of September 3, 2022. This information is current as Adaptive Immunity Natural Antibodies Bridge Innate and Saswati Panda and Jeak L. Ding http://www.jimmunol.org/content/194/1/13 doi: 10.4049/jimmunol.1400844 2015; 194:13-20; ; J Immunol References http://www.jimmunol.org/content/194/1/13.full#ref-list-1 , 47 of which you can access for free at: cites 124 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2014 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on September 3, 2022 http://www.jimmunol.org/ Downloaded from by guest on September 3, 2022 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of September 3, 2022.This information is current as
Adaptive ImmunityNatural Antibodies Bridge Innate and
Saswati Panda and Jeak L. Ding
http://www.jimmunol.org/content/194/1/13doi: 10.4049/jimmunol.1400844
2015; 194:13-20; ;J Immunol
Referenceshttp://www.jimmunol.org/content/194/1/13.full#ref-list-1
, 47 of which you can access for free at: cites 124 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2014 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

Natural Antibodies Bridge Innate and Adaptive ImmunitySaswati Panda1 and Jeak L. Ding
Natural Abs, belonging to isotypes IgM, IgG3, andIgA, were discovered nearly half a century ago. Despiteknowledge about the role of the polyreactive naturalIgM in pathogen elimination, B cell survival and ho-meostasis, inflammatory diseases, and autoimmunity,there is a lack of clarity about the physiological roleof natural IgG and natural IgA because they appearincapable of recognizing Ags on their own and areperceived as nonreactive. However, recent research re-vealed exciting functions of natural IgG in innate im-munity. Natural IgG:lectin collaboration swiftly andeffectively kills invading pathogens. These advancesprompt further examination of natural Abs in immunedefense and homeostasis, with the potential for devel-oping novel therapeutics. This review provides newinsights into the interaction between natural Abs andlectins, with implications on how interactions betweenmolecules of the innate and adaptive immune systemsbridge these two arms of immunity. The Journal ofImmunology, 2015, 194: 13–20.
Antibodies belong to the Ig superfamily. Ag-specificAbs are secreted by plasma cells in response to anAg (1). During the primary innate immune re-
sponse, APCs, such as dendritic cells (2, 3) and macrophages,recognize the pathogen through an array of pattern recogni-tion receptors (PRRs; e.g., TLRs) (4, 5) and present theprocessed Ag to B cells. These Ag-stimulated B cells undergosomatic hypermutation (6, 7) and clonal selection to becomelong-lived plasma cells that produce Ag-specific Abs in thesecondary adaptive immune response (8).Five isotypes of Ig exist (IgM, IgG, IgA, IgE, IgD) that differ
based on the H chain C region (9). IgM is the first isotypeproduced prior to class switching (10); it effectively recognizesand eliminates pathogens in the early stage of immune de-fense (11). Upon Ag stimulation, the B cells undergo classswitching (12, 13) and convert into plasma cells that producehigh-affinity IgG, IgA, IgE, and IgD. IgG is the major serumisotype and consists of four subtypes (IgG1, IgG2, IgG3, andIgG4), with further differences in the hinge region of the Hchain constant fragment. IgG is the only isotype that can crossthe placental barrier to provide immunity to the fetus (14).
IgA, present at the mucosal surfaces, such as the gut, respira-tory tract, and urogenital tract, plays an essential role in mu-cosal immunity (15–17). In humans, some B cells are knownto generate IgM2IgD+ plasmablasts through both T cell–de-pendent and T cell–independent (TI) pathways in the sec-ondary lymphoid organs of the upper respiratory mucosa uponAg exposure. These plasma cells produce IgD that is highlyspecific to respiratory commensals and pathogens (18). IgDalso was shown to be polyreactive and may play a role inanergy through tolerogenic pathways. However, there arecontradictory reports indicating that IgD may also protectB cells from tolerance (19). Despite these interesting findings,the biological role of IgD is still enigmatic. Further studies arerequired to provide insights into its role in innate immunity.In general, Ag-specific Abs elicit immune defense through theactivation of the classical complement system (20–22),pathogen neutralization (23, 24), phagocytosis of apoptoticcells (25–27), and priming of immune cells (28, 29).In addition to Ag-specific Abs, there are natural Abs that are
produced after birth in neonates and in adults prior to aninfection (30, 31). Extensive studies have been performed inmice to shed light on natural Abs. Natural Abs are reportedlyproduced by B-1 cells (32–34) in both mice (35) and humans(36, 37). Although natural Abs have the ability to recognizecertain self-antigens through their V region, they have beenperceived to lack specificity for any particular foreign Ag.However, reports indicate that certain natural Abs in micehave the ability to recognize foreign Ags. The T(EPC)-15 orT-15 idiotype natural Ab specifically recognizes phosphor-ylcholine on the surface of Gram-negative and Gram-positivebacteria, protozoa, fungi, and even modified low-density li-poprotein (38). The T-15 idiotype Ab was shown to conferprotection in atherosclerosis, apoptosis, and immunity againstpathogens (39). Natural IgM broadly and nonspecificallyrecognizes diverse microbial determinants and autoantigens.The significance of natural Abs is becoming increasingly ap-parent through decades of research on the function of naturalIgM, which have shed light on its involvement in multiplebiological processes, including infection, B cell homeostasis,inflammation, atherosclerosis, and autoimmunity (40).In this review, we first discuss the findings on natural Abs,
focusing on the role of natural IgM. This is followed by anexposition of the recent findings on natural IgG, which
Department of Biological Sciences, National University of Singapore, Singapore 117543
1Current address: Program in Molecular Pathogenesis, Department of Medicine, Skir-ball Institute of Biomolecular Medicine, New York University School of Medicine,New York, NY.
Received for publication April 7, 2014. Accepted for publication September 11, 2014.
This work was supported by the Ministry of Education, Singapore (Grants Tier 1,R-154-000-584-112, and MOE2013-T2-2-007).
Address correspondence and reprint requests to Dr. Jeak L. Ding, Department of Bio-logical Sciences, National University of Singapore, 14 Science Drive 4, Singapore117543. E-mail address: [email protected]
Abbreviations used in this article: CRP, C-reactive protein; DAMP, danger-associatedmolecular pattern; MBL, mannose-binding lectin; MZ, marginal zone; PRR, patternrecognition receptor; SLE, systemic lupus erythematosus; TI, T cell–independent.
Copyright� 2014 by TheAmerican Association of Immunologists, Inc. 0022-1767/14/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1400844
Journal ofTh e
ImmunologyBrief Reviews
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

collaborates with serum lectins, and how this immunecomplex links innate immunity to adaptive immunity. Wediscuss how the prevailing microenvironmental changes in-duced by infection–inflammation conditions might regulatethe antimicrobial action elicited by the natural IgG in hu-mans, although these results are restricted to in vitro and exvivo studies using human serum and are supported by in vivomice studies. Last, we provide a current opinion on the im-pending questions that lie ahead in this field and suggest fu-ture work that is needed to explore and exploit the role ofnatural Abs in health and disease.
