Activation of Mouse Natural Killer T Cells Accelerates Liver Regeneration After Partial Hepatectomy HIROYUKI NAKASHIMA,* TAKUO INUI,* YOSHIKO HABU,* MANABU KINOSHITA, ‡ SHIGEAKI NAGAO, § ATSUSHI KAWAGUCHI, § SOICHIRO MIURA, § NARIYOSHI SHINOMIYA,* HIDEO YAGITA, and SHUHJI SEKI* *Department of Immunology and Microbiology, National Defense Medical College, Tokorozawa, Japan; ‡ Division of Basic Traumatology, National Defense Medical College Research Institute, Tokorozawa, Japan; § Second Department of Internal Medicine, National Defense Medical College, Tokorozawa, Japan; and the Department of Immunology, Juntendo University School of Medicine, Hongo, Bunkyo-ku, Tokyo, Japan Background & Aims: Activation of natural killer T cells with the synthetic ligand -galactosylceramide (-GalCer) in- duced hepatotoxicity through the tumor necrosis factor (TNF) and Fas-ligand–mediated pathway in aged mice. The aim of this study was to elucidate how -GalCer–activated natural killer T cells function in hepatocyte proliferation and liver regeneration in partially hepatectomized (PHx) mice. Methods: Mice were injected with -GalCer at 36 hours after 70% PHx. Hepatocyte mitosis was evaluated by either mitotic figures or proliferating cell nuclear antigen staining. The role of TNF and Fas-ligand in hepatocyte mitosis also was assessed. Results: In PHx mice injected with -GalCer, hepatocyte mitosis was greatly en- hanced at 44 hours after surgery and the increase was more obvious in aged mice than in young mice. The expression of both TNF receptor 1 and Fas-ligand in liver natural killer T cells tended to increase after -GalCer injection in PHx mice. Treat- ment of mice with anti-NK1.1 Ab 3 days before and just after hepatectomy greatly inhibited the effect of -GalCer on hepa- tocyte mitosis and liver regeneration. Furthermore, pretreat- ment of PHx mice with either anti-TNF Ab or anti-FasL Ab 1 hour before -GalCer injection mostly abrogated the increase in hepatocyte proliferation. -GalCer injection did not accelerate hepatocyte proliferation in Fas-mutated lpr mice after PHx. CD1d-/- mice without -GalCer injection showed decreased hepatocyte mitosis after PHx. Conclusions: Activated natu- ral killer T cells help hepatocyte proliferation and liver regen- eration after PHx via the TNF and Fas/Fas-ligand–mediated pathway. C umulative evidence has shown the unique properties of mouse natural killer T (NKT) cells. These cells are depen- dent on a nonclassic major histocompatibility complex class I–like molecule, CD1d, for their development; they use an invariant V14/J281/V8 gene product for their T-cell recep- tors (TCRs) 1– 4 ; and they are abundant in the liver. 5–7 These cells, when activated by either endogenous or exogenous inter- leukin-12, produce interferon (IFN)- and acquire potent anti- tumor cytotoxicity: they inhibit tumor metastasis in the liver, lung, and kidney. 5,8 –10 -galactosylceramide (-GalCer), a re- cently found synthetic ligand of NKT cells, 11,12 has further advanced the functional analysis of these cells. NKT cells acti- vated by -GalCer promptly produce both IFN- and interleu- kin-4, 12 suggesting that NKT cells may be involved in either T-helper 1 response or T-helper 2 response. Because NKT cells preactivated by interleukin-12 greatly increase both IFN- and IL-4 production after -GalCer stimulation, they may produce these 2 cytokines to enhance cellular immunity and humoral immunity, respectively, against microbes. 13 In addition, NKT cells reportedly produce tumor necrosis factor (TNF) after -GalCer stimulation, 14,15 which is a critical cytokine for the host defense against infections. 16,17 The IFN- produced by -GalCer–stimulated NKT cells ac- tivates NK cells, and NK cells inhibit liver metastases of tumors in an IFN-– dependent manner, whereas activated NKT cells themselves induce hepatocyte injury via the TNF and Fas-ligand (FasL)-mediated pathways. 18 –20 The role of NKT cells in auto- immune diseases remains controversial; NKT cells activated by -GalCer reportedly inhibit autoimmune diabetes in nonobese diabetic mice, but they also aggravate diseases in other autoim- mune mouse models. 4,21,22 Based on these findings, NKT cells could be a double-edged sword; their function and the effects of their activation thus should be examined carefully. 19 Of inter- est, the -GalCer–mediated functions of NKT cells are en- hanced in mice in an age-dependent manner 19 ; the prolifera- tion, cytokine production, antitumor cytotoxicity, and hepatotoxicity of liver mononuclear cells (MNCs) mediated or triggered by -GalCer–activated liver NKT cells became higher as mice aged. Although hepatocyte injury was induced by -GalCer–acti- vated NKT cells through TNF and FasL in aged mice, recent reports also have indicated that either TNF or an agonistic anti-Fas antibody (Ab) increased the hepatocyte proliferation in regenerating liver after partial hepatectomy (PHx). 23–27 These molecules thus can be cytotoxic against steady-state hepato- cytes yet also augment proliferation of newly regenerating hepa- tocytes, suggesting that TNF and FasL function differently against hepatocytes depending on the condition of the cells. On the other hand, the number of NKT cells and NK-T cells (CD56 T cells) in the livers of mice and human beings, respec- tively, decreases with the progress of hepatitis and their pre- ponderance is finally lost in the liver with cirrhosis, 28,29 sug- gesting that intact hepatocytes are necessary for the maintenance of hepatic NKT cells. In addition, liver NKT cells greatly increased in number and proportion in the early phase Abbreviations used in this paper: Ab, antibody; -GalCer, -galacto- sylceramide; FasL, Fas-ligand; IFN, interferon; MNC, mononuclear cell; NKT, natural killer T; PCNA, proliferating cell nuclear antigen; PCR, polymerase chain reaction; PHx, partial hepatectomy or partially hep- atectomized; SE, standard error; TCR, T-cell receptor; TNF, tumor ne- crosis factor; TNFR1, tumor necrosis factor receptor 1. © 2006 by the AGA Institute 0016-5085/06/$32.00 doi:10.1053/j.gastro.2006.08.028 BASIC–LIVER, PANCREAS, AND BILIARY TRACT GASTROENTEROLOGY 2006;131:1573–1583

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AA
HA*C�
BwdasciimchihobtmhtmhhhChrep
CdIitcltlcavkTpI
GASTROENTEROLOGY 2006;131:1573–1583
ctivation of Mouse Natural Killer T Cells Accelerates Liver Regenerationfter Partial Hepatectomy
IROYUKI NAKASHIMA,* TAKUO INUI,* YOSHIKO HABU,* MANABU KINOSHITA,‡ SHIGEAKI NAGAO,§
TSUSHI KAWAGUCHI,§ SOICHIRO MIURA,§ NARIYOSHI SHINOMIYA,* HIDEO YAGITA,� and SHUHJI SEKI*Department of Immunology and Microbiology, National Defense Medical College, Tokorozawa, Japan; ‡Division of Basic Traumatology, National Defense Medicalollege Research Institute, Tokorozawa, Japan; §Second Department of Internal Medicine, National Defense Medical College, Tokorozawa, Japan; and the
Department of Immunology, Juntendo University School of Medicine, Hongo, Bunkyo-ku, Tokyo, Japan
tic�h
tit(i�dmctehthta
vrarmctat(tpgmg
sNpac
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
ackground & Aims: Activation of natural killer T cellsith the synthetic ligand �-galactosylceramide (�-GalCer) in-uced hepatotoxicity through the tumor necrosis factor (TNF)nd Fas-ligand–mediated pathway in aged mice. The aim of thistudy was to elucidate how �-GalCer–activated natural killer Tells function in hepatocyte proliferation and liver regenerationn partially hepatectomized (PHx) mice. Methods: Mice werenjected with �-GalCer at 36 hours after 70% PHx. Hepatocyte
itosis was evaluated by either mitotic figures or proliferatingell nuclear antigen staining. The role of TNF and Fas-ligand inepatocyte mitosis also was assessed. Results: In PHx mice
njected with �-GalCer, hepatocyte mitosis was greatly en-anced at 44 hours after surgery and the increase was morebvious in aged mice than in young mice. The expression ofoth TNF receptor 1 and Fas-ligand in liver natural killer T cellsended to increase after �-GalCer injection in PHx mice. Treat-
ent of mice with anti-NK1.1 Ab 3 days before and just afterepatectomy greatly inhibited the effect of �-GalCer on hepa-ocyte mitosis and liver regeneration. Furthermore, pretreat-
ent of PHx mice with either anti-TNF Ab or anti-FasL Ab 1our before �-GalCer injection mostly abrogated the increase inepatocyte proliferation. �-GalCer injection did not accelerateepatocyte proliferation in Fas-mutated lpr mice after PHx.D1d-/- mice without �-GalCer injection showed decreasedepatocyte mitosis after PHx. Conclusions: Activated natu-al killer T cells help hepatocyte proliferation and liver regen-ration after PHx via the TNF and Fas/Fas-ligand–mediatedathway.
