Short Communication Acute exercise preferentially redeploys NK-cells with a highly-differentiated phenotype and augments cytotoxicity against lymphoma and multiple myeloma target cells. Part II: Impact of latent cytomegalovirus infection and catecholamine sensitivity Austin B. Bigley a , Katayoun Rezvani b , Mira Pistillo a , Justin Reed a , Nadia Agha a , Hawley Kunz a , Daniel P. O’Connor a , Takuya Sekine b , Catherine M. Bollard c , Richard J. Simpson a,⇑ a Laboratory of Integrated Physiology, Department of Health and Human Performance, University of Houston, 3855 Holman Street, Houston, TX 77204, USA b Department of Stem Cell Transplantation, Division of Cancer Medicine, The University of Texas MD Anderson Cancer Center, Houston, TX 77030, USA c Program for Cell Enhancement and Technologies for Immunotherapy, Sheikh Zayed Institute for Pediatric Surgical Innovation, and Center for Cancer and Immunology Research, Children’s National Health System, Washington, DC 20010, USA article info Article history: Received 8 November 2014 Received in revised form 23 December 2014 Accepted 31 December 2014 Available online xxxx Keywords: Immunology NKG2C CD57 Isoproterenol Cyclic AMP Beta adrenergic receptor abstract We showed previously that acute exercise is associated with a preferential redeployment of highly-dif- ferentiated NK-cells and increased cytotoxicity against HLA-expressing tumor cell lines during exercise recovery. In this part II study, we retrospectively analyzed these findings in the context of latent cyto- megalovirus (CMV) infection and performed additional experiments to explore potential mechanisms underpinning the marked reduction in NK-cell redeployment with exercise in CMV-seropositive individ- uals. We show here that latent CMV infection impairs NK-cell mobilization with exercise, only when the intensity of the exercise bout exceeds the individual blood lactate threshold (BLT). This impaired mobi- lization is associated with increased proportions of poorly exercise-responsive NK-cell subsets (NKG2C+/ KIRÀ, NKG2C+/NKG2AÀ, and NKG2C+/CD57+) and decreased NK-cell b 2 -adrenergic receptor (AR) expres- sion in those with CMV. As a result, NK-cell production of cyclic AMP (cAMP) in response to in vitro iso- proterenol (synthetic b-agonist) stimulation was drastically lower in those with CMV (6.0 vs. 20.3 pmol/ mL, p < 0.001) and correlated highly with the proportion of NKG2C+/CD57+ NK-cells (R 2 = 0.97). More- over, NK-cell cytotoxic activity (NKCA) against the K562 (36.6% vs. 22.7%, p < 0.05), U266 (23.6% vs. 15.9%, p < 0.05), and 221.AEH (41.3% vs. 13.3%, p < 0.001) cell lines was increased at baseline in those infected with CMV; however, latent CMV infection abated the post-exercise increase in NKCA as a result of decreased NK-cell mobilization. Additionally, NKCA per cell against the U266 (0.24 vs. 0.12, p < 0.01), RPMI-8226 (0.17 vs. 0.11, p < 0.05), and 221.AEH (0.18 vs. 0.11, p < 0.05) cell lines was increased 1 h post- exercise (relative to baseline) in CMV-seronegative subjects, but not in those infected with CMV. Collec- tively, these data indicate that latent CMV infection may compromise NK-cell mediated immunosurveil- lance after acute exercise due to an increased proportion of ‘‘CMV-specific’’ NK-cell subsets with impaired b-adrenergic receptor signaling pathways. Ó 2015 Elsevier Inc. All rights reserved. 1. Introduction NK-cells are rapidly redeployed between the blood and tissues in response to a single bout of dynamic exercise. We showed in our Part I paper that the redeployment of NK-cell subsets is non- uniform (Supplementary Table 1); consistently evoking a stepwise redeployment of NK-cell subsets in accordance with differentiation status [highly-differentiated (KIR+/NKG2AÀ) > medium-differenti- ated (KIR+/NKG2A+) > low-differentiated (KIRÀ/NKG2A+)] across multiple exercise intensities (Bigley et al., 2014a). NK-cells expressing the putative terminal differentiation marker CD57 (Lopez-Verges et al., 2010) were also preferentially redeployed after exercise, while NK-cells expressing the activating receptor NKG2C responded poorly to exercise (Bigley et al., 2014a). These proportional changes in NK-cell subsets were associated with enhanced NKCA against HLA-expressing tumor cell lines during exercise recovery (Bigley et al., 2014a). In this Part II study, we http://dx.doi.org/10.1016/j.bbi.2014.12.027 0889-1591/Ó 2015 Elsevier Inc. All rights reserved. ⇑ Corresponding author. Tel.: +1 (713) 743 9270; fax: +1 (713) 743 9860. E-mail address: [email protected] (R.J. Simpson). Brain, Behavior, and Immunity xxx (2015) xxx–xxx Contents lists available at ScienceDirect Brain, Behavior, and Immunity journal homepage: www.elsevier.com/locate/ybrbi Please cite this article in press as: Bigley, A.B., et al. Acute exercise preferentially redeploys NK-cells with a highly-differentiated phenotype and augments cytotoxicity against lymphoma and multiple myeloma target cells. Part II: Impact of latent cytomegalovirus infection and catecholamine sensitivity. Brain Behav. Immun. (2015), http://dx.doi.org/10.1016/j.bbi.2014.12.027

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Brain, Behavior, and Immunity xxx (2015) xxx–xxx
Contents lists available at ScienceDirect
Brain, Behavior, and Immunity
journal homepage: www.elsevier .com/locate /ybrbi
Short Communication
Acute exercise preferentially redeploys NK-cells witha highly-differentiated phenotype and augments cytotoxicityagainst lymphoma and multiple myeloma target cells. Part II:Impact of latent cytomegalovirus infection and catecholamine sensitivity
http://dx.doi.org/10.1016/j.bbi.2014.12.0270889-1591/� 2015 Elsevier Inc. All rights reserved.
⇑ Corresponding author. Tel.: +1 (713) 743 9270; fax: +1 (713) 743 9860.E-mail address: [email protected] (R.J. Simpson).
