ACTIVATION TRANSCRIPTION FACTOR-3 ACTIVATION AND THE DEVELOPMENT OF SPINAL CORD DEGENERATION IN A RAT MODEL OF AMYOTROPHIC LATERAL SCLEROSIS A. MALASPINA,* S. F. A. NGOH, R. E. WARD, J. C. E. HALL, F. W. D. TAI, P. K. YIP, C. JONES, N. JOKIC, S. A. AVERILL, A. T. MICHAEL-TITUS AND J. V. PRIESTLEY Centre for Neuroscience and Trauma, Barts and the London School of Medicine and Dentistry, Queen Mary University of London, Blizard Institute, 4 Newark Street, London E1 2AT, UK Abstract—It has been reported that an early activation of glial fibrillary acid protein (GFAP) in astroglial cells occurs simul- taneously in peripheral nerves and spinal cord from the G93A SOD1 mouse model of amyotrophic lateral sclerosis (ALS), an invariably fatal neurodegenerative disorder. In ALS, the contribute to the pathological process of different cell types varies according to the disease stage, with a florid immune response in spinal cord at end stage disease. In this study, we have mapped in different anatomical sites the process of disease-induced functional perturbation from a pre-symp- tomatic stage using a marker of cellular distress expressed in neurons and glial cells, the activating transcription factor 3 (ATF-3), and applied large-scale gene expression analysis to define the pattern or transcriptional changes occurring in spinal cord from the G93A SOD1 rat model of ALS in parallel with ATF-3 neuronal activation. From the disease onset on- ward, transgenic lumbar spinal cord displayed ATF-3 tran- scriptional regulation and motor cells immunostaining in as- sociation with the over-expression of genes promoting cell growth, the functional integrity of cell organelles and in- volved in the modulation of immune responses. While spinal cord from the pre-symptomatic rat showed no detectable ATF-3 transcriptional regulation, ATF-3 activation was appre- ciated in large size neurofilament-rich, small size non-pepti- dergic and parvalbumin-positive neurons within the dorsal root ganglia (DRG), and in ventral roots Schwann cells along- side macrophages infiltration. This pattern of peripheral ATF-3 activation remained detectable throughout the disease process. In the G93A SOD1 rat model of ALS, signs of roots and nerves subtle distress preceded overt clinical-patholog- ical changes, involving both glial cells and neurons that func- tion as receptors of peripheral sensory stimuli from the mus- cle. In addition, factors previously described to be linked to ATF-3 activation under various experimental conditions of stress, become switched on in spinal cord from the end-stage transgenic rat model of ALS. © 2010 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: ALS, spinal cord, dorsal root ganglia, ATF-3, bead-array analysis. Amyotrophic lateral sclerosis (ALS) is a fatal neurodegen- erative disorder characterized by the rapid progression of muscular weakness and by the impairment of bulbar func- tions, leading to death from respiratory failure on average in 3–5 years from symptoms onset. The progressive loss of motor neurons is still considered the distinguishing patho- logical feature in ALS, but how and where the pathology initiates remains unresolved. A small percentage of familial ALS patients carry a mutation of the superoxide dismutase 1 gene (SOD1; Rosen et al., 1993). It is speculated that the involvement of motor neurons may not be the initial event but rather the consequence of unfavourable molecular changes originating elsewhere. Axons and synaptic junc- tions may be the sites from where the disease propagates or may simply bear the brunt of toxic alterations originating from motor cells, including an abnormal protein aggrega- tion, an alteration of the mechanisms regulating axonal flow and an increase in membrane excitability (Gould et al., 2006). While neurophysiological and histological observa- tions have confirmed that the sensory pathways are not spared by the disease process (Nagai et al., 2001; Hunt et al., 2004), more recent investigations on a rodent model of the disease engineered with the human G93A SOD1 gene mutation have demonstrated that the disease is initiated simultaneously in the spinal cord and in the peripheral nerves of the transgenic mouse (Keller et al., 2009). The detection of bioluminescence of the glial fibrillary acid pro- tein (GFAP) using an ALS-(GFAP-luciferase/SOD (G93A)) reporter mouse has allowed to stage GFAP disease-re- lated up-regulation in peripheral and central structures from a very early pre-symptomatic stage, and to observe a sharp increase of the expression of this glial marker in peripheral nerve Schwann cells. By using chimeric mice engineered to allow SOD1 mutant selective expression in specific cell types, it has been confirmed that disease onset is non-cell autonomous and that mutant SOD1 dam- age within cell types other than motor neurons is required for the initiation and progression of the pathological pro- cess in affected areas (Yamanaka et al., 2008b). GFAP activation reflects inflammatory events in cells that support functionally and structurally motor neurons. The choice of the most informative biomarker of cell dis- tress in the unravelling of the pathological process in ALS is clearly going to influence the interpretation of the dis- ease process. In particular, it would be important to study *Corresponding author. Tel: 44-20-7882-2288; fax: 44-20-7882-2180. E-mail address: [email protected] (A. Malaspina). Abbreviations: ALS, amyotrophic lateral sclerosis; ATF-3, activating transcription factor 3; CGRP, calcitonin gene-related peptide; DRG, dorsal root ganglia; FDR, false discovery rate; GFAP, glial fibrillary acid protein; GO, Gene Ontology; IB4, isolectin B4; MMP, matrix metalloproteinases; PBS, phosphate buffered saline; SOD1, superox- ide dismutase 1 gene; TLR, toll-like receptor; WT, wild type. Neuroscience 169 (2010) 812– 827 0306-4522/10 $ - see front matter © 2010 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2010.04.053 812

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ADO
AJNJ
CMI
AfitSacvrwdtn(dswwssgvcAcdrsApaitcAstE
*EAtdami
Neuroscience 169 (2010) 812–827
0d
CTIVATION TRANSCRIPTION FACTOR-3 ACTIVATION AND THEEVELOPMENT OF SPINAL CORD DEGENERATION IN A RAT MODEL
F AMYOTROPHIC LATERAL SCLEROSISKb
AemtimliA1ibctoftfl2tsatmsndtrlfspesoafc
tTti
. MALASPINA,* S. F. A. NGOH, R. E. WARD,. C. E. HALL, F. W. D. TAI, P. K. YIP, C. JONES,. JOKIC, S. A. AVERILL, A. T. MICHAEL-TITUS AND. V. PRIESTLEY
entre for Neuroscience and Trauma, Barts and the London School ofedicine and Dentistry, Queen Mary University of London, Blizard
nstitute, 4 Newark Street, London E1 2AT, UK
bstract—It has been reported that an early activation of glialbrillary acid protein (GFAP) in astroglial cells occurs simul-aneously in peripheral nerves and spinal cord from the G93AOD1 mouse model of amyotrophic lateral sclerosis (ALS),n invariably fatal neurodegenerative disorder. In ALS, theontribute to the pathological process of different cell typesaries according to the disease stage, with a florid immuneesponse in spinal cord at end stage disease. In this study,e have mapped in different anatomical sites the process ofisease-induced functional perturbation from a pre-symp-omatic stage using a marker of cellular distress expressed ineurons and glial cells, the activating transcription factor 3ATF-3), and applied large-scale gene expression analysis toefine the pattern or transcriptional changes occurring inpinal cord from the G93A SOD1 rat model of ALS in parallelith ATF-3 neuronal activation. From the disease onset on-ard, transgenic lumbar spinal cord displayed ATF-3 tran-criptional regulation and motor cells immunostaining in as-ociation with the over-expression of genes promoting cellrowth, the functional integrity of cell organelles and in-olved in the modulation of immune responses. While spinalord from the pre-symptomatic rat showed no detectableTF-3 transcriptional regulation, ATF-3 activation was appre-iated in large size neurofilament-rich, small size non-pepti-ergic and parvalbumin-positive neurons within the dorsaloot ganglia (DRG), and in ventral roots Schwann cells along-ide macrophages infiltration. This pattern of peripheralTF-3 activation remained detectable throughout the diseaserocess. In the G93A SOD1 rat model of ALS, signs of rootsnd nerves subtle distress preceded overt clinical-patholog-
cal changes, involving both glial cells and neurons that func-ion as receptors of peripheral sensory stimuli from the mus-le. In addition, factors previously described to be linked toTF-3 activation under various experimental conditions oftress, become switched on in spinal cord from the end-stageransgenic rat model of ALS. © 2010 IBRO. Published bylsevier Ltd. All rights reserved.
Corresponding author. Tel: �44-20-7882-2288; fax: �44-20-7882-2180.-mail address: [email protected] (A. Malaspina).bbreviations: ALS, amyotrophic lateral sclerosis; ATF-3, activating
ranscription factor 3; CGRP, calcitonin gene-related peptide; DRG,orsal root ganglia; FDR, false discovery rate; GFAP, glial fibrillarycid protein; GO, Gene Ontology; IB4, isolectin B4; MMP, matrix
eetalloproteinases; PBS, phosphate buffered saline; SOD1, superox-
de dismutase 1 gene; TLR, toll-like receptor; WT, wild type.
306-4522/10 $ - see front matter © 2010 IBRO. Published by Elsevier Ltd. All rightoi:10.1016/j.neuroscience.2010.04.053
812
ey words: ALS, spinal cord, dorsal root ganglia, ATF-3,ead-array analysis.
myotrophic lateral sclerosis (ALS) is a fatal neurodegen-rative disorder characterized by the rapid progression ofuscular weakness and by the impairment of bulbar func-
ions, leading to death from respiratory failure on averagen 3–5 years from symptoms onset. The progressive loss of
otor neurons is still considered the distinguishing patho-ogical feature in ALS, but how and where the pathologynitiates remains unresolved. A small percentage of familialLS patients carry a mutation of the superoxide dismutasegene (SOD1; Rosen et al., 1993). It is speculated that the
nvolvement of motor neurons may not be the initial eventut rather the consequence of unfavourable molecularhanges originating elsewhere. Axons and synaptic junc-ions may be the sites from where the disease propagatesr may simply bear the brunt of toxic alterations originatingrom motor cells, including an abnormal protein aggrega-ion, an alteration of the mechanisms regulating axonalow and an increase in membrane excitability (Gould et al.,006). While neurophysiological and histological observa-ions have confirmed that the sensory pathways are notpared by the disease process (Nagai et al., 2001; Hunt etl., 2004), more recent investigations on a rodent model ofhe disease engineered with the human G93A SOD1 geneutation have demonstrated that the disease is initiated
imultaneously in the spinal cord and in the peripheralerves of the transgenic mouse (Keller et al., 2009). Theetection of bioluminescence of the glial fibrillary acid pro-ein (GFAP) using an ALS-(GFAP-luciferase/SOD (G93A))eporter mouse has allowed to stage GFAP disease-re-ated up-regulation in peripheral and central structuresrom a very early pre-symptomatic stage, and to observe aharp increase of the expression of this glial marker ineripheral nerve Schwann cells. By using chimeric micengineered to allow SOD1 mutant selective expression inpecific cell types, it has been confirmed that diseasenset is non-cell autonomous and that mutant SOD1 dam-ge within cell types other than motor neurons is requiredor the initiation and progression of the pathological pro-ess in affected areas (Yamanaka et al., 2008b).
GFAP activation reflects inflammatory events in cellshat support functionally and structurally motor neurons.he choice of the most informative biomarker of cell dis-
ress in the unravelling of the pathological process in ALSs clearly going to influence the interpretation of the dis-
ase process. In particular, it would be important to studys reserved.

mditm“ic3fo2oodteotseatisiai
Arssspdiapt
A
SgWw(gowsglgwa2dt
(smrp
wwpgnAldshc(mti
B
TaApctafioeetlggpcswmH2cthAvi
tNupcduhWcwslem
A. Malaspina et al. / Neuroscience 169 (2010) 812–827 813
arkers whose early activation in motor cells signals cellistress and to ascertain whether the same molecular
ndicator can be detected in other cell populations poten-ially important in the disease process. In the G93A SOD1ouse model of ALS for example, a sub-population of
sick” motor neurons has been identified at disease onsetn the spinal cord by combined staining of phosphorylated-Jun (pc-Jun) and of activating transcription factor 3 (ATF-), a member of the ATF/CREB family of transcription
actors, even before any detectable caspase-3 activationr histological evidence of motor neuron loss (Vlug et al.,005). The cellular expression of ATF-3 has been previ-usly utilized in in vivo and in vitro studies as a biomarkerf early cell distress in response to different pathologicaleterminants. It is postulated that ATF-3 plays an impor-ant role in the regulation of cell growth, of axonal regen-ration and myelination, in the homeostasis of different cellrganelles, in the modulation of protein ubiquination and inhe inhibition of the macrophage-induced inflammatory re-ponses (Seijffers et al., 2007; Campbell et al., 2005; Huntt al., 2004; Khuu et al., 2007). The co-activation of ATF-3nd of other markers of cell distress like pc-Jun suggests
hat ATF-3 may condition cell’s fate in the progression fromnjury to death. Based on the above observations, wepeculate that the detection of ATF-3 may be a goodndication of cell functional distress, which can be used as
biomarker to study the different cell types potentiallynvolved in the disease process in animal models of ALS.
