Journal of Colloid and Interface Science 300 (2006) 131–140 www.elsevier.com/locate/jcis Adhesion force between calcium oxalate monohydrate crystal and kidney epithelial cells and possible relevance for kidney stone formation Yakov I. Rabinovich ∗ , Madhavan Esayanur, Saijit Daosukho, Karen J. Byer, Hassan E. El-Shall, Saeed R. Khan University of Florida, PO Box 6135, Gainesville, FL 32611-6135, USA Received 5 January 2006; accepted 28 March 2006 Available online 4 April 2006 Abstract AFM interaction force measurements have been performed between calcium oxalate monohydrate crystal (COM) colloidal probes and mono- layers of renal epithelial cells (on a polymer substrate) in artificial urine (AU) solutions. The adhesion force was measured for the COM/MDCK cell interaction, while no adhesion force was found for the COM/LLC-PK 1 cell interaction. Long-range repulsive forces for both lines of cells were measured in the range of 2–3 μm. After removal of the cell from the substrate by the AU flow, the basal membrane (BM), with a thickness of 100–200 nm, remained on the substrate. In this case, the shorter-range repulsive forces were found on the extending (approaching) portion of force/indentation curves. Similar to the COM/MDCK cell interaction, the retracting portions of curves for COM/basal membranes have shown the existence of the attractive force of adhesion for the interaction of COM with a BM of MDCK cells, while no adhesion was found for COM/BM LLC-PK 1 cells interaction. No adhesion force was found for the interaction of a BM (of any cells) with the silicon nitride tip. Besides the hy- drodynamic reasons, the adhesion difference between LLC-PK 1 and MDCK cells possibly explains the preferential deposition of crystals only in collecting ducts (lined with MDCK-type cells) and the lack of the crystal deposition in the proximal tubules (lined with LLC-PK 1 -type cells). Previous treatments of cells with oxalate alone increased the adhesion force COM/BM MDCK; however, even after oxalate treatment there was small or no adhesion between COM and BM LLC-PK 1 cells. Note that the adhesion force for COM/BM MDCK is practically independent of the probe velocity, i.e., does not have the viscous origin. Evaluation of the adhesion energy shows that this force should be related to the ionic or hydrogen bonds of samples. © 2006 Elsevier Inc. All rights reserved. Keywords: AFM, MDCK and LLC-PK 1 cells; Calcium oxalate monohydrate; Adhesion force and kidney stone 1. Introduction In the last ten years, several papers have appeared regard- ing measurement of the interaction between an AFM tip and biological objects, such as cells, DNA, proteins, antibodies, bacteria, microorganism, etc. (e.g., Refs. [1–15]). Some of these papers have dealt with renal epithelial cells, such as MDCK and LLC-PK 1 [4,5]. The properties of these cells were investigated in connection with the problem of forming of the kidney stones. In our previous paper [5] the interaction of an AFM tip with kidney epithelial cells (MDCK and LLC-PK 1 ) was measured. * Corresponding author. Corresponding address: 206 PERC Bldg, Bldg # 746, University of Florida, Gainesville, FL 32611-6135, USA. E-mail address: [email protected]fl.edu (Y.I. Rabinovich). Only repulsive forces were found on both the extending and retracting parts of the force/indentation curves (excluding the viscous attractive force for the large probe retracting rates). Calcium oxalate monohydrate (COM) is a major constituent in the majority of kidney stones; as a result, the interaction of the aforementioned cells with COM, rather than with the sili- con nitride tip of AFM, may provide some important clues to the pathology of kidney stone formation. Surprisingly, attrac- tive (adhesion) forces and binding affinity between COM and kidney cells have rarely been investigated although they may play a critical role in kidney stone nucleation and formation [16,17]. The measurement of COM/cell interaction forces in vitro is important because it simulates the approach and the detachment of the small COM particles (or aggregates) to and from the ep- 0021-9797/$ – see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.jcis.2006.03.070

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Colloid and Interface Science 300 (2006) 131–140www.elsevier.com/locate/jcis
Adhesion force between calcium oxalate monohydrate crystal and kidneyepithelial cells and possible relevance for kidney stone formation
Yakov I. Rabinovich ∗, Madhavan Esayanur, Saijit Daosukho, Karen J. Byer,Hassan E. El-Shall, Saeed R. Khan
University of Florida, PO Box 6135, Gainesville, FL 32611-6135, USA
Received 5 January 2006; accepted 28 March 2006
Available online 4 April 2006
Abstract
AFM interaction force measurements have been performed between calcium oxalate monohydrate crystal (COM) colloidal probes and mono-layers of renal epithelial cells (on a polymer substrate) in artificial urine (AU) solutions. The adhesion force was measured for the COM/MDCKcell interaction, while no adhesion force was found for the COM/LLC-PK1 cell interaction. Long-range repulsive forces for both lines of cellswere measured in the range of 2–3 µm. After removal of the cell from the substrate by the AU flow, the basal membrane (BM), with a thicknessof 100–200 nm, remained on the substrate. In this case, the shorter-range repulsive forces were found on the extending (approaching) portion offorce/indentation curves. Similar to the COM/MDCK cell interaction, the retracting portions of curves for COM/basal membranes have shown theexistence of the attractive force of adhesion for the interaction of COM with a BM of MDCK cells, while no adhesion was found for COM/BMLLC-PK1 cells interaction. No adhesion force was found for the interaction of a BM (of any cells) with the silicon nitride tip. Besides the hy-drodynamic reasons, the adhesion difference between LLC-PK1 and MDCK cells possibly explains the preferential deposition of crystals onlyin collecting ducts (lined with MDCK-type cells) and the lack of the crystal deposition in the proximal tubules (lined with LLC-PK1-type cells).Previous treatments of cells with oxalate alone increased the adhesion force COM/BM MDCK; however, even after oxalate treatment there wassmall or no adhesion between COM and BM LLC-PK1 cells. Note that the adhesion force for COM/BM MDCK is practically independent ofthe probe velocity, i.e., does not have the viscous origin. Evaluation of the adhesion energy shows that this force should be related to the ionic orhydrogen bonds of samples.© 2006 Elsevier Inc. All rights reserved.
Keywords: AFM, MDCK and LLC-PK1 cells; Calcium oxalate monohydrate; Adhesion force and kidney stone
1. Introduction
In the last ten years, several papers have appeared regard-ing measurement of the interaction between an AFM tip andbiological objects, such as cells, DNA, proteins, antibodies,bacteria, microorganism, etc. (e.g., Refs. [1–15]). Some of thesepapers have dealt with renal epithelial cells, such as MDCK andLLC-PK1 [4,5]. The properties of these cells were investigatedin connection with the problem of forming of the kidney stones.
In our previous paper [5] the interaction of an AFM tip withkidney epithelial cells (MDCK and LLC-PK1) was measured.
* Corresponding author. Corresponding address: 206 PERC Bldg, Bldg# 746, University of Florida, Gainesville, FL 32611-6135, USA.
E-mail address: [email protected] (Y.I. Rabinovich).
0021-9797/$ – see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.jcis.2006.03.070
Only repulsive forces were found on both the extending andretracting parts of the force/indentation curves (excluding theviscous attractive force for the large probe retracting rates).Calcium oxalate monohydrate (COM) is a major constituentin the majority of kidney stones; as a result, the interaction ofthe aforementioned cells with COM, rather than with the sili-con nitride tip of AFM, may provide some important clues tothe pathology of kidney stone formation. Surprisingly, attrac-tive (adhesion) forces and binding affinity between COM andkidney cells have rarely been investigated although they mayplay a critical role in kidney stone nucleation and formation[16,17].
The measurement of COM/cell interaction forces in vitro isimportant because it simulates the approach and the detachmentof the small COM particles (or aggregates) to and from the ep-

132 Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140
ithelial kidney cells. Strong repulsive forces indicate that theparticles will not adhere to the surface and, as a result, the di-rect contact of COM particles with cells is not likely to retainand promote the growth of the COM crystals on the epithelium.Taking into account the repulsive interaction force and energyto be proportional to the crystal radius, it should be noted thatonly very small crystals of quasi-molecular sizes can achievethe direct contact with the kidney walls. On the other hand,only if strong adhesive interactions are present then COM crys-tals are likely to stick to the tubule wall and may reside forextended periods of time in supersaturated solution conditions,allowing for growth. In vitro measurements allow for quantita-tive characteristics of the COM/cell interaction to be obtainedusing the different types of cells, with controlled solution treat-ments and compositions. The obtained force values will allowfor an understanding of the nature of the binding between cellsand COM crystals.