Please cite this article in press as: Bigley, A.B., et al. Acute exercise preferentially redeploys NK-cells with a highly-differentiated phenotype and aucytotoxicity against lymphoma and multiple myeloma target cells. Part II: Impact of latent cytomegalovirus infection and catecholamine sensitivitBehav. Immun. (2015), http://dx.doi.org/10.1016/j.bbi.2014.12.027
Austin B. Bigley a, Katayoun Rezvani b, Mira Pistillo a, Justin Reed a, Nadia Agha a, Hawley Kunz a,Daniel P. O’Connor a, Takuya Sekine b, Catherine M. Bollard c, Richard J. Simpson a,⇑a Laboratory of Integrated Physiology, Department of Health and Human Performance, University of Houston, 3855 Holman Street, Houston, TX 77204, USAb Department of Stem Cell Transplantation, Division of Cancer Medicine, The University of Texas MD Anderson Cancer Center, Houston, TX 77030, USAc Program for Cell Enhancement and Technologies for Immunotherapy, Sheikh Zayed Institute for Pediatric Surgical Innovation, and Center for Cancer and ImmunologyResearch, Children’s National Health System, Washington, DC 20010, USA
a r t i c l e i n f o
Article history:Received 8 November 2014Received in revised form 23 December 2014Accepted 31 December 2014Available online xxxx
Keywords:ImmunologyNKG2CCD57IsoproterenolCyclic AMPBeta adrenergic receptor
a b s t r a c t
We showed previously that acute exercise is associated with a preferential redeployment of highly-dif-ferentiated NK-cells and increased cytotoxicity against HLA-expressing tumor cell lines during exerciserecovery. In this part II study, we retrospectively analyzed these findings in the context of latent cyto-megalovirus (CMV) infection and performed additional experiments to explore potential mechanismsunderpinning the marked reduction in NK-cell redeployment with exercise in CMV-seropositive individ-uals. We show here that latent CMV infection impairs NK-cell mobilization with exercise, only when theintensity of the exercise bout exceeds the individual blood lactate threshold (BLT). This impaired mobi-lization is associated with increased proportions of poorly exercise-responsive NK-cell subsets (NKG2C+/KIR�, NKG2C+/NKG2A�, and NKG2C+/CD57+) and decreased NK-cell b2-adrenergic receptor (AR) expres-sion in those with CMV. As a result, NK-cell production of cyclic AMP (cAMP) in response to in vitro iso-proterenol (synthetic b-agonist) stimulation was drastically lower in those with CMV (6.0 vs. 20.3 pmol/mL, p < 0.001) and correlated highly with the proportion of NKG2C+/CD57+ NK-cells (R2 = 0.97). More-over, NK-cell cytotoxic activity (NKCA) against the K562 (36.6% vs. 22.7%, p < 0.05), U266 (23.6% vs.15.9%, p < 0.05), and 221.AEH (41.3% vs. 13.3%, p < 0.001) cell lines was increased at baseline in thoseinfected with CMV; however, latent CMV infection abated the post-exercise increase in NKCA as a resultof decreased NK-cell mobilization. Additionally, NKCA per cell against the U266 (0.24 vs. 0.12, p < 0.01),RPMI-8226 (0.17 vs. 0.11, p < 0.05), and 221.AEH (0.18 vs. 0.11, p < 0.05) cell lines was increased 1 h post-exercise (relative to baseline) in CMV-seronegative subjects, but not in those infected with CMV. Collec-tively, these data indicate that latent CMV infection may compromise NK-cell mediated immunosurveil-lance after acute exercise due to an increased proportion of ‘‘CMV-specific’’ NK-cell subsets with impairedb-adrenergic receptor signaling pathways.
� 2015 Elsevier Inc. All rights reserved.
1. Introduction
NK-cells are rapidly redeployed between the blood and tissuesin response to a single bout of dynamic exercise. We showed inour Part I paper that the redeployment of NK-cell subsets is non-uniform (Supplementary Table 1); consistently evoking a stepwiseredeployment of NK-cell subsets in accordance with differentiation
status [highly-differentiated (KIR+/NKG2A�) > medium-differenti-ated (KIR+/NKG2A+) > low-differentiated (KIR�/NKG2A+)] acrossmultiple exercise intensities (Bigley et al., 2014a). NK-cellsexpressing the putative terminal differentiation marker CD57(Lopez-Verges et al., 2010) were also preferentially redeployedafter exercise, while NK-cells expressing the activating receptorNKG2C responded poorly to exercise (Bigley et al., 2014a). Theseproportional changes in NK-cell subsets were associated withenhanced NKCA against HLA-expressing tumor cell lines duringexercise recovery (Bigley et al., 2014a). In this Part II study, we
gmentsy. Brain

2 A.B. Bigley et al. / Brain, Behavior, and Immunity xxx (2015) xxx–xxx
retrospectively analyzed our previously acquired data (Bigley et al.,2014a) in relation to CMV serostatus and performed additionalexperiments (plasma catecholamine and cortisol response to exer-cise and NK-cell b-AR expression/sensitivity) to provide insightinto potential mechanisms for the blunting effect of CMV on theredeployment of NK-cells with exercise (Bigley et al., 2012).
The aims of this study were twofold. First, we examined theimpact of exercise intensity and b-AR expression/sensitivity onthe known abating effects of latent CMV infection on the redeploy-ment of NK-cells after exercise (Bigley et al., 2012). Second, weexamined the impact of latent CMV infection on NKCA against tar-get cells with varying degrees of HLA expression before and afterexercise. We show that the mobilization of NK-cells in responseto exercise above BLT is lower in individuals with CMV and is asso-ciated with decreased NK-cell b-AR signaling. We also show thatlatent CMV infection abates the functional responses of NK-cellsto exercise. Overall, our findings indicate that latent CMV infectionmay be associated with impaired NK-cell immunosurveillance fol-lowing an acute bout of exercise.
2. Methods
2.1. Subjects
Data from twelve male and four female (n = 16) healthy,trained, non-smoking cyclists (age: 32.1 ± 4.3 year) who partici-pated in our part I study (Bigley et al., 2014a) was retrospectivelyanalyzed relative to CMV serostatus. All potential subjects wereassessed for serum anti-CMV IgG1 antibody titers prior to partici-pating in the study to ensure equal numbers of CMVpos and CMVneg
subjects. Subject physical characteristics and exercise performancemeasures contrasted by CMV serostatus are shown in Supplemen-tary Tables 2 and 3, respectively. Additional test subjects (n = 6,age: 34.0 ± 5.2 year), meeting the same inclusion/exclusion criteriadescribed for the exercise trials and counterbalanced for CMV IgGserostatus were recruited to determine the impact of latent CMVinfection on NK-cell b-AR signaling. Two subjects from our Part Ipaper who only had NKCA measured against K562 were retestedto determine NKCA against 721.221, 221.AEH, U266, and RPMI-8226 cells. The study protocol was approved by the Committeefor the Protection of Human Subjects (CPHS) at the University ofHouston.
