Biomaterials 28 (2007) 3834–3842 Adipose tissue engineering with naturally derived scaffolds and adipose-derived stem cells Lauren Flynn a,b , Glenn D. Prestwich c , John L. Semple d,e , Kimberly A. Woodhouse a,b, a Department of Chemical Engineering and Applied Chemistry, University of Toronto, 200 College Street, Toronto, Ont., Canada, M5S 3E5 b Institute of Biomaterials and Biomedical Engineering, University of Toronto, 4 Taddle Creek Road, Toronto, Ont., Canada, M5S 3G9 c Center for Therapeutic Biomaterials and Department of Medicinal Chemistry, University of Utah, 419 Wakara Way, Suite 205, Salt Lake City, Utah 84108-12, USA d Division of Plastic Surgery, Department of Surgery, University of Toronto, 100 College Street, Toronto, Ont., Canada, M5G 1L5 e Women’s College Hospital, 76 Grenville Street, Toronto, Ont., Canada, M5S 1B2 Received 28 February 2007; accepted 4 May 2007 Available online 16 May 2007 Abstract A tissue-engineered adipose substitute would have numerous applications in plastic and reconstructive surgery. This work involves the characterization of the in vitro cellular response of primary human adipose-derived stem cells (ASC) to three dimensional, naturally derived scaffolds. To establish a more thorough understanding of the influence of the scaffold environment on ASC, we have designed several different soft tissue scaffolds composed of decellularized human placenta and crosslinked hyaluronan (XLHA). The cellular organization within the scaffolds was characterized using confocal microscopy. Adipogenic differentiation was induced and the ASC response was characterized in terms of glycerol-3-phosphate dehydrogenase (GPDH) activity and intracellular lipid accumulation. The results indicate that the scaffold environment impacts the ASC response and that the adipogenic differentiation of the ASC was augmented in the non-adhesive XLHA gels. r 2007 Elsevier Ltd. All rights reserved. Keywords: Adipose tissue engineering; Scaffold; Extracellular matrix; Hyaluronan; Stem cells 1. Introduction Soft tissue augmentation is a major challenge for plastic and reconstructive surgeons. Subcutaneous adipose tissue loss is associated with numerous conditions including traumatic injury, oncologic resection and congenital birth defects [1]. The resultant scar tissue formation can lead to a loss of contour, as well as functional impairment, particularly if the defect occurs in proximity to a joint [2]. In 2005, approximately 5.4 million people underwent reconstructive surgery in the United States, with 3.9 million cases associated with tumor removal [3]. In addition, over 10.2 million cosmetic procedures were performed, includ- ing various forms of soft tissue augmentation with biological or synthetic fillers and implants [3]. The current treatment strategies for soft tissue recon- struction primarily involve tissue transplantation, includ- ing composite tissue flaps, or synthetic substitutes. The use of autologous tissues is associated with the creation of a donor site defect and, potentially, the need for multiple, complex and costly surgical procedures [4]. Moreover, the clinical outcome of adipose tissue transplantation is unpredictable, with graft resorption due to a lack of vascularization resulting in poor cosmesis and impaired functionality [5]. Synthetic implants are associated with immune rejection, implant migration and resorption, and a failure to integrate into the host tissues [6]. A tissue-engineered adipose substitute, that would promote regeneration, rather than repair, would be invaluable to plastic and reconstructive surgeons. The substitute should incorporate a biocompatible scaffold that ARTICLE IN PRESS www.elsevier.com/locate/biomaterials 0142-9612/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.biomaterials.2007.05.002 Corresponding author. Department of Chemical Engineering and Applied Chemistry, University of Toronto, 200 College Street, Toronto, Ont., Canada, M5S 3E5. Tel.: +416 978 3060; fax: +416 978 8605. E-mail address: [email protected] (K.A. Woodhouse).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
0142-9612/$ - se
doi:10.1016/j.bi
�CorrespondApplied Chemi
Ont., Canada,
E-mail addr
Biomaterials 28 (2007) 3834–3842
www.elsevier.com/locate/biomaterials
Adipose tissue engineering with naturally derived scaffolds andadipose-derived stem cells
Lauren Flynna,b, Glenn D. Prestwichc, John L. Sempled,e, Kimberly A. Woodhousea,b,�
aDepartment of Chemical Engineering and Applied Chemistry, University of Toronto, 200 College Street, Toronto, Ont., Canada, M5S 3E5bInstitute of Biomaterials and Biomedical Engineering, University of Toronto, 4 Taddle Creek Road, Toronto, Ont., Canada, M5S 3G9
cCenter for Therapeutic Biomaterials and Department of Medicinal Chemistry, University of Utah, 419 Wakara Way, Suite 205, Salt Lake City,
Utah 84108-12, USAdDivision of Plastic Surgery, Department of Surgery, University of Toronto, 100 College Street, Toronto, Ont., Canada, M5G 1L5
eWomen’s College Hospital, 76 Grenville Street, Toronto, Ont., Canada, M5S 1B2
Received 28 February 2007; accepted 4 May 2007
Available online 16 May 2007
Abstract
A tissue-engineered adipose substitute would have numerous applications in plastic and reconstructive surgery. This work involves the
characterization of the in vitro cellular response of primary human adipose-derived stem cells (ASC) to three dimensional, naturally
derived scaffolds. To establish a more thorough understanding of the influence of the scaffold environment on ASC, we have designed
several different soft tissue scaffolds composed of decellularized human placenta and crosslinked hyaluronan (XLHA). The cellular
organization within the scaffolds was characterized using confocal microscopy. Adipogenic differentiation was induced and the ASC
response was characterized in terms of glycerol-3-phosphate dehydrogenase (GPDH) activity and intracellular lipid accumulation. The
results indicate that the scaffold environment impacts the ASC response and that the adipogenic differentiation of the ASC was
augmented in the non-adhesive XLHA gels.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Adipose tissue engineering; Scaffold; Extracellular matrix; Hyaluronan; Stem cells
1. Introduction
Soft tissue augmentation is a major challenge for plasticand reconstructive surgeons. Subcutaneous adipose tissueloss is associated with numerous conditions includingtraumatic injury, oncologic resection and congenital birthdefects [1]. The resultant scar tissue formation can lead to aloss of contour, as well as functional impairment,particularly if the defect occurs in proximity to a joint[2]. In 2005, approximately 5.4 million people underwentreconstructive surgery in the United States, with 3.9 millioncases associated with tumor removal [3]. In addition, over10.2 million cosmetic procedures were performed, includ-
e front matter r 2007 Elsevier Ltd. All rights reserved.
omaterials.2007.05.002
ing author. Department of Chemical Engineering and
stry, University of Toronto, 200 College Street, Toronto,
M5S 3E5. Tel.: +416 978 3060; fax: +416 978 8605.
ess: [email protected] (K.A. Woodhouse).
