Toxicology Letters 129 (2002) 245 – 254 Acute cytotoxicity of the chemical carcinogen 2-acetylaminofluorene in cultured rat liver epithelial cells Lydie Sparfel, Maud Loewert, Laurence Huc, Le ´a Payen, Andre ´ Guillouzo, Dominique Lagadic-Gossmann, Olivier Fardel * INSERM U456, Faculte ´ de Pharmacie, 2 Aenue du Pr L. Bernard, 35043 Rennes, France Received 29 September 2001; received in revised form 7 January 2002; accepted 8 January 2002 Abstract Acute cytotoxic effects of 2-acetylaminofluorene (AAF), a powerful chemical carcinogen, were studied in cultured rat liver epithelial F258 cells. Acute treatment of these cells by AAF resulted in inhibition of cell proliferation, through an arrest in G 0 /G 1 phase of the cell cycle, and a loss of cell viability. By contrast, AAF failed to trigger apoptosis as demonstrated by flow cytometric analysis of hypoploid sub-G 1 cells. Cytochrome P4501A1 (CYP1A1), a drug metabolizing enzyme thought to play a major role in biotransformation of AAF, was induced in AAF-treated F258 cells as assessed by Northern blotting. AAF cytotoxic effects were however not blocked by the CYP1A1 inhibitor -naphtoflavone, thus suggesting that they did not require CYP1A1 activity. They were also not prevented by the antioxidant N-acetylcysteine, making unlikely a major contribution of AAF-related reactive oxygen species. These data therefore indicate that AAF can exert acute cellular toxicity, including inhibition of cell growth and cell death, in rat liver epithelial cells without triggering an apoptotic process. Such an acute toxicity may contribute to the well-known promoting effects of AAF. © 2002 Elsevier Science Ireland Ltd. All rights reserved. Keywords: 2-Acetylaminofluorene; Apoptosis; Cell cycle; Cytochrome P-4501A1; Liver cells www.elsevier.com/locate/toxlet 1. Introduction The arylamide 2-acetylaminofluorene (AAF) is a powerful carcinogen commonly used in experi- mental liver carcinogenesis models (Heflich and Neft, 1994). The classic Solt – Farber model of chemical tumorigenesis in rat liver thus involves sequential treatment of rats with diethylni- trosamine as an initiating agent and AAF as a promoter (Solt and Farber, 1976). The effects of AAF in this model are thought to be due to mito-inhibition of normal hepatocytes, leading to selective proliferation and expansion of initiated hepatocytes which further form preneoplastic le- sions (Farber, 1990). AAF can also act as a direct carcinogen; indeed, its metabolism upon the ac- tion of drug metabolizing enzymes, especially cy- * Corresponding author. Tel.: +33-19-2-23-23-48-80; fax: +33-19-2-23-33-47-94. E-mail address: [email protected] (O. Fardel). 0378-4274/02/$ - see front matter © 2002 Elsevier Science Ireland Ltd. All rights reserved. PII:S0378-4274(02)00015-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Toxicology Letters 129 (2002) 245–254
Acute cytotoxicity of the chemical carcinogen2-acetylaminofluorene in cultured rat liver epithelial cells
Lydie Sparfel, Maud Loewert, Laurence Huc, Lea Payen, Andre Guillouzo,Dominique Lagadic-Gossmann, Olivier Fardel *
INSERM U456, Faculte de Pharmacie, 2 A�enue du Pr L. Bernard, 35043 Rennes, France
Received 29 September 2001; received in revised form 7 January 2002; accepted 8 January 2002
Abstract
Acute cytotoxic effects of 2-acetylaminofluorene (AAF), a powerful chemical carcinogen, were studied in culturedrat liver epithelial F258 cells. Acute treatment of these cells by AAF resulted in inhibition of cell proliferation,through an arrest in G0/G1 phase of the cell cycle, and a loss of cell viability. By contrast, AAF failed to triggerapoptosis as demonstrated by flow cytometric analysis of hypoploid sub-G1 cells. Cytochrome P4501A1 (CYP1A1),a drug metabolizing enzyme thought to play a major role in biotransformation of AAF, was induced in AAF-treatedF258 cells as assessed by Northern blotting. AAF cytotoxic effects were however not blocked by the CYP1A1inhibitor �-naphtoflavone, thus suggesting that they did not require CYP1A1 activity. They were also not preventedby the antioxidant N-acetylcysteine, making unlikely a major contribution of AAF-related reactive oxygen species.These data therefore indicate that AAF can exert acute cellular toxicity, including inhibition of cell growth and celldeath, in rat liver epithelial cells without triggering an apoptotic process. Such an acute toxicity may contribute to thewell-known promoting effects of AAF. © 2002 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: 2-Acetylaminofluorene; Apoptosis; Cell cycle; Cytochrome P-4501A1; Liver cells
www.elsevier.com/locate/toxlet
1. Introduction
The arylamide 2-acetylaminofluorene (AAF) isa powerful carcinogen commonly used in experi-mental liver carcinogenesis models (Heflich andNeft, 1994). The classic Solt–Farber model of
chemical tumorigenesis in rat liver thus involvessequential treatment of rats with diethylni-trosamine as an initiating agent and AAF as apromoter (Solt and Farber, 1976). The effects ofAAF in this model are thought to be due tomito-inhibition of normal hepatocytes, leading toselective proliferation and expansion of initiatedhepatocytes which further form preneoplastic le-sions (Farber, 1990). AAF can also act as a directcarcinogen; indeed, its metabolism upon the ac-tion of drug metabolizing enzymes, especially cy-
* Corresponding author. Tel.: +33-19-2-23-23-48-80; fax:+33-19-2-23-33-47-94.