2.2. Exercise trials and CMV serostatus determination
The exercise protocols are described in detail in our Part I paper(Bigley et al., 2014a). Briefly, subjects underwent an incremental,discontinuous lactate threshold test using their personal road bicy-cle mounted on an indoor cycling ergometer (Computrainer, Racer-Mate Inc., Seattle WA). The main exercise trials consisted of threeseparate bouts of 30-min steady state cycling exercise at poweroutputs corresponding to �5%, +5%, and +15% of the individualBLT. Intravenous blood samples were collected before, immedi-ately after, and 1 h after exercise for each trial. Serum sampleswere frozen at �80 �C until analyses for CMV IgG antibodies (Gen-Way Biotech, San Diego, CA, USA), epinephrine/norepinephrine(LDN, Nordhorn, Germany), and cortisol (Abcam, Cambridge, UK)were performed using commercially available ELISA kits and a96-well microplate reader (Molecular Devices, Sunnyvale, CA,USA) in accordance with manufacturer instructions.
2.3. Flow cytometry
Whole blood was labeled with monoclonal antibodies and NK-cell phenotypes were assessed on a BD Accuri C6 flow cytometer
Please cite this article in press as: Bigley, A.B., et al. Acute exercise preferentiallcytotoxicity against lymphoma and multiple myeloma target cells. Part II: ImpaBehav. Immun. (2015), http://dx.doi.org/10.1016/j.bbi.2014.12.027
(Ann Arbor, MI, USA) by four-color flow cytometry as previouslydescribed (Bigley et al., 2014a). The lymphocyte count was deter-mined using a whole blood flow cytometric procedure (Bigleyet al., 2012). A four-color flow cytometric cytotoxicity assay wasused to quantify NK-cell killing of U266, RPMI-8226, 721.221,221.AEH, and K562 cells at 4 distinct NK-cell doses (1:1, 5:1,10:1, and 20:1 E:T) (Bigley et al., 2014a). The flow cytometryreagents are described in Bigley et al. (2014a) except for the PE-conjugated anti-b2-AR monoclonal antibody, which was purchasedfrom Santa Cruz Biotechnology (Dallas, TX, USA).
2.4. NK-cell catecholamine sensitivity assay
b-AR sensitivity was determined using magnetically sortedCD3�/CD56+ NK-cells (purity: 96 ± 1% NK-cells) obtained fromresting blood samples. First, PBMCs were negatively sorted usingCD3 MicroBeads (Miltenyi Biotec, Bergisch Gladbach, Germany)and then the CD3-depleted cells were positively sorted usingCD56 MicroBeads (Miltenyi). NK-cells were suspended in cold Dul-becco’s modified Eagle’s medium (DMEM) and incubations wereperformed in duplicate by adding 0.1 mL of 1.5, 3, 6, or 12x105 coldcells to 0.9 mL of 37 �C DMEM. Half of the tubes also contained10 lmol/L isoproterenol (Sigma–Aldrich, St. Louis, MO, USA). After5-min, the samples were frozen instantly in liquid Nitrogen to ter-minate the reaction (Benschop et al., 1994) and stored at �80 �Cuntil further use. Prior to determination of cAMP content, the sam-ples were heated in a boiling water bath for 5 min. The concentra-tion of cAMP was quantified using a cAMP ELISA kit (Enzo LifeSciences, Farmingdale, NY, USA) following the manufacturer’sinstructions.
2.5. Statistical analysis
Statistical analyses were performed using SPSS version 22 (Chi-cago, IL, USA). Two-way Repeated Measures ANOVA was used tocontrast physiological changes between the three exercise condi-tions (�5%, +5%, and +15% of BLT) in relation to CMV serostatus.One-way ANOVA was used to ensure there was no associationbetween CMV and subject characteristics. To examine the effectof CMV on NK-cell subset proportions and absolute counts, a max-imum likelihood linear mixed model (LMM) was built thatincluded main effects for CMV serostatus, time (pre, post, and 1 hpost-exercise), and exercise intensity. The impact of CMV and highNKG2C+ NK-cell proportion on exercise-induced NK-cell mobiliza-tion (% change, pre- to post-exercise) and egress (% change post- to1 h post-exercise) was determined using a LMM that includedmain effects for CMV/NKG2C status and exercise intensity, andan interaction effect between CMV/NKG2C status and exerciseintensity. A high NKG2C proportion was defined as a percentageof NKG2C+ NK-cells greater than the upper limit of the 95% confi-dence interval for CMV-seronegative subjects (16.4%). To examinethe effect of CMV on changes in NKCA with exercise, a LMM wasbuilt that included main effects for CMV serostatus, time, and dose(1�, 5�, 10�, and 20�), and an interaction effect of CMV serostatusx time. If the main effects of time or dose were significant, Bonfer-roni post hoc analysis was performed to determine the location ofthe significant effects. The effect of CMV on changes in plasma cat-echolamine concentrations with exercise was determined using aLMM that included main effects for CMV serostatus and time,and an interaction effect of CMV serostatus x time. To examinethe effect of CMV on NK-cell sensitivity to isoproterenol stimula-tion, a LMM was built that included main effects for CMV serosta-tus and dose, and an interaction effect of CMV serostatus � dose. Asimple regression model was used to determine the square of thecorrelation between isoproterenol-induced cAMP production andthe proportion of NKG2C+/CD57+ NK-cells. Statistical significance
y redeploys NK-cells with a highly-differentiated phenotype and augmentsct of latent cytomegalovirus infection and catecholamine sensitivity. Brain
A.B. Bigley et al. / Brain, Behavior, and Immunity xxx (2015) xxx–xxx 3
was taken at p < 0.05, except for the isoproterenol stimulationexperiment where a p-threshold of 0.001 was used due to low sam-ple size.