ing various forms of soft tissue augmentation withbiological or synthetic fillers and implants [3].The current treatment strategies for soft tissue recon-
struction primarily involve tissue transplantation, includ-ing composite tissue flaps, or synthetic substitutes. The useof autologous tissues is associated with the creation of adonor site defect and, potentially, the need for multiple,complex and costly surgical procedures [4]. Moreover, theclinical outcome of adipose tissue transplantation isunpredictable, with graft resorption due to a lack ofvascularization resulting in poor cosmesis and impairedfunctionality [5]. Synthetic implants are associated withimmune rejection, implant migration and resorption, and afailure to integrate into the host tissues [6].A tissue-engineered adipose substitute, that would
promote regeneration, rather than repair, would beinvaluable to plastic and reconstructive surgeons. Thesubstitute should incorporate a biocompatible scaffold that

ARTICLE IN PRESSL. Flynn et al. / Biomaterials 28 (2007) 3834–3842 3835
defines the appropriate three-dimensional tissue architec-ture and promotes host integration and implant vascular-ization [7]. Ultimately, the construct should degrade as it isreplaced by healthy host soft tissue. A number of differentsynthetic scaffolds have been investigated for adipose tissueengineering applications including polyethylene glycoldiacrylate (PEDGA) [8], polyglycolic acid (PGA) [9],polyethylene terephthalate (PET) [10], poly(lactic-co-gly-colic acid) (PLGA) [11] and polytetrafluoroethylene(PTFE) [12]. Naturally derived materials such as collagen[13], derivatives of hyaluronic acid [14], matrigel [15] andfibrin [16] have also been studied. There are many factorsto consider when designing a scaffold including themechanical properties, degradation characteristics, immu-nogenicity, cellular response to the material, ease ofhandling in the clinic and cost [17].
Adipose-derived stem cells (ASC) may be an idealautologous cell source for adipose tissue engineering [18].ASC are much more resistant to mechanical damage andischemic conditions than mature adipocytes [19]. Thecells, which can be readily harvested from excisedhuman subcutaneous fat or liposuction samples, havebeen shown to proliferate rapidly and differentiate intomature adipocytes both in vitro and in vivo [20–22].The development of methods to maintain the ASCdifferentiation potential in culture, while obtaining suffi-cient cell populations for transplantation, will be criticalto the clinical application of these cells [23]. Further,the optimization of the growth and differentiation condi-tions to maximize stable adipose tissue formation isrequired [24].
With a view to develop a tissue-engineered adiposesubstitute, we are investigating the response of human ASCto scaffolds comprised of placental decellular matrix(PDM) and crosslinked hyaluronan (XLHA). By investi-gating several different scaffolds, it is possible to obtain amore thorough understanding of the impact of the matrixenvironment on the ASC. We previously developed adetergent and enzymatic extraction protocol to fullydecellularize human placenta [25]. We believe that thePDM holds promise as a scaffold for adipose tissueengineering applications. The placenta is a rich source ofhuman extracellular matrix (ECM) components that can beharvested without harm to the donor. Constructs derivedfrom the ECM may mimic the native environment of thebody, promoting normal cellular organization and beha-vior. Natural materials also have advantages in terms ofease of processing, biodegradability and biocompatibility[26]. Hyaluronan (HA) is a highly conserved glycosami-noglycan (GAG) that functions in matrix stabilization, cellsignaling, adhesion, migration, proliferation and differen-tiation [27]. Incorporation of XLHA into the PDMscaffolds may improve the construct bulking propertiesand may influence cellular infiltration, differentiation andwound healing [28].
The primary objective of this research was to developeffective seeding protocols and in vitro culture conditions
for the ASC with the PDM and XLHA. The cellularorganization was characterized in each of the scaffolds.The influence of the scaffold environment on the differ-entiation of the ASC was also investigated.
2. Materials and methods
2.1. Materials
All chemicals used in the experiments, unless otherwise stated, were
purchased from Sigma-Aldrich Canada Ltd. (Oakville, Canada) and were
used as received. Water was distilled and deionized using a Millipore Milli-
RO 10 Plus filtration system at 18MO resistance.
2.2. Cell culture
Primary cultures of human ASC were established according to the
methods of Flynn et al. [25]. The ASC were isolated from freshly excised
subcutaneous abdominal adipose tissue from patients undergoing elective
surgery at Women’s College Hospital, Toronto, Canada, following the
ethical guidelines of the University of Toronto. The cells were plated in a
1:1 mixture of Dulbecco’s Modified Eagle’s Medium and Ham’s F-12
nutrient mixture (DMEM:Ham’s F-12), supplemented with 10% fetal
bovine serum (FBS), 100U/mL penicillin and 0.1mg/mL streptomycin.
The growth medium was changed every 2–3 days. To passage the cells,
cultures at 90% confluence were trypsin-released (0.25% trypsin/0.1%
EDTA, Gibco, Burlington, Canada), washed, counted and re-plated in
new flasks at 30,000 cells/cm2. Passage 2 cells were used for the seeding
experimentation.
2.3. ASC differentiation
To induce adipogenic differentiation, the cells were cultured in serum-
free DMEM:Ham’s F-12 supplemented with 15mM NaHCO3, 15mM
HEPES, 33mM biotin, 17mM pantothenate, 10 mg/mL transferrin, 100 nM
cortisol, 66 nM insulin, 1 nM triiodothyronine (T3), 100U/mL penicillin
and 0.1mg/mL streptomycin. For the first 72 h of differentiation, 0.25mM
isobutylmethylxanthine (IBMX) and 1mg/mL of troglitazone were added
to the differentiation medium [29].
2.4. Preparation of the PDM
Placentas were obtained, with informed consent, from normal-term
Caesarian-section deliveries at Women’s College Hospital, Toronto,
Canada and were decellularized as previously described [25]. In brief,
the perfusive and diffusive decellularization protocol involved treatment
with hypotonic and hypertonic solutions, enzymatic digestion, and
multiple detergent extractions. Research ethics board approval for this
study was obtained from Women’s College Hospital, Toronto, Canada
(REB # 9918). Histological analysis was conducted on representative
sections of the processed tissues to confirm the effectiveness of the
extraction protocol.
Following decellularization, the PDM was sectioned into samples by
mass, with each scaffold consisting of a 300mg portion of the villous tree
network. The scaffolds were decontaminated by three 30min rinses in
ethanol, re-hydrated with three washes in sterile phosphate buffered saline
(PBS) and stored at 4 1C in sterile PBS supplemented with 100U/mL
penicillin and 0.1mg/mL streptomycin.
The PDM scaffolds that were used in the confocal analyses were labeled
with an amine reactive Alexa Fluors 350 carboxylic acid, succinimidyl
ester (Molecular Probes, Burlington, Canada) to facilitate visualization.