In a recent paper [6] the adhesion force between the (100)face of COM crystals and the silicon nitride tip chemicallymodified by different functional groups was measured in CaOxsolutions. Forces were measured under solutions of CaOx andimportant results were obtained. A dependence of the adhesionforce on the type of modifying molecules was found. The max-imum adhesive force was found for the tip modified by an Aulayer and S(CH2)10COO− molecules. For this case, the valuesof the adhesion force were near 4 nN. In a new paper [7] theauthors showed that the same adhesion force acts between the(100)- face of the COM crystal and a tip modified by an Au-layer and S(CH2)2NHC(NH2) + NH2 molecules. In that paper,the role of the crystal faces was investigated and it was shownthat the order of the adhesion force is (100) > (121) > (010).
Other papers [16,17] have been dedicated to the investiga-tion of the binding affinity between COM crystals and MDCKcells. In these papers the value of the binding energy was evalu-ated qualitatively by measuring the amount of CaOx adhered tothe cells. A significant difference was found in the affinity forCOM and calcium oxalate dihydrate crystals. It was also shownthat magnesium and calcium at certain concentrations (�100and �50 mM for Mg and Ca, respectively) promoted the forcesof adhesion.
The interaction of a chemically modified tip with a COMcrystal [6] is not necessarily equal to the interaction COM crys-tals with biological cells, which is important for the process ofthe kidney stone formation. In this study the interaction forcebetween COM and renal epithelial cells, such as MDCK andLLC-PK1, were measured under the artificial urine solutions.Special attention was dedicated to attractive (adhesive) forces,which were found at the retracting portions of force/positioncurves. These measurements of the attractive force allowed usto draw certain conclusions about the possibility of the crystaladherence to the different types of the epithelial cells. Crystaladherence to the renal epithelium is considered an importantevent during crystal deposition in the kidney and developmentof the kidney stones. The measurements of the repulsive forceobtained were compared with previous results for tip/cell inter-action and conclusions were drawn about elastic properties ofthe cells and their basal lamina.
2. Experiment and materials
An Atomic Force Microscope (AFM) with a Nanoscope IIIcontroller from Digital Instruments, CA, was used for the forcemeasurements. Triangular cantilevers (NP-S type) from VeecoInstruments, CA, with a normal spring constant of 0.12 N/mwere used. All the force measurements were carried out inthe standard liquid chamber of the AFM. In distinction fromRef. [5], where the force was measured between a silicon ni-tride tip and the biological cell, in the present paper the forcewas measured between a cell and the COM crystal, mounted bythermal epoxy glue onto the tip, as it is shown in Fig. 1. TheCOM crystal were synthesized from super-saturated solutionof potassium oxalate and calcium chloride, as was describedin Refs. [18,19]. The usual size of crystals applied was near10 × 5 × 3 µm. For the force measurements, only the singlecrystals were used rather than twinned crystals. The orientationof the crystal on the cantilever was controlled by the opticalmicroscope and micromanipulator. Fig. 1 shows the usual ori-entation of the crystals. In the different experimental sets, thepossible deviation from this orientation was ca. several degrees.
The procedure of the calculation of indentation is standard,i.e., the indentation is calculated as the difference betweenthe piezo-crystal displacement and the deflection of the can-tilever. The final form of the data, represented as force, F ,versus indentation depth, δ, has been corrected for the dis-tance taking into account the “contact line” as related to thesoft body [20,21]. As in many published reports concerning themechanical properties of the cells, the results of the present pa-per are represented in the form of the force/indentation graphs(rather than force/distance graphs). The reason of represent-ing data using the force/indentation graphs is that they facili-tate easy comparison with the equations applied. These equa-tions used the mechanical properties of the deformed sam-ples, for example, the Young modulus of the biological cells.Here, the zero distance is taken as the samples position whenthe repulsion starts. This means actually that zero indentationis related to the first contact of the cell and the COM crys-tal.
Two cell lines were selected for the current study, LLC-PK1, a proximal tubular epithelial cell line derived from porcinekidneys and MDCK, a collecting duct line derived from ca-nine kidneys. The same COM crystals (and cantilevers) wereused for measurements with both cells lines. Both LLC-PK1
and MDCK have microvilli brush borders and form continu-ous monolayers in culture with uniform cell-to-cell contacts.The LLC-PK1 cells have longer, and a larger number of mi-crovilli compared to MDCK cells. Microvilli may affect therepulsive force and possibly the adhesion force. This effectcan be related to the chemical composition of microvilli (andbonding between COM and microvilli), and their mechanicalproperties (elasticity). Unfortunately, these effects could not beinvestigated in the present paper. We can recommend Ref. [14]for a better understanding the structure–function relationshipin the membrane and microvilli. The growth and acclimatiza-tion of LLC-PK1 (ATCC, Gaithersburg, MD, Cat # CL-101)

Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140 133
Fig. 1. COM crystal on the cantilever and the COM crystals photos.
and MDCK (ATCC, Gaithersburg, MD, Cat # CCR-34) cells isdescribed in Ref. [5].
To check the effect of the potassium oxalate on the cells,the latter were exposed for 4 h to 500 µM/ml potassium ox-alate (Ox) (Fisher Scientific, Atlanta, GA, Cat # P273-500).This study was made because the hyperoxaluria (the excess ofthe oxalate concentration) is one of the dangerous kidney dis-eases. Before exposure to Ox, the acclimatization media [5] wasremoved and fresh acclimatization media was added with orwithout Ox.
All measurements were conducted in an artificial urine (AU)solution. AU was prepared as it was described in Ref. [22]. Thecomposition of AU was, as follows, 0.1055 M NaCl; 0.0323 MNaH2PO4·2H2O; 0.00385 M MgSO4; 0.01695 M Na2SO4;0.0637 M KCl; 0.0363 M NH4Cl; 0.00062 M NH4OH. Thus,the total amount of the electrolyte was near 0.25 M (includingthe divalent ions). In our opinion, this large electrolyte con-centration excludes the action of the ion-electrostatic (electricdouble layer-) force due to the screening effect.
The values of the elasticity, k, elastic modulus, E, and adhe-sion (detachment) force, 〈Fad〉, have been found as the averageof experimental extending (k and E) or retracting (〈Fad〉) partsof the force/position curves for COM cell interaction. Theseforce/position curves were measured in at least three seriesof experiments, and each series was obtained with a certainCOM/crystal on the certain cantilever. In each series, at least20 different places of the cell were investigated, and at least 5–10 measurements performed in each place. Therefore, the totalnumber of values from which the average value was calculatedwas 300–600. The standard deviation of the resulting elastic-ity, k, adhesion force, 〈Fad〉, and the elastic modulus, E, wereabout ±30% and are given in Tables 1, 3 and 4. The large valuesof deviation may be related, first of all, to different propertiesof the different sets of the cells.
A brief description of the kidney and the renal epithelial cellsis given in Ref. [5]. The properties of these cells are written inmore detail in Refs. [23–25]. The size of the collecting ductscells could reach ca. 120 µm, which is larger than the size ofthe COM crystals and the size of the crystal top, which actuallyparticipates in the interaction with cells.
3. Theory
For small indentations, Refs. [1–5], the linear empirical for-mula was suggested to be
(1)F = kδ,
where δ is the indentation distance and k is the proportional-ity coefficient. The reference point for the indentation valueis the distance (position), at which the repulsive force starts toact. Unfortunately, this point cannot be found precisely enough,which contributes the additional error to the calculated valueof k. To decrease this error, the reference points for the indenta-tion were determined by the extrapolation of the linear portionof the force/distance curves to zero force line (abscissa).
In Ref. [5], our hypothesis suggests that this force is a resultof the surface pressure of the cell membrane. In this case, theresulting repulsive force should be independent of the probesize. Regardless of the accuracy of the proposed explanation, itshould be noted that in all experimental works, the linear part ofthe force/indentation curves was found for small indentations.
