Adipocyte morphology and leptin signaling in rat offspring from mothers supplemented with flaxseed during lactation Q1 Mariana Sarto Figueiredo Ph.D. a , Magna Cottini da Fonseca Passos Ph.D. a , Isis Hara Trevenzoli Ph.D. b , Aline Andrade Troina Ph.D. a , Aluana Santana Carlos M.S. c , Celly Cristina Alves Nascimento-Saba Ph.D. c , Mabel Carneiro Fraga Ph.D. a, d , Alex Christian Manh ~ aes Ph.D. d , Elaine de Oliveira Ph.D. a , Patr ıcia Cristina Lisboa Ph.D. a , Egberto Gaspar de Moura Ph.D. a, * a Laboratory of Endocrine Physiology, Biology Institute, State University of Rio de Janeiro, Rio de Janeiro, RJ, Brazil Q2 b Laboratory of Molecular Endocrinology, Federal University of Rio de Janeiro, Rio de Janeiro, RJ, Brazil c Laboratory of Morphofunctional Analysis, Biology Institute, State University of Rio de Janeiro, Rio de Janeiro, RJ, Brazil d Laboratory of Neurophysiology, Biology Institute, State University of Rio de Janeiro, Rio de Janeiro, RJ, Brazil article info Article history: Received 15 March 2011 Accepted 4 July 2011 Keywords: Adipose tissue Flaxseed Lactation Leptin signaling and programming abstract Objective: We have recently shown that maternal flaxseed supplementation during lactation induces insulin resistance in adult offspring. Here, we studied the effects of maternal dietary flaxseed during lactation on adipocyte morphology and leptin signaling in the hypothalamic- pituitary-thyroid axis as well as on behavioral traits in the adult progeny. Methods: Lactating rats were fed a control (C) diet or a diet with 25% flaxseed (F). After weaning, pups received a standard diet until postnatal day (PN) 180. Male offspring were killed at PN21 and 180. Data were considered significant at P < 0.05. Results: Weaned F rats presented a lower total and subcutaneous fat mass and higher subcutaneous adipocyte area (þ48%), but at adulthood they presented higher subcutaneous and visceral adipocyte areas (þ40% and 1.9-fold increase, respectively), with no change in body fat mass. At PN21, F pups had hyperleptinemia (þ69%), lower T 3 (33%), higher TSH (2.1-fold increase), higher pituitary leptin receptor (Ob-R, þ11%), signal transducer and activator of transcription 3 (STAT3, þ21%), and phosphorylated-STAT3 (p-STAT3, þ77%) protein content. Adult F offspring only showed lower T 4 (28%) and higher thyroid Ob-R (þ52%) expression. Maternal flaxseed intake during lactation did not result in behavioral changes in the adult offspring. Conclusions: Maternal flaxseed supplementation decreases offspring adiposity and increases pituitary leptin signaling at weaning, but it induces hypertrophic adipocytes and higher thyroid leptin receptor in adulthood. The present data suggest that extensive use of flaxseed during lactation is undesirable. Ó 2011 Elsevier Inc. All rights reserved. Introduction Adverse nutritional or hormonal environments during early life may change some physiological parameters at adulthood. Barker [1] reported low birth weight with diabetes, obesity, and hypertension in adult humans. This association has been named metabolic programming and has been confirmed by several epidemiologic and experimental studies [2,3]. Our group has shown that neonatal imprinting factors during lactation, such as malnutrition [4–6], overnutrition [7], thyroid hormones [8], leptin [9,10], and hypoprolactinemia [11,12], program changes in adiposity, leptin signaling, thyroid function, glucose homeo- stasis, and lipid profile. Leptin is a hormone secreted by white adipose tissue that crosses the blood-brain barrier by a saturable transport system and that affects feeding behavior, thermogenesis, and neuroen- docrine status by acting through the hypothalamic leptin receptor (Ob-R) and the Janus tyrosine kinase 2 (JAK2)/signal transducer and activator of transcription 3 (STAT3) intracellular pathway [13,14]. The long form (Ob-Rb) and the short form of the leptin receptor (Ob-Ra) are the most studied isoforms and Ob-Rb * Corresponding author. Tel.: (þ55.21) 25876434; fax: (þ55.21) 25876129. E-mail address: [email protected] (E. G. de Moura). 0899-9007/$ - see front matter Ó 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.nut.2011.07.002 Contents lists available at ScienceDirect Nutrition journal homepage: www.nutritionjrnl.com Please cite this article in press as: Figueiredo MS, et al., Adipocyte morphology and leptin signaling in rat offspring from mothers sup- plemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002 Nutrition xxx (2011) 1–9 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 NUT8595_proof ■ 23 July 2011 ■ 1/9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Q1
Q2
lable at ScienceDirect
Nutrition xxx (2011) 1–9
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
NUT8595_proof ■ 23 July 2011 ■ 1/9
Contents lists avai
Nutrition
journal homepage: www.nutr i t ionjrnl .com
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
Adipocyte morphology and leptin signaling in rat offspring from motherssupplemented with flaxseed during lactation
Mariana Sarto Figueiredo Ph.D. a, Magna Cottini da Fonseca Passos Ph.D. a, Isis Hara Trevenzoli Ph.D. b,Aline Andrade Troina Ph.D. a, Aluana Santana Carlos M.S. c,Celly Cristina Alves Nascimento-Saba Ph.D. c, Mabel Carneiro Fraga Ph.D. a,d,Alex Christian Manh~aes Ph.D. d, Elaine de Oliveira Ph.D. a, Patr�ıcia Cristina Lisboa Ph.D. a,Egberto Gaspar de Moura Ph.D. a,*a Laboratory of Endocrine Physiology, Biology Institute, State University of Rio de Janeiro, Rio de Janeiro, RJ, Brazilb Laboratory of Molecular Endocrinology, Federal University of Rio de Janeiro, Rio de Janeiro, RJ, Brazilc Laboratory of Morphofunctional Analysis, Biology Institute, State University of Rio de Janeiro, Rio de Janeiro, RJ, Brazild Laboratory of Neurophysiology, Biology Institute, State University of Rio de Janeiro, Rio de Janeiro, RJ, Brazil
71
72
73
74
75
76
77
78
79
80
a r t i c l e i n f o
Article history:Received 15 March 2011Accepted 4 July 2011
Keywords:Adipose tissueFlaxseedLactationLeptin signaling and programming
* Corresponding author. Tel.: (þ55.21) 25876434; fE-mail address: [email protected] (E. G. d
0899-9007/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.nut.2011.07.002
Please cite this article in press as: Figueiredplemented with..., Nutrition (2011), doi:10.1
81
82
83
84
85
86
87
88
89
90
91
a b s t r a c t
Objective: We have recently shown that maternal flaxseed supplementation during lactationinduces insulin resistance in adult offspring. Here, we studied the effects of maternal dietaryflaxseed during lactation on adipocyte morphology and leptin signaling in the hypothalamic-pituitary-thyroid axis as well as on behavioral traits in the adult progeny.Methods: Lactating rats were fed a control (C) diet or a diet with 25% flaxseed (F). After weaning,pups received a standard diet until postnatal day (PN) 180. Male offspring were killed at PN21 and180. Data were considered significant at P < 0.05.Results:Weaned F rats presented a lower total and subcutaneous fat mass and higher subcutaneousadipocyte area (þ48%), but at adulthood they presented higher subcutaneous and visceraladipocyte areas (þ40% and 1.9-fold increase, respectively), with no change in body fat mass. AtPN21, F pups had hyperleptinemia (þ69%), lower T3 (�33%), higher TSH (2.1-fold increase), higherpituitary leptin receptor (Ob-R, þ11%), signal transducer and activator of transcription 3 (STAT3,þ21%), and phosphorylated-STAT3 (p-STAT3, þ77%) protein content. Adult F offspring only showedlower T4 (�28%) and higher thyroid Ob-R (þ52%) expression. Maternal flaxseed intake duringlactation did not result in behavioral changes in the adult offspring.Conclusions: Maternal flaxseed supplementation decreases offspring adiposity and increasespituitary leptin signaling at weaning, but it induces hypertrophic adipocytes and higher thyroidleptin receptor in adulthood. The present data suggest that extensive use of flaxseed duringlactation is undesirable.
� 2011 Elsevier Inc. All rights reserved.
92
93
94
95
96
97
98
99
100
101
102
Introduction
Adverse nutritional or hormonal environments during earlylife may change some physiological parameters at adulthood.Barker [1] reported low birth weight with diabetes, obesity, andhypertension in adult humans. This association has been namedmetabolic programming and has been confirmed by severalepidemiologic and experimental studies [2,3]. Our group hasshown that neonatal imprinting factors during lactation, such as
ax: (þ55.21) 25876129.e Moura).
ll rights reserved.
o MS, et al., Adipocyte mo016/j.nut.2011.07.002
103
104
105
malnutrition [4–6], overnutrition [7], thyroid hormones [8],leptin [9,10], and hypoprolactinemia [11,12], program changes inadiposity, leptin signaling, thyroid function, glucose homeo-stasis, and lipid profile.
