D: When rats wereneither food restricted nor presented the cue, adults performed fewerperseverative pokes. Adolescents in this condition were more similar intheir perseveration to their counterparts exposed to a single motivationalfactor as in B and C. * � significant age-related differences.
20 STURMAN, MANDELL, AND MOGHADDAM

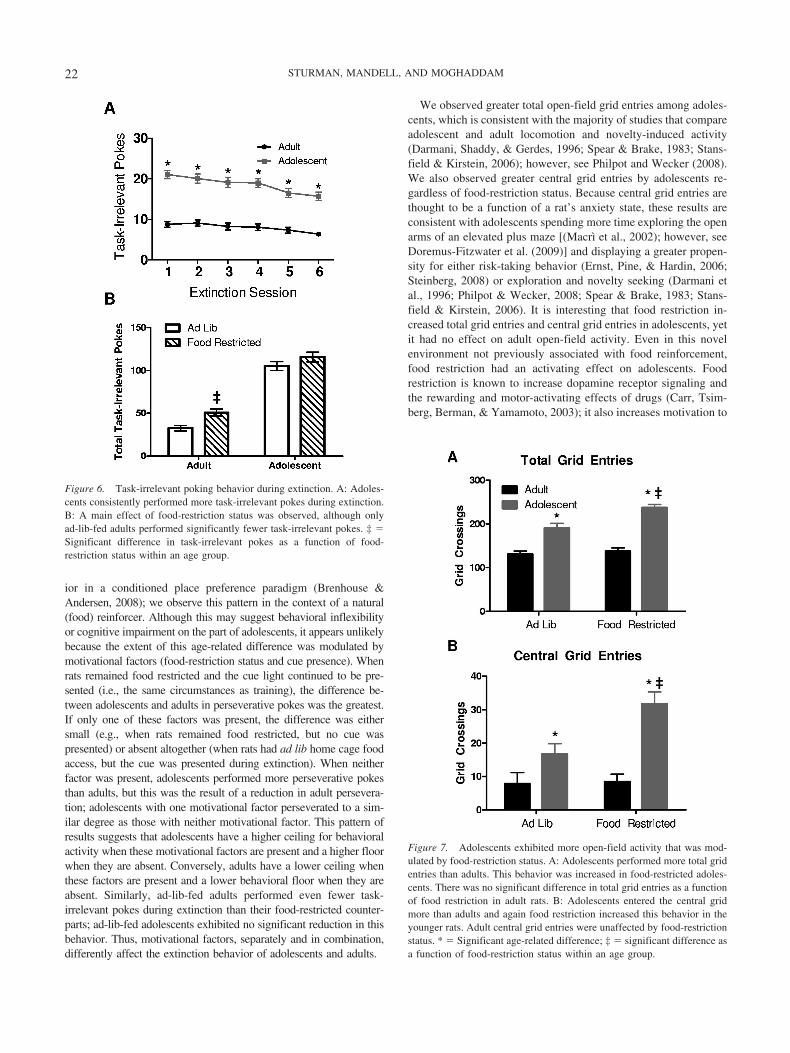
.05. Food restriction increased adolescents’ total grid entries, butadults were unaffected (Figure 7A). The effect was similar forcentral grid entries, with adolescents entering the central gridsmore than adults, F(1, 43) � 25.38, p � .001, and an Age �Food-Restriction Status interaction was observed, F(1, 43) � 5.03,p � .05. As with total grid entries, food restriction increasedcentral grid entries in adolescents but not in adults (Figure 7B).
Discussion
In this study, we compared the behavior of adolescent and adultrats during a simple instrumental task and extinction. There was noage-related difference in the total number of trials performedduring the first two sessions, although the within-session rate oftrial performance and the latencies from the trial-onset cue to theinstrumental response may have been slightly faster in adolescentsduring those sessions. By Sessions 3–4, adults and adolescentsreached a stable maximum in the total number of trials performed,with adults performing more total trials from Session 3 onward.We also found that the rate of trial performance was similar in theearly portions of these sessions, and an absence of latency differ-ences indicated similar performance once the task was welllearned. We did observe persistent differences in task-irrelevantpokes (performed more by adolescents) and premature pokes (per-formed more by adults). During extinction, we found that adoles-cents tended to perform more perseverative pokes than adults,although the presence and extent of this difference was modulatedby the combination of food restriction and continued cue presen-tation. Finally, we observed that adolescents were generally moreactive in an open field than adults and that this activity wasincreased by food restriction in the younger rats only.