For larger indentations, the Hertzian force between elasticcell and the probe was suggested. Analysis of this force for thedifferent shapes of probe was done in Refs. [26,27]. For exam-ple, for the conical shape of a probe with the opening angle α,the repulsive force will be equal
(2)F = (2/π)E tan(α)δ2/(1 − ν2),
where E is the elastic (Young’s) modulus of the cell materialand ν is the Poisson coefficient. In paper [5] we suggested thefollowing combining formula for the tip/cell repulsion
(3)F = kδ + (2/π)E tanα(δ − δcr
)2/(1 − ν2),
where δcr is the minimal indentation, above which the Hertzianforce starts to act.
Equations (2) and (3) are valid only for the conical probes.However in the case of crystal/cell interaction, as it is seen inFig. 1, the top of the COM crystal interacting with cells hasthe pyramidal shape rather than the conical one. Unfortunately,there is no formula for a pyramidal probe. For this reason,Eq. (3) was used as the first approximation for the calcula-tion of the elastic modulus, E. The angle of α = 45◦ was usedin Eq. (3); which is the average angle obtained from electron
134 Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140
micro-photographs of the COM crystals. Of course, the valuesof E for the cells obtained in the present study should be con-sidered as less accurate than the results of Ref. [5], where atip with a well known cone shape was used. However, resultsfor COM cells interaction are more closely related to the actualtask, i.e., the mechanism of the kidney stone formation.
Besides the absolute value of the elastic modulus, it is pos-sible to determine the ratio of the elastic moduli obtained fordifferent conditions or for different lines of the cells. This ap-proach avoids many systematic errors, including those relatedto the specific shape of the top of the crystal, and if two valuesof moduli are obtained for the same crystal. However, the ran-dom error of that ratio is approximately twice as big as the errorof each absolute value of the modulus, because their errors aresummed for calculation of the ratio. Note that one of the rea-sons of the possible error of the elastic modulus is the largenumber of the parameters in Eq. (5). The sensitivity of the re-sulting value of the elastic modulus to the choice of the value ofδcr is shown in the appendix. However, from this point of view,results of the present study are of the same level of accuracy asresults obtained in many other papers dealing with AFM mea-surements of the Hertz’s parameters of cells, as in Refs. [1,4,5].
It should also be mentioned that there are some papers thatsuggest alternative formulae (instead of Eq. (3)) for calculationof elastic modulus of soft samples. Particularly, theory was de-veloped in Ref. [28] for polyelectrolyte microcapsules, which,in theory, is able to calculate the elastic modulus of a thinmembrane from the force/deformation curves. Unfortunately,we could not compare our experimental results with the for-mulae developed in Ref. [28] or with experimental results ofRef. [29] because the thickness of plasma membrane and exactsize of cells are unknown. Moreover, for the interaction of thetip (and especially COM crystal) with renal cells, the model ofRef. [29] is non-applicable due to small radius of tip as com-pared with the cell radius. However, we hope that using thecolloidal probe method and measuring the cell radius and thick-ness of the plasma membrane will allow checks, if the theorysuggested for microcapsules with the thin membrane is validfor the cell surrounding by the plasma membrane.
4. Adhesion force
The investigation of adhesion force between COM and cellsis especially important from the point of view of the kidneystone formation. Adhesion force was measured as the detach-ment force required to pull COM crystals off the cells. Notethat the absolute value of Fad is not adequate enough to charac-terize the COM/cell (or COM/BM or cells) interaction, becausethis force depends on the crystal size. The actual characteris-tic of the interaction is the value of the specific (per unit area)surface energy of the adhesion, γ , which can be calculated, asfollows [30]
(4)Fad = 4πγR,
where Fad is the adhesion force between quasi-semisphericalsurface of the crystal edge (top) with radius R and the flat sur-face of the cell. In the detachment process, the flat surface of the
soft cells deforms, resulting in the following alternative formula[31], which is valid for soft materials and can be used insteadof Eq. (4),
(5)Fad = 3πγR.
There are certain criteria to find, which one from these equa-tions should be applied. Generally, Eq. (4) is valid for hardsolids, with the small deformation due to adhesion force, whileEq. (5) is applicable for soft solids, with the large deforma-tion of the surface due to adhesion force during the detachmentprocess.
Below we use Eq. (5) to calculate the surface energy, γ . Ofcourse, the application of Eq. (5) for calculation of the surfaceenergy is only the first approximation, because the top of theCOM crystal has pyramidal shape, while Eq. (5) is valid onlyfor the sphere/plate interaction. Moreover, the proteins mole-cules on the cells surface interacting with the crystal, do notform ideally smooth flat surface, as suggests Eq. (5).
5. Results and discussion
The design of the Nanoscope III does not allow us to see thepoint of contact of the COM crystal with cells precisely. For thisreason, the force measurements were made at many differentpoints and averaged, which we believe relates to the interactionwith the overall plasma membrane.
5.1. COM/cells interaction
As shown in Ref. [5] for tip/cell interaction, the COM/cellforce acts in the long-indentation range near several microns.
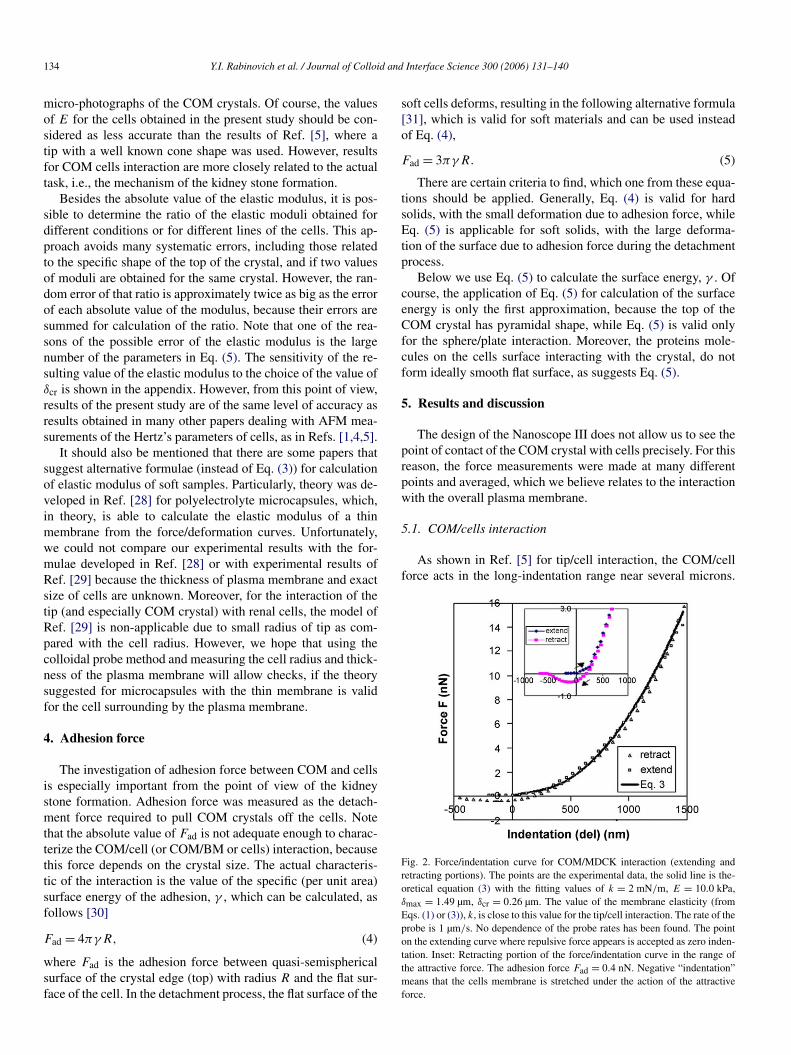
Fig. 2. Force/indentation curve for COM/MDCK interaction (extending andretracting portions). The points are the experimental data, the solid line is the-oretical equation (3) with the fitting values of k = 2 mN/m, E = 10.0 kPa,δmax = 1.49 µm, δcr = 0.26 µm. The value of the membrane elasticity (fromEqs. (1) or (3)), k, is close to this value for the tip/cell interaction. The rate of theprobe is 1 µm/s. No dependence of the probe rates has been found. The pointon the extending curve where repulsive force appears is accepted as zero inden-tation. Inset: Retracting portion of the force/indentation curve in the range ofthe attractive force. The adhesion force Fad = 0.4 nN. Negative “indentation”means that the cells membrane is stretched under the action of the attractiveforce.
Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140 135
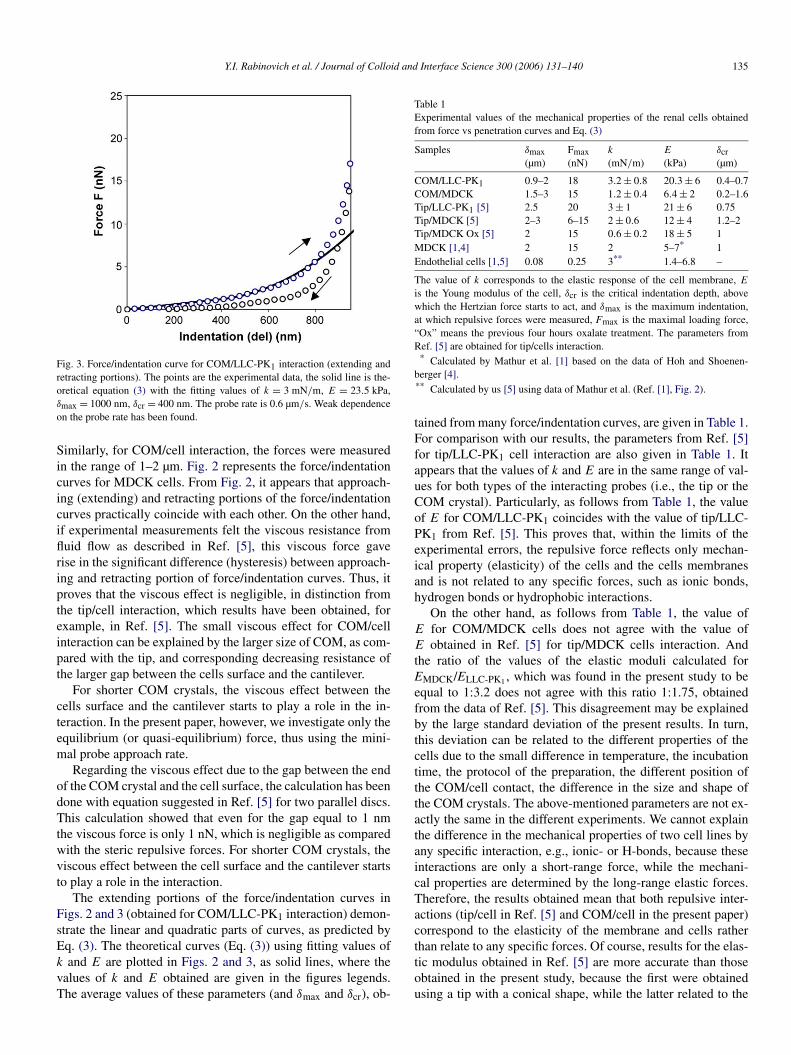
Fig. 3. Force/indentation curve for COM/LLC-PK1 interaction (extending andretracting portions). The points are the experimental data, the solid line is the-oretical equation (3) with the fitting values of k = 3 mN/m, E = 23.5 kPa,δmax = 1000 nm, δcr = 400 nm. The probe rate is 0.6 µm/s. Weak dependenceon the probe rate has been found.
Similarly, for COM/cell interaction, the forces were measuredin the range of 1–2 µm. Fig. 2 represents the force/indentationcurves for MDCK cells. From Fig. 2, it appears that approach-ing (extending) and retracting portions of the force/indentationcurves practically coincide with each other. On the other hand,if experimental measurements felt the viscous resistance fromfluid flow as described in Ref. [5], this viscous force gaverise in the significant difference (hysteresis) between approach-ing and retracting portion of force/indentation curves. Thus, itproves that the viscous effect is negligible, in distinction fromthe tip/cell interaction, which results have been obtained, forexample, in Ref. [5]. The small viscous effect for COM/cellinteraction can be explained by the larger size of COM, as com-pared with the tip, and corresponding decreasing resistance ofthe larger gap between the cells surface and the cantilever.
For shorter COM crystals, the viscous effect between thecells surface and the cantilever starts to play a role in the in-teraction. In the present paper, however, we investigate only theequilibrium (or quasi-equilibrium) force, thus using the mini-mal probe approach rate.
Regarding the viscous effect due to the gap between the endof the COM crystal and the cell surface, the calculation has beendone with equation suggested in Ref. [5] for two parallel discs.This calculation showed that even for the gap equal to 1 nmthe viscous force is only 1 nN, which is negligible as comparedwith the steric repulsive forces. For shorter COM crystals, theviscous effect between the cell surface and the cantilever startsto play a role in the interaction.
The extending portions of the force/indentation curves inFigs. 2 and 3 (obtained for COM/LLC-PK1 interaction) demon-strate the linear and quadratic parts of curves, as predicted byEq. (3). The theoretical curves (Eq. (3)) using fitting values ofk and E are plotted in Figs. 2 and 3, as solid lines, where thevalues of k and E obtained are given in the figures legends.The average values of these parameters (and δmax and δcr), ob-
Table 1Experimental values of the mechanical properties of the renal cells obtainedfrom force vs penetration curves and Eq. (3)
Samples δmax(µm)
Fmax(nN)
k
(mN/m)E
(kPa)δcr(µm)
COM/LLC-PK1 0.9–2 18 3.2 ± 0.8 20.3 ± 6 0.4–0.7COM/MDCK 1.5–3 15 1.2 ± 0.4 6.4 ± 2 0.2–1.6Tip/LLC-PK1 [5] 2.5 20 3 ± 1 21 ± 6 0.75Tip/MDCK [5] 2–3 6–15 2 ± 0.6 12 ± 4 1.2–2Tip/MDCK Ox [5] 2 15 0.6 ± 0.2 18 ± 5 1MDCK [1,4] 2 15 2 5–7* 1Endothelial cells [1,5] 0.08 0.25 3** 1.4–6.8 –
The value of k corresponds to the elastic response of the cell membrane, E
is the Young modulus of the cell, δcr is the critical indentation depth, abovewhich the Hertzian force starts to act, and δmax is the maximum indentation,at which repulsive forces were measured, Fmax is the maximal loading force,“Ox” means the previous four hours oxalate treatment. The parameters fromRef. [5] are obtained for tip/cells interaction.
* Calculated by Mathur et al. [1] based on the data of Hoh and Shoenen-berger [4].** Calculated by us [5] using data of Mathur et al. (Ref. [1], Fig. 2).
tained from many force/indentation curves, are given in Table 1.For comparison with our results, the parameters from Ref. [5]for tip/LLC-PK1 cell interaction are also given in Table 1. Itappears that the values of k and E are in the same range of val-ues for both types of the interacting probes (i.e., the tip or theCOM crystal). Particularly, as follows from Table 1, the valueof E for COM/LLC-PK1 coincides with the value of tip/LLC-PK1 from Ref. [5]. This proves that, within the limits of theexperimental errors, the repulsive force reflects only mechan-ical property (elasticity) of the cells and the cells membranesand is not related to any specific forces, such as ionic bonds,hydrogen bonds or hydrophobic interactions.
On the other hand, as follows from Table 1, the value ofE for COM/MDCK cells does not agree with the value ofE obtained in Ref. [5] for tip/MDCK cells interaction. Andthe ratio of the values of the elastic moduli calculated forEMDCK/ELLC-PK1 , which was found in the present study to beequal to 1:3.2 does not agree with this ratio 1:1.75, obtainedfrom the data of Ref. [5]. This disagreement may be explainedby the large standard deviation of the present results. In turn,this deviation can be related to the different properties of thecells due to the small difference in temperature, the incubationtime, the protocol of the preparation, the different position ofthe COM/cell contact, the difference in the size and shape ofthe COM crystals. The above-mentioned parameters are not ex-actly the same in the different experiments. We cannot explainthe difference in the mechanical properties of two cell lines byany specific interaction, e.g., ionic- or H-bonds, because theseinteractions are only a short-range force, while the mechani-cal properties are determined by the long-range elastic forces.Therefore, the results obtained mean that both repulsive inter-actions (tip/cell in Ref. [5] and COM/cell in the present paper)correspond to the elasticity of the membrane and cells ratherthan relate to any specific forces. Of course, results for the elas-tic modulus obtained in Ref. [5] are more accurate than thoseobtained in the present study, because the first were obtainedusing a tip with a conical shape, while the latter related to the
136 Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140
Table 2The presence (+) and the absence (−) of Fad for different interaction systems
Samples Fad Samples Fad
Tip/LLC-PK1 − COM/LLC-PK1 −Tip/MDCK − COM/MDCK +Tip/BM LLC-PK1 − COM/BM LLC-PK1 −Tip/BM MDCK − COM/BM MDCK +
pyramidal crystal top. However, results for COM crystal are im-portant because they characterize the interaction of the samples,which participate in the kidney stone formation.