umulative evidence has shown the unique properties ofmouse natural killer T (NKT) cells. These cells are depen-
ent on a nonclassic major histocompatibility complex class–like molecule, CD1d, for their development; they use annvariant V�14/J�281/V�8 gene product for their T-cell recep-ors (TCRs)1– 4; and they are abundant in the liver.5–7 Theseells, when activated by either endogenous or exogenous inter-eukin-12, produce interferon (IFN)-� and acquire potent anti-umor cytotoxicity: they inhibit tumor metastasis in the liver,ung, and kidney.5,8 –10 �-galactosylceramide (�-GalCer), a re-ently found synthetic ligand of NKT cells,11,12 has furtherdvanced the functional analysis of these cells. NKT cells acti-ated by �-GalCer promptly produce both IFN-� and interleu-in-4,12 suggesting that NKT cells may be involved in either-helper 1 response or T-helper 2 response. Because NKT cellsreactivated by interleukin-12 greatly increase both IFN-� and
L-4 production after �-GalCer stimulation, they may producehese 2 cytokines to enhance cellular immunity and humoralmmunity, respectively, against microbes.13 In addition, NKTells reportedly produce tumor necrosis factor (TNF) after-GalCer stimulation,14,15 which is a critical cytokine for theost defense against infections.16,17
The IFN-� produced by �-GalCer–stimulated NKT cells ac-ivates NK cells, and NK cells inhibit liver metastases of tumorsn an IFN-�– dependent manner, whereas activated NKT cellshemselves induce hepatocyte injury via the TNF and Fas-ligandFasL)-mediated pathways.18 –20 The role of NKT cells in auto-mmune diseases remains controversial; NKT cells activated by-GalCer reportedly inhibit autoimmune diabetes in nonobeseiabetic mice, but they also aggravate diseases in other autoim-une mouse models.4,21,22 Based on these findings, NKT cells
ould be a double-edged sword; their function and the effects ofheir activation thus should be examined carefully.19 Of inter-st, the �-GalCer–mediated functions of NKT cells are en-anced in mice in an age-dependent manner19; the prolifera-ion, cytokine production, antitumor cytotoxicity, andepatotoxicity of liver mononuclear cells (MNCs) mediated orriggered by �-GalCer–activated liver NKT cells became highers mice aged.
Although hepatocyte injury was induced by �-GalCer–acti-ated NKT cells through TNF and FasL in aged mice, recenteports also have indicated that either TNF or an agonisticnti-Fas antibody (Ab) increased the hepatocyte proliferation inegenerating liver after partial hepatectomy (PHx).23–27 These
olecules thus can be cytotoxic against steady-state hepato-ytes yet also augment proliferation of newly regenerating hepa-ocytes, suggesting that TNF and FasL function differentlygainst hepatocytes depending on the condition of the cells. Onhe other hand, the number of NKT cells and NK-T cellsCD56� T cells) in the livers of mice and human beings, respec-ively, decreases with the progress of hepatitis and their pre-onderance is finally lost in the liver with cirrhosis,28,29 sug-esting that intact hepatocytes are necessary for theaintenance of hepatic NKT cells. In addition, liver NKT cells
reatly increased in number and proportion in the early phase
Abbreviations used in this paper: Ab, antibody; �-GalCer, �-galacto-ylceramide; FasL, Fas-ligand; IFN, interferon; MNC, mononuclear cell;KT, natural killer T; PCNA, proliferating cell nuclear antigen; PCR,olymerase chain reaction; PHx, partial hepatectomy or partially hep-tectomized; SE, standard error; TCR, T-cell receptor; TNF, tumor ne-rosis factor; TNFR1, tumor necrosis factor receptor 1.
© 2006 by the AGA Institute0016-5085/06/$32.00
doi:10.1053/j.gastro.2006.08.028

aapi
matN
lS
(abIb(
t
J�
sF((dTfs
NpfGcPgqppa
F(fiwTrB
ASIC
–LIVER
,PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1574 NAKASHIMA ET AL GASTROENTEROLOGY Vol. 131, No. 5
fter PHx.30 These findings led us to hypothesize that NKT cellsnd hepatocytes mutually interact, and NKT cells may in turnositively participate in the growth/proliferation of hepatocytes
n regenerating liver.In the present study we show that �-GalCer injection into PHx
ice does not induce a significant hepatic injury, but insteadccelerates hepatocyte proliferation and liver regeneration throughhe TNF and FasL-mediated pathway, and we discuss the roles ofKT cells and NK cells in the homeostasis of hepatocytes.
Materials and MethodsThis study was conducted in accordance with the guide-
ines of the Institutional Review Board for the Care of Animalubjects at the National Defense Medical College, Japan.
MiceC57BL/6 (B6) mice were obtained from Japan SLC Inc.
Shizuoka, Japan) and were maintained until 50 or 75 weeks ofge at our institution. MRL lpr/lpr (lpr) mice, MRL�/�, B6g/bg (bg) mice, and BALB/c mice were obtained from Japan SLCnc. and used at 7 weeks of age. CD1d-/- mice with a BALB/cackground31 were purchased from The Jackson LaboratoryBar Harbor, ME) and used at 7 weeks of age.
Reagents�-GalCer (KRN7000) was provided by the Pharmaceu-
igure 1. Serum ALT levels and liver histopathology in PHx and sh50-week-old) mice were subjected to PHx or sham operation, and �-Grom the retro-orbital sinus of (A) young and (B) aged mice at the indicaten each group. *P � .05. ––�––, Sham �-GalCer; –□–, PHx �-GalCer;
ith �-GalCer shown at low (100�) magnification. (D) The liver of a PHwenty-four hours after �-GalCer injection, livers were obtained fromepresentative microscopic findings from among 4 mice in each group
ical Research Laboratory of Kirin Brewery Company (Tokyo, w
apan).11,32 The mice were injected intravenously (IV) with 100g/kg body weight of �-GalCer or with vehicle.
Oligonucleotide Primers and PolymeraseChain Reaction AnalysisThe sequences of oligonucleotide primers for reverse-tran-
cription polymerase chain reaction (PCR) analysis were as follows.or mouse TNF receptor 1 (TNFR1),33 the antisense primer
CAAGATAACCAGGGGCAACAG) and the sense primerCCAACGCTACCTGAGTGAGA) were used; for mouse glyceral-ehyde-3-phosphate dehydrogenase,13 antisense (GTCCAGGGTT-CTTACTCCT) and sense (ATGACCACAGTCCATGCCAT); and
or mouse FasL,34 antisense (TGGGCCACACTCCTCGGCTC) andense (TGG TGGCTCTGGTTGGAATGGG) were used.
For quantification, the band intensity was analyzed using theational Institutes of Health (Bethesda, MD) image softwareackage (version 1.60). An aliquot of extract containing total RNArom liver NKT cells (RNeasy Mini Kit [250]; Qiagen, Hilden,ermany) was used for the reverse-transcriptase reaction. Next, the
omplementary DNA of each sample was amplified by PCR. TheCR products were subjected to electrophoresis in a 2.4% agaroseel and then were stained with ethidium bromide. PCR bands wereuantitated based on the ratio of the signal intensity of the PCRroducts to those of the internal controls (glyceraldehyde-3-phos-hate dehydrogenase). The linearity of a reverse-transcription PCRnalysis was confirmed in advance, and an estimation of each band
perated mice after �-GalCer injection. Young (7-week-old) and agedor vehicle was injected (IV) 48 hours after surgery. Sera were obtainedrs after �-GalCer or vehicle injection. Data are means � SE from 5 mice- -, PHx vehicle. (C) The liver of a sham-operated aged mouse injecteded mouse injected with �-GalCer shown at low (100�) magnification.m-operated and PHx mice and fixed in 10% formalin. Shown areg similar findings (H&E staining).
am-oalCerd hou- -□x ag
shahavin
as performed within a linear range.
tlBpmj
p
iptP
nmC(�(fflu
f�am
mmoidw
itaasmwmhpsPwscn
mtl
tm
FFos�Naatps.