Natural Abs: B cell subsets and their contribution
In addition to Ag-specific Abs that are produced by B-2 cellsduring the adaptive immune response (41), a pool of spon-taneously occurring Abs is present in human cord blood (33),in normal healthy individuals (30, 31, 42), and in germ-free/Ag-free mice (43–46) that belongs to the IgM, IgG, and IgAisotypes. In mice, natural IgG typically belongs to the IgG3subclass. Although a study suggested that humans also pro-duce IgG3 subclass-specific natural IgG Abs (47), the subclassspecification of human natural IgG has not been confirmed.Mouse B-1 cells produce natural Abs in a TI pathway (48) onexposure to self-antigens, such as dsDNA (49) and nucleo-somes (50, 51), and TI Ags (52). B-1 cells are generatedpredominantly during fetal and neonatal development fromprecursor cells located in the liver and omentum (32); how-ever, it is unclear whether natural Abs exist before birth. Inadults, the B-1 cell population is maintained at a constant sizeas a result of its self-renewing capacity, thus sustaining thelevels of serum natural Abs (53–56).The B-1 Ig genes tend to have preferential usage of the JH-
proximal VH gene segments during V(D)J recombination,with fewer N-region insertions (32) and a lower rate of so-matic mutations (57) compared with the B-2 counterpart,thus giving rise to a restricted natural Ab repertoire. Like theirmurine counterparts, human peripheral blood B-1 cells pro-duce polyreactive Abs toward a limited range of self-antigens(57). However, it is unclear to what extent human B-1 cellsresemble mouse B-1 cells because of the lack of identifiablecell surface markers on human B-1 cells. Interestingly, B-1cells that produce natural IgM from unmutated or minimallymutated V(D)J genes can enter the follicular pathway toundergo somatic hypermutation and class switching to pro-duce higher-affinity natural IgG or IgA autoantibodies (42),particularly under autoimmune conditions, such as systemiclupus erythematosus (SLE) (58) and rheumatoid arthritis(59). In addition to B-1 cells, natural Abs are produced bya subset of B-2 cells, which may include marginal zone (MZ)B cells (60), transitional B cells (61), and/or an unidentifiedpopulation of B cells (62). However, there is a lack of in-formation on the origin and type of signals that stimulatenatural Ab production by these B-2 cell subsets. B-1 cellsrespond to certain altered-self components or danger-as-sociated molecular patterns (DAMPs), such as asialylatedglycoproteins with exposed terminal galactose residues. Thehuman anti-gal a 1-3 gal Ab is one such abundant naturalIgG that is a xenoreactive natural Ab (63). Other examplesinclude natural IgG Abs with specificities toward gal a 1-2gal, gal a 1-4 gal, and gal a 1-6 glc (melibiose), which are notxenoreactive (64). Isohemagglutinins, such as anti-A and anti-
B blood group Abs, are well-known examples of natural IgMAbs. These Abs play a critical role in blood transfusion,transplantation, and graft rejection through complement ac-tivation (65).Although earlier studies suggested that peritoneal B-1 cells
are the main source of natural Abs (66), B-1 cells in the bonemarrow (67) and splenic MZ B cells (52, 68–70) also wereshown to produce natural Abs. The bone marrow B cellprecursors, which are generally considered to be the progen-itors of conventional (B-2) B cells, also produce B-1 cells bothin the fetus (71, 72) and adult (56). Extensive research effortshave shed light on the developmental origin of B-1 cells. It isnow increasingly evident that B-1 cells arise at different timesof development from a distinct population of progenitor cellsaccording to the layered immune system model (reviewed inRef. 73). Further studies in this area should provide moreinsight into understanding B cell immunity and relatedpathological issues.
Functions of natural IgM in mice: hints on linkage between innate andadaptive immunity
The contribution of natural Abs to immunity has been broughtinto focus by studies on natural IgM. Pentameric natural IgM,with 10 potential Ag-binding sites (74), recognizes multiplephylogenetically conserved structures like nucleic acids,phospholipids, and carbohydrates (40). Natural IgM also mayexist in the monomeric form in patients suffering from au-toimmunity (75) and chronic liver disease (76). A protectiverole for natural IgM was found in numerous viral, bacterial,fungal, and parasitic infections (77). For example, vesicularstomatitis virus, lymphocytic choriomeningitis virus, influ-enza virus, Streptococcus pneumoniae, and Cryptococcus neo-formans (78, 79) are bound, neutralized, and cleared bynatural IgM (80–83). Natural IgM also alerts and primes theadaptive immune system against subsequent pathogen attackby directing Ags to the secondary lymphoid organs. Micedeficient in natural IgM succumb to infections as a result ofa lack of pathogen clearance, decreased neutrophil recruit-ment, and elevated proinflammatory serum cytokines (84).The level of natural IgM is unaffected by foreign Ags (45),ascertaining the innate immune character of natural IgM.Under the innate immunity arm, natural IgM limits the
spread of infection by using strategies like neutralization (80,81, 83), complement-mediated immune complex formationand pathogen elimination by complement activation (83, 85),and recruitment of Ags into secondary lymphoid organs, thuspriming the subsequent adaptive immune response (81, 86)and linking the innate and adaptive immune systems. NaturalIgM also plays a role in clearing apoptotic cells (87), main-taining B cell homeostasis (88), inflammation (89, 90), ath-erosclerosis (91), and autoimmunity (92, 93). Natural IgMwas proposed to bind strongly to complement factor C1q andto activate the complement cascade (85, 94). Natural IgMalso binds mannose-binding lectin (MBL), which is bound toapoptotic cells (95, 96), and directs the clearance of immunecomplexes by binding to the putative Fca/mR receptor onphagocytes. Conversely, natural IgM bound to apoptotic cellsmay recruit MBL, and the immune complex may be clearedby the phagocytic calreticulin receptor (89). Fig. 1 summa-rizes the diverse roles of natural IgM identified through studiesin mice.
14 BRIEF REVIEWS: NATURAL Abs BRIDGE INNATE AND ADAPTIVE IMMUNITY
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

Natural IgG is not quiescent: it collaborates with serum lectins infrontline immune defense
Despite vast knowledge about natural IgM, the existence andphysiological function of natural IgG and IgA, which arepredominant serum and mucosal natural Abs (32), are unclearand, hence, have been a subject of interest since their dis-covery almost 50 y ago (97). Because of their low affinity,nonspecificity, and low valency, natural IgG and natural IgAappeared to be unreactive and were deemed incapable ofrecognizing and eliminating pathogens.Recent studies exploring the untapped potential of these
natural Ab isotypes demonstrated that both mice and humannatural IgG is not nonreactive, rather it plays a proactivefundamental role in the systemic innate immune response (98).In vitro studies revealed that natural IgG purified fromuninfected/healthy human serum recognized a range ofGram-negative and Gram-positive bacteria with the aid ofserum lectin PRRs (e.g., ficolin and MBL), which are knownbind to sugar residues (e.g., N-acetylglucosamine) on themicrobes. The partnership between natural IgG and lectins(prebound on the microbe) efficiently drove phagocytosis ofthe bacteria via the FcgR1 receptor on human monocytes.The importance of natural IgG during infection was dem-onstrated further in vivo in AID2/2 mice (these mice lackclass switching and produce IgM but not other subclasses ofAbs). The susceptibility of AID2/2 mice to infection wasovercome by i.v. administration of purified natural IgG,which was shown to collaborate with the serum lectins toelicit effective antimicrobial activity (98). Future studies usingficolin and MBL double knockout mice and FcgR12/2 micemight provide insight into the collaboration between naturalIgG and lectins at the crossroad of the innate–adaptive im-mune defense. Fig. 2 summarizes the natural IgG–mediatedbacterial recognition and clearance in collaboration with se-rum lectins and highlights the gray areas that warrant furtherexploration. These efforts will pave the way for developingnatural IgG–mediated therapies.