Adolescents and adults performed different behaviors during theITI. The increased task-irrelevant pokes by adolescents may beconsistent with greater exploration and general activity in adoles-cents, described by others (Shalaby & Spear, 1980; Spear, 2000)and observed in our open-field experiment. It is notable, however,that we found no significant predictive relationship among indi-vidual rats between open-field activity and task-irrelevant poking.Because others have interpreted premature pokes in a different taskas a measure of impulsivity (Carli, Robbins, Evenden, & Everitt,1983; Robbins, 2002), we were initially surprised to observe thisbehavior more in adults than in adolescents. One important differencebetween those studies and this one is that in this study there was nopenalty for premature poking, and so rats had no reason to resist adesire to poke early. Thus, the behavior probably does not reflect poorimpulse control in the present task. Both adolescents and adultsincreased premature poking across training sessions; this suggests thatpremature pokes do not indicate a weaker cue–action association onthe part of adults because we would expect such an association toimprove over time. The observed age-related differences in thesebehaviors could represent a greater exclusive focus on the task at handby adults, whereas adolescents are more inclined to shift their atten-tion to the task-irrelevant holes during that period. It is noteworthythat once the trial-onset cue was presented, adolescents were at leastas quick to respond as adults. Thus, premature poking may indeedreflect the single-mindedness of task-performing rats, with adoles-cents tending to divert themselves more often (although to a lesserextent over time) to explore task-irrelevant holes, although this pro-pensity does not impair task performance.
During extinction, adolescents tended to perform more persevera-tive (previously reinforced) pokes. Others have demonstrated thatadolescents exhibit resistance to extinction of cocaine-seeking behav-
Figure 5. Interaction plots demonstrating differences between adolescents and adults in their sensitivities to thepresence of motivational factors in different combinations collapsed across extinction sessions. A: Amongfood-restricted rats, a significant Age � Cue Presence interaction was observed. Adolescents perseverated morethan food-restricted adults generally, but importantly, the presence of the cue had a stronger activating effect onadolescents than on adults. B: There was no Age � Cue Presence interaction in ad lib rats. C: A significantAge � Food-Restriction Status interaction was observed for rats presented the cue. Among these rats, only thefood-restricted adolescents perseverated more than adults. D: No Age � Food-Restriction Status interaction wasobserved in animals for which the cue was omitted. E: In adolescents, the combination of both motivationalfactors interacted to further increase perseveration. A single or no motivational factor led to similar lower levels.F: Unlike adolescents, in adults the combination of motivational factors did not interact synergistically to furtherincrease perseverative pokes. † � significant interaction.
21ADOLESCENT INSTRUMENTAL LEARNING AND EXTINCTION
ior in a conditioned place preference paradigm (Brenhouse &Andersen, 2008); we observe this pattern in the context of a natural(food) reinforcer. Although this may suggest behavioral inflexibilityor cognitive impairment on the part of adolescents, it appears unlikelybecause the extent of this age-related difference was modulated bymotivational factors (food-restriction status and cue presence). Whenrats remained food restricted and the cue light continued to be pre-sented (i.e., the same circumstances as training), the difference be-tween adolescents and adults in perseverative pokes was the greatest.If only one of these factors was present, the difference was eithersmall (e.g., when rats remained food restricted, but no cue waspresented) or absent altogether (when rats had ad lib home cage foodaccess, but the cue was presented during extinction). When neitherfactor was present, adolescents performed more perseverative pokesthan adults, but this was the result of a reduction in adult persevera-tion; adolescents with one motivational factor perseverated to a sim-ilar degree as those with neither motivational factor. This pattern ofresults suggests that adolescents have a higher ceiling for behavioralactivity when these motivational factors are present and a higher floorwhen they are absent. Conversely, adults have a lower ceiling whenthese factors are present and a lower behavioral floor when they areabsent. Similarly, ad-lib-fed adults performed even fewer task-irrelevant pokes during extinction than their food-restricted counter-parts; ad-lib-fed adolescents exhibited no significant reduction in thisbehavior. Thus, motivational factors, separately and in combination,differently affect the extinction behavior of adolescents and adults.
