Research Article Acyl-Coenzyme A–Binding Protein Regulates Beta-Oxidation Required for Growth and Survival of Non–Small Cell Lung Cancer Fredrick T. Harris 1,2 , S.M. Jamshedur Rahman 2 , Mohamed Hassanein 2 , Jun Qian 2 , Megan D. Hoeksema 2 , Heidi Chen 3 , Rosana Eisenberg 4 , Pierre Chaurand 8 , Richard M. Caprioli 6 , Masakazu Shiota 5 , and Pierre P. Massion 2,7 Abstract We identified acyl-coenzyme A–binding protein (ACBP) as part of a proteomic signature predicting the risk of having lung cancer. Because ACBP is known to regulate b-oxidation, which in turn controls cellular proliferation, we hypothesized that ACBP contributes to regulation of cellular proliferation and survival of non–small cell lung cancer (NSCLC) by modulating b-oxidation. We used matrix-assisted laser desorption/ ionization-imaging mass spectrometry (MALDI-IMS) and immunohistochemistry (IHC) to confirm the tissue localization of ABCP in pre-invasive and invasive NSCLCs. We correlated ACBP gene expression levels in NSCLCs with clinical outcomes. In loss-of-function studies, we tested the effect of the downregulation of ACBP on cellular proliferation and apoptosis in normal bronchial and NSCLC cell lines. Using tritiated- palmitate ( 3 H-palmitate), we measured b-oxidation levels and tested the effect of etomoxir, a b-oxidation inhibitor, on proliferation and apoptosis. MALDI-IMS and IHC analysis confirmed that ACBP is over- expressed in pre-invasive and invasive lung cancers. High ACBP gene expression levels in NSCLCs correlated with worse survival (HR ¼ 1.73). We observed a 40% decrease in b-oxidation and concordant decreases in proliferation and increases in apoptosis in ACBP-depleted NSCLC cells as compared with bronchial airway epithelial cells. Inhibition of b-oxidation by etomoxir in ACBP-overexpressing cells produced dose- dependent decrease in proliferation and increase in apoptosis (P ¼ 0.01 and P < 0.001, respectively). These data suggest a role for ACBP in controlling lung cancer progression by regulating b-oxidation. Cancer Prev Res; 7(7); 748–57. Ó2014 AACR. Introduction Early detection efforts in our laboratory have led to the discovery by matrix-assisted laser desorption/ionization- mass spectrometry (MALDI-MS) of acyl-coenzyme A–bind- ing protein (ACBP) as 1 of a 6-protein signature predicting the risk of having lung cancer in individuals with endo- bronchial lesions (1, 2). This novel observation prompted us to investigate its role in lung cancer progression. The ACBP gene encodes a 90–amino acid protein with molec- ular mass of 10 kDa that binds medium to long-chain acyl- CoA esters (C 12 –C 22 ) with high affinity and specificity (3, 4). ACBP was formerly known as the diazepam-binding inhibitor (DBI) by its ability to displace diazepam from its binding sites on type A gamma g -aminobutyric acid recep- tors (GABA A receptors; ref. 5). ACBP induces steroidogen- esis in isolated adrenal mitochondria, inhibits glucose- induced insulin secretion from the pancreas, induces medi- um-chain acyl-CoA ester synthesis, and affects cell growth (6–12). Mostly cytosolic, ACBP is localized to the endo- plasmic reticulum (ER) and Golgi apparatus (13). Fatty acyl-CoAs (ACBP’s substrate) are transported across the inner mitochondrial membrane by carnitine and play an important role in a host of cellular processes such as phospholipid synthesis, protein kinase C activation (PKC), and b-oxidation (7, 9, 12, 14). Cancer cells favor alternative pathways to meet the increased nutritional demand of cell growth (15). The mitochondrial metabolism can be repro- grammed to meet the macronutrient demand of sustained cellular growth (15). The catabolism or b-oxidation of acyl- CoAs is an important process in fulfilling these cellular energy requirements during metabolic stress. The process involves acyl-CoA moieties being broken down in the mitochondria to generate acetyl-CoA. The acetyl-CoA can Authors' Affiliations: 1 Department of Biochemistry and Cancer Biology, Meharry Medical College; 2 Division of Allergy, Pulmonary and Critical Care Medicine, Departments of 3 Biostatistics, 4 Pathology, Microbiology and Immunology, and 5 Molecular Physiology and Biophysics, 6 Mass Spec- trometry Research Center, Vanderbilt University School of Medicine; 7 Veterans Affairs Medical Center, Nashville, Tennessee; and 8 Department of Chemistry, Universit e de Montr eal, Montreal, Quebec, Canada Note: Supplementary data for this article are available at Cancer Prevention Research Online (http://cancerprevres.aacrjournals.org/). Corresponding Author: Pierre P. Massion, Vanderbilt-Ingram Cancer Center, PRB 640, 2220 Pierce Avenue, Nashville TN 37232. Phone: 615- 936-2256; Fax: 615-936-1790; E-mail: [email protected] doi: 10.1158/1940-6207.CAPR-14-0057 Ó2014 American Association for Cancer Research. Cancer Prevention Research Cancer Prev Res; 7(7) July 2014 748 Research. on May 18, 2020. © 2014 American Association for Cancer cancerpreventionresearch.aacrjournals.org Downloaded from Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Research Article
Acyl-CoenzymeA–Binding Protein Regulates Beta-OxidationRequired for Growth and Survival of Non–Small Cell LungCancer
Fredrick T. Harris1,2, S.M. Jamshedur Rahman2, Mohamed Hassanein2, Jun Qian2, Megan D. Hoeksema2,Heidi Chen3, Rosana Eisenberg4, Pierre Chaurand8, Richard M. Caprioli6, Masakazu Shiota5, andPierre P. Massion2,7
AbstractWe identified acyl-coenzyme A–binding protein (ACBP) as part of a proteomic signature predicting the
risk of having lung cancer. Because ACBP is known to regulate b-oxidation, which in turn controls cellular
proliferation, we hypothesized that ACBP contributes to regulation of cellular proliferation and survival of
non–small cell lung cancer (NSCLC) bymodulating b-oxidation. We usedmatrix-assisted laser desorption/
ionization-imaging mass spectrometry (MALDI-IMS) and immunohistochemistry (IHC) to confirm the
tissue localization of ABCP in pre-invasive and invasiveNSCLCs.We correlated ACBP gene expression levels
in NSCLCs with clinical outcomes. In loss-of-function studies, we tested the effect of the downregulation of
ACBP on cellular proliferation and apoptosis in normal bronchial and NSCLC cell lines. Using tritiated-
palmitate (3H-palmitate), we measured b-oxidation levels and tested the effect of etomoxir, a b-oxidationinhibitor, on proliferation and apoptosis. MALDI-IMS and IHC analysis confirmed that ACBP is over-
expressed in pre-invasive and invasive lung cancers. HighACBP gene expression levels inNSCLCs correlated
with worse survival (HR ¼ 1.73). We observed a 40% decrease in b-oxidation and concordant decreases in
proliferation and increases in apoptosis in ACBP-depleted NSCLC cells as compared with bronchial airway
epithelial cells. Inhibition of b-oxidation by etomoxir in ACBP-overexpressing cells produced dose-
dependent decrease in proliferation and increase in apoptosis (P ¼ 0.01 and P < 0.001, respectively).
These data suggest a role for ACBP in controlling lung cancer progression by regulating b-oxidation. CancerPrev Res; 7(7); 748–57. �2014 AACR.