In regards to the retracting portions of the force/positioncurves, they demonstrate the significant difference between re-sults obtained for COM/MDCK cell and tip/MDCK cell. Ac-tually, the retracting portion of COM/LLC-PK1 interaction isshown in Fig. 3. It appears that for these cells only repulsionforce was found. In turn, Fig. 2 proves the existence of theadhesion force (near 0.4 nN in this figure) for this case. Forclarity, the attractive part of the retracting curve is shown asan inset. From Fig. 2, it also appears that attractive force actsnot only at zero position (contact) between COM crystal andcell (where the force/distance curve intersect the abscissa axis),but also in the range of up to 500 nm in distance. This mayhowever produce the wrong impression with regards to dis-tance because the cell plasma membrane is not a hard boundaryand its position changes (due to plasma membrane stretching)with the interaction force. However, the membrane may stickto the crystal and follow it in the detachment process, as men-tioned above. Therefore, it is most probable that all valleyson the force/indentation curve are actually related to the con-tact position between various points of COM crystal and thecell membrane. The COM/MDCK cells adhesion is likely tobe different for different places of COM crystal and the cellmembrane. The scattering of the adhesion force in the differentplaces can be as much as 2 fold.
For tip/cell interaction, the existence of the adhesion force, ifany, could not be found on the background long-range viscousforces. Therefore, using COM crystal instead of the silicon ni-tride tip, allowed us to decrease the viscous background andmeasure the adhesion force between COM and cell more di-rectly. Moreover, from the point of view of highlighting themechanism of the kidney stone formation, the adhesion forceof COM/cell is much more important than possible adhesiontip/cell because the adhesion force can be related to the specificinteraction, for example, as ionic and hydrogen bonds.
The presence or absence of attractive (adhesion) force isgiven in Table 2. In the table the results are given both forCOM/cells and for COM/basal membrane (BM). The lat-ter results will be described below. The results for tip/cellsand tip/basal, obtained in Ref. [5] are also given in Table 2.We return to the discussion of the adhesion force betweenCOM/MDCK cells in the next section.
5.2. COM/basal membrane interaction
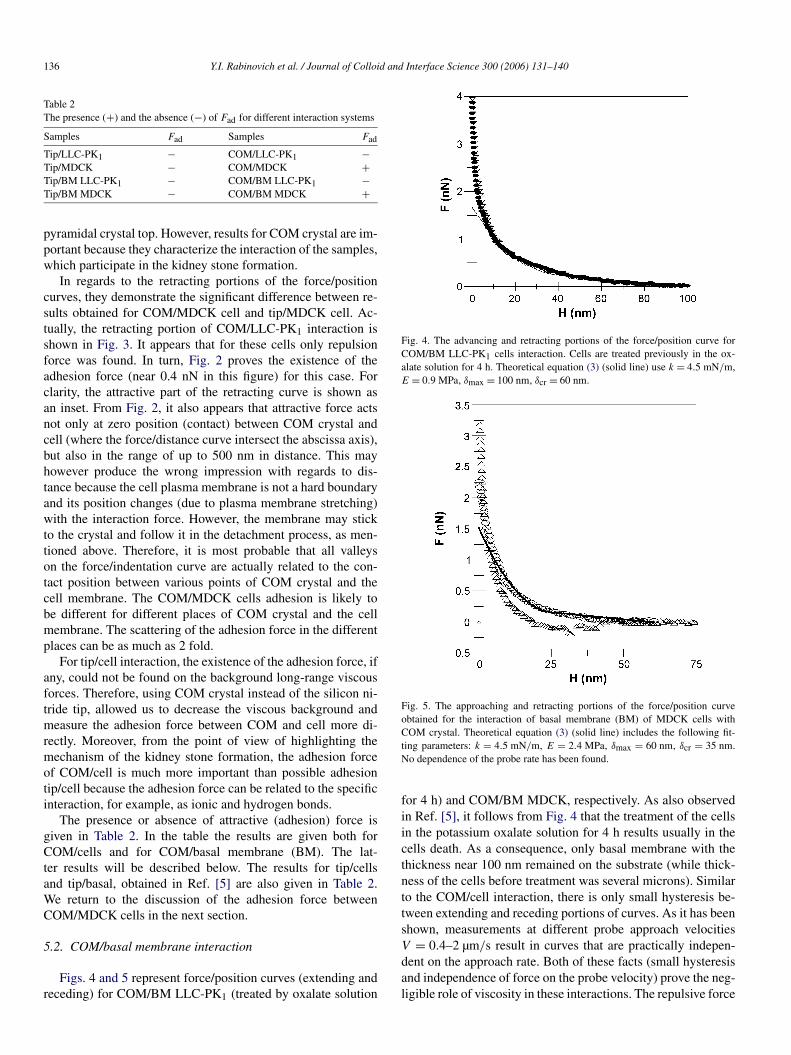
Figs. 4 and 5 represent force/position curves (extending andreceding) for COM/BM LLC-PK1 (treated by oxalate solution
Fig. 4. The advancing and retracting portions of the force/position curve forCOM/BM LLC-PK1 cells interaction. Cells are treated previously in the ox-alate solution for 4 h. Theoretical equation (3) (solid line) use k = 4.5 mN/m,E = 0.9 MPa, δmax = 100 nm, δcr = 60 nm.
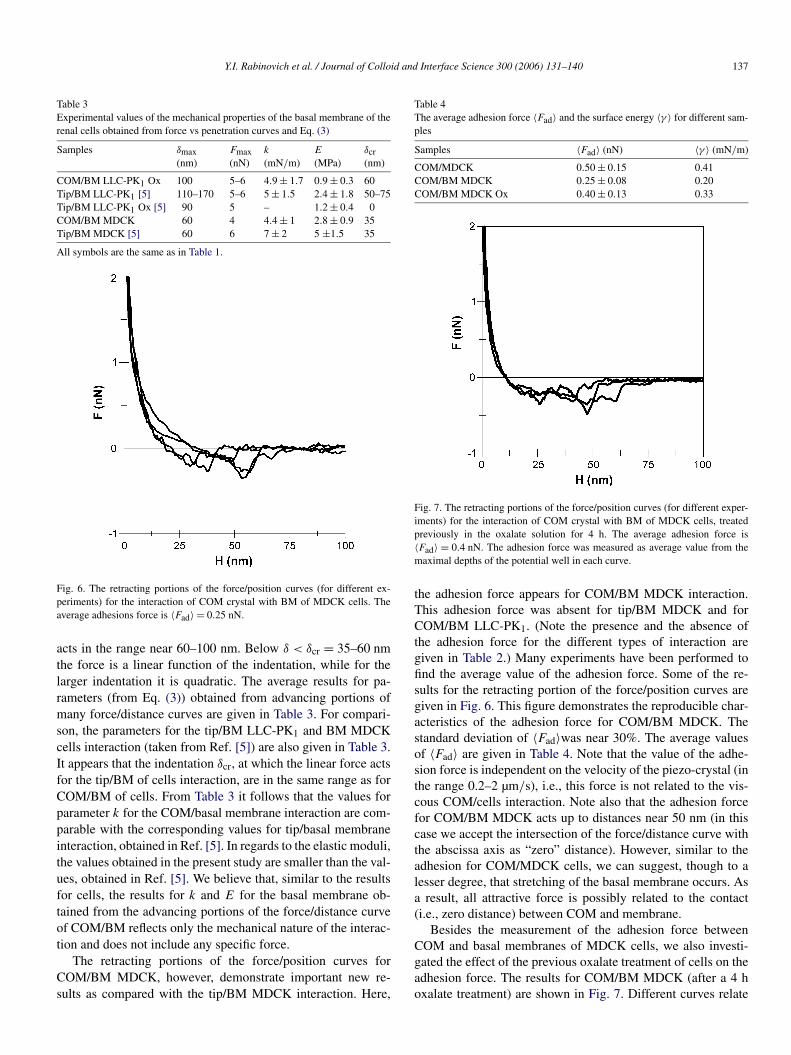
Fig. 5. The approaching and retracting portions of the force/position curveobtained for the interaction of basal membrane (BM) of MDCK cells withCOM crystal. Theoretical equation (3) (solid line) includes the following fit-ting parameters: k = 4.5 mN/m, E = 2.4 MPa, δmax = 60 nm, δcr = 35 nm.No dependence of the probe rate has been found.