We observed greater total open-field grid entries among adoles-cents, which is consistent with the majority of studies that compareadolescent and adult locomotion and novelty-induced activity(Darmani, Shaddy, & Gerdes, 1996; Spear & Brake, 1983; Stans-field & Kirstein, 2006); however, see Philpot and Wecker (2008).We also observed greater central grid entries by adolescents re-gardless of food-restriction status. Because central grid entries arethought to be a function of a rat’s anxiety state, these results areconsistent with adolescents spending more time exploring the openarms of an elevated plus maze [(Macrı̀ et al., 2002); however, seeDoremus-Fitzwater et al. (2009)] and displaying a greater propen-sity for either risk-taking behavior (Ernst, Pine, & Hardin, 2006;Steinberg, 2008) or exploration and novelty seeking (Darmani etal., 1996; Philpot & Wecker, 2008; Spear & Brake, 1983; Stans-field & Kirstein, 2006). It is interesting that food restriction in-creased total grid entries and central grid entries in adolescents, yetit had no effect on adult open-field activity. Even in this novelenvironment not previously associated with food reinforcement,food restriction had an activating effect on adolescents. Foodrestriction is known to increase dopamine receptor signaling andthe rewarding and motor-activating effects of drugs (Carr, Tsim-berg, Berman, & Yamamoto, 2003); it also increases motivation to
Figure 6. Task-irrelevant poking behavior during extinction. A: Adoles-cents consistently performed more task-irrelevant pokes during extinction.B: A main effect of food-restriction status was observed, although onlyad-lib-fed adults performed significantly fewer task-irrelevant pokes. ‡ �Significant difference in task-irrelevant pokes as a function of food-restriction status within an age group.
Figure 7. Adolescents exhibited more open-field activity that was mod-ulated by food-restriction status. A: Adolescents performed more total gridentries than adults. This behavior was increased in food-restricted adoles-cents. There was no significant difference in total grid entries as a functionof food restriction in adult rats. B: Adolescents entered the central gridmore than adults and again food restriction increased this behavior in theyounger rats. Adult central grid entries were unaffected by food-restrictionstatus. * � Significant age-related difference; ‡ � significant difference asa function of food-restriction status within an age group.
22 STURMAN, MANDELL, AND MOGHADDAM

work for food, which is why food-reinforced instrumental taskssuch as ours use it. It is possible that the novelty of the open-fieldenvironment maximally activated adults, such that food restrictiondid not further increase activity. In addition to exhibiting a higherbaseline of open-field activity, the increased food-restriction-induced activity of adolescents may be the result of a higherpotential behavioral ceiling in these younger rats.
We cannot be certain that the level of food restriction and itsmotivational effects were equal between age groups. This is particu-larly difficult because adolescents gain a great deal of weight (andmust do so to be healthy, even under food restriction), whereas adultslose weight under this condition. Although we cannot equate adoles-cent and adult food restriction, our lack of observed latency differ-ences and the similar early-trial performance rate in Sessions 3–6indicate that whatever baseline motivational differences might exist,they were not large enough to cause differences in these behavioralmeasures once the task was well learned. The exception to this is thewithin-session drop in trial performance rate by adolescents, whichcould reflect earlier satiety by these rats. Such differences, however,do not adequately explain the sustained disparities in task-irrelevantpoking and perseveration during extinction. Similarly, behavioraldifferences in the open field could partially reflect unequal foodrestriction. If this were to entirely account for the age-related differ-ences, we might still expect to observe adult open-field activity to beaffected to a lesser or greater extent. The lack of any change in adultopen-field behavior suggests that the age-related behavioral differ-ences are not solely the result of differences in the degree of food-restriction severity. Finally, the pattern of perseveration during ex-tinction depended strongly on the combination of both food restrictionand cue presence. In fact, those adolescents that were ad lib while thecue was presented, food restricted with the cue absent, or lacked boththe cue and food restriction all exhibited similar levels of persevera-tion. It was only the combination of food restriction and cue presencethat substantially increased adolescent perseveration. Thus, age-related differences in the motivational consequences of food restric-tion appear insufficient to account for all of these results, although weacknowledge that this is a difficulty in using any task that foodrestricts adolescents and adults.
Although care was taken to expose adolescents and adults tosimilar conditions before and during behavioral testing, there is al-ways the possibility that adolescents could respond to housing, ship-ping, or other conditions differently and in such a way that mightaffect measures of behavioral performance. For example, adolescentsand adults were shipped 4 days before handling. If their transport wasstressful, our findings could reflect age-related differential effects ofshipping stress on performance. Similarly, food restriction reduces therate of normal adolescent weight gain. The weights of our adolescentrats were within the normal to low-normal free-feeding range ofadolescent Sprague–Dawley rats at corresponding ages described byMcCutcheon and Marinelli (2009). Nevertheless, it is possible thatreduced growth rate or various external factors could differently affectadolescent physiology and alter behavior.