IntroductionEarly detection efforts in our laboratory have led to the
discovery by matrix-assisted laser desorption/ionization-mass spectrometry (MALDI-MS) of acyl-coenzyme A–bind-ing protein (ACBP) as 1 of a 6-protein signature predictingthe risk of having lung cancer in individuals with endo-bronchial lesions (1, 2). This novel observation promptedus to investigate its role in lung cancer progression. TheACBP gene encodes a 90–amino acid protein with molec-
ular mass of 10 kDa that binds medium to long-chain acyl-CoA esters (C12–C22) with high affinity and specificity (3,4). ACBP was formerly known as the diazepam-bindinginhibitor (DBI) by its ability to displace diazepam from itsbinding sites on type A gamma g-aminobutyric acid recep-tors (GABAA receptors; ref. 5). ACBP induces steroidogen-esis in isolated adrenal mitochondria, inhibits glucose-induced insulin secretion from the pancreas, induces medi-um-chain acyl-CoA ester synthesis, and affects cell growth(6–12). Mostly cytosolic, ACBP is localized to the endo-plasmic reticulum (ER) and Golgi apparatus (13). Fattyacyl-CoAs (ACBP’s substrate) are transported across theinner mitochondrial membrane by carnitine and play animportant role in a host of cellular processes such asphospholipid synthesis, protein kinase C activation (PKC),and b-oxidation (7, 9, 12, 14). Cancer cells favor alternativepathways to meet the increased nutritional demand of cellgrowth (15). The mitochondrial metabolism can be repro-grammed to meet the macronutrient demand of sustainedcellular growth (15). The catabolism or b-oxidation of acyl-CoAs is an important process in fulfilling these cellularenergy requirements during metabolic stress. The processinvolves acyl-CoA moieties being broken down in themitochondria to generate acetyl-CoA. The acetyl-CoA can
Authors' Affiliations: 1Department of Biochemistry and Cancer Biology,Meharry Medical College; 2Division of Allergy, Pulmonary and Critical CareMedicine, Departments of 3Biostatistics, 4Pathology, Microbiology andImmunology, and 5Molecular Physiology and Biophysics, 6Mass Spec-trometry Research Center, Vanderbilt University School of Medicine;7Veterans Affairs Medical Center, Nashville, Tennessee; and 8Departmentof Chemistry, Universit�e de Montr�eal, Montreal, Quebec, Canada
Note:Supplementary data for this article are available atCancer PreventionResearch Online (http://cancerprevres.aacrjournals.org/).
Corresponding Author: Pierre P. Massion, Vanderbilt-Ingram CancerCenter, PRB 640, 2220 Pierce Avenue, Nashville TN 37232. Phone: 615-936-2256; Fax: 615-936-1790; E-mail: [email protected]
doi: 10.1158/1940-6207.CAPR-14-0057
�2014 American Association for Cancer Research.
CancerPreventionResearch
Cancer Prev Res; 7(7) July 2014748
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

then serve as the substrate for the tricarboxylic acid (TCA)cycle for the generation of ATP. Carnitine palmitoyl trans-ferase-1 (CPT-1) is the key regulatory enzymemediating thetransport of acyl-CoAs with carnitine into the mitochon-dria. CPT-1 interacts with the acyl-CoA/ACBP complex, andthe activity of CPT-1 is correlated with the concentration ofACBP-bound acyl-CoAs, but not with levels of free acyl-CoAs (16). The b-oxidation pathway is a dominant bioe-nergetic pathway in prostate cancer which increases fattyacid utilization to provide increased ATP levels; however,little is known about its role in lung cancer (17). Wetherefore hypothesize that ACBP regulates proliferation andsurvival of non–small cell lung cancer (NSCLC) by mod-ulating b-oxidation. Determining the role of ACBP in lungcancer may enhance our understanding of lung cancerprogression and establish its implications in diagnosis andtreatment.
Material and MethodsMALDI–time-of-flight imaging MS (MALDI-TOF-IMS)proteomic analysisBronchial biopsy specimenswere frozen in liquidnitrogen
immediately after collection under an Institutional ReviewBoard (IRB)-approved protocol, embedded in Tissue-TekOptimal Cutting Temperature compound (IMEB), andstored at �80�C. To determine the location of proteinexpression byMS, we followed protocols described in earlierwork (1, 18–20). Briefly, a section from a unique bronchialtissue specimen containing progressive histologic stages ofNSCLC development, including normal bronchial epitheli-um, moderate dysplasia, severe dysplasia, carcinoma in situ(CIS), and invasive cancer, was cut at 12 mm and immedi-ately thaw-mountedonconductive indium-tinoxide–coatedglass slides (Delta Technologies Ltd). A sequential section
was cut and mounted onto a charged glass slide and stainedwith hematoxylin and eosin (H&E; Fig. 1A). The acousticroboticmicrospotterwas used to printmatrix (sinapinic acidprepared at 10 mg/mL in 60/40-acetonitrile/0.1% trifluor-oacetic acid) in an array format with a center-to-centerdistance of 300 mm covering the entire surface of the section(Fig. 1B). A total of 28 droplets of matrix were collected ateach array coordinate to form theMALDI spots. MALDI-IMSdata were acquired on an Applied Biosystems DE-STR TOFMS, equippedwith a nitrogen laser (N2, 337 nm)operated ata repletion rate of 20 Hz. Protein MS data were acquired inthe linear mode geometry with 25 kV of accelerating poten-tial under optimized delayed extraction conditions, focusingthe ions at �m/z 15,000. Under these conditions, optimalsignal resolutions were obtained throughout the studiedmass range from 2 to 25 kDa. MALDI-IMS data acquisitionwas automated using a custom-built plate geometry filecorresponding to the printedmatrix array. From eachmatrixspotof the array, amass spectrumwas generatedby summingthe signals from250 consecutive laser shots. Ion imageswereassembled using custom-built software and visualized usingBiomap (Novartis). After data acquisition, matrix wasremoved from the section by immersion in 70% ethanolfor 1minute. The sectionwas then immediately stainedwithH&E.
Validation of ACBP localization and expression byimmunohistochemistry
Five-micrometer tissue sections were cut from formalin-fixed, paraffin-embedded (FFPE) lung tissue blocks, depar-affinized by placing in 3 xylene baths, 10 minutes each,hydrated through a series of ethanol baths of decreasingconcentration, and placed in a buffer bath of TBS. Endog-enous peroxide was quenched using Dako Peroxidase
Inv
CIS
CIS
A B
C D2 m
m
Sev
dysp
0 720
640
560
480
400
320
240
160
80
09,900 9,910 9,920 9,930 9,940 9,950 9,960 9,970 9,980 9,990
9955,ACBP
100%
Mass/charge (m/z)
Invasive
In situ
Severe
Moderate
Normal
Ion
cu
rre
nt in
ten
sity
Mod
dysp N1 br ep
Figure 1. ACBP levels increasetoward invasive cancer andcorrelate with lower overall survivalrates. A, H&E stain of 2-mm tissuesection (total thickness) showingpathologic changes towardsinvasive carcinoma. This sectionwas used to obtain MALDI-IMSdata. B, array of matrix depositionon the entire tissue section. C,MALDI-IMS image of m/z9,955 corresponding to ACBPdemonstrating overexpression inhigh-grade pre-invasive andinvasive areas. Gray scale isprovided in the inset, with whitecorresponding to the highestintensity. D, MALDI-MS histogramof m/z 9,955 corresponding toACBP, demonstrating gradualincrease in high-grade pre-invasive and invasive areas of the2-mm tissue section. Inv, invasivecancer; Mod Dys, moderatedysplasia; Nl Br epithelium, normalbronchial epithelium; Sev Dys,severe dysplasia.