Leptin is a hormone secreted by white adipose tissue thatcrosses the blood-brain barrier by a saturable transport systemand that affects feeding behavior, thermogenesis, and neuroen-docrine status by acting through the hypothalamic leptinreceptor (Ob-R) and the Janus tyrosine kinase 2 (JAK2)/signaltransducer and activator of transcription 3 (STAT3) intracellularpathway [13,14]. The long form (Ob-Rb) and the short form of theleptin receptor (Ob-Ra) are the most studied isoforms and Ob-Rb
rphology and leptin signaling in rat offspring from mothers sup-
Q3
4
Table 1Composition of 100 g of diet used in the experimental during lactation phase
Ingredients Control Flaxseed
% kJ % kJ
Casein* 22.70 284.51 16.00 200.31Flaxseedy d d 25.00 513.38Corn starchz 50.30 701.45 44.00 617.10Sucrosex 10.00 167.36 10.00 167.36Mineral Mix AIN93G* 3.50 12.89 3.50 12.89Vitamin Mix AIN93G* 1.00 16.40 1.00 16.40Soybean oilk 7.00 263.59 d d
Cellulose (Fiber){ 5.00 d d d
Choline Bitartrate* 0.25 d 0.25 d
L-Cystine* 0.30 5.02 0.30 5.00Tert-Butylhydroquinone# 0.0014 d 0.0014 d
Macronutrient composition (100g/diet)Protein 17.40 17.30Carbohydrate 52.10 54.30Fat 7.60 8.90
Total energy (kJ/ 100 g) 1451.20 1532.44
* M Cassab Comercio & Industria LTDA (S~ao Paulo, SP, Brasil).y Arma Zen Produtos Naturais LTDA (Rio de Janeiro, RJ, Brasil).z Maisena, Unilever Best Foods Brasil LTDA (Mogi Guacu, SP, Brasil).x Uni~ao (Rio de Janeiro, RJ, Brasil).{ Microcel, Blanver LTDA (Cotia, SP, Brasil).k Liza Cargil Agricultura LTDA (Mairinque, SP, Brasil).# Vogler Ingredients (Eastman, USA).
M. S. Figueiredo et al. / Nutrition xxx (2011) 1–92
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
NUT8595_proof ■ 23 July 2011 ■ 2/9
is fully capable of activating intracellular signaling [15]. Leptinbinding to Ob-Rb initiates tyrosine phosphorylation by JAK2.Phosphorylated Ob-Rb recruits STAT3, which is activatedthrough phosphorylation by JAK2 [16]. Activated STAT3 dimer-izes and translocates to the nucleus to stimulate gene tran-scription [17]. The JAK2–STAT3 pathway stimulates thetranscription of the suppressor of cytokine signaling 3 (SOCS3),which is a negative regulator of leptin signaling [16,18].
Flaxseed (Linum usitatissimum L.) has been a focus of interestin the field of functional food because of its potential healthbenefits, such as the improvement of lipid profile, glycemia, andcardiovascular function [19–23]. Flaxseed is rich in protein, die-tary fat, and fiber fat; approximately 37% of its mass consists ofoil, of which 50% is a-linolenic acid, which is an (n-3) fatty acid[18:3(n-3), alpha-linolenic acid] [24]. Flaxseed is one of therichest sources of the plant lignan secoisolariciresinol digluco-side, which is present in a much higher proportion in the seedthan in other tissues and is responsible for the observed phy-toestrogenic, anticarcinogenic, and antiatherogenic effects[25–27].
Although flaxseed has several potential advantages whenconsumed throughout life, it may cause adverse effects when it isconsumed in critical periods of life, such as gestation and lactation[28,29]. We have shown that maternal flaxseed in the diet duringlactation in rats is associated with higher total and visceral fatmass, serum triglycerides, and cholesterol in adult femaleoffspring [30], whereas, in male offspring, there is no change inbody adiposity, but hypoadiponectinemia and hyperinsulinemiado occur, suggesting insulin resistance [31]. These effects seem toresult from changes observed in the milk composition, such ashigher leptin and 17b-estradiol contents [30].
Several studies have investigated the associations betweenleptin and thyroid hormones. Leptin seems to act on the thyroidsystem mainly by stimulating TRH secretion [32], but it also actsat the pituitary level [33,34] and thyroid level [35–37], and itinfluences thyroid hormone metabolism [38,39]. In rodents,Ob-R has also been identified in the thyroid and pituitary glands[5,35,40,41].
Both TRH and leptin have antidepressant effects [42], and oneclinical study has demonstrated the beneficial effects of flaxseedoil in the diet on bipolar disorder in children and adolescents[43]. Because flaxseed may influence adipogenesis and leptinproduction, and because leptin has profound effects on thehypothalamic-pituitary-thyroid (HPT) axis and antidepressanteffects [42], our aim was to evaluate the short- and long-termconsequences of maternal flaxseed in the diet during lactationon adipose tissue morphology, leptin signaling in the HPT, andbehavioral disturbances in rats.
Materials and methods
Animal care and diets
The use of the animals according to our experimental design was approvedby the Animal Care and Use Committee of the Biology Institute of the StateUniversity of Rio de Janeiro (protocol 230/2008), which based its analysis on theprinciples adopted and promulgated by the Brazilian law that concerns therearing and use of animals in teaching and research activities in Brazil [44].Three-month-old Wistar rats were maintained in a room with controlledtemperature (25 � 1�C) and dark–light cycle (lights on from 7:00 a.m. to 7:00p.m.). Virgin rats (200–220 g) were mated, and each female was placed in anindividual cage with free access to water and food until the birth of the offspring.
At birth, 16 lactating rats were randomly assigned to each of the followinggroups: control group (C, n¼ 8), with free access to a diet containing 20% protein,60.2% carbohydrate, and 19.8% lipid composing the energy value, with 5% ofcellulose fiber; and flaxseed group (F, n ¼ 8), with free access to a diet containing18.9% protein (13.9% casein and 5% flaxseed), 59.3% carbohydrate, and 21.8% lipid
Please cite this article in press as: Figueiredo MS, et al., Adipocyte moplemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002
exclusively from flaxseed composing the energy value, with 5% of fiber exclu-sively from flaxseed. Birth was defined as day 0 of lactation and weaning was atday 21. The lactating rats started to receive the experimental diet (control orflaxseed) on day 0 of lactation until day 21 of lactation. At birth, all of the litterswere randomly adjusted to six male pups for each mother to maximize lactationperformance [45]. After weaning, all pups received a standard laboratory diet(Nuvilab; Nuvital PR, Brazil Q) containing 22% protein and 66% carbohydrate until180 d of age.
Two pups of eachmother (16 pups) were chosen and killed at 21 d of age, andanother two pups from each mother (16 pups) were killed at 180 d of age witha lethal dose of pentobarbital (0.06g/kg/b.w.) for collection of blood by cardiacpuncture and collection of tissues.
The diets offered to lactating rats (Table 1) were prepared according to theAmerican Institute of Nutrition (AIN 93G) recommendation for rodent diets [46].The flaxseed diet contained 8.9% versus 7.6% lipid and 54.3% versus 52.1%carbohydrate in comparisonwith the control diet, but these differences were notsignificant. The flaxseed was ground in a blender before being added to the diet.The flaxseed diet contained 25% flaxseed, which completely provided the rec-ommended oil and fiber content [46]. The ingredients of the experimental dietswere carefully weighed and homogenized with boiling water to gelatinize thestarch in an industrial food mixer (Hobart). The resulting dough was formed intopellets and dried in a Fabbe-Primar ventilated oven at 60�C for 24 h, labeled, andstored in a refrigerator until use.
The body mass and food intake of the offspring were monitored every 4 dfrom weaning until they were 180 d old.
Behavioral studies
From PN150 to PN170, 40 C and 40 F adult offspring were submitted tothe behavioral tests. The following tests were used: 1) elevated plus-maze(EPM), 2) hole board arena (HB), and 3) radial arm water maze (RAWM).Full descriptions of the testing equipment and protocols can be found else-where [47,48]. All testing sessions were performed between 1:00 and 6:00p.m. in a sound-attenuated room. Behavioral analyses were performed usingvideo images of the tests.