E-mail address: [email protected] (O. Fardel).
0378-4274/02/$ - see front matter © 2002 Elsevier Science Ireland Ltd. All rights reserved.
PII: S 0378 -4274 (02 )00015 -2

L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254246
tochromes P-450 1A1/2 (CYP1A1/2), leads to theproduction of electrophilic compounds which re-act with DNA to form mutagenic adducts (Vernaet al., 1996). Besides its implication in chemicalcarcinogenesis, AAF has been shown to up-regu-late detoxifying pathways in hepatocytes. In par-ticular, it induces CYP1A1/2 expression, therebyincreasing its own metabolism and toxicity(Lecureur et al., 1996); it also augments expres-sion of drug efflux pumps such as P-glycoproteinand MRP2 (Kauffmann et al., 1997; Lecureur etal., 1996). AAF has also been shown to exertimmunosuppressive properties (Koh et al., 1994)and to enhance cellular production of reactiveoxygen species (ROS) (Deng et al., 2001).
Although exposure to AAF can lead to ex-tended cellular injury, cellular and molecularbases involved in acute toxicity caused by thiscompound remain incompletely understood.Moreover, studies aimed at investigating this sub-ject have resulted in some puzzling results. Indeed,AAF has been reported to inhibit DNA synthesisafter metabolic activation, possibly reflecting theCYP-dependent formation of DNA adducts(Verna et al., 1996). There is also evidence thatAAF has direct inhibitory effects on cell prolifera-tion through specific interactions with intracellu-lar signal transduction pathways (Skarpen et al.,2000); stimulation of cell growth in response toAAF has however also been reported (Tiwawechet al., 1991). In addition, AAF has been shown totrigger apoptosis in liver cells (Worner andSchrenk, 1996). In the present study, in order togain insights into AAF cellular toxicity, we haveused proliferating rat liver epithelial F258 cellsknown to be sensitive to chemical carcinogenssuch as polycyclic aromatic hydrocarbons (PAHs)(Payen et al., 2001a). Our results indicate thatacute treatment of these liver cells by AAF in-duced both a growth arrest in the G0/G1 phase ofthe cell cycle and a loss of cell viability but failedto trigger apoptosis. In addition, although expo-sure to AAF was found to up-regulate CYP1A1gene expression, co-treatment with the CYP1A1/2inhibitor �-naphtoflavone of AAF-exposed F258cells did not alter toxicity of the chemical carcino-gen, suggesting that AAF exerted direct toxiceffects in these cells.
2. Materials and methods
2.1. Chemicals
AAF, benzo(a)pyrene (BP), �-naphtoflavone,Hoechst 33342 and N-acetyl-L-cysteine (NAC)were purchased from Sigma Chemicals Co (StLouis, MO). AAF was used as a stock solution indimethylsulfoxide; the final concentration of thissolvent in culture medium was always less than0.2% (v/v) and control cultures received the samedose of solvent as AAF-treated cultures. All othercompounds were commercial products of thehighest purity available.
2.2. Cell culture
The F258 rat liver epithelial cell line and itsPAH-resistant F258/3MC variant cell line werecultured in Williams’E medium supplementedwith 10% fetal calf serum at 37 °C under a 5%CO2 atmosphere as previously described (Payen etal., 2001a). Cells were passaged every week usinga 0.1% trypsin solution. F258/3MC cells wereroutinely maintained in the presence of 5 �M3-methylcholanthrene; this compound was how-ever withdrawn at least one week before the useof the cells for experiments.
Rat hepatocytes were isolated from maleSprague–Dawley rats weighing 150–200 g byliver perfusion and cultured in Williams’Emedium as previously described (Payen et al.,2001b).
2.3. Cell proliferation/sur�i�al assay
The effects of AAF on cell proliferation andsurvival were analysed using the 3-(4-5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro-mide (MTT) colorimetric assay (Carmichael et al.,1987). Briefly, cells were cultured with variousconcentrations of AAF in 96-well microplates for96 h. Cells were then incubated with 100 �l ofMTT solution (0.5 mg/ml) for 2 h. Medium wasthereafter discarded and replaced by 100 �l ofdimethylsulfoxide. Blue formazan formed prod-ucts were further quantified by their absorbanceat 540 nm using a Titertek Multiskan (Flow Lab-

L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254 247
oratories, Puteaux, France). Effects of AAF oncell proliferation/survival was notably evaluatedas IC50, i.e. the AAF concentration providing a50% reduction in cell number or cell viability ascompared to control cells cultured in parallelwithout AAF.