Potential Role of ACBP in Tumor Progression in Lung Cells
www.aacrjournals.org Cancer Prev Res; 7(7) July 2014 749
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

Blocking Reagent and nonspecific staining blocked usingDako Serum-Free Protein Block according to manufac-turer’s instructions. IHC analysis was performed as previ-ously reported by our group (21). Results of the immuno-histochemical (IHC) staining were analyzed by a patholo-gist (R. Eisenberg).
Analysis of overall survival of patients with high ACBPexpression
Kaplan–Meier plotter on line survival analysis software(22) was used to test the relevance of changes in geneexpression to overall survival (kmplotter.com). The follow-ing publically available datasets were used: GSE31210,GSE29013, and GSE4573. Mixed adenocarcinomas andsquamous cell carcinomas (SCC), staged 1–3 (411 totalsamples), were represented. HR (and 95% confidence inter-vals) and log-rank P are provided in the online application;the association between ACBP and clinical variables wasanalyzed using a multivariant Cox regression excludingbiased arrays.
Cell cultureHuman lung cancer cell lines A549, adenocarcinoma,
H520, SCC, and the normal human bronchial epithelial(HBE) cell line, 16-HBE (ATCC) were maintained inRPMI-1640 medium (Gibco by Life Technologies) con-taining 10% heat-inactivated FBS (Gibco by Life Tech-nologies), at 37�C, 100% humidity, and 5% CO2. Cellswere tested every 6 months for mycoplasma using thePCR-based MycoAlert PLUS Mycoplasma Detection Kit(Lonza). For cells that had been infected with shRNAlentivirus, cells were maintained in additional media thatcontained 1.5 mg/mL puromycin (Sigma). Transfectedcells were of low-passage number (>15 passages), andfurther tested for all common murine pathogens by PCRat Charles River Research Animal Diagnostic Services.During experiments, cells were plated and allowed toadhere overnight. Cells were then washed with sterilePBS (8.1 mmol/L Na2HPO4, 1.47 mmol/L KH2PO4,137 mmol/L NaCl, and 2.68 mmol/L KCl; pH 7.4). Next,RPMI was added that contained no FBS but was supple-mented with EGF (25 ng/mL) and with 1� growth factorcocktail (Invitrogen) that includes insulin, selenium, andtransferrin. HEK293T cells were maintained in DMEMwith 10% FBS.
shRNA knockdownTo stably silence ACBP in H520, A549, and 16-HBE cells,
3 individual pGIPZ lentiviral shRNA-ACBP and 1 pGIPZnonsilencing shRNA lentiviral control vector were pur-chased from Open Biosystems. Transfection and transduc-tion were conducted according to manufacturer’s instruc-tions or as previously described (23). Briefly, low-passageHEK293T cells were plated and allowed to grow to about90% confluency. Lentiviral packaging mix and lentiviralvectors were introduced to the cells and allowed to integrateinto the genome. Media were changed and 2 days postinte-gration virus was collected. Virus was then purified by
filtering through a 0.4-mm filter. Cells to be infected wereplated the day before infection and subsequently incubatedwith virus in 8 mg/mL polybrene for 6 hours. Puromycin(1.5mg/mL)was added the following day for selection. Aftercolony selection, ACBP expression in various cell colonieswas measured using immunoblotting.
Proliferation assaysCells were seeded into 24-well plates at a density of 2 �
104 per well. Cells were allowed to adhere overnight, andthe next day cells were washed with PBS and 500 mL ofsupplemented media was added. At the respective timepoints, 50 mL of CellTiter 96 AQueous Non-RadioactiveCell ProliferationAssay (MTS) solutionwas added into eachwell and the experiment was carried out as per the manu-facturer’s instructions (Promega). Briefly, cells were gentlyshaken, and the plates were incubated at 37�C for 1 hour.The absorbance was detected at 490 nm with a MicroplateReader (Epoch Microplate Spectrophotometer, Biotek). Allexperiments were repeated 3 times. The WST-1 assay(Roche) was performed as previously reported by our group(24). Briefly, stably transfected H520 cells were seeded in96-well plates at 1.5� 103 cells per well. Cells were allowedto adhereovernight, and thenext day cellswerewashedwithPBS and 500 mL of supplemented media was added. WST-1reagent was used for spectrophotometric quantification ofcell proliferation.
ATP assayATP measurements were determined using the ATP Col-
orimetric/Fluorometric Assay Kit (Abcam). Measurementswere performedwith the fluorometric assay in 96-well blackbottom plates at the 48-hour time point, following themanufacturer’s instructions.
Reagents and antibodiesEtomoxir sodium salt were purchased from Sigma
Aldrich. ACBPantibodywaspurchased fromAbcam,where-as PARP and cleaved caspase-3 antibodies were from CellSignaling. All antibodies were used according to manufac-turers’ dilutions. Puromycin dihydrochloride (Invitrogen)was used at a concentration of 1.5 mg/mL for colonyselection.
b-Oxidation assayb-Oxidationmeasurementswere performedas previously
described with slightmodifications in cell media (25). Cellswere plated in RPMI culture medium containing 10% FBS.Cells were plated in 6-well dishes and allowed to adhereovernight, and media were replaced with defined culturemedia supplemented with EGF (25 ng/mL) and with 1�growth factor cocktail (Invitrogen). The media also con-tained albumin-bound [9,10-3H(N)] palmitic acid (4 mCi3H/mmol palmitate) at a final concentration of 400 mmol/L.The final volume of the media solution, including definedculture with palmitate, was calculated to be exactly 2mL perwell. After 48 hours of incubation, 1.5 mL of media wasremoved directly from each well and collected in individual
Harris et al.
Cancer Prev Res; 7(7) July 2014 Cancer Prevention Research750
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

round-bottom snap-top tubes. The remaining assay wasperformed as indicated in the cited protocol.
Mitochondrial potential assayThe lipophilic cation MitoProbe JC-1 Assay Kit (Cayman
Chemical) was used to assess the mitochondrial status ofNSCLC cells with etomoxir added. JC-1 (Invitrogen) is alipophilic membrane–permeant cation that selectivelyenters the mitochondria. JC-1 exists in a monomeric formproducing a green fluorescence or an aggregated formproducing a red fluorescence upon mitochondrial hyper-polarization. Briefly, cells were plated at a density of 5� 104
cells per 200mL in 96-well black, clear-bottom plates. Cellswere allowed to adhere overnight and subsequentlywashed. The respective concentrations of etomoxir mixedwith supplemented, defined RPMI media were then added.Sixteen hours later, the JC-1 assay was performed, andfluorescence was measured in a Flexstation II plate reader(Molecular Device) with wavelengths of excitation andemission (ex/em) 530/580 nm ("red") and then at ex/em485/535 nm ("green"). The ratio of green to red reflects theDym (membrane potential).
