ORIGINAL ARTICLE Activity-dependent structural plasticity of Purkinje cell spines in cerebellar vermis and hemisphere P. De Bartolo • F. Florenzano • L. Burello • F. Gelfo • L. Petrosini Received: 6 February 2014 / Accepted: 24 June 2014 Ó Springer-Verlag Berlin Heidelberg 2014 Abstract The environmental enrichment (EE) paradigm is widely used to study experience-dependent brain plas- ticity. In spite of a long history of research, the EE influ- ence on neuronal morphology has not yet been described in relation to the different regions of the cerebellum. Thus, aim of the present study was to characterize the EE effects on density and size of dendritic spines of Purkinje cell proximal and distal compartments in cerebellar vermian and hemispherical regions. Male Wistar rats were housed in an enriched or standard environment for 3.5 months from the 21st post-natal day onwards. The morphological features of Purkinje cell spines were visualized on calbin- din immunofluorescence-stained cerebellar vermian and hemispherical sections. Density, area, length and head diameter of spines were manually (ImageJ) or automati- cally (Imaris) quantified. Results demonstrated that the Purkinje cell spine density was higher in enriched rats than in controls on both proximal and distal dendrite compart- ments in the hemisphere, while it increased only on distal compartment in the vermis. As for spine size, a significant increase of area, length and head diameter was found in the distal dendrites in both vermis and hemisphere. Thus, the exposure to a complex environment enhances synapse formation and plasticity either in the vermis involved in balance and locomotion and in the hemisphere involved in complex motor adaptations and acquisition of new motor strategies. These data highlight the importance of cere- bellar activity-dependent structural plasticity underling the EE-related high-level performances. Keywords Environmental enrichment Á Proximal and distal dendrites Á Dendritic spines Á Cerebellum Á Rat Introduction The brain is strikingly responsive to environmental stim- ulations to the extent that its morphology is modified by experience. Interaction between brain and environment underlies adjustments allowing adaptation to an ever- changing environment. Brain morphological modifications triggered by the exposure to increased environmental complexity may be experimentally investigated through the environmental enrichment (EE) paradigm (Rosenzweig et al. 1962; Rosenzweig and Bennett 1996). EE is based on the enhancement of motor, sensory, cognitive and social stimulations in housing conditions (Van Praag et al. 2000; Nithianantharajah and Hannan 2006). Enhanced physical and cognitive stimulations induce structural modifications that strengthen neuronal connectivity (Nithianantharajah and Hannan 2006; Petrosini et al. 2009). The magnitude and persistence of the changes induced by the enriched experience depend on the age of the subjects and the length of the exposure (Tremml et al. 2002; Cutuli et al. 2011; De Bartolo et al. 2011). Interestingly, phenotypes acquired P. De Bartolo Á L. Burello Á F. Gelfo Á L. Petrosini IRCCS Santa Lucia Foundation, Via Del Fosso di Fiorano 64, 00143 Rome, Italy P. De Bartolo (&) Á L. Burello Á L. Petrosini Department of Psychology, University Sapienza of Rome, Via dei Marsi 78, 00185 Rome, Italy e-mail: [email protected] F. Florenzano Confocal Microscopy Unit, European Brain Research Institute (EBRI), Via del Fosso di Fiorano 64, 00143 Rome, Italy F. Gelfo Department of Systemic Medicine, University Tor Vergata of Rome, Via Oxford 81, 00133 Rome, Italy 123 Brain Struct Funct DOI 10.1007/s00429-014-0833-6

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Activity-dependent structural plasticity of Purkinje cell spinesin cerebellar vermis and hemisphere
P. De Bartolo • F. Florenzano • L. Burello •
F. Gelfo • L. Petrosini
Received: 6 February 2014 / Accepted: 24 June 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract The environmental enrichment (EE) paradigm
is widely used to study experience-dependent brain plas-
ticity. In spite of a long history of research, the EE influ-
ence on neuronal morphology has not yet been described in
relation to the different regions of the cerebellum. Thus,
aim of the present study was to characterize the EE effects
on density and size of dendritic spines of Purkinje cell
proximal and distal compartments in cerebellar vermian
and hemispherical regions. Male Wistar rats were housed
in an enriched or standard environment for 3.5 months
from the 21st post-natal day onwards. The morphological
features of Purkinje cell spines were visualized on calbin-
din immunofluorescence-stained cerebellar vermian and
hemispherical sections. Density, area, length and head
diameter of spines were manually (ImageJ) or automati-
cally (Imaris) quantified. Results demonstrated that the
Purkinje cell spine density was higher in enriched rats than
in controls on both proximal and distal dendrite compart-
ments in the hemisphere, while it increased only on distal
compartment in the vermis. As for spine size, a significant
increase of area, length and head diameter was found in the
distal dendrites in both vermis and hemisphere. Thus, the
exposure to a complex environment enhances synapse
formation and plasticity either in the vermis involved in
balance and locomotion and in the hemisphere involved in
complex motor adaptations and acquisition of new motor
strategies. These data highlight the importance of cere-
bellar activity-dependent structural plasticity underling the
EE-related high-level performances.
Keywords Environmental enrichment � Proximal and
distal dendrites � Dendritic spines � Cerebellum � Rat
Introduction
The brain is strikingly responsive to environmental stim-
ulations to the extent that its morphology is modified by
experience. Interaction between brain and environment
underlies adjustments allowing adaptation to an ever-
changing environment. Brain morphological modifications
triggered by the exposure to increased environmental
complexity may be experimentally investigated through the
environmental enrichment (EE) paradigm (Rosenzweig
et al. 1962; Rosenzweig and Bennett 1996). EE is based on
the enhancement of motor, sensory, cognitive and social
stimulations in housing conditions (Van Praag et al. 2000;
Nithianantharajah and Hannan 2006). Enhanced physical
and cognitive stimulations induce structural modifications
that strengthen neuronal connectivity (Nithianantharajah
and Hannan 2006; Petrosini et al. 2009). The magnitude
and persistence of the changes induced by the enriched
experience depend on the age of the subjects and the length
of the exposure (Tremml et al. 2002; Cutuli et al. 2011; De
Bartolo et al. 2011). Interestingly, phenotypes acquired
P. De Bartolo � L. Burello � F. Gelfo � L. Petrosini
IRCCS Santa Lucia Foundation, Via Del Fosso di Fiorano 64,
00143 Rome, Italy
P. De Bartolo (&) � L. Burello � L. Petrosini
Department of Psychology, University Sapienza of Rome,
Via dei Marsi 78, 00185 Rome, Italy
e-mail: [email protected]
F. Florenzano
Confocal Microscopy Unit, European Brain Research Institute
(EBRI), Via del Fosso di Fiorano 64, 00143 Rome, Italy
F. Gelfo
Department of Systemic Medicine, University Tor Vergata
of Rome, Via Oxford 81, 00133 Rome, Italy
123
Brain Struct Funct
DOI 10.1007/s00429-014-0833-6

from the exposure to enriched environment can even be
transmitted to the next generation (Arai and Feig 2011;
Caporali et al. 2014).