2.4. Flow cytometry analysis of cell cycle andapoptosis
F258 cells, growing in exponential phase, weretreated with 100 �M or 200 �M AAF for 48 h.Cells were then harvested and washed with phos-phate-buffer saline. Cell nuclei were isolated andstained with propidium iodide using the CycleTest™ PLUS DNA Reagent Kit (Becton Dickin-son, San Jose, CA). DNA content of 20 000 cells/analysis was monitored with a FACSCalibur flowcytometer (Becton Dickinson). Data wereanalysed using the Modfit software (Becton Dick-inson) when performing analysis of the cell cycleparameters or using the Cell Quest software (Bec-ton Dickinson) for peak detection of sub-G1 hy-poploid apoptotic cells.
2.5. Light microscopic detection of apoptosis
Microscopic detection of apoptosis was per-formed in floating cells using Hoechst 33342 la-belling. Briefly, culture media from AAF-treatedcell cultures were collected and centrifuged. Cellsfound in the pellets were further fixed, stained
with Hoechst 33342 and examined using a fluores-cence Olympus BX60 microscope. Cells withapoptotic nuclei, i.e. nuclei with condensed orfragmented chromatin, were then counted.
2.6. RNA isolation and northern blotting
Total RNA was extracted from cells using theSV Total RNA Isolation System (Promega,Madison, WI). Ten �g of total RNAs were sub-jected to electrophoresis in a denaturing 6% (v/v)formaldehyde 1% (w/v) agarose gel, and trans-ferred onto Hybond�N+ sheets (Amersham, LesUlis, France). The sheets were prehybridized andthen hybridized with a 32P-labeled CYP1A1 probe(Affolter et al., 1986), washed, dried and autora-diographied at −80 °C. Equal RNA loadingonto the gel and efficiency of the transfer werechecked by rehybridizing the blot with a 18SrRNA probe.
2.7. Statistical analysis
Data were analysed using the Student’s t-test.The level of significance was P�0.05.
3. Results
3.1. Cytotoxicity of AAF
We first investigated the effects of AAF onF258 liver epithelial cell proliferation using the
Fig. 1. Inhibition of cell proliferation in response to AAF. Growing F258 cells were treated with increasing concentrations of AAFfor 96 h. Cell growth was then determined using the MTT assay. Data are expressed as percentages of cell proliferation found inuntreated control cells and are the means�SEM of three independent experiments performed in triplicate. *, P�0.05 whencompared to untreated cells.
L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254248
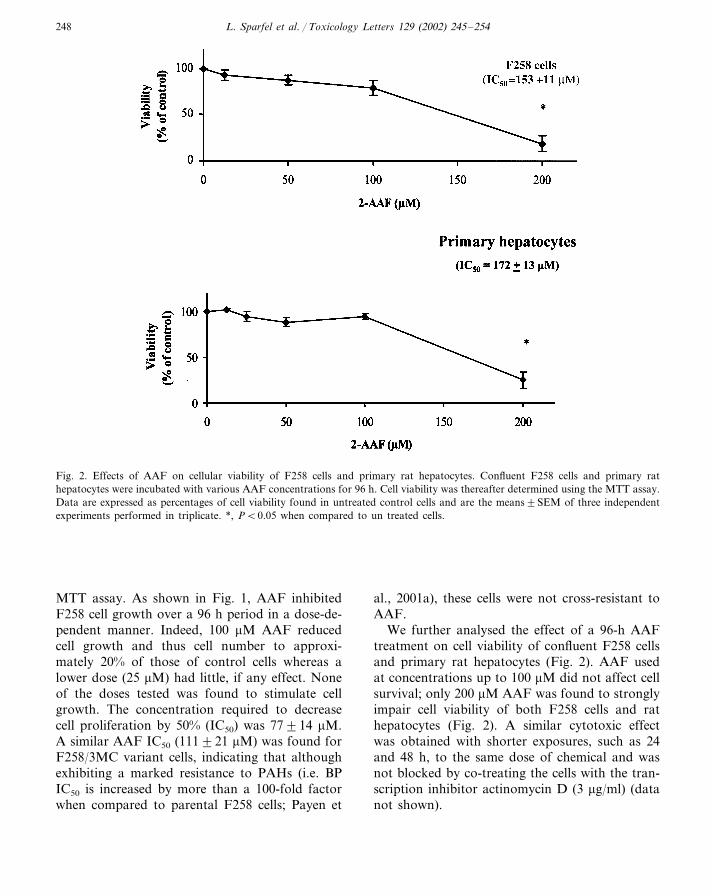
Fig. 2. Effects of AAF on cellular viability of F258 cells and primary rat hepatocytes. Confluent F258 cells and primary rathepatocytes were incubated with various AAF concentrations for 96 h. Cell viability was thereafter determined using the MTT assay.Data are expressed as percentages of cell viability found in untreated control cells and are the means�SEM of three independentexperiments performed in triplicate. *, P�0.05 when compared to un treated cells.
MTT assay. As shown in Fig. 1, AAF inhibitedF258 cell growth over a 96 h period in a dose-de-pendent manner. Indeed, 100 �M AAF reducedcell growth and thus cell number to approxi-mately 20% of those of control cells whereas alower dose (25 �M) had little, if any effect. Noneof the doses tested was found to stimulate cellgrowth. The concentration required to decreasecell proliferation by 50% (IC50) was 77�14 �M.A similar AAF IC50 (111�21 �M) was found forF258/3MC variant cells, indicating that althoughexhibiting a marked resistance to PAHs (i.e. BPIC50 is increased by more than a 100-fold factorwhen compared to parental F258 cells; Payen et
al., 2001a), these cells were not cross-resistant toAAF.