Statistical analysisStatistical analysis for the proliferation assays, b-oxidation
assay, ATP assay, and Dym indicator assay was performedusing the GraphPad Prism software (GraphPad Software).Data comparing2 experimental conditionswere analyzedby2-tailed Student t test. Only results with P < 0.05 or P < 0.01were considered to be statistically significant. All experimen-tal data are presented as a representative of 3 independentexperiments. All treatments within each experiment wereperformed in quadruplicate wells (proliferation assay, ATPassay, and Dym indicator assay) or triplicate fashion (b-oxi-dation assay) and repeated on 3 independent days.
ResultsCharacterization of ACBP expression and the clinicalrelevance to NSCLCs
To further assess the relevance of ABCP in lung cancerprogression, we obtained a unique 2-cm tissue sample thatdisplayed varying levels of cancer pathologic progression,including normal bronchial epithelium, moderate dyspla-sia, severe dysplasia, CIS, and invasive cancer (Fig. 1A). Thelevels of ACBP expression increased from normal to CIS(Fig. 1C). ACBP has an m/z value of 9,955 and the corre-sponding MALDI-MS peak amplitude varied in intensityacross the spectrum from normal to pre-invasive and inva-sive tissues (Fig. 1D). We next analyzed ACBP levels in apanel of paired primary lung SCCs, adenocarcinomas, andmatched normal adjacent lung tissues by Western blotanalysis. We discovered that ACBP is overexpressed in amajority of the SCCs and adenocarcinomas compared withthe adjacent normal lung tissue (Fig. 2A and B). To furthertest whether ACBP is associated with lung cancer progres-sion, we used IHC to stain additional tissue sections of pre-invasive lesions varying in severity to demonstrate anincreased staining pattern going in a continuum fromnormal bronchial epithelium to lung cancer (Fig. 2C). Theimmunostaining pattern was predominantly cytoplasmic,but exhibited some nuclear localization (Fig. 2C). We alsonote that the staining in the normal bronchial epithelium isconfined to inflammatory cells, pneumocytes, and macro-phages (Fig. 2C). Examining mRNA expression data in 411NSCLC samples (including 241 patients with stage I, 91with stage II, 23with stage III disease, and 56with unknownstaging), our multivariate Cox analysis revealed that ACBPexpression is correlated with NSCLC stage (HR ¼ 1.9; 95%CI, 1.49–2.41), patient smoking history (HR ¼ 0.53; 95%CI, 0.3–0.95), and overall poor survival in NSCLCs (HR ¼
NA B
CNI. br. epithelium
Moderate
dysplasia CIS ADC SCC
4×
40×
ACBP
α-Tubulin
ACBP
α-Tubulin
T N T N T N T N T N T N T N T
Figure 2. ACBP expression inhuman lung tumor development.A and B, Western blot analysesof ACBP protein levels inadenocarcinoma and SCCsamples (N, normal; T, tumor). C,IHC staining of patient tissues, withvarying magnifications. These datademonstrate a gradual increasein ACBP expression levelstoward invasive cancer. ADC,adenocarcinoma;NlBr. epithelium,normal bronchial epithelium.
Potential Role of ACBP in Tumor Progression in Lung Cells
www.aacrjournals.org Cancer Prev Res; 7(7) July 2014 751
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

1.73; 95% CI, 1.19–2.51; Fig. 3, Supplementary Table S1;ref. 22). Taken together, these results demonstrate thatACBP is differentially expressed in NSCLC cell lines andoverexpressed in lung cancer and in pre-invasive squamouslung tissues.
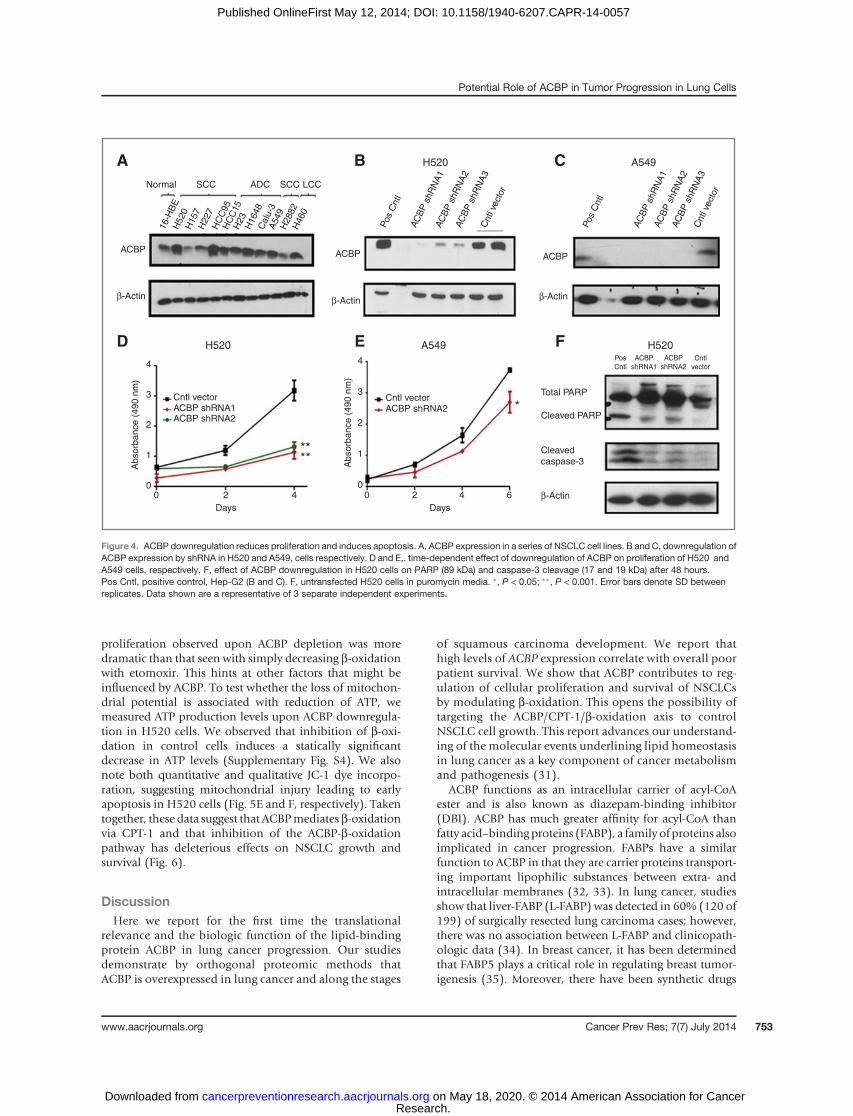
Loss of ACBP function reduces NSCLC proliferationand survival
We found that ACBP is differentially expressed in allNSCLC cell lines tested compared with noncancerous HBEcells (16-HBE in Fig. 4A and IHCSupplementary Fig. S1). Totest the hypothesis that high ACBP levels contribute toNSCLC proliferation and survival, we knocked down ACBPwith shRNA in H520 and A549 cells, 2 NSCLC cell linesstrongly expressing ACBP (Fig. 4A). We used the immor-talized human bronchial cell line 16-HBE as a control. Twoefficient knockdown clones were generated using single-colony selection under puromycin. Visually, both shRNAclones showed efficient (80%) knockdownof ACBP in the 3cell lines tested (H520 and A549 in Fig. 4B and C and 16-HBE in Supplementary Fig. S2A). To determine whetherACBPdepletion affectedNSCLCproliferation,wemeasuredcell proliferation in the ACBP shRNA–depleted cells. Silenc-ing ACBP significantly reduced the proliferation of H520and A549 (Fig. 4D and E). In contrast, silencing of ACBP in16-HBE cells had no significant effect on proliferation(Supplementary Fig. S2B). To determine whether the effectson cell proliferation are due to apoptosis, we examined thelevels of 2 apoptoticmarkers, cleaved caspase-3 and cleavedPARP. ACBP downregulation resulted in increases in bothcleaved caspase-3 and cleaved PARP in H520 cells demon-strating both early- and late-stage apoptosis, respectively
(Fig. 4F). In contrast, ACBP silencing by shRNA had nonoticeable effects on either cleaved caspase-3 or cleavedPARP in 16-HBE cells (Supplementary Fig. S2C). Together,these results confirmed that downregulation of ACBP inhi-bits NSCLC proliferation and induces apoptosis in thecancer cell lines tested.