Anxietylike behavior was assessed at PN150 in the EPM. Animals wereallowed 10 min to explore the maze. The number of entries and total timespent in the open and closed arms were recorded. The number of open-armentries (Entries OA) and the time spent in the open arms (Time OA) wereused as anxiety indices. Novelty-seeking behavior was assessed at PN157 in theHB. Animals were given 5 min to explore the arena. The number of nose pokes(head dips) was used as a measure of exploratory activity. Memory andlearning were assessed at PN165 in the RAWM. Animals were tested for fiveconsecutive days, four trials per day. Animals were given 2 min per trial toexplore the maze and find the hidden platform that was positioned at the endof one of the arms. The latency to find the hidden platform was used to assessmemory/learning performance.
rphology and leptin signaling in rat offspring from mothers sup-

M. S. Figueiredo et al. / Nutrition xxx (2011) 1–9 3
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
NUT8595_proof ■ 23 July 2011 ■ 3/9
Total, visceral, and subcutaneous fat masses
The total and visceral fat masses were determined at 21 and 180 d by carcassanalysis [6,9]. The carcasses of the C and F groups were weighed, autoclaved for1 h, and homogenized in distilled water (1:1). Samples of the homogenate werestored at 4�C for analysis. Three grams of homogenate was used to determine thefat mass content gravimetrically [49]. Samples were hydrolyzed in a shakingwater bath at 70�C for 2 h with 30% KOH and ethanol. Total fatty acids andnon-esterified cholesterol were removed using three successive washings withpetroleum ether. After drying overnight in a vacuum, the tubes were weighed,and the results are expressed as g fat/100 g carcass. The visceral (mesenteric,epididymal, and retroperitoneal) fat mass was excised and weighed to evaluatecentral adiposity [50]. The subcutaneous fat mass was estimated from thetotal fat mass minus the visceral fat mass, and the results are expressed aspercentages (%).
310
311
312
313
314
315
316
317
318
319
320
321
322
Adipose tissue morphology
Visceral and subcutaneous (inguinal) adipose tissue samples of the C and Fgroups at 21 and 180 d were fixed in buffered formaldehyde for 72 h, dehydrated,cleared, and then paraffin-embedded so that the plane of each section corre-spondedwith the that of thewider surface. Five-micrometer-thick sections at thesame level were obtained and stained with hematoxylin-eosin to assessmorphology. Tissue sections were observed with an Olympus BX40 lightmicroscope using a �10 objective, and digital images were captured with anOptronics CCD video camera system. The calibrationwas checked regularly usinga slide micrometer. At least 100 adipocyte profiles per animal were randomlyselected and captured. The area, perimeter, and number of adipocytes (cells/100mm2) were determined using the ImageJ 1.34s program (Wayne Rasband,National Institutes of Health, Bethesda, MD, USA) [51,52].
323
324
325
326
327
328
329
330
331
332
333
334
335
336
Serum hormone concentrations
Offspring blood samples were obtained at 21 and 180 d and centrifuged(1000 � g, 4�C, 20 min) to obtain the serum, which was individually kept at�20�C until assaying. All measurements were performed in one assay.
Leptin levels were measured using a murine leptin–specific radioimmuno-assay (RIA) kit (Linco Research, St. Charles, MO, USA), which measures both ratand mouse leptin with an assay sensitivity of 0.5 ng/mL and an intraassay coef-ficient of variation of 2.4%. Total serum T3 (TT3) and free T4 (FT4) levels weremeasured by RIA using commercial kits (Coat-A-Coat; DPC, Los Angeles, CA, USA).The coefficient of intraassay variation for the TT3 was 7.5% and that for FT4 was3.9%. Serum TSH was measured by specific RIA using a kit for rat TSH supplied bythe NIDDK (Bethesda, MD, USA). Data are expressed in terms of the referencepreparation provided (RP-3). The coefficient of intraassay variationwas 2.3%, andthe assay sensitivity was 0.18 ng/mL.
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
Western blot analysis
The amounts of leptin signaling pathway proteins (Ob-R, JAK2, STAT3,pSTAT3) in the hypothalamus, pituitary, and thyroid were evaluated by Westernblot as described before [5,41]. Briefly, to obtain cell extracts, tissues werehomogenized in ice-cold lysis buffer (50 mM HEPES, 1 mM MgCl2, 10 mM EDTA,Triton X-100 1%, pH 6.4) containing the following protease inhibitors: 10 mg/mLaprotinin, 10 mg/mL leupeptin, 2 mg/mL pepstatin, and 1 mM PMSF (Sigma-Aldrich,St. Louis, MO, USA). Actin was used as an internal control.
The total protein content in the homogenates was determined by the BCAprotein kit assay (Rockford, IL, USA), and cell lysates were denatured in samplebuffer (50 mM Tris-HCl, pH 6.8, 1% SDS, 5% 2-mercaptoethanol, 10% glycerol,0.001% bromophenol blue) and heated at 95�C for 5 min. Samples (30 mg totalprotein) were separated by 10% SDS-PAGE and transferred to polyvinylidenemembranes (PVDF, Hybond-P; Amersham Pharmacia Biotech, Piscataway, NJ,USA). Molecular weight markers (Amersham Biosciences, Uppsala, Sweden)wererun in parallel. Membranes were blocked with 5% nonfat milk in Tween-TBS (20mM Tris-HCl, pH 7.5, 500 mM NaCl, 0.1% Tween-20). The primary antibodies(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) used were anti-OB-R(1:500), anti-JAK2 (1:500), anti-STAT3 (1:500), anti-p-STAT3 (1:500), and anti-actin (1:500). The membranes were then washed three times with Tween-TBS(0.1%), followed by incubation for 1 h with the appropriate secondary antibodyconjugated to biotin (Santa Cruz Biotechnology). Then, the membranes wereincubated with streptavidin-conjugated horseradish peroxidase (Caltag Labora-tories, Burlingame, CA, USA). All Western blots were allowed to react withhorseradish peroxidase substrate (ECL-plus; Amersham Pharmacia Biotech) andthen were exposed to x-ray film for 10 s to 1 min. Images were obtained, and thebands were quantified by densitometry using ImageJ and normalized against thebands obtained for actin.
Please cite this article in press as: Figueiredo MS, et al., Adipocyte moplemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002
Statistical analysisdbiochemistry
Data are reported as the means � SEM. The statistical significance wasdetermined by two-way ANOVA to analyze body weight and food intake evolu-tion. The other experimental data were analyzed by Student’s unpaired t test.Differences were considered significant at P < 0.05.
Statistical analysisdbehavior
Data are reported as themeans� SEM. The Kolmogorov–Smirnovone-sampletest (K–S) was used to assess the normality of the distribution of each variable.Significance was assumed for P< 0.05. Differences between the C and F groups inthe Entries OA and Time OA measures were analyzed by means of multivariateanalyses of variance (mANOVA). TREATMENT (C or F) was used as the between-subject factor. Differences between the C and F groups in the number of nosepokes were analyzed by means of the repeated-measures ANOVA (rANOVA).Differences between the C and F groups in the latency to find the hidden platformin the RAWMwere analyzed by rANOVA. DAYand TRIAL were the within-subjectfactors. TREATMENT (C or F) was used as the between-subject factor.
Results
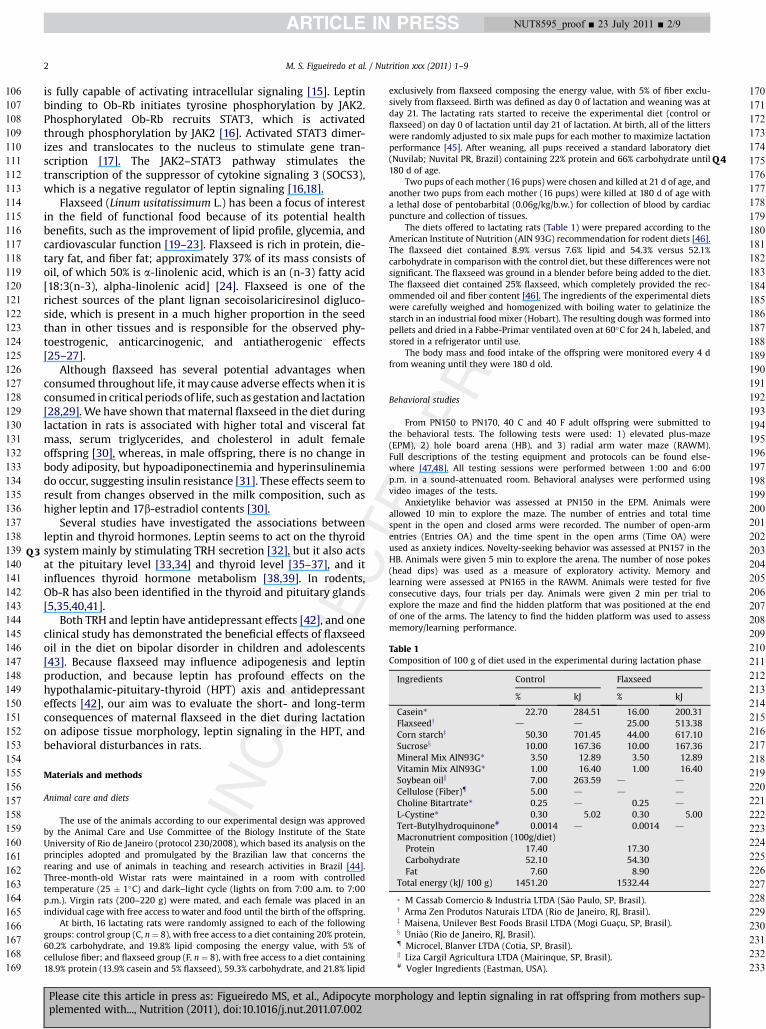
After weaning, pups from the F mothers had higher bodyweights at some points throughout their lives, with increases ofapproximately 10 to 15% (P < 0.05), and their food intake washigher (approximately þ10%, P < 0.05), which confirms previousfindings [31]. F pups presented a lower total fat mass at 21 d ofage (�17.8%, P < 0.05), but at 180 d of age, no changes wereobserved (Fig. 1A). No changes were observed in the visceral fatmass at 21 d (F ¼ 1.06 � 0.11 versus C ¼ 1.25 � 0.05 g) or 180d (F¼ 12.20� 1.03 versus C¼ 11.61�0.72 g). At 21 d, the F grouppresented a lower subcutaneous fat mass (approximately �23%,P < 0.05), and no changes were observed at 180 d old (Fig. 1B).