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
November 2006 NATURAL KILLER T CELLS AND LIVER REGENERATION 1575
PHxThe mice underwent 70% PHx under deep ether anes-
hesia by a resection of the anterior and left lateral lobes of theiver according to the method of Higgins and Anderson.35
efore surgical close of the peritoneal space, about 300 �L ofhosphate-buffered saline (PBS) or antibody aliquots were ad-inistered into the peritoneal space. Control mice were sub-
ected to a sham operation and received PBS.
Isolation of Mononuclear CellsHepatic MNCs were prepared essentially as described
reviously.36
Flow Cytometric Analysis and Sorting ofNKT CellsFor an analysis of the FasL expression, liver MNCs were
solated 1 hour after the injection of �-GalCer or vehicle. Next, asreviously described,19 the cells were stained with fluorescein iso-hiocyanate–conjugated monoclonal Ab to TCR�� (H57-597; BD
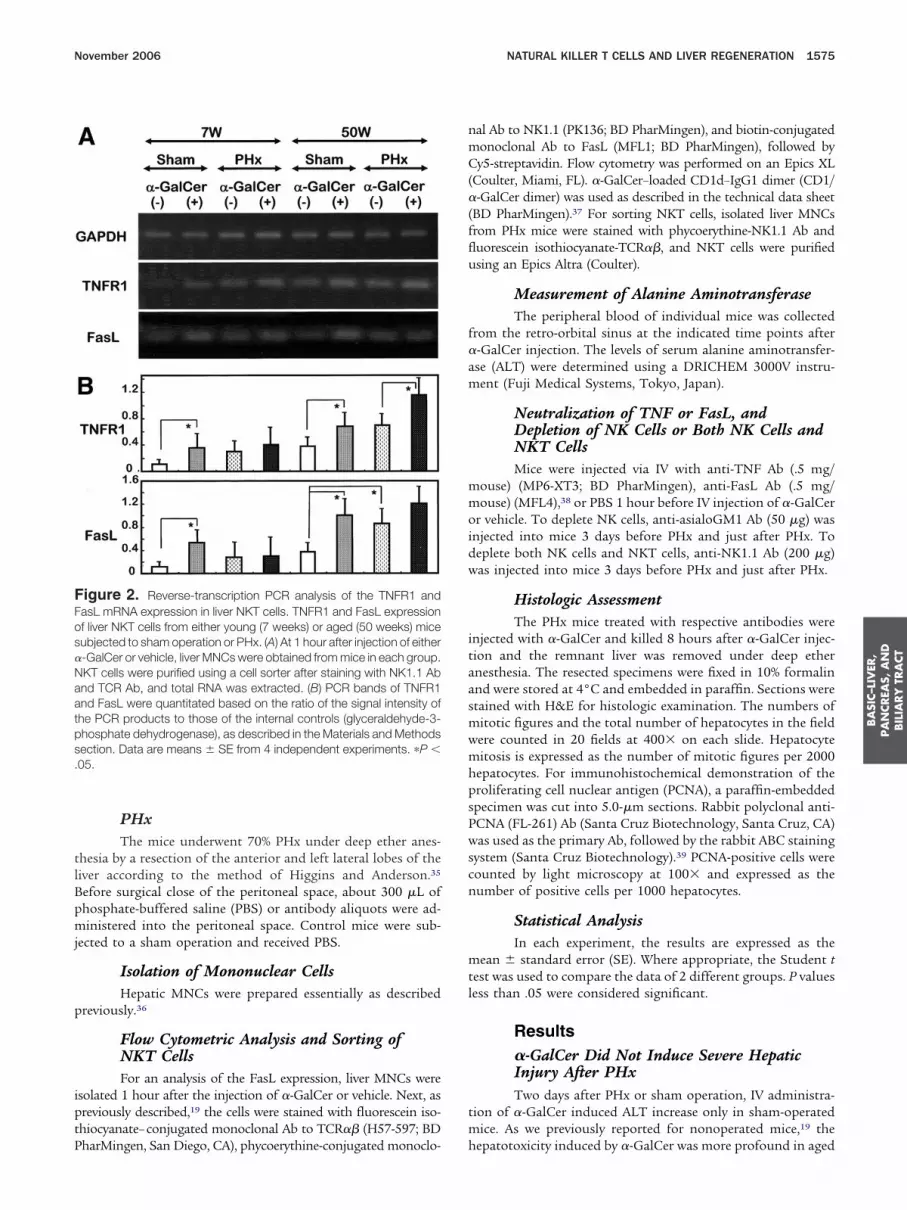
igure 2. Reverse-transcription PCR analysis of the TNFR1 andasL mRNA expression in liver NKT cells. TNFR1 and FasL expressionf liver NKT cells from either young (7 weeks) or aged (50 weeks) miceubjected to sham operation or PHx. (A) At 1 hour after injection of either-GalCer or vehicle, liver MNCs were obtained from mice in each group.KT cells were purified using a cell sorter after staining with NK1.1 Abnd TCR Ab, and total RNA was extracted. (B) PCR bands of TNFR1nd FasL were quantitated based on the ratio of the signal intensity ofhe PCR products to those of the internal controls (glyceraldehyde-3-hosphate dehydrogenase), as described in the Materials and Methodsection. Data are means � SE from 4 independent experiments. �P �05.
harMingen, San Diego, CA), phycoerythine-conjugated monoclo- h
al Ab to NK1.1 (PK136; BD PharMingen), and biotin-conjugatedonoclonal Ab to FasL (MFL1; BD PharMingen), followed byy5-streptavidin. Flow cytometry was performed on an Epics XL
Coulter, Miami, FL). �-GalCer–loaded CD1d–IgG1 dimer (CD1/-GalCer dimer) was used as described in the technical data sheet
BD PharMingen).37 For sorting NKT cells, isolated liver MNCsrom PHx mice were stained with phycoerythine-NK1.1 Ab anduorescein isothiocyanate-TCR��, and NKT cells were purifiedsing an Epics Altra (Coulter).
Measurement of Alanine AminotransferaseThe peripheral blood of individual mice was collected
rom the retro-orbital sinus at the indicated time points after-GalCer injection. The levels of serum alanine aminotransfer-se (ALT) were determined using a DRICHEM 3000V instru-ent (Fuji Medical Systems, Tokyo, Japan).
Neutralization of TNF or FasL, andDepletion of NK Cells or Both NK Cells andNKT CellsMice were injected via IV with anti-TNF Ab (.5 mg/
ouse) (MP6-XT3; BD PharMingen), anti-FasL Ab (.5 mg/ouse) (MFL4),38 or PBS 1 hour before IV injection of �-GalCer
r vehicle. To deplete NK cells, anti-asialoGM1 Ab (50 �g) wasnjected into mice 3 days before PHx and just after PHx. Toeplete both NK cells and NKT cells, anti-NK1.1 Ab (200 �g)as injected into mice 3 days before PHx and just after PHx.
Histologic AssessmentThe PHx mice treated with respective antibodies were
njected with �-GalCer and killed 8 hours after �-GalCer injec-ion and the remnant liver was removed under deep ethernesthesia. The resected specimens were fixed in 10% formalinnd were stored at 4°C and embedded in paraffin. Sections weretained with H&E for histologic examination. The numbers of
itotic figures and the total number of hepatocytes in the fieldere counted in 20 fields at 400� on each slide. Hepatocyteitosis is expressed as the number of mitotic figures per 2000
epatocytes. For immunohistochemical demonstration of theroliferating cell nuclear antigen (PCNA), a paraffin-embeddedpecimen was cut into 5.0-�m sections. Rabbit polyclonal anti-CNA (FL-261) Ab (Santa Cruz Biotechnology, Santa Cruz, CA)as used as the primary Ab, followed by the rabbit ABC staining
ystem (Santa Cruz Biotechnology).39 PCNA-positive cells wereounted by light microscopy at 100� and expressed as theumber of positive cells per 1000 hepatocytes.