for 4 h) and COM/BM MDCK, respectively. As also observedin Ref. [5], it follows from Fig. 4 that the treatment of the cellsin the potassium oxalate solution for 4 h results usually in thecells death. As a consequence, only basal membrane with thethickness near 100 nm remained on the substrate (while thick-ness of the cells before treatment was several microns). Similarto the COM/cell interaction, there is only small hysteresis be-tween extending and receding portions of curves. As it has beenshown, measurements at different probe approach velocitiesV = 0.4–2 µm/s result in curves that are practically indepen-dent on the approach rate. Both of these facts (small hysteresisand independence of force on the probe velocity) prove the neg-ligible role of viscosity in these interactions. The repulsive force
Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140 137
Table 3Experimental values of the mechanical properties of the basal membrane of therenal cells obtained from force vs penetration curves and Eq. (3)
Samples δmax(nm)
Fmax(nN)
k
(mN/m)E
(MPa)δcr(nm)
COM/BM LLC-PK1 Ox 100 5–6 4.9 ± 1.7 0.9 ± 0.3 60Tip/BM LLC-PK1 [5] 110–170 5–6 5 ± 1.5 2.4 ± 1.8 50–75Tip/BM LLC-PK1 Ox [5] 90 5 – 1.2 ± 0.4 0COM/BM MDCK 60 4 4.4 ± 1 2.8 ± 0.9 35Tip/BM MDCK [5] 60 6 7 ± 2 5 ±1.5 35
All symbols are the same as in Table 1.
Fig. 6. The retracting portions of the force/position curves (for different ex-periments) for the interaction of COM crystal with BM of MDCK cells. Theaverage adhesions force is 〈Fad〉 = 0.25 nN.
acts in the range near 60–100 nm. Below δ < δcr = 35–60 nmthe force is a linear function of the indentation, while for thelarger indentation it is quadratic. The average results for pa-rameters (from Eq. (3)) obtained from advancing portions ofmany force/distance curves are given in Table 3. For compari-son, the parameters for the tip/BM LLC-PK1 and BM MDCKcells interaction (taken from Ref. [5]) are also given in Table 3.It appears that the indentation δcr, at which the linear force actsfor the tip/BM of cells interaction, are in the same range as forCOM/BM of cells. From Table 3 it follows that the values forparameter k for the COM/basal membrane interaction are com-parable with the corresponding values for tip/basal membraneinteraction, obtained in Ref. [5]. In regards to the elastic moduli,the values obtained in the present study are smaller than the val-ues, obtained in Ref. [5]. We believe that, similar to the resultsfor cells, the results for k and E for the basal membrane ob-tained from the advancing portions of the force/distance curveof COM/BM reflects only the mechanical nature of the interac-tion and does not include any specific force.
The retracting portions of the force/position curves forCOM/BM MDCK, however, demonstrate important new re-sults as compared with the tip/BM MDCK interaction. Here,
Table 4The average adhesion force 〈Fad〉 and the surface energy 〈γ 〉 for different sam-ples
Samples 〈Fad〉 (nN) 〈γ 〉 (mN/m)
COM/MDCK 0.50 ± 0.15 0.41COM/BM MDCK 0.25 ± 0.08 0.20COM/BM MDCK Ox 0.40 ± 0.13 0.33
Fig. 7. The retracting portions of the force/position curves (for different exper-iments) for the interaction of COM crystal with BM of MDCK cells, treatedpreviously in the oxalate solution for 4 h. The average adhesion force is〈Fad〉 = 0.4 nN. The adhesion force was measured as average value from themaximal depths of the potential well in each curve.
the adhesion force appears for COM/BM MDCK interaction.This adhesion force was absent for tip/BM MDCK and forCOM/BM LLC-PK1. (Note the presence and the absence ofthe adhesion force for the different types of interaction aregiven in Table 2.) Many experiments have been performed tofind the average value of the adhesion force. Some of the re-sults for the retracting portion of the force/position curves aregiven in Fig. 6. This figure demonstrates the reproducible char-acteristics of the adhesion force for COM/BM MDCK. Thestandard deviation of 〈Fad〉was near 30%. The average valuesof 〈Fad〉 are given in Table 4. Note that the value of the adhe-sion force is independent on the velocity of the piezo-crystal (inthe range 0.2–2 µm/s), i.e., this force is not related to the vis-cous COM/cells interaction. Note also that the adhesion forcefor COM/BM MDCK acts up to distances near 50 nm (in thiscase we accept the intersection of the force/distance curve withthe abscissa axis as “zero” distance). However, similar to theadhesion for COM/MDCK cells, we can suggest, though to alesser degree, that stretching of the basal membrane occurs. Asa result, all attractive force is possibly related to the contact(i.e., zero distance) between COM and membrane.
Besides the measurement of the adhesion force betweenCOM and basal membranes of MDCK cells, we also investi-gated the effect of the previous oxalate treatment of cells on theadhesion force. The results for COM/BM MDCK (after a 4 hoxalate treatment) are shown in Fig. 7. Different curves relate

138 Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140
to the different experiments. It appears that the adhesion forceexists in this case.
Note that the saw-like character of the adhesion in Figs. 6and 7 indicates possibly a rupture of the multiple contacts dur-ing separation of the COM/cell surfaces. Similar force/distancebehavior has also been observed in Ref. [32] for attractive forcebetween hydrophobic surfaces under water where it was at-tributed to the effect of the micro-bubbles between samples.
The average values of the adhesion force for COM/MDCKand COM/BM MDCK are given in Table 4. The detail analysisof the different possible interaction forces in the biological sys-tems is given in the review [33]. Below we consider the possibleeffect of some of these forces.
It is difficult to compare the experimental values of the adhe-sion force with the possible theoretical value of van der Waalsforce, because the cell surface is covered with macromoleculesof different molecular weight and chemical structure. However,we believe that attractive van der Waals force is negligible inthis case due to the steric repulsion of the proteins chains,preventing direct contact between the solid COM crystal andplasma membrane of the cells. The protein chains form loopsand not the molecularly planar surface. As a result, van derWaals force between these chains and COM is significantlysmaller than between the smooth solid (nonporous) bodies. Thishypothesis is confirmed by the absence of the adhesion forcebetween COM and LLC-PK1 cells, while van der Waals forceshould exist for any line of the cells in the case of the direct con-tact of plasma membrane with COM. Additional confirmationof the negligible role of van der Waals force in the consideredcase is taken to be the absence of the adhesion between tip/cellsand tip/basal membrane of the cells.
The theoretical value of ion-electrostatic interaction is dif-ficult to calculate as it depends on a number of chemical pa-rameters, which characterize the behavior of the electric chargeand potential. However, the long-range ion-electrostatic (doublelayer) force should be completely screened by the electrolytewith a concentration near 0.25 M [30]. On the other hand, thedirect contact between the oppositely charged phospho-lipidhead groups on the surface of the plasma membranes [34] and
Fig. 8. SEM-picture of the COM crystal top.
COM crystal can give rise to strong adhesion between cells andCOM.
To better characterize adhesive phenomena, we can calculatethe energy of adhesion using experimental values of the adhe-sion force. The surface energy, γ , is evaluated using Eq. (5)(i.e., suggesting the spherical shape of the edge of the top of theCOM crystal). To experimentally find the radius of the top ofthe COM crystals, which interacts with the cells, SEM imagesof the top of the COM crystal have been taken (Fig. 8). Theaverage radius of the top was found to be R = 130 ± 15 nm.Using this value of radius and the experimental values of theadhesion force (Table 4) in Eq. (5), the values of γ are found tolie in the range γ = 0.2–0.4 mN/m. These results are includedin Table 4.