Briefly, the dye was dissolved in dimethyl sulfoxide (DMSO) at a
concentration of 10mg/mL. This stock solution was diluted with 0.15M
NaHCO3 (pH 8.3) to obtain a working concentration of 0.3mg/mL. The
PDM scaffolds were agitated in the dye solution for 1 h at room

ARTICLE IN PRESSL. Flynn et al. / Biomaterials 28 (2007) 3834–38423836
temperature (0.3mg dye/300mg PDM). To stop the labelling reaction, the
PDM was incubated for an additional hour in 1.5M hydroxylamine (pH
8.5). The scaffolds were then rinsed three times in sterile PBS and stored at
4 1C in sterile PBS supplemented with 100U/mL penicillin and 0.1mg/mL
streptomycin. All work was conducted under minimal lighting conditions.
2.5. Thiolated HA preparation
Thiol-modified HA (HA-DTPH) was synthesized according to the
methods of Shu et al. [30]. GPC and 1H NMR was used to assess the
purity and degree of substitution (SD) of the HA-DTPH. The HA-DTPH
used in the experimentation had an SD ¼ 57%.
2.6. Scaffold fabrication
Four different scaffold groups were investigated to characterize the
influence of the scaffold environment on the ASC population: the PDM
alone, the XLHA alone and the PDM in combination with the XLHA
using two different seeding methodologies.
Prior to cell seeding, the PDM was incubated for 24 h in serum-free
DMEM:Ham’s F-12 (37 1C, 5% CO2) supplemented with 100U/mL
penicillin and 0.1mg/mL streptomycin. Passage 2 ASC at 90% confluence
were trypsin-released, washed and counted using trypan blue exclusion.
For each of the scaffold groups, the cellular seeding density was 1� 106
cells/scaffold. This seeding density was selected based on our previous in
vitro work that indicated that higher seeding densities promoted ASC
viability on the PDM scaffolds in culture [25].
2.6.1. PDM scaffold fabrication
The ASC were suspended in growth medium at a concentration of
2� 107 cells/mL. Each PDM scaffold was transferred to a 24-well plate
and seeded with 1� 106ASC in 50mL of growth medium. The scaffolds
were incubated for 3 h (37 1C, 5% CO2) to facilitate cell adhesion and then
transferred into 6-well plates containing growth medium (5mL/well).
2.6.2. XLHA scaffold fabrication
The XLHA hydrogels were fabricated according to the methods of Shu
et al [31]. This methodology uses polyethylene glycol-diacrylate (PEGDA;
MW 3400; Nektar Transforming Therapeutics, Huntsville AL) to rapidly
crosslink the HA-DTPH to form a gel in situ. Briefly, a 12.5mg/mL
solution of HA–DTPH was prepared in DMEM:Ham’s F-12 growth
medium and the pH was adjusted to 7.4 with 1.0M NaOH. A 4.5% (w/v)
Fig. 1. The two seeding methodologies for th
solution of the PEGDA was prepared in PBS. The solutions were sterilized
by syringe filtration with a 0.22mm filter. To initiate crosslinking, the
PEGDA solution was added to the HA–DTPH solution in a ratio of 1:4
(v:v) and mixed thoroughly. Immediately following mixing, the ASC were
resuspended in the HA–DTPH–PEGDA solution at a concentration of
3.33� 106 cells/mL. To fabricate each scaffold with a seeding density of
1� 106 cells, 300mL of the cell-seeded HA–DTPH–PEGDA was added to
a well in a 24-well plate and allowed to gel for 1 h (37 1C, 5% CO2). The
scaffolds were then gently detached from the surface using blunt forceps
and transferred into 6-well plates containing growth medium (5mL/well).
2.6.3. PDM with XLHA scaffold fabrication
Two different seeding methodologies (Fig. 1) were investigated to assess
whether there was a significant impact on the organization and
differentiation of the ASC in the combined PDM with XLHA scaffolds.
The first method was termed the PDM with XLHA ‘‘Encapsulation’’
methodology. The 300mg PDM scaffolds were placed in a 24-well plate
and an HA–DTPH–PEGDA solution was prepared, as described in the
previous section. Immediately following mixing, the ASC were resus-
pended in the HA–DTPH–PEGDA at a concentration of 4� 106 cells/mL.
For each PDM sample, 250mL of the cell-seeded HA–DTPH–PEGDA
was added and allowed to gel for 1 h (37 1C, 5% CO2), to fabricate a
combined construct seeded with 1� 106 cells. The scaffolds were then
transferred into 6-well plates containing growth medium (5mL/well).
The second seeding method was defined as the PDM with XLHA
‘‘Adhesion’’ methodology. The ASC were suspended in growth medium at
a concentration of 2� 107 cells/mL. The 300mg PDM scaffolds were
placed in a 24-well plate and each scaffold was seeded with 1� 106ASC in
50mL of growth medium. The PDM scaffolds were incubated (37 1C, 5%
CO2) for 2 h to facilitate cell adhesion. Following this, an HA–DTPH–
PEGDA solution was prepared, as described in the previous section. A
250mL volume of the HA–DTPH–PEGDA solution was added to each
cell-seeded PDM, and the constructs were incubated (37 1C, 5% CO2) for
an additional hour to facilitate gelation. The combined scaffolds were then
transferred into 6-well plates containing growth medium (5mL/well).
2.6.4. Scaffold culture
Following seeding, the scaffolds were incubated (37 1C, 5% CO2) under
agitation on a rotomix in the 6-well plates containing growth medium. The
medium was changed 24 h after seeding and every 2 days thereafter. To
investigate cellular differentiation within the scaffolds, after a 72-h growth
period, the scaffolds were rinsed in sterile PBS and transferred into the
previously defined differentiation medium. The constructs were incubated
e combined PDM with XLHA scaffolds.

ARTICLE IN PRESS
Fig. 2. The macroscopic architecture of the PDM scaffolds, XLHA
scaffolds and PDM combined with XLHA scaffolds. Bars represent 5mm.
L. Flynn et al. / Biomaterials 28 (2007) 3834–3842 3837
(37 1C, 5% CO2) under agitation for up to 14 days after the induction of
differentiation.
2.7. Cellular organization
Cellular organization in the scaffolds and on tissue culture polystyrene
(TCPS) controls was assessed at 24 and 72 h after seeding using confocal
microscopy (Zeiss LSM510, FLUAR 20x/0.75 NA objective lens,
excitation with an Argon laser at 488 nm and an Enterprise laser at
351 nm). The ASC were labeled prior to seeding with Cell TrackerTM
Green 5-chloromethylfluorescein diacetate (CMFDA) (Invitrogen, Bur-
lington, Canada) according to the manufacturer’s instructions. The PDM
scaffolds were labeled with the Alexa Fluors 350 carboxylic acid,
succinimidyl ester, as described in Section 2.4. All work was conducted
under minimal lighting conditions.
