1664 P ulmonary artery hypertension (PH) occurring as an idiopathic condition or associated with an underlying disease is an unexplained disorder whose severe forms in adults and neonates are fatal and for which no satisfactory treatment is available. Hyperplasia of pulmonary-artery smooth muscle cells (PA-SMCs) is the primary determinant of the pulmonary vessel remodeling process that under- lies PH. 1 Similarities exist between cancer and PH. Thus, cultured PA-SMCs and pulmonary-artery endothelial cells (PA-ECs) from patients with PH show growth dysregula- tion, 2–5 PA-SMCs from patients with PH have decreased susceptibility to apoptosis, 6 remodeled pulmonary vessels express cancer biomarkers, 7 and drugs designed to treat malignant proliferations exert beneficial effects in animal models of PH. 8 Clinical Perspective on p 1676 One common abnormality in cancer is an inactivating mutation in the gene encoding the powerful growth-suppressive and proapoptotic transcription factor p53. 9 Also common is posttranslational p53 inactivation via interaction of the p53 protein with its negative regulator MDM2 (murine double minute 2), a specific p53 ubiquitin ligase and transcriptional inhibitor. 10,11 MDM2 is overexpressed in many human tumors, suggesting that p53-MDM2 interactions may hold promise as a target for cancer therapy. 12 Nutlins are cis-imidazoline analogs recently developed as anticancer agents. 13 These selective small-molecule p53-MDM2 binding inhibitors stabilize p53, thereby increasing the expression of genes targeted by p53 including the antiproliferative gene BTG2, the prosenescent gene p21, and the proapoptotic genes Bax and PUMA. 14 The most potent Nutlin, Nutlin-3a, exhibits antitumor activity via p53 activation in various cancer cells and induces tumor regression when administered chronically to mice. 13 Nutlin-3a Background—Induction of cellular senescence through activation of the p53 tumor suppressor protein is a new option for treating proliferative disorders. Nutlins prevent the ubiquitin ligase MDM2 (murine double minute 2), a negative p53 regulator, from interacting with p53. We hypothesized that cell senescence induced by Nutlin-3a exerted therapeutic effects in pulmonary hypertension (PH) by limiting the proliferation of pulmonary artery smooth muscle cells (PA-SMCs). Methods and Results—Nutlin-3a treatment of cultured human PA-SMCs resulted in cell growth arrest with the induction of senescence but not apoptosis; increased phosphorylated p53 protein levels; and expression of p53 target genes including p21, Bax, BTG2, and MDM2. Daily intraperitoneal Nutlin-3a treatment for 3 weeks dose-dependently reduced PH, right ventricular hypertrophy, and distal pulmonary artery muscularization in mice exposed to chronic hypoxia or SU5416/ hypoxia. Nutlin-3a treatment also partially reversed PH in chronically hypoxic or transgenic mice overexpressing the serotonin-transporter in SMCs (SM22-5HTT + mice). In these mouse models of PH, Nutlin-3a markedly increased senescent p21-stained PA-SMCs; lung p53, p21, and MDM2 protein levels; and p21, Bax, PUMA, BTG2, and MDM2 mRNA levels; but induced only minor changes in control mice without PH. Marked MDM2 immunostaining was seen in both mouse and human remodeled pulmonary vessels, supporting the use of Nutlins as a PH-targeted therapy. PH prevention or reversal by Nutlin-3a required lung p53 stabilization and increased p21 expression, as indicated by the absence of Nutlin-3a effects in hypoxia-exposed p53 −/− and p21 −/− mice. Conclusions—Nutlin-3a may hold promise as a prosenescence treatment targeting PA-SMCs in PH. (Circulation. 2013;127:1664-1676.) Key Words: cell aging ◼ hypertension ◼ muscles ◼ pulmonary artery ◼ ventricular remodeling © 2013 American Heart Association, Inc. Circulation is available at http://circ.ahajournals.org DOI: 10.1161/CIRCULATIONAHA.113.002434 Received February 1, 2012; accepted March 8, 2013. From INSERM U955 and Département de Physiologie (N.M., E.M., S.A., G.G.-B., M.S., A.H., L.B., J.B., V.A., S.A.) and Service de Cardiologie (J.-L.D.-R.), Hôpital Henri Mondor, AP-HP, Université Paris-Est Creteil (UPEC), Créteil, France and DHU ATVB; and CarMeN INSERM Unit 1060, Université de Lyon, Lyon, France (G.D.). The online-only Data Supplement is available with this article at http://circ.ahajournals.org/lookup/suppl/doi:10.1161/CIRCULATIONAHA. 113.002434/-/DC1. Correspondence to Serge Adnot, MD, PhD, Hôpital Henri Mondor, Service de Physiologie-Explorations Fonctionnelles, 94010, Créteil, France. E-mail [email protected] Activation of Lung p53 by Nutlin-3a Prevents and Reverses Experimental Pulmonary Hypertension Nathalie Mouraret, MSC; Elisabeth Marcos, MSC; Shariq Abid, MSC; Guillaume Gary-Bobo, PhD; Mirna Saker, MSC; Amal Houssaini, MSC; Jean-Luc Dubois-Rande, MD; Laurent Boyer, MD; Jorge Boczkowski, MD, PhD; Geneviève Derumeaux, MD; Valérie Amsellem, PhD; Serge Adnot, MD, PhD Hypertension by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from by guest on February 8, 2016 http://circ.ahajournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1664
Pulmonary artery hypertension (PH) occurring as an idiopathic condition or associated with an underlying
disease is an unexplained disorder whose severe forms in adults and neonates are fatal and for which no satisfactory treatment is available. Hyperplasia of pulmonary-artery smooth muscle cells (PA-SMCs) is the primary determinant of the pulmonary vessel remodeling process that under-lies PH.1 Similarities exist between cancer and PH. Thus, cultured PA-SMCs and pulmonary-artery endothelial cells (PA-ECs) from patients with PH show growth dysregula-tion,2–5 PA-SMCs from patients with PH have decreased susceptibility to apoptosis,6 remodeled pulmonary vessels express cancer biomarkers,7 and drugs designed to treat malignant proliferations exert beneficial effects in animal models of PH.8
Clinical Perspective on p 1676
One common abnormality in cancer is an inactivating mutation in the gene encoding the powerful growth-suppressive and proapoptotic transcription factor p53.9 Also common is posttranslational p53 inactivation via interaction of the p53 protein with its negative regulator MDM2 (murine double minute 2), a specific p53 ubiquitin ligase and transcriptional inhibitor.10,11 MDM2 is overexpressed in many human tumors, suggesting that p53-MDM2 interactions may hold promise as a target for cancer therapy.12 Nutlins are cis-imidazoline analogs recently developed as anticancer agents.13 These selective small-molecule p53-MDM2 binding inhibitors stabilize p53, thereby increasing the expression of genes targeted by p53 including the antiproliferative gene BTG2, the prosenescent gene p21, and the proapoptotic genes Bax and PUMA.14 The most potent Nutlin, Nutlin-3a, exhibits antitumor activity via p53 activation in various cancer cells and induces tumor regression when administered chronically to mice.13 Nutlin-3a
Background—Induction of cellular senescence through activation of the p53 tumor suppressor protein is a new option for treating proliferative disorders. Nutlins prevent the ubiquitin ligase MDM2 (murine double minute 2), a negative p53 regulator, from interacting with p53. We hypothesized that cell senescence induced by Nutlin-3a exerted therapeutic effects in pulmonary hypertension (PH) by limiting the proliferation of pulmonary artery smooth muscle cells (PA-SMCs).
Methods and Results—Nutlin-3a treatment of cultured human PA-SMCs resulted in cell growth arrest with the induction of senescence but not apoptosis; increased phosphorylated p53 protein levels; and expression of p53 target genes including p21, Bax, BTG2, and MDM2. Daily intraperitoneal Nutlin-3a treatment for 3 weeks dose-dependently reduced PH, right ventricular hypertrophy, and distal pulmonary artery muscularization in mice exposed to chronic hypoxia or SU5416/hypoxia. Nutlin-3a treatment also partially reversed PH in chronically hypoxic or transgenic mice overexpressing the serotonin-transporter in SMCs (SM22-5HTT+ mice). In these mouse models of PH, Nutlin-3a markedly increased senescent p21-stained PA-SMCs; lung p53, p21, and MDM2 protein levels; and p21, Bax, PUMA, BTG2, and MDM2 mRNA levels; but induced only minor changes in control mice without PH. Marked MDM2 immunostaining was seen in both mouse and human remodeled pulmonary vessels, supporting the use of Nutlins as a PH-targeted therapy. PH prevention or reversal by Nutlin-3a required lung p53 stabilization and increased p21 expression, as indicated by the absence of Nutlin-3a effects in hypoxia-exposed p53−/− and p21−/− mice.
Conclusions—Nutlin-3a may hold promise as a prosenescence treatment targeting PA-SMCs in PH. (Circulation. 2013;127:1664-1676.)
Key Words: cell aging ◼ hypertension ◼ muscles ◼ pulmonary artery ◼ ventricular remodeling
© 2013 American Heart Association, Inc.
Circulation is available at http://circ.ahajournals.org DOI: 10.1161/CIRCULATIONAHA.113.002434
Received February 1, 2012; accepted March 8, 2013.From INSERM U955 and Département de Physiologie (N.M., E.M., S.A., G.G.-B., M.S., A.H., L.B., J.B., V.A., S.A.) and Service de Cardiologie
(J.-L.D.-R.), Hôpital Henri Mondor, AP-HP, Université Paris-Est Creteil (UPEC), Créteil, France and DHU ATVB; and CarMeN INSERM Unit 1060, Université de Lyon, Lyon, France (G.D.).
The online-only Data Supplement is available with this article at http://circ.ahajournals.org/lookup/suppl/doi:10.1161/CIRCULATIONAHA. 113.002434/-/DC1.
Correspondence to Serge Adnot, MD, PhD, Hôpital Henri Mondor, Service de Physiologie-Explorations Fonctionnelles, 94010, Créteil, France. E-mail [email protected]
Activation of Lung p53 by Nutlin-3a Prevents and Reverses Experimental Pulmonary Hypertension
Nathalie Mouraret, MSC; Elisabeth Marcos, MSC; Shariq Abid, MSC; Guillaume Gary-Bobo, PhD; Mirna Saker, MSC; Amal Houssaini, MSC; Jean-Luc Dubois-Rande, MD; Laurent Boyer, MD;
Jorge Boczkowski, MD, PhD; Geneviève Derumeaux, MD; Valérie Amsellem, PhD; Serge Adnot, MD, PhD
115,18,162
Hypertension
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from

Mouraret et al Role of Cell Senescence in Pulmonary Hypertension 1665
is being tested for the treatment of lymphoblastic leukemia in humans.15
Treatments that activate p53 in PA-SMCs might be capable of limiting or reversing the pulmonary vascular remodeling process that characterizes PH. The ability of Nutlin-3a to sta-bilize and activate p53 in target cells depends on the expres-sion or activity of MDM2 in these cells. MDM2 dysregulation has been reported in SMCs at sites of vascular injury,16 as well as in human atherosclerotic tissues and in vascular SMCs from patients with primary aldosteronism.17,18 Moreover, MDM2 expression can be induced by hypoxia or by p53.11,19 Whether MDM2 expression or activity is altered during progression of hypoxic or nonhypoxic PH remains unexplored. The poten-tial effects of p53 activation in target cells may also vary according to cell status or environmental conditions, leading to cell quiescence, senescence, or apoptosis.20–23 Both senes-cence and apoptosis of PA-SMCs may provide therapeutic benefits in PH, although the accumulation of senescent cells within the vessel wall may potentially affect the remodeling process.24 Until now, prosenescence therapy for PH has not been evaluated.