We further analysed the effect of a 96-h AAFtreatment on cell viability of confluent F258 cellsand primary rat hepatocytes (Fig. 2). AAF usedat concentrations up to 100 �M did not affect cellsurvival; only 200 �M AAF was found to stronglyimpair cell viability of both F258 cells and rathepatocytes (Fig. 2). A similar cytotoxic effectwas obtained with shorter exposures, such as 24and 48 h, to the same dose of chemical and wasnot blocked by co-treating the cells with the tran-scription inhibitor actinomycin D (3 �g/ml) (datanot shown).

L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254 249
3.2. Cell cycle-related response to AAF
To investigate the basis of the inhibition ofF258 cell proliferation due to AAF, we examinedthe relative percentages of cells in the different cellcycle phases after a 48 h exposure to 100 �M or200 �M AAF using flow cytometry. As indicatedin Table 1, about 50% of untreated cells were in Sphase, likely reflecting the high proliferative statusof these liver epithelial cells. In the presence of100 �M AAF, the proportion of cells in the Sphase markedly decreased whereas the contingentof cells in the G0/G1 phase concomitantly in-creased. This accumulation of cells in G0/G1 waseven more pronounced in cells treated by 200 �MAAF (Table 1); indeed, almost 90% of the cellswere identified in the G0/G1 phase whereas thecontingents of cells in the S and G2/M phaseswere dramatically reduced when compared totheir untreated counterparts.
3.3. Apoptosis in response to AAF
To explore a putative apoptotic response inAAF-treated cells, we have first investigated theproportion of sub-G1 hypoploid apoptotic cells inAAF-exposed cultures. As shown in Fig. 3, AAFused at 100 or 200 �M did not, or only mar-ginally, increase the number of sub-G1 cells whencompared to controls. By contrast, treatment byBP, a chemical carcinogen which has been previ-ously shown to trigger apoptosis in liver cells (Lei
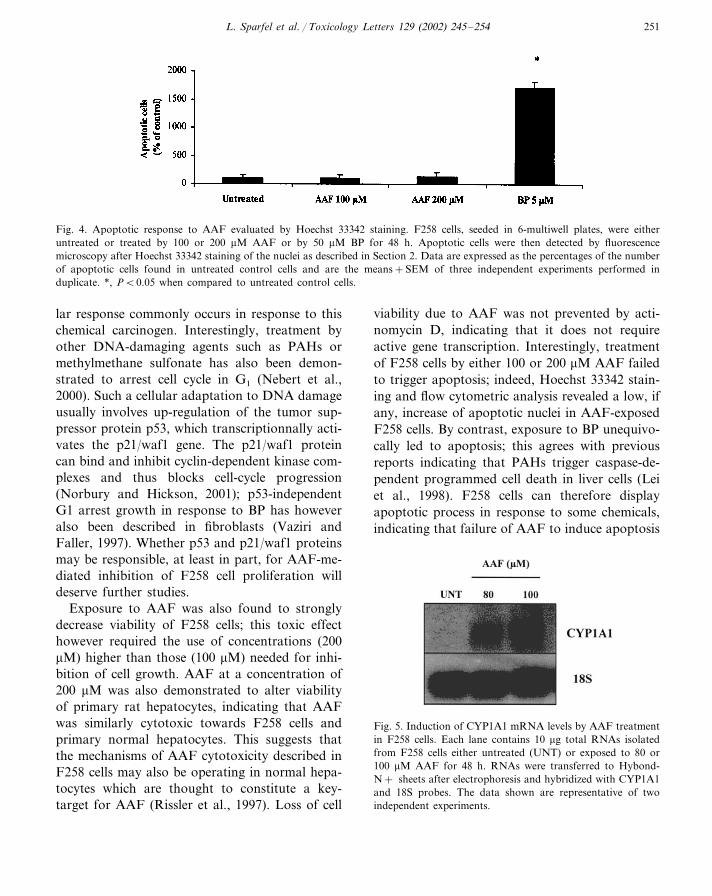
et al., 1998), resulted in a marked increase in thepercentage of hypoploid cells. Similarly, BP in-creased the number of F258 cells exhibiting apop-totic nuclei, i.e. condensed or fragmentedchromatin after Hoechst 33342 staining, whereasAAF had no effect (Fig. 4).