The connection between adolescent neurodevelopmental andbehavioral changes are of great clinical relevance, especially inlight of the associated increased risk taking (Spear, 2000; Stein-berg, 2008) and addiction vulnerability of this period (Chambers etal., 2003; Compton, Thomas, Conway, & Colliver, 2005; Khuder,Dayal, & Mutgi, 1999). Significant components of the circuitrythat underlies motivated, goal-directed behavior undergo substan-
tial changes during adolescence (Doremus-Fitzwater, Varlinskaya,& Spear, in press; Spear, 2000). These regions subserve aspects ofinstrumental learning and extinction. The dorsal striatum is criticalto the expression of action–outcome associations of the sortformed during instrumental learning (Balleine, Liljeholm, &Ostlund, 2009). The prefrontal cortex (PFC) mediates the initialencoding of action–outcome learning and plays a central role incognitive flexibility, such as during extinction (Corbit & Balleine,2003; Jung, Baeg, Kim, Kim, & Kim, 2008). The amygdala andextended amygdala are thought to provide valence informationnecessary for learning the initial rewarding or antirewarding con-tingencies of an operant behavior and for allowing flexibility whencontingencies change (Koob, 2009). Finally, the nucleus accum-bens (NAc), which receives a convergence of information from thePFC, amygdala, thalamus, and other regions, is critical toincentive-motivated behavior, instrumental learning, and food in-take (Kelley, 2004; Mogenson, Jones, & Yim, 1980). These crucialbrain regions undergo numerous changes during adolescence.There is increasing dopaminergic innervation of the PFC duringadolescence (Benes et al., 2000; Rosenberg & Lewis, 1994).Dopamine D1, D2, and D4 receptor expression peaks duringadolescence in the dorsal striatum (Seeman et al., 1987; Tarazi &Baldessarini, 2000; Teicher et al., 1995; Teicher, Krenzel, Thomp-son, & Andersen, 2003) and PFC (Andersen et al., 2000) beforebeing pruned to lower adult levels. Others have also found asimilar pattern for the NAc (Tarazi & Baldessarini, 2000), al-though this has not always been observed (Teicher et al., 1995).During adolescence, there is also increasing glutamatergic connec-tivity from the basolateral amygdala to the PFC (Cunningham etal., 2002; Cunningham, Bhattacharyya, & Benes, 2008) and fromthe PFC to the NAc among D1-expressing neurons, before this isreduced in adulthood (Brenhouse, Sonntag, & Andersen, 2008).
Recently, the triadic node hypothesis has been proposed to explainthe elevated risk-taking behavior of adolescents in terms of underlyingneurodevelopment (Ernst & Fudge, 2009; Ernst et al., 2006). Thishypothesis posits that in adolescents, NAc-mediated approach is outof balance with amygdala-mediated avoidance. The third “node” ofthis model is the PFC, which normally maintains equilibrium betweenthese countervailing forces. This intriguing framework may accountfor adolescent proclivities to both take risks and behave impulsively.The idea that these circuits associated with reward and motivation areshifted in their sensitivities during adolescence is consistent withage-related regional neural activation differences in anticipation, re-ceipt, and omission of reward observed in functional MRI studies(Galvan et al., 2006; Van Leijenhorst et al., 2010) and greater ado-lescent preferences for natural (Douglas, Varlinskaya, & Spear, 2004;Wilmouth & Spear, 2009) and drug reinforcers (Badanich, Adler, &Kirstein, 2006; Brenhouse & Andersen, 2008; Brenhouse et al., 2008;Philpot, Badanich, & Kirstein, 2003; Shram, Funk, Li, & Le, 2006;Vastola, Douglas, Varlinskaya, & Spear, 2002). Thus, both the neuralcircuitry of motivated behavior and the sensitivity and preference forreinforcing stimuli are altered during adolescence. This framework isconsistent with our finding that adolescent perseveration during ex-tinction is differentially affected by the presence of certain motiva-tional factors.