In the present study, we tested the hypothesis that Nutlins hold therapeutic potential in PH via prosenescence effects. To this end, we first examined the effects of Nutlin-3a on cul-tured human PA-SMCs. Second, we investigated the effects of chronic Nutlin-3a treatment in normoxic mice free of PH; mice with hypoxia-induced PH; chronically hypoxic mice simultaneously treated with SU5416 (SU5416/hypoxia)25; and transgenic mice overexpressing the serotonin transporter in SMCs (SM22-5HTT+ mice), which develop spontaneous PH in normoxia.26 Finally, we examined the mechanisms of action of Nutlin-3a by investigating p53 and p21 knockout mice exposed to chronic hypoxia.
Materials and MethodsMiceAdult male mice (C57Bl/6j) were used according to institutional guidelines that complied with national and international regulations. All animal experiments were approved by the Institutional Animal Care and Use Committee of the French National Institute of Health and Medical Research (INSERM)-Unit 955, Créteil, France. Transgenic mice overexpressing 5-HTT in smooth muscle cells (SMCs) under the control of the SM22 promoter (SM22-5HTT+) were produced and bred as previously described.26 These SM22-5HTT+ mice are fertile and have a normal life span and normal growth.5 Mice with deletion of the p53 (p53−/−) or p21 (p21−/−) gene were obtained from Jackson Laboratory (Bar Harbor, ME). Only male mice were used for the ex-periments. Nutlin-3a was administered by intraperitoneal injection in a dosage of 6, 12, or 25 mg/Kg/d. At treatment completion, the lungs were removed and prepared for histological or Western blot analyses.
Exposure to Chronic HypoxiaMale mice aged 15 to 20 weeks were exposed to chronic hypoxia (9% O
2) in a ventilated chamber (Biospherix, New York, NY).27 To
establish the hypoxic environment, the chamber was flushed with a mixture of room air and nitrogen. The chamber was opened every other day for 1 hour to clean the cages, administer drugs, and replenish food and water supplies. Normoxic mice were kept in the same room, with the same light-dark cycle. Mice subjected to SU5416/hypoxia re-ceived an intraperitoneal injection of SU5416 20 mg/Kg once a week during a 3-week period of hypoxia exposure.25
Assessment of Pulmonary HypertensionWe anesthetized mice previously exposed to hypoxia, room air, or SU5416/hypoxia; as well as SM22-5HTT+ mice. After incision of the abdomen, a 26-gauge needle connected to a pressure transducer was inserted into the right ventricle through the diaphragm, and right ven-tricular systolic pressure was recorded immediately. Then, the thorax was opened and the lungs and heart were removed. The right ventricle (RV) was dissected from the left ventricle plus septum (LV+S), and these dissected samples were weighed for determination of Fulton’s index (RV/LV+S). The lungs were fixed by intratracheal infusion of 4% aqueous buffered formalin. A midsagittal slice of the right lung was processed for paraffin embedding. Sections 5 µm in thickness were cut and stained with hematoxylin-phloxine-saffron for examina-tion by light microscopy. In each mouse, a total of 20 to 30 intraacinar vessels accompanying either alveolar ducts or alveoli were examined by an observer who was blinded to the treatment or genotype. Each vessel was categorized as nonmuscular (no evidence of vessel wall muscularization) or muscular (ie, partially muscular [SMCs identifi-able in less than three-fourths of the vessel circumference] or fully muscular [SMCs in more than three-fourths of the vessel circumfer-ence]). The percentage of muscularized pulmonary vessels was deter-mined by dividing the number of partially or fully muscular vessels by the total number of vessels in the relevant group of animals. In addition, medial wall thickness of fully muscularized intraacinar arteries was calculated in SU5416/hypoxia mice and expressed as follows: index (%)=(external diameter − internal diameter)/exter-nal diameter×100. Cell proliferation (Ki67-stained cells), apoptosis (TUNEL-positive cells), and senescence (p21-stained cells) were as-sessed in the walls of distal pulmonary vessels and expressed as the number of stained nuclei over the total number of nuclei counted in the media of at least 20 muscularized vessels per mouse.
EchocardiographyClosed-chest transthoracic echocardiography was performed in non-sedated mice as described in the expanded methods section available in the online-only Data Supplement. Images were acquired using a 13-MHz linear-array transducer with a digital ultrasound system (Vivid 7, GE Medical Systems). All measures were averaged on 5 cardiac cycles, and the reader was blinded to treatment group.
Studies on Cultured Human PA-SMCsCultured PA-SMCs were collected from pulmonary arteries of patients undergoing lung surgery for localized lung tumors.24 To as-sess the effects of Nutlin-3a on PA-SMC proliferation, we exposed PA-SMCs to Nutlin-3a (2.5–10 µmol/L) or vehicle in serum-free me-dium then added platelet-derived growth factor-BB (50 ng/mL). After 48 hours, tetrazolium salt (MTT, Sigma, Lyon, France) was added to each well (0.2 mg/mL). After 4 hours’ incubation at 37°C, the culture medium was removed and formazan crystals were solubilized by add-ing 500 µL of DMSO. Tetrazolium salt reduction to formazan within the cells was quantified by spectrophotometry at 520 nm and taken as an indicator of the number of cells.
To assess the effects of Nutlin-3a on PA-SMC apoptosis, cells were trypsinized and resuspended in binding buffer 1X then incubated with annexin V-FITC–conjugated antibody and stained with propidium iodide according to the manufacturer’s instructions (Sigma-Aldrich, St Louis, MO). Annexin V staining and propidium iodide staining were detected by FACS (Becton Dickinson, Franklin Lakes, NJ). Apoptotic cells were propidium iodide–positive cells and annexin V/propidium iodide–positive cells.
To assess the effects of Nutlin-3a on cell senescence, we deter-mined the percentage of β-galactosidase–positive cells after 48 hours’ incubation with Nutlin-3a with or without vehicle or platelet-derived growth factor.
In addition, cell transfection studies were performed using a p53 Cignal Reporter Assay (Qiagen, ZA Courtaboeuf, France) for assessing p53 signaling in response to Nutlin, as described in the online supplement.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from

1666 Circulation April 23, 2013
Biological Measurements in Mouse Tissues and Cultured Human PA-SMCsWestern blotting was used to detect and quantitate p53, p21, and MDM2 proteins in mouse tissues or human cells as described in the expanded Methods section in the online-only Data Supplement. Complementary experiments were performed in mouse tissue to as-sess phosphorylated p53 levels by using a coimmunoprecipitation procedure or to determine the nuclear and cytoplasmic fractions of total p53 protein. Levels of p21, Bax and PUMA mRNAs in lung tissue and cells were determined using RT-qPCR. Total mRNA was extracted from PA-SMCs using the RNeasy Mini Kit (Qiagen, ZA Courtaboeuf, France). First-strand cDNA was synthesized in reversed transcribed samples, as follows: 1 µg total RNA isolated from cells or lung tissues, 200 U/µL SuperScript II reverse transcriptase, 100 ng Random primers, and 10 mmol/L mixed dNTP (Invitrogen, Life Technologies, Cergy-Pontoise, France). Quantitative PCR was per-formed in a 7900HT Real-Time PCR system (Applied Biosystems, ZA Courtaboeuf, France), using SYBR green Mix from Invitrogen as described in the expanded Methods section in the online-only Data Supplement.
ImmunohistochemistryParaffin-embedded sections were incubated with antibodies against Ki67 or p21 for immunostaining identification of vascular prolifera-tive or senescent cells, respectively. Apoptotic nuclei were labeled with a TUNEL immunostaining assay (Roche, Meylan, France), as described in the expanded Methods section available in the online-only Data Supplement. For immunofluorescence, slides were incu-bated with anti-MDM2 SMP-14 mouse antibody (1:50, Santa Cruz, Santa Cruz, CA) and anti-αSMA rabbit antibody (1:200, Abcam, Cambridge, UK) then exposed to antimouse Alexa Fluor (1:1000, Cell Signaling Technology, Boston, MA) and antirabbit Alexa Fluor (1:1000, Invitrogen, Cergy-Pontoise, France) antibodies. Nuclei were stained with Hoechst 333342 (1 µg/mL, Cell Signaling Technology).
Chemicals and DrugsNutlin-3a was purchased from Bertin Pharma (Montigny-le-Bretonneux, France) and diluted in vehicle (0.9% NaCl and 30% DMSO) to obtain concentrations of 6 mg/Kg, 12 mg/Kg, and 25 mg/Kg. Mice received daily intraperitoneal injections of Nutlin-3a for 21 days. SU5416 was purchased from Sigma (Saint Quentin Fallavier, France).
Statistical AnalysisThe data are described as mean±SEM. Parametric tests were used after verification that the variables in each group were normally dis-tributed. One-way analysis of variance (ANOVA) was performed to compare treatment effects obtained with Nutlin-3a in various doses or at various postdose times. One-way ANOVA was used to com-pare hypoxia-exposed vehicle-treated mice with normoxia-exposed control mice, as well as to compare transgenic mice with wild-type mice during normoxia or hypoxia. P values <0.05 were considered significant for the initial ANOVA, and Bonferroni correction was then used for multiple comparisons (P<0.017).
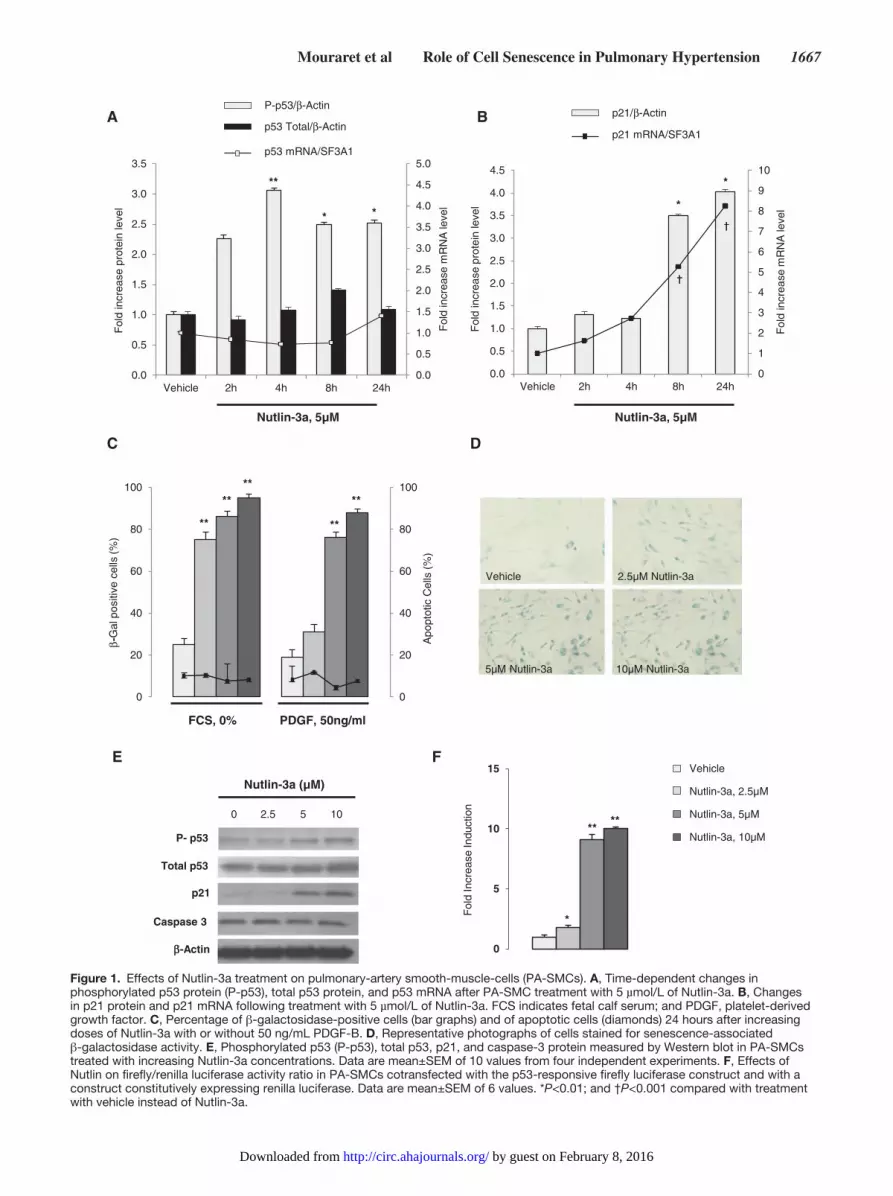
ResultsEffects of Nutlin-3a Treatment on Cultured Human PA-SMCsNutlin-3a treatment of cultured human PA-SMCs was fol-lowed by a marked rise in phosphorylated p53 protein levels within 2 hours after treatment, with a peak after 4 hours and persistent elevation until 24 hours (Figure 1A); total p53 pro-tein and p53 mRNA levels remained unchanged (Figure 1A).