Samples from the XLHA scaffold group were also stained with calcein-
AM and ethidium homodimer-1 (LIVE/DEADs Viability/Cytotoxicity
Kit, Invitrogen, Burlington, Canada) to differentiate between live and
dead cells, respectively. Live/dead staining was not conducted on the
scaffolds incorporating the PDM, as the ethidium homodimer-1 bound
strongly to the PDM matrix, interfering with the imaging process.
2.8. Glycerol-3-phosphate dehydrogenase (GPDH) activity
To assess the impact of the scaffold microenvironment on the
differentiation of the cells, the cellular GPDH activity levels in each of
the scaffold groups were measured at 72 h, 7 and 14 days after the
induction of differentiation (n ¼ 3, N ¼ 6). The positive control was ASC
differentiated on TCPS and the negative control was undifferentiated ASC
on TCPS. For comparative purposes, the assay was also conducted on
samples of freshly-excised human subcutaneous adipose tissue from 4
different donors.
A GPDH activity measurement kit (Kamiya Biomedical Corporation)
was used in the quantification. Briefly, this kit provides a substrate reagent
containing NADH, which is a co-enzyme for GPDH, and dihydroxyace-
tone phosphate (DHAP). The activity level was quantified by spectro-
photometrically measuring the decrease in the NADH concentration as
the DHAP was converted into glycerol-3-phosphate by the GPDH. Within
each sample, the absorbance (340 nm) was measured over a 10min period
at 25 1C using a microplate reader. The change in absorbance (DOD/min)
was determined from the linear portion of the kinetic curve and each
sample was assayed in duplicate. The activity levels were normalized in
terms of the total cytosolic protein content within each sample, measured
in triplicate using the Bio-Rad protein assay, with an albumin standard.
The data was expressed in terms of mUnits/mg protein (mU/mg), where 1
unit was defined as the GPDH activity required to oxidize 1 mmol of
NADH in 1min.
In preparation for the GPDH and protein assays, each scaffold or
adipose tissue sample was minced in 1mL of 0.25M sucrose solution at
4 1C and disrupted with 3 five-s bursts of sonication, with intervals of
cooling on ice. The samples were centrifuged at 16,000g for 10min (4 1C).
The supernatant was transferred into a new 2mL tube and centrifuged for
an additional hour (16,000g, 4 1C) to isolate the cytosolic protein fraction,
including the GPDH. The TCPS control samples were disrupted with
sonication in the enzyme extraction buffer provided in the GPDH assay
kit, and centrifuged at 16,000g (4 1C) for 10min. All supernatant samples
were immediately assayed for GPDH activity and total protein content
according to the manufacturers’ instructions.
2.9. Oil red O staining
Lipid accumulation within the scaffolds was visually assessed using oil
red O staining at 7 and 14 days after the induction of differentiation. The
PDM scaffolds were labeled prior to seeding with the Alexa Fluors 350
carboxylic acid, succinimidyl ester, as described in Section 2.4. At each
time point, two scaffolds from each group were fixed for 24 h in 10%
neutral buffered formalin and rinsed thoroughly with PBS. Control
samples of differentiated and undifferentiated ASC on TCPS were
formalin-fixed for 5min, followed by extensive PBS rinsing.
An oil red O stock solution (3 g/L) was prepared in isopropanol and
was diluted 3:2 (v:v) with deionized water. This solution was agitated at
room temperature for 10min and then filtered through a type 1 qualitative
filter paper. Each scaffold was incubated in 5mL of the prepared stain for
2 h under agitation at room temperature. The TCPS control samples were
stained for 5min. All samples were rinsed thoroughly overnight in
deionized water, with three solution changes to reduce non-specific
staining. The scaffolds were analyzed using confocal microscopy (Zeiss
LSM510, C-APO 63x/1.2 NA water immersion (DIC) objective lens,
excitation with a HeNe laser at 543 nm and an Enterprise laser at 351 nm)
and the control samples were visualized using light microscopy (Zeiss
Axiovert 200M).
3. Results
3.1. Macroscopic scaffold architecture
The macroscopic architecture of each of the scaffoldgroups is shown in Fig. 2. All of the scaffolds wereapproximately 1.5 cm in diameter immediately followingfabrication. The PDM alone scaffolds expanded duringculture into a more random architecture. The XLHA aloneconstructs were difficult to handle and tore easily. Thecombined PDM with XLHA scaffolds were significantlystronger, could be easily handled with forceps andmaintained their structure throughout the entire cultureperiod.
3.2. Cellular organization
Representative images of the cellular organization at 24and 72 h post-seeding are shown in Fig. 3. On the TCPS,the cells had the expected fibroblastic morphology. On thePDM scaffolds, the cells adhered to the matrix andextended. Qualitatively, there were more adherent cells at72 h, potentially due to cell proliferation. In the XLHAgels, the cells were rounded and formed self-aggregates.Live/dead staining revealed that the majority of these cellswere alive at the 24-h time point, despite the roundedcellular morphology (Fig. 4). By the 72-h time point, therewere more dead cells present, both individually and withinthe aggregates. In the PDM with XLHA Encapsulationsamples, most of the cells could be found within the XLHAportion of the scaffold, with only a small percentageadhering to the PDM. In contrast, when the PDM withXLHA Adhesion seeding methodology was used, the

ARTICLE IN PRESS
Fig. 3. The cellular organization of the ASC in the scaffolds at 24 and 72 h after seeding, before the induction of differentiation. Prior to seeding, the cells
were labeled with Cell TrackerTM Green and the PDM was labeled with an amine reactive Alexa Fluors 350 carboxylic acid, succinimidyl ester.
Visualization was conducted using confocal microscopy (blue ¼ PDM, green ¼ cells). The PDM with XLHA Adhesion seeding methodology
(PDM+XLHA Adh.) promoted greater adhesion of the ASC to the PDM scaffold than the PDM with XLHA Encapsulation methodology
(PDM+XLHA Enc.). Bars represent 50mm.
Fig. 4. LIVE/DEAD staining of the ASC in the XLHA gels at 24 and 72 h
after seeding. Visualization using confocal microscopy (green ¼ live cells
(calcein-AM), red ¼ dead cells (ethidium homodimer-1)). In general, more
dead cells were present both individually and within the cellular aggregates
at the 72-h time point. Bars represent 50mm.
L. Flynn et al. / Biomaterials 28 (2007) 3834–38423838
majority of the cells were located in close proximity to thePDM and many had an extended morphology. With bothseeding methods, more adherent and extended cells couldbe visualized on the PDM at 72 h (Fig. 3).