Infection–inflammation condition triggers the interaction betweennatural IgG:ficolin and enhances the immune response
Infection or injury causes an influx of inflammatory cells tothe site, leading to inflammation (99, 100). Inflammation ischaracterized by a drop in the pH (101–103) and calcium(104–106) levels in the microenvironment of infection/injury.In vitro and ex vivo studies with human samples, usinga simulated uninfected normal condition (typically, pH 7.4and 2.5 mM calcium) and an infection-inflammation con-dition (pH 6.5 and 2 mM calcium) (98, 107) explored var-ious PRR:PRR interactions under infection-induced changesin the microenvironment. The infection–inflammation con-dition increases PRR:PRR interaction affinity and boosts theimmune responses. Examples include the C-reactive protein(CRP):ficolin interaction, which synergistically enhances theclassical and lectin complement pathways (107), and thenatural IgG:ficolin interaction, which facilitates bacterialphagocytosis (98, 108). These studies demonstrate how per-turbations in the serum that result in mild local acidosis andhypocalcaemia change the conformation of PRRs, exposingnovel interaction motifs between them and leading to moreeffective pathogen binding and clearance. The identificationof the amino acids involved in the molecular interplay be-
tween natural IgG:ficolin (109) showed that infection–in-flammation–induced lower pH regulates the intermolecularelectrostatic interaction. By hydrogen-deuterium exchangemass spectrometry, Panda et al. (109) delineated the bindinginterfaces of the CH2–CH3 region of natural IgG Fc and theP-subdomain of the ficolin FBG domain. Notably, histidine iscritical for the molecular interaction. Mild acidosis at the siteof infection–inflammation probably resulted in protonationof histidine side chains that increased the interaction affinityoverall (109). These studies also imply that the immunesystem exploits the prevailing physicochemical changes at theinfection site for effective immune defense.The immunomodulatory activities resulting from the pH
change ensure a specific immune response rather than a ran-dom PRR:PRR interaction, which could lead to immuneoveractivation that manifests as autoimmune diseases. Inter-estingly, formation of the natural IgG:ficolin immune com-plex is similar in humans and mice. During infection, thehuman and mouse ficolins bind to the pathogen and recruitnatural IgG to form an immune complex. Conservation of thisphenomenon suggests the fundamental significance of naturalIgG:ficolin-mediated innate immune defense.Natural IgG specifically collaborates with pathogen-
associated lectins to elicit an effective antimicrobial actionagainst opportunistic Pseudomonas aeruginosa and Staphylo-coccus aureus (98). The immune evasiveness of such pathogensis a significant barrier to their clearance and resolution of theinfection and inflammation; hence, a continuing effort todevise new and more effective therapies is needed to combatsuch infections. Because natural IgG (specifically IgG3 iso-type) interacts with pathogen-associated ficolins through itsconserved CH2–CH3 region in the constant Fc region, it isplausible that IgG3 derived from plasma cells in bone marrowinteracts with ficolins in a similar manner once the ficolinshave bound to the pathogen. Further studies are required toinvestigate this hypothesis. Nevertheless, new insights into themechanism of interaction between the innate and adaptiveimmune proteins should advance our understanding of howthe host might counter the immune evasiveness of thesepathogens.
Natural IgG bridges innate and adaptive immunity and boostsimmune response
The mechanisms underlying the ligand specificity, signalingpathways, and cellular trafficking of PRR immune complexeshave been characterized extensively. Interestingly, multiplepathogen-associated molecular patterns that simultaneouslyactivate numerous PRRs (4, 110) were shown to inducePRR:PRR interactions as a prerequisite for effective immuneresponses. For example, CRP and ficolins, which are ini-tiators of the classical and lectin complement pathways, re-spectively, interact with each other to boost the immuneresponse against pathogens (107). Second, TLRs in collab-oration with other PRRs orchestrate both pathogen- and celltype–specific host responses to fight infections (111–113).Third, interactions between C1q and CRP were observedduring the immune response (114). Last, several studiesshowed the Ab-like behavior of pentraxins, which bind toFcgRs on macrophages and other immune cells to elicitimmune responses. For example, serum amyloid P, an acute-phase pentraxin, was shown to bind to FcgRIIA (115).
The Journal of Immunology 15
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
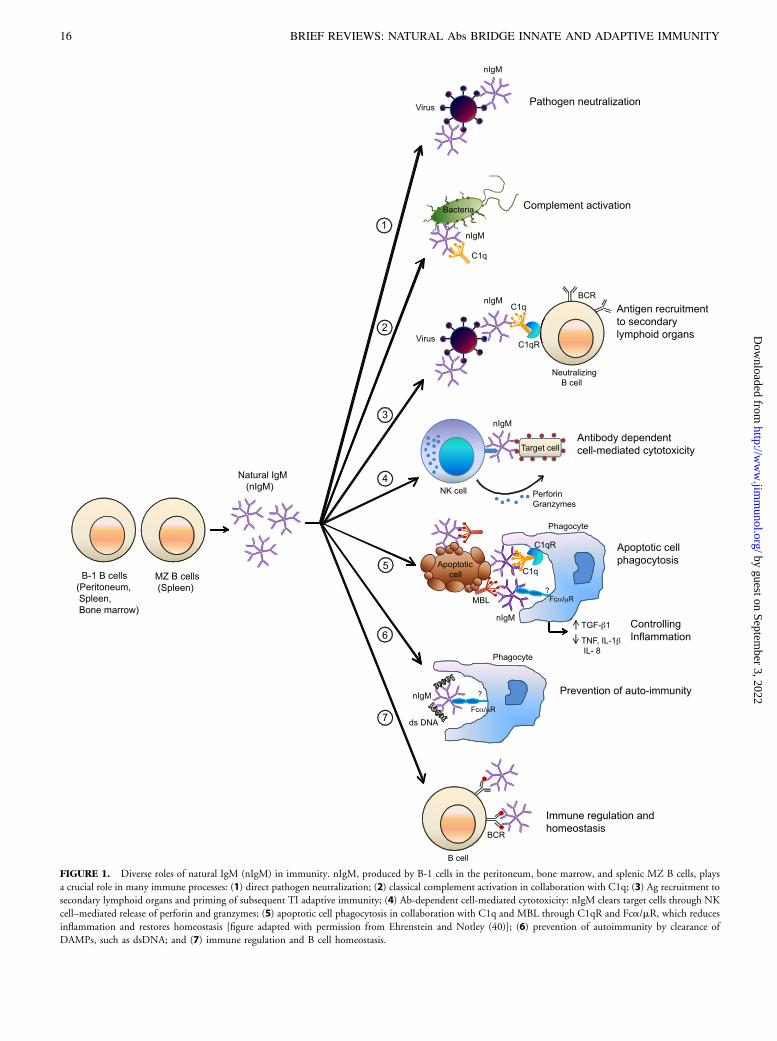
FIGURE 1. Diverse roles of natural IgM (nIgM) in immunity. nIgM, produced by B-1 cells in the peritoneum, bone marrow, and splenic MZ B cells, plays
a crucial role in many immune processes: (1) direct pathogen neutralization; (2) classical complement activation in collaboration with C1q; (3) Ag recruitment to
secondary lymphoid organs and priming of subsequent TI adaptive immunity; (4) Ab-dependent cell-mediated cytotoxicity: nIgM clears target cells through NK
cell–mediated release of perforin and granzymes; (5) apoptotic cell phagocytosis in collaboration with C1q and MBL through C1qR and Fca/mR, which reduces
inflammation and restores homeostasis [figure adapted with permission from Ehrenstein and Notley (40)]; (6) prevention of autoimmunity by clearance of
DAMPs, such as dsDNA; and (7) immune regulation and B cell homeostasis.
16 BRIEF REVIEWS: NATURAL Abs BRIDGE INNATE AND ADAPTIVE IMMUNITY
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
Another pentraxin, PTX3, binds to FcgRIII (116), sug-gesting its potential role in neutrophil and NK cell activa-tion. These PRR:PRR interaction–mediated mechanisms inducestronger and rapid immune responses that prime and shapeadaptive immunity.Natural IgG was recently shown to interact with pathogen-
associated lectins, such as ficolin and MBL, and to formimmune complexes to clear the pathogens (98, 108). Axi-omatically, Ag-specific IgG that binds directly to pathogens isalso expected to interact with other PRRs, such as C1q (117),to activate the classical complement pathway. Notably, boththe Ag-specific IgG (117) and natural IgG interact with PRRsthrough their H chain C region (109), indicating that the Fcis conserved in relaying the host defense information down-stream, irrespective of the origin and specificity of the IgGAbs. Identification of the interaction domains involved in theformation of immune complexes between natural IgG:ficolinand between Ag-specific IgG:C1q indicated that differentPRRs (ficolins and C1q, in these instances) interact withunique regions of the IgG Fc (98, 117). These findings evokecuriosity about why such subtle differences exist and how theymay fine-tune the immune response. The interaction betweennatural IgG:lectins corroborates the direct functional linkbetween the two arms of immunity, which boosts antimi-crobial response. Although this is crucial to prevent subver-
sion of evasive pathogens, immune overreaction is an Achilles’heel to autoimmunity and inflammatory diseases if left un-controlled.