3. Results
3.1. CMV serostatus blunts the mobilization of NK-cells in response toexercise intensities exceeding BLT through accumulation of poorlyexercise responsive NK-cells
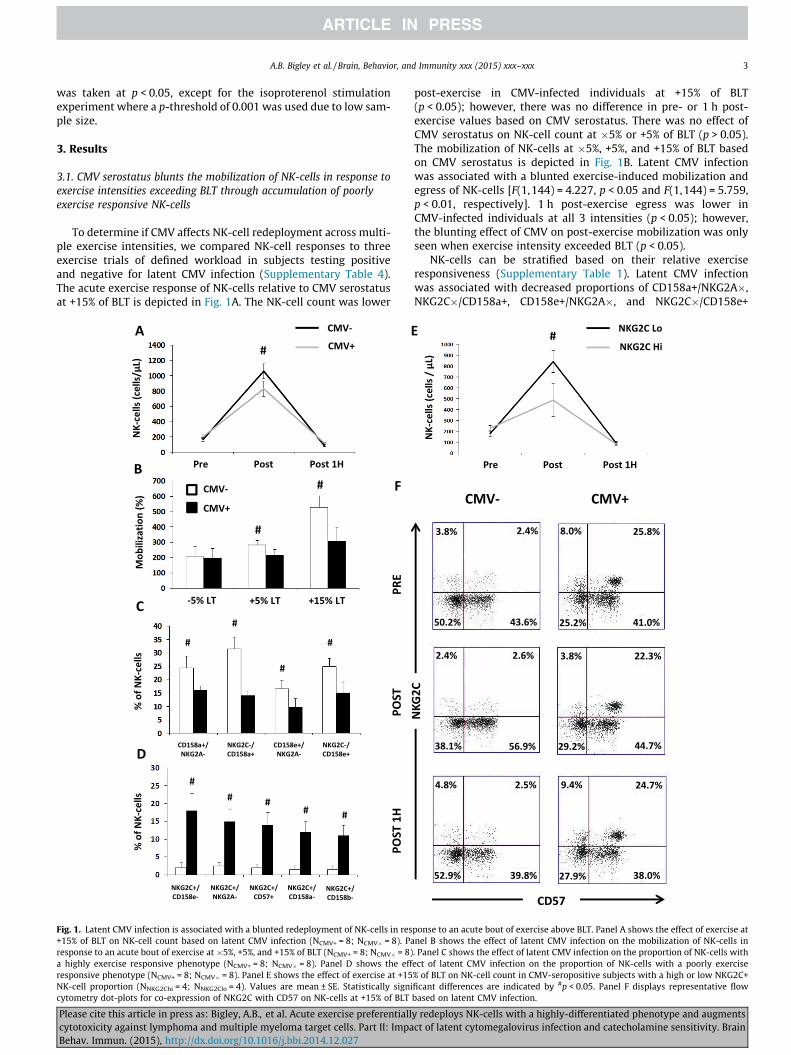
To determine if CMV affects NK-cell redeployment across multi-ple exercise intensities, we compared NK-cell responses to threeexercise trials of defined workload in subjects testing positiveand negative for latent CMV infection (Supplementary Table 4).The acute exercise response of NK-cells relative to CMV serostatusat +15% of BLT is depicted in Fig. 1A. The NK-cell count was lower
Fig. 1. Latent CMV infection is associated with a blunted redeployment of NK-cells in res+15% of BLT on NK-cell count based on latent CMV infection (NCMV+ = 8; NCMV� = 8). Paresponse to an acute bout of exercise at �5%, +5%, and +15% of BLT (NCMV+ = 8; NCMV� = 8)a highly exercise responsive phenotype (NCMV+ = 8; NCMV� = 8). Panel D shows the effresponsive phenotype (NCMV+ = 8; NCMV� = 8). Panel E shows the effect of exercise at +15NK-cell proportion (NNKG2Chi = 4; NNKG2Clo = 4). Values are mean ± SE. Statistically signicytometry dot-plots for co-expression of NKG2C with CD57 on NK-cells at +15% of BLT
Please cite this article in press as: Bigley, A.B., et al. Acute exercise preferentiallcytotoxicity against lymphoma and multiple myeloma target cells. Part II: ImpaBehav. Immun. (2015), http://dx.doi.org/10.1016/j.bbi.2014.12.027
post-exercise in CMV-infected individuals at +15% of BLT(p < 0.05); however, there was no difference in pre- or 1 h post-exercise values based on CMV serostatus. There was no effect ofCMV serostatus on NK-cell count at �5% or +5% of BLT (p > 0.05).The mobilization of NK-cells at �5%, +5%, and +15% of BLT basedon CMV serostatus is depicted in Fig. 1B. Latent CMV infectionwas associated with a blunted exercise-induced mobilization andegress of NK-cells [F(1,144) = 4.227, p < 0.05 and F(1,144) = 5.759,p < 0.01, respectively]. 1 h post-exercise egress was lower inCMV-infected individuals at all 3 intensities (p < 0.05); however,the blunting effect of CMV on post-exercise mobilization was onlyseen when exercise intensity exceeded BLT (p < 0.05).
NK-cells can be stratified based on their relative exerciseresponsiveness (Supplementary Table 1). Latent CMV infectionwas associated with decreased proportions of CD158a+/NKG2A�,NKG2C�/CD158a+, CD158e+/NKG2A�, and NKG2C�/CD158e+
ponse to an acute bout of exercise above BLT. Panel A shows the effect of exercise atnel B shows the effect of latent CMV infection on the mobilization of NK-cells in. Panel C shows the effect of latent CMV infection on the proportion of NK-cells withect of latent CMV infection on the proportion of NK-cells with a poorly exercise% of BLT on NK-cell count in CMV-seropositive subjects with a high or low NKG2C+ficant differences are indicated by #p < 0.05. Panel F displays representative flowbased on latent CMV infection.
y redeploys NK-cells with a highly-differentiated phenotype and augmentsct of latent cytomegalovirus infection and catecholamine sensitivity. Brain

4 A.B. Bigley et al. / Brain, Behavior, and Immunity xxx (2015) xxx–xxx
NK-cells (p < 0.05) (Fig. 1C), and increased proportions of NKG2C+/CD158e�, NKG2C+/NKG2A�, NKG2C+/CD57+, NKG2C+/CD158a�,and NKG2C+/CD158b� NK-cells (p < 0.001) (Fig. 1D). The effect ofNKG2C+ NK-cell proportion on NK-cell redeployment at +15% ofBLT is depicted in Fig. 1E. In CMV-seropositive subjects, a high pro-portion of NKG2C+ NK-cells was associated with reduced exercise-induced mobilization and egress of NK-cells at all three intensities[F(1,48) = 21.42, p < 0.001 and F(1,48) = 24.43, p < 0.001, respec-tively]; however, the attenuating effect of NKG2C+ NK-cell propor-tion on NK-cell mobilization grew with increasing exerciseintensity [F(2,48) = 3.78, p < 0.05]. Representative flow cytometrydot-plots illustrating the CMV-driven accumulation of poorly exer-cise responsive NKG2C+/CD57+ NK-cells are shown in Fig. 1F.