Statistical AnalysisIn each experiment, the results are expressed as the
ean � standard error (SE). Where appropriate, the Student test was used to compare the data of 2 different groups. P valuesess than .05 were considered significant.
Results�-GalCer Did Not Induce Severe HepaticInjury After PHxTwo days after PHx or sham operation, IV administra-
ion of �-GalCer induced ALT increase only in sham-operatedice. As we previously reported for nonoperated mice,19 the
epatotoxicity induced by �-GalCer was more profound in aged

m(tIaiIm(Pta
anN44�ccmmLa
cwIw�sAsmsctsTtTa(
cpaeh
F(Ap re m
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1576 NAKASHIMA ET AL GASTROENTEROLOGY Vol. 131, No. 5
ice than in young mice when both had the sham operationFigure 1A, B). Surprisingly, however, in PHx mice, the hepato-oxicity was greatly reduced even in aged mice (Figure 1A, B).ncreased ALT levels in PHx mice after �-GalCer injection rel-tive to that of PHx mice after vehicle injection suggests thenjury of nonproliferating quiescent hepatocytes (Figure 1A, B).n histologic examination of the aged mice, sham-operated
ice showed massive necrotic areas in the liver parenchymaFigure 1C); however, such necrotic areas rarely were found inHx mice (Figure 1D). These findings suggest that young hepa-ocytes after mitosis and those undergoing mitosis in aged micere resistant to the FasL-mediated hepatic injury.
�-GalCer Induced TNFR1 and FasLExpression in Liver NKT Cells of PHx MiceBecause TNFR1 and FasL messenger RNAs (mRNAs)
re confined mainly to NKT cells after �-GalCer injection inonoperated mice,19,20 we examined their expression in liverKT cells. Because hepatocyte mitosis is most prominent at
0 – 44 hours after PHx,40 �-GalCer or vehicle was administered0 hours after PHx; the remnant liver was resected 1 hour after-GalCer administration and MNCs were obtained; then NKTells were isolated by sorting. TNFR1 and FasL mRNAs in NKTells were expressed more prominently in liver MNCs of oldice than in young mice with or without PHx (Figure 2), andRNAs tended to increase by �-GalCer injection (Figure 2).
iver MNCs other than NKT cells showed much lower TNFR1
igure 3. Surface FasL expression of liver NKT cells. At 1 hour after the75 weeks of age) were isolated and subjected to flow cytometric analysnti-TNF Ab was injected 12 hours before �-GalCer injection. The datercentages of FasL expression of old PHX mice with or without TNF Ab a
nd FasL mRNA levels than NKT cells (data not shown). Be- t
ause FasL expression of NKT cells increased age-dependently,19
e used 75-week-old mice for flow cytometric analysis of FasL.n PHx mice, FasL-expressing CD1/�-GalCer dimer� NKT cellsere induced almost comparably in aged mice 1 hour after-GalCer or vehicle administration, but the surface FasL inten-ity tended to increase with �-GalCer administration (Figure 3).nti-TNF Ab pretreatment (12 hours before �-GalCer injection)
ignificantly decreased FasL expression of NKT cells in PHxice injected with either �-GalCer or vehicle (Figure 3, right-
ided panels). Similarly to the case of NK1.1 Ag expressing NKTells,13 CD1/�-GalCer� NKT cells started to decrease propor-ionally at 2 hours after �-GalCer administration (data nothown), which probably is associated with internalization ofCRs from the surface of NKT cells after �-GalCer injec-
ion.41,42 Most NK cells and conventional ��T cells with highCRs did not express FasL at 1 hour after �-GalCer or vehicledministration and thereafter until 48 hours after injectiondata not shown).
Enhancement of Hepatocyte Mitosis and LiverRegeneration by �-GalCer After PHxTo investigate whether �-GalCer activation of NKT
ells could accelerate the proliferation of hepatocytes, we com-ared the frequencies of mitotic hepatocytes by histologic ex-mination. Considering the beginning of the hepatocyte prolif-ration and mitosis period, �-GalCer was administered 36ours after surgery and the number of mitotic figures in hepa-
lCer or vehicle treatment, liver MNCs of young and aged PHx or sham micesL expression on gated CD1/�-GalCer� NK1.1� NKT cells was shown.representative of 4 independent experiments with similar results. The
eans � SE from 4 mice. *P � .05, PHx group vs PHx �TNF Ab group.
�-Gais. Faa are
ocytes was counted 8 hours later. The number of mitotic
fictioi(dPcoemsd
p
wi(PmuTtNcat(cblct
mmurrt
F(trFqc1sTntwa(apducBmaPN8s
FtohmimfP*
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
November 2006 NATURAL KILLER T CELLS AND LIVER REGENERATION 1577
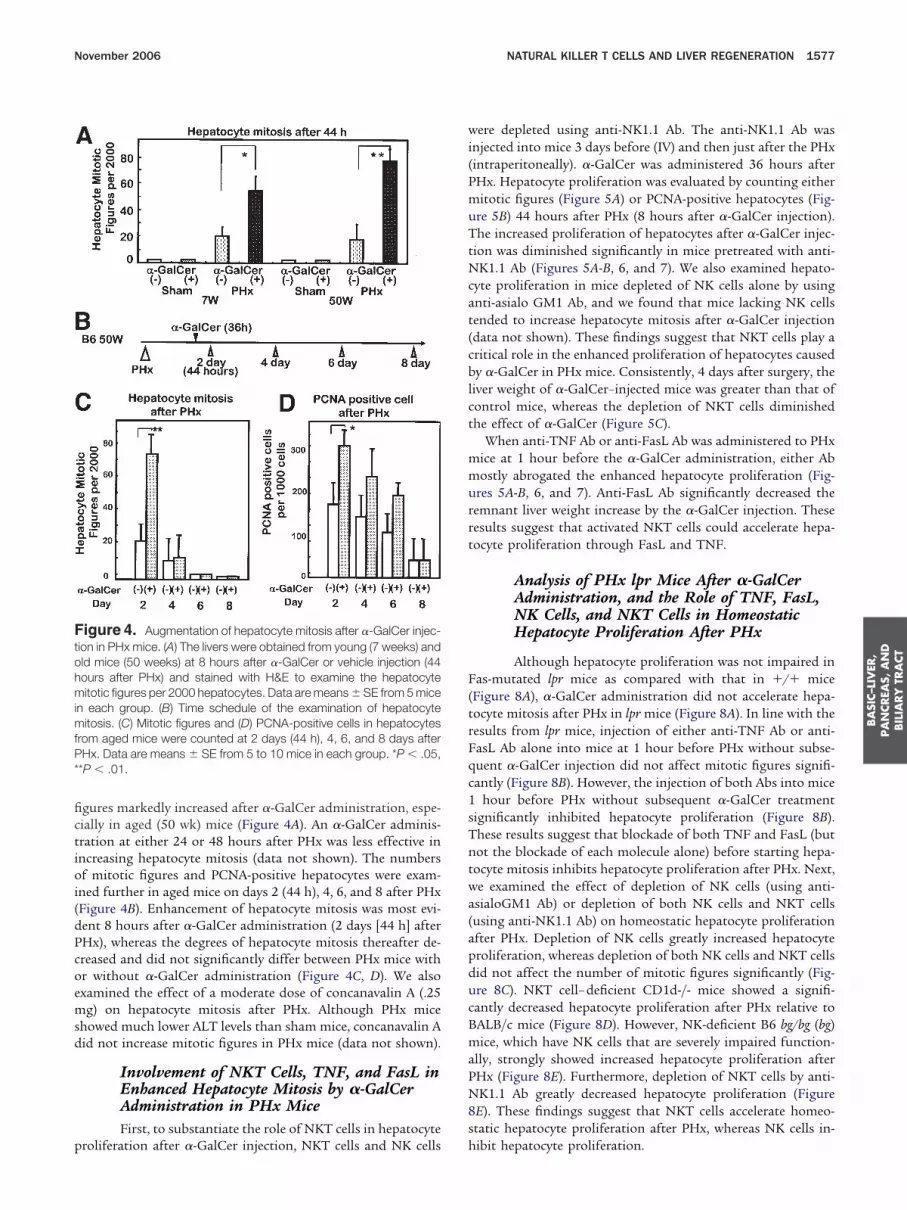
gures markedly increased after �-GalCer administration, espe-ially in aged (50 wk) mice (Figure 4A). An �-GalCer adminis-ration at either 24 or 48 hours after PHx was less effective inncreasing hepatocyte mitosis (data not shown). The numbersf mitotic figures and PCNA-positive hepatocytes were exam-
ned further in aged mice on days 2 (44 h), 4, 6, and 8 after PHxFigure 4B). Enhancement of hepatocyte mitosis was most evi-ent 8 hours after �-GalCer administration (2 days [44 h] afterHx), whereas the degrees of hepatocyte mitosis thereafter de-reased and did not significantly differ between PHx mice withr without �-GalCer administration (Figure 4C, D). We alsoxamined the effect of a moderate dose of concanavalin A (.25g) on hepatocyte mitosis after PHx. Although PHx mice
howed much lower ALT levels than sham mice, concanavalin Aid not increase mitotic figures in PHx mice (data not shown).