The importance of studying the EE effects on brain
plasticity arises from the evidence that in humans the
exposure to complex-enriched stimulations (as high edu-
cational or occupational attainment, high IQ, and an active
lifestyle) leads to great tolerance of pathological abnor-
malities (Scarmeas and Stern 2003; Whalley et al. 2000,
2004), a concept known as brain (structural) and cognitive
(functional) reserve theory (Stern 2002, 2003, 2006).
Brain stores information by modulating the strength of
the existing synapses and enlarging or shrinking dendritic
spines, principal postsynaptic target of excitatory afferents
(Gray 1959; Harris and Kater 1994; Fiala et al. 2002).
Intrinsic factors, as neurotrophins, and extrinsic factors, as
environmental conditions, modulate number, shape and
function of dendritic spines (Bae et al. 2012; Petrinovic
et al. 2013). In neocortical and hippocampal neurons
increased spine density is demonstrated in relation to EE
exposure (Moser et al. 1994; Kozorovitskiy et al. 2005;
Mandolesi et al. 2008; Liu et al. 2012), neuronal activity
(Bundman and Gall 1994; Hosokawa et al. 1995; Zhao
et al. 2012), learning (Gonzalez-Ramırez et al. 2013;
Muhammad et al. 2013), and physical exercise (Klintsova
and Greenough 1999; Yau et al. 2011).
Even in the cerebellar circuitries, the increased envi-
ronmental stimulation facilitates neuronal remodeling in the
cells directly involved in the experience-dependent activity.
Namely, exposure to EE produces an enlargement of the
Purkinje cell synaptic buttons and an enhancement of the
mossy and climbing fiber terminal density in deep nuclear
neurons (Foscarin et al. 2011). These observations add to
the previously reported EE-dependent structural remodeling
in the cerebellar cortex. Namely, the number of parallel
fiber–Purkinje cell glutamatergic synapses as well as stel-
late cell–Purkinje cell GABAergic synapses is strongly
increased under EE conditions (Lonetti et al. 2010). EE
increases cerebellar production of neurotrophins (Angelucci
et al. 2009; Vazquez-Sanroman et al. 2013) but not of
synaptophysin, as conversely it occurs in forebrain, hippo-
campus, and thalamus (Nithianantharajah et al. 2004).
Additionally, learning complex acrobatic tasks (but not
merely running) increases the number of synapses in Pur-
kinje cells (Federmeier et al. 2002; Kim et al. 2002; Lee
et al. 2007; Gonzalez-Burgos et al. 2011). Purkinje cells
represent an excellent model to investigate EE-induced
synaptic changes given the dynamic changes they exhibit
under diverse conditions, such as physical activity (Pysh
and Weiss 1979; Gonzalez-Burgos et al. 2011 ), block of
electrical activity (Bravin et al. 1999; Morando et al. 2001),
deafferentation (Sotelo et al. 1975), presence of ataxia
(Rhyu et al. 1999a, b) or olivo-ponto-cerebellar atrophy
(Ferrer et al. 1988, 1994). Considering that the responses to
environmental requests may occur through spine changes
regulating in turn synaptic strength and signaling properties,
the present research was aimed at analyzing the effects of a
prolonged exposure to complex environment on density and
size of Purkinje cell spines to provide information on syn-
aptogenesis and circuit complexity (Koch and Poggio 1983;
Turner 1984; Lee et al. 2012). The EE effects on Purkinje
cell spines have been analyzed both in the vermis and
hemisphere, two regions differently involved in the motor
functions of the cerebellum. The vermis plays a primary
role in regulating extensor tone, maintaining dynamic bal-
ance control, modulating the rhythmic flexor and extensor
muscle activity during locomotion (Chambers and Sprague
1955; Thach et al. 1992). The hemispheres have less
importance in controlling balance and walking, but play a
pivotal role in motor control when complex motor adapta-
tions are required, as in novel and attention-demanding
contexts (Manto and Oulad Ben Taib 2010). Given the
functionally different cortico-cerebellar networks in which
vermian and hemispherical regions are engaged (Morton
and Bastian 2004), EE could differently impact on them.
Spine characterization was performed in proximal and
distal dendritic compartments of the same Purkinje cells.
Namely, the proximal compartment consisting of large
dendrites on which the climbing fibers impinge, displays a
rather low number of spines, while conversely, the distal
compartment made up of thin branchlets contacted by the
parallel fibers displays a very high spine density. Accord-
ingly, the two compartments are differently regulated by the
activity of the specific afferents, and thus could differently
respond to environmental drives (Bravin et al. 1999; Mor-
ando et al. 2005; Sugawara et al. 2013). Noteworthy, in
spite of the described differences in cerebellar functioning,
to our knowledge this is the first study addressing the EE-
induced morphological effects in relation to the differential
functional engagement of the cerebellar vermian and
hemispherical regions as well as of the two Purkinje cell
dendritic compartments.
Materials and methods
Animals and experimental groups
Fourteen male Wistar rats (Harlan Laboratories S.r.l.,
Udine, Italy) were used. They were maintained according
to the guidelines for ethical conduct developed by the
European Communities Council Directive of 22 September
2010 (2010/63/EU). All animal protocols used have been
approved by the Ethical Committee on animal experiments
of ‘‘Sapienza’’ University of Rome. All efforts were made
to minimize animal suffering and to reduce the number of
Brain Struct Funct
123

animals used. Animals were randomly assigned to one of
two experimental groups: standard reared animals, used as
controls (group name: C; 4 animals); enriched reared ani-
mals (group name: E; 10 animals).
Rearing conditions
Starting from the 21st post-natal day till the day of killing
(about the 120th post-natal day), the enriched rats were
housed in groups of ten animals in a large cage (100 9
50 9 70 cm). The cage had an additional floor made of a
galvanized wire mesh and connected by ramps of the same
material to create two interconnected levels. The cage
contained wood shavings, a running wheel, shelters and
plastic toys. Throughout the enrichment period, the running
wheel and shelters were always kept in the cage, while the
toys were changed twice a week. Once a week, the feeding
boxes and water bottles were moved to different cage
points to encourage foraging and explorative behavior.
Control rats reared in standard condition were housed in
groups of two animals in standard cages (40 9 26 9
18 cm) containing wood shaving bedding and only a
plastic tube as shelter. Feeding boxes and water bottles
were kept in a fixed position. The animals of both exper-
imental groups received the same type of food. Food and
water were provided ad libitum.
Tissue processing
On the 4th post-natal month, three animals of the E group
and three animals of the C group randomly selected were
deeply anesthetized (33 % chloral hydrate in water) to be
transcardially perfused with saline followed by 4 % para-
formaldehyde in phosphate-buffered saline (PBS; 4 �C, pH
7.5). Brains were removed, cryoprotected in 30 % buffered
sucrose at ?4 �C, frozen to be cut on a freezing microtome.
The posterior part of the brains embracing the cerebellum
was cut into 30-lm sagittal sections for calbindin immu-
nofluorescence to allow visualization of the entire dendritic
tree of Purkinje cells. Namely, sections were collected at
the level of left cerebellar hemisphere (3.90/3.40 mm lat-
eral to the midline) and cerebellar vermis (0.90/0.40 mm
lateral to the midline).