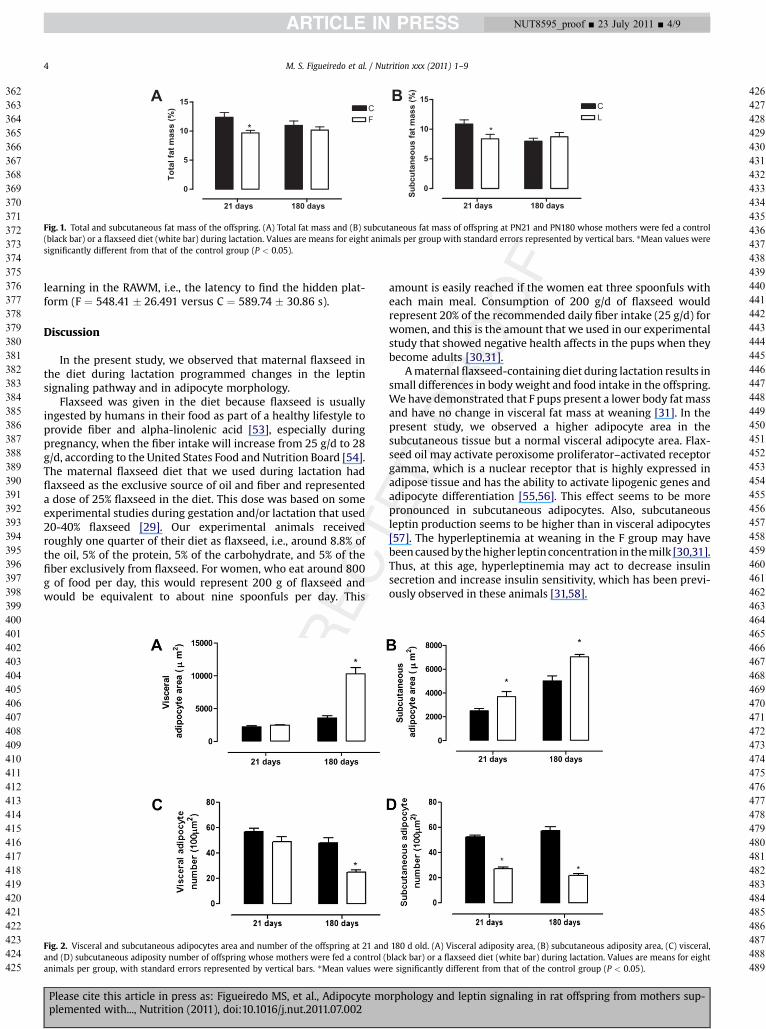
TheFgrouppresentedahigher subcutaneous adipocyte areaat21 (þ48%, P< 0.05) and 180 d of age (þ40%, P< 0.05), as shown inFigure 2A. No changeswere observed in visceral adipocyte area inweaned F pups, but at adulthood, this group showed a highervisceral adipocyte area (1.9-fold increase, P < 0.05; Fig. 2B). Asexpected, the number of adipocytes per areawas lower when thearea of the adipocyteswas higher (Fig. 2C and 2D). Figure 3 showsrepresentative micrographs of the adipocytes.
Leptinemia was higher in the 21-d-old F pups (F ¼ 1.83 � 0.9versus C¼ 1.09� 0.16 ng/mL;þ69%, P< 0.05), but no changewasobserved in adult F rats (F ¼ 1.84 � 0.24 versus C ¼ 1.60 � 0.16ng/mL). At 21 d, F offspring showed lower serum TT3 (F ¼ 75.1 �5.2 versus C ¼ 113.1 � 5.2 ng/dL; P < 0.05), no change in serumFT4, and higher serum TSH (F ¼ 0.72 � 0.03 versus C ¼ 0.35 �0.07 ng/mL; P < 0.05). At 180 d, the F group presented normallevels of TT3 and TSH and lower FT4 (F ¼ 0.66 � 0.05 versus C ¼0.92 � 0.01 ng/dL; P < 0.05).
The levels of leptin signaling pathway proteins (Ob-R, JAK2,STAT3, and p-STAT3) in the hypothalamus of C and F groups weresimilar at weaning and adulthood. In the pituitary gland at 21 d,weaned F pups had a higher Ob-R content (þ11%, P < 0.05;Fig. 4B) and consequently higher STAT3 and pSTAT3 (þ21% andþ77%, respectively, P < 0.05; Fig. 4D and 4E). In contrast, nochanges were detected at adulthood in the pituitary gland.Concerning the leptin pathway in the thyroid gland, no changewas observed at 21 d, but Ob-R expression was higher in Foffspring than in C offspring (þ52%, P < 0.05; Fig. 5A) at 180 d.
The maternal flaxseed-containing diet during lactation hadno effect on behavioral traits in adult progeny. No difference wasobserved between groups in 1) anxiety-like behavior in the EPM,i.e., Entries OA (F¼ 2.03� 0.389 versus C¼ 2.44� 0.416) or TimeOA (F¼ 15794.55� 3673.41 versus C¼ 23508.44 � 5318.29 ms);2) novelty-seeking behavior in the HB, i.e., the number of nosepokes (F ¼ 5.56 � 0.47 versus C ¼ 6.3 � 0.466); or 3) memory/
rphology and leptin signaling in rat offspring from mothers sup-
A B
21 days 180 days
0
5
10
15
CL
*
Su
bc
uta
ne
ou
s fa
t m
as
s (%
)
21 days 180 days
0
5
10
15
CF
*
To
tal fat m
ass (%
)
Fig. 1. Total and subcutaneous fat mass of the offspring. (A) Total fat mass and (B) subcutaneous fat mass of offspring at PN21 and PN180 whose mothers were fed a control(black bar) or a flaxseed diet (white bar) during lactation. Values are means for eight animals per group with standard errors represented by vertical bars. *Mean values weresignificantly different from that of the control group (P < 0.05).
M. S. Figueiredo et al. / Nutrition xxx (2011) 1–94
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
NUT8595_proof ■ 23 July 2011 ■ 4/9
learning in the RAWM, i.e., the latency to find the hidden plat-form (F ¼ 548.41 � 26.491 versus C ¼ 589.74 � 30.86 s).
Discussion
In the present study, we observed that maternal flaxseed inthe diet during lactation programmed changes in the leptinsignaling pathway and in adipocyte morphology.
Flaxseed was given in the diet because flaxseed is usuallyingested by humans in their food as part of a healthy lifestyle toprovide fiber and alpha-linolenic acid [53], especially duringpregnancy, when the fiber intake will increase from 25 g/d to 28g/d, according to the United States Food and Nutrition Board [54].The maternal flaxseed diet that we used during lactation hadflaxseed as the exclusive source of oil and fiber and representeda dose of 25% flaxseed in the diet. This dose was based on someexperimental studies during gestation and/or lactation that used20-40% flaxseed [29]. Our experimental animals receivedroughly one quarter of their diet as flaxseed, i.e., around 8.8% ofthe oil, 5% of the protein, 5% of the carbohydrate, and 5% of thefiber exclusively from flaxseed. For women, who eat around 800g of food per day, this would represent 200 g of flaxseed andwould be equivalent to about nine spoonfuls per day. This
Fig. 2. Visceral and subcutaneous adipocytes area and number of the offspring at 21 andand (D) subcutaneous adiposity number of offspring whose mothers were fed a control (animals per group, with standard errors represented by vertical bars. *Mean values wer
Please cite this article in press as: Figueiredo MS, et al., Adipocyte moplemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002
amount is easily reached if the women eat three spoonfuls witheach main meal. Consumption of 200 g/d of flaxseed wouldrepresent 20% of the recommended daily fiber intake (25 g/d) forwomen, and this is the amount that we used in our experimentalstudy that showed negative health affects in the pups when theybecome adults [30,31].