In this study, we have defined the temporal profile ofTF-3 activation in the spinal cord from the G93A SOD1
at model of ALS, employing a large-scale gene expres-ion analysis and defined the parallel activation in theame tissue of several molecular pathways known to actynergistically with ATF-3. We have characterized the tem-oral and anatomical pattern of ATF-3 expression andefined its co-localization with other cell-specific markers
n the spinal cord, dorsal root ganglia (DRG), and in dorsalnd ventral roots of the G93A SOD1 rat model of ALS,roviding further insight into the origin and cell specificity ofhe pathological process in this animal model of ALS.
EXPERIMENTAL PROCEDURES
nimals and behavioural analysis
prague–Dawley male rats heterozygous for the G93A SOD1ene mutation (Emerging Model 2148 Het Male, Taconic USA;yeth and Amyotrophic Lateral Sclerosis Association, 2002) andild type female rats were bred at Taconic Breeding Services
USA). Pups from this colony project were tailed-sampled forenomic analysis and were shipped to our laboratories at 6 weeksf age. A total of 20 heterozygous and 20 wild type females ratsere retained for the experimental procedures detailed in thistudy. Following shipment, rats were housed in a specific patho-en-free animal facility at room temperature (21 °C), under a 12 h
ight–dark cycle. The rat model of ALS carrying the G93A SOD1ene mutation is known to develop signs of motor impairment,ith either a hindlimb or a forelimb initial disease presentation,nd to progress rapidly to widespread paralysis in approximatelyweeks after disease onset. In this study, the disease onset was
efined as the identification of early signs of motor limb dysfunc-
ion using a serial evaluation of locomotor activity in an open field f80�30�30 cm3 polypropylene box) (Jokic et al., 2007). End-tage disease was defined either as a reduction of 25% in theaximum body weight or as the consistent loss of the righting
eflex (i.e. the failure of the animal to right itself within 30 s whenlaced on its side).
Groups of five G93A SOD1 mutant (transgenic) rats and fiveild type (WT) age-matched littermates were sacrificed at 10eeks of age (a time point in which transgenic rats are stillre-symptomatic), at disease onset and at disease end-stage, forene expression analysis. The thoraco–lumbar region of the spi-al cord was harvested and immediately frozen in liquid nitrogen.dditional groups of three transgenic and three WT age-matched
ittermates were sacrificed at a pre-symptomatic stage and atisease end-stage, for histochemical analyses. All transgenic ratsacrificed at onset and at end-stage disease presented with aindlimb paralysis at disease onset. All animal procedures wereonducted according to the Animals Scientific Procedures Act1986), approved by the United Kingdom Home Office. Only fe-ale rats have been used to ensure consistence in rat gender and
o avoid any potential gender-related confounding factors in thenterpretation of the results.
ead array gene expression analysis
he RNA expression profile of the spinal cord from transgenic ratsnd from WT age-matched littermates was analyzed using a Beadrray analysis system (Illumina, San Diego, USA). For the pur-ose of this gene expression analysis, spinal cord samples wereollected from a total of 30 animals, including five samples fromransgenic rats at a pre-symptomatic stage, five samples from ratst disease onset, five samples from rats at disease end-stage andve samples from age-matched WT littermates sacrificed at eachf the above time points. We have performed two types of genexpression analysis: (1) a standard Bead array differential genexpression study comparing spinal cord samples from femaleransgenic rats to spinal cord samples from age and sex-matchedittermates, to obtain a profile of the most differentially regulatedenes in transgenic spinal cord, (2) a pathway analysis of the mainene expression changes detected by Bead array analysis ofools of homogeneous cord samples (each pool containing spinalord samples of the same genetic type dissected from animalsacrificed at the same time points). The study on pooled samplesas performed to obtain an ontological description of the mainolecular changes occurring in transgenic spinal cord, usingigh-Throughput GOMiner (Malaspina et al., 2008; Zeeber et al.,005) to assemble the differentially expressed genes within spe-ific Gene Ontology (GO) categories. We have also identifiedhose differentially regulated gene candidates already reported toave functional interaction with AT-3 and we have also identifiedTF-3-containing gene categories that appear to become acti-ated in the time points of the disease process that have beennvestigated.
RNA was extracted from the spinal cord samples using the SVotal RNA isolation system (Promega, UK) and quantified using aanodrop ND-1000 spectrophotometer. RNA quality was checkedsing the Agilent bioanalyser system (Agilent). Three RNA sam-les from transgenic and three RNA samples from WT lumbarords for each of the time-points considered (pre-symptomatic,isease onset and disease end-stage) were tested individuallysing Bead-array analysis. In addition, in a second experiment weave pooled the RNA samples obtained from either transgenic orT rats sacrificed at the same time points (a total of six pools
ontaining five RNA samples each). cRNA labeling was performedith 750 ng of total RNA from each RNA samples or pool of RNAamples, using the Ambion Total Prep kit. cRNA purity and label-ing was checked using the Nanodrop and Agilent bioanalyser. Tonsure homogeneous conditions for the gene expression experi-ents, 500 ng from each of the nine labelled RNA samples and
rom each of the six labelled RNA pools were hybridized simulta-

nsUpdautctuya
51sdiaDiscpfi2
O
W(biocdp(ogtbtBaeewctPfesg(
At
Wcdaipgi
dIwiissotc
I
Ttgetm0(24aomSssswfsDabsfn(SwtlSdi(E(iaibwlsuP
wiLtHDe
A. Malaspina et al. / Neuroscience 169 (2010) 812–827814
eously to 15 separate arrays contained in two RatRef-12 Expres-ion BeadChip, as per the Illumina protocol (Illumina, San Diego,SA). Using RNA from one of the pools (transgenic rats at there-symptomatic stage), we have performed a technical replica inuplicate, probing two separate arrays available in one of the12-rray Bead-chip, and under the same experimental conditionstilized for the gene expression analyses reported above. A scat-
er plot analysis tool in logarithmic scale was used to calculateorrelation coefficients for the gene expression data generated byhe technical replica experiments. Scatter plot analysis was alsosed to correlate the results of differential gene expression anal-sis between single samples and pooled samples obtained fromnimals of the same genetic type sacrificed at the same time point.
Output files were analyzed using the Illumina BeadArray00GX Reader the Illumina BeadStudio software (version.5.0.34). The detection signal for each probe in the Bead-arrayystem was expressed in pixel intensities and with a P-value ofetection generated by the Bead-array workflow station. Normal-
zed expression data were obtained subtracting local backgroundnd using a rank-invariant algorithm (BeadStudio-3, Illumina, Saniego, USA) to remove systematic signal variation of non-biolog-
cal origin. BeadStudio-3 was used to test differential gene expres-ion between transgenic and WT spinal cord RNA samples at thehosen time points, including RNA from single tissue samples andools of RNAs of the same genetic type and from animals sacri-ced at the same time point as described above (Malaspina et al.,008).
ntological analysis by High-Throughput GO-Miner
e have used High-Throughput GOMiner as previously reportedMalaspina et al., 2008; Zeeber et al., 2005), to characterize theiological significance of the gene expression changes identified
n transgenic spinal cord at a pre-symptomatic stage, at diseasenset and at disease end-stage. High-Throughput GOMiner pro-esses (and integrates) simultaneously multiple gene expressionatasets. The program identifies those categories within the ap-roximately 8000 gene categories archived in the GO databasehttp://www.geneontology.org/), which present a significant levelf enrichment in the differentially regulated genes under investi-ation. Submission to High-Throughput GoMiner of the differen-ially expressed genes (each represented by a flat �1 or �1inary code, to indicate up-regulation or down-regulation respec-ively) and of the list of the probes contained in the Illumina rateadchip, was undertaken as previously reported (Malaspina etl., 2008). GO categories were identified according to their level ofnrichment of “changed genes” and to their statistical significance,xpressed by a false discovery rate (FDR) (only gene categoriesith FDR�0.05 were selected). The FDR addresses the multipleomparison problems that occur when calculating the P-values forhousands of categories, and protects against over-interpreting-values that do not have a biological meaning. The FDR value
or a specific GO category found to be significantly differentiallyxpressed was automatically computed for all the samples intudy, allowing an estimation of the relative importance of eachene category across all time points and experimental groupsMalaspina et al., 2008).
TF-3 activation and the gene expression profile inhe G93A SOD1 spinal cord
e have used the gene expression data from transgenic spinalord from pre-symptomatic animals and from animal models atisease onset and at end-stage, to identify molecular pathwaysnd genes that show a pattern of co-regulation with ATF-3, includ-
ng those that have functional synergy with ATF-3. For this pur-ose, we have identified in our gene expression datasets, all thoseene categories computed by High-Throughput GOMiner which
nclude ATF-3. We have also reviewed the scientific literature and w
ifferent biological databases including NCBI, Gene Card, OMIM,HOP (information hyperlink over proteins) to identify those genesith an ATF-3 down-stream activation under conditions of exper-
mentally-induced stress, ranging from mechanical trauma, genet-cally-induced neurodegeneration and other experimental stres-ors like serum-deprivation. We have then performed a cross-earch in our gene expression datasets, to ascertain whether anyf the genes previously linked to ATF-3 is reported to be differen-ially regulated in transgenic spinal cord at any of the time pointsonsidered.
mmunohistochemistry
he immunohistochemical expression of ATF-3 was analyzed inhe L5 lumbar spinal cord and DRGs obtained from additionalroups of WT and transgenic rats at both 10 weeks of age and atnd-stage disease (three animals per group). Rats were anesthe-ized with sodium pentobarbital (Danisco A/S, Brabrand, Den-ark; 60 mg/kg, i.p.) and perfused transcardially with ice-cold.9% NaCl followed by 4% paraformaldehyde in phosphate bufferpH 7.4). The spinal cord and DRGs were removed, post-fixed forh and further cryoprotected in 30% sucrose solution overnight at°C. Tissues were embedded in Tissue-Tek O.C.T. compound
nd stored at �80 °C. Serial transverse sections (15 �m) were cutn a cryostat through the lumbar (L3–L5) spinal cord enlarge-ents and through the DRG (8 �m sections) and mounted ontouperFrost Plus slides (VWR International, UK). The spinal cordections included attached dorsal and ventral roots, and the DRGections included attached dorsal and ventral roots and proximalpinal nerves. Prior to immunohistochemical staining, the sectionsere washed in phosphate buffered saline (PBS, 10 mM, pH 7.4)
our times for 5 min and then blocked in 10% normal donkeyerum for 2 h at room temperature. Sections from spinal cord andRGs were incubated for 48 h at 4 °C with a primary antiserumgainst ATF-3 (1:200, Chemicon), and subsequently double-la-eled with the following polyclonal primary antibodies: for thepinal cord sections, sheep polyclonal anti-choline acetyltrans-erase (anti-ChAT, 1:1000, Chemicon, UK; a marker for motoreurons), and mouse monoclonal anti-glial fibrillary acidic proteinanti-GFAP, 1:8000, Chemicon, UK) or goat anti-S-100 (1:1000,igma) to label glial cells. Double labeling of the DRG sectionsas achieved by processing the tissues for ATF-3 immunoreac-
ivity, together with the following primary antisera: anti-N52 to stainarge diameter neurons rich in 200 kD neurofilament (1:16000,igma), anti-calcitonin gene-related peptide (CGRP) to label smalliameter peptidergic neurons (1:100, Santa Cruz), biotinylated
solectin B4 (IB4) to label small diameter non-peptidergic neurons1:200, Sigma), S-100 to label Schwann cells (1:1000, Sigma),D1 to label macrophages (1:1000, Serotec), and parvalbumin
1:500, Sigma), to label proprioceptive DRG neurons. After wash-ng four times for 5 min in PBS, the sections were incubated withppropriate secondary antibodies and labeled with fluorescein
sothiocyanate (FITC) or tetra-rhodamine isothiocyanate (TRITC),oth at 1:600, Jackson Laboratories. IB4 sections were taggedith avidin-FITC (1:400, Jackson Laboratories). All sections were
abeled with bisbenzimide Hoechst 33342 (2 �g/mL). Finally, thelides were mounted with 1:8 PBS-glycerol. Images were takensing a Leica microscope and plates assembled using Adobehotoshop.