Involvement of NKT Cells, TNF, and FasL inEnhanced Hepatocyte Mitosis by �-GalCerAdministration in PHx MiceFirst, to substantiate the role of NKT cells in hepatocyte
igure 4. Augmentation of hepatocyte mitosis after �-GalCer injec-ion in PHx mice. (A) The livers were obtained from young (7 weeks) andld mice (50 weeks) at 8 hours after �-GalCer or vehicle injection (44ours after PHx) and stained with H&E to examine the hepatocyteitotic figures per 2000 hepatocytes. Data are means � SE from 5 mice
n each group. (B) Time schedule of the examination of hepatocyteitosis. (C) Mitotic figures and (D) PCNA-positive cells in hepatocytes
rom aged mice were counted at 2 days (44 h), 4, 6, and 8 days afterHx. Data are means � SE from 5 to 10 mice in each group. *P � .05,
*P � .01.
roliferation after �-GalCer injection, NKT cells and NK cells h
ere depleted using anti-NK1.1 Ab. The anti-NK1.1 Ab wasnjected into mice 3 days before (IV) and then just after the PHxintraperitoneally). �-GalCer was administered 36 hours afterHx. Hepatocyte proliferation was evaluated by counting eitheritotic figures (Figure 5A) or PCNA-positive hepatocytes (Fig-
re 5B) 44 hours after PHx (8 hours after �-GalCer injection).he increased proliferation of hepatocytes after �-GalCer injec-
ion was diminished significantly in mice pretreated with anti-K1.1 Ab (Figures 5A-B, 6, and 7). We also examined hepato-
yte proliferation in mice depleted of NK cells alone by usingnti-asialo GM1 Ab, and we found that mice lacking NK cellsended to increase hepatocyte mitosis after �-GalCer injectiondata not shown). These findings suggest that NKT cells play aritical role in the enhanced proliferation of hepatocytes causedy �-GalCer in PHx mice. Consistently, 4 days after surgery, the
iver weight of �-GalCer–injected mice was greater than that ofontrol mice, whereas the depletion of NKT cells diminishedhe effect of �-GalCer (Figure 5C).
When anti-TNF Ab or anti-FasL Ab was administered to PHxice at 1 hour before the �-GalCer administration, either Abostly abrogated the enhanced hepatocyte proliferation (Fig-
res 5A-B, 6, and 7). Anti-FasL Ab significantly decreased theemnant liver weight increase by the �-GalCer injection. Theseesults suggest that activated NKT cells could accelerate hepa-ocyte proliferation through FasL and TNF.
Analysis of PHx lpr Mice After �-GalCerAdministration, and the Role of TNF, FasL,NK Cells, and NKT Cells in HomeostaticHepatocyte Proliferation After PHx
Although hepatocyte proliferation was not impaired inas-mutated lpr mice as compared with that in �/� mice
Figure 8A), �-GalCer administration did not accelerate hepa-ocyte mitosis after PHx in lpr mice (Figure 8A). In line with theesults from lpr mice, injection of either anti-TNF Ab or anti-asL Ab alone into mice at 1 hour before PHx without subse-uent �-GalCer injection did not affect mitotic figures signifi-antly (Figure 8B). However, the injection of both Abs into mice
hour before PHx without subsequent �-GalCer treatmentignificantly inhibited hepatocyte proliferation (Figure 8B).hese results suggest that blockade of both TNF and FasL (butot the blockade of each molecule alone) before starting hepa-ocyte mitosis inhibits hepatocyte proliferation after PHx. Next,e examined the effect of depletion of NK cells (using anti-sialoGM1 Ab) or depletion of both NK cells and NKT cellsusing anti-NK1.1 Ab) on homeostatic hepatocyte proliferationfter PHx. Depletion of NK cells greatly increased hepatocyteroliferation, whereas depletion of both NK cells and NKT cellsid not affect the number of mitotic figures significantly (Fig-re 8C). NKT cell– deficient CD1d-/- mice showed a signifi-antly decreased hepatocyte proliferation after PHx relative toALB/c mice (Figure 8D). However, NK-deficient B6 bg/bg (bg)ice, which have NK cells that are severely impaired function-
lly, strongly showed increased hepatocyte proliferation afterHx (Figure 8E). Furthermore, depletion of NKT cells by anti-K1.1 Ab greatly decreased hepatocyte proliferation (Figure
E). These findings suggest that NKT cells accelerate homeo-tatic hepatocyte proliferation after PHx, whereas NK cells in-
ibit hepatocyte proliferation.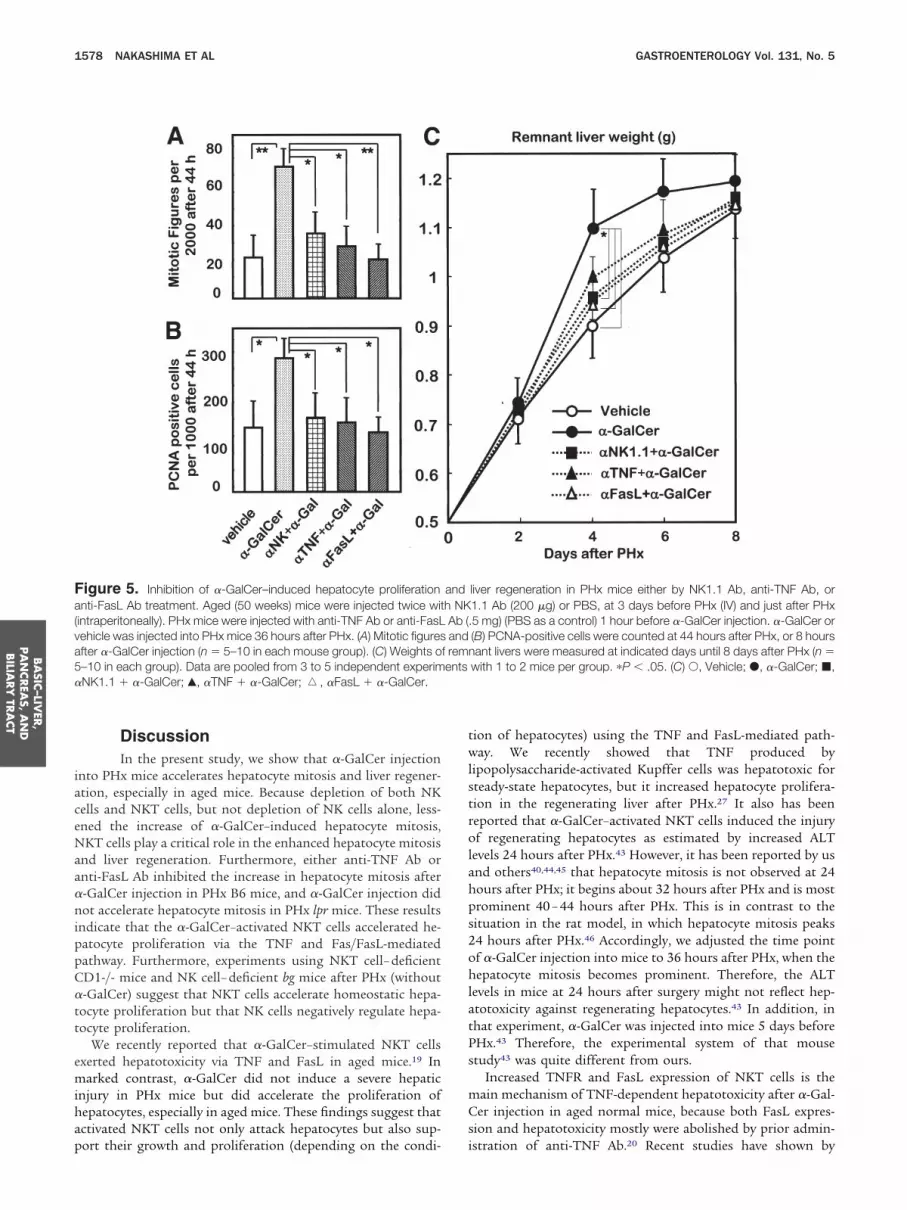
iaceNaa�nippC�tt
emihap
twlstrolahps2ohlatPs
mCs
Fa(va5�
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1578 NAKASHIMA ET AL GASTROENTEROLOGY Vol. 131, No. 5
DiscussionIn the present study, we show that �-GalCer injection
nto PHx mice accelerates hepatocyte mitosis and liver regener-tion, especially in aged mice. Because depletion of both NKells and NKT cells, but not depletion of NK cells alone, less-ned the increase of �-GalCer–induced hepatocyte mitosis,KT cells play a critical role in the enhanced hepatocyte mitosis
nd liver regeneration. Furthermore, either anti-TNF Ab ornti-FasL Ab inhibited the increase in hepatocyte mitosis after-GalCer injection in PHx B6 mice, and �-GalCer injection didot accelerate hepatocyte mitosis in PHx lpr mice. These results
ndicate that the �-GalCer–activated NKT cells accelerated he-atocyte proliferation via the TNF and Fas/FasL-mediatedathway. Furthermore, experiments using NKT cell– deficientD1-/- mice and NK cell– deficient bg mice after PHx (without-GalCer) suggest that NKT cells accelerate homeostatic hepa-
ocyte proliferation but that NK cells negatively regulate hepa-ocyte proliferation.