Amaternal flaxseed-containing diet during lactation results insmall differences in body weight and food intake in the offspring.We have demonstrated that F pups present a lower body fat massand have no change in visceral fat mass at weaning [31]. In thepresent study, we observed a higher adipocyte area in thesubcutaneous tissue but a normal visceral adipocyte area. Flax-seed oil may activate peroxisome proliferator–activated receptorgamma, which is a nuclear receptor that is highly expressed inadipose tissue and has the ability to activate lipogenic genes andadipocyte differentiation [55,56]. This effect seems to be morepronounced in subcutaneous adipocytes. Also, subcutaneousleptin production seems to be higher than in visceral adipocytes[57]. The hyperleptinemia at weaning in the F group may havebeencausedby thehigher leptinconcentration in themilk [30,31].Thus, at this age, hyperleptinemia may act to decrease insulinsecretion and increase insulin sensitivity, which has been previ-ously observed in these animals [31,58].
180 d old. (A) Visceral adiposity area, (B) subcutaneous adiposity area, (C) visceral,black bar) or a flaxseed diet (white bar) during lactation. Values are means for eighte significantly different from that of the control group (P < 0.05).
rphology and leptin signaling in rat offspring from mothers sup-
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
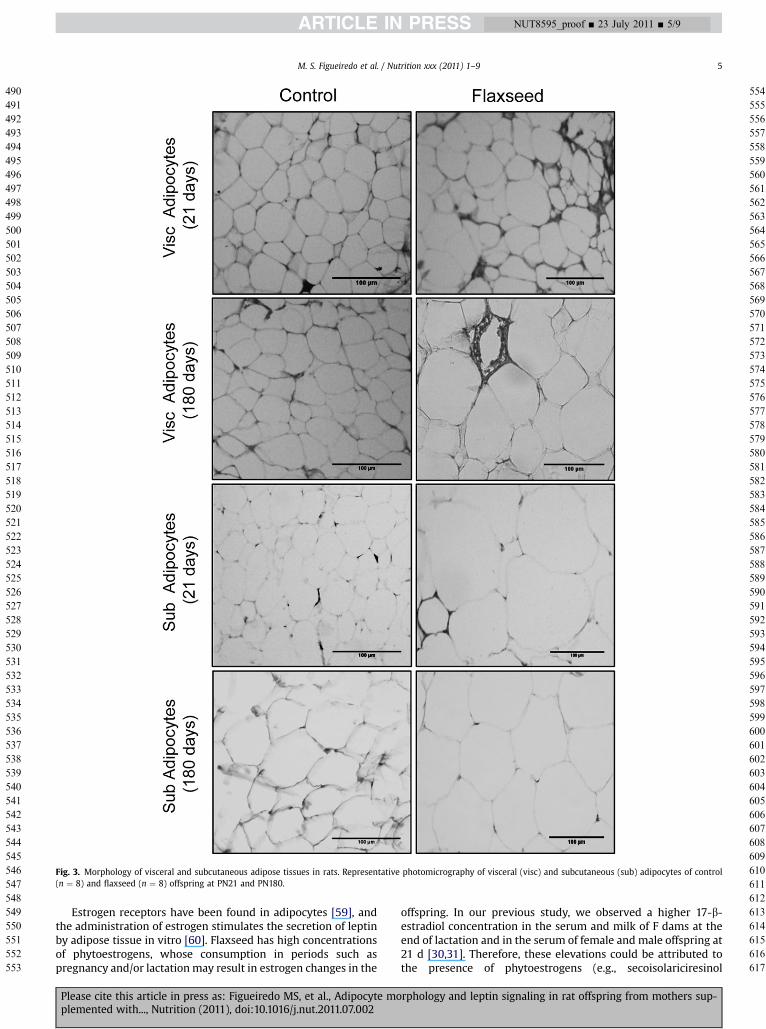
Fig. 3. Morphology of visceral and subcutaneous adipose tissues in rats. Representative photomicrography of visceral (visc) and subcutaneous (sub) adipocytes of control(n ¼ 8) and flaxseed (n ¼ 8) offspring at PN21 and PN180.
M. S. Figueiredo et al. / Nutrition xxx (2011) 1–9 5
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543
544
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
560
561
562
563
564
565
566
567
568
569
570
571
572
573
574
575
576
577
578
579
580
581
582
583
584
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
616
617
NUT8595_proof ■ 23 July 2011 ■ 5/9
Estrogen receptors have been found in adipocytes [59], andthe administration of estrogen stimulates the secretion of leptinby adipose tissue in vitro [60]. Flaxseed has high concentrationsof phytoestrogens, whose consumption in periods such aspregnancy and/or lactationmay result in estrogen changes in the
Please cite this article in press as: Figueiredo MS, et al., Adipocyte moplemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002
offspring. In our previous study, we observed a higher 17-b-estradiol concentration in the serum and milk of F dams at theend of lactation and in the serum of female and male offspring at21 d [30,31]. Therefore, these elevations could be attributed tothe presence of phytoestrogens (e.g., secoisolariciresinol
rphology and leptin signaling in rat offspring from mothers sup-
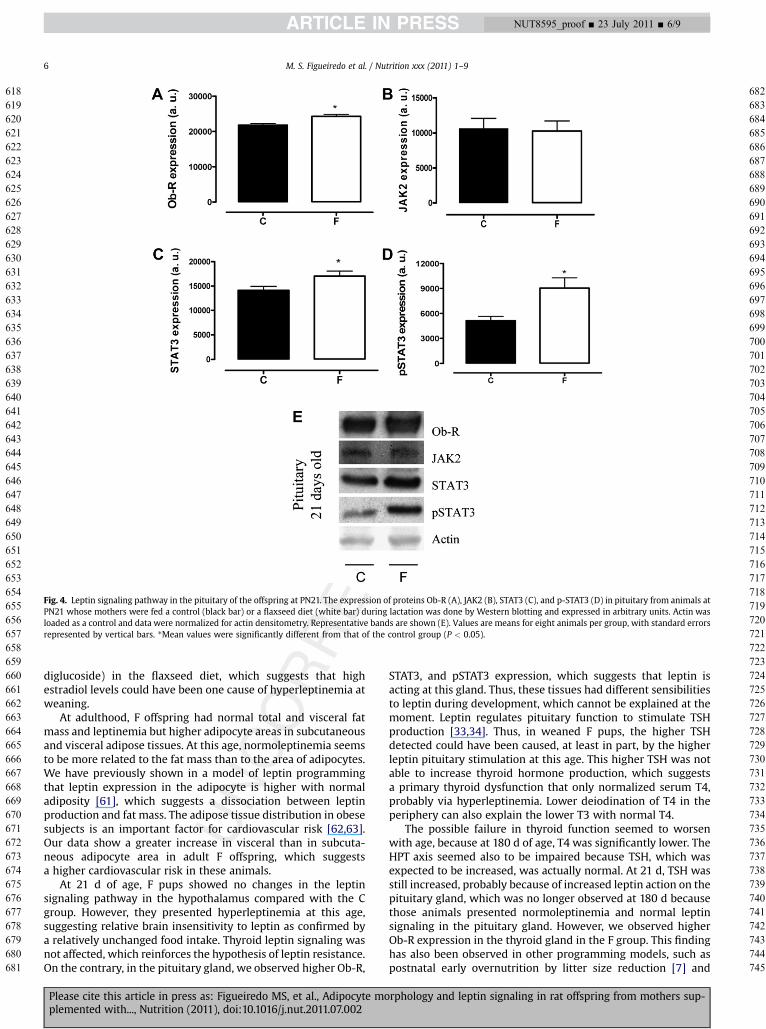
Fig. 4. Leptin signaling pathway in the pituitary of the offspring at PN21. The expression of proteins Ob-R (A), JAK2 (B), STAT3 (C), and p-STAT3 (D) in pituitary from animals atPN21 whose mothers were fed a control (black bar) or a flaxseed diet (white bar) during lactation was done by Western blotting and expressed in arbitrary units. Actin wasloaded as a control and data were normalized for actin densitometry. Representative bands are shown (E). Values are means for eight animals per group, with standard errorsrepresented by vertical bars. *Mean values were significantly different from that of the control group (P < 0.05).