3.4. AAF toxicity is not modulated by�-naphtofla�one
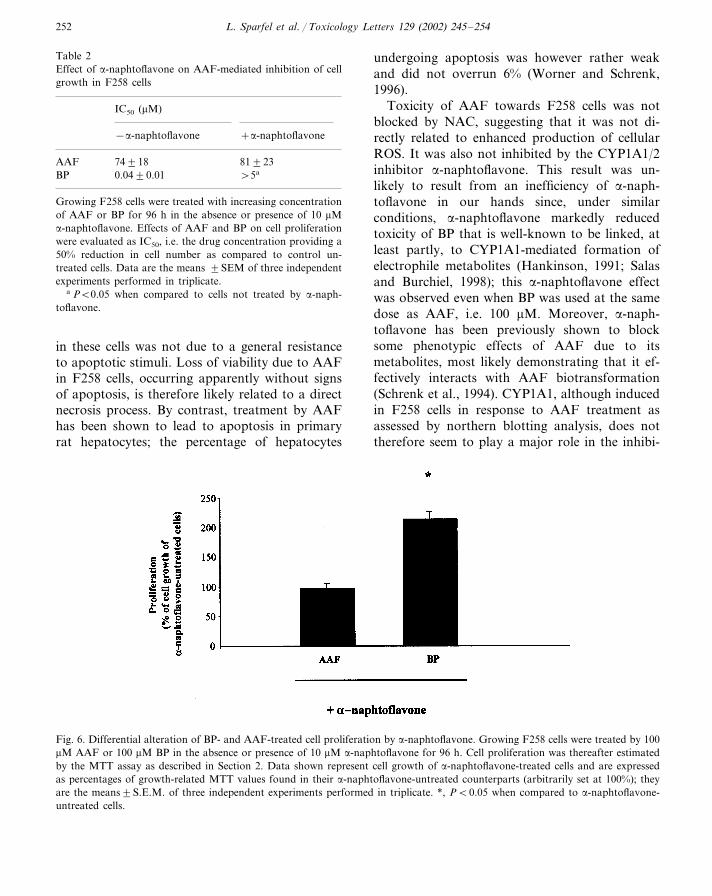
CYP1A1/2 is thought to play an important rolein the occurrence of toxic effects of chemicalcarcinogens, including AAF and BP. This drugmetabolizing enzyme, not detected at the mRNAlevel in untreated F258 cells, was however inducedin response to 80 or 100 �M AAF as demon-strated by Northern blotting (Fig. 5); BP treat-ment also resulted in CYP1A1 up-regulation (datanot shown) in agreement with previous data(Payen et al., 2001a). The addition of �-naph-toflavone, used at a concentration (10 �M) knownto nearly fully inhibit CYP1A1/2 activity (Blanket al., 1987), however failed to modulate signifi-cantly the toxicity of AAF towards F258 cellgrowth as revealed by measurements of AAF IC50
through MTT assays (Table 2); it also did notabolish the loss of viability occurring in confluentF258 cells treated by 200 �M AAF, even when�-naphtoflavone was used at a higher concentra-tion (25 �M) (data not shown). By contrast, �-naphtoflavone markedly reduced the toxicity ofBP. Indeed, the IC50 of this PAH was increasedby more than a 130-fold factor in the presence ofthe CYP1A1/2 inhibitor (Table 2). Moreover,proliferation of F258 cells exposed to 100 �M BP,a highly cytotoxic concentration, was notably in-creased by co-treatment with 10 �M �-naph-toflavone (Fig. 6), demonstrating that�-naphtoflavone still protects cells, at least inpart, against elevated doses of BP; however, �-naphtoflavone did not act at all on cell growth ofF258 cells exposed to 100 �M AAF (Fig. 6).
3.5. AAF toxicity is not pre�ented by NAC
Since the antioxidant NAC is known to preventor decrease toxic effects of xenobiotics actingthrough enhanced production of ROS (Cotgreave,
Table 1Flow cytometric analysis of AAF effects on the relative distri-bution of cell cycle phases in F258 cells
% of cells
S G2/MG0/G1
Control 30.7�11.4 49.6�8.9 19.7�6.150.5�4.8a 16.8�1.132.7�3.9a100 �M AAF86.2�15.6a 7.4�6.6a 6.4�9.4a200 �M AAF
Cells were treated with 100 or 200 �M AAF for 48 h. Nucleiwere then stained by prodium iodide and analysed by flowcytometry. Data are expressed as percentage of cells in theG0/G1, S and G2/M phases of cell cycle; they are the means�SD of three independent experiments.
a P�0.05 when compared to control untreated cells.

L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254250
Fig. 3. Apoptotic response to AAF assessed by flow cytometric analysis of hypoploid sub-G1 cells. F258 cells were either untreatedor treated by 100 or 200 �M AAF or by 5 �M BP for 48 h. Hypoploid sub-G1 cells were then detected by flow cytometry asindicated in Section 2. The apoptotic sub-G1 cell numbers are expressed as percentages of total cell count and are the means�SEMof three independent experiments.
1997) and since AAF has been reported to in-crease cellular levels of ROS (Deng et al., 2001;Verna et al., 1996), we have analysed the effects of10 mM NAC over a 24 h period on AAF toxicityin F258 cells. As shown in Fig. 7, NAC failed toprevent the loss of viability due to AAF. NACwas nevertheless fully active in our F258 cellmodel since it totally blocked the toxicity of H2O2
(Fig. 7).