Implications of natural Abs in autoimmunity and inflammatorydiseases
Natural IgM was reported to control autoimmunity and in-flammatory diseases. Mouse models of SLE have reduced levelsof natural IgM (118) with an increase in the B-1 cell subsetand an increase in IgG autoantibodies (92, 93). Decreasednatural IgM apparently makes these mice susceptible tonumerous infections. Natural IgM with C1q is known torecognize, bind, and clear autoantigens [e.g., oxidized low-density lipoprotein in atherosclerotic plaques (119, 120)and phospholipids and dsDNA expressed on apoptotic cells(87)]. Hence, a reduction in natural IgM levels is associatedwith ineffective clearance of apoptotic cells, resulting in au-toimmune diseases. Conversely, stronger and persistent rec-ognition of apoptotic cells by natural Abs may lead tooveractivation of the immune system and chronic inflamma-tion. Further studies on the extent of apoptotic cell uptake,immunomodulation by infection-induced microenvironmen-tal changes, and cytokine responses by natural Abs in auto-immune patients will provide clarity about the overall con-tribution of natural Abs in autoimmunity.
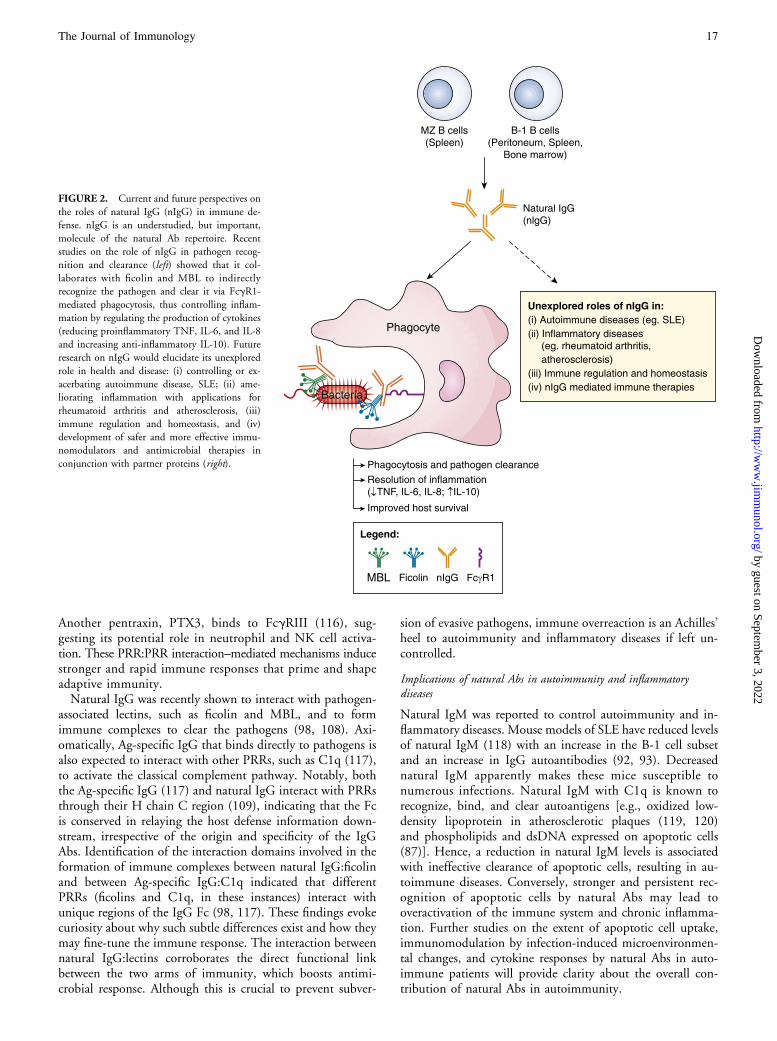
FIGURE 2. Current and future perspectives on
the roles of natural IgG (nIgG) in immune de-
fense. nIgG is an understudied, but important,
molecule of the natural Ab repertoire. Recent
studies on the role of nIgG in pathogen recog-
nition and clearance (left) showed that it col-
laborates with ficolin and MBL to indirectly
recognize the pathogen and clear it via FcgR1-
mediated phagocytosis, thus controlling inflam-
mation by regulating the production of cytokines
(reducing proinflammatory TNF, IL-6, and IL-8
and increasing anti-inflammatory IL-10). Future
research on nIgG would elucidate its unexplored
role in health and disease: (i) controlling or ex-
acerbating autoimmune disease, SLE; (ii) ame-
liorating inflammation with applications for
rheumatoid arthritis and atherosclerosis, (iii)
immune regulation and homeostasis, and (iv)
development of safer and more effective immu-
nomodulators and antimicrobial therapies in
conjunction with partner proteins (right).
The Journal of Immunology 17
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

How do natural Abs control inflammatory diseases? On onehand, natural IgM reduces the inflammatory response by re-moving the apoptotic cells and preventing inflammatory ar-thritis (121) and atherosclerosis (91). On the other hand,natural IgM enhances inflammation through increasing theprecipitation of urate crystals, which recruits neutrophils andworsens inflammation (122). Natural IgM is also involved inischemia–reperfusion injury, including myocardial infarctionand intestinal ischemia–reperfusion injury, wherein it targetsautoantigen-expressing endothelial cells (123, 124). It is likelythat these two opposing and parallel processes of the naturalIgM–mediated immune response maintain homeostasis. Thepotential role of natural IgG in controlling inflammation wasrecently uncovered based on the existence of the natural IgG:hemoglobin complex that is formed under hemolytic con-ditions: natural IgG facilitated uptake of the redox activehemoglobin by immune cells (125), leading to resolution ofinflammation and restoration of homeostasis. Recently, Hesset al. (126) also reported that Abs produced in a TI manner,similar to natural Abs, elicit anti-inflammatory properties.This is suggested to be attributable to differences in theirglycosylation patterns compared with conventional Abs,which lead to a proinflammatory response. Natural Abs ex-hibit a sialylated pattern, which confers an anti-inflammatorynature. In contrast, Ag-specific Abs are either agalactosylatedor asialylated and exhibit proinflammatory properties. Thisfinding has important implications in the immunoregulatorynature of Abs during a TI response to self-antigens in auto-immunity or inflammatory diseases initiated by TI Ags. Inaddition, this information could aid in vaccine design fordeveloping specific Abs with unique glycosylation patterns toprotect against infections without provoking excessive proin-flammatory responses.Uncontrolled/chronic infection may also result in over-
activation of the immune system, leading to autoimmunity(82). Hence, a timely and well-regulated immune response isnecessary to resolve infection and restore normalcy. Furtherwork is required to fully understand the mechanisms of actionof natural Abs in autoimmunity and chronic infection/inflammation. Moreover, knowledge of the role of naturalIgG and IgA in immunoregulation and homeostasis is still inits infancy, and much remains to be explored.