The determination of ATF-3 positive neurons in DRG sectionsas carried out by counting the number of neurons with an ATF-3
mmunoreactive nucleus at 20� objective magnitude using aeica Microscope. This was expressed as a percentage of theotal number of neurons in the field of view, visualized usingoechst 33342 counterstaining. The same number of sections ofRG from pre-symptomatic (10-week of age) and end-stage dis-ase were used for quantitation analysis. At least 500 neurons
ere counted per animal. Data are expressed as mean�SEM.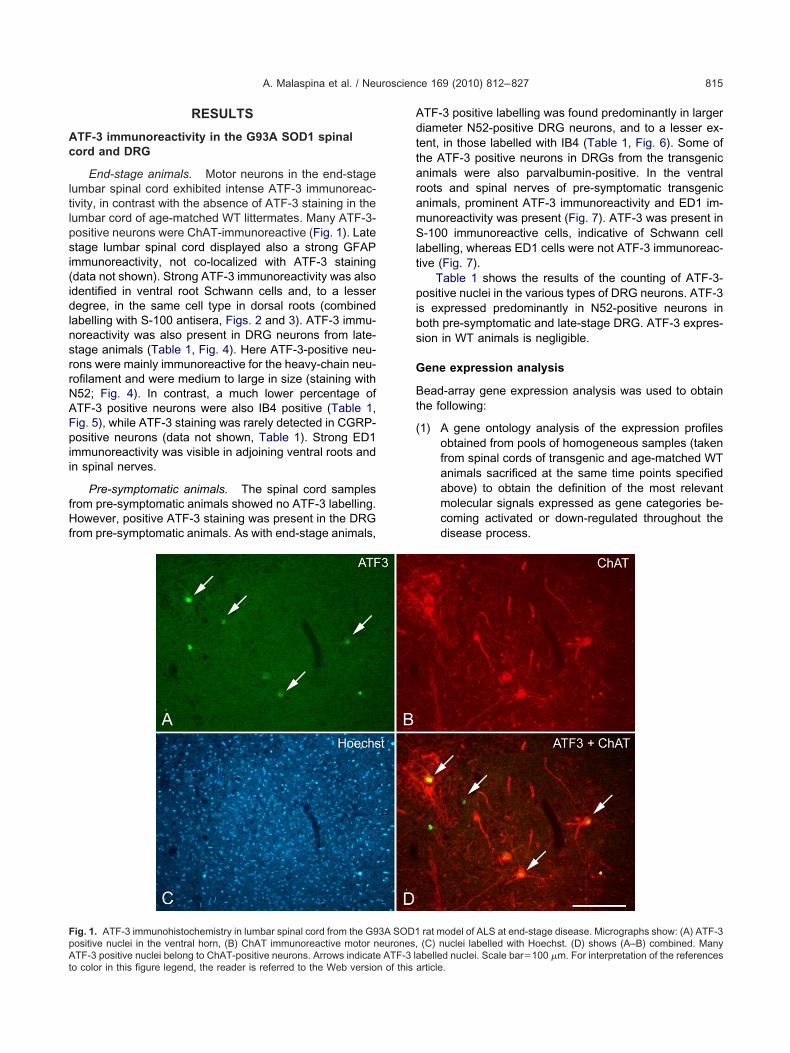
Ac
ltlpsi(idlnsrrNAFpii
fHf
AdttaramSlt
pibs
G
Bt
(
FpAt
A. Malaspina et al. / Neuroscience 169 (2010) 812–827 815
RESULTS
TF-3 immunoreactivity in the G93A SOD1 spinalord and DRG
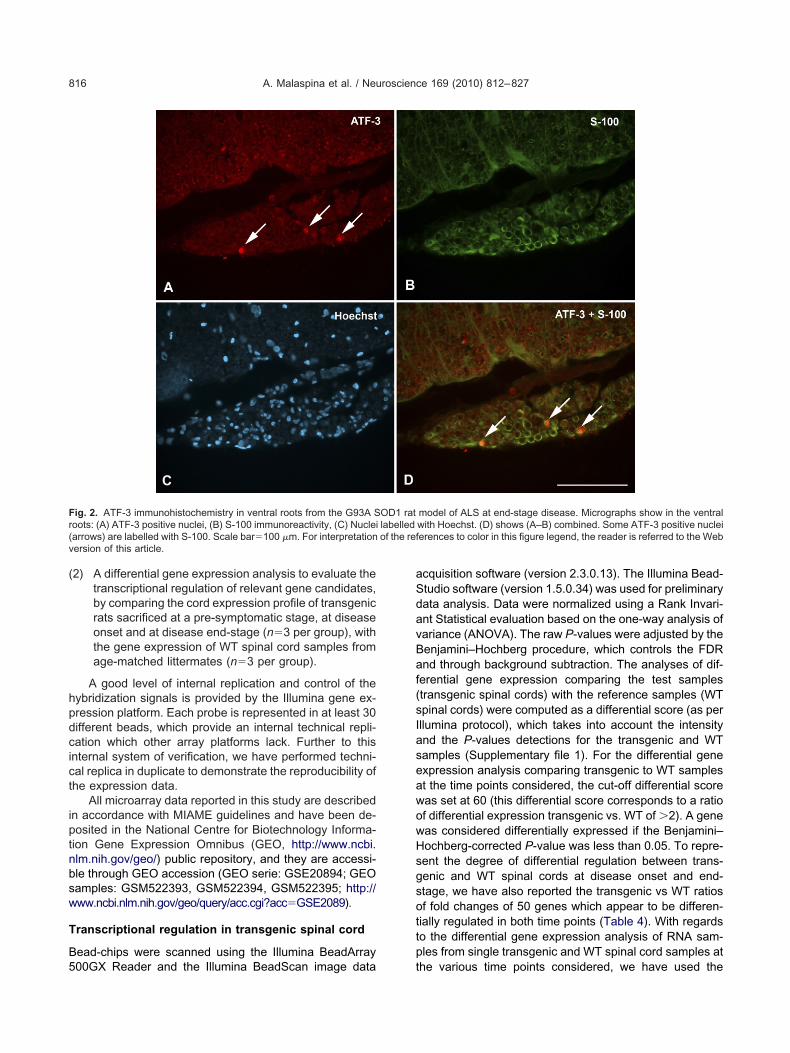
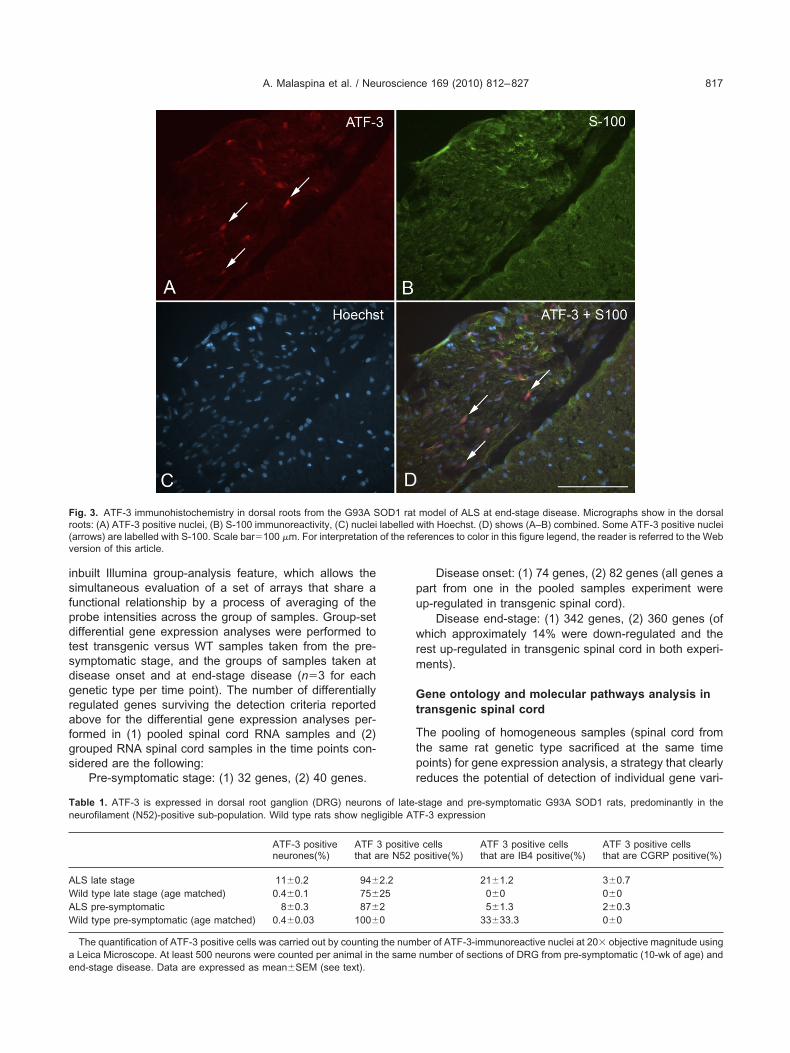
End-stage animals. Motor neurons in the end-stageumbar spinal cord exhibited intense ATF-3 immunoreac-ivity, in contrast with the absence of ATF-3 staining in theumbar cord of age-matched WT littermates. Many ATF-3-ositive neurons were ChAT-immunoreactive (Fig. 1). Latetage lumbar spinal cord displayed also a strong GFAPmmunoreactivity, not co-localized with ATF-3 stainingdata not shown). Strong ATF-3 immunoreactivity was alsodentified in ventral root Schwann cells and, to a lesseregree, in the same cell type in dorsal roots (combined
abelling with S-100 antisera, Figs. 2 and 3). ATF-3 immu-oreactivity was also present in DRG neurons from late-tage animals (Table 1, Fig. 4). Here ATF-3-positive neu-ons were mainly immunoreactive for the heavy-chain neu-ofilament and were medium to large in size (staining with52; Fig. 4). In contrast, a much lower percentage ofTF-3 positive neurons were also IB4 positive (Table 1,ig. 5), while ATF-3 staining was rarely detected in CGRP-ositive neurons (data not shown, Table 1). Strong ED1
mmunoreactivity was visible in adjoining ventral roots andn spinal nerves.
Pre-symptomatic animals. The spinal cord samplesrom pre-symptomatic animals showed no ATF-3 labelling.owever, positive ATF-3 staining was present in the DRG
rom pre-symptomatic animals. As with end-stage animals,
ig. 1. ATF-3 immunohistochemistry in lumbar spinal cord from the G9ositive nuclei in the ventral horn, (B) ChAT immunoreactive motor n
TF-3 positive nuclei belong to ChAT-positive neurons. Arrows indicate ATF-3 lao color in this figure legend, the reader is referred to the Web version of this a
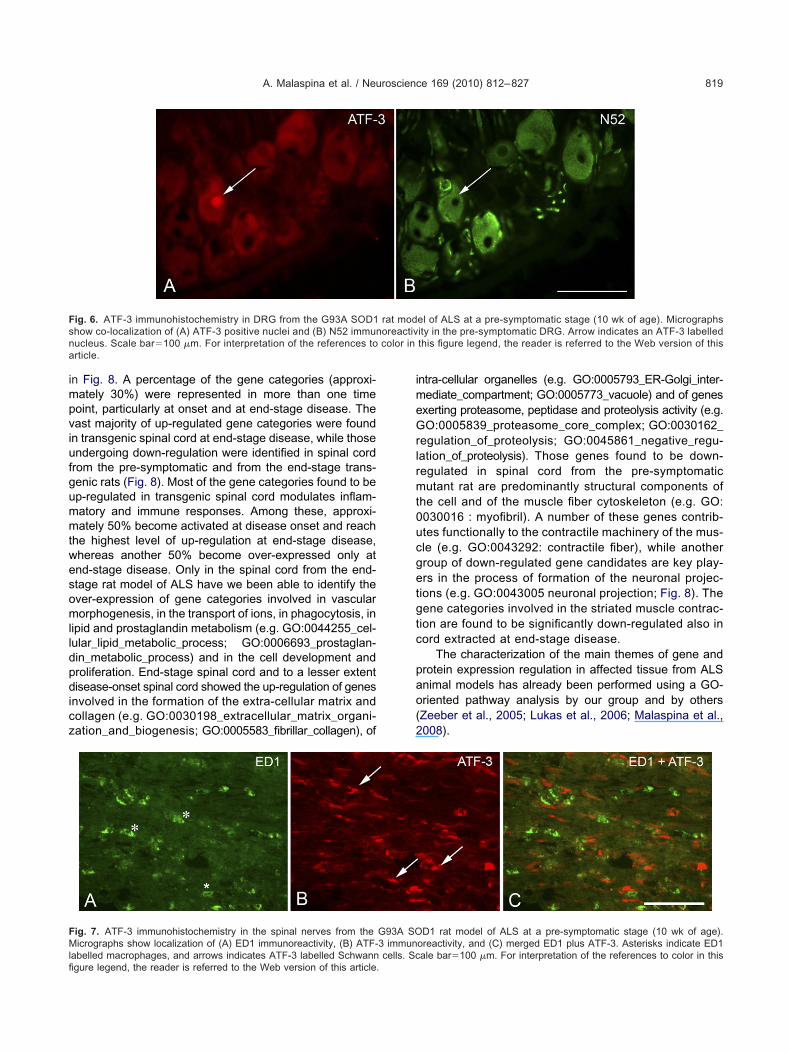
TF-3 positive labelling was found predominantly in largeriameter N52-positive DRG neurons, and to a lesser ex-ent, in those labelled with IB4 (Table 1, Fig. 6). Some ofhe ATF-3 positive neurons in DRGs from the transgenicnimals were also parvalbumin-positive. In the ventraloots and spinal nerves of pre-symptomatic transgenicnimals, prominent ATF-3 immunoreactivity and ED1 im-unoreactivity was present (Fig. 7). ATF-3 was present in-100 immunoreactive cells, indicative of Schwann cell
abelling, whereas ED1 cells were not ATF-3 immunoreac-ive (Fig. 7).