4. Discussion
The present study was designed to analyseacute toxicity of AAF in liver epithelial cells
which have been previously shown to be targetsfor chemical carcinogens such as PAHs (Payen etal., 2001a). Our results demonstrate that in vitroexposure to AAF exerted inhibitory effects to-wards F258 cell proliferation through arrestingcells in the G0/G1 phase of the cell cycle. Simi-larly, administration of AAF to partially hepatec-tomised rats has been reported to blockhepatocytes in G1 phase. This effect is believed todirectly contribute to the promoting effects ofAAF (Rissler et al., 1997) and it involves lack ofcyclin E expression at the G1/S phase restrictionpoint (Trautwein et al., 1999). Thus, both in vitroand in vivo exposure to AAF results in cell cycleblockage in G1, likely indicating that such a cellu-
L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254 251
Fig. 4. Apoptotic response to AAF evaluated by Hoechst 33342 staining. F258 cells, seeded in 6-multiwell plates, were eitheruntreated or treated by 100 or 200 �M AAF or by 50 �M BP for 48 h. Apoptotic cells were then detected by fluorescencemicroscopy after Hoechst 33342 staining of the nuclei as described in Section 2. Data are expressed as the percentages of the numberof apoptotic cells found in untreated control cells and are the means+SEM of three independent experiments performed induplicate. *, P�0.05 when compared to untreated control cells.
lar response commonly occurs in response to thischemical carcinogen. Interestingly, treatment byother DNA-damaging agents such as PAHs ormethylmethane sulfonate has also been demon-strated to arrest cell cycle in G1 (Nebert et al.,2000). Such a cellular adaptation to DNA damageusually involves up-regulation of the tumor sup-pressor protein p53, which transcriptionnally acti-vates the p21/waf1 gene. The p21/waf1 proteincan bind and inhibit cyclin-dependent kinase com-plexes and thus blocks cell-cycle progression(Norbury and Hickson, 2001); p53-independentG1 arrest growth in response to BP has howeveralso been described in fibroblasts (Vaziri andFaller, 1997). Whether p53 and p21/waf1 proteinsmay be responsible, at least in part, for AAF-me-diated inhibition of F258 cell proliferation willdeserve further studies.
Exposure to AAF was also found to stronglydecrease viability of F258 cells; this toxic effecthowever required the use of concentrations (200�M) higher than those (100 �M) needed for inhi-bition of cell growth. AAF at a concentration of200 �M was also demonstrated to alter viabilityof primary rat hepatocytes, indicating that AAFwas similarly cytotoxic towards F258 cells andprimary normal hepatocytes. This suggests thatthe mechanisms of AAF cytotoxicity described inF258 cells may also be operating in normal hepa-tocytes which are thought to constitute a key-target for AAF (Rissler et al., 1997). Loss of cell
viability due to AAF was not prevented by acti-nomycin D, indicating that it does not requireactive gene transcription. Interestingly, treatmentof F258 cells by either 100 or 200 �M AAF failedto trigger apoptosis; indeed, Hoechst 33342 stain-ing and flow cytometric analysis revealed a low, ifany, increase of apoptotic nuclei in AAF-exposedF258 cells. By contrast, exposure to BP unequivo-cally led to apoptosis; this agrees with previousreports indicating that PAHs trigger caspase-de-pendent programmed cell death in liver cells (Leiet al., 1998). F258 cells can therefore displayapoptotic process in response to some chemicals,indicating that failure of AAF to induce apoptosis
Fig. 5. Induction of CYP1A1 mRNA levels by AAF treatmentin F258 cells. Each lane contains 10 �g total RNAs isolatedfrom F258 cells either untreated (UNT) or exposed to 80 or100 �M AAF for 48 h. RNAs were transferred to Hybond-N+ sheets after electrophoresis and hybridized with CYP1A1and 18S probes. The data shown are representative of twoindependent experiments.
L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254252
Table 2Effect of �-naphtoflavone on AAF-mediated inhibition of cellgrowth in F258 cells
IC50 (�M)
−�-naphtoflavone +�-naphtoflavone
AAF 81�2374�18BP 0.04�0.01 �5a
Growing F258 cells were treated with increasing concentrationof AAF or BP for 96 h in the absence or presence of 10 �M�-naphtoflavone. Effects of AAF and BP on cell proliferationwere evaluated as IC50, i.e. the drug concentration providing a50% reduction in cell number as compared to control un-treated cells. Data are the means �SEM of three independentexperiments performed in triplicate.
a P�0.05 when compared to cells not treated by �-naph-toflavone.
undergoing apoptosis was however rather weakand did not overrun 6% (Worner and Schrenk,1996).