We recently reported that �-GalCer–stimulated NKT cellsxerted hepatotoxicity via TNF and FasL in aged mice.19 Inarked contrast, �-GalCer did not induce a severe hepatic
njury in PHx mice but did accelerate the proliferation ofepatocytes, especially in aged mice. These findings suggest thatctivated NKT cells not only attack hepatocytes but also sup-
igure 5. Inhibition of �-GalCer–induced hepatocyte proliferationnti-FasL Ab treatment. Aged (50 weeks) mice were injected twice wit
intraperitoneally). PHx mice were injected with anti-TNF Ab or anti-FasLehicle was injected into PHx mice 36 hours after PHx. (A) Mitotic figuresfter �-GalCer injection (n � 5–10 in each mouse group). (C) Weights o–10 in each group). Data are pooled from 3 to 5 independent experimNK1.1 � �-GalCer; Œ, �TNF � �-GalCer; �, �FasL � �-GalCer.
ort their growth and proliferation (depending on the condi- i
ion of hepatocytes) using the TNF and FasL-mediated path-ay. We recently showed that TNF produced by
ipopolysaccharide-activated Kupffer cells was hepatotoxic forteady-state hepatocytes, but it increased hepatocyte prolifera-ion in the regenerating liver after PHx.27 It also has beeneported that �-GalCer–activated NKT cells induced the injuryf regenerating hepatocytes as estimated by increased ALT
evels 24 hours after PHx.43 However, it has been reported by usnd others40,44,45 that hepatocyte mitosis is not observed at 24ours after PHx; it begins about 32 hours after PHx and is mostrominent 40 – 44 hours after PHx. This is in contrast to theituation in the rat model, in which hepatocyte mitosis peaks4 hours after PHx.46 Accordingly, we adjusted the time pointf �-GalCer injection into mice to 36 hours after PHx, when theepatocyte mitosis becomes prominent. Therefore, the ALT
evels in mice at 24 hours after surgery might not reflect hep-totoxicity against regenerating hepatocytes.43 In addition, inhat experiment, �-GalCer was injected into mice 5 days beforeHx.43 Therefore, the experimental system of that mousetudy43 was quite different from ours.
Increased TNFR and FasL expression of NKT cells is theain mechanism of TNF-dependent hepatotoxicity after �-Gal-er injection in aged normal mice, because both FasL expres-
ion and hepatotoxicity mostly were abolished by prior admin-
liver regeneration in PHx mice either by NK1.1 Ab, anti-TNF Ab, or1.1 Ab (200 �g) or PBS, at 3 days before PHx (IV) and just after PHx5 mg) (PBS as a control) 1 hour before �-GalCer injection. �-GalCer or(B) PCNA-positive cells were counted at 44 hours after PHx, or 8 hoursant livers were measured at indicated days until 8 days after PHx (n �
with 1 to 2 mice per group. �P � .05. (C) Œ, Vehicle; �, �-GalCer; �,
andh NKAb (.and
f remnents
stration of anti-TNF Ab.20 Recent studies have shown by

i�dNeeiipc
Pi
eaaame
F
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
November 2006 NATURAL KILLER T CELLS AND LIVER REGENERATION 1579
ntracellular staining that NKT cells may produce TNF after-GalCer injection.14,15 These findings suggest that TNF pro-uced and secreted by NKT cells and Kupffer cells activatesKT cells through their own TNFR and induce their FasL
xpression. In the present study, we have found that TNFR1xpression in liver MNCs was more profound in PHx mice thann sham-operated mice, and �-GalCer administration furtherncreased the expression of TNFR1. In addition, anti-TNF Abartially but significantly decreased FasL expression from NKT
igure 6. Mitotic figures of hepatocytes in the livers of aged mice in
ells in PHx mice. These results suggest that at 40 hours after o
Hx, when the hepatocyte mitosis is most active, NKT cellsncrease their sensitivity to TNF and express FasL.
It was recently reported that the engagement of Fas antigenxpressed on hepatocytes by an agonistic anti-Fas Ab acceler-ted hepatocyte proliferation in PHx mice.25,26 In sham-oper-ted mice, Fas stimulation by anti-Fas Ab increased hepatocytepoptosis and caused fulminant hepatic failure, whereas in PHxice, hepatic regeneration (rather than hepatic damage) was
nhanced.25,26 The involvement of Fas on hepatocytes and FasL
h group. Mitotic figures are indicated by white arrowheads.
eacn liver MNCs in liver regeneration also could explain why

Fp
NTFbfits
afsal
sat
es in
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1580 NAKASHIMA ET AL GASTROENTEROLOGY Vol. 131, No. 5
as-deficient lpr mice and FasL-deficient gld mice showed im-aired liver regeneration after anti-Fas Ab injection.26
�-GalCer–activated NKT cells exert antitumor activity via theK cell/IFN-�/perforin pathway and hepatotoxicity via theNF/FasL pathway because neutralization of TNF inhibitedasL expression in NKT cells and the hepatic injury is inducedy �-GalCer without affecting the antitumor effect.18 –20 Thesendings imply that NKT cells are a double-edged sword andhey exert a harmful effect on the host through the TNF/FasL
Figure 7. PCNA-positive hepatocyt
ystem. However, our present study indicates that NKT cells o
nd the TNF/FasL system may function in a beneficial manneror the host. Although TNF is considered a villain in septichock and organ failure, it also is essential for the host defensegainst infections16,17,47 and for hepatocyte proliferation andiver regeneration after PHx.23,48
Hepatocytes are newly generated in the portal area andenescent hepatocytes gradually move to the central vein areand undergo apoptosis after 200 days in rats.49 NKT cells andhe TNF/FasL/Fas system may normally regulate the turnover
the livers of aged mice in each group.
f hepatocytes. Whether TNF and/or FasL induces an apoptotic

otodoipi
FiTpsc
ammm
echc
itddiNfw
Fwph1o�amae
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
November 2006 NATURAL KILLER T CELLS AND LIVER REGENERATION 1581
r an anti-apoptotic signal in hepatocytes through their recep-ors may depend on the physiologic and pathologic conditionsf the hepatocytes.26,50 Indeed, an anti-apoptotic molecule wasecreased in hepatocytes of control mice but not in hepatocytesf PHx mice after anti-Fas Ab injection.26 The age-dependent
ncrease of FasL and TNFR expression in NKT cells may com-ensate for the depressed hepatocyte proliferation after dimin-
shed secretion of growth hormone in aged hosts.45
Our present results also show that the absence of TNF orasL/Fas alone did not significantly impair hepatocyte mitosis
n PHx mice without �-GalCer treatment, but depletion of bothNF and FasL did impair hepatocyte mitosis. Therefore, in therocess of homeostatic hepatocyte proliferation after PHx,ome other growth factors involved in hepatocyte mitosis mayompensate for the lack of TNF or FasL.