Immunofluorescence
To visualize Purkinje cells, sections were pre-incubated at
room temperature in 0.25 % Triton X-100 (Sigma-Aldrich
Chemie, Steinheim, Germany) and 10 % Normal Donkey
Serum (Vector Laboratories Inc, Burlingame, CA) in PBS
for 1 h. Sections then received a 2-day incubation at ?4 �C
in a solution composed of 0.25 % Triton X-100, 1 %
Normal Donkey Serum and the primary anti-calbindin
antibody (made in mouse, 1:2,000; Sigma-Aldrich Chemie,
Steinheim, Germany). After three rinses in PBS (10 min
each), sections were dark-incubated in a solution of 0.25 %
Triton X-100 and the Cy3-conjugated secondary anti-
mouse antibody (made in donkey, 1:200; Jackson Immu-
noresearch, BA) in PBS. After an incubation of 2 h at room
temperature, sections were mounted on slides and covers-
lipped by the anti-fading mounting medium Fluoromount
(Sigma-Aldrich Chemie, Steinheim, Germany). Specificity
of the staining was checked in sample sections by omitting
the primary antibody.
Confocal imaging
Imaging was performed through a LSM 700 confocal laser
scanning microscope (CLSM; Zeiss, Germany). Two dif-
ferent approaches were used: z-stack series with an
objective 639, allowing the visualization of the entire
dendritic tree of Purkinje cells, and z-stack series with an
objective 1009, allowing the visualization of the dendritic
tree distal portion. For the visualization of the entire den-
dritic tree, the images were acquired starting from the
Purkinje cell body to the pial surface. Such images were
used for spine density analysis on proximal and distal
dendritic compartments. For the visualization of the distal
portion of the dendritic tree, images were acquired starting
immediately below the pial surface. Such images were used
to analyze spine size. The following acquisition settings
were used for the entire dendritic tree: objective 639 oil
immersion NA 1.4; zoom factor 0.5; image format
1024 9 1024; sampled image area was x: 203.03 lm, y:
203.03 lm; pinhole 1 producing an optical section thick-
ness of 1.2 lm; z-spacing 0.5 lm; number of optical sec-
tions was about 10; pixel dwell time 50.4 ls. For the distal
dendritic tree we used: objective 1009 oil immersion NA
1.2; zoom factor 1.5; image format 1024 9 1024; sampled
image area was x: 46 lm, y: 46 lm; pinhole 1 producing an
optical section thickness of 0.8 lm; z-spacing 0.5 lm;
number of optical sections was about 16; pixel dwell time
50.4 ls. To generate projection images, the maximum
intensity algorithm was used, and the first and last images
of the z-series were cut so that only the central images were
used to generate the projection image. This procedure
allowed discarding images of the tissue surfaces eventually
containing broken or disconnected dendritic segments,
allowing a better image definition and a more reliable
analysis. The confocal image acquisitions were performed
so that all samples were imaged using consistent settings
for laser power and detector gain. To increase the resolu-
tion (Vecellio et al. 2000; Wallace et al. 2001; Wallace and
Bear 2004), the images were deconvolved by means of
blind deconvolution by using AutoQuant X3 (Media
Cybernetics Inc., Rockville, MD). For the production of
Brain Struct Funct
123

figures, a .tiff file of projection images was exported,
brightness and contrast of images were adjusted and final
figures were assembled using Adobe Photoshop 6 and
Adobe Illustrator 10. Image analysis was performed by
directly opening the acquisition file (.lsm), which retained
all the information about acquisition setting.
Image analysis
Image analysis was performed on proximal and distal
dendrites of Purkinje cells of both vermis and hemisphere.
Proximal and distal dendrites were identified according to
specific morphological features (Larramendi and Victor
1967; Palay and Chan-Palay 1974), and only those having a
diameter above and below 2 lm, respectively, were
retained for analysis (Bravin et al. 1999). Length and cal-
iber of sampled proximal and distal dendrites were mea-
sured using ImageJ 1.42q software (Abramoff et al. 2004).
Dendritic spines were identified as small protrusions from
the parent dendrite and their density was manually counted
offline in maximum intensity projection of the z-stacks by
using ImageJ 1.42q software. Care was taken to ensure that
each spine was counted only once by following its pro-
jection course through the stack of z-sections. Evaluation
of spine density was performed on segments of about
20 lm in length of dendrites of a single Purkinje cell.
Since significant changes in spine density were found
only in Purkinje cell distal dendrites both in vermis and
hemisphere, the evaluation of spine size was executed only
on distal compartment. Spine size was evaluated using the
Filament Tracer module of the Imaris Suite 7.4� (Bitplane
A.G., Zurich, Switzerland) software to generate spine
reconstructions of the acquired z-stack images. The ana-
lysis procedure was performed under visual control to
determine thresholds subtracting background noise and
taking into account dendritic structures. The spine length
was set at a minimum of 0.2 lm and a maximum of 1.5 lm
(i.e., the shortest quantifiable spines were 0.2 lm and the
longest ones 1.5 lm in length). These thresholds were
settled on the basis of previous not automated manual
measurements performed on the same areas revealing that
no protrusion to be considered as a spine longer than
1.2–1.3 lm was ever observed. In fact, the occasionally
encountered apparently longer protrusions had to be
referred to two or more spines located too closely or even
overlapping on different focus plans, as evidenced by a
detailed visual inspection. During image processing, the
images were compared with the original raw data to make
sure that no structure not seen in the original data was
introduced or vice versa that a structure present in the
original data was erroneously removed. Once determined
thresholds that preserve dendrite and spine features, they
were applied to all images. Projection images were created
by applying maximum intensity projection algorithm to
z-series and volume rendering of the z-stacks were gener-
ated with Filament modules. Analyzed immunofluorescent
images were overlaid with volume rendering images gen-
erated by the filament analysis module to visually verify
the accuracy of the dendrites and spines reconstruction.
Notably, the high spine density in Purkinje cell distal
dendrites creates a challenge for analysis, as single spines
may overlap to form a confounding multi-spine protrusion.
To overcome such a trouble as much as possible, image
analyses were separately performed by two experimenters
blind to the experimental treatment. Measurements of spine
values were considered reliable only when their measure
evaluations were consistent.
As parameters, we evaluated mean spine density (num-
ber of spines per lm of dendritic length), mean spine area
(surface of the spine from the dendritic surface to the spine
ending), mean spine length (extension of the entire spine
from the dendritic surface to the spine ending), and mean
spine head diameter (diameter of the spine bulbous ending).
Statistical analysis
Data are presented as mean ± SEM. Statistical analyses
were performed using the software SPSS 8. Data were first
tested for normality (Will–Shapiro’s test) and homosce-
dasticity (Levene’s test). To verify the appropriateness of
applying nested-design ANOVAs taking into account the
influence of the random factor ‘‘animal’’ on the data, the
significance level of the interaction between the random
factor ‘‘animal’’ and the fixed factor ‘‘group’’ in two-way
ANOVAs was preliminarily evaluated. Since no significant
interaction was found for any parameter, no nested-design
ANOVA was applied. One-way ANOVAs with group (C,
E) as fixed factor were then performed on spine density, for
the cerebellar region (vermis or hemisphere) and neuronal
compartment (proximal or distal dendrites). Size parame-
ters were analyzed by MANOVA with group (C, E) as
fixed factor and spine area and length as dependent vari-
ables for distal dendrites of both cerebellar regions (vermis
or hemisphere). Tukey’s test was used for post hoc com-
parisons. p values B0.05 were considered statistically sig-
nificant. Furthermore, to reveal the magnitude of the
eventual changes, percentage differences of spine values
between E and C groups were calculated.