Human adolescence is often considered a period of storm and stressbecause of tendencies toward heightened interpersonal conflict, emo-tional reactivity, and risk behavior (Arnett, 1999). Although mostadolescents do not experience psychiatric problems, it is at this time
23ADOLESCENT INSTRUMENTAL LEARNING AND EXTINCTION

that problems often arise (Pine, 2002; Sisk & Zehr, 2005; Spear, 2000;Volkmar, 1996). The changing cognitive and affective milieu of thedeveloping brain may cause adolescents to process and react tointernal and external stimuli differently. This in turn could be relevantto their increased neuropsychiatric vulnerabilities and tendencies to-ward risk behavior. We observed that although certain measures ofcognitive performance were similar among adolescent rats performingan instrumental learning task, persistent differences were observedduring training (e.g., task-irrelevant and premature pokes) and extinc-tion, with adolescents exhibiting more perseverative behavior andmore sensitivity to the activating effects of internal and externalmotivational factors. By studying how adolescents respond to suchstimuli differently, we may learn more about the unique propensitiesand neuropsychiatric vulnerabilities of the period.
References
Adriani, W., Chiarotti, F., & Laviola, G. (1998). Elevated novelty seekingand peculiar d-amphetamine sensitization in periadolescent mice com-pared with adult mice. Behavioral Neuroscience, 112, 1152–1166.
Adriani, W., & Laviola, G. (2003). Elevated levels of impulsivity andreduced place conditioning with d-amphetamine: Two behavioral fea-tures of adolescence in mice. Behavioral Neuroscience, 117, 695–703.
Adriani, W., & Laviola, G. (2004). Windows of vulnerability to psycho-pathology and therapeutic strategy in the adolescent rodent model.Behavioural Pharmacology, 15, 341–352.
Andersen, S. L., Thompson, A. T., Rutstein, M., Hostetter, J. C., &Teicher, M. H. (2000). Dopamine receptor pruning in prefrontal cortexduring the periadolescent period in rats. Synapse, 37, 167–169.
Andrucci, G. L., Archer, R. P., Pancoast, D. L., & Gordon, R. A. (1989).The relationship of MMPI and Sensation Seeking Scales to adolescentdrug use. Journal of Personality Assessment, 53, 253–266.
Arnett, J. J. (1999). Adolescent storm and stress, reconsidered. AmericanPsychologist, 54, 317–326.
Badanich, K. A., Adler, K. J., & Kirstein, C. L. (2006). Adolescents differfrom adults in cocaine conditioned place preference and cocaine-induceddopamine in the nucleus accumbens septi. European Journal of Phar-macology, 550, 95–106.
Badanich, K. A., Maldonado, A. M., & Kirstein, C. L. (2008). Earlyadolescents show enhanced acute cocaine-induced locomotor activity incomparison to late adolescent and adult rats. Developmental Psychobi-ology, 50, 127–133.
Balleine, B. W., Liljeholm, M., & Ostlund, S. B. (2009). The integrativefunction of the basal ganglia in instrumental conditioning. BehaviouralBrain Research, 199, 43–52.
Benes, F. M., Taylor, J. B., & Cunningham, M. C. (2000). Convergenceand plasticity of monoaminergic systems in the medial prefrontal cortexduring the postnatal period: Implications for the development of psy-chopathology. Cerebral Cortex, 10, 1014–1027.
Benes, F. M., Turtle, M., Khan, Y., & Farol, P. (1994). Myelination of akey relay zone in the hippocampal formation occurs in the human brainduring childhood, adolescence, and adulthood. Archives of GeneralPsychiatry, 51, 477–484.
Berridge, K. C. (2007). The debate over dopamine’s role in reward: The casefor incentive salience. Psychopharmacology (Berlin), 191, 391–431.
Bolanos, C. A., Glatt, S. J., & Jackson, D. (1998). Subsensitivity to dopami-nergic drugs in periadolescent rats: A behavioral and neurochemical anal-ysis. Brain Research: Developmental Brain Research, 111, 25–33.
Brenhouse, H. C., & Andersen, S. L. (2008). Delayed extinction and strongerreinstatement of cocaine conditioned place preference in adolescent rats,compared to adults. Behavioral Neuroscience, 122, 460–465.
Brenhouse, H. C., Sonntag, K. C., & Andersen, S. L. (2008). Transient D1dopamine receptor expression on prefrontal cortex projection neurons:
Relationship to enhanced motivational salience of drug cues in adoles-cence. Journal of Neuroscience, 28, 2375–2382.