3.3. GPDH activity
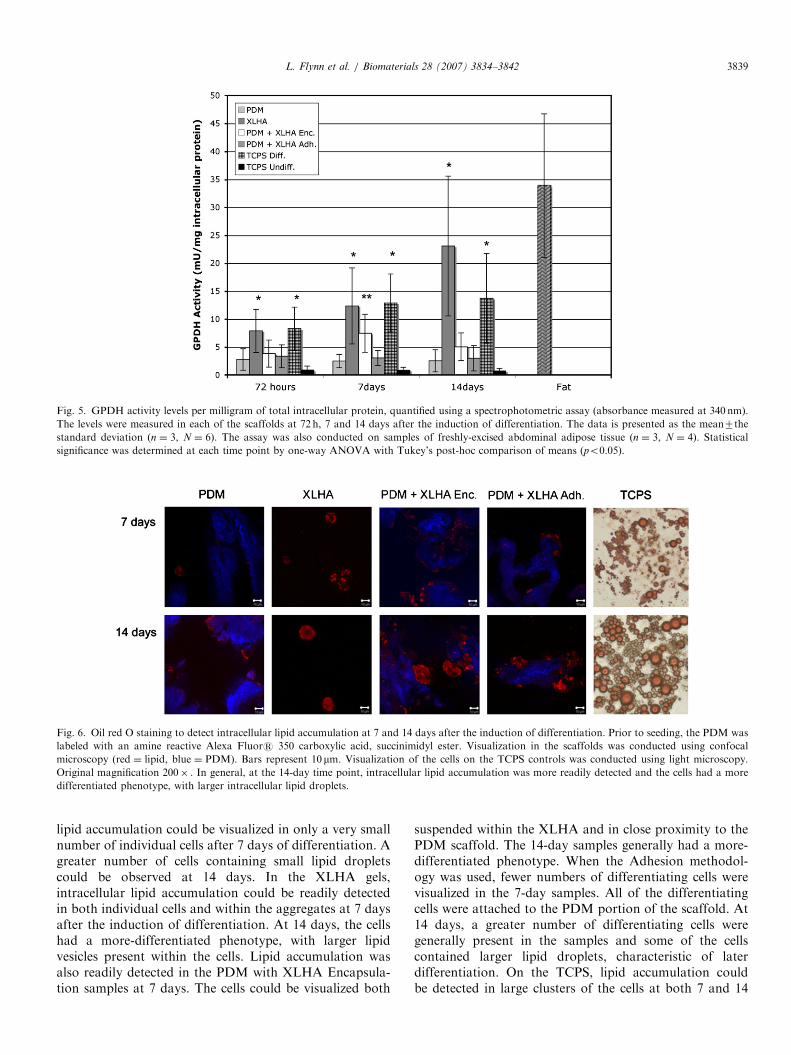
The average measured GPDH activity levels are shownin Fig. 5. The values are expressed as the mean7thestandard deviation. Statistical significance was determinedusing a one-way ANOVA with a Tukey’s post-hoccomparison of means (po0.05). Donor-specific activitylevels of 20.4772.63, 30.6375.67, 34.6179.23 and56.5376.13mU/mg were measured in the mature adiposetissue samples, with a mean value of 33.86712.84mU/mg.
At 72 h after the induction of differentiation, the cells inthe XLHA scaffolds (7.9073.85mU/mg) and on the TCPSpositive controls (8.2873.87mU/mg) had statistically
higher activity levels than the other groups. Whilenot statistically significant, the cells in the PDM(2.7871.04mU/mg), the PDM with XLHA Encapsulation(3.8473.42mU/mg) and the PDM with XLHA Adhesion(3.3472.04mU/mg) scaffolds, all had higher mean activitylevels than the non-induced TCPS negative control samples(0.8870.74mU/mg).At 7 days after the induction of differentiation, increased
activity levels, as compared to the 72-h samples, wereobserved in the XLHA, PDM with XLHA Encapsulationand TCPS positive control samples. The cells in the XLHAscaffolds (12.3576.80mU/mg) and the TCPS positivecontrols (12.8975.19mU/mg) had statistically higheractivity levels than all of the other sample groups. Thecells in the PDM with XLHA Encapsulation scaffolds(7.4773.38mU/mg) had statistically higher activity levelsthan the cells in the PDM (2.5071.18mU/mg), PDM withXLHA Adhesion (3.0671.35mU/mg) and TCPS negativecontrol (0.8470.54mU/mg) samples.At 14 days after the induction of differentiation, the
highest activity levels were observed in the cells fromthe XLHA scaffolds (23.07712.50mU/mg), approachingthe levels observed in mature adipose tissue. The XLHAand TCPS positive control (13.7478.00mU/mg) sampleshad statistically higher activity levels than all of theother groups. While not statistically significant, thecells in the PDM with XLHA Encapsulation scaffolds(5.0572.49mU/mg) had a higher mean activity level thanthe cells in the PDM (2.5371.98mU/mg), PDM withXLHA Adhesion (3.0072.28mU/mg) and TCPS negativecontrol (0.7170.47mU/mg) samples.
3.4. Oil red O staining
Representative images of the Oil Red O stained scaffoldsand controls are shown in Fig. 6. In the PDM scaffolds,
ARTICLE IN PRESS
Fig. 5. GPDH activity levels per milligram of total intracellular protein, quantified using a spectrophotometric assay (absorbance measured at 340 nm).
The levels were measured in each of the scaffolds at 72 h, 7 and 14 days after the induction of differentiation. The data is presented as the mean7the
standard deviation (n ¼ 3, N ¼ 6). The assay was also conducted on samples of freshly-excised abdominal adipose tissue (n ¼ 3, N ¼ 4). Statistical
significance was determined at each time point by one-way ANOVA with Tukey’s post-hoc comparison of means (po0.05).
Fig. 6. Oil red O staining to detect intracellular lipid accumulation at 7 and 14 days after the induction of differentiation. Prior to seeding, the PDM was
labeled with an amine reactive Alexa Fluors 350 carboxylic acid, succinimidyl ester. Visualization in the scaffolds was conducted using confocal
microscopy (red ¼ lipid, blue ¼ PDM). Bars represent 10 mm. Visualization of the cells on the TCPS controls was conducted using light microscopy.
Original magnification 200� . In general, at the 14-day time point, intracellular lipid accumulation was more readily detected and the cells had a more
differentiated phenotype, with larger intracellular lipid droplets.
L. Flynn et al. / Biomaterials 28 (2007) 3834–3842 3839
lipid accumulation could be visualized in only a very smallnumber of individual cells after 7 days of differentiation. Agreater number of cells containing small lipid dropletscould be observed at 14 days. In the XLHA gels,intracellular lipid accumulation could be readily detectedin both individual cells and within the aggregates at 7 daysafter the induction of differentiation. At 14 days, the cellshad a more-differentiated phenotype, with larger lipidvesicles present within the cells. Lipid accumulation wasalso readily detected in the PDM with XLHA Encapsula-tion samples at 7 days. The cells could be visualized both
suspended within the XLHA and in close proximity to thePDM scaffold. The 14-day samples generally had a more-differentiated phenotype. When the Adhesion methodol-ogy was used, fewer numbers of differentiating cells werevisualized in the 7-day samples. All of the differentiatingcells were attached to the PDM portion of the scaffold. At14 days, a greater number of differentiating cells weregenerally present in the samples and some of the cellscontained larger lipid droplets, characteristic of laterdifferentiation. On the TCPS, lipid accumulation couldbe detected in large clusters of the cells at both 7 and 14

ARTICLE IN PRESSL. Flynn et al. / Biomaterials 28 (2007) 3834–38423840
days. In general, the cells had a more-differentiatedphenotype at the 14-day time point.