Fatty acid oxidation is mediated by ACBP in NSCLCsACBP is important for a host of biologic responses,
including regulation of the intracellular acyl-CoA pool sizeand donation of acyl-CoA esters for b-oxidation (26).Because cancer cells rely on altered metabolism and repro-gramming of metabolic pathways for survival (27), wehypothesized that adenocarcinomas and SCCs rely onmitochondrial ACBP-mediated b-oxidation for overall sur-vival. Using tritiated-palmitate (3H-palmitate) at normalphysiological concentrations of 40 mmol/L as a precursorsubstrate, we measured b-oxidation levels using scintilla-tion counts of ion exchange column fragments inH520 and16-HBE cells. Because b-oxidation increases when cellsundergo metabolic stress, cells were cultured without glu-cose and we measured b-oxidation levels. In ACBP controlvector cells, we discovered that without glucose there is astatistically significant increase in b-oxidation (P < 0.05; Fig.5A). In contrast, in ACBP-depleted cells, in the presence ofglucose, there is a statistically significant decrease in b-oxi-dation compared with control cells (P < 0.05; Fig. 5A).Moreover, we did not observe any significant increase inb-oxidation in our ACBP-depleted cells when glucose wasremoved. This suggests that ACBP mediates b-oxidation inH520 cells. In contrast to H520, b-oxidation levels wereunaffected in 16-HBE cells upon ACBP depletion (Fig. 5B).These results indicate that the downregulation of ACBPaffects lung cancer cell metabolism.
Fatty acid oxidation is required for NSCLC growth andsurvival
To further determine whether b-oxidation is a key playerin this ACBP-mediated regulation of cell proliferation andsurvival, we inhibited b-oxidation pharmacologically usingetomoxir, a potent inhibitor of CPT-1 (28). We first con-firmed that etomoxir could in fact inhibit b-oxidation inNSCLCs at a relatively lowdose, that is, 10mmol/L (Fig. 5C).To further elucidate the effect of inhibition of b-oxidationon NSCLC cells mitochondrial integrity, the membrane-permeant JC-1 dye was used to quantify alterations inmitochondrial potential that can serve as an early sign ofapoptosis (29, 30). Using the JC-1 assay, we found a dose-dependent decrease in mitochondrial potential (Dym) inboth H520 and A549 cells by etomoxir (Fig. 5E; Supple-mentary Fig. S3). The effect was more pronounced at lowerconcentrations in theH520 cells than in the A549 cells. Thisis consistent with the prosurvival role of the b-oxidationpathway in NSCLCs. To further test the importance of theb-oxidation pathway on growth and survival, we examinedproliferation when b-oxidation is inhibited. Etomoxirinduced a dose-dependent decrease in proliferation inH520 cells with intact ACBP (Fig. 5D). The decrease in
Gene expression data
HR = 1.73 (1.19–2.51)
log-rank P = 0.0039
LowHigh
0 20
N= 206
N= 205
188
165
130
117
74
79
25
34
16
22
0
10
0
2
40 60 80
Time (mo)
100 120 140
1.0
0.8
0.6
Pro
ba
bili
ty
0.4
0.2
0.0
Figure 3. ACBP overexpression correlates with poor overall survival inpatients with lung cancer. Overall survival of patients with NSCLCs withhigh expression (red line) or low expression (black line) of ACBP.Data are derived from 3 publically available gene expression data sets(GSE31210, GSE29013, and GSE4573; n ¼ 411) using a meanexpression of 3 probes that match the annotated ACBP transcript.
Harris et al.
Cancer Prev Res; 7(7) July 2014 Cancer Prevention Research752
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057
proliferation observed upon ACBP depletion was moredramatic than that seen with simply decreasing b-oxidationwith etomoxir. This hints at other factors that might beinfluenced by ACBP. To test whether the loss of mitochon-drial potential is associated with reduction of ATP, wemeasured ATP production levels upon ACBP downregula-tion in H520 cells. We observed that inhibition of b-oxi-dation in control cells induces a statically significantdecrease in ATP levels (Supplementary Fig. S4). We alsonote both quantitative and qualitative JC-1 dye incorpo-ration, suggesting mitochondrial injury leading to earlyapoptosis in H520 cells (Fig. 5E and F, respectively). Takentogether, these data suggest that ACBPmediates b-oxidationvia CPT-1 and that inhibition of the ACBP-b-oxidationpathway has deleterious effects on NSCLC growth andsurvival (Fig. 6).
DiscussionHere we report for the first time the translational
relevance and the biologic function of the lipid-bindingprotein ACBP in lung cancer progression. Our studiesdemonstrate by orthogonal proteomic methods thatACBP is overexpressed in lung cancer and along the stages
of squamous carcinoma development. We report thathigh levels of ACBP expression correlate with overall poorpatient survival. We show that ACBP contributes to reg-ulation of cellular proliferation and survival of NSCLCsby modulating b-oxidation. This opens the possibility oftargeting the ACBP/CPT-1/b-oxidation axis to controlNSCLC cell growth. This report advances our understand-ing of the molecular events underlining lipid homeostasisin lung cancer as a key component of cancer metabolismand pathogenesis (31).
ACBP functions as an intracellular carrier of acyl-CoAester and is also known as diazepam-binding inhibitor(DBI). ACBP has much greater affinity for acyl-CoA thanfatty acid–binding proteins (FABP), a family of proteins alsoimplicated in cancer progression. FABPs have a similarfunction to ACBP in that they are carrier proteins transport-ing important lipophilic substances between extra- andintracellular membranes (32, 33). In lung cancer, studiesshow that liver-FABP (L-FABP) was detected in 60% (120 of199) of surgically resected lung carcinoma cases; however,there was no association between L-FABP and clinicopath-ologic data (34). In breast cancer, it has been determinedthat FABP5 plays a critical role in regulating breast tumor-igenesis (35). Moreover, there have been synthetic drugs
Normal
ACBP
4
3
2
1
0
4
3
2
1
0
β-Actin
ACBP
β-Actin
ACBP
β-Actin
SCC
H520A B C
D E F
A549
H520
Absorb
ance
(4
90
nm
)
Absorb
an
ce
(4
90
nm
)
0 2 4
Days
Cntl vectorACBP shRNA1ACBP shRNA2
Cntl vector
Pos
Cntl
ACBP
shRNA1
ACBP
shRNA2
Cntl
vector
ACBP shRNA2
Total PARP
Cleaved PARP
Cleaved
caspase-3
β-Actin0 2 4 6
Days
H520A549
****
*
16-H
BE
Pos
Cntl
AC
BP
shR
NA
1A
CB
P s
hR
NA
2A
CB
P s
hR
NA
3C
ntl
vect
or
Pos
Cntl
AC
BP
shR
NA
1A
CB
P s
hR
NA
2A
CB
P s
hR
NA
3C
ntl
vect
or
H520
H157
H227
HC
C95
HC
C15
H23
H1648
Calu
-3A
549
H2882
H460
ADC SCC LCC
Figure 4. ACBP downregulation reduces proliferation and induces apoptosis. A, ACBP expression in a series of NSCLC cell lines. B and C, downregulation ofACBP expression by shRNA in H520 and A549, cells respectively. D and E,. time-dependent effect of downregulation of ACBP on proliferation of H520 andA549 cells, respectively. F, effect of ACBP downregulation in H520 cells on PARP (89 kDa) and caspase-3 cleavage (17 and 19 kDa) after 48 hours.Pos Cntl, positive control, Hep-G2 (B and C). F, untransfected H520 cells in puromycin media. �, P < 0.05; ��, P < 0.001. Error bars denote SD betweenreplicates. Data shown are a representative of 3 separate independent experiments.