Nutlin-3a treatment also induced marked but delayed increases in p21 mRNA and protein levels, which peaked at 24 hours (Figure 1B) and were accompanied with increased expression of other p53-target genes, including Bax, BTG2, and MDM2 (data not shown). After 24 hours of Nutlin-3a treatment, -galactosidase–stained cells increased in percentage from 20% to 90% and exhibited a typical senescent phenotype char-acterized by a flat shape and increased size, in the absence or presence of platelet-derived growth factor (Figure 1C and 1D). The number of annexin-V–positive cells was not affected by Nutlin-3a (Figure 1C). Nutlin-3a treatment dose-dependently increased the phosphorylated p53 protein and p21 protein lev-els without altering the caspase-3 protein level (Figure 1E), indicating that Nutlin-3a treatment induced senescence but not apoptosis of PA-SMCs. In addition, Nutlin-3a applied to cells transfected with the luciferase p53 reporter led to a marked increase in luciferase activity compared with vehicle-treated control cells (Figure 1F).
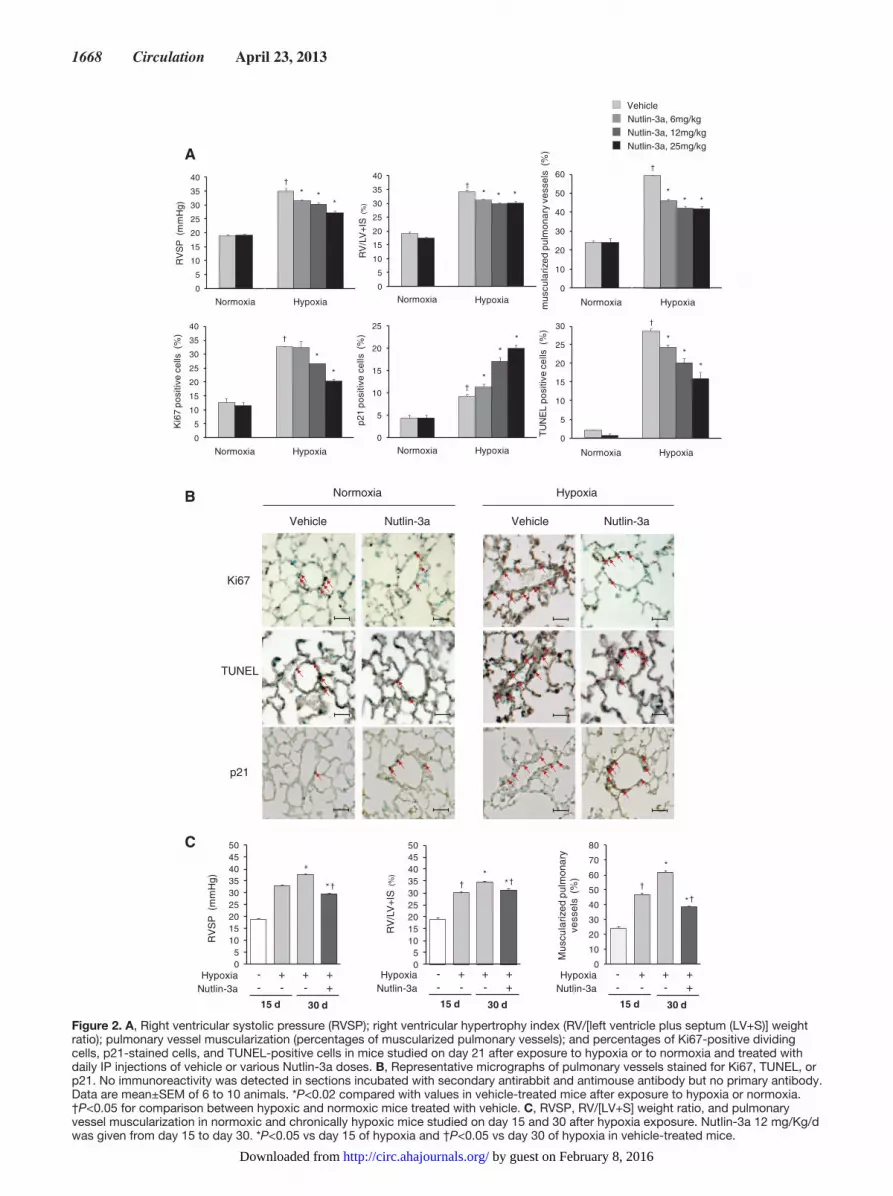
Effects of Treatment With Nutlin-3a in Mice Exposed to Chronic HypoxiaIntraperitoneal treatment of chronically hypoxic mice with 6 to 25 mg/Kg/d of Nutlin-3a attenuated the development of PH, as judged based on right ventricular systolic pressure, RV hypertrophy, and distal pulmonary artery muscularization (Figure 2A). This protective effect of Nutlin-3a was accompanied with simultaneous decreases in PA-SMC proliferation and apoptosis, as assessed by the percentage of Ki67- and TUNEL-positive cells, respectively, contrasting with an increased number of senescent p21-positive PA-SMCs (Figure 2A and 2B). Cardiac output and left ventricular ejection fraction remained unchanged in mice treated with 25 mg/Kg/d of Nutlin-3a, whereas pulmonary acceleration time and the pulmonary acceleration time/ejection time ratio, 2 alternative indices of pulmonary artery pressure, improved in chronically hypoxic mice (Table I in the online-only Data Supplement). Nutlin-3a, 12 mg/Kg/d, from day 15 to day 30 partially reversed PH in chronically hypoxic mice (Figure 2C).
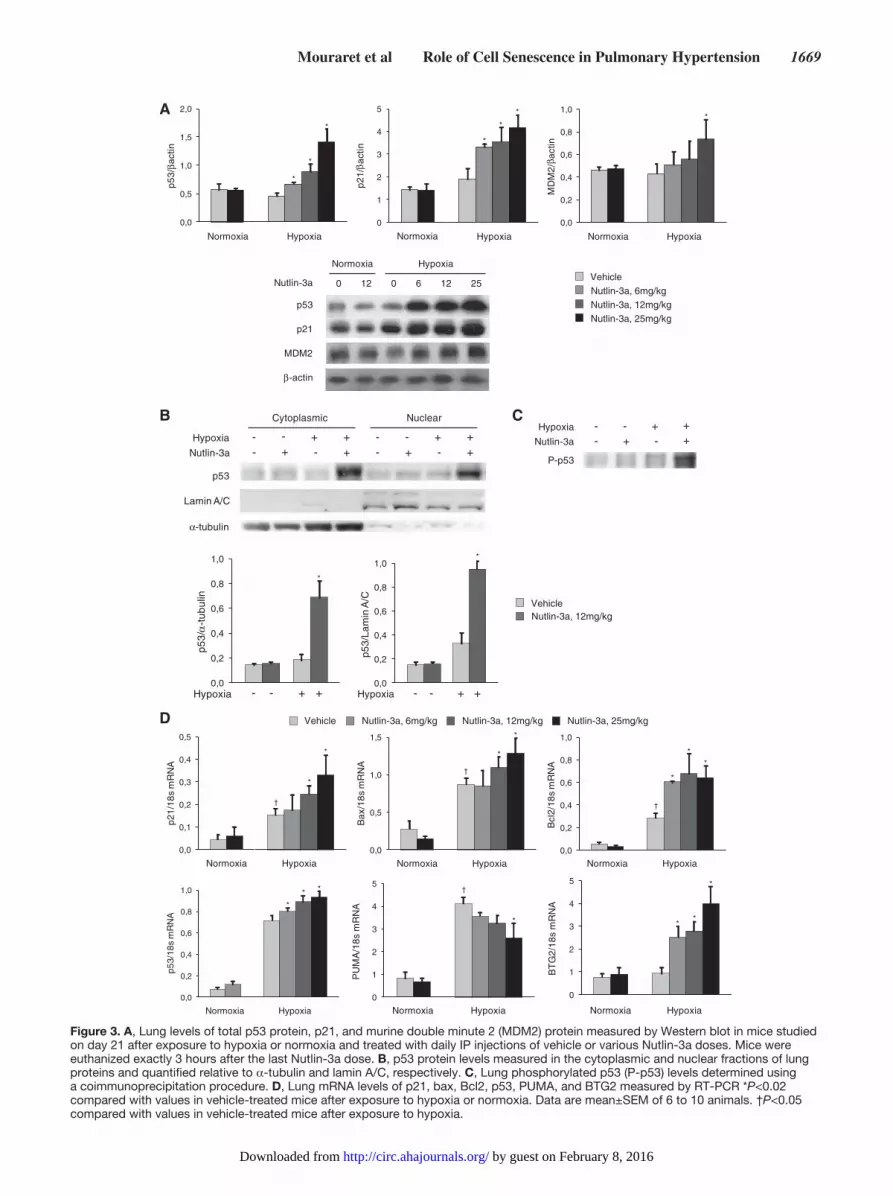
Nutlin-3a treatment markedly increased lung levels of p53, p21, and MDM2 protein and of p21, Bax, Bcl2, and BTG2 mRNAs in chronically hypoxic mice but not in normoxic animals (Figure 3A and 3D). Phosphorylated p53 protein levels measured in whole lung proteins and total p53 protein levels measured in the nuclear and cytoplasmic fractions of lung proteins increased after Nutlin-3a treatment in hypoxic but not normoxic mice (Figure 3B and 3C). Because lung samples were collected exactly 3 hours after the last Nutlin-3a injection in both normoxic and hypoxic mice, the dissimilar response to Nutlin-3a in hypoxic and normoxic mice was interpreted as reflecting increased MDM2 activity during hypoxia. This increased MDM2 activity was measured, although total lung MDM2 protein levels did not change from normoxia to hypoxia (Figure 3A). In contrast to the dose-dependent increase in lung p21, Bax, Bcl2, and BTG2 mRNA levels induced by Nutlin-3a, lung expression of the proapoptotic gene PUMA, which increased from normoxia to hypoxia, was reduced by Nutlin-3a treatment (Figure 3D). The increased p53 protein levels induced by Nutlin-3a also occurred in the kidneys, heart, spleen, and liver from normoxic
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from
Mouraret et al Role of Cell Senescence in Pulmonary Hypertension 1667
0
5
10
15
Fol
dIn
crea
seIn
duct
ion
0
1
2
3
4
5
6
7
8
9
10
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
Vehicle 2h 4h 8h 24h
Fol
d in
crea
se m
RN
A le
vel
Fol
d in
crea
se p
rote
in le
vel
A
Nutlin-3a, 5µM
0
20
40
60
80
100
0
20
40
60
80
100
Apo
ptot
ic C
ells
(%
)
-Gal
pos
itive
cel
ls (%
)β
PDGF, 50ng/mlFCS, 0%
E
Vehicle 2.5µM Nutlin-3a
5µM Nutlin-3a 10µM Nutlin-3a
F
Nutlin-3a, 5µM
p21/ -Actin
p21 mRNA/SF3A1
C
0 2.5 5 10
Nutlin-3a (µM)
Total p53
-Actinβ
P- p53
p21
Caspase 3
D
Vehicle
Nutlin-3a, 2.5µM
Nutlin-3a, 10µM
Nutlin-3a, 5µM
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Vehicle 2h 4h 8h 24h
Fol
d in
crea
se m
RN
A le
vel
Fol
d in
crea
se p
rote
in le
vel
P-p53/ -Actinββ
βp53 Total/ -Actin
p53 mRNA/SF3A1
**
* **
*
†
†
**
****
**
**
****
*
B
Figure 1. Effects of Nutlin-3a treatment on pulmonary-artery smooth-muscle-cells (PA-SMCs). A, Time-dependent changes in phosphorylated p53 protein (P-p53), total p53 protein, and p53 mRNA after PA-SMC treatment with 5 µmol/L of Nutlin-3a. B, Changes in p21 protein and p21 mRNA following treatment with 5 µmol/L of Nutlin-3a. FCS indicates fetal calf serum; and PDGF, platelet-derived growth factor. C, Percentage of β-galactosidase-positive cells (bar graphs) and of apoptotic cells (diamonds) 24 hours after increasing doses of Nutlin-3a with or without 50 ng/mL PDGF-B. D, Representative photographs of cells stained for senescence-associated β-galactosidase activity. E, Phosphorylated p53 (P-p53), total p53, p21, and caspase-3 protein measured by Western blot in PA-SMCs treated with increasing Nutlin-3a concentrations. Data are mean±SEM of 10 values from four independent experiments. F, Effects of Nutlin on firefly/renilla luciferase activity ratio in PA-SMCs cotransfected with the p53-responsive firefly luciferase construct and with a construct constitutively expressing renilla luciferase. Data are mean±SEM of 6 values. *P<0.01; and †P<0.001 compared with treatment with vehicle instead of Nutlin-3a.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from
1668 Circulation April 23, 2013
0
10
20
30
40
50
60
*
mus
cula