Results
Methodological approach
Calbindin-immunostained cerebellar sections showed an
intense fluorescence of Purkinje cell bodies, dendrites and
Brain Struct Funct
123

spines. As visible in Fig. 1, the superimposition of the
projection images of z-stack series taken through the
Purkinje cell distal dendritic region with the volume ren-
dering generated by the Filament Analysis module showed
the accuracy of the digital reconstruction relative to the
native image. Broken dendrites or dendritic branches not
perfectly comprised in the acquisition volumes were
excluded from the analysis. Closer analysis of dendrite
small segments (Fig. 1d–f) revealed accurate identification
of spines. In several cases, the immunostained spines
appeared as fluorescent point-like regions not connected to
the dendrite, while the volume rendering showed spines
attached to the dendrite. By increasing the brightness of
the image to fluorescence saturation levels, the spines
were shown to be really connected to the dendrite, indi-
cating that the fluorescence intensity levels of the spines
in the native image were too low to be well appreciated by
the human eye. A clear advantage of the described
approach was the possibility to include in the analysis the
high majority of fluorescent spines present in the imaging
field.
Morphological changes in vermian and hemispheric
regions
Rearing in an enriched environment provoked an
enhancement of spine density on both proximal and distal
compartments of Purkinje cell dendrites in the hemisphere
and only on distal compartment in the vermis (Fig. 2). As
for spine size, a significant increase of area, length and
head diameter was found in the spines on distal dendrites of
Purkinje cells in both vermis and hemisphere (Fig. 3).
Fig. 1 Representative deconvolved images and rendering of distal
dendrites of a calbindin-immunostained Purkinje cell. a Maximum
projection of a confocal z-stack series showing intense spine labeling.
The pial surface is on the left upper part of the image. b Volume
rendering of the maximum projection image created by Filament
Tracer module of Imaris software package. c Overlay of the
maximum projection image and the relative volume rendering. Note
that the areas where the calbindin labeling was not sufficiently intense
or the dendrites that were damaged by cutting procedures were not
included in the analysis. d–f Higher magnification of the boxed area
in a–c to illustrate the accuracy of the automated spine identification
procedure. Scale bars a = 13 lm; d = 0.8 lm
Fig. 2 Effects of the exposure to a standard (C) or an enriched
(E) environment on Purkinje cell spine density. Histograms show the
mean spine density of Purkinje cell proximal and distal dendrites in
the vermis (a1; a2) and the hemisphere (b1; b2). Data are shown as
mean ± SEM. Asterisks indicate significant differences between
groups (*p \ 0.05; ***p \ 0.0005)
Brain Struct Funct
123
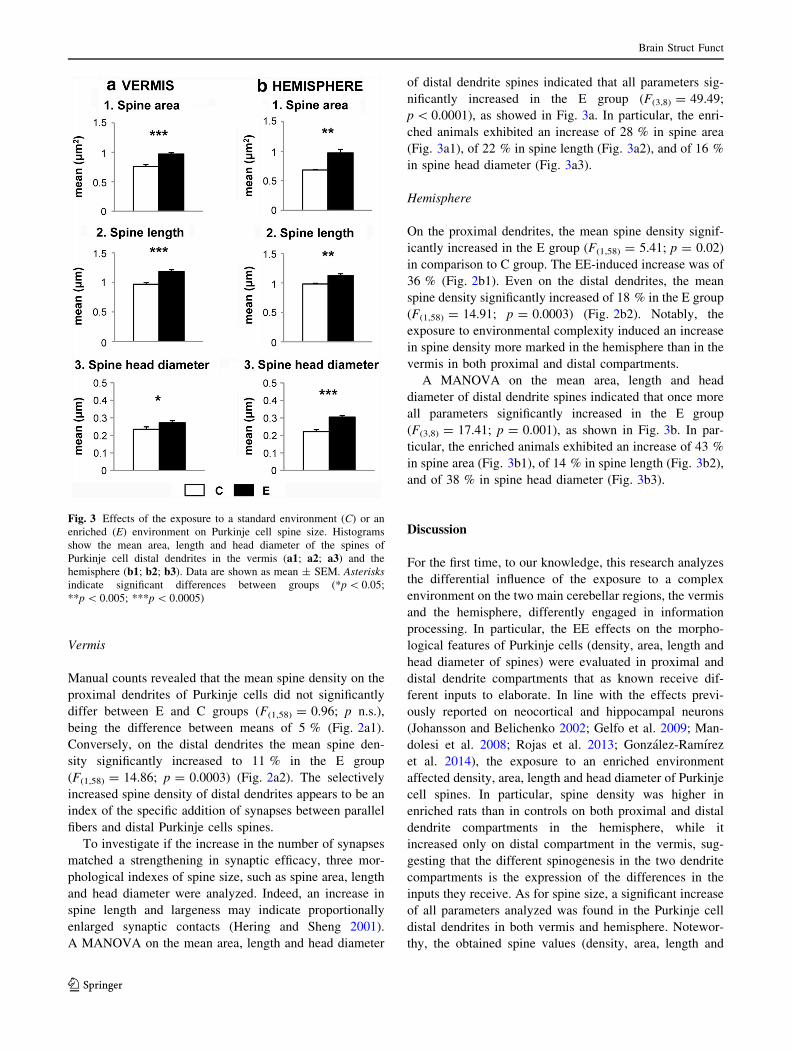
Vermis
Manual counts revealed that the mean spine density on the
proximal dendrites of Purkinje cells did not significantly
differ between E and C groups (F(1,58) = 0.96; p n.s.),
being the difference between means of 5 % (Fig. 2a1).
Conversely, on the distal dendrites the mean spine den-
sity significantly increased to 11 % in the E group
(F(1,58) = 14.86; p = 0.0003) (Fig. 2a2). The selectively
increased spine density of distal dendrites appears to be an
index of the specific addition of synapses between parallel
fibers and distal Purkinje cells spines.
To investigate if the increase in the number of synapses
matched a strengthening in synaptic efficacy, three mor-
phological indexes of spine size, such as spine area, length
and head diameter were analyzed. Indeed, an increase in
spine length and largeness may indicate proportionally
enlarged synaptic contacts (Hering and Sheng 2001).
A MANOVA on the mean area, length and head diameter
of distal dendrite spines indicated that all parameters sig-
nificantly increased in the E group (F(3,8) = 49.49;
p \ 0.0001), as showed in Fig. 3a. In particular, the enri-
ched animals exhibited an increase of 28 % in spine area
(Fig. 3a1), of 22 % in spine length (Fig. 3a2), and of 16 %
in spine head diameter (Fig. 3a3).
Hemisphere
On the proximal dendrites, the mean spine density signif-
icantly increased in the E group (F(1,58) = 5.41; p = 0.02)
in comparison to C group. The EE-induced increase was of
36 % (Fig. 2b1). Even on the distal dendrites, the mean
spine density significantly increased of 18 % in the E group
(F(1,58) = 14.91; p = 0.0003) (Fig. 2b2). Notably, the
exposure to environmental complexity induced an increase
in spine density more marked in the hemisphere than in the
vermis in both proximal and distal compartments.