3.2. Latent CMV infection impairs NK-cell sensitivity to b-adrenergicstimulation in association with decreased b2-AR expression
To assess the effect of CMV on blood catecholamine and gluco-corticoid responses to exercise, we evaluated the acute exerciseresponse of plasma epinephrine, norepinephrine, and cortisol con-centrations relative to CMV serostatus (Fig. 2A). There was no maineffect of latent CMV infection on epinephrine, norepinephrine, orcortisol concentration [F(1,24) = 0.01, p = 0.99; F(1,24) = 0.23,p = 0.64; and F(1,24) = 0.44, p = 0.51, respectively]. The concentra-tion of epinephrine and norepinephrine increased with exercise at+15% of BLT [F(1,24) = 11.82, p < 0.01 and F(1,24) = 39.03,p < 0.001, respectively] independently of CMV [F(1,24) = 0.03,p = 0.855 and F(1,24) = 0.06, p = 0.815, respectively]. We next eval-uated if CMV was associated with a lower expression of b2-AR onthe surface of total and CD57+ NK-cells (Fig. 2B). The percentageof b2-ARpos NK-cells [F(1,15) = 5.37, p < 0.05] and the expression(GMFI) of b2-AR on total and CD57+ NK-cells was lower in those
Epi
neph
rine
(mM
)
Nor
epin
ephr
ine
(mM
)
Pre
cA
MP
(pm
ol/m
L)
A
C
# #
#
#
150 300 600 1,200
D
NKG2 103 NK-cells / mL
*
Pre Post
*
CMV-
CMV+
0 10
R2
cA
MP
(pm
ol/m
L)
Fig. 2. Latent CMV infection inhibits b-AR sensitivity of NK-cells through downregulnorepinephrine concentrations in relation to CMV serostatus (NCMV+ = 8; NCMV� = 8). P(NCMV+ = 8; NCMV� = 8). Panel C shows the blunted isoproterenol (10 lmol/L) response ocorrelation between isoproterenol-induced cAMP production (at an NK-cell dose ofNCMV� = 3). Values are mean ± SE. Statistically significant differences based on CMV serospost-exercise values are indicated by *p < 0.05. Panel E displays a representative flow cy
Please cite this article in press as: Bigley, A.B., et al. Acute exercise preferentiallcytotoxicity against lymphoma and multiple myeloma target cells. Part II: ImpaBehav. Immun. (2015), http://dx.doi.org/10.1016/j.bbi.2014.12.027
infected with CMV [F(1,15) = 6.25, p < 0.05 and F(1,15) = 6.10,p < 0.05, respectively]. We also evaluated isoproterenol-inducedcAMP production by NK-cells relative to CMV serostatus and theproportion of NKG2C+/CD57+ NK-cells. Latent CMV infection wasassociated with a markedly decreased isoproterenol-induced cAMPproduction by NK-cells at all 4 doses [F(1,24) = 24.869, p < 0.001](Fig. 2C). The proportion of NKG2C+/CD57+ NK-cells was stronglycorrelated (R2 = 0.97) with isoproterenol-induced cAMP (Fig. 2D).A representative flow cytometry histogram that illustrates NK-cellb2-AR expression is shown in Fig. 2E.
3.3. NKCA per cell against HLA-expressing tumor cell lines increasesduring exercise recovery in CMV-seronegative individuals only
The effect of CMV serostatus on NKCA (specific lysis) at +15% ofBLT is depicted in Fig. 3A. The dose response for the NK-cell assay ispresented in Supplementary Table 5. There was a main effect ofCMV on NKCA against K562 [F(1,192) = 6.33, p < 0.05], 221.AEH[F(1,120) = 10.98, p < 0.01], and RPMI-8226 cells [F(1,120) = 8.95,p < 0.01] but not 721.221 [F(1,120) = 0.01, p = 0.937] or U266[F(1,120) = 0.02, p = 0.901] cells. Latent CMV infection was associ-ated with increased pre-exercise NKCA against K562 and U266cells [F(1,64) = 4.46, p < 0.05 and F(1,40) = 5.29, p < 0.05, respec-tively], and decreased post-exercise NKCA against 721.221 cells[F(1,40) = 4.24, p < 0.05]. Latent CMV infection was associated withincreased NKCA against 221.AEH cells and decreased NKCA againstRPMI-8226 cells at all three time points (p < 0.05).
The effect of CMV serostatus on NKCA per cell at +15% of BLT isdepicted in Fig. 3B. There was a main effect of CMV on NKCA per cellagainst K562 [F(1,192) = 9.40, p < 0.01], 221.AEH [F(1,120) = 8.00,p < 0.01], and RPMI-8226 [F(1,120) = 19.02, p < 0.001] cells, but not721.221 [F(1,120) = 1.77, p = 0.188] or U266 [F(1,120) = 3.01,
Post
β2-
AR
Expr
essi
on (G
MFI
)
CMV+ CMV-
B
C+/CD57+ (%)
# * *
CMV-
CMV+
20 30 40 50
= 0.97
13.1%
β2-AR
E
ation of b2-AR. Panel A shows the effect of exercise on serum epinephrine andanel B shows the effect of latent CMV infection on b2-AR expression by NK-cellsf NK-cells in CMV-seropositive subjects (NCMV+ = 3; NCMV� = 3). Panel D shows the
6 � 105 NK-cells/mL) and the proportion of NKG2C+/CD57+ NK-cells (NCMV+ = 3;tatus are indicated by #p < 0.05. Statistically significant differences from pre- and 1 htometry histogram of NK-cell b2-AR expression.
y redeploys NK-cells with a highly-differentiated phenotype and augmentsct of latent cytomegalovirus infection and catecholamine sensitivity. Brain

K562 U266 721.221 221 AEH
Spec
ific
Lysi
s (%
)
A
B
PI
CD71
CMV
+CM
V-
PRE POST POST 1H
C i
221 AEH
Pre Post 1H
#
Pre Post 1H
#
Pre Post 1H
#
Pre Post 1H
# #
Pre Post 1H Pre Post 1H Pre Post 1H Pre Post 1H
# # # #
#* #
NKC
A p
er c
ell
20.7% 29.7% 12.4%
8.5% 19.6% 12.5%
CMV+
CMV-
#
#
#
*
RPMI-8226
Pre Post 1H
# #
#
Pre Post 1H
# #*
Pre Post 1H
NKG
2C+/
NKG
2A- (
%)
D
#
# #
*
CMV+
CMV-
Fig. 3. Latent CMV infection abrogates the 1 h post-exercise increase in NKCA per cell against the U266, RPMI-8226, and 221.AEH cell lines. Panel A shows the effect of latentCMV infection on specific lysis (%) of K562 (NCMV+ = 8; NCMV� = 8), U266 (NCMV+ = 5; NCMV� = 5), 721.221 (NCMV+ = 5; NCMV� = 5), 221.AEH (NCMV+ = 5; NCMV� = 5), and RPMI-8226 cells (NCMV+ = 5; NCMV� = 5) at a 20:1 effector-to-target cell ratio before, immediately after, and 1-hour after exercise at +15% of BLT. Panel B shows the effect of latentCMV infection on the number of dead K562 (NCMV+ = 8; NCMV� = 8), U266 (NCMV+ = 5; NCMV� = 5), 721.221 (NCMV+ = 5; NCMV� = 5), 221.AEH (NCMV+ = 5; NCMV� = 5), and RPMI-8226 cells (NCMV+ = 5; NCMV� = 5) per NK-cell (NKCA per cell) before, immediately after, and 1-hour after exercise at +15% of BLT (given a suspension of 10,000 target cells witha 20:1 effector to target cell ratio). Panel C shows the effect of latent CMV infection on the proportion of NKG2C+/NKG2A� NK-cells at +15% of BLT (NCMV+ = 8; NCMV� = 8).Values are mean ± SE. Statistically significant differences based on CMV status are indicated by #p < 0.05. Statistically significant differences from pre- and post-exercisevalues are indicated by *p < 0.05. Panel D displays representative flow cytometry dot-plots illustrating NKCA against 221.AEH cells at +15% of BLT.