Another possible method of the adhesion energy calcula-tion is using integral of the potential well on the force/distancecurve. However, this would give overestimated values, as itwould also include the energy of the deformation of the cellsand membrane.
Calculation of the energy per bond (or per mole) dependson the density of bonding sites. If the adhesion is determinedby H-bonds, we can suggest that, in turn, the density of thesebonds depends on the density of COO-groups on the surfaceof the COM crystals. Knowing the elementary unit area to benear 2 nm2 [35], we obtain the energy per bond to be equal to0.12, 0.06 and 0.10 kcal/mol for COM/MDCK cells, COM/BMMDCK, and COM/BM MDCK after oxalate treatment, respec-tively. Comparing these energies with the energy of the weakhydrogen bonds (1–5 kcal/mol), we find that only one of the25 COO− groups forms a hydrogen bond with the protein onthe MDCK cells surfaces. It is possible that the formation of H-bonds is determined by the proteins on the cells rather than bythe COM crystal. This hypothesis agrees qualitatively with theresults of Refs. [6,7], where the adhesion force between differ-ent faces of the COM crystal and the tip modified by COO- orNH2NH2-groups (and the same radius as in the present study[36]) was measured. In these reports the adhesion force (andcorresponding surface energy) was 2–5 times larger than in thepresent study. This can be explained by the different density ofthe interacting groups on the cell-surface (in the present paper)and on the modified tip (in the papers [6,7]).
In addition, besides the H-bonds, the formation of the ionicCa2 +–COO− bonds is also possible. The density of the dis-sociated ions COO− on the COM crystal was calculated fromζ potential of the COM crystal. ζ potential was determinedexperimentally in 2 mM electrolyte solution to be equal toζ = −14 mV [37]. From this value, the density of the ele-mentary charge, i.e., the density of dissociated COO− ions,was calculated to be 8.81 × 1015 m−2 = 1.46 × 10−8 mol/m2.From this value density and the specific adhesion energy forCOM/MDCK cells (Table 4), we obtain the energy per mole ofdissociated COO− ions equal to 13.4 kcal/mol. Knowing theenergy of this reaction to be near 100 kcal/mol [38], we canconclude only one from eight dissociated COO− ions on theCOM surface forms ionic bond with the cell.
One of the facts supporting the ideas about the role of H-and ionic-bonds in the COM/BM MDCK adhesion is that the
Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140 139
oxalate treatment of cells gives rise to an increase in Fad (Ta-ble 4). This increase can be explained by the oxalate-inducedmembrane disruption and the release of the organic cellularcomponents [39,40]. This will change the elastic properties bythe possible appearance of additional binding sites on the mem-brane. These sites can provide the ionic- and H-bonds withCOM or short-range (contact) electrical attraction (Coulombforce). The increase of the adhesion force after the oxalate treat-ment agrees well with the known fact that the probability ofthe kidney stone formation increases in the presence of oxalate[40,41]. However, kidney stone formation under urine solutionswith excess oxalate can be related not only to the increase ofthe adhesion force between COM and cells, but also to the in-creased COM crystallization in the presence of oxalate [18,19].Particularly, it was found in Ref. [42] that the epithelial injuryleads to increased crystal binding.
Therefore, the experimental values of the adhesion force canbe explained, in the first approximation, by the different typesof forces; namely, by H- and ionic-bonds. The adhesion forcehas been observed and measured for MDCK/COM interaction,while it was negligibly small for LLC-PK1/COM interaction.There was also no adhesion between the tip and renal epithe-lial cells. This difference shows that the adhesion force is notconnected with the mechanical properties of the membrane andshould be related to the certain specific short-range forces. Thehypothesis on the presence/absence of specific forces as the rea-son of the adhesion force can explain the existence of adhesionfor COM/MDCK and its absence for COM/LLC-PK1. This dif-ference can be related to the difference between the chemicalcomposition of two cell lines used and possibly to the longermicrovilli on the plasma membrane of the LLC-PK1 cells.
As mentioned in the introduction, only the large attractiveforce can provide the formation of the adhesion contacts be-tween COM crystals and renal epithelial cells (inside renaltubules). The further growth of these crystals leads to the kidneystone formation. Therefore, the AFM results for the adhesionforce predict the possible formation of the kidney stones on theMDCK cells and the stone absence on the LLC-PK1 cells. Thisphenomenon agrees with the known fact that the kidney stoneformation begins in collecting ducts rather than in the renaltubules. The conclusion about the important role of the adhe-sion between COM crystals and certain chemical groups in theprocess of the kidney stone formation agrees with similar con-clusion made in Ref. [36] on the crucial role of adhesion inthe pathological activity of CaOx hydrates in this process. Ofcourse, the difference in the kidney stone formation on the dif-ferent epithelial cells can be explained not only by the differentadhesion force, but also by the hydrodynamic reason. In addi-tion, the composition of urine is also different in the differentsections of the renal tubules [41].
6. Conclusions
The force between COM crystal and the kidney epithelialcells are directly measured by AFM in artificial urine. For LLC-PK1 only a repulsive force was found, while for MDCK theattractive force was measured on the retracting portion of the
force/position curve. The average adhesion force was near 0.5,0.25 and 0.4 nN for MDCK cells, for non-treated and oxalatetreated basal membranes of MDCK cells, respectively. Theoxalate treatment of cells yields an increase of the adhesionforce for MDCK cells. This is explained by disruption of thecell membrane and the corresponding appearance of bondingsites for calcium oxalate monohydrate as a result of the ox-alate treatment. No adhesion was found for the interaction ofMDCK cells with the silicon nitride tip. This proves the exis-tence of specific interactions, including ionic- and H-bonds, orshort range Coulomb interaction, on the adhesion force betweenCOM crystal and MDCK cells.
Acknowledgments
The authors acknowledge the financial support of the Na-tional Institute of Health (NIH Grant 4509-802-12), the Na-tional Science Foundation (NSF Grant EEC-94-02989), the Par-ticle Engineering Research Center (PERC) at the University ofFlorida, and the Industrial Partners of the PERC for supportof this research. The authors thank Dr. Daniel Talham for use-ful discussion and Miss Maryam El-Shall, Mr. Scott Brown,Dr. Suresh Yeruva and Mr. Stephen Tedeschi for editing thepaper. Any opinions, findings, and conclusions or recommen-dations expressed in this material are those of the author(s) anddo not necessarily reflect those of the NIH and/or the NSF.