ConclusionsExtensive research has highlighted the role of natural IgM inhealth, infection, inflammation, and autoimmune diseases.Recent studies revealed the early protective role of natural IgGin infection, demonstrating that natural IgG:lectin interactionled to enhanced pathogen clearance (98). This collaborationbetween molecules of the innate and adaptive immune sys-tems blurs the boundaries between the seemingly discretearms of immunity and opens up a vast area to explore fun-damental immunology: what lies at and ahead of the cross-road of these immune pathways?Elucidation of the precise contact points between natural
IgG and ficolins (109) offers the potential for development ofeffective immunomodulatory drugs to intercept the immunecomplexes of natural IgG and lectins, to ameliorate immuneoverreaction, and to pre-empt chronic inflammatory diseases,such as rheumatoid arthritis, atherosclerosis, and SLE. Suchtherapies could also be tunable to the prevailing pH and
calcium conditions in the infection–inflammation microen-vironment to modulate the downstream immune response.To this end, various questions need to be addressed. What isthe spectrum of apoptotic cell DAMPs and autoantigensrecognized by the natural Ab:PRR immune complexes? Whatmechanism does natural IgG use to ameliorate inflammation?Is it through the removal of pathogens, apoptotic cell debris,and/or harmful DAMPs? Which immune cell type(s) doesnatural IgG regulate? What are the dynamics of the cytokineprofile during such a response? How can i.v. Ig, involvingnatural IgG and/or lectins, aid in controlling inflammation?How do natural IgG Abs regulate ongoing immune processesand specifically control autoimmunity-related inflammation?Is there a potential feedback/autoregulation of the productionof natural IgG? Some of these questions may be answeredwith in vivo studies on IgG-deficient or conditional knockoutmice to provide mechanistic insights.Ig infusions, consisting primarily of IgG, have been used to
treat inflammatory and autoimmune diseases, with limitedsuccess. Additionally, the finding that differential glycosylationpatterns in the Fc region of Abs determines the immune re-sponse (sialylated leading to anti-inflammatory response andagalactosylated or asialylated leading to proinflammatory re-sponse) (126) could aid in efficient vaccine design and i.v. Igtherapy to treat these inflammatory diseases. There are noreports on the use of natural IgG Abs in i.v. Ig therapy forrecurrent infections in patients with primary immune defi-ciencies. However, recent findings on the association betweennatural IgG and ficolins (lectins that bind to microbial sugarresidues) regulated by perturbations in pH and calciumconditions in the microenvironment (98) suggest the poten-tial use of i.v. Ig treatment for patients affected by primary Abdeficiencies. With a view to using the protective outcome ofnatural IgG:ficolin interaction in i.v. Ig therapy, it will beparticularly interesting to study the levels of natural IgG andlectins and the prevailing microenvironmental conditionsduring recurrent infections in these patients. It will also becrucial to take into account the role of other serum factorsthat may contribute through new and yet-to-be-exploredcompensatory mechanisms under such conditions. Thus, theexisting and exciting new findings in the field of natural Absprovide a better understanding of their role (in collaborationwith partner proteins) and extend the possibilities for devel-oping effective drugs to effectively counteract these diseases.
DisclosuresThe authors have no financial conflicts of interest.
References1. Burton, D. R., and J. M. Woof. 1992. Human antibody effector function. Adv.
Immunol. 51: 1–84.2. Banchereau, J., and R. M. Steinman. 1998. Dendritic cells and the control of
immunity. Nature 392: 245–252.3. Zitvogel, L. 2002. Dendritic and natural killer cells cooperate in the control/switch
of innate immunity. J. Exp. Med. 195: F9–F14.4. Janeway, C. A., Jr., and R. Medzhitov. 2002. Innate immune recognition. Annu.
Rev. Immunol. 20: 197–216.5. Dempsey, P. W., S. A. Vaidya, and G. Cheng. 2003. The art of war: Innate and
adaptive immune responses. Cell. Mol. Life Sci. 60: 2604–2621.6. Jacob, J., G. Kelsoe, K. Rajewsky, and U. Weiss. 1991. Intraclonal generation of
antibody mutants in germinal centres. Nature 354: 389–392.7. Berek, C., A. Berger, and M. Apel. 1991. Maturation of the immune response in
germinal centers. Cell 67: 1121–1129.8. Tarlinton, D. M., and K. G. Smith. 2000. Dissecting affinity maturation: a model
explaining selection of antibody-forming cells and memory B cells in the germinalcentre. Immunol. Today 21: 436–441.
18 BRIEF REVIEWS: NATURAL Abs BRIDGE INNATE AND ADAPTIVE IMMUNITY
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

9. Stavnezer, J., J. E. Guikema, and C. E. Schrader. 2008. Mechanism and regulationof class switch recombination. Annu. Rev. Immunol. 26: 261–292.
10. Fellah, J. S., M. V. Wiles, J. Charlemagne, and J. Schwager. 1992. Evolution ofvertebrate IgM: complete amino acid sequence of the constant region of Ambys-toma mexicanum mu chain deduced from cDNA sequence. Eur. J. Immunol. 22:2595–2601.
11. Boes, M. 2000. Role of natural and immune IgM antibodies in immune responses.Mol. Immunol. 37: 1141–1149.
12. Revy, P., T. Muto, Y. Levy, F. Geissmann, A. Plebani, O. Sanal, N. Catalan,M. Forveille, R. Dufourcq-Labelouse, A. Gennery, et al. 2000. Activation-inducedcytidine deaminase (AID) deficiency causes the autosomal recessive form of theHyper-IgM syndrome (HIGM2). Cell 102: 565–575.
13. Muramatsu, M., K. Kinoshita, S. Fagarasan, S. Yamada, Y. Shinkai, and T. Honjo.2000. Class switch recombination and hypermutation require activation-inducedcytidine deaminase (AID), a potential RNA editing enzyme. Cell 102: 553–563.
14. Schur, P. H. 1987. IgG subclasses—a review. Ann. Allergy 58: 89–96, 99.15. Woof, J. M., and M. A. Kerr. 2006. The function of immunoglobulin A in im-
munity. J. Pathol. 208: 270–282.16. Underdown, B. J., and J. M. Schiff. 1986. Immunoglobulin A: strategic defense
initiative at the mucosal surface. Annu. Rev. Immunol. 4: 389–417.17. Yan, H., M. E. Lamm, E. Bjorling, and Y. T. Huang. 2002. Multiple functions of
immunoglobulin A in mucosal defense against viruses: an in vitro measles virusmodel. J. Virol. 76: 10972–10979.
18. Chen, K., W. Xu, M. Wilson, B. He, N. W. Miller, E. Bengten, E. S. Edholm,P. A. Santini, P. Rath, A. Chiu, et al. 2009. Immunoglobulin D enhances immunesurveillance by activating antimicrobial, proinflammatory and B cell-stimulatingprograms in basophils. Nat. Immunol. 10: 889–898.
19. Chen, K., and A. Cerutti. 2010. New insights into the enigma of immunoglobulinD. Immunol. Rev. 237: 160–179.
20. Carroll, M. C., and M. B. Fischer. 1997. Complement and the immune response.Curr. Opin. Immunol. 9: 64–69.
21. Carroll, M. C. 1998. The role of complement and complement receptors in in-duction and regulation of immunity. Annu. Rev. Immunol. 16: 545–568.
22. Boross, P., and J. H. Leusen. 2012. Boosting antibody therapy with complement.Blood 119: 5945–5947.
23. Haigwood, N. L., and L. Stamatatos. 2003. Role of neutralizing antibodies in HIVinfection. AIDS 17(Suppl. 4): S67–S71.
24. Corti, D., and A. Lanzavecchia. 2013. Broadly neutralizing antiviral antibodies.Annu. Rev. Immunol. 31: 705–742.
25. Sutterwala, F. S., G. J. Noel, P. Salgame, and D. M. Mosser. 1998. Reversal ofproinflammatory responses by ligating the macrophage Fcgamma receptor type I.J. Exp. Med. 188: 217–222.
26. Kurosaka, K., N. Watanabe, and Y. Kobayashi. 2002. Potentiation by humanserum of anti-inflammatory cytokine production by human macrophages in re-sponse to apoptotic cells. J. Leukoc. Biol. 71: 950–956.
27. Hart, S. P., C. Jackson, L. M. Kremmel, M. S. McNeill, H. Jersmann,K. M. Alexander, J. A. Ross, and I. Dransfield. 2003. Specific binding of anantigen-antibody complex to apoptotic human neutrophils. Am. J. Pathol. 162:1011–1018.
28. Mota, G., M. Manciulea, E. Cosma, I. Popescu, M. Hirt, E. Jensen-Jarolim,A. Calugaru, C. Galatiuc, T. Regalia, D. Tamandl, et al. 2003. Human NK cellsexpress Fc receptors for IgA which mediate signal transduction and target cellkilling. Eur. J. Immunol. 33: 2197–2205.
29. Perussia, B. 1998. Fc receptors on natural killer cells. Curr. Top. Microbiol.Immunol. 230: 63–88.
30. Avrameas, S. 1991. Natural autoantibodies: from ‘horror autotoxicus’ to ‘gnothiseauton’. Immunol. Today 12: 154–159.
31. Coutinho, A., M. D. Kazatchkine, and S. Avrameas. 1995. Natural autoantibodies.Curr. Opin. Immunol. 7: 812–818.
32. Kantor, A. B., and L. A. Herzenberg. 1993. Origin of murine B cell lineages.Annu. Rev. Immunol. 11: 501–538.
33. Hardy, R. R., and K. Hayakawa. 1994. CD5 B cells, a fetal B cell lineage. Adv.Immunol. 55: 297–339.
34. Hardy, R. R., C. E. Carmack, Y. S. Li, and K. Hayakawa. 1994. Distinctive devel-opmental origins and specificities of murine CD5+ B cells. Immunol. Rev. 137: 91–118.