A.B. Bigley et al. / Brain, Behavior, and Immunity xxx (2015) xxx–xxx 5
p = 0.086] cells. In addition, there were interaction effects betweenCMV and time for NKCA per cell against U266 [F(2,120) = 3.214,p < 0.05], 221.AEH [F(2,120) = 3.432, p < 0.05], and RPMI-8226 cells[F(2,120) = 3.565, p < 0.05]. In CMV-seronegative subjects, NKCA percell was greater 1 h post-exercise than pre- or post-exercise forU266, 221.AEH, and RPMI-8226 cells (p < 0.05). Latent CMV infectionwas associated with increased pre-exercise and decreased 1 h post-exercise NKCA per cell against U266 cells [F(1,40) = 4.151, p < 0.05and F(1,40) = 7.676, p < 0.01, respectively], while NKCA per cellagainst RPMI-8226 cells was decreased pre- and 1 h post-exercise[F(1,40) = 9.111, p < 0.01 and F(1,40) = 9.989, p < 0.01, respectively].Latent CMV infection was associated with increased NKCA per cellagainst 221.AEH and K562 cells at all three time points (p < 0.05).The effect of CMV serostatus on the proportion of NKG2C+/NKG2A�NK-cells at +15% of BLT is depicted in Fig. 3C. The proportion ofNKG2C+/NKG2A� NK-cells was greater in CMV-seropositive sub-jects (p < 0.05); however, only CMV-seronegative subjects showeda 1 h post-exercise increase in NKG2C+/NKG2A�NK-cell proportion(p < 0.05). Representative flow cytometry dot-plots illustrating the
Please cite this article in press as: Bigley, A.B., et al. Acute exercise preferentiallcytotoxicity against lymphoma and multiple myeloma target cells. Part II: ImpaBehav. Immun. (2015), http://dx.doi.org/10.1016/j.bbi.2014.12.027
effects of latent CMV infection and exercise on NKCA are shown inFig. 3D.
There were no statistically significant interaction effectsbetween sex (male or female) and CMV or sex and exercise forany of the immune parameters measured in this study (p > 0.05).
4. Discussion
In this study, we report that latent CMV infection profoundlyimpairs NK-cell redeployment and functional responses to high-intensity exercise. The CMV-driven accumulation of NKG2C+ NK-cells was associated with a marked reduction in NK-cell b-AR sig-naling and impaired mobilization of NK-cells in response to exer-cise, but only when the intensity of exercise exceeded theindividual BLT. Latent CMV infection was also associated withmarked alterations in baseline NK-cell cytotoxicity and NK-cellfunctional responses to exercise. NKCA against K562, U266, and221.AEH cells was increased at baseline in those infected withCMV; however, latent CMV infection abated the post-exercise
y redeploys NK-cells with a highly-differentiated phenotype and augmentsct of latent cytomegalovirus infection and catecholamine sensitivity. Brain

6 A.B. Bigley et al. / Brain, Behavior, and Immunity xxx (2015) xxx–xxx
increase in NKCA due to decreased NK-cell mobilization. Addition-ally, NKCA per cell against U266, RPMI-8226, and 221.AEH cellswas increased 1 h post-exercise in CMV-seronegative subjects,but not in those infected with CMV. Collectively, these data indi-cate that latent CMV infection may impair NK-cell-mediatedimmunosurveillance after an acute bout of physical exercise.
We previously reported that latent CMV infection is associatedwith a blunted redeployment of NK-cells with exercise (Bigleyet al., 2012). We show here that this blunting effect is exerciseintensity-dependent as mobilization of NK-cells is lower in CMV-infected individuals at +15% and +5% of BLT, but not at �5% ofBLT. The BLT is a reliable proxy measure of the catecholaminethreshold; the point at which epinephrine and norepinephrine lev-els begin to increase exponentially during graded exercise (Podolinet al., 1991). NK-cells express more b-AR than other lymphocytes(Van Tits et al., 1990) and the mobilization of NK-cells with exer-cise can be mimicked with epinephrine infusion (Crary et al.,1983) via a b2-AR-dependent mechanism (Schedlowski et al.,1996). Given the exercise-intensity dependence of the CMV effect,we hypothesized that CMV impairs NK-cell mobilization by inhib-iting the catecholamine/b-AR axis. While CMV serostatus did notimpact plasma catecholamine responses to exercise, it was associ-ated with impaired NK-cell b-AR signaling as evidenced by mark-edly decreased production of the secondary messenger cAMPfollowing stimulation with the b-agonist isoproterenol. This effectwas striking at all 4 doses, which made for a significant result evenwith a conservative p-threshold and small sample size. Interest-ingly, epinephrine and norepinephrine have been linked to CMVreactivation through catecholamine-responsive elements in theCMV promoter (Prosch et al., 2000) and stress hormone levelsare known to correlate strongly with CMV reactivation in Astro-nauts before and after spaceflight (Mehta et al., 2000). Thus, itcould be argued that decreased sensitivity to b-AR stimulation isan adaptation to prevent frequent reactivation of latent CMV infec-tion. It is likely, however, that any effect of CMV on catecholaminesensitivity is dependent on the composition of cellular subtypes.For instance, the abated exercise-induced mobilization of NK-cellsstands in stark contrast to the enhanced redeployment of highly-differentiated CD8+ (Spielmann et al., 2014; Turner et al., 2010)and cd T-cells (Pistillo et al., 2013) that is seen in young, CMV-infected adults. Future studies should determine why CMV exertsapparently dichotomous effects on T-cells and NK-cells in theirresponse to exercise. Additional studies should also rule out thepossibility that CMV affects exercise-related shear stress, anotherfactor that increases concomitantly with exercise intensity.
In our Part I study, we showed that highly-differentiated NK-cells lacking NKG2C are preferentially mobilized by exercise(Bigley et al., 2014a). It is shown here that CMV alters NK-cell phe-notype in the direction of lower exercise responsiveness. Specifi-cally, there is an increased proportion of NKG2C+ NK-cells and adecreased proportion of KIR+/NKG2A� NK-cells. These data arein line with earlier mouse studies that report ‘‘unlicensed’’ NK-cells(those lacking inhibitory KIR for self-MHC molecules) expressingLy49 (mouse equivalent of NKG2C) are preferentially expandedin response to CMV infection (Orr et al., 2010). Of particular impor-tance, latent CMV infection was associated with a far greater per-centage of NKG2C+/CD57+ NK-cells, which was in turn associatedwith a marked, intensity-dependent reduction in the post-exercisemobilization of NK-cells. Moreover, the expression of b2-AR isreduced on CD57+ NK-cells in those infected with CMV and therewas a strong negative correlation between the proportion ofNKG2C+/CD57+ NK-cells and isoproterenol-induced cAMP produc-tion by NK-cells. Thus, it is likely that the CMV-driven accumula-tion of NKG2C+/CD57+ NK-cells plays a critical role in theimpaired exercise responsiveness of NK-cells through decreasedb-AR expression and sensitivity.