A MANOVA on the mean area, length and head
diameter of distal dendrite spines indicated that once more
all parameters significantly increased in the E group
(F(3,8) = 17.41; p = 0.001), as shown in Fig. 3b. In par-
ticular, the enriched animals exhibited an increase of 43 %
in spine area (Fig. 3b1), of 14 % in spine length (Fig. 3b2),
and of 38 % in spine head diameter (Fig. 3b3).
Discussion
For the first time, to our knowledge, this research analyzes
the differential influence of the exposure to a complex
environment on the two main cerebellar regions, the vermis
and the hemisphere, differently engaged in information
processing. In particular, the EE effects on the morpho-
logical features of Purkinje cells (density, area, length and
head diameter of spines) were evaluated in proximal and
distal dendrite compartments that as known receive dif-
ferent inputs to elaborate. In line with the effects previ-
ously reported on neocortical and hippocampal neurons
(Johansson and Belichenko 2002; Gelfo et al. 2009; Man-
dolesi et al. 2008; Rojas et al. 2013; Gonzalez-Ramırez
et al. 2014), the exposure to an enriched environment
affected density, area, length and head diameter of Purkinje
cell spines. In particular, spine density was higher in
enriched rats than in controls on both proximal and distal
dendrite compartments in the hemisphere, while it
increased only on distal compartment in the vermis, sug-
gesting that the different spinogenesis in the two dendrite
compartments is the expression of the differences in the
inputs they receive. As for spine size, a significant increase
of all parameters analyzed was found in the Purkinje cell
distal dendrites in both vermis and hemisphere. Notewor-
thy, the obtained spine values (density, area, length and
Fig. 3 Effects of the exposure to a standard environment (C) or an
enriched (E) environment on Purkinje cell spine size. Histograms
show the mean area, length and head diameter of the spines of
Purkinje cell distal dendrites in the vermis (a1; a2; a3) and the
hemisphere (b1; b2; b3). Data are shown as mean ± SEM. Asterisks
indicate significant differences between groups (*p \ 0.05;
**p \ 0.005; ***p \ 0.0005)
Brain Struct Funct
123

head diameter) are in agreement with the values reported in
previous studies (Harris and Stevens 1988; Lee et al. 2007;
Sdrulla and Linden 2007; Petrinovic et al. 2013).
As previously demonstrated, the exposure to EE evokes
a different extent of spinogenesis in the apical and basal
arborizations of neocortical pyramidal neurons. Namely,
enriched rats showed an increased spine density in both
apical and basal arborizations of pyramidal neurons of the
layer III in the parietal cortex and mainly in apical arbor-
ization of the pyramidal neurons of layers III and V in
frontal cortex (Gelfo et al. 2009). Thus, the different spine
density in the two dendritic compartments of the vermian
and hemispherical regions seems to be related to the EE-
potentiated locomotor activity, voluntary motility and
motor learning resulting in different morphological effects
at cerebellar level. Rearing in large groups, foraging wide
spaces, experiencing multiple sensorial stimuli require
advanced sensory-motor integration, effective planning of
motor strategies, fine matching of actions with goal, con-
tinuous adaptation of behavior to the multifarious context.
Thus, environmental complexity primarily influences those
neural circuits that have access to sensory information and
at same time modulate motor output. The cerebellar net-
works do meet such a requirement. The strengthening of
the cerebellar synapses has positive repercussions on the
wide cortico-subcortical networks, supporting the EE-
induced high-level performances. Indeed, effective infor-
mation processing, highly coordinated movements and
complex motor adjustments were previously demonstrated
in animals exposed to the very same enrichment protocol.
Namely, the enriched animals quickly acquire tuned navi-
gational strategies, improved procedural competencies
and working memory abilities. Furthermore, when the
requirements of the context change, they promptly are able
to reorganize their strategies by shifting from prevalently
using spatial procedures to applying mnesic competencies
(Leggio et al. 2005; De Bartolo et al. 2008; Mandolesi et al.
2008; Petrosini et al. 2009; Foti et al. 2011). The beneficial
effects of complex experiences appear to be linked to the
strengthening of not only the neocortical and hippocampal
synapses, as already demonstrated, but also of the Purkinje
cell synapses. Specifically, the parallel fiber–Purkinje cell
synapses (distal dendrites) provide contextual information
and the climbing fiber–Purkinje cell synapses (proximal
dendrites) provides signal ‘‘errors’’ adapting the behavior
to requirement of ever-changing environments (Ito 1984;
Hansel and Linden 2000, 2001; Ramnani 2006). It is
intriguing that the exposure to complex environments
enhances synapse formation and plasticity either in the
vermis involved in balance and locomotion and in the
hemisphere involved in complex motor adaptations and
acquisition of new motor strategies. Interestingly, while in
the vermis there was an increased spine density only in
Purkinje cell distal dendrites, in the hemisphere such an
increase was present also in proximal dendrites. This
finding suggests that in the hemisphere, the environmental
complexity specifically strengthened the teaching line
provided by the climbing fibers (Marr 1969; Albus 1971).
The bidirectional connections of cerebellar hemispheres
with neocortical areas, as the prefrontal, premotor, primary
motor and parietal cortices, involved in all facets of motor
function (Voogd 1995; Schmahmann 1996; Morton and
Bastian 2004; Manto and Oulad Ben Taib 2010) are con-
sistent with the striking EE influence on Purkinje cell distal
and proximal dendrites of hemispherical regions. The
present results complement the increased number of syn-
apses per Purkinje cell or the synaptic efficacy changes
following motor learning or classical conditioning previ-
ously described (Sakurai 1987; Black et al. 1990; Schreurs
et al. 1998; Velazquez-Zamora et al. 2011).
As described, the enhanced spine density was accom-
panied by an increased spine size in distal dendrites of both
vermis and hemisphere of enriched rats. An increase in
spine length and largeness may be considered an indication
of proportionally enlarged synaptic contacts (Hering and
Sheng 2001). As evidenced by the findings provided by
biophysics and molecular biology studies, large spines
abundantly express AMPA receptors and are associated
with large postsynaptic density, producing consequently
large AMPA receptor-mediated currents (Matsuzaki et al.
2001, 2004; Kasai et al. 2003; Holtmaat et al. 2005;
Noguchi et al. 2005; Bourne and Harris 2007). On such a
basis, it is intriguing to suggest that the larger spines of
enriched animals are remarkably persistent and function-
ally stronger in their response to glutamate and in local
regulation of intracellular calcium, and that the spine
enlargement may be essential for an increased postsynaptic
response in LTP (Grutzendler et al. 2002; Kasai et al. 2003;
Holtmaat et al. 2005). As advanced by Matsuzaki et al.
(2004), large spines act as ‘‘memory units’’ and are the
structural basis of long-term memory (Kasai et al. 2003;
Matsuzaki et al. 2004; Gonzalez-Burgos et al. 2011). Thus,
enlarged spines can stabilize and retain long-term infor-
mation and become critical for information storage (Kasai
et al. 2003). Spine density and morphology are widely
modulated by environmental inputs in the form of synaptic
activity, which is central to mnesic traces formation and
other adaptive brain changes. The new spines persist at
long last after EE and the performance of enriched animals
is positively associated with the extent of new spine for-
mation, as we previously demonstrated following the
exposure to the same EE paradigm (Mandolesi et al. 2008).