Carli, M., Robbins, T. W., Evenden, J. L., & Everitt, B. J. (1983). Effectsof lesions to ascending noradrenergic neurones on performance of a5-choice serial reaction task in rats: Implications for theories of dorsalnoradrenergic bundle function based on selective attention and arousal.Behavioural Brain Research, 9, 361–380.
Carr, K. D., Tsimberg, Y., Berman, Y., & Yamamoto, N. (2003). Evidenceof increased dopamine receptor signaling in food-restricted rats. Neuro-science, 119, 1157–1167.
Chambers, R. A., Taylor, J. R., & Potenza, M. N. (2003). Developmentalneurocircuitry of motivation in adolescence: A critical period of addic-tion vulnerability. American Journal of Psychiatry, 160, 1041–1052.
Compton, W. M., Thomas, Y. F., Conway, K. P., & Colliver, J. D. (2005).Developments in the epidemiology of drug use and drug use disorders.American Journal of Psychiatry, 162, 1494–1502.
Cools, R. (2008). Role of dopamine in the motivational and cognitivecontrol of behavior. Neuroscientist, 14, 381–395.
Corbit, L. H., & Balleine, B. W. (2003). The role of prelimbic cortex ininstrumental conditioning. Behavioural Brain Research, 146, 145–157.
Csikszentmihalyi, M., Larson, R., & Prescott, S. (1977). The ecology ofadolescent activity and experience. Journal of Youth and Adolescence, 6,281–294.
Cunningham, M. G., Bhattacharyya, S., & Benes, F. M. (2002). Amygdalo-cortical sprouting continues into early adulthood: Implications for thedevelopment of normal and abnormal function during adolescence.Journal of Comparative Neurology, 453, 116–130.
Cunningham, M. G., Bhattacharyya, S., & Benes, F. M. (2008). Increasinginteraction of amygdalar afferents with GABAergic interneurons be-tween birth and adulthood. Cerebral Cortex, 18, 1529–1535.
Darmani, N. A., Shaddy, J., & Gerdes, C. F. (1996). Differential ontogen-esis of three DOI-induced behaviors in mice. Physiology and Behavior,60, 1495–1500.
Doremus-Fitzwater, T. L., Varlinskaya, E. I., & Spear, L. P. (2009). Socialand non-social anxiety in adolescent and adult rats after repeated re-straint. Physiology and Behavior, 97, 484–494.
Doremus-Fitzwater, T. L., Varlinskaya, E. I., & Spear, L. P. (in press).Motivational systems in adolescence: Possible implications for agedifferences in substance abuse and other risk-taking behaviors. Brainand Cognition.
Douglas, L. A., Varlinskaya, E. I., & Spear, L. P. (2004). Rewardingproperties of social interactions in adolescent and adult male and femalerats: Impact of social versus isolate housing of subjects and partners.Developmental Psychobiology, 45, 153–162.
Ernst, M., & Fudge, J. L. (2009). A developmental neurobiological modelof motivated behavior: Anatomy, connectivity and ontogeny of thetriadic nodes. Neuroscience and Biobehavioral Reviews, 33, 367–382.
Ernst, M., Pine, D. S., & Hardin, M. (2006). Triadic model of the neuro-biology of motivated behavior in adolescence. Psychological Medicine,36, 299–312.
Floresco, S. B., & Magyar, O. (2006). Mesocortical dopamine modulationof executive functions: Beyond working memory. Psychopharmacology(Berlin), 188, 567–585.
Galvan, A., Hare, T. A., Parra, C. E., Penn, J., Voss, H., Glover, G., &Casey, B. J. (2006). Earlier development of the accumbens relative toorbitofrontal cortex might underlie risk-taking behavior in adolescents.Journal of Neuroscience, 26, 6885–6892.
Giedd, J. N., Blumenthal, J., Jeffries, N. O., Castellanos, F. X., Liu, H.,Zijdenbos, A., . . . Rapoport, J. L. (1999). Brain development duringchildhood and adolescence: A longitudinal MRI study. Nature Neuro-science, 2, 861–863.
Jung, M. W., Baeg, E. H., Kim, M. J., Kim, Y. B., & Kim, J. J. (2008).Plasticity and memory in the prefrontal cortex. Reviews in the Neuro-sciences, 19, 29–46.