Table 1 shows the results of the counting of ATF-3-ositive nuclei in the various types of DRG neurons. ATF-3
s expressed predominantly in N52-positive neurons inoth pre-symptomatic and late-stage DRG. ATF-3 expres-ion in WT animals is negligible.
ene expression analysis
ead-array gene expression analysis was used to obtainhe following:
1) A gene ontology analysis of the expression profilesobtained from pools of homogeneous samples (takenfrom spinal cords of transgenic and age-matched WTanimals sacrificed at the same time points specifiedabove) to obtain the definition of the most relevantmolecular signals expressed as gene categories be-coming activated or down-regulated throughout thedisease process.
rat model of ALS at end-stage disease. Micrographs show: (A) ATF-3(C) nuclei labelled with Hoechst. (D) shows (A–B) combined. Many
3A SOD1eurones,
belled nuclei. Scale bar�100 �m. For interpretation of the referencesrticle.
(
hpdcict
iptnbsw
T
B5
aSdavBaf(sIaseawowHsgsottp
Fr( of the refv
A. Malaspina et al. / Neuroscience 169 (2010) 812–827816
2) A differential gene expression analysis to evaluate thetranscriptional regulation of relevant gene candidates,by comparing the cord expression profile of transgenicrats sacrificed at a pre-symptomatic stage, at diseaseonset and at disease end-stage (n�3 per group), withthe gene expression of WT spinal cord samples fromage-matched littermates (n�3 per group).
A good level of internal replication and control of theybridization signals is provided by the Illumina gene ex-ression platform. Each probe is represented in at least 30ifferent beads, which provide an internal technical repli-ation which other array platforms lack. Further to thisnternal system of verification, we have performed techni-al replica in duplicate to demonstrate the reproducibility ofhe expression data.
All microarray data reported in this study are describedn accordance with MIAME guidelines and have been de-osited in the National Centre for Biotechnology Informa-ion Gene Expression Omnibus (GEO, http://www.ncbi.lm.nih.gov/geo/) public repository, and they are accessi-le through GEO accession (GEO serie: GSE20894; GEOamples: GSM522393, GSM522394, GSM522395; http://ww.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc�GSE2089).
ranscriptional regulation in transgenic spinal cord
ead-chips were scanned using the Illumina BeadArray
ig. 2. ATF-3 immunohistochemistry in ventral roots from the G93A Soots: (A) ATF-3 positive nuclei, (B) S-100 immunoreactivity, (C) Nucleiarrows) are labelled with S-100. Scale bar�100 �m. For interpretationersion of this article.
00GX Reader and the Illumina BeadScan image data t
cquisition software (version 2.3.0.13). The Illumina Bead-tudio software (version 1.5.0.34) was used for preliminaryata analysis. Data were normalized using a Rank Invari-nt Statistical evaluation based on the one-way analysis ofariance (ANOVA). The raw P-values were adjusted by theenjamini–Hochberg procedure, which controls the FDRnd through background subtraction. The analyses of dif-erential gene expression comparing the test samplestransgenic spinal cords) with the reference samples (WTpinal cords) were computed as a differential score (as perllumina protocol), which takes into account the intensitynd the P-values detections for the transgenic and WTamples (Supplementary file 1). For the differential genexpression analysis comparing transgenic to WT samplest the time points considered, the cut-off differential scoreas set at 60 (this differential score corresponds to a ratiof differential expression transgenic vs. WT of �2). A geneas considered differentially expressed if the Benjamini–ochberg-corrected P-value was less than 0.05. To repre-ent the degree of differential regulation between trans-enic and WT spinal cords at disease onset and end-tage, we have also reported the transgenic vs WT ratiosf fold changes of 50 genes which appear to be differen-ially regulated in both time points (Table 4). With regardso the differential gene expression analysis of RNA sam-les from single transgenic and WT spinal cord samples at
model of ALS at end-stage disease. Micrographs show in the ventralwith Hoechst. (D) shows (A–B) combined. Some ATF-3 positive nucleierences to color in this figure legend, the reader is referred to the Web
OD1 ratlabelled
he various time points considered, we have used the
isfpdtsdgrafgs
pu
wrm
Gt
Ttpr
Fr( of the refv
Tn
AWAW
ae
A. Malaspina et al. / Neuroscience 169 (2010) 812–827 817
nbuilt Illumina group-analysis feature, which allows theimultaneous evaluation of a set of arrays that share aunctional relationship by a process of averaging of therobe intensities across the group of samples. Group-setifferential gene expression analyses were performed to
est transgenic versus WT samples taken from the pre-ymptomatic stage, and the groups of samples taken atisease onset and at end-stage disease (n�3 for eachenetic type per time point). The number of differentiallyegulated genes surviving the detection criteria reportedbove for the differential gene expression analyses per-ormed in (1) pooled spinal cord RNA samples and (2)rouped RNA spinal cord samples in the time points con-idered are the following:
Pre-symptomatic stage: (1) 32 genes, (2) 40 genes.
ig. 3. ATF-3 immunohistochemistry in dorsal roots from the G93A Soots: (A) ATF-3 positive nuclei, (B) S-100 immunoreactivity, (C) nucleiarrows) are labelled with S-100. Scale bar�100 �m. For interpretationersion of this article.
able 1. ATF-3 is expressed in dorsal root ganglion (DRG) neuroneurofilament (N52)-positive sub-population. Wild type rats show neg
ATF-3 positiveneurones(%)
ATFthat a
LS late stage 11�0.2 94�
ild type late stage (age matched) 0.4�0.1 75�
LS pre-symptomatic 8�0.3 87�
ild type pre-symptomatic (age matched) 0.4�0.03 100�
The quantification of ATF-3 positive cells was carried out by countingLeica Microscope. At least 500 neurons were counted per animal in t
nd-stage disease. Data are expressed as mean�SEM (see text).Disease onset: (1) 74 genes, (2) 82 genes (all genes aart from one in the pooled samples experiment werep-regulated in transgenic spinal cord).
Disease end-stage: (1) 342 genes, (2) 360 genes (ofhich approximately 14% were down-regulated and the
est up-regulated in transgenic spinal cord in both experi-ents).
ene ontology and molecular pathways analysis inransgenic spinal cord
he pooling of homogeneous samples (spinal cord fromhe same rat genetic type sacrificed at the same timeoints) for gene expression analysis, a strategy that clearlyeduces the potential of detection of individual gene vari-
model of ALS at end-stage disease. Micrographs show in the dorsalwith Hoechst. (D) shows (A–B) combined. Some ATF-3 positive nucleierences to color in this figure legend, the reader is referred to the Web
stage and pre-symptomatic G93A SOD1 rats, predominantly in theF-3 expression
cellsositive(%)
ATF 3 positive cellsthat are IB4 positive(%)
ATF 3 positive cellsthat are CGRP positive(%)
21�1.2 3�0.70�0 0�05�1.3 2�0.3
33�33.3 0�0
ber of ATF-3-immunoreactive nuclei at 20� objective magnitude usingnumber of sections of DRG from pre-symptomatic (10-wk of age) and
OD1 ratlabelled
s of late-ligible AT
3 positivere N52 p
2.22520
the numhe same

aptd(pdrpp
Hcbcftmind
Fnt the Web
FrHa
A. Malaspina et al. / Neuroscience 169 (2010) 812–827818
tion across samples, has been performed solely for theurpose of studying by High-Throughput GOMiner the on-
ology of the most relevant molecular responses occurringuring neurodegeneration in transgenic spinal cord,Malaspina et al., 2008; Zeeber et al., 2005). High-through-ut Go Miner has computed 140 up-regulated and 11own-regulated GO categories containing the differentiallyegulated genes in transgenic spinal cord in the three timeoints considered (FDR�0.05, enrichment �2%). As inreviously reported analyses of gene expression data by
ig. 4. ATF-3 immunohistochemistry of DRG from the G93A SOD1 ruclei and (B) N52 positive cells. Arrows indicate ATF-3 labelled nucle
he references to color in this figure legend, the reader is referred to
ig. 5. ATF-3 immunohistochemistry in dorsal roots and DRG from tharely belonged to IB4 positive neurones. Micrographs show in DRG: (A
oechst. (D) shows (A–C) combined. For interpretation of the references to colorticle.igh-Throughput GOMiner (Malaspina et al., 2008), a per-entage of the selected gene categories was representedy general “parent” categories (e.g. GO:0008150 : biologi-al_process; GO:0065007 : biological regulation). There-ore, 78 general gene categories were excluded from fur-her analysis, as not considered informative of specificolecular mechanisms. The remaining 73 were grouped
nto functionally-related subsets of gene categories; theumber of categories in each functional group and theiristribution across the time points considered is displayed
of ALS at end-stage disease. Micrographs show (A) ATF-3 positivere immunoreactive for N52. Scale bar�100 �m. For interpretation ofversion of this article.
SOD1 rat model of ALS at end-stage disease. ATF-3 positive nuclei-3 positive nucleus, (B) IB4 positive neurones, (C) nuclei labeled with
at modeli which a
e G93A) An ATF
r in this figure legend, the reader is referred to the Web version of this
impviufgummtwesomlldpdicz
imeGrlrmt0ucgetgtc
pao(2
Fsna
FMlfi
A. Malaspina et al. / Neuroscience 169 (2010) 812–827 819
n Fig. 8. A percentage of the gene categories (approxi-ately 30%) were represented in more than one timeoint, particularly at onset and at end-stage disease. Theast majority of up-regulated gene categories were foundn transgenic spinal cord at end-stage disease, while thosendergoing down-regulation were identified in spinal cordrom the pre-symptomatic and from the end-stage trans-enic rats (Fig. 8). Most of the gene categories found to bep-regulated in transgenic spinal cord modulates inflam-atory and immune responses. Among these, approxi-ately 50% become activated at disease onset and reach
he highest level of up-regulation at end-stage disease,hereas another 50% become over-expressed only atnd-stage disease. Only in the spinal cord from the end-tage rat model of ALS have we been able to identify thever-expression of gene categories involved in vascularorphogenesis, in the transport of ions, in phagocytosis, in
ipid and prostaglandin metabolism (e.g. GO:0044255_cel-ular_lipid_metabolic_process; GO:0006693_prostaglan-in_metabolic_process) and in the cell development androliferation. End-stage spinal cord and to a lesser extentisease-onset spinal cord showed the up-regulation of genes
nvolved in the formation of the extra-cellular matrix andollagen (e.g. GO:0030198_extracellular_matrix_organi-ation_and_biogenesis; GO:0005583_fibrillar_collagen), of
ig. 6. ATF-3 immunohistochemistry in DRG from the G93A SOD1how co-localization of (A) ATF-3 positive nuclei and (B) N52 immunucleus. Scale bar�100 �m. For interpretation of the references torticle.
ig. 7. ATF-3 immunohistochemistry in the spinal nerves from theicrographs show localization of (A) ED1 immunoreactivity, (B) ATF-
abelled macrophages, and arrows indicates ATF-3 labelled Schwann cells. Scgure legend, the reader is referred to the Web version of this article.
ntra-cellular organelles (e.g. GO:0005793_ER-Golgi_inter-ediate_compartment; GO:0005773_vacuole) and of genesxerting proteasome, peptidase and proteolysis activity (e.g.O:0005839_proteasome_core_complex; GO:0030162_
egulation_of_proteolysis; GO:0045861_negative_regu-ation_of_proteolysis). Those genes found to be down-egulated in spinal cord from the pre-symptomaticutant rat are predominantly structural components of
he cell and of the muscle fiber cytoskeleton (e.g. GO:030016 : myofibril). A number of these genes contrib-tes functionally to the contractile machinery of the mus-le (e.g. GO:0043292: contractile fiber), while anotherroup of down-regulated gene candidates are key play-rs in the process of formation of the neuronal projec-
ions (e.g. GO:0043005 neuronal projection; Fig. 8). Theene categories involved in the striated muscle contrac-
ion are found to be significantly down-regulated also inord extracted at end-stage disease.