rized
pulm
onar
y ve
ssel
s (
%)
**
Normoxia Hypoxia
†
0
5
10
15
20
25
Normoxia Hypoxia
p21
posi
tive
cells
(%
)
*
**
†
0
5
10
15
20
25
30
35
40
Normoxia Hypoxia
Ki6
7 po
sitiv
e ce
lls (
%)
*
*
†
0
5
10
15
20
25
30
35
40
Normoxia Hypoxia
*R
VS
P (
mm
Hg)
**
†
0
5
10
15
20
25
30
35
40
Normoxia Hypoxia
RV
/LV
+IS
(%)
* * *†
Normoxia Hypoxia
Ki67
TUNEL
Vehicle VehicleNutlin-3a Nutlin-3a
p21
VehicleNutlin-3a, 6mg/kgNutlin-3a, 12mg/kgNutlin-3a, 25mg/kg
A
B
TU
NE
L po
sitiv
e ce
lls (
%)
0
5
10
15
20
25
30
Normoxia Hypoxia
*
*
*
†
05
101520253035404550
†*
RV
/LV
+IS
(%)
Nutlin-3aHypoxia - + + +
- - - +15 d 30 d
* †
05
101520253035404550
Nutlin-3aHypoxia
RV
SP
(m
mH
g)
- + + +- - - +15 d 30 d
*†
*
0
10
20
30
40
50
60
70
80
†
*
Mus
cula
rized
pulm
onar
y ve
ssel
s (
%)
Nutlin-3aHypoxia - + + +
- - - +15 d 30 d
*†
C
Figure 2. A, Right ventricular systolic pressure (RVSP); right ventricular hypertrophy index (RV/[left ventricle plus septum (LV+S)] weight ratio); pulmonary vessel muscularization (percentages of muscularized pulmonary vessels); and percentages of Ki67-positive dividing cells, p21-stained cells, and TUNEL-positive cells in mice studied on day 21 after exposure to hypoxia or to normoxia and treated with daily IP injections of vehicle or various Nutlin-3a doses. B, Representative micrographs of pulmonary vessels stained for Ki67, TUNEL, or p21. No immunoreactivity was detected in sections incubated with secondary antirabbit and antimouse antibody but no primary antibody. Data are mean±SEM of 6 to 10 animals. *P<0.02 compared with values in vehicle-treated mice after exposure to hypoxia or normoxia. †P<0.05 for comparison between hypoxic and normoxic mice treated with vehicle. C, RVSP, RV/[LV+S] weight ratio, and pulmonary vessel muscularization in normoxic and chronically hypoxic mice studied on day 15 and 30 after hypoxia exposure. Nutlin-3a 12 mg/Kg/d was given from day 15 to day 30. *P<0.05 vs day 15 of hypoxia and †P<0.05 vs day 30 of hypoxia in vehicle-treated mice.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from
Mouraret et al Role of Cell Senescence in Pulmonary Hypertension 1669
P-p53
Nutlin-3a
Hypoxia - - + +- + - +
p53
Lamin A/C
Nutlin-3a
Hypoxia
Cytoplasmic Nuclear
- - + +- + - +
- - +- + -
++
-tubulinα
Nutlin-3a
Normoxia
0 12 0 6 12 25
p53
p21
MDM2
-β actin
Hypoxia
0,0
0,2
0,4
0,6
0,8
1,0
MD
M2/
actin
Normoxia Hypoxia
0
1
2
3
4
5
p21/
actin
Normoxia Hypoxia
0,0
0,5
1,0
1,5
2,0
p53/
actin
β β β
Normoxia Hypoxia
Vehicle
Nutlin-3a, 6mg/kgNutlin-3a, 12mg/kg
Nutlin-3a, 25mg/kg
A
B C
VehicleNutlin-3a, 12mg/kg
0,0
0,2
0,4
0,6
0,8
1,0
p53/
-tub
ulin
α
Hypoxia - - + +0,0
0,2
0,4
0,6
0,8
1,0
- - + +
p53/
Lam
inA
/C
Hypoxia
Vehicle Nutlin-3a, 6mg/kg Nutlin-3a, 12mg/kg Nutlin-3a, 25mg/kg
0
1
2
3
4
5
PU
MA
/18s
mR
NA
Normoxia Hypoxia
†
0,0
0,2
0,4
0,6
0,8
1,0
Bcl
2/18
s m
RN
A
Normoxia Hypoxia
†
0,0
0,5
1,0
1,5
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
***
**
*
Bax
/18s
mR
NA
Normoxia Hypoxia
†
0,0
0,1
0,2
0,3
0,4
0,5
p21/
18s
mR
NA
Normoxia Hypoxia
†
0
1
2
3
4
5
BT
G2/
18s
mR
NA
Normoxia Hypoxia0,0
0,2
0,4
0,6
0,8
1,0
p53/
18s
mR
NA
Normoxia Hypoxia
D
Figure 3. A, Lung levels of total p53 protein, p21, and murine double minute 2 (MDM2) protein measured by Western blot in mice studied on day 21 after exposure to hypoxia or normoxia and treated with daily IP injections of vehicle or various Nutlin-3a doses. Mice were euthanized exactly 3 hours after the last Nutlin-3a dose. B, p53 protein levels measured in the cytoplasmic and nuclear fractions of lung proteins and quantified relative to α-tubulin and lamin A/C, respectively. C, Lung phosphorylated p53 (P-p53) levels determined using a coimmunoprecipitation procedure. D, Lung mRNA levels of p21, bax, Bcl2, p53, PUMA, and BTG2 measured by RT-PCR *P<0.02 compared with values in vehicle-treated mice after exposure to hypoxia or normoxia. Data are mean±SEM of 6 to 10 animals. †P<0.05 compared with values in vehicle-treated mice after exposure to hypoxia.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from
1670 Circulation April 23, 2013
and hypoxic mice, although at different levels (online Figure 1). A marked (3.5-fold) increase in p53 protein was observed only in the lungs and kidneys of chronically hypoxic mice.
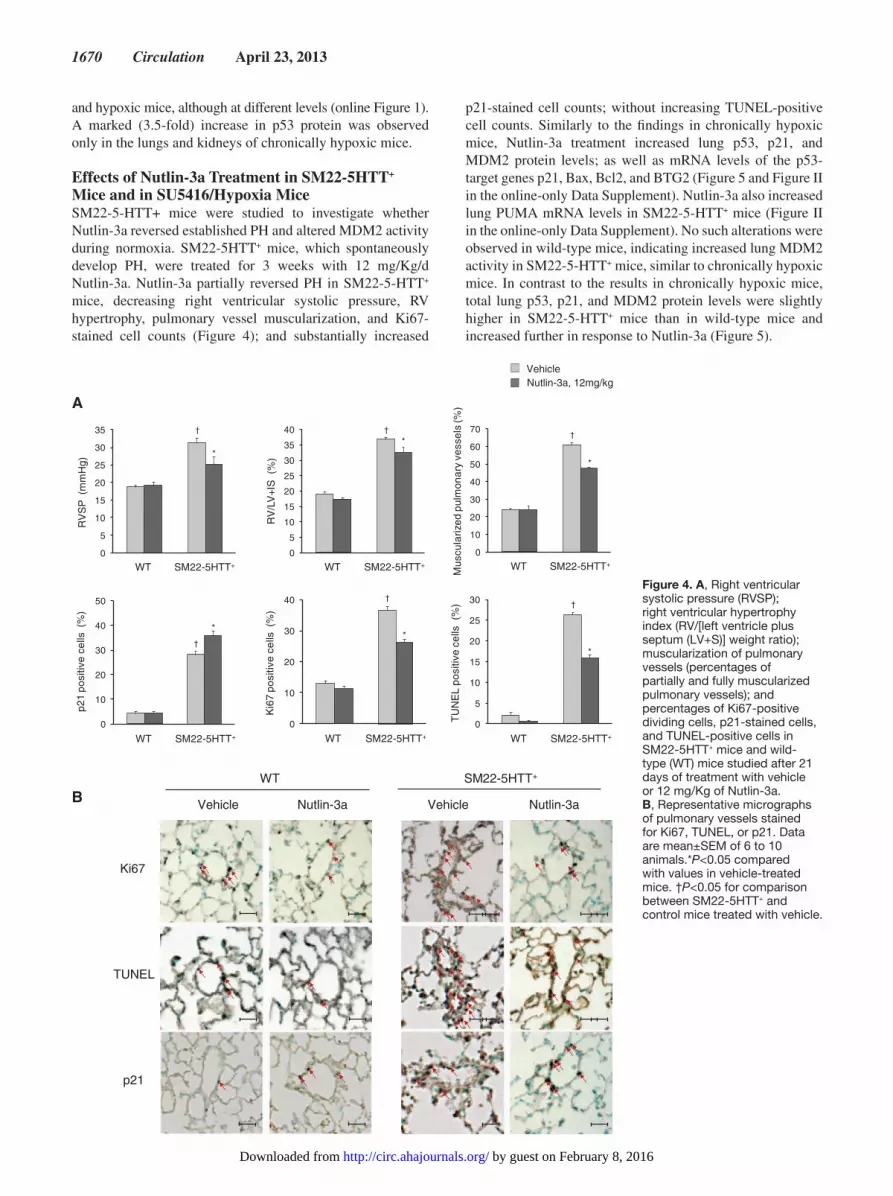
Effects of Nutlin-3a Treatment in SM22-5HTT+ Mice and in SU5416/Hypoxia MiceSM22-5-HTT+ mice were studied to investigate whether Nutlin-3a reversed established PH and altered MDM2 activity during normoxia. SM22-5HTT+ mice, which spontaneously develop PH, were treated for 3 weeks with 12 mg/Kg/d Nutlin-3a. Nutlin-3a partially reversed PH in SM22-5-HTT+ mice, decreasing right ventricular systolic pressure, RV hypertrophy, pulmonary vessel muscularization, and Ki67-stained cell counts (Figure 4); and substantially increased
p21-stained cell counts; without increasing TUNEL-positive cell counts. Similarly to the findings in chronically hypoxic mice, Nutlin-3a treatment increased lung p53, p21, and MDM2 protein levels; as well as mRNA levels of the p53-target genes p21, Bax, Bcl2, and BTG2 (Figure 5 and Figure II in the online-only Data Supplement). Nutlin-3a also increased lung PUMA mRNA levels in SM22-5-HTT+ mice (Figure II in the online-only Data Supplement). No such alterations were observed in wild-type mice, indicating increased lung MDM2 activity in SM22-5-HTT+ mice, similar to chronically hypoxic mice. In contrast to the results in chronically hypoxic mice, total lung p53, p21, and MDM2 protein levels were slightly higher in SM22-5-HTT+ mice than in wild-type mice and increased further in response to Nutlin-3a (Figure 5).