24 STURMAN, MANDELL, AND MOGHADDAM

Juraska, J. M., & Markham, J. A. (2004). The cellular basis for volumechanges in the rat cortex during puberty: White and gray matter. Annalsof the New York Academy of Sciences, 1021, 431–435.
Kelley, A. E. (2004). Ventral striatal control of appetitive motivation: Rolein ingestive behavior and reward-related learning. Neuroscience andBiobehavioral Reviews, 27, 765–776.
Khuder, S. A., Dayal, H. H., & Mutgi, A. B. (1999). Age at smoking onsetand its effect on smoking cessation. Addictive Behaviors, 24, 673–677.
Koob, G. F. (2009). Dynamics of neuronal circuits in addiction: Reward,antireward, and emotional memory. Pharmacopsychiatry, 42(Suppl. 1),S32–S41.
Laviola, G., Adriani, W., Terranova, M. L., & Gerra, G. (1999). Psycho-biological risk factors for vulnerability to psychostimulants in humanadolescents and animal models. Neuroscience and Biobehavioral Re-views, 23, 993–1010.
Lewis, D. A. (1997). Development of the prefrontal cortex during adoles-cence: Insights into vulnerable neural circuits in schizophrenia. Neuro-psychopharmacology, 16, 385–398.
Little, P. J., Kuhn, C. M., Wilson, W. A., & Swartzwelder, H. S. (1996).Differential effects of ethanol in adolescent and adult rats. Alcoholism:Clinical and Experimental Research, 20, 1346–1351.
Macrı̀, S., Adriani, W., Chiarotti, F., & Laviola, G. (2002). Risk takingduring exploration of a plus-maze is greater in adolescent than injuvenile or adult mice. Animal Behaviour, 64, 541–546.
Mathews, I. Z., & McCormick, C. M. (2007). Female and male rats in lateadolescence differ from adults in amphetamine-induced locomotor ac-tivity, but not in conditioned place preference for amphetamine. Behav-ioural Pharmacology, 18, 641–650.
McCutcheon, J. E., & Marinelli, M. (2009). Age matters. European Jour-nal of Neuroscience, 29, 997–1014.
Meyer, G., Ferres-Torres, R., & Mas, M. (1978). The effects of puberty andcastration on hippocampal dendritic spines of mice: A Golgi study.Brain Research, 155, 108–112.
Mogenson, G. J., Jones, D. L., & Yim, C. Y. (1980). From motivation toaction: Functional interface between the limbic system and the motorsystem. Progress in Neurobiology, 14, 69–97.
Moy, S. S., Duncan, G. E., Knapp, D. J., & Breese, G. R. (1998).Sensitivity to ethanol across development in rats: Comparison to[3H]zolpidem binding. Alcoholism: Clinical and Experimental Re-search, 22, 1485–1492.
Nunez, J. L., Sodhi, J., & Juraska, J. M. (2002). Ovarian hormones afterpostnatal day 20 reduce neuron number in the rat primary visual cortex.Journal of Neurobiology, 52, 312–321.
Paus, T. (2005). Mapping brain maturation and cognitive developmentduring adolescence. Trends in Cognitive Sciences, 9, 60–68.
Paus, T., Collins, D. L., Evans, A. C., Leonard, G., Pike, B., & Zijdenbos,A. (2001). Maturation of white matter in the human brain: A review ofmagnetic resonance studies. Brain Research Bulletin, 54, 255–266.
Pautassi, R. M., Myers, M., Spear, L. P., Molina, J. C., & Spear, N. E.(2008). Adolescent but not adult rats exhibit ethanol-mediated appetitivesecond-order conditioning. Alcoholism: Clinical and Experimental Re-search, 32, 2016–2027.
Philpot, R. M., Badanich, K. A., & Kirstein, C. L. (2003). Place condi-tioning: Age-related changes in the rewarding and aversive effects ofalcohol. Alcoholism: Clinical and Experimental Research, 27, 593–599.
Philpot, R. M., & Wecker, L. (2008). Dependence of adolescent novelty-seeking behavior on response phenotype and effects of apparatus scal-ing. Behavioral Neuroscience, 122, 861–875.
Pine, D. S. (2002). Brain development and the onset of mood disorders.Seminars in Clinical Neuropsychiatry, 7, 223–233.
Pinos, H., Collado, P., Rodriguez-Zafra, M., Rodriguez, C., Segovia, S., &Guillamon, A. (2001). The development of sex differences in the locuscoeruleus of the rat. Brain Research Bulletin, 56, 73–78.