Toxicity of AAF towards F258 cells was notblocked by NAC, suggesting that it was not di-rectly related to enhanced production of cellularROS. It was also not inhibited by the CYP1A1/2inhibitor �-naphtoflavone. This result was un-likely to result from an inefficiency of �-naph-toflavone in our hands since, under similarconditions, �-naphtoflavone markedly reducedtoxicity of BP that is well-known to be linked, atleast partly, to CYP1A1-mediated formation ofelectrophile metabolites (Hankinson, 1991; Salasand Burchiel, 1998); this �-naphtoflavone effectwas observed even when BP was used at the samedose as AAF, i.e. 100 �M. Moreover, �-naph-toflavone has been previously shown to blocksome phenotypic effects of AAF due to itsmetabolites, most likely demonstrating that it ef-fectively interacts with AAF biotransformation(Schrenk et al., 1994). CYP1A1, although inducedin F258 cells in response to AAF treatment asassessed by northern blotting analysis, does nottherefore seem to play a major role in the inhibi-
in these cells was not due to a general resistanceto apoptotic stimuli. Loss of viability due to AAFin F258 cells, occurring apparently without signsof apoptosis, is therefore likely related to a directnecrosis process. By contrast, treatment by AAFhas been shown to lead to apoptosis in primaryrat hepatocytes; the percentage of hepatocytes
Fig. 6. Differential alteration of BP- and AAF-treated cell proliferation by �-naphtoflavone. Growing F258 cells were treated by 100�M AAF or 100 �M BP in the absence or presence of 10 �M �-naphtoflavone for 96 h. Cell proliferation was thereafter estimatedby the MTT assay as described in Section 2. Data shown represent cell growth of �-naphtoflavone-treated cells and are expressedas percentages of growth-related MTT values found in their �-naphtoflavone-untreated counterparts (arbitrarily set at 100%); theyare the means�S.E.M. of three independent experiments performed in triplicate. *, P�0.05 when compared to �-naphtoflavone-untreated cells.

L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254 253
Fig. 7. Effect of NAC on AAF cytotoxicity. Confluent F258 cells were either untreated or treated with 200 �M AAF or 1.3 mMH2O2 in the absence or presence of 10 mM NAC for 24 h. Cell viability was thereafter determined using the MTT assay. Data areexpressed as percentages of cell viability compared to untreated control cells and are the means�S.E.M of triplicate cultures. *,P�0.05 when compared to NAC-untreated cells.
tion of cell growth and the loss of cell viabilitydue to AAF. Such a conclusion is moreover fullysupported by the lack of cross resistance to AAFof PAH-resistant F258/3MC cells, which havebeen previously shown to exhibit altered up-regu-lation of CYP1A1 in response to chemicals(Payen et al., 2001a). This suggests that AAFexerted direct toxic effects towards F258 cells.Involvement of other metabolizing enzymes can-not be however completely excluded, althoughunlikely due to the crucial role of CYP1A1/2 inthe metabolism of AAF (Verna et al., 1996). Inthe same way, AAF is thought to display directinhibitory effects on hepatocyte proliferationthrough specific interactions with intracellular sig-nal transduction pathways; notably, AAF hasbeen shown to alter mitogen signalling throughinterfering with some mitogen-activated proteinkinases such as extracellular signal regulatedprotein kinases (Skarpen et al., 2000). It is how-ever noteworthy that some properties of AAF,especially up-regulation of the drug transporterP-glycoprotein, are likely due to its metabolites(Schrenk et al., 1994); similarly, mutagenic actionof AAF requires the formation of electrophilemetabolites (Rudo et al., 1987). Taken together,these data indicate that the different phenotypiceffects of AAF can be due to the compound itselfor to its metabolites.
In conclusion, acute exposure to AAF of ratliver epithelial cells was shown to inhibit cell
growth through inducing an arrest in the G1
phase and to lead to necrosis without apparentconcomitant features of apoptosis. These effectswere not prevented by the CYP1A1/2 inhibitor�-naphtoflavone, indicating that AAF may exertdirect cellular toxicity. Whether such an acutetoxicity may participate to the promoting effectsof AAF deserves further studies.
Acknowledgements
This work was supported by the Institut Na-tional de Recherche et de Securite (INRS).
References
Affolter, M., Labbe, D., Jean, A., Raymond, M., Noel, D.,Labelle, Y., Parent-Vaugeois, C., Lambert, M., Bo-janowski, R., Anderson, A., 1986. cDNA clones for livercytochrome P-450s from individual aroclor-treated rats:constitutive expression of a new P-450 gene related tophenobarbital-inducible forms. DNA 5, 209.
Blank, J.A., Tucker, A.N., Sweatlock, J., Gasiewicz, T.A.,Luster, M.I., 1987. alpha-Naphthoflavone antagonism of2,3,7,8-tetrachlorodibenzo-p-dioxin-induced murine lym-phocyte ethoxyresorufin-O-deethylase activity and im-munosuppression. Mol. Pharmacol. 32, 169.
Carmichael, J., DeGraff, W.G., Gazdar, A.F., Minna, J.D.,Mitchell, J.B., 1987. Evaluation of a tetrazolium-basedsemiautomated colorimetric assay: assessment of radiosen-sitivity. Cancer Res. 47, 943.

L. Sparfel et al. / Toxicology Letters 129 (2002) 245–254254
Cotgreave, I.A., 1997. N-acetylcysteine: pharmacological con-siderations and experimental and clinical applications.Adv. Pharmacol. 38, 205.
Deng, L., Lin-Lee, Y.C., Claret, F.X., Kuo, M.T., 2001.2-acetylaminofluorene up-regulates rat mdr1b expressionthrough generating reactive oxygen species that activateNF-kappa B pathway. J. Biol. Chem. 276, 413.
Farber, E., 1990. Clonal adaptation during carcinogenesis.Biochem. Pharmacol. 39, 1837.
Hankinson, O., 1991. Selections for and against cells possess-ing cytochrome P450IA1-dependent aryl hydrocarbon hy-droxylase activity. Methods Enzymol. 206, 381.