A
0
10
20
30
40
50
60
70
Mus
cula
rized
pulm
onar
y ve
ssel
s (%
)
WT SM22-5HTT+
*
†
0
10
20
30
40
WT SM22-5HTT+
Ki6
7 po
sitiv
e ce
lls(%
)
*
†
0
10
20
30
40
50
*
p21
posi
tive
cells
(%)
WT SM22-5HTT+
†
0
5
10
15
20
25
30
35
40
WT SM22-5HTT+
RV
/LV
+IS
(%
)
*†
0
5
10
15
20
25
30
35
RV
SP
(m
mH
g)
WT SM22-5HTT+
*
†
WT SM22-5HTT+
Ki67
TUNEL
Vehicle VehicleNutlin-3a Nutlin-3a
p21
VehicleNutlin-3a, 12mg/kg
B
0
5
10
15
20
25
30
*
†
TU
NE
L po
sitiv
e ce
lls(%
)
WT SM22-5HTT+
Figure 4. A, Right ventricular systolic pressure (RVSP); right ventricular hypertrophy index (RV/[left ventricle plus septum (LV+S)] weight ratio); muscularization of pulmonary vessels (percentages of partially and fully muscularized pulmonary vessels); and percentages of Ki67-positive dividing cells, p21-stained cells, and TUNEL-positive cells in SM22-5HTT+ mice and wild-type (WT) mice studied after 21 days of treatment with vehicle or 12 mg/Kg of Nutlin-3a. B, Representative micrographs of pulmonary vessels stained for Ki67, TUNEL, or p21. Data are mean±SEM of 6 to 10 animals.*P<0.05 compared with values in vehicle-treated mice. †P<0.05 for comparison between SM22-5HTT+ and control mice treated with vehicle.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from
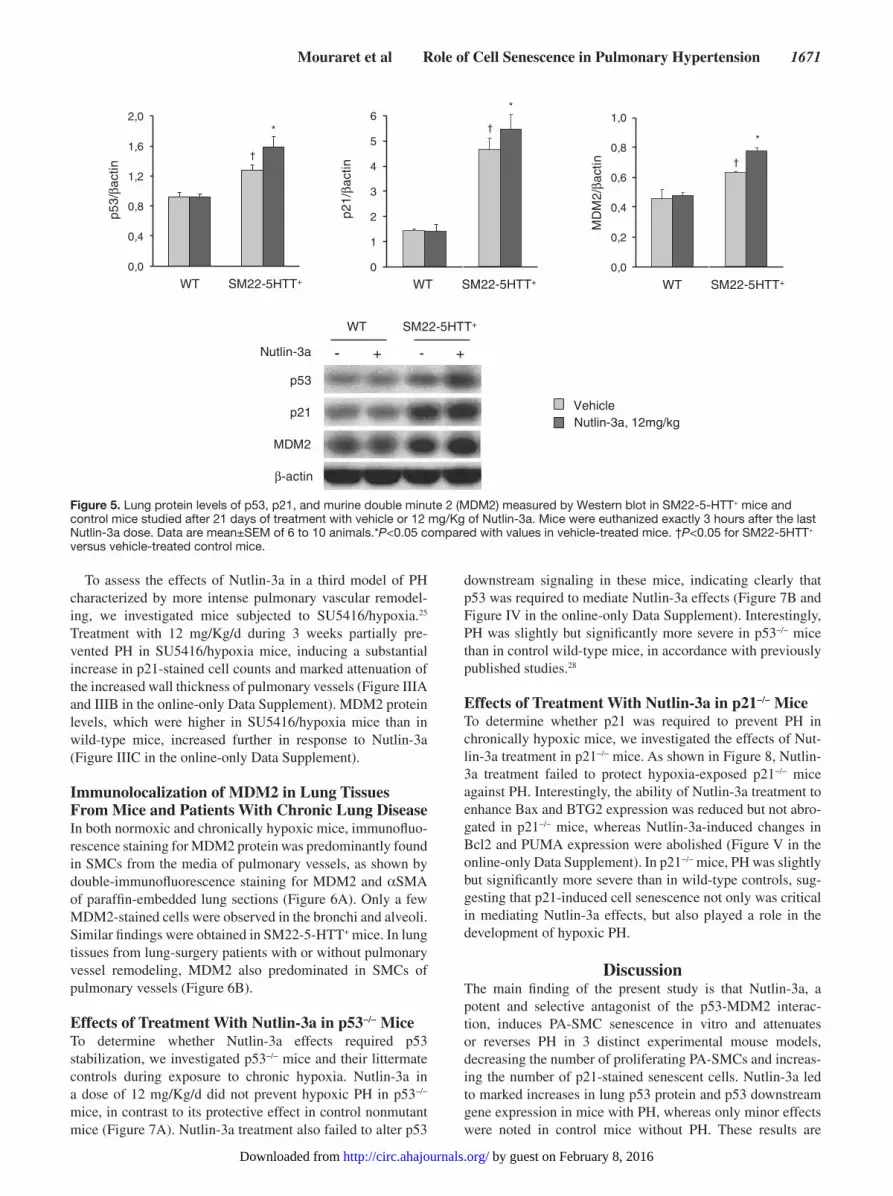
Mouraret et al Role of Cell Senescence in Pulmonary Hypertension 1671
To assess the effects of Nutlin-3a in a third model of PH characterized by more intense pulmonary vascular remodel-ing, we investigated mice subjected to SU5416/hypoxia.25 Treatment with 12 mg/Kg/d during 3 weeks partially pre-vented PH in SU5416/hypoxia mice, inducing a substantial increase in p21-stained cell counts and marked attenuation of the increased wall thickness of pulmonary vessels (Figure IIIA and IIIB in the online-only Data Supplement). MDM2 protein levels, which were higher in SU5416/hypoxia mice than in wild-type mice, increased further in response to Nutlin-3a (Figure IIIC in the online-only Data Supplement).
Immunolocalization of MDM2 in Lung Tissues From Mice and Patients With Chronic Lung DiseaseIn both normoxic and chronically hypoxic mice, immunofluo-rescence staining for MDM2 protein was predominantly found in SMCs from the media of pulmonary vessels, as shown by double-immunofluorescence staining for MDM2 and αSMA of paraffin-embedded lung sections (Figure 6A). Only a few MDM2-stained cells were observed in the bronchi and alveoli. Similar findings were obtained in SM22-5-HTT+ mice. In lung tissues from lung-surgery patients with or without pulmonary vessel remodeling, MDM2 also predominated in SMCs of pulmonary vessels (Figure 6B).
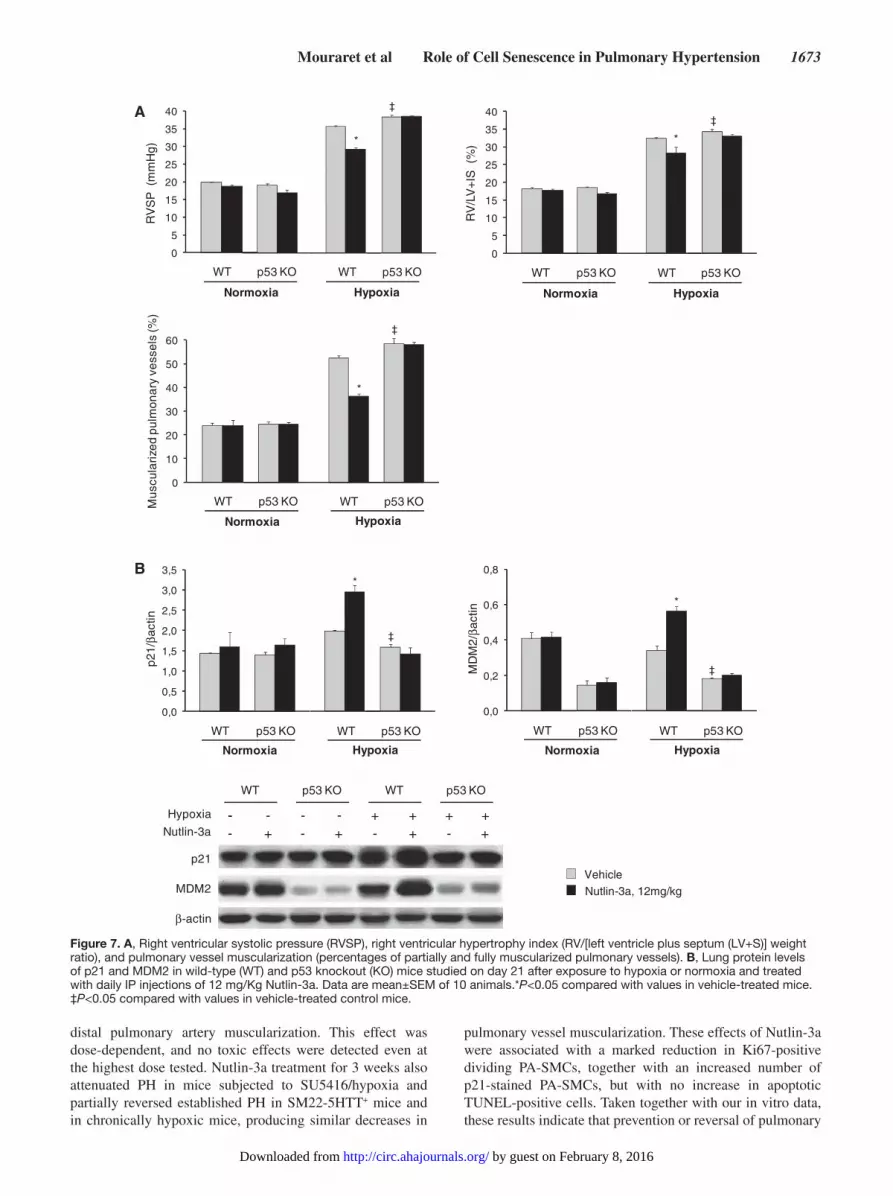
Effects of Treatment With Nutlin-3a in p53−/− MiceTo determine whether Nutlin-3a effects required p53 stabilization, we investigated p53−/− mice and their littermate controls during exposure to chronic hypoxia. Nutlin-3a in a dose of 12 mg/Kg/d did not prevent hypoxic PH in p53−/− mice, in contrast to its protective effect in control nonmutant mice (Figure 7A). Nutlin-3a treatment also failed to alter p53
downstream signaling in these mice, indicating clearly that p53 was required to mediate Nutlin-3a effects (Figure 7B and Figure IV in the online-only Data Supplement). Interestingly, PH was slightly but significantly more severe in p53−/− mice than in control wild-type mice, in accordance with previously published studies.28
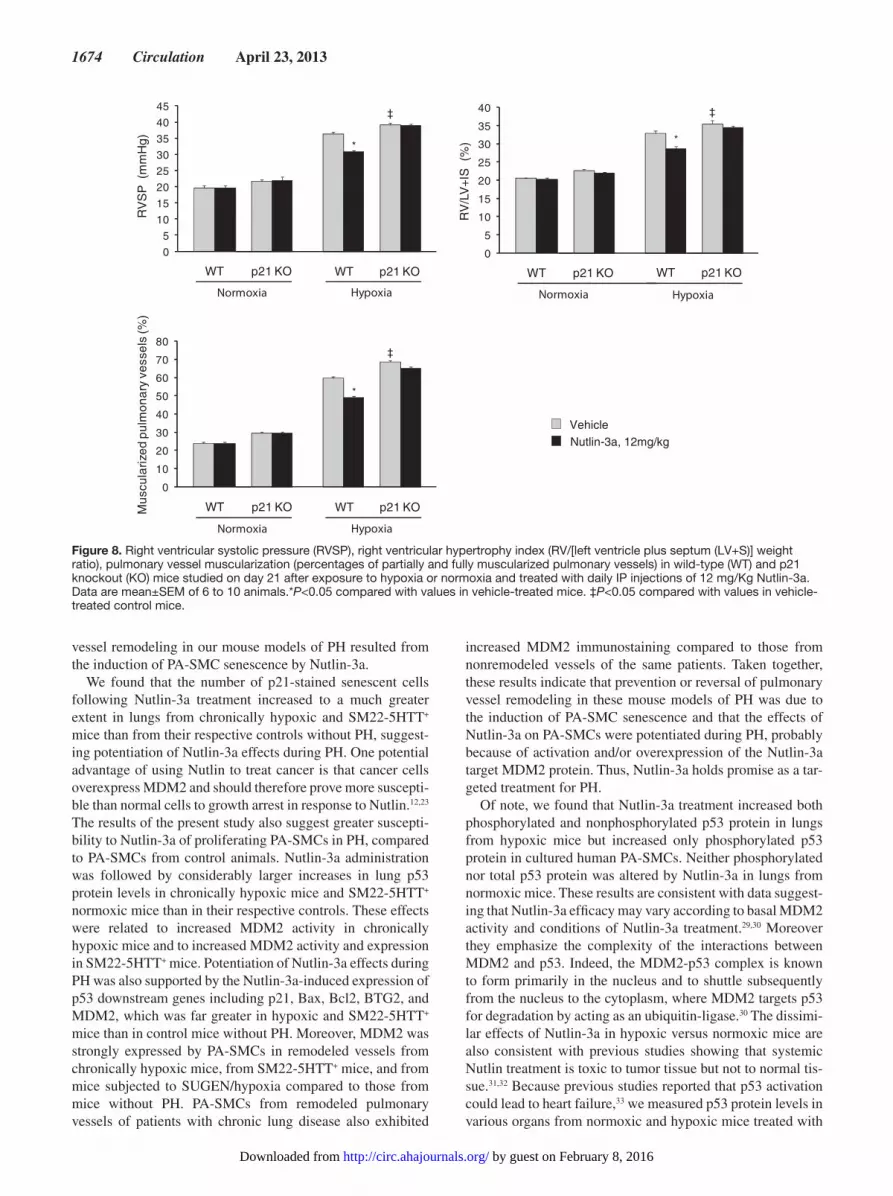
Effects of Treatment With Nutlin-3a in p21−/− MiceTo determine whether p21 was required to prevent PH in chronically hypoxic mice, we investigated the effects of Nut-lin-3a treatment in p21−/− mice. As shown in Figure 8, Nutlin-3a treatment failed to protect hypoxia-exposed p21−/− mice against PH. Interestingly, the ability of Nutlin-3a treatment to enhance Bax and BTG2 expression was reduced but not abro-gated in p21−/− mice, whereas Nutlin-3a-induced changes in Bcl2 and PUMA expression were abolished (Figure V in the online-only Data Supplement). In p21−/− mice, PH was slightly but significantly more severe than in wild-type controls, sug-gesting that p21-induced cell senescence not only was critical in mediating Nutlin-3a effects, but also played a role in the development of hypoxic PH.