M. S. Figueiredo et al. / Nutrition xxx (2011) 1–96
618
619
620
621
622
623
624
625
626
627
628
629
630
631
632
633
634
635
636
637
638
639
640
641
642
643
644
645
646
647
648
649
650
651
652
653
654
655
656
657
658
659
660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
678
679
680
681
682
683
684
685
686
687
688
689
690
691
692
693
694
695
696
697
698
699
700
701
702
703
704
705
706
707
708
709
710
711
712
713
714
715
716
717
718
719
720
721
722
723
724
725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
NUT8595_proof ■ 23 July 2011 ■ 6/9
diglucoside) in the flaxseed diet, which suggests that highestradiol levels could have been one cause of hyperleptinemia atweaning.
At adulthood, F offspring had normal total and visceral fatmass and leptinemia but higher adipocyte areas in subcutaneousand visceral adipose tissues. At this age, normoleptinemia seemsto be more related to the fat mass than to the area of adipocytes.We have previously shown in a model of leptin programmingthat leptin expression in the adipocyte is higher with normaladiposity [61], which suggests a dissociation between leptinproduction and fat mass. The adipose tissue distribution in obesesubjects is an important factor for cardiovascular risk [62,63].Our data show a greater increase in visceral than in subcuta-neous adipocyte area in adult F offspring, which suggestsa higher cardiovascular risk in these animals.
At 21 d of age, F pups showed no changes in the leptinsignaling pathway in the hypothalamus compared with the Cgroup. However, they presented hyperleptinemia at this age,suggesting relative brain insensitivity to leptin as confirmed bya relatively unchanged food intake. Thyroid leptin signaling wasnot affected, which reinforces the hypothesis of leptin resistance.On the contrary, in the pituitary gland, we observed higher Ob-R,
Please cite this article in press as: Figueiredo MS, et al., Adipocyte moplemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002
STAT3, and pSTAT3 expression, which suggests that leptin isacting at this gland. Thus, these tissues had different sensibilitiesto leptin during development, which cannot be explained at themoment. Leptin regulates pituitary function to stimulate TSHproduction [33,34]. Thus, in weaned F pups, the higher TSHdetected could have been caused, at least in part, by the higherleptin pituitary stimulation at this age. This higher TSH was notable to increase thyroid hormone production, which suggestsa primary thyroid dysfunction that only normalized serum T4,probably via hyperleptinemia. Lower deiodination of T4 in theperiphery can also explain the lower T3 with normal T4.
The possible failure in thyroid function seemed to worsenwith age, because at 180 d of age, T4 was significantly lower. TheHPT axis seemed also to be impaired because TSH, which wasexpected to be increased, was actually normal. At 21 d, TSH wasstill increased, probably because of increased leptin action on thepituitary gland, which was no longer observed at 180 d becausethose animals presented normoleptinemia and normal leptinsignaling in the pituitary gland. However, we observed higherOb-R expression in the thyroid gland in the F group. This findinghas also been observed in other programming models, such aspostnatal early overnutrition by litter size reduction [7] and
rphology and leptin signaling in rat offspring from mothers sup-
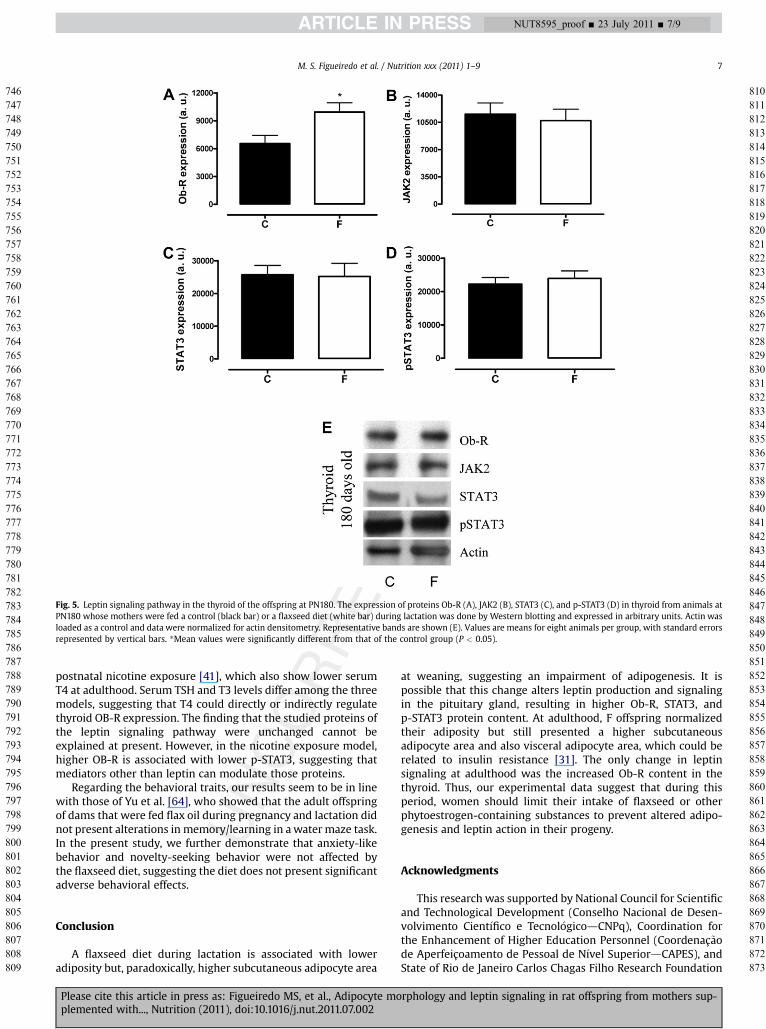
Fig. 5. Leptin signaling pathway in the thyroid of the offspring at PN180. The expression of proteins Ob-R (A), JAK2 (B), STAT3 (C), and p-STAT3 (D) in thyroid from animals atPN180 whose mothers were fed a control (black bar) or a flaxseed diet (white bar) during lactation was done by Western blotting and expressed in arbitrary units. Actin wasloaded as a control and data were normalized for actin densitometry. Representative bands are shown (E). Values are means for eight animals per group, with standard errorsrepresented by vertical bars. *Mean values were significantly different from that of the control group (P < 0.05).
M. S. Figueiredo et al. / Nutrition xxx (2011) 1–9 7
746
747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
763
764
765
766
767
768
769
770
771
772
773
774
775
776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
793
794
795
796
797
798
799
800
801
802
803
804
805
806
807
808
809
810
811
812
813
814
815
816
817
818
819
820
821
822
823
824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
865
866
867
NUT8595_proof ■ 23 July 2011 ■ 7/9
postnatal nicotine exposure [41], which also show lower serumT4 at adulthood. Serum TSH and T3 levels differ among the threemodels, suggesting that T4 could directly or indirectly regulatethyroid OB-R expression. The finding that the studied proteins ofthe leptin signaling pathway were unchanged cannot beexplained at present. However, in the nicotine exposure model,higher OB-R is associated with lower p-STAT3, suggesting thatmediators other than leptin can modulate those proteins.
Regarding the behavioral traits, our results seem to be in linewith those of Yu et al. [64], who showed that the adult offspringof dams that were fed flax oil during pregnancy and lactation didnot present alterations in memory/learning in awater maze task.In the present study, we further demonstrate that anxiety-likebehavior and novelty-seeking behavior were not affected bythe flaxseed diet, suggesting the diet does not present significantadverse behavioral effects.
868
869
870
871
872
873
Conclusion
A flaxseed diet during lactation is associated with loweradiposity but, paradoxically, higher subcutaneous adipocyte area
Please cite this article in press as: Figueiredo MS, et al., Adipocyte moplemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002
at weaning, suggesting an impairment of adipogenesis. It ispossible that this change alters leptin production and signalingin the pituitary gland, resulting in higher Ob-R, STAT3, andp-STAT3 protein content. At adulthood, F offspring normalizedtheir adiposity but still presented a higher subcutaneousadipocyte area and also visceral adipocyte area, which could berelated to insulin resistance [31]. The only change in leptinsignaling at adulthood was the increased Ob-R content in thethyroid. Thus, our experimental data suggest that during thisperiod, women should limit their intake of flaxseed or otherphytoestrogen-containing substances to prevent altered adipo-genesis and leptin action in their progeny.
Acknowledgments
This research was supported by National Council for Scientificand Technological Development (Conselho Nacional de Desen-volvimento Cient�ıfico e Tecnol�ogicodCNPq), Coordination forthe Enhancement of Higher Education Personnel (Coordenac~aode Aperfeicoamento de Pessoal de N�ıvel SuperiordCAPES), andState of Rio de Janeiro Carlos Chagas Filho Research Foundation
rphology and leptin signaling in rat offspring from mothers sup-

M. S. Figueiredo et al. / Nutrition xxx (2011) 1–98
874
875
876
877
878
879
880
881
882
883
884
885
886
887
888
889
890
891
892
893
894
895
896
897
898
899
900
901
902
903
904
905
906
907
908
909
910
911
912
913
914
915
916
917
918
919
920
921
922
923
924
925
926
927
928
929
930
931
932
933
934
935
936
937
938
939
940
941
942
943
944
945
946
947
948
949
950
951
952
953
954
955
956
957
958
959
960
961
962
963
964
965
966
967
968
969
970
971
972
973
974
975
976
977
978
979
980
981
982
983
984
985
986
987
988
989
990
991
992
993
994
995
996
997
998
999
1000
NUT8595_proof ■ 23 July 2011 ■ 8/9
(Fundac~ao Carlos Chagas Filho de Amparo �a Pesquisa do Estadodo Rio de JaneirodFAPERJ). E.O. was recipient of the CapesFellowship; M.S.F., I.H.T., and A.S.C. were recipients of the FAPERJFellowship; and A.A.T. was a recipient of a CNPq fellowship.