35. Conrad, K., M. P. Bachmann, E. Matsuura, and Y. Shoenfeld. 2005. From animalmodels to human genetics: research on the induction and pathogenicity of auto-antibodies. Autoimmun. Rev. 4: 178–187.
36. Harindranath, N., H. Ikematsu, A. L. Notkins, and P. Casali. 1993. Structure of theVH and VL segments of polyreactive and monoreactive human natural antibodies toHIV-1 and Escherichia coli beta-galactosidase. Int. Immunol. 5: 1523–1533.
37. Schettino, E. W., S. K. Chai, M. T. Kasaian, H. W. Schroeder, Jr., and P. Casali.1997. VHDJH gene sequences and antigen reactivity of monoclonal antibodiesproduced by human B-1 cells: evidence for somatic selection. J. Immunol. 158:2477–2489.
38. Kearney, J. F. 2000. Immune recognition of OxLDL in atherosclerosis. J. Clin.Invest. 105: 1683–1685.
39. Shaw, P. X., S. Horkko, M. K. Chang, L. K. Curtiss, W. Palinski, G. J. Silverman,and J. L. Witztum. 2000. Natural antibodies with the T15 idiotype may act inatherosclerosis, apoptotic clearance, and protective immunity. J. Clin. Invest. 105:1731–1740.
40. Ehrenstein, M. R., and C. A. Notley. 2010. The importance of natural IgM:scavenger, protector and regulator. Nat. Rev. Immunol. 10: 778–786.
41. Baumgarth, N., O. C. Herman, G. C. Jager, L. Brown, L. A. Herzenberg, andL. A. Herzenberg. 1999. Innate and acquired humoral immunities to influenzavirus are mediated by distinct arms of the immune system. Proc. Natl. Acad. Sci.USA 96: 2250–2255.
42. Casali, P., and E. W. Schettino. 1996. Structure and function of natural anti-bodies. Curr. Top. Microbiol. Immunol. 210: 167–179.
43. Sidman, C. L., L. D. Shultz, R. R. Hardy, K. Hayakawa, and L. A. Herzenberg.1986. Production of immunoglobulin isotypes by Ly-1+ B cells in viablemotheaten and normal mice. Science 232: 1423–1425.
44. Bos, N. A., H. Kimura, C. G. Meeuwsen, H. De Visser, M. P. Hazenberg,B. S. Wostmann, J. R. Pleasants, R. Benner, and D. M. Marcus. 1989. Serumimmunoglobulin levels and naturally occurring antibodies against carbohydrateantigens in germ-free BALB/c mice fed chemically defined ultrafiltered diet. Eur. J.Immunol. 19: 2335–2339.
45. Haury, M., A. Sundblad, A. Grandien, C. Barreau, A. Coutinho, and A. Nobrega.1997. The repertoire of serum IgM in normal mice is largely independent of ex-ternal antigenic contact. Eur. J. Immunol. 27: 1557–1563.
46. Hooijkaas, H., R. Benner, J. R. Pleasants, and B. S. Wostmann. 1984. Isotypes andspecificities of immunoglobulins produced by germ-free mice fed chemically de-fined ultrafiltered “antigen-free” diet. Eur. J. Immunol. 14: 1127–1130.
47. Okech, B. A., A. Nalunkuma, D. Okello, X. L. Pang, K. Suzue, J. Li, T. Horii, andT. G. Egwang. 2001. Natural human immunoglobulin G subclass responses toPlasmodium falciparum serine repeat antigen in Uganda. Am. J. Trop. Med. Hyg.65: 912–917.
48. Berland, R., and H. H. Wortis. 2002. Origins and functions of B-1 cells with noteson the role of CD5. Annu. Rev. Immunol. 20: 253–300.
49. Hirose, S., M. Wakiya, Y. Kawano-Nishi, J. Yi, R. Sanokawa, S. Taki,T. Shimamura, T. Kishimoto, H. Tsurui, H. Nishimura, et al. 1993. Somaticdiversification and affinity maturation of IgM and IgG anti-DNA antibodies inmurine lupus. Eur. J. Immunol. 23: 2813–2820.
50. Kantor, A. B., A. M. Stall, S. Adams, K. Watanabe, and L. A. Herzenberg. 1995.De novo development and self-replenishment of B cells. Int. Immunol. 7: 55–68.
51. Gioud, M., B. L. Kotzin, R. L. Rubin, F. G. Joslin, and E. M. Tan. 1983. In vivoand in vitro production of anti-histone antibodies in NZB/NZW mice.J. Immunol. 131: 269–274.
52. Martin, F., A. M. Oliver, and J. F. Kearney. 2001. Marginal zone and B1 B cellsunite in the early response against T-independent blood-borne particulate antigens.Immunity 14: 617–629.
53. Lalor, P. A., L. A. Herzenberg, S. Adams, and A. M. Stall. 1989. Feedback reg-ulation of murine Ly-1 B cell development. Eur. J. Immunol. 19: 507–513.
54. Deenen, G. J., and F. G. Kroese. 1992. Murine peritoneal Ly-1 B cells do not turnover rapidly. Ann. N. Y. Acad. Sci. 651: 70–71.
55. D€uber, S., M. Hafner, M. Krey, S. Lienenklaus, B. Roy, E. Hobeika, M. Reth,T. Buch, A. Waisman, K. Kretschmer, and S. Weiss. 2009. Induction of B-celldevelopment in adult mice reveals the ability of bone marrow to produce B-1acells. Blood 114: 4960–4967.
56. Holodick, N. E., K. Repetny, X. Zhong, and T. L. Rothstein. 2009. Adult BMgenerates CD5+ B1 cells containing abundant N-region additions. Eur. J.Immunol. 39: 2383–2394.
57. Griffin, D. O., N. E. Holodick, and T. L. Rothstein. 2011. Human B1 cells inumbilical cord and adult peripheral blood express the novel phenotype CD20+CD27+ CD43+ CD702. J. Exp. Med. 208: 67–80.
58. Kasaian, M. T., H. Ikematsu, J. E. Balow, and P. Casali. 1994. Structure of theVH and VL segments of monoreactive and polyreactive IgA autoantibodies toDNA in patients with systemic lupus erythematosus. J. Immunol. 152: 3137–3151.
59. Harindranath, N., I. S. Goldfarb, H. Ikematsu, S. E. Burastero, R. L. Wilder,A. L. Notkins, and P. Casali. 1991. Complete sequence of the genes encoding theVH and VL regions of low- and high-affinity monoclonal IgM and IgA1 rheu-matoid factors produced by CD5+ B cells from a rheumatoid arthritis patient. Int.Immunol. 3: 865–875.
60. Cerutti, A., M. Cols, and I. Puga. 2013. Marginal zone B cells: virtues of innate-like antibody-producing lymphocytes. Nat. Rev. Immunol. 13: 118–132.
61. Capolunghi, F., S. Cascioli, E. Giorda, M. M. Rosado, A. Plebani, C. Auriti,G. Seganti, R. Zuntini, S. Ferrari, M. Cagliuso, et al. 2008. CpG drives humantransitional B cells to terminal differentiation and production of natural antibodies.J. Immunol. 180: 800–808.
62. Descatoire, M., J. C. Weill, C. A. Reynaud, and S. Weller. 2011. A humanequivalent of mouse B-1 cells? J. Exp. Med. 208: 2563–2564.
63. Ohdan, H., K. G. Swenson, H. S. Kruger Gray, Y. G. Yang, Y. Xu, A. D. Thall,and M. Sykes. 2000. Mac-1-negative B-1b phenotype of natural antibody-producing cells, including those responding to Gal alpha 1,3Gal epitopes in al-pha 1,3-galactosyltransferase-deficient mice. J. Immunol. 165: 5518–5529.