The characterization of the main themes of gene androtein expression regulation in affected tissue from ALSnimal models has already been performed using a GO-riented pathway analysis by our group and by othersZeeber et al., 2005; Lukas et al., 2006; Malaspina et al.,008).
el of ALS at a pre-symptomatic stage (10 wk of age). Micrographsity in the pre-symptomatic DRG. Arrow indicates an ATF-3 labelledthis figure legend, the reader is referred to the Web version of this
D1 rat model of ALS at a pre-symptomatic stage (10 wk of age).oreactivity, and (C) merged ED1 plus ATF-3. Asterisks indicate ED1
rat modoreactivcolor in
G93A SO3 immun
ale bar�100 �m. For interpretation of the references to color in this

Gr
Ttpdocrtetecsgtttm(ts(oidtsrtashea
iktCfedaatct
Fs(rerica
Ftaaddf
A. Malaspina et al. / Neuroscience 169 (2010) 812–827820
roup analysis of transgenic spinal cord tissues:elevant differentially regulated genes
he gene expression analysis of spinal cord tissues fromransgenic and age-matched WT rats using the Illuminalatform has identified differentially regulated gene candi-ates whose function is mostly expressed by the genentology categories identified testing the pooled spinalord samples (Table 4, Fig. 8). Most of the differentiallyegulated gene candidates are detected in spinal cordissues from end-stage disease and appear to be over-xpressed in transgenic spinal cord (Table 4; Supplemen-ary file 1). 50 genes are up-regulated in both onset andnd-stage spinal cords and their level of transgenic spinalord up-regulation seem to increase significantly at end-tage disease (Table 4). As expected from the preliminaryene ontology analysis performed in the same pooledissues, the vast majority of molecular signals undergoingranscriptional regulation play a role in the modulation ofhe immune response, in cell proliferation, in the lipidetabolism and in the regulation of the proteasome activity
Tables 3 and 4, Fig. 8). ATF-3 mRNA expression appearso increase significantly at disease onset, with a furtherurge at end-stage when compared to WT spinal cordTable 4, in bold; Fig. 9), along with the up-regulation ofther gene candidates involved in the regulation of the
mmune response, cell growth and the proteasome. As theisease progresses, the tissue expression of this transcrip-ion factor increases more than 20 folds over the expres-ion level detected in spinal cord from age-matched WTats (Table 2). Among those gene candidate which formhe structure and function of the striated muscle machinerynd that appear to become down-regulated in transgenicpinal cord from the pre-symptomatic transgenic rat, weave identified Creatine kinase (CK), Actin alpha-1 (skel-tal muscle), Actinin alpha-3 (ACTN3; skeletal muscle)
-20
-10
0
10
20
30
40
50
60
Imm
une
resp
onse
Prol
ifera
tion
Extr
acel
lula
r mat
rix
Prot
ein
hom
eost
asis
*
Lipi
d/pr
osta
glan
dins
met
abol
ism
Up-regulated
Number of GO categories
Down-regulated
ig. 8. High-Throughput GO Miner ontological characterization of the damples extracted from the transgenic rat model of ALS at 10 wk of aFDR�0.05) with a high level of enrichment of the differentially regulaeports the number of gene categories for each biological function annd-stage disease and to a lesser extent, at disease onset, and containole in the modulation of the immune response and to a lesser extent, oncluding proteasome activity and proteolysis. ** Genes particularly reontractile functioning of the muscle. For interpretation of the referencerticle.
nd Actin beta (ACTB) (Supplementary file 1). The admin-ia
stration of creatine to the G93A mouse model of ALS isnown to produce a dose-dependent improvement in mo-or performance and an extended survival, by acting onK, on its substrates creatine and phosphocreatine and on
unctionally related actins to improve the impaired cellularnergy buffering at sites of energy production (mitochon-ria) and consumption (Klivenyi et al., 1999). Actins havelready been under the radar of the scientific communitys they appear to contribute to the disease-specific cy-oskeletal changes observed in ALS. TDP-43, an importantomponent of the cell inclusions found in ALS affectedissues, has been shown to co-localize with ACTA1, while
Org
anel
le
Vaso
gene
sis
Taxi
s
Ion
chan
nels
Phag
ocyt
osis
Com
plem
ent
Stria
ted
mus
cle*
*
Cel
l pro
ject
ion
10-weeks
Onset
Late-stage
lly regulated genes obtained by Bead-array analysis of the cord tissueease onset and at end-stage disease. The selected gene categorieshave been grouped according to their biological function. The graph
int. The majority of selected gene categories have been identified atlated genes. Most of the gene categories identified plays an importantth. * gene categories involved in protein metabolism and homeostasisd in striated muscle. Some of the gene categories are central to ther in this figure legend, the reader is referred to the Web version of this
0
100
200
300
400
500
600
700
800
900
egats-etaLtesnOskeew-01
Disease �me points
Inte
nsity
det
ectio
n va
lue
G93ASOD1WT
ig. 9. Changes in ATF-3 RNA expression in spinal cord from theransgenic rat model of ALS at a pre-symptmatic stage (10 wk of age),t disease onset and at disease end-stage (blue code) and fromge-matched wild type littermates (red code). Normalized intensityetection values for ATF-3 have been taken from the gene expressionata obtained by Bead-array gene expression analysis of spinal cord
rom the animals in study. For interpretation of the references to color
Bin
ding
Tran
spor
t
ifferentiage, at disted genesd time po
up-reguf cell growpresentes to colo
n this figure legend, the reader is referred to the Web version of thisrticle.

io
SpsaRfr
M
ApfgeptcaGca03it0to
tAadu
ptto(saH2ctapdJCrsiBTpslaatm(piro(op
Tow
G
GGGGGGGGGGGG
*g
A. Malaspina et al. / Neuroscience 169 (2010) 812–827 821
nteracting with another protein linked to the pathogenesisf ALS, angiogenin (Seilhean et al., 2009).
The r2-value obtained by correlation analysis (Bead-tudio-3 scatter plot) of the technical replica experimenterformed probing two separate bead-arrays with theame RNA sample was 0.9876. The r2 of the correlationnalysis between two Bead arrays probed with spinal cordNA from a single pre-symptomatic transgenic rat and
rom a pool of five spinal cord RNAs from pre-symptomaticats was 0.9754.
olecular responses co-regulated with ATF-3
mong those gene categories identified by High-Through-ut GOMiner which show a significant enrichment of dif-erentially regulated genes in the spinal cord from trans-enic rats (FDR�0.05; enrichment�2%), 12 were found atnd-stage disease to contain ATF-3 (Table 2). In line withrevious data on the role of ATF-3 activation under condi-ions of stress, the biological role of these categories en-ompasses the regulation of cell development, differentiationnd proliferation (e.g. GO:0032502_developmental_process;O:0008283_cell_proliferation; GO:0042127_regulation_of_ell_proliferation) and protein homeostasis including bindingnd dimerization (e.g. GO:0005515_protein_binding; GO:046983_protein_dimerization_activity). Some of the ATF--containing gene categories include genes that code for
mportant components of the cell architecture, or that con-ribute to the function of different cell organelles (GO:005575_cellular_component). Three of these ATF-3-con-aining gene categories were also up-regulated at diseasenset (Table 2).
Our literature search and database mining have iden-ified a number of genes known to act in synergism withTF-3 or to be down-stream targets of its transcriptionalctivity, under different condition of experimentally-in-uced stress. These include gene candidates which mod-
able 2. ATF-3-containing GO categories identified as up-regulated af the differentially expressed genes found in transgenic spinal cord (uith a significant level of differential regulation (FDR�0.05) at the tim
O category On
En
O:0032502_developmental_process 1.8O:0048518_positive_regulation_of_biological_process 2O:0008284_positive_regulation_of_cell_proliferation 3.2O:0008283_cell_proliferation 1.8O:0042127_regulation_of_cell_proliferation 1.8O:0005515_protein_binding 0.7O:0046983_protein_dimerization_activity 1.4O:0065007_biological_regulation 1.1O:0048522_positive_regulation_of_cellular_process 1.3O:0030154_cell_differentiation 0.9O:0005488_binding 0.6O:0005575_cellular_component 0.7
FDR, false discovery rate. Highly significant FDRs are approximateThe enrichment expresses the percentage of differentially regulated gene category.
late the cell cycle, cell proliferation and differentiation, the S
rocess of protein binding and dimerization and the pro-easome activity. Other ATF-3-responsive genes are struc-ural components of the Golgi apparatus and of otherrganelles, or regulate macrophage, Toll-like receptorTLR)-mediated and extra-cellular matrix inflammatory re-ponses (Ho et al., 2008; Kiryu-Seo et al., 2008; Song etl., 2008; Beyrouthy et al., 2008; Seijffers et al., 2007;sin-Mei et al., 2007; Gilchrist et al., 2006; Vlug et al.,005; Chen and Wang, 2004; Jiang et al., 2004). A per-entage of these gene candidates appear to be differen-ially regulated in the spinal cord from the transgenic rat,ccording to our gene expression analysis (Table 3; Sup-lementary file 1). Among these, Growth Arrest and DNAamage-inducible gene 45 (Gadd45), Cyclin B2 (CCNB2),unB and DNA damage-inducible transcript 4 (DDIT4-HOP) have been reported to play an important role in the
egulation of the cell cycle and survival under conditions oftress, including amino acid and serum deprivation, UV
rradiation and proteasome inhibition (Jiang et al., 2007;eyrouthy et al., 2008; Gonzalez de Aguilar et al., 2008).heir activity as transcription factors and as DNA bindingroteins in concert with ATF-3 has been extensively de-cribed. Gadd45 is a stress-responsive protein that regu-
ates genome stability, apoptosis and immune responses,nd acts on the mitotic cell-cycle, preventing cell cyclerrest. Gadd153, Gadd45 and Sectm1 are also included inhe gene categories GO:0005793_ER-Golgi_compart-ent, GO:0005773_vacuole and GO:0005764_lysosome
Table 3). The genes listed in these gene categories su-ervise the maintenance of the functional and structural
ntegrity of the Golgi apparatus and of the endoplasmiceticulum and may become implicated in the process ofrganelle fragmentation in neurodegenerative conditionsLehotský et al., 2009). Up-regulation and co-localizationf CHOP/Gadd153 and of ATF-3 have been reported in aopulation of motor neurons in spinal cord from the G93A
nd at end-stage disease using High-Thoroughput GO-Miner analysisspinal cord tissue as reference). In grey shade those gene categoriesconsidered
Late stage
* % FDR Enrichment* % FDR
0.0086 4.2 00.0033 5.2 00.0128 7.7 00.0670 5.9 00.1302 6.1 0.00080.7647 3.18 0.00160.5632 5.9 0.00380.0715 3.4 0.00360.1690 4.3 0.00730.6515 3.7 0.01420.7621 2.85 0.01530.4676 2.84 0.0433
cording to our gene expression analysis, that are included in a specific
t onset asing WTe points
set
richment
518
9
d to 0.enes, ac
OD1 mouse model of ALS at disease onset, along with

s(
cwwtaknmiai
doomdroimawthmsuIoT
TtmmChfcbarsftrmpAntple
3.G
ene
cand
idat
espr
evio
usly
repo
rted
toin
tera
ctw
ithA
TF
-3or
tobe
dow
n-st
ream
targ
ets
(und
erdi
ffere
ntex
perim
enta
lco
nditi
ons
ofst
ress
,re
fere
nces
inth
ela
stco
lum
n)fo
und
tobe
eren
tially
regu
late
dat
onse
tan
dat
end-
stag
edi
seas
ein
spin
alco
rdfr
omth
eG
93A
SO
D1
rat
mod
elof
ALS
.T
hege
neca
ndid
ates
are
grou
ped
acco
rdin
gto
thei
rbi
olog
ical
func
tion.