The increase in spine density is obviously related to a
major proximity of the spines. This is particularly inter-
esting, since neighboring spines are retained to function
within the same neural circuit and transmit similar
Brain Struct Funct
123

information to the postsynaptic neuron as it has been pro-
posed to occur in clustered spines (Yang et al. 2009; Fu
et al. 2012).
In conclusion, the interaction with a complex environ-
ment appears to induce tuned neuronal activity that facil-
itates the remodeling of synaptic connectivity mainly in the
structures more directly involved in the experience-
dependent events. In this way, the external stimuli drive a
primarily instructive action that determines the actual
number and size of the adaptive spines. The influence of
external stimuli on the cellular/molecular mechanisms that
regulate spine metabolism has been object of many studies
(Klintsova and Greenough 1999; Lee et al. 2012; Lai and
Ip 2013). In particular, the EE stimulates changes in the
expression of neuronal growth genes, regulatory substances
(as myelin-associated proteins and extracellular matrix
components) and neurotrophins acting on neuronal plas-
ticity and connection patterns. As previously demonstrated,
in enriched rats the cerebellum was the brain region where
BDNF and NGF levels were more increased as compared
to the levels of frontal cortex, hippocampus and striatum
(Angelucci et al. 2009). Thus, the EE-induced effects on
Purkinje cell spines observed in the present research might
be linked also to the modification of neurotrophic factor
levels, dampening of growth-inhibitory mechanisms and
shifting of the balance between synthesis and removal of
matrix components to reorganize the cerebellar output to
the cerebral cortex, as recently described in a different
model of cerebellar plasticity (Foscarin et al. 2011).
Acknowledgments We are grateful to Mr. Maurizio Abbate for the
technical help with images deconvolution. The research was sup-
ported by MIUR fund to LP.
Conflict of interest The authors declare that they have no conflict
of interest.
Ethical standards The animals used in the study described in this
manuscript were maintained according to the guidelines for ethical
conduct developed by the European Communities Council Directive
of 22 September 2010 (2010/63/EU). All animal protocols used have
been approved by the Ethical Committee on animal experiments of
‘‘Sapienza’’ University of Rome. The manuscript does not contain
clinical studies or patient data.
References
Abramoff MD, Magalhaes PJ, Ram SJ (2004) Image processing with
ImageJ. Biophotonics Int 11:36–42
Albus JS (1971) A theory of cerebellar function. Math Biosci
10:25–61
Angelucci F, De Bartolo P, Gelfo F, Foti F, Cutuli D, Bossu P,
Caltagirone C, Petrosini L (2009) Increased concentrations of
nerve growth factor and brain-derived neurotrophic factor in the
rat cerebellum after exposure to environmental enrichment.
Cerebellum 8:499–506
Arai JA, Feig LA (2011) Long-lasting and transgenerational effects of
an environmental enrichment on memory formation. Brain Res
Bull 85:30–35
Bae J, Sung BH, Cho IH, Kim S-M, Song WK (2012) NESH regulates
dendritic spine morphology and synapse formation. PLoS One
7:e34677
Black JE, Isaacs KR, Anderson BJ, Alcantara AA, Greenough WT
(1990) Learning causes synaptogenesis, whereas motor activity
causes angiogenesis, in cerebellar cortex of adult rats. Proc Natl
Acad Sci USA 87:5568–5572
Bourne J, Harris KM (2007) Do thin spines learn to be mushroom
spines that remember? Curr Opin Neurobiol 17:381–386
Bravin M, Morando L, Vercelli A, Rossi F, Strata P (1999) Control of
spine formation by electrical activity in the adult rat cerebellum.
Proc Natl Acad Sci USA 96:1704–2709
Bundman MC, Gall CM (1994) Ultrastructural plasticity of the
dentate gyrus granule cells following recurrent limbic seizures:
II. Alterations in somatic synapses. Hippocampus 4:611–622
Caporali P, Cutuli D, Gelfo F, Laricchiuta D, Foti F, De Bartolo P,
Mancini L, Angelucci F, Petrosini L (2014) Pre-reproductive
maternal enrichment influences offspring developmental trajec-
tories: motor behavior and neurotrophin expression. Front Behav
Neurosci 8:195
Chambers WW, Sprague JM (1955) Functional localization in the
cerebellum. I. Organization in longitudinal cortico-nuclear zones
and their contribution to the control of posture, both extrapy-
ramidal and pyramidal. J Comp Neurol 103:105–129
Cutuli D, Rossi S, Burello L, Laricchiuta D, De Chiara V, Foti F, De
Bartolo P, Musella A, Gelfo F, Centonze D, Petrosini L (2011)
Before or after does it matter? Different protocols of environ-
mental enrichment differently influence motor, synaptic and
structural deficits of cerebellar origin. Neurobiol Dis 42:9–20
De Bartolo P, Leggio MG, Mandolesi L, Foti F, Gelfo F, Ferlazzo F,
Petrosini L (2008) Environmental enrichment mitigates the
effects of basal forebrain lesions on cognitive flexibility.
Neuroscience 154:444–453
De Bartolo P, Gelfo F, Burello L, De Giorgio A, Petrosini L, Granato
A (2011) Plastic changes in striatal fast-spiking interneurons
following hemicerebellectomy and environmental enrichment.
Cerebellum 10:624–632
Federmeier KD, Kleim JA, Greenough WT (2002) Learning-induced
multiple synapse formation in rat cerebellar cortex. Neurosci
Lett 332:180–184
Ferrer I, Kulisevski J, Vazquez J, Gonzalez G, Pineda M (1988)
Purkinje cells in degenerative disease of the cerebellum and its
connections: a Golgi study. Clin Neuropathol 7:22–28
Ferrer I, Genıs D, Davalos A, Bernado L, Sant F, Serrano T (1994)
The Purkinje cell in olivopontocerebellar atrophy. A Golgi and
immunocytochemical study. Neuropathol Appl Neurobiol
20:38–46
Fiala JC, Spacek J, Harris KM (2002) Dendritic spine pathology:
cause or consequence of neurological disorders? Brain Res Rev
39:29–54
Foscarin S, Ponchione D, Pajaj E, Leto K, Gawlak M, Wilczynski
GM, Rossi F, Carulli D (2011) Experience-dependent plasticity
and modulation of growth regulatory molecules at central
synapses. PLoS One 6:e16666
Foti F, Laricchiuta D, Cutuli D, De Bartolo P, Gelfo F, Angelucci F,
Petrosini L (2011) Exposure to an enriched environment
accelerates recovery from cerebellar lesion. Cerebellum
10:104–119
Fu M, Yu X, Lu J, Zuo Y (2012) Repetitive motor learning induces
coordinated formation of clustered dendritic spines in vivo.
Nature 483:92–95
Gelfo F, De Bartolo P, Giovine A, Petrosini L, Leggio MG (2009)
Layer and regional effects of environmental enrichment on the
Brain Struct Funct
123

pyramidal neuron morphology of the rat. Neurobiol Learn Mem
91:353–365
Gonzalez-Burgos I, Gonzalez-Tapia D, Zamora DA, Feria-Velasco A,
Beas-Zarate C (2011) Guided motor training induces dendritic
spine plastic changes in adult rat cerebellar Purkinje cells.