Please cite this article in press as: Bigley, A.B., et al. Acute exercise preferentiallcytotoxicity against lymphoma and multiple myeloma target cells. Part II: ImpaBehav. Immun. (2015), http://dx.doi.org/10.1016/j.bbi.2014.12.027
We also found that latent CMV infection has a profound impacton NK-cell function before and after exercise. CMV was associatedwith increased baseline NKCA against the K562, U266, and221.AEH cell lines, while NKCA against RPMI-8226 was lower inthose infected with CMV and NKCA against 721.221 cells was unaf-fected. This is intuitive, because latent CMV infection is associatedwith an increased proportion of NKG2C+/NKG2A� NK-cells.NKG2C is an activating receptor and NKG2A an inhibitory receptorthat interacts with HLA-E (Vales-Gomez et al., 1999), a non-classi-cal HLA molecule that is expressed by all of the target cell linestested here except 721.221 and RPMI-8226 (Bigley et al., 2014b;Lee et al., 1998). The lower NKCA against RPMI-8226 is likely dueto a lower proportion of licensed NK-cells in CMV-infected individ-uals (Orr et al., 2010) and high expression of classical HLA mole-cules by RPMI-8226 (Heidenreich et al., 2012). An increasedproportion of ‘‘CMV-specific’’ NKG2C+/NKG2A� NK-cells in peoplewith CMV could have major clinical implications as multiple hema-tologic malignancies, including acute myeloid leukemia (AML), arecharacterized by high HLA-E expression (Nguyen et al., 2005; LoMonaco et al., 2011; Bigley et al., 2013). Besides cancer, HIV(Nattermann et al., 2005), CMV (Prod’homme et al., 2007), andHantavirus (Björkström et al., 2011) have all been shown to upreg-ulate HLA-E expression on infected cells, which protects them fromNK-cell-mediated cytotoxicity. Future studies should investigatethe potential of NKG2C+/NKG2A� NK-cells in immunotherapy.
Due to the blunted exercise-induced mobilization of NK-cells,the baseline effect of CMV on NKCA against K562 and U266 cellsdisappeared after exercise, and post-exercise NKCA against721.221 cells was actually greater in CMV-seronegative subjects.Our Part I study reported that proportional shifts in NK-cell subsetswith exercise result in increased 1 h post-exercise NKCA per cellagainst HLA-expressing target cells, such as U266 (HLA-C1) and221.AEH (HLA-E) (Bigley et al., 2014a). We show here that the1 h post-exercise increase in NKCA per cell against HLA-expressingtumor cell lines (U266, RPMI-8226, and 221 AEH) is absent in thosewith CMV, due in part, perhaps, to an accumulation of poorly exer-cise responsive NKG2C+ NK-cells. Interestingly, the proportion ofhighly cytotoxic NKG2C+/NKG2A� NK-cells is elevated 1 h post-exercise in CMV-seronegative individuals; however, it falls belowbaseline in those with CMV. Thus, CMV not only abates the exer-cise-induced redeployment of NK-cells, but also renders NK-cellsinsusceptible to exercise-induced increases in NK-cell cytotoxicity.We did not find any sex differences in NK-cell functional responsesto CMV or exercise, which is in line with other studies that showedno difference in NK-cell cytotoxicity between men and pre-meno-pausal women (Souza et al., 2001; Yovel et al., 2001).
In summary, latent CMV infection is associated with a markedreduction in the mobilization of NK-cells in response to exerciseat intensities above BLT, and renders NK-cells impervious to theaugmenting effect of exercise on NK-cell cytotoxicity that is seenin non-infected people. These effects were associated with pro-found alterations in NK-cell phenotype and b-AR signaling, indicat-ing that the CMV effect might be due to reduced NK-cellcatecholamine sensitivity. We conclude that latent CMV infectionmay compromise NK-cell mediated immunosurveillance followingan acute stress response due to an increased proportion of ‘‘CMV-specific’’ NK-cell subsets with impaired b-AR signaling pathways.
Acknowledgments
The authors thank Dr. Hanspeter Pircher for providing theAlexa488-conjugated anti-KLRG1 (clone 13F12F2) monoclonalantibody and Dr. Dan Geraghty for providing the 721.221 and221 AEH cells. This work was supported by NASA GrantNNX12AB48G to R.J. Simpson.
y redeploys NK-cells with a highly-differentiated phenotype and augmentsct of latent cytomegalovirus infection and catecholamine sensitivity. Brain

A.B. Bigley et al. / Brain, Behavior, and Immunity xxx (2015) xxx–xxx 7
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.bbi.2014.12.027.
References
Benschop, R.J., Nijkamp, F.P., Ballieux, R.E., Heijnen, C.J., 1994. The effects of beta-adrenoceptor stimulation on adhesion of human natural killer cells to culturedendothelium. Br. J. Pharmacol. 113, 1311–1316.
Bigley, A.B., Lowder, T.W., Spielmann, G., Rector, J.L., Pircher, H., Woods, J.A.,Simpson, R.J., 2012. NK-cells have an impaired response to acute exercise and alower expression of the inhibitory receptors KLRG1 and CD158a in humans withlatent cytomegalovirus infection. Brain Behav. Immun. 26, 177–186.
Bigley, A.B., Spielmann, G., LaVoy, E.C., Simpson, R.J., 2013. Can exercise-relatedimprovements in immunity influence cancer prevention and prognosis in theelderly? Maturitas 76, 51–56.
Bigley, A.B., Rezvani, K., Chew, C., Sekine, T., Pistillo, M., Crucian, B., Bollard, C.M.,Simpson, R.J., 2014a. Acute exercise preferentially redeploys NK-cells with ahighly-differentiated phenotype and augments cytotoxicity against lymphomaand multiple myeloma target cells. Brain Behav. Immun. 39, 160–171.
Bigley, A.B., Spielmann, G., LaVoy, E.C., Pistillo, M., Kunz, H., Agha, N., Simpson, R.J.,2014b. Latent cytomegalovirus infection enhances baseline anti-tumorcytotoxicity but impairs NK-cell responses to acute exercise throughpreferential expansion of NKG2C+ NK-cells in healthy humans. Brain Behav.Immun. 40S, e44.
Björkström, N.K., Lindgren, T., Stoltz, M., Fauriat, C., Braun, M., et al., 2011. Rapidexpansion and long-term persistence of elevated NK cell numbers in humansinfected with hantavirus. J. Exp. Med. 208, 13–21.