Potential Role of ACBP in Tumor Progression in Lung Cells
www.aacrjournals.org Cancer Prev Res; 7(7) July 2014 753
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

designed to inhibit inflammation that could potentiallyinhibit substrate binding to the FABPs (36).
Similar to our assertion that ACBP is overexpressed inlung cancer and inflammatory cells, and associated withpoorer patient survival, in bladder cancer, a study suggeststhat ACBP could be predictive of poor survival and of poorresponse to chemotherapy (37). Lee and colleagues alsodemonstrated upregulation of the ACBP gene in their DNAmicroarray dataset of normal, neoplastic, and invasiveepithelial ovarian tissues (38). Using IMS, we had a uniqueopportunity to assess the expression of ACBP in a patientspecimen harboring various types of pre-invasive lesionsthat recapitulate the natural history of squamous carcinomaof the lung. We further validated ACBP expression in inde-pendent tissue specimens by IHC. These studies promptedus to further investigate the biologic implications of ACBPoverexpression in lung cancer.
We show that ACBP depletion reduces cell proliferationand induces apoptosis in the NSCLC cell lines tested. This
observation was strengthened as we demonstrated thatACBP mediates b-oxidation in lung cancer cells. b-Oxida-tion plays a crucial role in providing the acyl-CoA substratefor the b-oxidation pathway and controlling cell growth.b-Oxidation is the process by which fatty acids are brokendown in the mitochondria to generate acetyl-CoA. Theacetyl-CoA can serve as the substrate for the TCA cycle andas a substrate for fatty acid synthesis. One of the keyregulators of b-oxidation is CPT-1, for which ACBP servesas an endogenous ligand. CPT-1 mediates the transport ofacyl-CoAs into the mitochondria for b-oxidation. More-over, CPT-1 prefers the acyl-CoA complexed with ACBPsubstrate rather than free-acyl-CoAs (39). Pharmacologicinhibition of CPT-1 using the drug etomoxir results in ATPdepletion and cell death in glioblastomas and sensitizeshuman leukemia cells to apoptosis (40, 41). Recently,inhibition of CPT-1 protein levels were shown to reducecell growth in the in vitro models of lung, breast, andprostate cancers and in in vivomodels of breast cancer under
20,000
15,000
10,000
5,000
0
1.0
0.8
0.6
0.4
0.2
0.0
20,000
15,000
10,000
5,000
0
15,000
10,000
5,000
0
–5,000
H520A B C
D E FH520 H520
**
**
**
**
*
***
H52016-HBE
3H
-pa
lmita
te, C
PM
ACBP
shRNA1
ACBP
shRNA1
(–Glc)
Cntl
vector
0 2 4 6
Days
Absorb
ance (
49
0 n
m)
25
20
15
10
5
0
Mitochondri
al po
ten
tia
l, R
FU
ACBP
shRNA1
No
etomoxir
10 μmol/L
etomoxir
10 μmol/L etomoxir
0 mmol/L etomoxir
10 mmol/L etomoxir
100 mmol/L etomoxir
0 mmol/L etomoxir
10 mmol/L etomoxir
100 mmol/L etomoxir
100 μmol/L etomoxir
Cntl
vector
Cntl
10 μmol/L
etomoxir
100 μmol/L
etomoxir
Cntl
Cntl
vector
(–Glc)
Figure 5. ACBP downregulation decreases b-oxidation in H520 cells. A, downregulation of ACBP by shRNA inhibits b-oxidation in cancer cells. In theseexperiments, 3H-pamitate levels in cells expressing control vector (Cntl Vector) are compared with the ACBP shRNA. B, downregulation of ACBPhas no significant effect on b-oxidation in16-HBE cells. The results in A and B were normalized to protein levels. Glucose was removed (�Glc) in both controlvector and ACBP-depleted cells to ensure that we were measuring b-oxidation levels. C, decrease in b-oxidation levels using 10 mmol/L of CPT-1inhibitor, etomoxir, in H520 cells. D, dose-dependent effect of etomoxir on the proliferation in H520 cells. Proliferation wasmeasured by theMTS proliferationassay. E, dose-dependent increase of etomoxir on mitochondrial potential measured. Alterations in mitochondrial potential were measured by JC-1accumulation in H520 cells. F, qualitative assessment of mitochondrial injury asmeasured by JC-1 (red, healthy viable cells; green, unhealthy, early apoptoticcells). �, P < 0.05; ��, P < 0.001. Error bars denote SD between replicates. Data, representative of 3 separate, independent experiments. CPM, countsper minutes; RFU, relative fluorescent units.
Harris et al.
Cancer Prev Res; 7(7) July 2014 Cancer Prevention Research754
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

metabolic stressors (42). Our data suggest that the b-oxi-dation pathway is required to meet macromoleculardemands. It has been largely accepted that cancer cellsrequire fatty acid synthesis, coupled with the key enzymefatty acid synthase for growth and survival (43–47). It mayseem counterintuitive that cancer cells would use b-oxida-tion, a degradation pathway, during their rapid growth andexpansion. However, whenwe consider that the cancer cellscan quickly deplete other nutritional sources, such as glu-cose and glutamine, b-oxidation becomes an ideal sourcefor energy. A recent report demonstrates that b-oxidation isupregulated in alveolar cells isolated from mice exposed tocigarette smoke (48). There may be a truemetabolic shift inalveolar type II cells from glucose utilization (glycolysis) topalmitate utilization (b-oxidation) for energy productionafter cigarette smoke exposure. This is in line with ourhypothesis that b-oxidation, regulated by ACBP, is impor-tant for lung cancer growth.In summary, we identified a set of lung cancers over-
expressing ACBP that have worse overall survival implicat-ing ACBP in cancer progression. Our results suggest thatACBP plays a role in lung cancer growth and survival byregulating b-oxidation. Understanding of the role of ACBPin lung cancer progression could play a pivotal role in futurestudies investigating the ACBP/CPT-1/b-oxidation pathwayin controlling NSCLC pathophysiology and treatment.
Disclosure of Potential Conflicts of InterestR.M. Caprioli is the Editor-in-Chief of the Journal of Mass Spectrometry. No
potential conflicts of interest were disclosed by the other authors.