Neurosci Lett 491:216–220
Gonzalez-Ramırez MM, Velazquez-Zamora DA, Olvera-Cortes ME,
Gonzalez-Burgos I (2014) Changes in the plastic properties of
hippocampal dendritic spines underlie the attenuation of place
learning in healthy aged rats. Neurobiol Learn Mem 109:94–103
Gray EG (1959) Electron microscopy of synaptic contacts on dendrite
spines of the cerebral cortex. Nature 183:1592–15933
Grutzendler J, Kasthuri N, Gan WB (2002) Long-term dendritic spine
stability in the adult cortex. Nature 420:812–816
Hansel C, Linden DJ (2000) Long-term depression of the cerebellar
climbing fiber–Purkinje neuron synapse. Neuron 26:473–482
Hansel C, Linden DJ, D’Angelo E (2001) Beyond parallel fiber LTD:
the diversity of synaptic and non-synaptic plasticity in the
cerebellum. Nat Neurosci 4:467–475
Harris KM, Kater SB (1994) Dendritic spines: cellular specializations
imparting both stability and flexibility to synaptic function. Annu
Rev Neurosci 17:341–371
Harris KM, Stevens JK (1988) Dendritic spines of rat cerebellar
Purkinje cells: serial electron microscopy with reference to their
biophysical characteristics. J Neurosci 8:4455–4469
Hering H, Sheng M (2001) Dendritic spines: structure, dynamics and
regulation. Nat Rev Neurosci 2:880–888
Holtmaat AJ, Trachtenberg JT, Wilbrecht L, Shepherd GM, Zhang X,
Knott GW, Svoboda K (2005) Transient and persistent dendritic
spines in the neocortex in vivo. Neuron 45:279–291
Hosokawa T, Rusakov DA, Bliss TVP, Fine A (1995) Repeated
confocal imaging of individual dendritic spines in the living
hippocampal slice: evidence for changes in length and orienta-
tion associated with chemically induced LTP. J Neurosci
15:5560–5573
Ito M (1984) The modifiable neuronal network of the cerebellum. Jpn
J Physiol 34:781–792
Johansson BB, Belichenko PV (2002) Neuronal plasticity and
dendritic spines: effect of environmental enrichment on intact
and post-ischemic rat brain. J Cereb Blood Flow Metab
22:89–96
Kasai H, Matsuzaki M, Noguchi J, Yasumatsu N, Nakahara H (2003)
Structure-stability-function relationships of dendritic spines.
Trends Neurosci 26:360–368
Kim HT, Kim IH, Lee KJ, Lee JR, Park SK, Chun YH, Kim H, Rhyu
IJ (2002) Specific plasticity of parallel fiber/Purkinje cell spine
synapses by motor skill learning. Neuro Rep 13:1607–1610
Klintsova AY, Greenough WT (1999) Synaptic plasticity in cortical
systems. Curr Opin Neurobiol 9:203–208
Koch C, Poggio T (1983) A theoretical analysis of electrical
properties of spines. Proc R Soc Lond B Biol Sci 218:455–477
Kozorovitskiy Y, Gross CG, Kopil C, Battaglia L, McBreen M,
Stranahan AM, Gould E (2005) Experience induces structural
and biochemical changes in the adult primate brain. Proc Natl
Acad Sci USA 102:17478–17482
Lai KO, Ip NY (2013) Structural plasticity of dendritic spines: the
underlying mechanisms and its dysregulation in brain disorders.
Biochim Biophys Acta 1832:2257–2263
Larramendi EM, Victor T (1967) Synapses on the Purkinje cell spines
in the mouse. An electronmicroscopic study. Brain Res 5:15–30
Lee KJ, Jung JG, Arii T, Imoto K, Rhyu IJ (2007) Morphological
changes in dendritic spines of Purkinje cells associated with
motor learning. Neurobiol Learn Mem 88:445–450
Lee KF, Soares C, Beıque JC (2012) Examining form and function of
dendritic spines. Neural Plast 2012:704103
Leggio MG, Mandolesi L, Federico F, Spirito F, Ricci B, Gelfo F,
Petrosini L (2005) Environmental enrichment promotes
improved spatial abilities and enhanced dendritic growth in the
rat. Behav Brain Res 163:78–90
Liu N, He S, Yu X (2012) Early natural stimulation through
environmental enrichment accelerates neuronal development in
the mouse dentate gyrus. PLoS One 7:e30803
Lonetti G, Angelucci A, Morando L, Boggio EM, Giustetto M,
Pizzorusso T (2010) Early environmental enrichment moderates
the behavioral and synaptic phenotype of MeCP2 null mice. Biol
Psychiatry 67:657–665
Mandolesi L, De Bartolo P, Foti F, Gelfo F, Federico F, Leggio MG,
Petrosini L (2008) Environmental enrichment provides a cogni-
tive reserve to be spent in the case of brain lesion. J Alzheimers
Dis 15:11–28
Manto M, Oulad Ben Taib N (2010) Cerebellar nuclei: key roles for
strategically located structures. Cerebellum 9:17–21
Marr D (1969) A theory of cerebellar cortex. J Physiol 202:437–470
Matsuzaki M, Ellis-Davies GC, Nemoto T, Miyashita Y, Iino M,
Kasai H (2001) Dendritic spine geometry is critical for AMPA
receptor expression in hippocampal CA1 pyramidal neurons. Nat
Neurosci 4:1086–1092
Matsuzaki M, Honkura N, Ellis-Davies GC, Kasai H (2004) Structural
basis of long-term potentiation in single dendritic spines. Nature
429:761–766
Morando L, Cesa R, Rasetti R, Harvey R, Strata P (2001) Role of
glutamate delta -2 receptors in activity-dependent competition
between heterologous afferent fibers. Proc Natl Acad Sci USA
98:9954–9959
Morando L, Cesa R, Harvey RJ, Strata P (2005) Spontaneous
electrical activity and structural plasticity in the mature
cerebellar cortex. Ann N Y Acad Sci 1048:131–140
Morton SM, Bastian AJ (2004) Cerebellar control of balance and
locomotion. Neuroscientist 10:247–259
Moser MB, Trommald M, Andersen P (1994) An increase in dendritic
spine density on hippocampal CA1 pyramidal cells following
spatial learning in adult rats suggests the formation of new
synapses. Proc Natl Acad Sci USA 91:12673–12675
Muhammad A, Mychasiuk R, Hosain S, Nakahashi A, Carroll C, Gibb
R, Kolb B (2013) Training on motor and visual spatial learning
tasks in early adulthood produces large changes in dendritic
organization of prefrontal cortex and nucleus accumbens in rats
given nicotine prenatally. Neuroscience 252:178–189
Nithianantharajah J, Hannan AJ (2006) Enriched environments,
experience-dependent plasticity and disorders of the nervous
system. Nat Rev Neurosci 7:697–709
Nithianantharajah J, Levis H, Murphy M (2004) Environmental
enrichment results in cortical and subcortical changes in levels of
synaptophysin and PSD-95 proteins. Neurobiol Learn Mem
81:200–210
Noguchi J, Matsuzaki M, Ellis-Davies GC, Kasai H (2005) Spine-
neck geometry determines NMDA receptor-dependent Ca2?