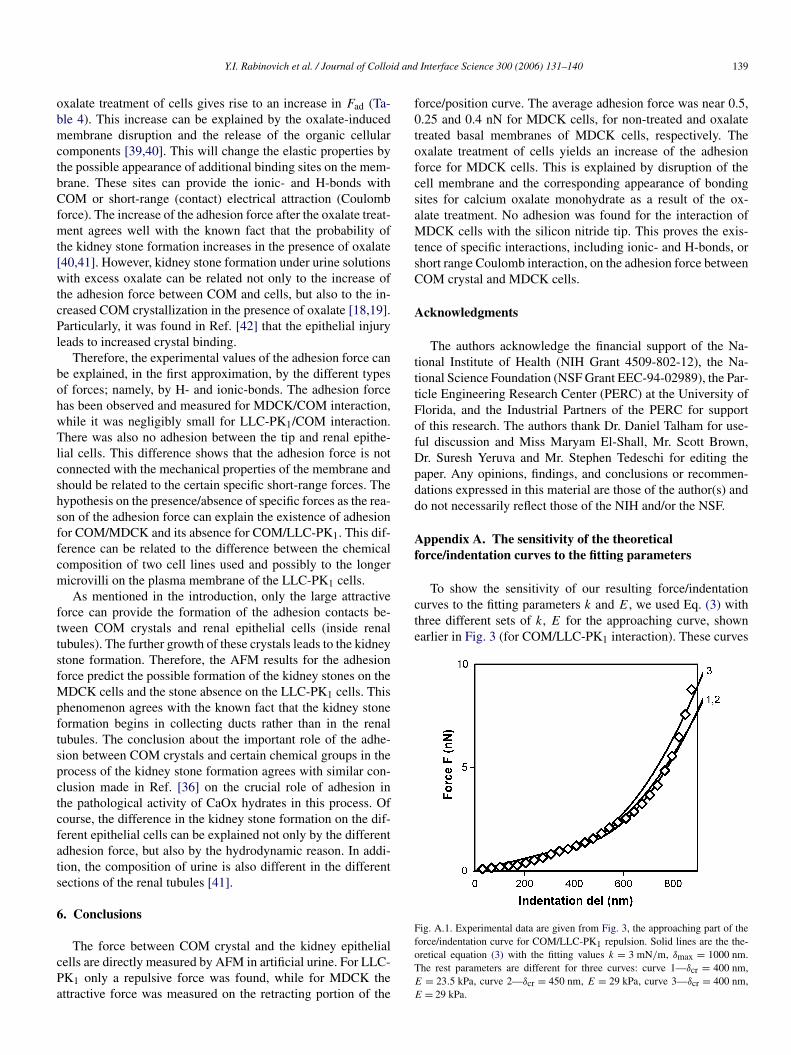
Appendix A. The sensitivity of the theoreticalforce/indentation curves to the fitting parameters
To show the sensitivity of our resulting force/indentationcurves to the fitting parameters k and E, we used Eq. (3) withthree different sets of k, E for the approaching curve, shownearlier in Fig. 3 (for COM/LLC-PK1 interaction). These curves
Fig. A.1. Experimental data are given from Fig. 3, the approaching part of theforce/indentation curve for COM/LLC-PK1 repulsion. Solid lines are the the-oretical equation (3) with the fitting values k = 3 mN/m, δmax = 1000 nm.The rest parameters are different for three curves: curve 1—δcr = 400 nm,E = 23.5 kPa, curve 2—δcr = 450 nm, E = 29 kPa, curve 3—δcr = 400 nm,E = 29 kPa.

140 Y.I. Rabinovich et al. / Journal of Colloid and Interface Science 300 (2006) 131–140
are shown in Fig. A.1. The sets of values of k and E are givenin figure legend. From curves 1 and 2 in Fig. A.1, it followsthat we can get different values of E (E = 23.5 and 29 kPa)changing the value of δcr from 400 to 450 nm. On the otherhand, using the certain value of δcr = 400 nm, the change ofE from 23.5 to 29 kPa (by 25%) (curve 3) gives rise to thelarge deviation of the theoretical curve obtained from experi-mental data. Therefore, despite large number of parameters inEq. (3), the analysis of experimental curves made in the presentpaper has sense and the values of parameters are meaning-ful.
References
[1] A.B. Mathur, A.M. Collinsworth, W.M. Reichert, W.E. Kraus, G.A.Truskey, J. Biomech. 34 (2001) 1545.
[2] W. Xu, P.J. Mulhern, B.L. Blackford, M.H. Jericho, M. Firtel, T.J. Bev-eridge, J. Bacteriol. (1996) 3106.
[3] A.E. Smith, K.E. Moxham, A.P. Middelberg, J. Chem. Eng. Sci. 55 (2000)2043.
[4] J.H. Hoh, C.A. Shoenenberger, J. Cell Sci. 107 (1994) 1105.[5] Y. Rabinovich, M. Esayanur, S. Daosukho, K. Byer, H. El-Shall, S. Khan,
J. Colloid Interface Sci. 285 (2005) 125.[6] X. Sheng, M.D. Ward, J.A. Wesson, J. Am. Chem. Soc. 125 (2003) 2854.[7] X. Sheng, T. Jung, J.A. Wesson, M.D. Ward, Proc. Natl. Acad. Sci.
USA 102 (2005) 267.[8] H.W. Wu, T. Kuhn, V.T. Moy, Scanning 20 (5) (1998) 389.[9] Y.F. Dufrene, C.J.P. Boonaert, H.C. van der Mei, et al., Ultrami-
croscopy 86 (1–2) (2001) 113.[10] G.A. Burks, S.B. Velegol, E. Paramonova, et al., Langmuir 19 (6) (2003)
2366.[11] Y.S. Lo, Y.J. Zhu, T.P. Beebe, Langmuir 17 (12) (2001) 3741.[12] T. Hugel, M. Seitz, Macromol. Rapid Comm. 22 (13) (2001) 989.[13] J. Wang, A.J. Bard, Anal. Chem. 73 (10) (2001) 2207.[14] E. Lesniewska, M.C. Giocondi, V. Vie, E. Finot, J.P. Goudonnet, C. Le
Grimellec, Kidney Int. Suppl. 65 (1998) S42.[15] E. Lesniewska, P.E. Milhiet, M.C. Giocondi, et al., Methods Cell Biol. 68
(2002) 51.
[16] J.A. Wesson, E.M. Worcester, J.H. Wiessner, N.S. Mandel, Kidney Int. 53(1998) 952.
[17] J.C. Lieske, G. Farell, S. Deganello, Urol. Res. 32 (2004) 117.[18] H. El-Shall, J. Jeon, E.A. Abdel-Aal, S. Khan, L. Gower, Y. Rabinovich,
Cryst. Res. Technol. 39 (2003) 212.[19] H. El-Shall, J. Jeon, E.A. Abdel-Aal, S. Khan, L. Gower, Y. Rabinovich,
Cryst. Res. Technol. 39 (2003) 222.[20] I.U. Vakarelski, A. Toritani, M. Nakayama, K. Higashitani, Langmuir 17
(2001) 4739, Langmuir 19 (2003) 110.[21] A. Touhami, B. Nyste, Y.F. Dufrene, Langmuir 19 (2003) 4539.[22] J.R. Burns, B. Finlayson, Invest. Urol. 18 (1980) 167.[23] C.C. Tisher, K.M. Madsen, in: F.C. Rector (Ed.), The Kidney, Saunders,
Philadelphia, 1986, pp. 3–60.[24] E. Koushanpour, Renal Physiology: Principles and Functions, Saunders,
Philadelphia, 1976.[25] F.C.J. Rector, in: R.W. Berliner (Ed.), Handbook of Physiology, American
Physiological Society, Washington, DC, 1983, pp. 431–454.[26] A.L. Weisenhorn, M. Khorsandi, S. Kasas, V. Gotzos, H.J. Butt, Nanotech-
nology 4 (1993) 106.[27] K.D. Costa, F.C.P. Yin, J. Biomech. Eng. 121 (1999) 462.[28] V.V. Lulevich, D. Andrienko, O.I. Vinogradova, J. Chem. Phys. 120 (2004)
3822.[29] V.V. Lulevich, I.L. Radtchenko, G.B. Sukhorukov, O.I. Vinogradova,
J. Phys. Chem. B 107 (2003) 2735.[30] B.V. Derjaguin, N.V. Churaev, V.M. Muller, Surface Force, Nauka,
Moscow, 1985, 400 pp.[31] K.L. Johnson, K. Kendall, A.D. Roberts, Proc. R. Soc. London A 324
(1971) 301.[32] J. Parker, P.M. Claesson, P. Attard, J. Phys. Chem. 98 (1994) 8468.[33] D. Leckband, J.N. Israelachvili, Quart. Rev. Biophys. 34 (2001) 105.[34] S.R. Khan, P.A. Glenton, R. Backov, D.R. Talham, Kidney Int. 62 (2002)
2062.[35] V. Tazzoli, C. Domeneghetti, Am. Mineral. 65 (1980) 327.[36] X. Sheng, M.D. Ward, F.A. Wesson, J. Am. Soc. Nephrol. 16 (2005) 1904.[37] K.G. Christmas, Ph.D. Thesis, University of Florida, 2001.[38] http://www.webelements.com/webelements/elements/text/Ca/enth.html.[39] S.R. Khan, Nephron Exp. Nephrol. 98 (2004) 55.[40] S.R. Khan, Clin. Exp. Nephrol. 8 (2004) 75.[41] J.M. Fasano, S.R. Khan, Kidney Int. 59 (2001) 169.[42] C.F. Verkoelen, B.G. van der Boom, A.B. Houtsmuller, F.H. Shroeder, J.C.
Romijn, Am. J. Physiol. Renal Physiol. 274 (1998) F958.
Related Documents