Authors' ContributionsConception and design: F.T. Harris, S.M.J. Rahman, M. Hassanein, J. Qian,R.M. Caprioli, P.P. MassionDevelopment of methodology: F.T. Harris, S.M.J. Rahman, M. Hassanein,P. Chaurand, M. ShiotaAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): F.T. Harris, S.M.J. Rahman, M.D. Hoeksema,P. Chaurand, M. Shiota, P.P. MassionAnalysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis): F.T. Harris, S.M.J. Rahman, M. Hassa-nein, J. Qian, H. Chen, P. Chaurand, P.P. MassionWriting, review, and or revision of the manuscript: F.T. Harris, S.M.J.Rahman, M. Hassanein, P. Chaurand, R.M. Caprioli, P.P. MassionAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): F.T. Harris, S.M.J. Rahman,M. ShiotaStudy supervision: F.T. Harris, S.M.J. Rahman, P.P. MassionReview and interpretation of pathologic material: R. Eisenberg
AcknowledgmentsThe authors thank Drs. LaMonica V. Stewart and Ifeanyi J. Arinze
(Meharry Medical College, Nashville, TN) for their helpful review of thearticle. They also thank Bradford K. Harris and Dr. Xiangming Ji (VanderbiltUniversity, Nashville, TN) for their technical assistance.
Grant SupportThis study was supported by NIH grants R25CA092043 (to F.T. Harris)
and R01-CA102353-05 and SPORE in Lung Cancer CA90949 (to P.P.Massion).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received February 18, 2014; revised April 18, 2014; acceptedMay 6, 2014;published OnlineFirst May 12, 2014.
ACBP
ATP
Cell survival
Cell
proliferation
Acyl-
CoA
Acety-CoA
TCA
cycle
Electron
transport
chain
Cytoplasm
Acyl-
CoA
CPT-1
Etomoxir
Mitochondria
β-oxidation
Figure 6. Schematic representationof the mechanism by which ACBPregulates b-oxidation and controlsgrowth and survival in lung cancer.ACBP serves to sequester the fattyacyl-CoAs. The ACBP/fatty acyl-CoA complex binds to CPT-1 totransport acyl-CoAs to the innermitochondrial membrane forb-oxidation. The product, acetyl-CoA, enters the TCA cycle and theresulting electrons can then enterthe electron transport channel forATP production. ATP productionregulates cellular proliferation andsurvival. ATP, adenosinetriphosphate; ROS, reactiveoxygen species.
Potential Role of ACBP in Tumor Progression in Lung Cells
www.aacrjournals.org Cancer Prev Res; 7(7) July 2014 755
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

References1. RahmanSM,Gonzalez AL, LiM, Seeley EH, Zimmerman LJ, Zhang XJ,
et al. Lung cancer diagnosis from proteomic analysis of preinvasivelesions. Cancer Res 2011;71:3009–17.
2. Rahman SM, Shyr Y, Yildiz PB, Gonzalez AL, Li H, Zhang X, et al.Proteomic patterns of preinvasive bronchial lesions. Am J Respir CritCare Med 2005;172:1556–62.
3. Rasmussen JT, Borchers T, Knudsen J. Comparison of the bindingaffinities of acyl-CoA-binding protein and fatty-acid-binding proteinfor long-chain acyl-CoA esters. Biochem J 1990;265:849–55.
4. Rosendal J, Ertbjerg P, Knudsen J. Characterization of ligand bindingto acyl-CoA-binding protein. Biochem J 1993;290:321–6.
5. Guidotti A, Forchetti CM, Corda MG, Konkel D, Bennett CD, Costa E.Isolation, characterization, and purification to homogeneity of anendogenous polypeptide with agonistic action on benzodiazepinereceptors. Proc Natl Acad Sci U S A 1983;80:3531–5.
6. Marquardt H, Todaro GJ, ShoyabM. Complete amino acid sequencesof bovine and human endozepines. Homology with rat diazepambinding inhibitor. J Biol Chem 1986;261:9727–31.
7. Guidotti A, Konkel DR, Ebstein B, Corda MG, Wise BC, Krutzsch H,et al. Isolation, characterization, and purification to homogeneity of arat brain protein (GABA-modulin). Proc Natl Acad Sci U S A 1982;79:6084–8.
8. Shoyab M, Gentry LE, Marquardt H, Todaro GJ. Isolation andcharacterization of a putative endogenous benzodiazepineoid(endozepine) from bovine and human brain. J Biol Chem 1986;261:11968–73.
9. Mogensen IB, Schulenberg H, Hansen HO, Spener F, Knudsen J. Anovel acyl-CoA-binding protein from bovine liver. Effect on fatty acidsynthesis. Biochem J 1987;241:189–92.
10. Yanagibashi K, Ohno Y, Kawamura M, Hall PF. The regulation ofintracellular transport of cholesterol in bovine adrenal cells: purificationof a novel protein. Endocrinology 1988;123:2075–82.
11. BesmanMJ, Yanagibashi K, Lee TD, KawamuraM, Hall PF, Shively JE.Identification of des-(Gly-Ile)-endozepine as an effector of corticotro-pin-dependent adrenal steroidogenesis: stimulation of cholesteroldelivery is mediated by the peripheral benzodiazepine receptor. ProcNatl Acad Sci U S A 1989;86:4897–901.
12. Chen ZW, Agerberth B, Gell K, Andersson M, Mutt V, Ostenson CG,et al. Isolation and characterization of porcine diazepam-bindinginhibitor, a polypeptide not only of cerebral occurrence but alsocommon in intestinal tissues and with effects on regulation of insulinrelease. Eur J Biochem 1988;174:239–45.
13. Hansen HO, Andreasen PH, Mandrup S, Kristiansen K, Knudsen J.Induction of acyl-CoA-binding protein and itsmRNA in 3T3-L1 cells byinsulin during preadipocyte-to-adipocyte differentiation. Biochem J1991;277:341–4.
14. Rasmussen JT, Faergeman NJ, Kristiansen K, Knudsen J. Acyl-CoA-binding protein (ACBP) can mediate intermembrane acyl-CoA trans-port and donate acyl-CoA for beta-oxidation and glycerolipid synthe-sis. Biochem J 1994;299:165–70.
15. Ward PS, Thompson CB. Metabolic reprogramming: a cancerhallmark even Warburg did not anticipate. Cancer Cell 2012;21:297–308.
16. Corda MG, Ferrari M, Guidotti A, Konkel D, Costa E. Isolation, puri-fication and partial sequence of a neuropeptide (diazepam-bindinginhibitor) precursor of an anxiogenic putative ligand for benzodiaze-pine recognition site. Neurosci Lett 1984;47:319–24.
17. Liu Y. Fatty acid oxidation is a dominant bioenergetic pathway inprostate cancer. Prostate Cancer Prostatic Dis 2006;9:230–4.
18. Cornett DS, Reyzer ML, Chaurand P, Caprioli RM: MALDI imagingmass spectrometry:molecular snapshots of biochemical systems.NatMethods 2007;4:828–33.
19. Chaurand P, Norris JL, Cornett DS, Mobley JA, Caprioli RM. Newdevelopments in profiling and imaging of proteins from tissuesections by MALDI mass spectrometry. J Proteome Res 2006;5:2889–900.
20. Stoeckli M, Chaurand P, Hallahan DE, Caprioli RM. Imaging massspectrometry: a new technology for the analysis of protein expressionin mammalian tissues. Nat Med 2001;7:493–6.
21. Hassanein M, Hoeksema MD, Shiota M, Qian J, Harris BK, Chen H,et al. SLC1A5 mediates glutamine transport required for lung cancercell growth and survival. Clin Cancer Res 2013;19:560–70.