It has been reported that polyriboinosinic polyribocytidyliccid–activated NK cells inhibited liver regeneration in PHxice.51 We also examined the effect of �-GalCer on hepatocyteitosis in mice depleted of NK cells and found that hepatocyte
igure 8. (A) The effect of �-GalCer injection on hepatocyte mitosiseeks of age) at 8 hours after �-GalCer or vehicle injection (44 hours afteer 2000 hepatocytes. Data are means � SE from 5 lpr mice and 3 �/epatocyte mitosis in PHx B6 mice. B6 mice (n � 3) (50 weeks of age)hour before PHx, and hepatocyte mitosis was examined at 44 hours ar of both NK and NKT cells on hepatocyte mitosis in PHx B6 mice. Micg), or PBS at 3 days before PHx (IV) and just after PHx (intraperitoneallfter PHx. CD1d-/- mice (n � 5) and BALB/c mice (n � 5) (7 weeks of agitosis after PHx, and depletion of NKT and NK cells decreased hepat
t 3 days before PHx (IV) and just after PHx (intraperitoneally), B6 mice (xamined. *P � .05, **P � .01.
itosis tended to increase in these mice. Furthermore, our
xperiments using CD1d-/- mice, NK-deficient bg mice, and NKell– depleted mice suggest that NKT cells also accelerate theomeostatic proliferation of hepatocytes after PHx but that NKells are inhibitory.
Finally, it should be noted that NK-T cells in human livernfected with hepatitis C virus gradually decrease as the hepa-itis progresses, and the preponderance of NK-T cells in the liverisappears in patients with cirrhosis.29 Mouse NKT cells alsoisappear in cirrhotic liver induced by CCl4.28 These findings
ndicate that intact hepatocytes are needed to maintain liverKT cells. Taken together with the results in this study, it is
ascinating to suggest that NKT cells and hepatocytes interactith and affect each other.
References
1. Bendelac A. Positive selection of mouse NK1� T cells by CD1-expressing cortical thymocytes. J Exp Med 1995;182:2091.
2. Makino Y, Koseki H, Adachi Y, Akasaka T, Tsuchida K, TaniguchiM. Extrathymic differentiation of a T cell bearing invariant V alpha
PHx in lpr mice. The livers were obtained from MRL�/� or lpr mice (7x) and were stained with H&E to examine the hepatocyte mitotic figuresce. (B) The effect of anti-TNF Ab and/or anti-FasL Ab pretreatment ontreated with anti-TNF Ab, anti-FasL Ab, or both Abs (PBS as a control)Hx without �-GalCer injection. (C) The effect of the depletion of NK cells
3) were injected with anti-asialoGM1 Ab (50 �g), anti-NK1.1 Ab (200) NKT cell–deficient CD1d-/- mice show decreased hepatocyte mitosisre examined. (E) bg mice (7 weeks of age) show increased hepatocytemitosis. bg mice were injected twice with NK1.1 Ab (200 �g) or PBS,
eks of age) also were injected with PBS. Four mice of each group were
afterr PH� miwerefter Pe (n �y). (De) we
ocyte7 we
14J alpha 281 TCR. Int Rev Immunol 1994;11:31–46.

1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1582 NAKASHIMA ET AL GASTROENTEROLOGY Vol. 131, No. 5
3. Lantz O, Bendelac A. An invariant T cell receptor alpha chain isused by a unique subset of major histocompatibility complexclass I-specific CD4� and CD4-8- T cells in mice and humans. JExp Med 1994;180:1097–1106.
4. Van Kaer L. Alpha-galactosylceramide therapy for autoimmunediseases: prospects and obstacles. Nat Rev Immunol 2005;5:31–42.
5. Hashimoto W, Takeda K, Anzai R, Ogasawara K, Sakihara H,Sugiura K, Seki S, Kumagai K. Cytotoxic NK1.1 Ag� alpha beta Tcells with intermediate TCR induced in the liver of mice by IL-12.J Immunol 1995;154:4333–4340.
6. Tsukahara A, Seki S, Iiai T, Moroda T, Watanabe H, Suzuki S,Tada T, Hiraide H, Hatakeyama K, Abo T. Mouse liver T cells: theirchange with aging and in comparison with peripheral T cells.Hepatology 1997;26:301–309.
7. Seki S, Habu Y, Kawamura T, Takeda K, Dobashi H, Ohkawa T,Hiraide H. The liver as a crucial organ in the first line of hostdefense: the roles of Kupffer cells, natural killer (NK) cells andNK1.1 Ag� T cells in T helper 1 immune responses. Immunol Rev2000;174:35–46.
8. Seki S, Hashimoto W, Ogasawara K, Satoh M, Watanabe H, HabuY, Hiraide H, Takeda K. Antimetastatic effect of NK1� T cells onexperimental haematogenous tumour metastases in the liver andlungs of mice. Immunology 1997;92:561–566.
9. Cui J, Shin T, Kawano T, Sato H, Kondo E, Toura I, Kaneko Y,Koseki H, Kanno M, Taniguchi M. Requirement for Valpha14 NKTcells in IL-12-mediated rejection of tumors. Science 1997;278:1623–1626.
0. Takahashi M, Ogasawara K, Takeda K, Hashimoto W, SakiharaH, Kumagai K, Anzai R, Satoh M, Seki S. LPS induces NK1.1�alpha beta T cells with potent cytotoxicity in the liver of mice viaproduction of IL-12 from Kupffer cells. J Immunol 1996;156:2436–2442.
1. Kobayashi E, Motoki K, Uchida T, Fukushima H, Koezuka Y.KRN7000, a novel immunomodulator, and its antitumor activi-ties. Oncol Res 1995;7:529–534.
2. Kawano T, Cui J, Koezuka Y, Toura I, Kaneko Y, Motoki K, UenoH, Nakagawa R, Sato H, Kondo E, Koseki H, Taniguchi M. CD1d-restricted and TCR-mediated activation of V alpha14 NKT cells byglycosylceramides. Science 1997;278:1626–1629.
3. Habu Y, Uchida T, Inui T, Nakashima H, Fukasawa M, Seki S.Enhancement of the synthetic ligand-mediated function of liverNK1.1Ag� T cells in mice by interleukin-12 pretreatment. Immu-nology 2004;113:35–43.
4. Minagawa M, Deng Q, Liu ZX, Tsukamoto H, Dennert G. Activatednatural killer T cells induce liver injury by Fas and tumor necrosisfactor-alpha during alcohol consumption. Gastroenterology2004;126:1387–1399.
5. Biburger M, Tiegs G. Alpha-galactosylceramide-induced liver in-jury in mice is mediated by TNF-alpha but independent of Kupffercells. J Immunol 2005;175:1540–1550.
6. Rothe J, Mackay F, Bluethmann H, Zinkernagel R, Lesslauer W.Phenotypic analysis of TNFR1-deficient mice and characterizationof TNFR1-deficient fibroblasts in vitro. Circ Shock 1994;44:51–56.
7. Muller M, Althaus R, Frohlich D, Frei K, Eugster HP. Reducedantilisterial activity of TNF-deficient bone marrow-derived macro-phages is due to impaired superoxide production. Eur J Immunol1999;29:3089–3097.
8. Nakagawa R, Nagafune I, Tazunoki Y, Ehara H, Tomura H, IijimaR, Motoki K, Kamishohara M, Seki S. Mechanisms of the anti-metastatic effect in the liver and of the hepatocyte injury inducedby alpha-galactosylceramide in mice. J Immunol 2001;166:6578–6584.
9. Inui T, Nakagawa R, Ohkura S, Habu Y, Koike Y, Motoki K,Kuranaga N, Fukasawa M, Shinomiya N, Seki S. Age-associated
augmentation of the synthetic ligand-mediated function of mouseNK1.1 ag(�) T cells: their cytokine production and hepatotoxicityin vivo and in vitro. J Immunol 2002;169:6127–6132.
0. Inui T, Nakashima H, Habu Y, Nakagawa R, Fukasawa M, Ki-noshita M, Shinomiya N, Seki S. Neutralization of tumor necrosisfactor abrogates hepatic failure induced by alpha-galactosylcer-amide without attenuating its antitumor effect in aged mice.J Hepatol 2005;43:670–678.