4. Discussion
Advances in obesity and metabolic research have helpedto elucidate the cellular process of adipogenesis [32,33].While this knowledge could benefit the field of adiposetissue engineering, most of these studies were conductedunder two-dimensional (2D) growth conditions, with thecells grown in monolayer culture on TCPS. However, thisenvironment does not accurately mimic the complex 3Dnetwork of the body [34]. Significant research remains to beconducted in order to understand the ideal scaffoldenvironment for adipose tissue engineering applications.While a number of different synthetic and naturally derivedmaterials have been studied both in vitro and in vivo, theoptimal scaffold composition and architecture to promotethe formation of stable adipose tissue remains unclear. Ourwork focuses on the development of 3D constructsincorporating scaffolds derived from the ECM.
Cell adhesion to the ECM plays a critical role in theregulation of cellular behavior. Cell–matrix interactionsthrough transmembrane receptors can induce signaltransduction pathways that regulate a variety of cellularresponses including survival, proliferation, differentiation,migration, and trafficking [35]. Naturally derived materialsbased on collagen and HA have been widely studied asscaffolds for tissue engineering applications [36]. Collagengels support the attachment and proliferation of manytypes of cells, mediated by cellular integrins that bindcollagen [37]. In contrast, the hydrophilic and polyanionicnature of HA does not facilitate cellular adhesion andprotein adsorption [38]. Fibroblasts seeded in crosslinkedHA–DTPH hydrogels were previously shown to surviveand proliferate, but cellular adhesion and migration wasnot supported [30,39]. Similar to the results with the ASC,the fibroblasts were observed to have a rounded morphol-ogy and tended to form aggregates. A number oftechniques have been developed to improve the celladhesion to the HA–DTPH hydrogels, including theincorporation of crosslinked gelatin [40], fibronectindomains [41], and RGD peptides [42,43]. These strategieswere shown to promote cellular attachment and spreadingin a concentration dependent manner.
The study of the ASC response in collagen-based PDMscaffolds and XLHA scaffolds has yielded interestingresults. Due to the lack of an adhesive matrix, the cellsseeded in the XLHA gels were rounded and tended to formself-aggregates. The live/dead staining results qualitativelyindicated that there was a loss of cellular viability withtime. In assaying the total intracellular protein content tonormalize the GPDH expression levels, there was lessprotein present in the XLHA alone constructs, suggestiveof a reduced cellular population (results not shown). TheASC could be observed to have an extended morphology inthe scaffolds that contained the PDM. In the combined
PDM with XLHA scaffolds, it was possible to alter thecellular organization by modifying the seeding methodol-ogy. The PDM with XLHA Adhesion methodologypromoted cellular adhesion to the PDM. In contrast, whenthe PDM with XLHA Encapsulation methodology wasused, a large percentage of the seeded cells could be foundwithin the XLHA portion of the construct. It is probablethat the ASC were trapped within the XLHA and wereunable to migrate unless they were in close proximity to thePDM at the time of gelation. In all of the constructsincorporating the PDM, the qualitative presence of morecells at the 72-h time point suggests that the PDM scaffoldsmay support the proliferation of the ASC. Similar to nativeadipose tissue, an engineered substitute should incorporatea mixture of mature adipocytes and undifferentiatedprogenitors to facilitate regeneration. The PDM scaffoldmay support the maintenance of this stem cell pool.We hypothesize that cellular adhesion may play an
important role in the differentiation response of the ASC.This theory may be supported by the fact that matureadipocytes maintain their phenotype when cultured insuspension and have been shown to de-differentiate into aprecursor state when cultured under cell-adhesive condi-tions [44]. In addition, a study conducted by Spiegelmanand Ginty showed that adipogenic differentiation in themurine 3T3-F422A cell line was inhibited when the cellsadhered and spread on fibronectin matrices [45]. Theobserved inhibitory effects could be reversed by disruptingthe actin cytoskeleton and culturing the cells in a roundedstate. In another study, McBeath et al. observed that cellshape influenced lineage commitment in human mesench-ymal stem cells (hMSC) [46]. More specifically, adipogenicdifferentiation was observed in unspread hMSC that had arounded phenotype (i.e. low adhesion conditions). Incontrast, osteogenic differentiation was observed in cultureconditions that promoted cellular attachment and spread-ing (i.e. high adhesion conditions).In the current work, the cells in the non-adhesive XLHA
alone constructs had significantly higher GPDH activitylevels, approaching the values observed for mature adiposetissue. The Oil Red O staining results support theaugmentation of differentiation in the XLHA aloneconstructs. Cells containing intracellular lipid accumula-tion could be readily detected in the gels at both 7 days and14 days. The cells in the 14-day samples had a moredifferentiated phenotype with larger intracellular lipiddroplets, approaching the unilocular morphology ofmature adipocytes. This augmentation in the differentia-tion response may involve the interaction of the ASC withthe XLHA through the hyaluronan receptor CD44 [47].However, our experimental findings with the two combinedscaffolds suggest a role for adhesion in the modulation ofthe differentiation response. Higher GPDH activity levels(statistically significant at 7 days) were found when thePDM with XLHA encapsulation methodology was used.As previously discussed, the majority of the cells weresuspended within the XLHA portion of the scaffold with

ARTICLE IN PRESSL. Flynn et al. / Biomaterials 28 (2007) 3834–3842 3841
this seeding strategy. The Oil Red O staining results alsoindicate that the encapsulation methodology promotedintracellular lipid accumulation and a more differentiatedphenotype. These results clearly show that the seedingmethodologies and the scaffold environment influence theASC response. Moreover, promoting ASC adhesion maynot be the best strategy for facilitating adipogenicdifferentiation.
Overall, we believe that the PDM and XLHA scaffoldshold promise as scaffolds for adipose tissue engineering.Differentiating cells could be detected using Oil Red Ostaining in all of the scaffold conditions. In general, thecells seeded in the scaffolds had higher GPDH activitylevels than the non-induced TCPS control samples. Thelarge standard deviations in the activity levels can beattributed to the use of primary cells from a variety ofdonors.