DiscussionThe main finding of the present study is that Nutlin-3a, a potent and selective antagonist of the p53-MDM2 interac-tion, induces PA-SMC senescence in vitro and attenuates or reverses PH in 3 distinct experimental mouse models, decreasing the number of proliferating PA-SMCs and increas-ing the number of p21-stained senescent cells. Nutlin-3a led to marked increases in lung p53 protein and p53 downstream gene expression in mice with PH, whereas only minor effects were noted in control mice without PH. These results are
p53
p21
MDM2
β-actin
Nutlin-3a
WT
-
SM22-5HTT+
- + - +
0,0
0,2
0,4
0,6
0,8
1,0
MD
M2/
βact
in
WT SM22-5HTT+
*
†
0
1
2
3
4
5
6
WT SM22-5HTT+
p21/
βact
in
*
†
0,0
0,4
0,8
1,2
1,6
2,0p5
3/βa
ctin
WT SM22-5HTT+
*
†
VehicleNutlin-3a, 12mg/kg
Figure 5. Lung protein levels of p53, p21, and murine double minute 2 (MDM2) measured by Western blot in SM22-5-HTT+ mice and control mice studied after 21 days of treatment with vehicle or 12 mg/Kg of Nutlin-3a. Mice were euthanized exactly 3 hours after the last Nutlin-3a dose. Data are mean±SEM of 6 to 10 animals.*P<0.05 compared with values in vehicle-treated mice. †P<0.05 for SM22-5HTT+ versus vehicle-treated control mice.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from
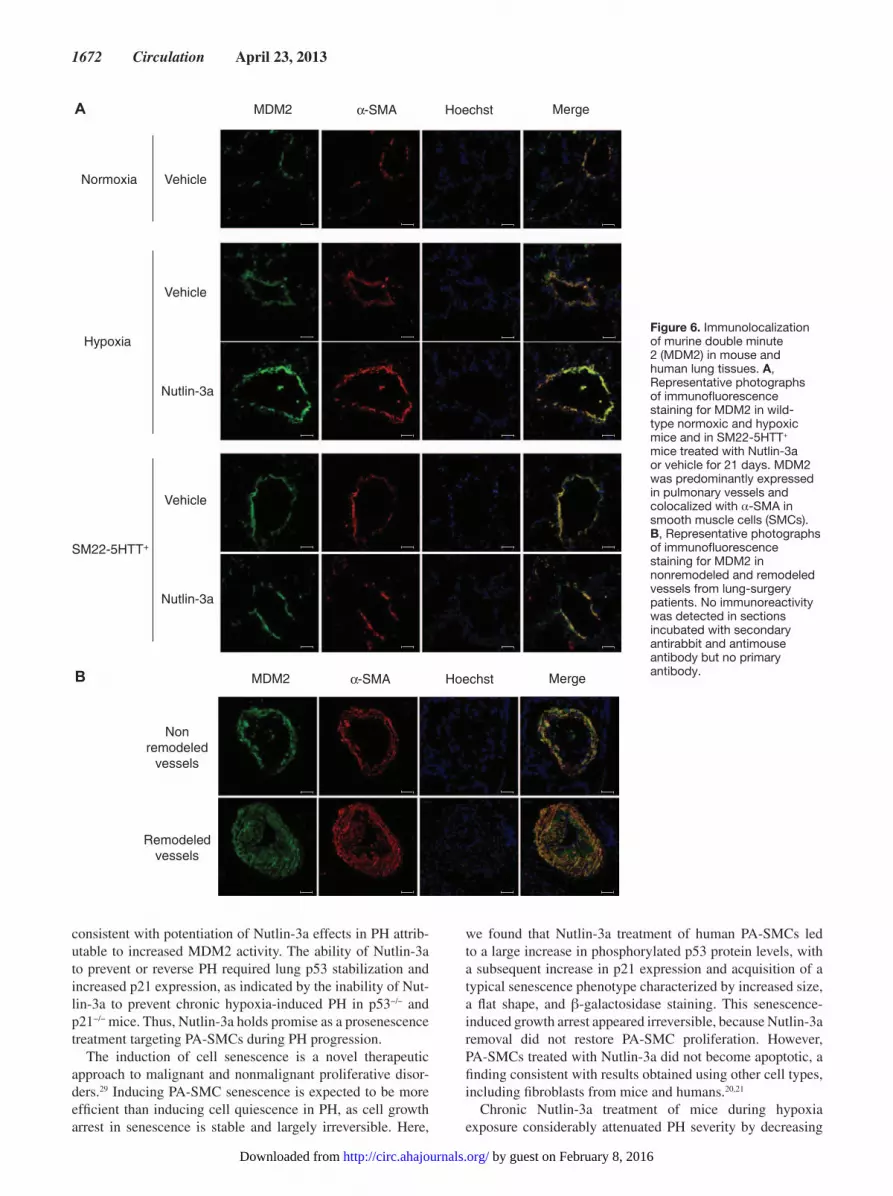
1672 Circulation April 23, 2013
consistent with potentiation of Nutlin-3a effects in PH attrib-utable to increased MDM2 activity. The ability of Nutlin-3a to prevent or reverse PH required lung p53 stabilization and increased p21 expression, as indicated by the inability of Nut-lin-3a to prevent chronic hypoxia-induced PH in p53−/− and p21−/− mice. Thus, Nutlin-3a holds promise as a prosenescence treatment targeting PA-SMCs during PH progression.
The induction of cell senescence is a novel therapeutic approach to malignant and nonmalignant proliferative disor-ders.29 Inducing PA-SMC senescence is expected to be more efficient than inducing cell quiescence in PH, as cell growth arrest in senescence is stable and largely irreversible. Here,
we found that Nutlin-3a treatment of human PA-SMCs led to a large increase in phosphorylated p53 protein levels, with a subsequent increase in p21 expression and acquisition of a typical senescence phenotype characterized by increased size, a flat shape, and β-galactosidase staining. This senescence-induced growth arrest appeared irreversible, because Nutlin-3a removal did not restore PA-SMC proliferation. However, PA-SMCs treated with Nutlin-3a did not become apoptotic, a finding consistent with results obtained using other cell types, including fibroblasts from mice and humans.20,21
Chronic Nutlin-3a treatment of mice during hypoxia exposure considerably attenuated PH severity by decreasing
MDM2 α-SMA Hoechst Merge
Vehicle
Vehicle
Nutlin-3a
Normoxia
Hypoxia
Vehicle
Nutlin-3a
SM22-5HTT+
A
B MDM2 α-SMA Hoechst Merge
Non remodeled
vessels
Remodeledvessels
Figure 6. Immunolocalization of murine double minute 2 (MDM2) in mouse and human lung tissues. A, Representative photographs of immunofluorescence staining for MDM2 in wild-type normoxic and hypoxic mice and in SM22-5HTT+ mice treated with Nutlin-3a or vehicle for 21 days. MDM2 was predominantly expressed in pulmonary vessels and colocalized with α-SMA in smooth muscle cells (SMCs). B, Representative photographs of immunofluorescence staining for MDM2 in nonremodeled and remodeled vessels from lung-surgery patients. No immunoreactivity was detected in sections incubated with secondary antirabbit and antimouse antibody but no primary antibody.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from
Mouraret et al Role of Cell Senescence in Pulmonary Hypertension 1673
distal pulmonary artery muscularization. This effect was dose-dependent, and no toxic effects were detected even at the highest dose tested. Nutlin-3a treatment for 3 weeks also attenuated PH in mice subjected to SU5416/hypoxia and partially reversed established PH in SM22-5HTT+ mice and in chronically hypoxic mice, producing similar decreases in
pulmonary vessel muscularization. These effects of Nutlin-3a were associated with a marked reduction in Ki67-positive dividing PA-SMCs, together with an increased number of p21-stained PA-SMCs, but with no increase in apoptotic TUNEL-positive cells. Taken together with our in vitro data, these results indicate that prevention or reversal of pulmonary
0
10
20
30
40
50
60
Mu
scul
ariz
edpu
lmon
ary
vess
els
(%)
WT p53 KO WT p53 KO
Normoxia Hypoxia
*
‡
0
5
10
15
20
25
30
35
40‡
*
RV
/LV
+IS
(%
)
WT p53 KO WT p53 KO
Normoxia Hypoxia
*
‡
0
5
10
15
20
25
30
35
40
WT p53 KO WT p53 KO
Normoxia Hypoxia
RV
SP
(m
mH
g)
0,0
0,2
0,4
0,6
0,8
MD
M2/
βact
in
WT p53 KO WT p53 KO
Normoxia Hypoxia
*
‡
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
p21/
βact
in
WT p53 KO WT p53 KO
Normoxia Hypoxia
*
‡
Nutlin-3a
p21
Hypoxia
WT
-- +- - -
- +- +
- ++
- ++ +
p53 KO WT p53 KO
MDM2
β-actin
VehicleNutlin-3a, 12mg/kg
B
A
Figure 7. A, Right ventricular systolic pressure (RVSP), right ventricular hypertrophy index (RV/[left ventricle plus septum (LV+S)] weight ratio), and pulmonary vessel muscularization (percentages of partially and fully muscularized pulmonary vessels). B, Lung protein levels of p21 and MDM2 in wild-type (WT) and p53 knockout (KO) mice studied on day 21 after exposure to hypoxia or normoxia and treated with daily IP injections of 12 mg/Kg Nutlin-3a. Data are mean±SEM of 10 animals.*P<0.05 compared with values in vehicle-treated mice. ‡P<0.05 compared with values in vehicle-treated control mice.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from
1674 Circulation April 23, 2013
vessel remodeling in our mouse models of PH resulted from the induction of PA-SMC senescence by Nutlin-3a.