Crary, B., Hauser, S.L., Borysenko, M., Kutz, I., Hoban, C., Ault, K.A., Weiner, H.L.,Benson, H., 1983. Epinephrine-induced changes in the distribution oflymphocyte subsets in peripheral blood of humans. J. Immunol. 131, 1178–1181.
Heidenreich, S., ZuEulenburg, C., Hildebrandt, Y., Stubig, T., Sierich, H., Badbaran, A.,Eiermann, T.H., Binder, T.M.C., Kroger, N., 2012. Impact of the NK cell receptorLIR-1 (ILT-2/CD85j/LILRB1) on cytotoxicity against multiple myeloma. Clin. Dev.Immunol. 2012, 1–13 (Article ID 652130).
Lee, N., Goodlett, D.R., Ishitani, A., Marquardt, H., Geraghty, D.E., 1998. HLA-Esurface expression depends on binding of TAP-dependent peptides derivedfrom certain HLA class I signal sequences. J. Immunol. 160, 4951–4960.
Lo Monaco, E., Tremante, E., Cerboni, C., Melucci, E., Sibilio, L., Zingoni, A., Nicotra,M.R., Natali, P.G., Giacomini, P., 2011. Human leukocyte antigen E contributes toprotect tumor cells from lysis by natural killer cells. Neoplasia 13 (9), 822–830.
Lopez-Verges, S., Milush, J.M., Pandey, S., York, V.A., Arakawa-Hoyt, J., Pircher, H.,Norris, P.J., Nixon, D.F., Lanier, L.L., 2010. CD57 defines a functionally distinctpopulation of mature NK cells in the human CD56dimCD16+ NK-cell subset.Blood 116, 3865–3874.
Mehta, S.K., Stowe, R.P., Feiveson, A.H., Tyring, S.K., Pierson, D.L., 2000. Reactivationand shedding of cytomegalovirus in astronauts during spaceflight. J. Infect. Dis.182, 1761–1764.
Please cite this article in press as: Bigley, A.B., et al. Acute exercise preferentiallcytotoxicity against lymphoma and multiple myeloma target cells. Part II: ImpaBehav. Immun. (2015), http://dx.doi.org/10.1016/j.bbi.2014.12.027
Nattermann, J., Nischalke, H.D., Hofmeister, V., Kupfer, B., Ahlenstiel, G., Feldmann,G., Rockstroh, J., Weiss, E.H., Sauerbruch, T.S.U., 2005. HIV-1 infection leads toincreased HLA-E expression resulting in impaired function of natural killer cells.Antivir. Ther. 10, 95–107.
Nguyen, S., Dhedin, N., Vernant, J.P., Kuentz, M., Al Jijakli, A., Rouas-Freiss, N.,Carosella, E.D., Boudifa, A., Debre, P., Vieillard, V., 2005. NK-cell reconstitutionafter haploidentical hematopoietic stem-cell transplantations: immaturity ofNK cells and inhibitory effect of NKG2A override GvL effect. Blood 105 (10),4135–4142.
Orr, M.T., Murphy, W.J., Lanier, L.L., 2010. ‘Unlicensed’ natural killer cells dominatethe response to cytomegalovirus infection. Nat. Immunol. 11, 321–327.
Pistillo, M., Bigley, A.B., Spielmann, G., LaVoy, E.C., Morrison, M.R., Kunz, H.,Simpson, R.J., 2013. The effects of age and viral serology on gammadelta T-cellnumbers and exercise responsiveness in humans. Cell. Immunol. 284, 91–97.
Podolin, D.A., Munger, P.A., Mazzeo, R.S., 1991. Plasma catecholamine and lactateresponse during graded exercise with varied glycogen conditions. J. Appl.Physiol. 71, 1427–1433.
Prodhomme, V., Griffin, C., Aicheler, R.J., Wang, E.C., McSharry, B.P., Rickards, C.R.,Stanton, R.J., Borysiewicz, L.K., Lopez-Botet, M., Wilkinson, G.W., Tomasec, P.,2007. The human cytomegalovirus MHC class I homolog UL18 inhibits LIR-1+but activates LIR-1- NK cells. J. Immunol. 178 (7), 4473–4481.
Prosch, S., Wendt, C.E., Reinke, P., Priemer, C., Oppert, M., Kruger, D.H., Volk, H.D.,Docke, W.D., 2000. A novel link between stress and human cytomegalovirus(HCMV) infection: sympathetic hyperactivity stimulates HCMV activation.Virology 272, 357–365.
Schedlowski, M., Hosch, W., Oberbeck, R., Benschop, R.J., Jacobs, R., Raab, H.R.,Schmidt, R.E., 1996. Catecholamines modulate human NK cell circulation andfunction via spleen-independent beta 2-adrenergic mechanisms. J. Immunol.156, 93–99.
Souza, S.S., Castro, F.A., Mendonca, H.C., Palma, P.V., Morais, F.R., Ferriani, R.A.,Voltarelli, J.C., 2001. Influence of menstrual cycle on NK activity. J. Reprod.Immunol. 50, 151–159.
Spielmann, G., Bollard, C.M., Bigley, A.B., Hanley, P.J., Blaney, J.W., LaVoy, E.C.,Pircher, H., Simpson, R.J., 2014. The effects of age and latent cytomegalovirusinfection on the redeployment of CD8+ T cell subsets in response to acuteexercise in humans. Brain Behav. Immun. 39, 142–151.
Turner, J.E., Aldred, S., Witard, O.C., Drayson, M.T., Moss, P.M., Bosch, J.A., 2010.Latent cytomegalovirus infection amplifies CD8 T-lymphocyte mobilisation andegress in response to exercise. Brain Behav. Immun. 24, 1362–1370.
Vales-Gomez, M., Reyburn, H.T., Erskine, R.A., Lopez-Botet, M., Strominger, J.L.,1999. Kinetics and peptide dependency of the binding of the inhibitory NKreceptor CD94/NKG2-A and the activating receptor CD94/NKG2-C to HLA-E.EMBO J. 18, 4250–4260.
Van Tits, L.J., Michel, M.C., Grosse-Wilde, H., Happel, M., Eigler, F.W., Soliman, A.,Brodde, O.E., 1990. Catecholamines increase lymphocyte beta 2-adrenergicreceptors via a beta 2-adrenergic, spleen-dependent process. Am. J. Physiol. 258,E191–E202.
Yovel, G., Shakhar, K., Ben Eliyahu, S., 2001. The effects of sex, menstrual cycle, andoral contraceptives on the number and activity of natural killer cells. Gynecol.Oncol. 81, 254–262.
y redeploys NK-cells with a highly-differentiated phenotype and augmentsct of latent cytomegalovirus infection and catecholamine sensitivity. Brain
Related Documents