There are a number of advantages to including the PDMscaffold in the construct. The PDM significantly improvesthe surgical handlabililty and mechanical integrity of thescaffold. Additionally, the PDM contains a similar ECMcomposition to adipose tissue, including collagens type I,III, IV, V and VI [48,49]. Hence, the PDM may mimic thenative adipose tissue environment and help to maintain thepool of adipogenic progenitors following implantation.The PDM may also help to promote construct vascular-ization following implantation, which will be critical to thesuccess of the device. The vascular network of the placentais preserved during the decellularization processing, andmay facilitate angiogenesis by providing the appropriate3D cues to stimulate proper cellular organization andinfiltration. Vascularization could also be promoted by HAoligosaccharides produced during the degradation of theXLHA by hyaluronidases [41], and the acceleration ofvascularization is possible through the addition of single ormultiple growth factors within the XLHA materials [50,51].
We are continuing our investigation of the ASC responseto the PDM and XLHA scaffolds in order to develop amore thorough understanding of the influence of thescaffold environment on the cellular behavior. We arecurrently conducting studies to quantify the cellularproliferation and metabolic activity in the differentscaffolds. Our ongoing work also focuses on characterizingthe changes in the ASC gene expression during adipogenicdifferentiation within the scaffolds using reverse transcrip-tase polymerase chain reaction (RT–PCR).
5. Conclusions
Fabrication techniques and culture methods were devel-oped to investigate primary human ASC in severalnaturally-derived scaffolds incorporating decellularizedhuman placenta and XLHA. The results with the combinedPDM with XLHA constructs indicate that cellularorganization and adipogenic differentiation can be alteredthrough variations in the cell seeding methodology. Celladhesion may impact the ASC proliferation and differ-
entiation responses. The PDM scaffolds facilitated cellularattachment and spreading. In contrast, adipogenic differ-entiation was augmented when the cells were encapsulatedin the non-cell-adhesive XLHA gels. The results of ourinvestigation clearly show that the scaffold environmentgreatly influences the organization and differentiation ofthe seeded ASC.
Acknowledgments
The authors would like to acknowledge Mary Boyle andthe delivery suite staff at Women’s College Hospital fortheir assistance with the placental procurement. We thankDr. X.Z. Shu for providing the HA–DTPH, and the UtahCenters of Excellence Program and the NIH (DC 004336)for financial support at Utah. This work was supported byfunding from the Natural Sciences and EngineeringResearch Council (NSERC) of Canada, the CanadianInstitutes of Health Research (CIHR) and the Province ofOntario through the Advanced Regenerative TissueEngineering Centre (ARTEC).
References
[1] Katz AJ, Llull R, Hedrick MH, Futrell JW. Emerging approaches to
the tissue engineering of fat. Clin Plast Surg 1999;26(4):587–603 viii.
[2] Patrick Jr. CW. Tissue engineering strategies for adipose tissue repair.
Anat Rec 2001;263(4):361–6.
[3] The American Society of Plastic Surgeons. /www.plasticsurgery.orgS.[4] Shenaq SM, Yuksel E. New research in breast reconstruction:
adipose tissue engineering. Clin Plast Surg 2002;29(1):111–25 vi.
[5] Patrick Jr CW. Adipose tissue engineering: the future of breast and
soft tissue reconstruction following tumor resection. Semin Surg
Oncol 2000;19(3):302–11.
[6] Patrick CW. Breast tissue engineering. Annu Rev Biomed Eng
2004;6:109–30.
[7] Beahm EK, Walton RL, Patrick Jr CW. Progress in adipose tissue
construct development. Clin Plast Surg 2003;30(4):547–58 viii.
[8] Alhadlaq A, Tang M, Mao JJ. Engineered adipose tissue from human
mesenchymal stem cells maintains predefined shape and dimension:
implications in soft tissue augmentation and reconstruction. Tissue
Eng 2005;11(3-4):556–66.
[9] Fischbach C, Spruss T, Weiser B, Neubauer M, Becker C, Hacker M,
et al. Generation of mature fat pads in vitro and in vivo utilizing 3-D
long-term culture of 3T3-L1 preadipocytes. Exp Cell Res
2004;300(1):54–64.
[10] Kang X, Xie Y, Kniss DA. Adipose tissue model using three-
dimensional cultivation of preadipocytes seeded onto fibrous polymer
scaffolds. Tissue Eng 2005;11(3-4):458–68.
[11] Patrick Jr CW, Zheng B, Johnston C, Reece GP. Long-term
implantation of preadipocyte-seeded PLGA scaffolds. Tissue Eng
2002;8(2):283–93.
[12] Kral JG, Crandall DL. Development of a human adipocyte synthetic
polymer scaffold. Plast Reconstr Surg 1999;104(6):1732–8.
[13] Gentleman E, Nauman EA, Livesay GA, Dee KC. Collagen
composite biomaterials resist contraction while allowing development
of adipocytic soft tissue in vitro. Tissue Eng 2006.
[14] Halbleib M, Skurk T, de Luca C, von Heimburg D, Hauner H. Tissue
engineering of white adipose tissue using hyaluronic acid-based
scaffolds. I: in vitro differentiation of human adipocyte precursor
cells on scaffolds. Biomaterials 2003;24(18):3125–32.
[15] Kawaguchi N, Toriyama K, Nicodemou-Lena E, Inou K, Torii S,
Kitagawa Y. De novo adipogenesis in mice at the site of injection of

ARTICLE IN PRESSL. Flynn et al. / Biomaterials 28 (2007) 3834–38423842
basement membrane and basic fibroblast growth factor. Proc Natl
Acad Sci USA 1998;95(3):1062–6.
[16] Schoeller T, Lille S, Wechselberger G, Otto A, Mowlavi A, Piza-
Katzer H. Histomorphologic and volumetric analysis of implanted
autologous preadipocyte cultures suspended in fibrin glue: a potential
new source for tissue augmentation. Aesthetic Plast Surg 2001;25(1):
57–63.
[17] Kim BS, Baez CE, Atala A. Biomaterials for tissue engineering.
World J Urol 2000;18(1):2–9.
[18] De Ugarte DA, Ashjian PH, Elbarbary A, Hedrick MH. Future of
fat as raw material for tissue regeneration. Ann Plast Surg 2003;
50(2):215–9.
[19] von Heimburg D, Hemmrich K, Zachariah S, Staiger H, Pallua N.
Oxygen consumption in undifferentiated versus differentiated adipo-
genic mesenchymal precursor cells. Respir Physiol Neurobiol 2005;
146(2-3):107–16.
[20] Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, et al.
Multilineage cells from human adipose tissue: implications for cell-
based therapies. Tissue Eng 2001;7(2):211–28.
[21] Cho SW, Kim SS, Rhie JW, Cho HM, Choi CY, Kim BS.
Engineering of volume-stable adipose tissues. Biomaterials 2005;
26(17):3577–85.