64. Parker, W., S. S. Lin, P. B. Yu, A. Sood, Y. C. Nakamura, A. Song, M. L. Everett,and J. L. Platt. 1999. Naturally occurring anti-alpha-galactosyl antibodies: rela-tionship to xenoreactive anti-alpha-galactosyl antibodies. Glycobiology 9: 865–873.
65. Parker, W., K. Lundberg-Swanson, Z. E. Holzknecht, J. Lateef, S. A. Washburn,S. J. Braedehoeft, and J. L. Platt. 1996. Isohemagglutinins and xenoreactive antibodies:members of a distinct family of natural antibodies. Hum. Immunol. 45: 94–104.
66. Baumgarth, N., J. W. Tung, and L. A. Herzenberg. 2005. Inherent specificities innatural antibodies: a key to immune defense against pathogen invasion. SpringerSemin. Immunopathol. 26: 347–362.
67. Choi, Y. S., J. A. Dieter, K. Rothaeusler, Z. Luo, and N. Baumgarth. 2012. B-1 cells inthe bone marrow are a significant source of natural IgM. Eur. J. Immunol. 42: 120–129.
68. Martin, F., and J. F. Kearney. 2002. Marginal-zone B cells. Nat. Rev. Immunol. 2:323–335.
69. Kretschmer, K., A. Jungebloud, J. Stopkowicz, T. Kleinke, R. Hoffmann, andS. Weiss. 2003. The selection of marginal zone B cells differs from that of B-1acells. J. Immunol. 171: 6495–6501.
70. Carey, J. B., C. S. Moffatt-Blue, L. C. Watson, A. L. Gavin, and A. J. Feeney.2008. Repertoire-based selection into the marginal zone compartment duringB cell development. J. Exp. Med. 205: 2043–2052.
The Journal of Immunology 19
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

71. Dorshkind, K., and E. Montecino-Rodriguez. 2007. Fetal B-cell lymphopoiesisand the emergence of B-1-cell potential. Nat. Rev. Immunol. 7: 213–219.
72. Montecino-Rodriguez, E., H. Leathers, and K. Dorshkind. 2006. Identification ofa B-1 B cell-specified progenitor. Nat. Immunol. 7: 293–301.
73. Montecino-Rodriguez, E., and K. Dorshkind. 2012. B-1 B cell development in thefetus and adult. Immunity 36: 13–21.
74. Czajkowsky, D. M., and Z. Shao. 2009. The human IgM pentamer is a mushroom-shaped molecule with a flexural bias. Proc. Natl. Acad. Sci. USA 106: 14960–14965.
75. Bonagura, V. R., L. Mendez, N. Agostino, and B. Pernis. 1987. Monomeric (7S)IgM found in the serum of rheumatoid arthritis patients share idiotypes withpentameric (19S) monoclonal rheumatoid factors. J. Clin. Invest. 79: 813–818.
76. Fakunle, Y. M., F. Aranguibel, D. de Villiers, H. C. Thomas, and S. Sherlock. 1979.Monomeric (7S) IgM in chronic liver disease. Clin. Exp. Immunol. 38: 204–210.
77. Zhou, Z. H., Y. Zhang, Y. F. Hu, L. M. Wahl, J. O. Cisar, and A. L. Notkins.2007. The broad antibacterial activity of the natural antibody repertoire is due topolyreactive antibodies. Cell Host Microbe 1: 51–61.
78. Briles, D. E., M. Nahm, K. Schroer, J. Davie, P. Baker, J. Kearney, andR. Barletta. 1981. Antiphosphocholine antibodies found in normal mouse serumare protective against intravenous infection with type 3 Streptococcus pneumoniae.J. Exp. Med. 153: 694–705.
79. Subramaniam, K. S., K. Datta, E. Quintero, C. Manix, M. S. Marks, andL. A. Pirofski. 2010. The absence of serum IgM enhances the susceptibility of mice topulmonary challenge with Cryptococcus neoformans. J. Immunol. 184: 5755–5767.
80. Ochsenbein, A. F., T. Fehr, C. Lutz, M. Suter, F. Brombacher, H. Hengartner,and R. M. Zinkernagel. 1999. Control of early viral and bacterial distribution anddisease by natural antibodies. Science 286: 2156–2159.
81. Baumgarth, N., O. C. Herman, G. C. Jager, L. E. Brown, L. A. Herzenberg, andJ. Chen. 2000. B-1 and B-2 cell-derived immunoglobulin M antibodies arenonredundant components of the protective response to influenza virus infection.J. Exp. Med. 192: 271–280.
82. Choi, Y. S., and N. Baumgarth. 2008. Dual role for B-1a cells in immunity toinfluenza virus infection. J. Exp. Med. 205: 3053–3064.
83. Jayasekera, J. P., E. A. Moseman, and M. C. Carroll. 2007. Natural antibody andcomplement mediate neutralization of influenza virus in the absence of priorimmunity. J. Virol. 81: 3487–3494.
84. Boes, M., A. P. Prodeus, T. Schmidt, M. C. Carroll, and J. Chen. 1998. A criticalrole of natural immunoglobulin M in immediate defense against systemic bacterialinfection. J. Exp. Med. 188: 2381–2386.
85. Ogden, C. A., R. Kowalewski, Y. Peng, V. Montenegro, and K. B. Elkon. 2005.IGM is required for efficient complement mediated phagocytosis of apoptotic cellsin vivo. Autoimmunity 38: 259–264.
86. Boes, M., C. Esau, M. B. Fischer, T. Schmidt, M. Carroll, and J. Chen. 1998.Enhanced B-1 cell development, but impaired IgG antibody responses in micedeficient in secreted IgM. J. Immunol. 160: 4776–4787.
87. Chen, Y., Y. B. Park, E. Patel, and G. J. Silverman. 2009. IgM antibodies toapoptosis-associated determinants recruit C1q and enhance dendritic cell phago-cytosis of apoptotic cells. J. Immunol. 182: 6031–6043.
88. Notley, C. A., N. Baker, and M. R. Ehrenstein. 2010. Secreted IgM enhancesB cell receptor signaling and promotes splenic but impairs peritoneal B cell sur-vival. J. Immunol. 184: 3386–3393.
89. Chen, Y., S. Khanna, C. S. Goodyear, Y. B. Park, E. Raz, S. Thiel, C. Gronwall,J. Vas, D. L. Boyle, M. Corr, et al. 2009. Regulation of dendritic cells andmacrophages by an anti-apoptotic cell natural antibody that suppresses TLRresponses and inhibits inflammatory arthritis. J. Immunol. 183: 1346–1359.
90. Fabrizio, K., A. Groner, M. Boes, and L. A. Pirofski. 2007. A human monoclonalimmunoglobulin M reduces bacteremia and inflammation in a mouse model ofsystemic pneumococcal infection. Clin. Vaccine Immunol. 14: 382–390.
91. Lewis, M. J., T. H. Malik, M. R. Ehrenstein, J. J. Boyle, M. Botto, and D. O. Haskard.2009. Immunoglobulin M is required for protection against atherosclerosis in low-density lipoprotein receptor-deficient mice. Circulation 120: 417–426.
92. Boes, M., T. Schmidt, K. Linkemann, B. C. Beaudette, A. Marshak-Rothstein, andJ. Chen. 2000. Accelerated development of IgG autoantibodies and autoimmunedisease in the absence of secreted IgM. Proc. Natl. Acad. Sci. USA 97: 1184–1189.
93. Ehrenstein, M. R., H. T. Cook, and M. S. Neuberger. 2000. Deficiency in serumimmunoglobulin (Ig)M predisposes to development of IgG autoantibodies. J. Exp.Med. 191: 1253–1258.
94. Quartier, P., P. K. Potter, M. R. Ehrenstein, M. J. Walport, and M. Botto. 2005.Predominant role of IgM-dependent activation of the classical pathway in theclearance of dying cells by murine bone marrow-derived macrophages in vitro. Eur.J. Immunol. 35: 252–260.