22. Gyorffy B, Surowiak P, Budczies J, Lanczky A. Online survival analysissoftware to assess the prognostic value of biomarkers using tran-scriptomic data in non-small-cell lung cancer. PLoS ONE 2013;8:e82241.
23. WangJ,Qian J,HoeksemaMD,ZouY, EspinosaAV,RahmanSM, et al.Integrative genomics analysis identifies candidate drivers at 3q26–29amplicon in squamous cell carcinoma of the lung. Clin Cancer Res2013;19:5580–90.
24. Ocak S, Chen H, Callison C, Gonzalez AL, Massion PP. Expression offocal adhesion kinase in small-cell lung carcinoma. Cancer 2012;118:1293–301.
25. Egnatchik RA, Leamy AK, Noguchi Y, Shiota M, Young JD. Palmitate-induced activation of mitochondrial metabolism promotes oxidativestress and apoptosis in H4IIEC3 rat hepatocytes. Metabolism 2014;63:283–95.
26. Knudsen J, Mandrup S, Rasmussen JT, Andreasen PH, Poulsen F,Kristiansen K. The function of acyl-CoA-binding protein (ACBP)/diazepam binding inhibitor (DBI). Mol Cell Biochem 1993;123:129–38.
27. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation.Cell 2011;144:646–74.
28. Selby PL, Sherratt HS. Substituted 2-oxiranecarboxylic acids: a newgroup of candidate hypoglycaemic drugs. Trends Pharmacol Sci1989;10:495–500.
29. Reers M, Smiley ST, Mottola-Hartshorn C, Chen A, Lin M, Chen LB.Mitochondrial membrane potential monitored by JC-1 dye. MethodsEnzymol 1995;260:406–17.
30. Green DR, Reed JC. Mitochondria and apoptosis. Science 1998;281:1309–12.
31. Menendez JA, Lupu R. Fatty acid synthase and the lipogenic pheno-type in cancer pathogenesis. Nat Rev Cancer 2007;7:763–77.
32. Smathers RL, Petersen DR. The human fatty acid-binding proteinfamily: evolutionary divergences and functions. Hum Genomics2011;5:170–91.
33. Chmurzynska A. The multigene family of fatty acid-binding proteins(FABPs): function, structure and polymorphism. J Appl Genet 2006;47:39–48.
34. Kawamura T, Kanno R, Fujii H, Suzuki T. Expression of liver-type fatty-acid-binding protein, fatty acid synthase and vascular endothelialgrowth factor in human lung carcinoma. Pathobiology 2005;72:233–40.
35. Levi L, LoboG, DoudMK, von Lintig J, Seachrist D, Tochtrop GP, et al.Genetic ablation of the fatty acid-binding protein FABP5 suppressesHER2-induced mammary tumorigenesis. Cancer Res 2013;73:4770–80.
36. Berger WT, Ralph BP, Kaczocha M, Sun J, Balius TE, Rizzo RC, et al.Targeting fatty acid binding protein (FABP) anandamide transporters -a novel strategy for development of anti-inflammatory and anti-noci-ceptive drugs. PLoS ONE 2012;7:e50968.
37. Pollard C, Nitz M, Baras A, Williams P, Moskaluk C, Theodorescu D.Genoproteomic mining of urothelial cancer suggests {gamma}-gluta-myl hydrolase and diazepam-binding inhibitor as putative urinarymarkers of outcome after chemotherapy. Am J Pathol 2009;175:1824–30.
38. Lee BC, Cha K, Avraham S, Avraham HK. Microarray analysis ofdifferentially expressed genes associated with human ovarian cancer.Int J Oncol 2004;24:847–51.
39. Abo-HashemaKA, CakeMH, LukasMA, Knudsen J. The interaction ofacyl-CoA with acyl-CoA binding protein and carnitine palmitoyltrans-ferase I. Int J Biochem Cell Biol 2001;33:807–15.
40. Pike LS, Smift AL, Croteau NJ, Ferrick DA, Wu M. Inhibition of fattyacid oxidation by etomoxir impairs NADPH production andincreases reactive oxygen species resulting in ATP depletion andcell death in human glioblastoma cells. Biochim Biophys Acta 2011;1807:726–34.
41. Samudio I, Harmancey R, Fiegl M, Kantarjian H, KonoplevaM, KorchinB, et al. Pharmacologic inhibition of fatty acid oxidation sensitizes
Harris et al.
Cancer Prev Res; 7(7) July 2014 Cancer Prevention Research756
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

human leukemia cells to apoptosis induction. J Clin Invest 2010;120:142–56.
42. ZauggK,YaoY,Reilly PT,KannanK,KiarashR,MasonJ, et al. Carnitinepalmitoyltransferase 1C promotes cell survival and tumor growth underconditions of metabolic stress. Genes Dev 2011;25:1041–51.
43. Kuhajda FP. Fatty acid synthase and cancer: new application of an oldpathway. Cancer Res 2006;66:5977–80.
44. Menendez JA, Lupu R. Fatty acid synthase-catalyzed de novo fattyacid biosynthesis: from anabolic-energy-storage pathway in normaltissues to jack-of-all-trades in cancer cells. Arch Immunol Ther Exp2004;52:414–26.
45. Menendez JA, Lupu R. Oncogenic properties of the endogenous fattyacid metabolism: molecular pathology of fatty acid synthase in cancercells. Curr Opin Clin Nutr Metab Care 2006;9:346–57.
46. Swinnen JV, Brusselmans K, Verhoeven G. Increased lipogenesis incancer cells: new players, novel targets. Curr Opin Clin Nutr MetabCare 2006;9:358–65.
47. KuhajdaFP. Fatty-acid synthase andhumancancer: newperspectiveson its role in tumor biology. Nutrition 2000;16:202–8.
48. Agarwal AR, Yin F, Cadenas E. Short-term cigarette smoke exposureleads to metabolic alterations in lung alveolar cells. Am J Respir CellMol Biol 2014 Mar 13. [Epub ahead of print].
www.aacrjournals.org Cancer Prev Res; 7(7) July 2014 757
Potential Role of ACBP in Tumor Progression in Lung Cells
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057

2014;7:748-757. Published OnlineFirst May 12, 2014.Cancer Prev Res Fredrick T. Harris, S.M. Jamshedur Rahman, Mohamed Hassanein, et al.
Small Cell Lung Cancer−Required for Growth and Survival of Non Binding Protein Regulates Beta-Oxidation−Acyl-Coenzyme A
Updated version
10.1158/1940-6207.CAPR-14-0057doi:
Access the most recent version of this article at:
Material
Supplementary
1
http://cancerpreventionresearch.aacrjournals.org/content/suppl/2014/05/12/1940-6207.CAPR-14-0057.DCAccess the most recent supplemental material at:
Cited articles
http://cancerpreventionresearch.aacrjournals.org/content/7/7/748.full#ref-list-1
This article cites 47 articles, 17 of which you can access for free at:
Citing articles
http://cancerpreventionresearch.aacrjournals.org/content/7/7/748.full#related-urls
This article has been cited by 2 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerpreventionresearch.aacrjournals.org/content/7/7/748To request permission to re-use all or part of this article, use this link
Research. on May 18, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst May 12, 2014; DOI: 10.1158/1940-6207.CAPR-14-0057
http://cancerpreventionresearch.aacrjournals.org/content/suppl/2014/05/12/1940-6207.CAPR-14-0057.DC1
Related Documents