References
[1] Barker DJ. The developmental origins of adult disease. Eur J Epidemiol2003;18:733–6.
[2] Moura EG, Passos MCF. Neonatal programming of body weight regulationand energetic metabolism. Biosci Rep 2005;25:251–69.
[3] de Moura EG, Lisboa PC, Passos MC. Neonatal programming ofneuroimmunomodulation-role of adipocytokines and neuropeptides.Neuroimmunomodulation 2008;15:176–88.
[4] Passos MCF, Ramos CF, Moura EG. Short and long term effects of malnu-trition in rats during lactation on the body weight of offspring. Nutr Res2000;20:1603–12.
[5] Fagundes AT, Moura EG, Passos MC, Oliveira E, Toste FP, Bonomo IT, et al.Maternal low-protein diet during lactation programmes body compo-sition and glucose homeostasis in the adult rat offspring. Br J Nutr2007;98:922–8.
[6] Lisboa PC, Fagundes AT, Denolato AT, Oliveira E, Bonomo IT, Alves SB, et al.Neonatal low-protein diet changes deiodinase activities and pituitary TSHresponse to TRH in adult rats. Exp Biol Med 2008;233:57–63.
[7] Rodrigues AL, de Moura EG, Passos MC, Dutra SC, Lisboa PC. Postnatal earlyovernutrition changes the leptin signalling pathway in the hypothalamic-pituitary-thyroid axis of young and adult rats. J Physiol 2009;587:2647–61.
[8] Moura EG, Santos RS, Lisboa PC, Alves SB, Bonomo IT, Fagundes AT, et al.Thyroid function and body weight programming by neonatal hyperthy-roidism in ratsdthe role of leptin and deiodinase activities. Horm MetabRes 2008;40:1–7.
[9] Toste FP, Moura EG, Lisboa PC, Fagundes AT, Oliveira E, Passos MC. Neonatalleptin treatment programs leptin hypothalamic resistance and interme-diary metabolic parameters in adult rats. Br J Nutr 2006;95:830–7.
[10] Toste FP, Alves SB, Dutra SC, Bonomo IT, Lisboa PC, Moura EG, et al.Temporal evaluation of the thyroid function of rats programmed by leptintreatment on the neonatal period. Horm Metab Res 2006;38:827–31.
[11] Bonomo IT, Lisboa PC, Pereira AR, Passos MCF, Moura EG. Prolactin inhi-bition in dams during lactation programs for overweight and leptin resis-tance in adult offspring. J Endocrinol 2007;192:339–44.
[12] de Moura EG, Bonomo IT, Nogueira-Neto JF, de Oliveira E, Trevenzoli IH,Reis AM, et al. Maternal prolactin inhibition during lactation programs formetabolic syndrome in adult progeny. J Physiol 2009;587:4919–29.
[13] Elmquist JK, Maratos-Flier E, Saper CB, Flier JS. Unraveling the centralnervous system pathways underlying responses to leptin. Nat Neurosci1998;1:445–50.
[14] Bjørbaek CJ. Central leptin receptor action and resistance in obesity. JInvestig Med 2009;57:789–94.
[15] Sahu A. Minireview: a hypothalamic role in energy balance with specialemphasis on leptin. Endocrinology 2004;145:2613–20.
[16] Vaisse C, Halaas JL, Horvath CM, Darnell JE Jr, Stoffel M, Friedman JM. Leptinactivation of Stat3 in the hypothalamus of wild-type and ob/ob mice butnot db/db mice. Nat Genet 1996;14:95–7.
[17] Hekerman P, Zeidler J, Bamberg-Lemper S, Knobelspies H, Lavens D,Tavernier J, et al. Pleiotropy of leptin receptor signalling is defined bydistinct roles of the intracellular tyrosines. FEBS J 2005;272:109–19.
[18] Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positionalcloning of the mouse obese gene and its human homologue. Nature1994;372:425–32.
[19] Cunnane SC, Hamadeh MJ, Liede AC, Thompson LU, Wolever TM, Jenkins DJ.Nutritional attributes of traditional flaxseed in healthy young adults. Am JClin Nutr 1995;61:62–8.
[20] Nestel PJ, Pomeroy SE, Sasahara T, Yamashita T, Liang YL, Dart AM, et al.Arterial compliance in obese subjects is improved with dietary plant n-3fatty acid from flaxseed oil despite increased LDL oxidizability. ArteriosclerThromb Vasc Biol 1997;17:1163–70.
[21] Thompson LU. Experimental studies on lignans and cancer. Baillieres ClinEndocrinol Metab 1998;12:691–705.
[22] Craig WJ. Health-promoting properties of common herbs. Am J Clin Nutr1999;70:491–9.
[23] Jenkins DJA, Kendall CWW, Vidgen E, Agarwal S, Rao AV, Rosenberg RS,et al. Health aspects of partially defatted flaxseed, including effects onserum lipids, oxidative measures, and ex vivo androgen and progestinactivity: a controlled crossover trial. Am J Clin Nutr 1999;69:395–402.
[24] Oomah BD. Flaxseed as a funtional food source. J Sci Food Agric2001;81:889–94.
[25] Bhathena SJ, Ali AA, Mohamed AI, Hansen CT, Velasquez MT. Differentialeffects of dietary flaxseed protein and soy protein on plasma triglycerideand uric acid levels in animal models. J Nutr Biochem 2002;13:684–9.
[26] Frank J, Eliasson C, Leroy-Nivard D, Budek A, Lundh T, Vessby B, et al.Dietary secoisolariciresinol diglucoside and its oligomers with 3-hydroxy-
Please cite this article in press as: Figueiredo MS, et al., Adipocyte moplemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002
3-methyl glutaric acid decrease vitamin E levels in rats. Br J Nutr2004;92:169–76.
[27] Fukumitsu S, Aida K, Ueno N, Ozama S, Takahashi Y, Kobori M. Flaxseedlignan attenuates high-fat diet induced fat accumulation and inducesadiponectin expression in mice. Br J Nutr 2008;100:669–76.
[28] Tou JCL, Chen J, Thompson LU. Flaxseed and its lignan precursor, secoiso-lariciresinol diglycoside, affect pregnancy outcome and reproductivedevelopment in rats. J Nutr 1998;128:1861–8.
[29] Collins TFX, Sprando RL, Black TN, Olejnik N, Wiesenfeld PW, Babu US, et al.Effects of flaxseed and defatted flaxseed meal on reproduction anddevelopment in rats. Food Chem Toxicol 2003;41:819–34.
[30] Troina AA, Figueiredo MS, Moura EG, Boaventura GT, Soares LL, Cardozo LF,et al. Maternal flaxseed diet during lactation alters milk composition andprograms the offspring body composition, lipid profile and sexual function.Food Chem Toxicol 2010;48:697–703.
[31] Figueiredo MS, de Moura EG, Lisboa PC, Troina AA, Trevenzoli IH, Oliveira E,et al. Flaxseed supplementation of rats during lactation changes theadiposity and glucose homeostasis of their offspring. LifeSci 2009;85:365–71.
[32] L�egr�adi G, Emerson CH, Ahima RS, Flier JS, Lechan RM. Leptin preventsfasting-induced suppression of prothyrotropin-releasing hormonemessenger ribonucleic acid in neurons of the hypothalamic paraventricularnucleus. Endocrinology 1997;138:2569–76.
[33] Seoane LM, Carro E, Tovar S, Casanueva FF, Dieguez C. Regulation of in vivoTSH secretion by leptin. Regul Pept 2000;25:25–9.
[34] Ortiga-Carvalho TM, Oliveira KJ, Soares BA, Pazos-Moura CC. The role ofleptin in the regulation of TSH secretion in the fed state: in vivo and in vitrostudies. J Endocrinol 2002;174:121–5.
[35] Nowak KW, Kaczmarek P, Mackowiak P, Ziolkowska A, Albertin G,Ginda WJ, et al. Rat thyroid gland expresses the long form of leptinreceptors, and leptin stimulates the function of the gland in euthyroid non-fasted animals. Int J Mol Med 2002;9:31–4.