Heflich, R.H., Neft, R.E., 1994. Genetic toxicity of 2-acety-laminofluorene, 2-aminofluorene and some of theirmetabolites and model metabolites. Mutat. Res. 318, 73.
Kauffmann, H.M., Keppler, D., Kartenbeck, J., Schrenk, D.,1997. Induction of cMrp/cMoat gene expression by cis-platin, 2-acetylaminofluorene, or cycloheximide in rat hep-atocytes. Hepatology 26, 980.
Koh, W.S., Chun, Y.J., Yang, K.H., 1994. Suppressive effectsof 2-acetylaminofluorene on concanavalin A- stimulatedmurine splenocyte proliferation in vitro: inhibition of inter-leukin-2 receptor expression. Biochem. Mol. Biol. Int. 32,501.
Lecureur, V., Guillouzo, A., Fardel, O., 1996. Differentialregulation of mdr genes in response to 2-acety-laminofluorene treatment in cultured rat and human hepa-tocytes. Carcinogenesis 17, 1157.
Lei, W., Yu, R., Mandlekar, S., Kong, A.N., 1998. Inductionof apoptosis and activation of interleukin 1beta-convertingenzyme/Ced-3 protease (caspase-3) and c-Jun NH2-termi-nal kinase 1 by benzo(a)pyrene. Cancer Res. 58, 2102.
Nebert, D.W., Roe, A.L., Dieter, M.Z., Solis, W.A., Yang, Y.,Dalton, T.P., 2000. Role of the aromatic hydrocarbonreceptor and [Ah] gene battery in the oxidative stressresponse, cell cycle control, and apoptosis. Biochem. Phar-macol. 59, 65.
Norbury, C.J., Hickson, I.D., 2001. Cellular responses toDNA damage. Annu. Rev. Pharmacol. Toxicol. 41, 367.
Payen, L., Courtois, A., Langouet, S., Guillouzo, A., Fardel,O., 2001a. Unaltered expression of multidrug resistancetransporters in polycyclic aromatic hydrocarbon-resistantrat liver cells. Toxicology 156, 109.
Payen, L., Courtois, A., Loewert, M., Guillouzo, A., Fardel,O., 2001b. Reactive oxygen species-related induction ofmultidrug resistance-associated protein 2 expression in pri-
mary hepatocytes exposed to sulforaphane. Biochem. Bio-phys. Res. Commun. 282, 257.
Rissler, P., Torndal, U.B., Eriksson, L.C., 1997. Induced drugresistance inhibits selection of initiated cells and cancerdevelopment. Carcinogenesis 18, 649.
Rudo, K., Meyers, W.C., Dauterman, W., Langenbach, R.,1987. Comparison of human and rat hepatocytemetabolism and mutagenic activation of 2-acety-laminofluorene. Cancer Res. 47, 5861.
Salas, V.M., Burchiel, S.W., 1998. Apoptosis in Daudi humanB cells in response to benzo[a]pyrene and benzo[a]pyrene-7,8-dihydrodiol. Toxicol. Appl. Pharmacol. 151, 367.
Schrenk, D., Gant, T.W., Michalke, A., Orzechowski, A.,Silverman, J.A., Battula, N., Thorgeirsson, S.S., 1994.Metabolic activation of 2-acetylaminofluorene is requiredfor induction of multidrug resistance gene expression in ratliver cells. Carcinogenesis 15, 2541.
Skarpen, E., Lindeman, B., Thoresen, G.H., Guren, T.K.,Oksvold, M.P., Christoffersen, T., Huitfeldt, H.S., 2000.Impaired nuclear accumulation and shortened phosphory-lation of ERK after growth factor stimulation in culturedhepatocytes from rats exposed to 2-acetylaminofluorene.Mol. Carcinog. 28, 84.
Solt, D., Farber, E., 1976. New principle for the analysis ofchemical carcinogenesis. Nature 263, 701.
Tiwawech, D., Hasegawa, R., Kurata, Y., Tatematsu, M.,Shibata, M.A., Thamavit, W., Ito, N., 1991. Dose-depen-dent effects of 2-acetylaminofluorene on hepatic foci devel-opment and cell proliferation in rats. Carcinogenesis 12,985.
Trautwein, C., Will, M., Kubicka, S., Rakemann, T., Flem-ming, P., Ito, M.P., 1999. 2-Acetylaminofluorene blockscell cycle progression after hepatectomy by p21 inductionand lack of Cyclin E expression. Oncogene 18, 6443.
Vaziri, C., Faller, D.V., 1997. A benzo[a]pyrene-induced cellcycle checkpoint resulting in p53-independent G1 arrest in3T3 fibroblasts. J. Biol. Chem. 272, 2762.
Verna, L., Whysner, J., Williams, G.M., 1996. 2-Acety-laminofluorene mechanistic data and risk assessment:DNA reactivity, enhanced cell proliferation and tumorinitiation. Pharmacol. Ther. 71, 83.
Worner, W., Schrenk, D., 1996. Influence of liver tumorpromoters on apoptosis in rat hepatocytes induced by2-acetylaminofluorene, ultraviolet light, or transforminggrowth factor beta 1. Cancer Res. 56, 1272.
Related Documents