We found that the number of p21-stained senescent cells following Nutlin-3a treatment increased to a much greater extent in lungs from chronically hypoxic and SM22-5HTT+ mice than from their respective controls without PH, suggest-ing potentiation of Nutlin-3a effects during PH. One potential advantage of using Nutlin to treat cancer is that cancer cells overexpress MDM2 and should therefore prove more suscepti-ble than normal cells to growth arrest in response to Nutlin.12,23 The results of the present study also suggest greater suscepti-bility to Nutlin-3a of proliferating PA-SMCs in PH, compared to PA-SMCs from control animals. Nutlin-3a administration was followed by considerably larger increases in lung p53 protein levels in chronically hypoxic mice and SM22-5HTT+ normoxic mice than in their respective controls. These effects were related to increased MDM2 activity in chronically hypoxic mice and to increased MDM2 activity and expression in SM22-5HTT+ mice. Potentiation of Nutlin-3a effects during PH was also supported by the Nutlin-3a-induced expression of p53 downstream genes including p21, Bax, Bcl2, BTG2, and MDM2, which was far greater in hypoxic and SM22-5HTT+ mice than in control mice without PH. Moreover, MDM2 was strongly expressed by PA-SMCs in remodeled vessels from chronically hypoxic mice, from SM22-5HTT+ mice, and from mice subjected to SUGEN/hypoxia compared to those from mice without PH. PA-SMCs from remodeled pulmonary vessels of patients with chronic lung disease also exhibited
increased MDM2 immunostaining compared to those from nonremodeled vessels of the same patients. Taken together, these results indicate that prevention or reversal of pulmonary vessel remodeling in these mouse models of PH was due to the induction of PA-SMC senescence and that the effects of Nutlin-3a on PA-SMCs were potentiated during PH, probably because of activation and/or overexpression of the Nutlin-3a target MDM2 protein. Thus, Nutlin-3a holds promise as a tar-geted treatment for PH.
Of note, we found that Nutlin-3a treatment increased both phosphorylated and nonphosphorylated p53 protein in lungs from hypoxic mice but increased only phosphorylated p53 protein in cultured human PA-SMCs. Neither phosphorylated nor total p53 protein was altered by Nutlin-3a in lungs from normoxic mice. These results are consistent with data suggest-ing that Nutlin-3a efficacy may vary according to basal MDM2 activity and conditions of Nutlin-3a treatment.29,30 Moreover they emphasize the complexity of the interactions between MDM2 and p53. Indeed, the MDM2-p53 complex is known to form primarily in the nucleus and to shuttle subsequently from the nucleus to the cytoplasm, where MDM2 targets p53 for degradation by acting as an ubiquitin-ligase.30 The dissimi-lar effects of Nutlin-3a in hypoxic versus normoxic mice are also consistent with previous studies showing that systemic Nutlin treatment is toxic to tumor tissue but not to normal tis-sue.31,32 Because previous studies reported that p53 activation could lead to heart failure,33 we measured p53 protein levels in various organs from normoxic and hypoxic mice treated with
VehicleNutlin-3a, 12mg/kg
0
10
20
30
40
50
60
70
80
Mus
cula
rized
pulm
onar
y ve
ssel
s (%
)
*
WT p21 KO WT p21 KO
Normoxia Hypoxia
‡
0
5
10
15
20
25
30
35
40
*
RV
/LV
+IS
(%
)
WT p21 KO WT p21 KO
Normoxia Hypoxia
‡
0
5
10
15
20
25
30
35
40
45
*R
VS
P (
mm
Hg)
WT p21 KO WT p21 KO
Normoxia Hypoxia
‡
Figure 8. Right ventricular systolic pressure (RVSP), right ventricular hypertrophy index (RV/[left ventricle plus septum (LV+S)] weight ratio), pulmonary vessel muscularization (percentages of partially and fully muscularized pulmonary vessels) in wild-type (WT) and p21 knockout (KO) mice studied on day 21 after exposure to hypoxia or normoxia and treated with daily IP injections of 12 mg/Kg Nutlin-3a. Data are mean±SEM of 6 to 10 animals.*P<0.05 compared with values in vehicle-treated mice. ‡P<0.05 compared with values in vehicle-treated control mice.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from

Mouraret et al Role of Cell Senescence in Pulmonary Hypertension 1675
Nutlin-3a. We observed only minimal changes in p53 protein levels in the heart, spleen, and liver, and found that cardiac function remained unaltered. Major increases in p53 protein were seen only in the lungs and in the kidneys. Further stud-ies are therefore needed to assess the renal effects of Nutlins in conditions associated with PH, although Nutlins have been reported to improve kidney function in other conditions.34
Although p53 regulation appears to be the central focus of MDM2 activity, the p53-binding domain of MDM2 binds to other proteins that are potentially involved in cell growth and survival.35 To investigate whether Nutlin-3a effects were mediated solely by stabilization and overexpression of p53, we investigated p53−/− mice. Nutlin-3a did not prevent PH in p53−/− mice simultaneously exposed to chronic hypoxia. Neither did Nutlin-3a alter p53 downstream signaling in these mice, clearly indicating that p53 was required to mediate the effects of Nutlin-3a. Another point of interest was whether p53 played a pivotal role in PH development in our mouse models. As previously reported, we found that PH was more severe in p53−/− mice than in control wild-type mice.28 However, in contrast to these previous studies showing marked increases in lung p53 protein levels from normoxia to hypoxia,28 we observed only minimal alterations in baseline p53 levels in hypoxic versus normoxic mice and in SM22-5HTT+ mice ver-sus control wild-type mice. The reason for this difference is unclear but probably involves differences in hypoxia exposure conditions. The relative stability of p53 protein levels from normoxia to hypoxia in our study is probably ascribable to increased p53 protein degradation attributable to increased MDM2 ubiquitin ligase activity. Suppression of this adap-tive response by Nutlin-3a led to major increases in p53 in PA-SMCs, thereby strongly inhibiting the pulmonary vascular remodeling process.
Among the transcriptional targets of p53, p21 has been shown to play a major role as a regulator of cell-cycle pro-gression and to be involved in cell senescence.36 We found that the protective action of Nutlin-3a against PH required p21, as indicated by the inability of Nutlin-3a to prevent chronic hypoxia-induced PH in p21−/− mice, despite signifi-cant increases in mRNA levels of the antiproliferative protein BTG2 and of the proapoptotic protein Bax. These results indi-cate a prominent role for p21 in our experimental PH model and support cell-senescence induction as the mechanism by which Nutlin-3a protects against PH. Moreover, the slight increase in PH severity noted in p21−/− mice compared with their hypoxic controls suggests involvement of p21 in the con-trol of hypoxic PH progression.
The present results supporting a beneficial effect of cell-senescence induction in PH may appear to contradict our previous studies supporting a role for senescent cells in mediating pulmonary vascular remodeling. In these previous studies, we showed that the accumulation of senescent cells within pulmonary vessels contributed to neointima formation in patients with chronic lung diseases.24 Indeed, senescent cells are still metabolically active and release various factors, including cytokines, growth factors, and matrix components; which promote the proliferation of neighboring cells.24,36,37 Similar observations have been made for cancer cells, which can be stimulated by neighboring senescent cells.36 Thus, an
important issue regarding PH is whether senescent cells can accumulate for long periods in pulmonary vessels. In theory, such accumulation is prevented via senescent-cell clearance by the immune system.38 However, in age-related diseases such as chronic obstructive lung disease, combined mecha-nisms, including increased susceptibility to cell senescence and an inability of the immune system to clear senescent cells, probably promote the accumulation of senescent cells within the vascular wall. Whether this may occur in response to Nutlin treatment may depend on various factors such as age, immune system performance, and environmental condi-tions.38 Thus, inducing cell senescence might hold promise as a treatment against pulmonary vascular remodeling, pro-vided the senescent cells are simultaneously cleared by the immune system.
Further studies are therefore needed to identify the mecha-nisms responsible for senescent-cell clearance in PH. Whether prosenescence strategies such as Nutlin therapy hold promise for the treatment of PH will need to be evaluated.
Sources of Funding This study was supported by grants from the INSERM, Fondation pour la Recherche Médicale, Ministère de la Recherche, Chancellerie des universities de Paris, and Fondation Caresen.
DisclosuresNone.
References 1. Morrell NW, Adnot S, Archer SL, Dupuis J, Jones PL, MacLean MR,
McMurtry IF, Stenmark KR, Thistlethwaite PA, Weissmann N, Yuan JX, Weir EK. Cellular and molecular basis of pulmonary arterial hyperten-sion. J Am Coll Cardiol. 2009;54(1 Suppl):S20–S31.
2. Eddahibi S, Humbert M, Fadel E, Raffestin B, Darmon M, Capron F, Simonneau G, Dartevelle P, Hamon M, Adnot S. Serotonin trans-porter overexpression is responsible for pulmonary artery smooth muscle hyperplasia in primary pulmonary hypertension. J Clin Invest. 2001;108:1141–1150.
3. Yeager ME, Halley GR, Golpon HA, Voelkel NF, Tuder RM. Microsatellite instability of endothelial cell growth and apoptosis genes within plexiform lesions in primary pulmonary hypertension. Circ Res. 2001;88:E2–E11.
4. Lee SD, Shroyer KR, Markham NE, Cool CD, Voelkel NF, Tuder RM. Monoclonal endothelial cell proliferation is present in primary but not sec-ondary pulmonary hypertension. J Clin Invest. 1998;101:927–934.
5. Eddahibi S, Guignabert C, Barlier-Mur AM, Dewachter L, Fadel E, Dartevelle P, Humbert M, Simonneau G, Hanoun N, Saurini F, Hamon M, Adnot S. Cross talk between endothelial and smooth muscle cells in pul-monary hypertension: critical role for serotonin-induced smooth muscle hyperplasia. Circulation. 2006;113:1857–1864.
6. Atkinson C, Stewart S, Upton PD, Machado R, Thomson JR, Trembath RC, Morrell NW. Primary pulmonary hypertension is associated with reduced pulmonary vascular expression of type II bone morphogenetic protein receptor. Circulation. 2002;105:1672–1678.
7. McMurtry MS, Archer SL, Altieri DC, Bonnet S, Haromy A, Harry G, Bonnet S, Puttagunta L, Michelakis ED. Gene therapy targeting survivin selectively induces pulmonary vascular apoptosis and reverses pulmonary arterial hypertension. J Clin Invest. 2005;115:1479–1491.
8. Schermuly RT, Dony E, Ghofrani HA, Pullamsetti S, Savai R, Roth M, Sydykov A, Lai YJ, Weissmann N, Seeger W, Grimminger F. Reversal of experimental pulmonary hypertension by PDGF inhibition. J Clin Invest. 2005;115:2811–2821.
9. Petitjean A, Achatz MI, Borresen-Dale AL, Hainaut P, Olivier M. TP53 mutations in human cancers: functional selection and impact on cancer prognosis and outcomes. Oncogene. 2007;26:2157–2165.