[36] Isozaki O, Tsushima T, Nozoe Y, Miyakawa M, Takano K. Leptin regulation ofthe thyroids: negative regulation on thyroid hormone levels in euthyroidsubjects and inhibitory effects on iodide uptake and Naþ/I- symportermRNA expression in rat FRTL-5 cells. Endocr J 2004;51:415–23.
[37] Oliveira E, Fagundes AT, Alves SB, Pazos-Moura CC, Moura EG, Passos MC,et al. Chronic leptin treatment inhibits liver mitochondrial alpha-glycerol-beta-phosphate dehydrogenase in euthyroid rats. Horm Metab Res2007;39:867–70.
[38] Lisboa PC, Passos MC, Dutra SC, Santos RS, Bonomo IT, Cabanelas AP, et al.Increased 59-iodothyronine deiodinase activity is a maternal adaptivemechanism in response to protein restriction during lactation. J Endocrinol2003;177:261–7.
[39] Cabanelas A, Lisboa PC, Moura EG, Pazos-Moura CC. Leptin acute modu-lation of the 5’-deiodinase activities in hypothalamus, pituitary and brownadipose tissue of fed rats. Horm Metab Res 2006;38:481–5.
[40] Vicente LL, de Moura EG, Lisboa PC, Monte Alto Costa A, Amadeu T,Mandarim-de-Lacerda CA, et al. Malnutrition during lactation in rats isassociated with higher expression of leptin receptor in the pituitary ofadult offspring. Nutrition 2004;20:924–8.
[41] Santos-Silva AP, Moura EG, Pinheiro CR, Rios AS, Abreu-Villaca Y,Passos MC, et al. Neonatal nicotine exposure alters leptin signaling in thehypothalamus-pituitary-thyroid axis in the late postnatal period andadulthood in rats. Life Sci 2010;87:187–95.
[42] Pekary AE, Sattin A, Blood J. Rapid modulation of TRH and TRH-like peptiderelease in rat brain and peripheral tissues by leptin. Brain Res2010;1345:9–18.
[43] Gracious BL, Chirieac MC, Costescu S, Finucane TL, Youngstrom EA. HibbelnJR.Randomized, placebo-controlled trial of flax oil in pediatric bipolardisorder. Bipolar Disord 2010;12:142–54.
[44] Marques RG, Morales MM, Petroianu A. Brazilian law for scientific use ofanimals. Acta Cir Bras 2009;24:69–74.
[45] Fishbeck KL, Rasmussen KM. Effect of repeated cycles on maternal nutri-tional status, lactational performance and litter growth in ad libitum-fedand chronically food-restricted rats. J Nutr 1987;117:1967–75.
[46] Reeves PG, Nielsen FH, Fahey GC Jr. AIN-93 purified diets for laboratoryrodents: final report of the American Institute of Nutrition ad hoc writingcommittee on the reformulation of the AIN-76 rodent diet. J Nutr1993;123:1939–51.
[47] Fraga-Marques MC, Moura EG, Claudio-Neto S, Trevenzoli IH, Toste FP,Passos MCF, et al. Neonatal hyperleptinaemia programmes anxiety-like andnovelty seeking behaviours but not memory/learning in adult rats. HormBehav 2009;55:272–9.
[48] Fraga-Marques MC, Moura EG, Silva JO, Claudio-Neto S, Pereira-Toste F, Passos MC, et al. Effects of maternal hyperleptinaemia duringlactation on short-term memory/learning, anxiety-like and novelty-seeking behavioral traits of adult male rats. Behav Brain Res2010;206:147–50.
[49] Stansbie D, Browsey RW, Crettaz M, Demton RM. Acute effects in vivo ofanti-insulin serum on rates of acids synthesis and activities of acetyl-coenzyme A carboxilase and pyruvate dehydrogenase in liver and epidid-ymal adipose tissue of fed rats. Biochem J 1976;160:413–6.
rphology and leptin signaling in rat offspring from mothers sup-
1001

Q5
M. S. Figueiredo et al. / Nutrition xxx (2011) 1–9 9
1002
1003
1004
1005
1006
1007
1008
1009
1010
1011
1012
1013
1014
1015
1016
1017
1018
1019
1020
1021
1022
1023
1024
1025
1026
1027
1028
1029
1030
1031
1032
1033
1034
NUT8595_proof ■ 23 July 2011 ■ 9/9
[50] Hansen PA, Han DH, Nolte LA, Chen M, Holloszy JO. DHEA protects againstvisceral obesity and muscle insulin resistance in rats fed a high-fat diet. AmJ Physiol 1997;273:1704–8.
[51] Chen HC, Farese RV Jr. Determination of adipocyte size by computer imageanalysis. J Lipid Res 2002;43:986–9.
[52] de Oliveira E, Moura EG, Santos-Silva AP, Pinheiro CR, Lima NS, Nogueira-Neto JF, et al. Neonatal nicotine exposure causes insulin and leptin resis-tance and inhibits hypothalamic leptin signaling in adult rat offspring. JEndocrinol 2010;206:55–63.
[53] Carter JF. Potential of flaxseed and flaxseed oil in baked goods and otherproducts in human nutrition. Cereal Foods World 1993;38:753–9.
[54] Food and Nutrition Board. Dietary references intakes for energy,carbohydrate, fiber, fat, fatty acids, cholesterol, protein, and amino acids.Washington, DC: National Academy Press; 2005.
[55] Bugge A, Mandrup S. Molecular mechanisms and genome-wide aspects ofPPAR subtype specific transactivation. PPAR Res; 2010:1–12.
[56] Chechi K, Yasui N, Ikeda K, Yamori Y, Cheemas SK. Flax oil-mediated acti-vation of PPAR-g correlates with reduction of hepatic lipid accumulation inobese spontaneously hypertensive/NDmcr-cp rats, a model of the meta-bolic syndrome. Br J Nutr 2010;104:1313–21.
[57] Ahima RS, Osei SY. Leptin signaling. Physiol Behav 2004;81:223–41.
Please cite this article in press as: Figueiredo MS, et al., Adipocyte moplemented with..., Nutrition (2011), doi:10.1016/j.nut.2011.07.002
[58] Kieffer TJ, Habener JF. The adipoinsular axis effects of leptin on pancreaticbeta-cells. Am J Physiol 2000;278:E1–14.
[59] Pedersen SB, Hansen PS, Lund S, Andersen PH, Odgaard A, Richelsen B.Identification of oestrogen receptors and oestrogen receptor mRNA inhuman adipose tissue. Eur J Clin Invest 1996;26:262–9.
[60] Kristensen K, Pedersen SB, Richelsen B. Regulation of leptin by steroidhormones in rat adipose tissue. Biochem Biophys Res Commun1999;259:624–30.
[61] Trevenzoli IH, Rodrigues AL, Oliveira E, Thole AA, Carvalho L,Figueiredo MS, et al. Leptin treatment during lactation programs leptinsynthesis, intermediate metabolism, and liver microsteatosis in adult rats.Horm Metab Res 2010;42:483–90.
[62] Velkoska E, Cole TJ, Morris MJ. Early dietary intervention: long-term effectson blood pressure, brain neuropeptide Y, and adiposity markers. Am JPhysiol Endocrinol Metab 2005;288:1236–43.
[63] Mathieu P, Pibarot P, Larose E, Poirier P, Marette A, Despr�es JP. Visceralobesity and the heart. Int J Biochem Cell Biol 2008;40:821–36.
[64] Yu H, Bi Y, Ma W, He L, Yuan L, Feng J, et al. Long-term effects of highlipid and high energy diet on serum lipid, brain fatty acid composition,and memory and learning ability in mice. Int J Dev Neurosci2010;28:271–6.
rphology and leptin signaling in rat offspring from mothers sup-
1035

Our reference: NUT 8595 P-authorquery-v9
AUTHOR QUERY FORM
Journal: NUT
Article Number: 8595
Please e-mail or fax your responses and any corrections to:
E-mail: [email protected]
Fax: (717) 738-9478; (717) 738-9479
Dear Author,
Please check your proof carefully and mark all corrections at the appropriate place in the proof (e.g., by using on-screen
annotation in the PDF file) or compile them in a separate list. To ensure fast publication of your paper please return your
corrections within 48 hours.
For correction or revision of any artwork, please consult http://www.elsevier.com/artworkinstructions.
Any queries or remarks that have arisen during the processing of your manuscript are listed below and highlighted by flags in
the proof.
Location
in articleQuery / Remark: Click on the Q link to find the query’s location in text
Please insert your reply or correction at the corresponding line in the proof
Q1 Please supply section head.
Q2 Please verify addresses in all affilations are correct and complete.
Q3 Please spell out TRH.
Q4 Please verify manufacturer and location are correct for Nuvilab.
Q5 Please provide vol. no. for ref. 55.
Thank you for your assistance.
Related Documents