1. Morshed SR, Mannoor K, Halder RC, Kawamura H, Bannai M,Sekikawa H, Watanabe H, Abo T. Tissue-specific expansion ofNKT and CD5�B cells at the onset of autoimmune disease in(NZBxNZW)F1 mice. Eur J Immunol 2002;32:2551–2561.
2. Chiba A, Kaieda S, Oki S, Yamamura T, Miyake S. The involve-ment of V(alpha)14 natural killer T cells in the pathogenesis ofarthritis in murine models. Arthritis Rheum 2005;52:1941–1948.
3. Akerman P, Cote P, Yang SQ, McClain C, Nelson S, Bagby GJ,Diehl AM. Antibodies to tumor necrosis factor-alpha inhibit liverregeneration after partial hepatectomy. Am J Physiol 1992;263:G579–G585.
4. Webber EM, Bruix J, Pierce RH, Fausto N. Tumor necrosis factorprimes hepatocytes for DNA replication in the rat. Hepatology1998;28:1226–1234.
5. Takehara T, Hayashi N, Mita E, Kanto T, Tatsumi T, Sasaki Y,Kasahara A, Hori M. Delayed Fas-mediated hepatocyte apoptosisduring liver regeneration in mice: hepatoprotective role of TNFalpha. Hepatology 1998;27:1643–1651.
6. Desbarats J, Newell MK. Fas engagement accelerates liver re-generation after partial hepatectomy. Nat Med 2000;6:920–923.
7. Kinoshita M, Uchida T, Nakashima H, Ono S, Seki S, Hiraide H.Opposite effects of enhanced tumor necrosis factor-alpha pro-duction from Kupffer cells by gadolinium chloride on liver injury/mortality in endotoxemia of normal and partially hepatectomizedmice. Shock 2005;23:65–72.
8. Kawachi Y, Arai K, Moroda T, Kawamura T, Umezu H, Naito M,Ohtsuka K, Hasegawa K, Takahashi-Iwanaga H, Iwanaga T, et al.Supportive cellular elements for hepatic T cell differentiation: Tcells expressing intermediate levels of the T cell receptor arecytotoxic against syngeneic hepatoma, and are lost after hepa-tocyte damage. Eur J Immunol 1995;25:3452–3459.
9. Kawarabayashi N, Seki S, Hatsuse K, Ohkawa T, Koike Y, AiharaT, Habu Y, Nakagawa R, Ami K, Hiraide H, Mochizuki H. Decreaseof CD56(�)T cells and natural killer cells in cirrhotic livers withhepatitis C may be involved in their susceptibility to hepatocellu-lar carcinoma. Hepatology 2000;32:962–969.
0. Minagawa M, Oya H, Yamamoto S, Shimizu T, Bannai M,Kawamura H, Hatakeyama K, Abo T. Intensive expansion ofnatural killer T cells in the early phase of hepatocyte regenerationafter partial hepatectomy in mice and its association with sym-pathetic nerve activation. Hepatology 2000;31:907–915.
1. Smiley ST, Kaplan MH, Grusby MJ. Immunoglobulin E productionin the absence of interleukin-4-secreting CD1-dependent cells.Science 1997;275:977–979.
2. Morita M, Motoki K, Akimoto K, Natori T, Sakai T, Sawa E, YamajiK, Koezuka Y, Kobayashi E, Fukushima H. Structure-activity rela-tionship of alpha-galactosylceramides against B16-bearing mice.J Med Chem 1995;38:2176–2187.
3. Walter U, Franzke A, Sarukhan A, Zober C, von Boehmer H, BuerJ, Lechner O. Monitoring gene expression of TNFR family mem-bers by beta-cells during development of autoimmune diabetes.Eur J Immunol 2000;30:1224–1232.
4. Medana I, Li Z, Flugel A, Tschopp J, Wekerle H, Neumann H. Fasligand (CD95L) protects neurons against perforin-mediated Tlymphocyte cytotoxicity. J Immunol 2001;167:674–681.
5. Higgins GM, Anderson RM. Restoration of the liver of the white
rat following partial surgical removal. Arch Pathol 1931;12:186.
3
3
3
3
4
4
4
4
4
4
4
4
4
4
5
5
INf
(
November 2006 NATURAL KILLER T CELLS AND LIVER REGENERATION 1583
6. Ogasawara K, Takeda K, Hashimoto W, Satoh M, Okuyama R,Yanai N, Obinata M, Kumagai K, Takada H, Hiraide H, Seki S.Involvement of NK1� T cells and their IFN-gamma production inthe generalized Shwartzman reaction. J Immunol 1998;160:3522–3527.
7. Schumann J, Voyle RB, Wei BY, MacDonald HR. Cutting edge:influence of the TCR V beta domain on the avidity of CD1d:alpha-galactosylceramide binding by invariant V alpha 14 NKT cells.J Immunol 2003;170:5815–5819.
8. Kayagaki N, Yamaguchi N, Nagao F, Matsuo S, Maeda H, Oku-mura K, Yagita H. Polymorphism of murine Fas ligand that affectsthe biological activity. Proc Natl Acad Sci U S A 1997;94:3914–3919.
9. Bravo R, Macdonald-Bravo H. Existence of two populations ofcyclin/proliferating cell nuclear antigen during the cell cycle:association with DNA replication sites. J Cell Biol 1987;105:1549–1554.
0. Inui T, Shinomiya N, Fukasawa M, Kobayashi M, Kuranaga N,Ohkura S, Seki S. Growth-related signaling regulates activation oftelomerase in regenerating hepatocytes. Exp Cell Res 2002;273:147–156.
1. Wilson MT, Johansson C, Olivares-Villagomez D, Singh AK, StanicAK, Wang CR, Joyce S, Wick MJ, Van Kaer L. The response ofnatural killer T cells to glycolipid antigens is characterized bysurface receptor down-modulation and expansion. Proc Natl AcadSci U S A 2003;100:10913–10918.
2. Crowe NY, Uldrich AP, Kyparissoudis K, Hammond KJ, HayakawaY, Sidobre S, Keating R, Kronenberg M, Smyth MJ, Godfrey DI.Glycolipid antigen drives rapid expansion and sustained cytokineproduction by NK T cells. J Immunol 2003;171:4020–4027.
3. Ito H, Ando K, Nakayama T, Taniguchi M, Ezaki T, Saito K,Takemura M, Sekikawa K, Imawari M, Seishima M, Moriwaki H.Role of Valpha 14 NKT cells in the development of impaired liver
regeneration in vivo. Hepatology 2003;38:1116–1124. N4. Noguchi S, Ohba Y, Oka T. Influence of epidermal growth factoron liver regeneration after partial hepatectomy in mice. J Endo-crinol 1991;128:425–431.
5. Krupczak-Hollis K, Wang X, Dennewitz MB, Costa RH. Growthhormone stimulates proliferation of old-aged regenerating liverthrough forkhead box m1b. Hepatology 2003;38:1552–1562.
6. Michalopoulos GK, DeFrances MC. Liver regeneration. Science1997;276:60–66.
7. Bopst M, Garcia I, Guler R, Olleros ML, Rulicke T, Muller M, WyssS, Frei K, Le Hir M, Eugster HP. Differential effects of TNF andLTalpha in the host defense against M. bovis BCG. Eur J Immunol2001;31:1935–1943.
8. Yamada Y, Kirillova I, Peschon JJ, Fausto N. Initiation of livergrowth by tumor necrosis factor: deficient liver regeneration inmice lacking type I tumor necrosis factor receptor. Proc Natl AcadSci U S A 1997;94:1441–1446.
9. Zajicek G, Oren R, Weinreb M Jr. The streaming liver. Liver1985;5:293–300.
0. Yuan J. Transducing signals of life and death. Curr Opin Cell Biol1997;9:247–251.
1. Sun R, Gao B. Negative regulation of liver regeneration by innateimmunity (natural killer cells/interferon-gamma). Gastroenterol-ogy 2004;127:1525–1539.
Received December 6, 2005. Accepted July 28, 2006.Address requests for reprints to: Shuhji Seki, MD, Department of
mmunology and Microbiology, National Defense Medical College, 3-2amiki, Tokorozawa 359-8513, Japan. e-mail: [email protected];
ax: (81) 42-996-5194.Supported in part by a grant-in-aid for a Special Research Program
Host Stress Responses to Internal and External Factors) from the
ational Defense Medical College (N.S. and S.S.).BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
Related Documents