The
tabl
eo
repo
rts
the
gene
cate
gorie
sw
hich
cont
ain
the
repo
rted
AT
F-3
-res
pons
ive
gene
s,fo
und
tobe
diffe
rent
ially
regu
late
din
our
onto
logi
cala
naly
sis
ofth
edi
ffere
ntia
llyre
gula
ted
gene
sin
tran
sgen
icd
usin
gH
igh-
Tho
roug
hput
GO
-Min
er
ctio
nG
enes
GO
cate
gorie
sR
efer
ence
s
Ons
etE
nd-s
tage
Ons
etE
nd-s
tage
llpr
olife
ratio
n,yc
leC
cnd1
,S
P1
DIN
E,
Cyc
linD
2C
cnd1
,Ju
nB,
GA
DD
45a,
SP
1D
INE
,P
irb—
GO
:004
2127
_reg
ulat
ion_
of_c
ell_
prol
ifera
tioG
O:0
0434
88_r
egul
atio
n_of
_mR
NA
_sta
bilit
yG
O:0
0015
68_b
lood
_ves
sel_
deve
lopm
ent
Kiry
u-S
eoet
al.,
2008
;O
hba
etal
.,20
04
tein
omeo
stas
is—
Psm
b8,
Mm
p11,
Mm
p12,
Mm
p23
DIN
E,
Ada
mts
1
—G
O:0
0058
39_p
rote
asom
e_co
re_c
ompl
exG
O:0
0301
62_r
egul
atio
n_of
_pro
teol
ysis
GO
:000
4175
_end
opep
tidas
e_ac
tivity
Ho
etal
.,20
08;
Ohb
aet
al.,
2004
anel
les
Sec
tm1
GA
DD
153,
Gad
d45a
,S
ectm
1G
O:0
0057
73_v
acuo
leG
O:0
0057
64_l
ysos
ome
GO
:000
5793
_ER
-Gol
gi_c
ompa
rtm
ent
GO
:000
5773
_vac
uole
GO
:000
5764
_lys
osom
e
Leho
tský
etal
.,20
09;
Vlu
get
al.,
2005
une
espo
nse
Mm
p12
IL-1
8,M
mp1
1M
mp1
2,M
mp2
3G
O:0
0450
87_i
nnat
e_im
mun
e_re
spon
seG
O:0
0069
59_h
umor
al_i
mm
une_
resp
onse
GO
:001
9724
_B_c
ell_
med
iate
d_im
mun
ity
GO
:000
5581
_col
lage
nG
O:0
0310
12_e
xtra
cellu
lar_
mat
rixG
O:0
0055
83_fi
brill
ar_c
olla
gen
GO
:000
5125
_cyt
okin
e_ac
tivity
GO
:000
6959
_hum
oral
_im
mun
e_re
spon
seG
O:0
0197
24_B
_cel
l_m
edia
ted_
imm
unity
Whi
tmor
eet
al.,
2007
;G
ilchr
ist
etal
.,20
06;
Ho
etal
.,20
08;
Hos
sain
-Ibr
ahim
etal
.,20
06;
Che
nan
dW
ang,
2004
;S
lent
z-K
esle
ret
al.,
1998
A. Malaspina et al. / Neuroscience 169 (2010) 812–827822
omato–dendritic ubiquitination and Golgi fragmentationVlug et al., 2005).
Our transcriptomic analysis of the transgenic spinalord has also identified the differential regulation of factorshich may modulate protein homeostasis in partnershipith ATF-3, via proteolysis, endopeptidase activity and
hrough the proteasome complex (Tables 3 and 4). ATF-3ctivation has been reported to occur in co-regulation withey proteins in this family, including the damage-inducedeuronal endopeptidase (DINE), a nerve-injury associatedolecule. It has been reported that nerve damage co-
nduces ATF-3, c-Jun, STAT3, and Sp1, which becomessociated with the GC box region of the DINE gene in
njured neurons (Kiryu-Seo et al., 2008; Ohba et al., 2004).The molecular profile of the transgenic cord at end-stage
isease shows also a substantial increase in the expressionf molecular signals linked to the unravelling of different typesf immune response, including innate, TLR-dependent, hu-oral, B-cell mediated, extracellular matrix and cytokine-in-uced inflammatory processes (GO:0045087innate immuneesponse; GO:0002460_adaptive_immune_response_based_n_somatic_recombination_of_immune_receptors_built_from_
mmunoglobulin_superfamily_domains; GO:0008009_che-okine_activity both onset and late stage; Fig. 8, Tables 3nd 4). ATF-3 is known to behave as a transcription factorith a negative regulatory effect on TLR pathways, altering
he responses that innate immunity and host defenses mayave to various types of immunological challenge (Whit-ore et al., 2007). Recent data suggest that ATF-3 down-
tream effects on inflammation may also involve the reg-lation of extra-cellular matrix metalloproteases and ofL-18, both found to be up-regulated in the spinal cord fromur animal model of ALS (Hossain-Ibrahim et al., 2006;able 3; Supplementary file 1).
DISCUSSION
his study provides further insight into the early stages ofhe pathological process in a well-characterized animalodel of ALS, when the disease end-point represented byotor neurons degeneration is still not fully reached.hanges in the expression of ATF-3 and/or of ATF-3/c-Junave been described in both motor neurons and glial cellsrom spinal cord subjected to various types of stress, in-luding mechanical injury and neurodegeneration inducedy a mutation of the SOD1 gene (Hunt et al., 2004; Vlug etl., 2005). Previously, the evaluation of GFAP differentialegulation has proved a good indicator of the timing andites of onset of the disease in both nerves and spinal cordrom the G93A SOD1 mouse model of ALS but the detec-ion of this disease-related signal was limited to non-neu-onal cells (Keller et al., 2009). The use of ATF-3 as aarker of cell functional perturbation in our study makes itossible to extend this preliminary observation to includeLS-related molecular signals in different neuronal andon-neuronal cell types, which may be potential targets ofhe disease process from a very early stage. Looking atatterns of co-regulated genes at different time points
during the disease progression, we have also obtained aTab
diff
als
cor
Fun
Ce c
Pro h
Org
Imm r

Tesio(raNcio
A
XNNXXXNNXXXNXXNNXXNNNNNNNXNNCNNXNNNXNXNNXNNNXXNN
A. Malaspina et al. / Neuroscience 169 (2010) 812–827 823
able 4. Gene candidates found to be differentially expressed by Bead-array analysis of transgenic spinal cord at both disease onset and at diseasen-stage (using spinal cord tissue from age-matched WT rats as reference). The rest of the genes found to be significantly differentially regulated inpinal cord from transgenic rats at disease onset (n�19), at disease end-stage (n�276) and at a pre-symptomatic stage (n�31) have been reportedn the Supplementary file 1 attached to this paper. The genes displaying the highest ratios of fold changes (transgenic vs. control) and the highest levelf up-regulation in the end-stage spinal cord are reported at the top of the table, with a descending order. In this table, ratios replace differential scorestransgenic vs. WT determination). Differential score values are generated by the BeadStudio analysis software and represent the level of differentialegulation when both intensity and P-value detections are taken into account. Each gene candidate gene ontology (GO) category is reported ifvailable, which corresponds to those identified by High-Throughput GO-Miner analysis of the gene expression profiled obtained from pooled samples.ote the significant increase of the level of differential regulation in end-stage transgenic spinal cord for almost all the gene candidates. The lastolumn reports a broad description of the gene’s function. The vast majority of the differentially regulated genes are involved in the modulation of themmune response, of cell growth, of the extracellular matrix and in the control of lipid metabolism. Gene candidates reporting the suffix LOC are partf the annotation system of the Illumina platform used in this study
ccession Symbol DetectionP Value*
Transgenic/Control foldchanges
GO Function
Onset End-stage
M_213372.3 Cd68_predicted 0 15 22 Immune responseM_031530.1 Ccl2 �0.0283 4.9 22 7259 Immune responseM_172333.1 Cthrc1 0 8 21 Extracellular matrixM_227485.3 Msr2_predicted 0 18 20 Immune responseM_345360.2 Galnt3_predicted �0.0283 7 20 ProteasomeM_342245.2 Postn_predicted 0 9 20 Cell growthM_012912.1 Atf3 0 8 20 Transcription factorM_145672.3 Cxcl9 0 5.5 18 6955 Immune responseM_216299.3 RGD1308734 0 10 15 Cell growthM_215117.3 Ifitm1_predicted 0 10 15 Immune responseM_215013.3 Folr2 0 8 15 ProliferationM_012610.1 Ngfr 0 7.5 14 7417 Axonal guidanceM_579554.1 Fabp4 0 7 13 Lipid metabolismM_573561.1 LOC498335 0 6 12 Immune responseM_182952.2 Cxcl11 0 4 12 6955 Immune responseM_031832.1 Lgals3 0 8 11 7157 Immune responseM_576419.1 Loxl1_predicted 0 3.8 10 Lysyl oxidaseM_215598.3 S100a11 0 3 8 Calcium bindingM_012824.1 Apoc1 0 6 8 6869 Lipid transportM_133624.1 Gbp2 0 4 8 6955 Immune responseM_001006975.1 Ms4a6b 0 3 8 Extracellular matrixM_053484.1 Gas7 0 3 8 7399 NeurogenesisM_053819.1 Timp1 0 3 8 6700 Extracellular matrixM_053734.2 Ncf1 0 7 8 6691 Immune responseM_144750.1 LOC246266 0 5 7 4622 ProteasomeM_228072.3 Itgb2_predicted �0.0086 7 7 Immune responseM_012843.2 Emp1 0 3 7 16049 Cell growthM_212525.1 Tyrobp 0 7 7 Proteasomeyp1b1 Cyp1b1 0 3 6.5 MitochondrialM_001009645.1 Kif22 0 3 6 6281 Axonal transportM_012523.1 Cd53 0 3.8 6 19735 Immune responseM_342027.1 Ms4a4a_predicted 0 5.5 6 Immune responseM_012687.1 Tbxas1 0 5 6 1516 ProstaglandinsM_019316.1 Mafb 0 4.5 6 45944 Transcription factorM_171992.2 Ccnd1 0 3.2 5 8236 Cell cycleM_573130.1 LOC497942 0 3 5 NKM_138826.2 Mt1a 0 2.6 5 6882 Zinc ion bindingM_343169.1 Adn 0 4.5 5 ProteasomeM_139341.1 Slc15a3 0 4 4.8 6857 Protein catabolismM_001009646.1 Qprt 0 3 4.5 9435 Synaptic transmissionM_217335.3 Trem2_predicted 0 5 4.3 Immune responseM_019354.1 Ucp2 0 2.4 4 159992 Immune responseM_019262.1 C1qb 0 4.5 4 6956 Immune responseM_013069.1 Cd74 0 4 4 6457 Immune responseM_573502.1 LOC498276 0 4 3.2 NKM_215541.2 LOC294942 0 3 3 NKM_024160.1 Cyba 0 3 3 16491 Oxidoreductase
M_053635.2 St14 0 3.2 3 16491 Cell migration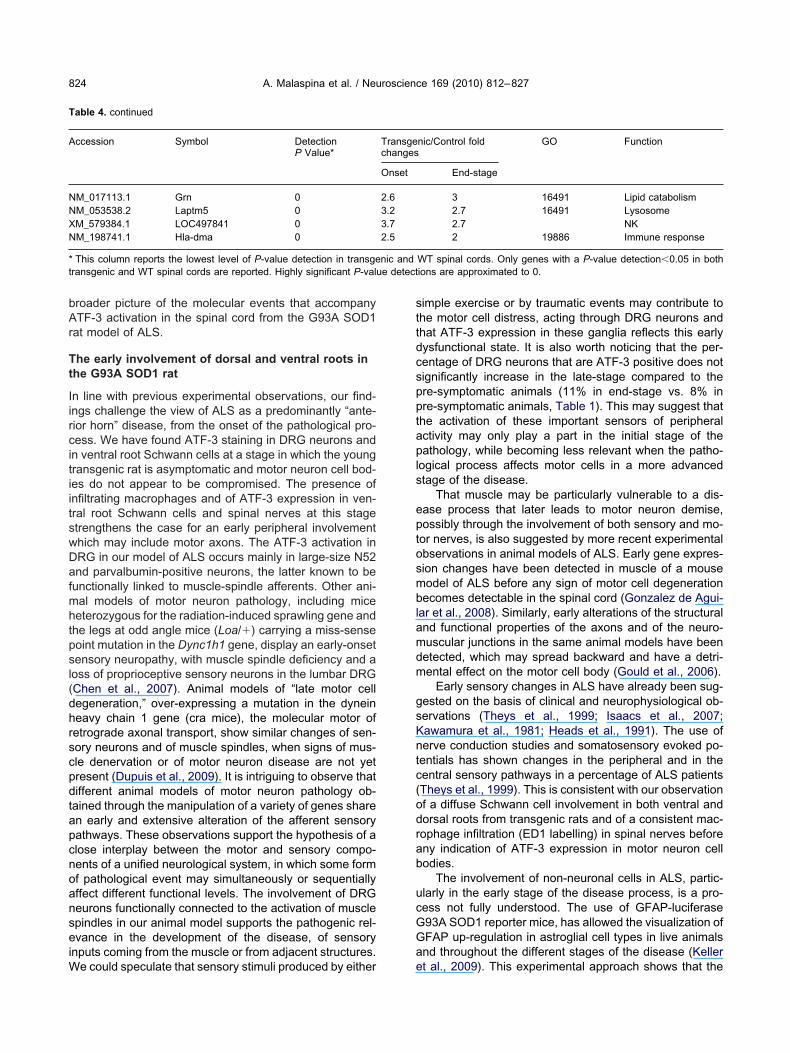
bAr
Tt
IircitiitswDafmhtpsl(dhrscpdtapcnoanseiW
sttdcspptapls
eptosmblamdm
gsKntc(odrab
ucGGa
T
A
NNXN
*t e detect
A. Malaspina et al. / Neuroscience 169 (2010) 812–827824
roader picture of the molecular events that accompanyTF-3 activation in the spinal cord from the G93A SOD1
at model of ALS.