Robbins, T. W. (2002). The 5-choice serial reaction time task: Behaviouralpharmacology and functional neurochemistry. Psychopharmacology(Berlin), 163, 362–380.
Rosenberg, D. R., & Lewis, D. A. (1994). Changes in the dopaminergicinnervation of monkey prefrontal cortex during late postnatal develop-ment: A tyrosine hydroxylase immunohistochemical study. BiologicalPsychiatry, 36, 272–277.
Salamone, J. D., & Correa, M. (2002). Motivational views of reinforce-ment: Implications for understanding the behavioral functions of nucleusaccumbens dopamine. Behavioural Brain Research, 137, 3–25.
Seeman, P., Bzowej, N. H., Guan, H. C., Bergeron, C., Becker, L. E.,Reynolds, G. P., . . . Tourtellotte, W. W. (1987). Human brain dopaminereceptors in children and aging adults. Synapse, 1, 399–404.
Shalaby, I. A., & Spear, L. P. (1980). Chronic administration of haloperidol duringdevelopment: Later psychopharmacological responses to apomorphine andarecoline. Pharmacology, Biochemistry and Behavior, 13, 685–690.
Shram, M. J., Funk, D., Li, Z., & Le, A. D. (2006). Periadolescent and adultrats respond differently in tests measuring the rewarding and aversiveeffects of nicotine. Psychopharmacology (Berlin), 186, 201–208.
Sisk, C. L., & Zehr, J. L. (2005). Pubertal hormones organize the adoles-cent brain and behavior. Frontiers in Neuroendocrinology, 26, 163–174.
Spear, L. P. (2000). The adolescent brain and age-related behavioralmanifestations. Neuroscience and Biobehavioral Reviews, 24, 417–463.
Spear, L. P., & Brake, S. C. (1983). Periadolescence: Age-dependentbehavior and psychopharmacological responsivity in rats. Developmen-tal Psychobiology, 16, 83–109.
Stansfield, K. H., & Kirstein, C. L. (2006). Effects of novelty onbehavior in the adolescent and adult rat. Developmental Psychobiol-ogy, 48, 10 –15.
Steinberg, L. (2008). A social neuroscience perspective on adolescentrisk-taking. Developmental Review, 28, 78–106.
Tarazi, F. I., & Baldessarini, R. J. (2000). Comparative postnatal develop-ment of dopamine D(1), D(2) and D(4) receptors in rat forebrain.International Journal of Developmental Neuroscience, 18, 29–37.
Teicher, M. H., Andersen, S. L., & Hostetter, J. C., Jr. (1995). Evidence fordopamine receptor pruning between adolescence and adulthood in stri-atum but not nucleus accumbens. Brain Research: Developmental BrainResearch, 89, 167–172.
Teicher, M. H., Krenzel, E., Thompson, A. P., & Andersen, S. L. (2003). Dopa-mine receptor pruning during the peripubertal period is not attenuated byNMDA receptor antagonism in rat. Neuroscience Letters, 339, 169–171.
Van Leijenhorst, L., Zanolie, K., Van Meel, C. S., Westenberg, P. M.,Rombouts, S. A., & Crone, E. A. (2010). What motivates the adolescent?Brain regions mediating reward sensitivity across adolescence. CerebralCortex, 20, 61–69.
Varlinskaya, E. I., & Spear, L. P. (2006). Ontogeny of acute tolerance toethanol-induced social inhibition in Sprague-Dawley rats. Alcoholism:Clinical and Experimental Research, 30, 1833–1844.
Vastola, B. J., Douglas, L. A., Varlinskaya, E. I., & Spear, L. P. (2002).Nicotine-induced conditioned place preference in adolescent and adultrats. Physiology and Behavior, 77, 107–114.
Volkmar, F. R. (1996). Childhood and adolescent psychosis: A review ofthe past 10 years. Journal of the American Academy of Child & Ado-lescent Psychiatry, 35, 843–851.
Wilmouth, C. E., & Spear, L. P. (2009). Hedonic sensitivity in adolescentand adult rats: Taste reactivity and voluntary sucrose consumption.Pharmacology, Biochemistry and Behavior, 92, 566–573.
Received November 6, 2009Revision received December 1, 2009
Accepted December 2, 2009 �
25ADOLESCENT INSTRUMENTAL LEARNING AND EXTINCTION
Related Documents