signaling in dendrites. Neuron 46:609–622
Palay SL, Chan-Palay V (1974) Cerebellar cortex: cytology and
organization. Springer, Berlin
Petrinovic MM, Hourez R, Aloy EM, Dewarrat G, Gall D, Weinmann
O, Gaudias J, Bachmann LC, Schiffmann SN, Vogt KE, Schwab
ME (2013) Neuronal Nogo-A negatively regulates dendritic
morphology and synaptic transmission in the cerebellum. Proc
Natl Acad Sci USA 110:1083–1088
Petrosini L, De Bartolo P, Foti F, Gelfo F, Cutuli D, Leggio MG,
Mandolesi L (2009) On whether the environmental enrichment may
provide cognitive and brain reserves. Brain Res Rev 61:221–239
Pysh JJ, Weiss GM (1979) Exercise during development induces an
increase in Purkinje cell dendritic tree size. Science 206:230–232
Brain Struct Funct
123

Ramnani N (2006) The primate cortico-cerebellar system: anatomy
and function. Nat Rev Neurosci 7:511–522
Rhyu IJ, Oda S, Uhm CS, Kim H, Suh YS, Abbott LC (1999a)
Morphologic investigation of rolling mouse Nagoya (tg(rol)/
tg(rol)) cerebellar Purkinje cells: an ataxic mutant, revisited.
Neurosci Lett 266:49–52
Rhyu IJ, Abbott LC, Walker DB, Sotelo C (1999b) An ultrastructural
study of granule cell/Purkinje cell synapses in tottering (tg/tg),
leaner (tg(la)/tg(la)) and compound heterozygous tottering/leaner
(tg/tg(la)) mice. Neuroscience 90:717–728
Rojas JJ, Deniz BF, Miguel PM, Diaz R, Hermel Edo E, Achaval M,
Netto CA, Pereira LO (2013) Effects of daily environmental
enrichment on behavior and dendritic spine density in hippo-
campus following neonatal hypoxia-ischemia in the rat. Exp
Neurol 241:25–33
Rosenzweig MR, Bennett EL (1996) Psychobiology of plasticity:
effects of training and experience on brain and behavior. Behav
Brain Res 78:57–65
Rosenzweig MR, Krech D, Bennet EL, Diamond MC (1962) Effects
of environmental complexity and train on brain chemistry and
anatomy. J Comp Physiol Psychol 55:429–437
Sakurai M (1987) Synaptic modification of parallel fiber-Purkinje cell
transmission in in vitro guinea-pig cerebellar slices. J Physiol
394:463–480
Scarmeas N, Stern Y (2003) Cognitive reserve and lifestyle. J Clin
Exp Neuropsychol 25:625–633
Schmahmann JD (1996) From movement to thought: anatomic
substrates of the cerebellar contribution to cognitive processing.
Hum Brain Mapp 4:174–198
Schreurs BG, Gusev PA, Tomsic D, Alkon DL, Shi T (1998)
Intracellular correlates of acquisition and long-term memory of
classical conditioning in Purkinje cell dendrites in slices of rabbit
cerebellar lobule HVI. J Neurosci 18:5498–5507
Sdrulla AD, Linden DJ (2007) Double dissociation between long-term
depression and dendritic spine morphology in cerebellar Purkinje
cells. Nat Neurosci 10:546–548
Sotelo C, Hillman DE, Zamora AJ, Llinas R (1975) Climbing fiber
deafferentation: its action on Purkinje cell dendritic spines. Brain
Res 98:574–581
Stern Y (2002) What is cognitive reserve? Theory and research
application of the reserve concept. J Int Neuropsychol Soc
8:448–460
Stern Y (2003) The concept of cognitive reserve: a catalyst for
research. J Clin Exp Neuropsychol 25:589–593
Stern Y (2006) Cognitive reserve and Alzheimer disease. Alzheimer
Dis Assoc Disord 20:112–117
Sugawara T, Hisatsune C, Le TD, Hashikawa T, Hirono M, Hattori
M, Nagao S, Mikoshiba K (2013) Type 1 inositol trisphosphate
receptor regulates cerebellar circuits by maintaining the spine
morphology of Purkinje cells in adult mice. J Neurosci
33:12186–12196
Thach WT, Goodkin HP, Keating JG (1992) The cerebellum and the
adaptive coordination of movement. Annu Rev Neurosci
15:403–442
Tremml P, Lipp HP, Muller U, Wolfer DP (2002) Enriched early
experiences of mice underexpressing the beta-amyloid precursor
protein restore spatial learning capabilities but not normal open
field behavior of adult animals. Genes Brain Behav 1:230–241
Turner DA (1984) Conductance transients onto dendritic spines in a
segmental cable model of hippocampal neurons. Biophys J
46:85–96
Van Praag H, Kempermann G, Gage FH (2000) Neural consequences
of environmental enrichment. Nature Rev Neurosci 1:191–198
Vazquez-Sanroman D, Sanchis-Segura C, Toledo R, Hernandez ME,
Manzo J, Miquel M (2013) The effects of enriched environment
on BDNF expression in the mouse cerebellum depending on the
length of exposure. Behav Brain Res 243:118–128
Vecellio M, Schwaller B, Meyer M, Hunziker W, Celio MR (2000)
Alterations in Purkinje cell spines of calbindin D-28k and
parvalbumin knock-out mice. Eur J Neurosci 12:945–954
Velazquez-Zamora DA, Martınez-Degollado M, Gonzalez-Burgos I
(2011) Morphological development of dendritic spines on rat
cerebellar Purkinje cells. Int J Dev Neurosci 29:515–520
Voogd J (1995) Cerebellum. In: Paxinos G (ed) The rat nervous
system, 2nd edn. Academic press, San Diego, pp 277–308
Wallace W, Bear MF (2004) A morphological correlate of synaptic
scaling in visual cortex. J Neurosci 24:6928–6938
Wallace W, Schaefer LH, Swedlow JR (2001) A workingperson’s
guide to deconvolution in light microscopy. Biotechniques
31:1076–1078 1080, 1082 passim
Whalley L, Starr J, Athawes R, Hunter D, Pattie A, Deary IJ
(2000) Childhood mental ability and dementia. Neurology
55:1455–1459
Whalley LJ, Deary IJ, Appleton CL, Starr JM (2004) Cognitive
reserve and the neurobiology of cognitive aging. Ageing Res
Rev 3:369–382
Yang G, Pan F, Gan WB (2009) Stably maintained dendritic spines
are associated with lifelong memories. Nature 462:920–924
Yau SY, Lau BW, Tong JB, Wong R, Ching YP, Qiu G, Tang SW,
Lee TM, So KF (2011) Hippocampal neurogenesis and dendritic
plasticity support running-improved spatial learning and depres-
sion-like behaviour in stressed rats. PLoS One 6:e24263
Zhao C, Warner-Schmidt J, Duman RS, Gage FH (2012) Electro-
convulsive seizure promotes spine maturation in newborn
dentate granule cells in adult rat. Dev Neurobiol 72:937–942
Brain Struct Funct
123
Related Documents