he early involvement of dorsal and ventral roots inhe G93A SOD1 rat
n line with previous experimental observations, our find-ngs challenge the view of ALS as a predominantly “ante-ior horn” disease, from the onset of the pathological pro-ess. We have found ATF-3 staining in DRG neurons andn ventral root Schwann cells at a stage in which the youngransgenic rat is asymptomatic and motor neuron cell bod-es do not appear to be compromised. The presence ofnfiltrating macrophages and of ATF-3 expression in ven-ral root Schwann cells and spinal nerves at this stagetrengthens the case for an early peripheral involvementhich may include motor axons. The ATF-3 activation inRG in our model of ALS occurs mainly in large-size N52nd parvalbumin-positive neurons, the latter known to be
unctionally linked to muscle-spindle afferents. Other ani-al models of motor neuron pathology, including miceeterozygous for the radiation-induced sprawling gene and
he legs at odd angle mice (Loa/�) carrying a miss-senseoint mutation in the Dync1h1 gene, display an early-onsetensory neuropathy, with muscle spindle deficiency and aoss of proprioceptive sensory neurons in the lumbar DRGChen et al., 2007). Animal models of “late motor cellegeneration,” over-expressing a mutation in the dyneineavy chain 1 gene (cra mice), the molecular motor ofetrograde axonal transport, show similar changes of sen-ory neurons and of muscle spindles, when signs of mus-le denervation or of motor neuron disease are not yetresent (Dupuis et al., 2009). It is intriguing to observe thatifferent animal models of motor neuron pathology ob-ained through the manipulation of a variety of genes sharen early and extensive alteration of the afferent sensoryathways. These observations support the hypothesis of alose interplay between the motor and sensory compo-ents of a unified neurological system, in which some formf pathological event may simultaneously or sequentiallyffect different functional levels. The involvement of DRGeurons functionally connected to the activation of musclepindles in our animal model supports the pathogenic rel-vance in the development of the disease, of sensory
nputs coming from the muscle or from adjacent structures.
able 4. continued
ccession Symbol DetectionP Value*
M_017113.1 Grn 0M_053538.2 Laptm5 0M_579384.1 LOC497841 0M_198741.1 Hla-dma 0
This column reports the lowest level of P-value detection in transgeransgenic and WT spinal cords are reported. Highly significant P-valu
e could speculate that sensory stimuli produced by either e
imple exercise or by traumatic events may contribute tohe motor cell distress, acting through DRG neurons andhat ATF-3 expression in these ganglia reflects this earlyysfunctional state. It is also worth noticing that the per-entage of DRG neurons that are ATF-3 positive does notignificantly increase in the late-stage compared to there-symptomatic animals (11% in end-stage vs. 8% inre-symptomatic animals, Table 1). This may suggest thathe activation of these important sensors of peripheralctivity may only play a part in the initial stage of theathology, while becoming less relevant when the patho-
ogical process affects motor cells in a more advancedtage of the disease.
That muscle may be particularly vulnerable to a dis-ase process that later leads to motor neuron demise,ossibly through the involvement of both sensory and mo-or nerves, is also suggested by more recent experimentalbservations in animal models of ALS. Early gene expres-ion changes have been detected in muscle of a mouseodel of ALS before any sign of motor cell degenerationecomes detectable in the spinal cord (Gonzalez de Agui-
ar et al., 2008). Similarly, early alterations of the structuralnd functional properties of the axons and of the neuro-uscular junctions in the same animal models have beenetected, which may spread backward and have a detri-ental effect on the motor cell body (Gould et al., 2006).
Early sensory changes in ALS have already been sug-ested on the basis of clinical and neurophysiological ob-ervations (Theys et al., 1999; Isaacs et al., 2007;awamura et al., 1981; Heads et al., 1991). The use oferve conduction studies and somatosensory evoked po-entials has shown changes in the peripheral and in theentral sensory pathways in a percentage of ALS patientsTheys et al., 1999). This is consistent with our observationf a diffuse Schwann cell involvement in both ventral andorsal roots from transgenic rats and of a consistent mac-ophage infiltration (ED1 labelling) in spinal nerves beforeny indication of ATF-3 expression in motor neuron cellodies.
The involvement of non-neuronal cells in ALS, partic-larly in the early stage of the disease process, is a pro-ess not fully understood. The use of GFAP-luciferase93A SOD1 reporter mice, has allowed the visualization ofFAP up-regulation in astroglial cell types in live animalsnd throughout the different stages of the disease (Keller
nic/Control fold GO Function
End-stage
3 16491 Lipid catabolism2.7 16491 Lysosome2.7 NK2 19886 Immune response
WT spinal cords. Only genes with a P-value detection�0.05 in bothions are approximated to 0.
Transgechanges
Onset
2.63.23.72.5
nic and
t al., 2009). This experimental approach shows that the

dichbf2
Tc
OAtmraaeccteglAeitTtm
AteoroegiaitiwhOdqrAlcootsS
mtsiBicaApcmwlsec22taismpsldttaflr(voefiamd(sstcabgps
tafpmv
A. Malaspina et al. / Neuroscience 169 (2010) 812–827 825
isease in the G93A SOD1 mice causes a particularlyntense induction of GFAP in peripheral nerve Schwannells at disease initiation. Astrocytes and Schwann cellsave been found to strongly promote synapse formationetween spinal motor neurons and to be able to release
actors that can affect motor neuron survival (Ullian et al.,003).
he ATF-3 activation and the gene expressionhanges in the G93A SOD1spinal cord
ur study describes the time-dependent activation ofTF-3 in the spinal cord motor neurons from the onset of
he locomotor decline observed in the G93A SOD1 ratodel of ALS. ATF-3 activation in the spinal cord has been
eported following mechanical injury (Huang et al., 2007),nd in a subset of motor neurons in a SOD1-mutant micet disease onset (Vlug et al., 2005). Central axotomyvokes a transient increase in ATF-3 expression which isonfined to large diameter, neurofilament-immunoreactiveells (Huang et al., 2006). Previous studies have shownhat ATF-3 is highly induced in DRG neurons after periph-ral axonal injury and becomes down-regulated when tar-et re-innervation occurs (Tsujino et al., 2000; Bloech-
inger et al., 2004). There is no conclusive evidence thatTF-3 activation has a pro-survival effect, although ATF-3xpression in DRG cells in response to peripheral nerve
njury has been shown to increase after administration ofhe neuroprotective NGF and GDNF (Averill et al., 2004).he above observations in different conditions of cell dis-
ress may indicate that regulation of this transcription factoray be determinant for the cell’s fate.
Previous investigations conducted on animal models ofLS carrying a deletable SOD1 mutant gene have shown
hat while the expression within motor neurons of the del-terious gene mutation is a primary determinant of diseasenset and of an early phase of disease progression, theeduction of the levels of mutated microglia had little effectn the early disease phase but sharply slowed later dis-ase progression (Boillée et al., 2006). In the same trans-enic mice, the diminished expression of the mutant gene
n astrocytes did not affect onset, but delayed microglialctivation and slowed significantly the disease progression
n a later stage (Yamanaka et al., 2008a). These observa-ions indicate that onset and progression of the patholog-cal process may represent distinct disease phases, inhich diverse cell types and molecular responses mayave a timely contribute to the neuropathological process.ur gene expression analysis has also uncovered twoistinct phases in the disease process, with regard to theualitative and quantitative aspects of the transcriptionalegulation taking place in the affected spinal cord from theLS model in study. While the transgenic spinal cord mo-
ecular response in a very early stage of the disease pro-ess, when the transgenic rats demonstrate no overt signsf disease initiation, is dominated by the down-regulationf a large number of structural and cytoskeletal genes, withhe progression of the disease, we have observed theignificant activation of other types of molecular response.
imilarly to other array studies in spinal cord from animal Aodels of ALS, the vast majority of differentially regulatedranscripts at an advanced stage of the disease are repre-ented by factors contributing to the surge of the ALS-
nduced tissue inflammatory response (Malaspina and deelleroche, 2004). A broad range of molecular signals
nvolved in the maintenance of the extracellular matrix, ofell proliferation and of the proteasome activity becomelso activated (Fig. 8). The progressive up-regulation ofTF-3 in spinal cord neurons as appreciated by combiningrotein and gene expression analysis (Fig. 8, Table 4)o-exists with the expansion of different modalities of im-une responses. A broad range of molecular pathways,hich have already been described to be functionally re-
ated to AT3 under different conditions of experimentaltress, become also activated (Ho et al., 2008; Kiryu-Seot al., 2008; Song et al., 2008; Seijffers et al., 2007; Gil-hrist et al., 2006; Beyrouthy et al., 2008; Hsin-Mei et al.,007; Vlug et al., 2005; Chen and Wang, 2004; Jiang et al.,004). The significant level of differential regulation iden-ified through spinal cord tissue array analysis (Table 4)nd the fact that ATF-3 protein expression is only identified
n neuronal cells of affected spinal cord suggest that at thistage of the disease, ATF-3 plays a part in neuronal cellaintenance. This seems not to be the case in an earlierhase of the disease, when non-motor neuronal stainingeems to be a feature of the transgenic cord in a still
argely a-symptomatic rat. In a molecular environmentominated by a widespread activation of immune signals,he activation of ATF-3 seems not to be out of context. Thisranscription factor has been shown to modulate TLR-4nd a range of mediators of the extra-cellular matrix in-ammatory processes. For example, the lipopolysaccha-ide-induced expression of matrix metalloproteinasesMMP) has been reported to be suppressed by interferon-�ia induction of ATF-3 (Ho et al., 2008). Equally, the nitricxide inhibition of MMP-2 occurs via ATF-3 activation inndothelial cells (Chen and Wang, 2004). One of the mainndings in our gene expression analysis is the activation ofnumber of metalloproteases (MMP 11, 12, 23; Supple-entary file 1), which are known to possess also an en-opeptidase activity within the matrix inflammatory milieuTable 3). Another important feature of the gene expres-ion profile we have obtained from the spinal cord dis-ected from disease onset and end-stage mutant rats ishe significant up-regulation of gene categories involved inell proliferation and growth (Fig. 8). It is worth noticing thatmong those ATF-3-containing gene categories identifiedy ontological analysis of the differentially expressedenes, a fair percentage has a role in cell development,roliferation and differentiation and display a high level oftatistical significance and gene enrichment (Table 2).
The identification in our study of a temporal co-regula-ion of ATF-3 with multiple genes involved in cell prolifer-tion and in the immune response does not resolve theundamental question of whether ATF-3 activation has aro-survival or a detrimental effect for motor neurons in ourodel of genetically-induced degeneration. Previous in-
estigations have shown that experimentally-induced
TF-3 over-expression increases the intrinsic growth state
oeofAwwtctacacaeriefdsn
Atrs
A
B
B
B
C
C
C
D
G
G
G
H
H
H
H
H
H
H
I
J
J
J
K
K
K
K
K
A. Malaspina et al. / Neuroscience 169 (2010) 812–827826
f injured neurons, creating a more permissive growthnvironment (Liu et al., 2008; Seijffers et al., 2007). Basedn the current understanding of the role of this transcriptionactor in different experimental contexts, we could describeTF-3 as a “hub” at the centre of different biological net-orks, which may have diverging effects on cell survivalhen activated. Thus, the description of ATF-3 up-regula-
ion as an adaptive response appears appropriate, be-ause it may reflect a number of events causing perturba-ion of cellular homeostasis and the attempt of the cell todapt to extra- and/or intra-cellular changes. There is nolear and conclusive indication that ATF-3 could be useds disease-specific marker of the neurodegenerative pro-ess induced by ALS. Although the detection of an earlyctivation of ATF-3 in areas normally affected by the dis-ase process in animal models of ALS is a promisingesult, this preliminary observation should be extended tonclude studies on other models of neurodegeneration andventually, investigations into post-mortem tissue obtainedrom individuals affected by different neurodegenerativeisorders. This will allow a better characterization of thepecificity of ATF-3 activation as a molecular fingerprint ofeurodegeneration.
cknowledgments—We are grateful to The Royal London Hospi-al Charitable Foundation for its financial support to the ongoingesearch projects on ALS. We also thank the MND Association forponsoring our research in MND.