95. Nauta, A. J., N. Raaschou-Jensen, A. Roos, M. R. Daha, H. O. Madsen,M. C. Borrias-Essers, L. P. Ryder, C. Koch, and P. Garred. 2003. Mannose-binding lectin engagement with late apoptotic and necrotic cells. Eur. J. Immu-nol. 33: 2853–2863.
96. Arnold, J. N., M. R. Wormald, D. M. Suter, C. M. Radcliffe, D. J. Harvey,R. A. Dwek, P. M. Rudd, and R. B. Sim. 2005. Human serum IgM glycosylation:identification of glycoforms that can bind to mannan-binding lectin. J. Biol. Chem.280: 29080–29087.
97. Boyden, S. V. 1966. Natural antibodies and the immune response. Adv. Immunol. 5:1–28.
98. Panda, S., J. Zhang, N. S. Tan, B. Ho, and J. L. Ding. 2013. Natural IgG antibodiesprovide innate protection against ficolin-opsonized bacteria. EMBO J. 32: 2905–2919.
99. Craig, A., J. Mai, S. Cai, and S. Jeyaseelan. 2009. Neutrophil recruitment to thelungs during bacterial pneumonia. Infect. Immun. 77: 568–575.
100. Zhou, J., S. A. Stohlman, D. R. Hinton, and N. W. Marten. 2003. Neutrophilspromote mononuclear cell infiltration during viral-induced encephalitis.J. Immunol. 170: 3331–3336.
101. Baranov, D., and P. Neligan. 2007. Trauma and aggressive homeostasis man-agement. Anesthesiol. Clin. 25: 49–63, viii.
102. Simmen, H. P., H. Battaglia, P. Giovanoli, and J. Blaser. 1994. Analysis of pH, pO2and pCO2 in drainage fluid allows for rapid detection of infectious complicationsduring the follow-up period after abdominal surgery. Infection 22: 386–389.
103. Bessman, A. N., J. Page, and L. J. Thomas. 1989. In vivo pH of induced soft-tissueabscesses in diabetic and nondiabetic mice. Diabetes 38: 659–662.
104. TranVan Nhieu, G., C. Clair, G. Grompone, and P. Sansonetti. 2004. Calciumsignalling during cell interactions with bacterial pathogens. Biol. Cell 96: 93–101.
105. Prince, A. S., J. P. Mizgerd, J. Wiener-Kronish, and J. Bhattacharya. 2006. Cellsignaling underlying the pathophysiology of pneumonia. Am. J. Physiol. Lung Cell.Mol. Physiol. 291: L297–L300.
106. Eichstaedt, S., K. Gabler, S. Below, C. M€uller, C. Kohler, S. Engelmann,P. Hildebrandt, U. Volker, M. Hecker, and J. P. Hildebrandt. 2009. Effects ofStaphylococcus aureus-hemolysin A on calcium signalling in immortalized humanairway epithelial cells. Cell Calcium 45: 165–176.
107. Zhang, J., J. Koh, J. Lu, S. Thiel, B. S. Leong, S. Sethi, C. Y. He, B. Ho, andJ. L. Ding. 2009. Local inflammation induces complement crosstalk whichamplifies the antimicrobial response. PLoS Pathog. 5: e1000282.
108. Puga, I., and A. Cerutti. 2013. Protection by natural IgG: a sweet partnership withsoluble lectins does the trick! EMBO J. 32: 2897–2899.
109. Panda, S., J. Zhang, L. Yang, G. S. Anand, and J. L. Ding. 2014. Molecular in-teraction between natural IgG and ficolin–mechanistic insights on adaptive-innateimmune crosstalk. Sci. Rep. 4: 3675.
110. Medzhitov, R., and C. A. Janeway, Jr. 2002. Decoding the patterns of self andnonself by the innate immune system. Science 296: 298–300.
111. Beutler, B., Z. Jiang, P. Georgel, K. Crozat, B. Croker, S. Rutschmann, X. Du,and K. Hoebe. 2006. Genetic analysis of host resistance: Toll-like receptor sig-naling and immunity at large. Annu. Rev. Immunol. 24: 353–389.
112. Brown, G. D. 2006. Dectin-1: a signalling non-TLR pattern-recognition receptor.Nat. Rev. Immunol. 6: 33–43.
113. Hoebe, K., P. Georgel, S. Rutschmann, X. Du, S. Mudd, K. Crozat, S. Sovath,L. Shamel, T. Hartung, U. Zahringer, and B. Beutler. 2005. CD36 is a sensor ofdiacylglycerides. Nature 433: 523–527.
114. McGrath, F. D., M. C. Brouwer, G. J. Arlaud, M. R. Daha, C. E. Hack, andA. Roos. 2006. Evidence that complement protein C1q interacts with C-reactiveprotein through its globular head region. J. Immunol. 176: 2950–2957.
115. Lu, J., K. D. Marjon, C. Mold, T. W. Du Clos, and P. D. Sun. 2012. Pentraxinsand Fc receptors. Immunol. Rev. 250: 230–238.
116. Mantovani, A., S. Valentino, S. Gentile, A. Inforzato, B. Bottazzi, andC. Garlanda. 2013. The long pentraxin PTX3: a paradigm for humoral patternrecognition molecules. Ann. N. Y. Acad. Sci. 1285: 1–14.
117. Duncan, A. R., and G. Winter. 1988. The binding site for C1q on IgG. Nature332: 738–740.
118. Senaldi, G., R. Ireland, A. J. Bellingham, D. Vergani, K. Veerapan, and F. Wang.1988. IgM reduction in systemic lupus erythematosus. Arthritis Rheum. 31: 1213.
119. Shaw, P. X., C. S. Goodyear, M. K. Chang, J. L. Witztum, and G. J. Silverman.2003. The autoreactivity of anti-phosphorylcholine antibodies for atherosclerosis-associated neo-antigens and apoptotic cells. J. Immunol. 170: 6151–6157.
120. Tuominen, A., Y. I. Miller, L. F. Hansen, Y. A. Kesaniemi, J. L. Witztum, andS. Horkko. 2006. A natural antibody to oxidized cardiolipin binds to oxidized low-density lipoprotein, apoptotic cells, and atherosclerotic lesions. Arterioscler.Thromb. Vasc. Biol. 26: 2096–2102.
121. Gray, M., K. Miles, D. Salter, D. Gray, and J. Savill. 2007. Apoptotic cells protectmice from autoimmune inflammation by the induction of regulatory B cells. Proc.Natl. Acad. Sci. USA 104: 14080–14085.
122. Kanevets, U., K. Sharma, K. Dresser, and Y. Shi. 2009. A role of IgM antibodies inmonosodium urate crystal formation and associated adjuvanticity. J. Immunol.182: 1912–1918.
123. Kulik, L., S. D. Fleming, C. Moratz, J. W. Reuter, A. Novikov, K. Chen,K. A. Andrews, A. Markaryan, R. J. Quigg, G. J. Silverman, et al. 2009. Patho-genic natural antibodies recognizing annexin IV are required to develop intestinalischemia-reperfusion injury. J. Immunol. 182: 5363–5373.
124. Haas, M. S., E. M. Alicot, F. Schuerpf, I. Chiu, J. Li, F. D. Moore, andM. C. Carroll. 2010. Blockade of self-reactive IgM significantly reduces injury ina murine model of acute myocardial infarction. Cardiovasc. Res. 87: 618–627.
125. Subramanian, K., R. Du, N. S. Tan, B. Ho, and J. L. Ding. 2013. CD163 and IgGcodefend against cytotoxic hemoglobin via autocrine and paracrine mechanisms.J. Immunol. 190: 5267–5278.
126. Hess, C., A. Winkler, A. K. Lorenz, V. Holecska, V. Blanchard, S. Eiglmeier,A. L. Schoen, J. Bitterling, A. D. Stoehr, D. Petzold, et al. 2013. T cell-independent B cell activation induces immunosuppressive sialylated IgG anti-bodies. J. Clin. Invest. 123: 3788–3796.
20 BRIEF REVIEWS: NATURAL Abs BRIDGE INNATE AND ADAPTIVE IMMUNITY
by guest on September 3, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
Related Documents