[22] Zuk PA, Zhu M, Ashjian P, De Ugarte DA, Huang JI, Mizuno H,
et al. Human adipose tissue is a source of multipotent stem cells. Mol
Biol Cell 2002;13(12):4279–95.
[23] Ryden M, Dicker A, Gotherstrom C, Astrom G, Tammik C, Arner P,
et al. Functional characterization of human mesenchymal stem cell-
derived adipocytes. Biochem Biophys Res Commun 2003;311(2):
391–7.
[24] Hemmrich K, von Heimburg D, Cierpka K, Haydarlioglu S, Pallua
N. Optimization of the differentiation of human preadipocytes in
vitro. Differentiation 2005;73(1):28–35.
[25] Flynn L, Semple JL, Woodhouse KA. Decellularized placental
matrices for adipose tissue engineering. J Biomed Mater Res A
2006;79(2):359–69.
[26] Schmidt CE, Baier JM. Acellular vascular tissues: natural biomater-
ials for tissue repair and tissue engineering. Biomaterials 2000;
21(22):2215–31.
[27] Prestwich GD, Marecak DM, Marecek JF, Vercruysse KP, Ziebell
MR. Controlled chemical modification of hyaluronic acid: synthesis,
applications, and biodegradation of hydrazide derivatives. J Control
Release 1998;53(1–3):93–103.
[28] Shu XZ, Prestwich GD. Therapeutic biomaterials from chemically
modified hyaluronan. In: Garg HG, Hales CA, editors. Chemistry
and biology of hyaluronan. Amsterdam: Elsevier Press; 2004.
p. 475–504.
[29] Hauner H, Skurk T, Wabitsch M. Cultures of human adipose
precursor cells. Methods Mol Biol 2001;155:239–47.
[30] Shu XZ, Liu Y, Luo Y, Roberts MC, Prestwich GD. Disulfide cross-
linked hyaluronan hydrogels. Biomacromolecules 2002;3(6):1304–11.
[31] Shu X, Liu Y, Palumbo FS, Luo Y, Prestwich GD. In situ
crosslinkable hyaluronan hydrogels for tissue engineering. Biomater-
ials 2004;25(7–8):1339–48.
[32] Halvorsen YD, Bond A, Sen A, Franklin DM, Lea-Currie YR,
Sujkowski D, et al. Thiazolidinediones and glucocorticoids synergis-
tically induce differentiation of human adipose tissue stromal cells:
biochemical, cellular, and molecular analysis. Metabolism 2001;
50(4):407–13.
[33] Rodriguez AM, Elabd C, Delteil F, Astier J, Vernochet C, Saint-
Marc P, et al. Adipocyte differentiation of multipotent cells
established from human adipose tissue. Biochem Biophys Res
Commun 2004;315(2):255–63.
[34] Abbott A. Cell culture: biology’s new dimension. Nature 2003;
424(6951):870–2.
[35] Gumbiner BM. Cell adhesion: the molecular basis of tissue
architecture and morphogenesis. Cell 1996;84(3):345–57.
[36] Park SN, Park JC, Kim HO, Song MJ, Suh H. Characterization of
porous collagen/hyaluronic acid scaffold modified by 1-ethyl-3-
(3-dimethylaminopropyl)carbodiimide cross-linking. Biomaterials
2002;23(4):1205–12.
[37] Lee RT, Berditchevski F, Cheng GC, Hemler ME. Integrin-mediated
collagen matrix reorganization by cultured human vascular smooth
muscle cells. Circ Res 1995;76(2):209–14.
[38] Liu Y, Zheng Shu X, Prestwich GD. Biocompatibility and stability of
disulfide-crosslinked hyaluronan films. Biomaterials 2005;26(23):
4737–46.
[39] Liu Y, Shu XZ, Gray SD, Prestwich GD. Disulfide-crosslinked
hyaluronan–gelatin sponge: growth of fibrous tissue in vivo.
J Biomed Mater Res A 2004;68(1):142–9.
[40] Shu XZ, Ahmad S, Liu Y, Prestwich GD. Synthesis and evaluation of
injectable, in situ crosslinkable synthetic extracellular matrices for
tissue engineering. J Biomed Mater Res A 2006;79(4):902–12.
[41] Ghosh K, Ren XD, Shu XZ, Prestwich GD, Clark RA. Fibronectin
functional domains coupled to hyaluronan stimulate adult human
dermal fibroblast responses critical for wound healing. Tissue Eng
2006;12(3):601–13.
[42] Shu XZ, Liu Y, Palumbo F, Prestwich GD. Disulfide-crosslinked
hyaluronan-gelatin hydrogel films: a covalent mimic of the extra-
cellular matrix for in vitro cell growth. Biomaterials 2003;24(21):
3825–34.
[43] Shu XZ, Ghosh K, Liu Y, Palumbo FS, Luo Y, Clark RA, et al.
Attachment and spreading of fibroblasts on an RGD peptide-
modified injectable hyaluronan hydrogel. J Biomed Mater Res A
2004;68(2):365–75.
[44] Tholpady SS, Aojanepong C, Llull R, Jeong JH, Mason AC, Futrell
JW, et al. The cellular plasticity of human adipocytes. Ann Plast Surg
2005;54(6):651–6.
[45] Spiegelman BM, Ginty CA. Fibronectin modulation of cell shape and
lipogenic gene expression in 3T3-adipocytes. Cell 1983;35(3 Part 2):
657–66.
[46] McBeath R, Pirone DM, Nelson CM, Bhadriraju K, Chen CS. Cell
shape, cytoskeletal tension, and RhoA regulate stem cell lineage
commitment. Dev Cell 2004;6(4):483–95.
[47] Gimble J, Guilak F. Adipose-derived adult stem cells: isolation,
characterization, and differentiation potential. Cytotherapy 2003;
5(5):362–9.
[48] Amenta PS, Gay S, Vaheri A, Martinez-Hernandez A. The
extracellular matrix is an integrated unit: ultrastructural localization
of collagen types I, III, IV, V, VI, fibronectin, and laminin in human
term placenta. Coll Relat Res 1986;6(2):125–52.
[49] Nakajima I, Yamaguchi T, Ozutsumi K, Aso H. Adipose tissue
extracellular matrix: newly organized by adipocytes during differ-
entiation. Differentiation 1998;63(4):193–200.
[50] Cai S, Liu Y, Zheng Shu X, Prestwich GD. Injectable glycosami-
noglycan hydrogels for controlled release of human basic fibroblast
growth factor. Biomaterials 2005;26(30):6054–67.
[51] Pike DB, Cai S, Pomraning KR, Firpo MA, Fisher RJ, Shu XZ, et al.
Heparin-regulated release of growth factors in vitro and angiogenic
response in vivo to implanted hyaluronan hydrogels containing
VEGF and bFGF. Biomaterials 2006;27(30):5242–51.
Related Documents