REFERENCES
verill A, Gregory J, Shortland PJ, Leavesley RC, King V, BradburyEJ, Mcmahon SB, Priestley JV (2004) NGF and GDNF amelioratethe increase in ATF3 expression which occurs in dorsal root gan-glion cells in response to peripheral nerve injury. Eur J Neurosci19:1437–1445.
eyrouthy MJ, Alexander KE, Baldwin A, Whitfield ML, Bass HW,McGee D, Hurt MM (2008) Identification of G1-regulated genes innormally cycling human cells. PLoS One 3:e3943.
loechlinger S, Karchewski LA, Woolf CJ (2004) Dynamic changes inglypican-1 expression in dorsal root ganglion neurons after periph-eral and central axonal injury. Eur J Neurosci 19:1119–1132.
oillée S, Yamanaka K, Lobsiger CS, Copeland NG, Jenkins NA,Kassiotis G, Kollias G, Cleveland DW (2006) Onset and progres-sion in inherited ALS determined by motor neurons and microglia.Science 312:1389–1392.
ampbell G, Hutchins K, Winterbottom J, Grenningloh G, LiebermanAR, Anderson PN (2005) Upregulation of activating transcriptionfactor 3 (ATF3) by intrinsic CNS neurons regenerating axons intoperipheral nerve grafts. Exp Neurol 192:340–347.
hen HH, Wang DL (2004) Nitric oxide inhibits matrix metalloprotein-ase-2 expression via the induction of activating transcription factor3 in endothelial cells. Mol Pharmacol 65:1130–1140.
hen XJ, Levedakou EN, Millen KJ, Wollmann RL, Soliven J, Popko B(2007) Proprioceptive sensory neuropathy in mice with a mutationin the cytoplasmic dynein heavy chain 1 gene. J Neurosci 27:14515–14524.
upuis L, Fergani A, Braunstein KE et al. (2009) Mice with a mutationin the dynein heavy chain 1 gene display sensory neuropathy butlack motor neuron disease. Exp Neurol 215:146–152.
ilchrist M, Thorsson V, Li B, Rust AG, Korb M, Kennedy K, Hai T,Bolouri H, Aderem A (2006) Systems biology approaches identifyATF3 as a negative regulator of Toll-like receptor 4. Nature 441:
173–178.onzalez de Aguilar JL, Niederhauser-Wiederkehr C, Halter B, DeTapia M, Di Scala F, Demougin P, Dupuis L, Primig M, MeiningerV, Loeffler JP (2008) Gene profiling of skeletal muscle in anamyotrophic lateral sclerosis mouse model. Physiol Genomics32:207–218.
ould TW, Buss RR, Vinsant S, Prevette D, Sun W, Knudson CM,Milligan CE, Oppenheim RW (2006) Complete dissociation of mo-tor neuron death from motor dysfunction by Bax deletion in amouse model of ALS. J Neurosci 26:8774–8786.
eads T, Pollock M, Robertson A, Sutherland WH, Allpress S (1991)Sensory nerve pathology in amyotrophic lateral sclerosis. ActaNeuropathol 82:316–320.
o HH, Antoniv TT, Ji JD, Ivashkiv LB (2008) Lipopolysaccharide-induced expression of matrix metalloproteinases in human mono-cytes is suppressed by IFN-gamma via superinduction of ATF-3and suppression of AP-1. J Immunol 181:5089–5097.
ossain-Ibrahim MK, Rezajooi K, MacNally JK, Mason MR, LiebermanAR, Anderson PN (2006) Effects of lipopolysaccharide-inducedinflammation on expression of growth-associated genes by corti-cospinal neurons. BMC Neurosci 7:8.
sin-Mei C, Lulu W, Santosh D’Mello R (2007) Inhibition of ATF-3expression by B-Raf mediates the neuroprotective action ofGW5074. J Neurochem 105:1300–1312.
uang WL, George KJ, Ibba V, Liu MC, Averill S, Quartu M, HamlynPJ, Priestley JV (2007) The characteristics of neuronal injury in astatic compression model of spinal cord injury in adult rats. EurJ Neurosci 25:362–372.
uang WL, Robson D, Liu MC, Averill S, Shortland PJ, Priestley JV(2006) Spinal cord compression and dorsal root injury causesup-regulation of activating transcription factor-3 in large diameterdorsal root ganglion neurons. Eur J Neurosci 23:273–278.
unt D, Hossain-Ibrahim K, Mason MR, Coffin RS, Lieberman AR,Winterbottom J, Anderson PN (2004) ATF3 upregulation in gliaduring Wallerian degeneration: differential expression in peripheralnerves and CNS white matter. BMC Neurosci 4:5–9.
saacs JD, Dean AF, Shaw CE, Al-Chalabi A, Mills KR, Leigh PN(2007) Amyotrophic lateral sclerosis with sensory neuropathy: partof a multisystem disorder? J Neurol Neurosurg Psychiatry 78:750–753.
iang HJ, Jiang L, Wek RC (2007) The eukaryotic initiation factor 2kinase pathway facilitates differential GADD45a expression in re-sponse to environmental stress. J Biol 282:3755–3765.
iang HY, Wek SA, McGrath BC, Lu D, Hai T, Harding HP, Wang X,Ron D, Cavener DR, Wek RC (2004) Activating transcription factor3 is integral to the eukaryotic initiation factor 2 kinase stressresponse. Mol Cell Biol 24:1365–1377.
okic N, Ling YY, Ward RR, Michael-Titus AT, Priestley JV, MalaspinaA (2007) Retinoid receptors in chronic degeneration of the spinalcord: observations in a rat model of amyotrophic lateral sclerosis.J Neurochem 103:1821–1833.
awamura Y, Dyck PJ, Shimono M, Okazaki H, Tateishi J, Doi H(1981) Morphometric comparison of the vulnerability of peripheralmotor and sensory neurons in amyotrophic lateral sclerosis. J Neu-ropathol Exp Neurol 40:667–675.
eller AF, Gravel M, Kriz J (2009) Live imaging of amyotrophic lateralsclerosis pathogenesis: disease onset is characterized by markedinduction of GFAP in Schwann cells. Glia 57:1130–1142.
huu CH, Barrozo RM, Hai T, Weinstein SL (2007) Activating tran-scription factor 3 (ATF3) represses the expression of CCL4 inmurine macrophages. Mol Immunol 44:1598–1605.
iryu-Seo S, Kato R, Ogawa T, Nakagomi S, Nagata K, Kiyama H(2008) Neuronal injury-inducible gene is synergistically regulatedby ATF3, c-Jun, and STAT3 through the interaction with Sp1 indamaged neurons. J Biol Chem 83:6988–6996.
livenyi P, Ferrante RJ, Matthews RT, Bogdanov MB, Klein K, Andre-assen OA, Mueller G, Wermer M, Kaddurah-Daouk R, Beal MF(1999) Neuroprotective effects of creatine in a transgenic animal
model of amyotrophic lateral sclerosis. Nat Med 5:347–350.
L
L
L
M
M
N
O
R
S
S
S
S
T
T
U
V
W
Y
Y
Z
S
S
A. Malaspina et al. / Neuroscience 169 (2010) 812–827 827
ehotský J, Urban P, Pavlíková M, Tatarková Z, Kaminska B, KaplánP (2009) Molecular mechanisms leading to neuroprotection/isch-emic tolerance: effect of preconditioning on the stress reaction ofendoplasmic reticulum. Cell Mol Neurobiol 29:917–925.
iu R, Chen XP, Tao LY (2008) Regulation of axonal regenerationfollowing the central nervous system injury in adult mammalian.Neurosci Bull 24:395–400.
ukas TJ, Luo WW, Mao H, Cole N, Siddique T (2006) Informatics-assisted protein profiling in a transgenic mouse model of amyotro-phic lateral sclerosis. Mol Cell Proteomics 5:1233–1244.
alaspina A, de Belleroche J (2004) Spinal cord molecular profilingprovides a better understanding of amyotrophic lateral sclerosispathogenesis. Brain Res Rev 45:213–229.
alaspina A, Jokic N, Huang WL, Priestley JV (2008) Comparativeanalysis of the time-dependent functional and molecular changesin spinal cord degeneration induced by the G93A SOD1 genemutation and by mechanical compression. BMC Genomics 9:500.
agai M, Aoki M, Miyoshi I, Kato M, Pasinelli P, Kasai N, Brown RH,Itoyama Y (2001) Rats expressing human cytosolic copper-zincsuperoxide dismutase transgenes with amyotrophic lateral sclero-sis: associated mutations develop motor neuron disease. J Neu-rosci 21:9246–9254.
hba N, Kiryu-Seo S, Maeda M, Muraoka M, Ishii M, Kiyama H (2004)Expression of damage-induced neuronal endopeptidase (DINE)mRNA in peri-infarct cortical and thalamic neurons following middlecerebral artery occlusion. J Neurochem 91:956–964.
osen DR, Siddique T, Patterson D, Figlewicz DA, Sapp P, Hentati A,Donaldson D, Goto J, O’Regan JP, Deng HX et al. (1993) Muta-tions in Cu/Zn superoxide dismutase gene are associated withfamilial amyotrophic lateral sclerosis. Nature 362:59–62.
eijffers R, Mills CD, Woolf CJ (2007) ATF3 increases the intrinsicgrowth state of DRG neurons to enhance peripheral nerve regen-eration. J Neurosci 27:7911–7920.
eilhean D, Cazeneuve C, Thuriès V, Russaouen O, Millecamps S,Salachas F, Meininger V, Leguern E, Duyckaerts C (2009) Accu-mulation of TDP-43 and alpha-actin in an amyotrophic lateral scle-rosis patient with the K17I ANG mutation. Acta Neuropathol118:561–573.
lentz-Kesler KA, Hale LP, Kaufman RE (1998) Identification andcharacterization of K12 (SECTM1), a novel human gene that en-codes a Golgi-associated protein with transmembrane and se-
creted isoforms. Genomics 47:327–340. tong D, Young YY, Shin DH, Shue S, Kim YS, Lee BH, Tong H, ChoBP (2008) Axotomy-induced dopaminergic neurodegeneration isaccompanied with c-Jun phosphorylation and activation transcrip-tion factor 3 expression. Exp Neurol 209:268–278.
heys PA, Peeters E, Robberecht W (1999) Evolution of motor andsensory deficits in amyotrophic lateral sclerosis estimated by neu-rophysiological techniques. J Neurol 246:438–442.
sujino H, Kondo E, Fukuoka T, Dai Y, Tokunaga A, Miki K, YonenobuK, Ochi T, Noguchi K (2000) Activating transcription factor 3(ATF3) induction by axotomy in sensory and motoneurons: a novelneuronal marker of nerve injury. Mol Cell Neurosci 15:170–182.
llian EM, Harris BT, Wu A, Chan JR, Barres BA (2003) Schwann cellsand astrocytes induce synapse formation by spinal motor neuronsin culture. Mol Cell Neurosci 25:241–251.
lug AS, Teuling E, Haasdijk ED, French P, Hoogenraad CC, JaarsmaD (2005) ATF3 expression precedes death of spinal motoneuronsin amyotrophic lateral sclerosis-SOD1 transgenic mice and corre-lates with c-Jun phosphorylation, CHOP expression, somato-den-dritic ubiquitination and Golgi fragmentation. Eur J Neurosci22:1881–1894.
hitmore MM, Iparraguirre A, Kubelka L, Weninger W, Hai T, WilliamsBR (2007) Negative regulation of TLR-signaling pathways by acti-vating transcription factor-3. J Immunol 179:3622–3630.
amanaka K, Chun SJ, Boillee S, Fujimori-Tonou N, Yamashita H,Gutmann DH, Takahashi R, Misawa H Cleveland DW (2008a)Astrocytes as determinants of disease progression in inheritedamyotrophic lateral sclerosis. Nat Neurosci 11:251–253.
amanaka K, Boillee S, Roberts EA, Garcia ML, McAlonis-Downes M,Mikse OR, Cleveland DW, Goldstein LS (2008b) Mutant SOD1 incell types other than motor neurons and oligodendrocytes accel-erates onset of disease in ALS mice. Proc Natl Acad Sci U S A105:7594–7599.
eeber BR, Qin H, Narasimhan S et al. (2005) High-ThroughputGoMiner, an ’industrial-strength’ integrative gene ontology tool forinterpretation of multiple-microarray experiments, with applicationto studies of Common Variable Immune Deficiency (CVID). BMCBioinformatics 6:168.
APPENDIX
upplementary data
upplementary data associated with this article can be found in
he online version, at doi:10.1016/j.neuroscience.2010.04.053.(Accepted 23 April 2010)(Available online 12 May 2010)
Related Documents