10. Kubbutat MH, Jones SN, Vousden KH. Regulation of p53 stability by Mdm2. Nature. 1997;387:299–303.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from

1676 Circulation April 23, 2013
11. Mayo LD, Donner DB. The PTEN, Mdm2, p53 tumor suppressor-oncoprotein network. Trends Biochem Sci. 2002;27:462–467.
12. Vargas DA, Takahashi S, Ronai Z. Mdm2: A regulator of cell growth and death. Adv Cancer Res. 2003;89:1–34.
13. Vassilev LT, Vu BT, Graves B, Carvajal D, Podlaski F, Filipovic Z, Kong N, Kammlott U, Lukacs C, Klein C, Fotouhi N, Liu EA. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science. 2004;303:844–848.
14. Vousden KH. Outcomes of p53 activation–spoilt for choice. J Cell Sci. 2006;119(Pt 24):5015–5020.
15. Alimonti A, Nardella C, Chen Z, Clohessy JG, Carracedo A, Trotman LC, Cheng K, Varmeh S, Kozma SC, Thomas G, Rosivatz E, Woscholski R, Cognetti F, Scher HI, Pandolfi PP. A novel type of cellular senescence that can be enhanced in mouse models and human tumor xenografts to sup-press prostate tumorigenesis. J Clin Invest. 2010;120:681–693.
16. Hashimoto T, Ichiki T, Ikeda J, Narabayashi E, Matsuura H, Miyazaki R, Inanaga K, Takeda K, Sunagawa K. Inhibition of MDM2 attenuates neointimal hyperplasia via suppression of vascular proliferation and in-flammation. Cardiovasc Res. 2011;91:711–719.
17. Ihling C, Haendeler J, Menzel G, Hess RD, Fraedrich G, Schaefer HE, Zeiher AM. Co-expression of p53 and MDM2 in human atherosclerosis: implications for the regulation of cellularity of atherosclerotic lesions. J Pathol. 1998;185:303–312.
18. Nakamura Y, Suzuki S, Suzuki T, Ono K, Miura I, Satoh F, Moriya T, Saito H, Yamada S, Ito S, Sasano H. MDM2: a novel mineralocorticoid-responsive gene involved in aldosterone-induced human vascular struc-tural remodeling. Am J Pathol. 2006;169:362–371.
19. An WG, Kanekal M, Simon MC, Maltepe E, Blagosklonny MV, Neckers LM. Stabilization of wild-type p53 by hypoxia-inducible factor 1alpha. Nature. 1998;392:405–408.
20. Efeyan A, Ortega-Molina A, Velasco-Miguel S, Herranz D, Vassilev LT, Serrano M. Induction of p53-dependent senescence by the MDM2 antagonist nutlin-3a in mouse cells of fibroblast origin. Cancer Res. 2007;67:7350–7357.
21. Kumamoto K, Spillare EA, Fujita K, Horikawa I, Yamashita T, Appella E, Nagashima M, Takenoshita S, Yokota J, Harris CC. Nutlin-3a activates p53 to both down-regulate inhibitor of growth 2 and up-regulate mir-34a, mir-34b, and mir-34c expression, and induce senescence. Cancer Res. 2008;68:3193–3203.
22. Huang B, Deo D, Xia M, Vassilev LT. Pharmacologic p53 activation blocks cell cycle progression but fails to induce senescence in epithelial cancer cells. Mol Cancer Res. 2009;7:1497–1509.
23. Miyachi M, Kakazu N, Yagyu S, Katsumi Y, Tsubai-Shimizu S, Kikuchi K, Tsuchiya K, Iehara T, Hosoi H. Restoration of p53 pathway by nutlin-3 induces cell cycle arrest and apoptosis in human rhabdomyosarcoma cells. Clin Cancer Res. 2009;15:4077–4084.
24. Noureddine H, Gary-Bobo G, Alifano M, Marcos E, Saker M, Vienney N, Amsellem V, Maitre B, Chaouat A, Chouaid C, Dubois-Rande JL, Damotte D, Adnot S. Pulmonary artery smooth muscle cell senescence is a pathogenic mechanism for pulmonary hypertension in chronic lung disease. Circ Res. 2011;109:543–553.
25. Ciuclan L, Bonneau O, Hussey M, Duggan N, Holmes AM, Good R, Stringer R, Jones P, Morrell NW, Jarai G, Walker C, Westwick J, Thomas M. A novel murine model of severe pulmonary arterial hypertension. Am J Respir Crit Care Med. 2011;184:1171–1182.
26. Guignabert C, Izikki M, Tu LI, Li Z, Zadigue P, Barlier-Mur AM, Hanoun N, Rodman D, Hamon M, Adnot S, Eddahibi S. Transgenic mice over-expressing the 5-hydroxytryptamine transporter gene in smooth muscle develop pulmonary hypertension. Circ Res. 2006;98:1323–1330.
27. Gary-Bobo G, Houssaini A, Amsellem V, Rideau D, Pacaud P, Perrin A, Brégeon J, Marcos E, Dubois-Randé JL, Sitbon O, Savale L, Adnot S. Effects of HIV protease inhibitors on progression of monocro-taline- and hypoxia-induced pulmonary hypertension in rats. Circulation. 2010;122:1937–1947.
28. Mizuno S, Bogaard HJ, Kraskauskas D, Alhussaini A, Gomez-Arroyo J, Voelkel NF, Ishizaki T. p53 Gene deficiency promotes hypoxia-induced pulmonary hypertension and vascular remodeling in mice. Am J Physiol Lung Cell Mol Physiol. 2011;300:L753–L761.
29. Nardella C, Clohessy JG, Alimonti A, Pandolfi PP. Pro-senescence therapy for cancer treatment. Nat Rev Cancer. 2011;11:503–511.
30. Wade M, Wang YV, Wahl GM. The p53 orchestra: Mdm2 and Mdmx set the tone. Trends Cell Biol. 2010;20:299–309.
31. Shangary S, Qin D, McEachern D, Liu M, Miller RS, Qiu S, Nikolovska-Coleska Z, Ding K, Wang G, Chen J, Bernard D, Zhang J, Lu Y, Gu Q, Shah RB, Pienta KJ, Ling X, Kang S, Guo M, Sun Y, Yang D, Wang S. Temporal activation of p53 by a specific MDM2 inhibitor is selectively toxic to tumors and leads to complete tumor growth inhibition. Proc Natl Acad Sci USA. 2008;105:3933–3938.
32. Lane DP, Brown CJ, Verma C, Cheok CF. New insights into p53 based therapy. Discov Med. 2011;12:107–117.
33. Sano M, Minamino T, Toko H, Miyauchi H, Orimo M, Qin Y, Akazawa H, Tateno K, Kayama Y, Harada M, Shimizu I, Asahara T, Hamada H, Tomita S, Molkentin JD, Zou Y, Komuro I. p53-induced inhibition of Hif-1 causes cardiac dysfunction during pressure overload. Nature. 2007;446:444–448.
34. Wei Q, Dong G, Yang T, Megyesi J, Price PM, Dong Z. Activation and involvement of p53 in cisplatin-induced nephrotoxicity. Am J Physiol Renal Physiol. 2007;293:F1282–F1291.
35. Meek DW, Knippschild U. Posttranslational modification of MDM2. Mol Cancer Res. 2003;1:1017–1026.
36. Campisi J. Senescent cells, tumor suppression, and organismal aging: good citizens, bad neighbors. Cell. 2005;120:513–522.
37. Amsellem V, Gary-Bobo G, Marcos E, Maitre B, Chaar V, Validire P, Stern JB, Noureddine H, Sapin E, Rideau D, Hue S, Le Corvoisier P, Le Gouvello S, Dubois-Randé JL, Boczkowski J, Adnot S. Telomere dys-function causes sustained inflammation in chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2011;184:1358–1366.
38. Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, Kirkland JL, van Deursen JM. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011;479:232–236.
CLINICAL PERSPECTIvEHyperplasia of pulmonary-artery smooth muscle cells (PA-SMCs) is the primary determinant of the pulmonary vessel remodeling process that underlies pulmonary hypertension (PH). Because similarities exist between cancer and PH, drugs designed to treat malignant proliferative disorders might prove useful for treating PH. Nutlins are anticancer agents that disrupt the interaction between p53 and its negative regulator murine double minute 2, a specific p53 ubiquitin ligase and transcriptional inhibitor. Here, we showed that Nutlin-3a attenuated or reversed PH in 3 distinct experimental mouse mod-els, decreased the number of proliferating PA-SMCs, induced PA-SMC senescence in vitro, and increased the number of p21-stained senescent cells. Interestingly, Nutlin-3a markedly increased lung p53 protein and downstream p53 gene expres-sion in mice with PH, whereas only minor effects were noted in control mice without PH. The absence of nutlin toxicity for normal tissue may be ascribable to differential activities of murine double minute 2 in proliferative versus nonproliferative cells. Thus, Nutlin-3a holds promise as a prosenescence treatment targeting PA-SMCs during PH progression.
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from

Valérie Amsellem and Serge AdnotHoussaini, Jean-Luc Dubois-Rande, Laurent Boyer, Jorge Boczkowski, Geneviève Derumeaux, Nathalie Mouraret, Elisabeth Marcos, Shariq Abid, Guillaume Gary-Bobo, Mirna Saker, Amal
HypertensionActivation of Lung p53 by Nutlin-3a Prevents and Reverses Experimental Pulmonary
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2013 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/CIRCULATIONAHA.113.002434
2013;127:1664-1676; originally published online March 19, 2013;Circulation.
http://circ.ahajournals.org/content/127/16/1664World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circ.ahajournals.org/content/suppl/2013/03/19/CIRCULATIONAHA.113.002434.DC1.htmlData Supplement (unedited) at:
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialCirculationin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on February 8, 2016http://circ.ahajournals.org/Downloaded from

CIRCULATIONAHA/2012/097238 R3
1
SUPPLEMENTAL MATERIAL
Activation of Lung p53 by Nutlin-3a Prevents and Reverses Experimental Pulmonary
Hypertension
Mouraret; Role of Cell Senescence in Pulmonary Hypertension
Mouraret N MSC, Marcos E MSC, Abid S MSC, Gary-Bobo G PhD, Saker M MSC,
Houssaini A MSC, Dubois-Rande JL MD, Boyer L MD, Boczkowski J MD-PhD, Derumeaux
G MD, Amsellem V PhD, Adnot S MD-PhD

CIRCULATIONAHA/2012/097238 R3
2
Supplemental methods
Echocardiography
Closed-chest transthoracic echocardiography was performed in nonsedated mice as previously
described1. Images were acquired using a 13-MHz linear-array transducer with a digital
ultrasound system (Vivid 7, GE Medical Systems). Conventional measurements [LV
diameters, wall thickness and thickening] were obtained serially from M-mode tracings at the
level of the papillary muscles at each time point. LV end-systolic and end-diastolic volumes
(LVESV and LVEDV, respectively) were calculated from the M-mode image by the “D3”
respectively as LVESD3 and LVEDD3. Stroke volume (SV) was derived from the difference
between LVEDV and LVESV. Cardiac output (CO) was calculated as the product of SV and
heart rate2. The pulsed-wave Doppler recording of pulmonary flow was obtained from the
parasternal short-axis view at the level of the aortic valve. The pulsed-wave Doppler sample
was positioned within the pulmonary infundibulum at the tip of the pulmonary leaflets. The
following variables were measured: systolic time-velocity integral of pulmonary flow (VTI,
cm), pulmonary acceleration time (PAT, ms: time from pulmonary flow onset to peak
velocity), ejection time (ET, ms: time from onset to end of systolic flow velocity), and the
PAT/ET ratio (%)) 1. All measures were averaged over five cardiac cycles and the reader was
blinded to the treatment of the mice.
Real-time reverse-transcription quantitative PCR (RT-qPCR)
The levels of p53 downstream genes were determined by real-time reverse-transcription (RT)-
qPCR using SYBR Green (Invitrogen, Cergy-Pontoise, France). To normalize for cDNA input
load, mouse 18s was used as an endogenous standard. Specific primers were as follows: p21
forward, 5’-CAGATCCACAGCGATATCCA-3’ and p21 reverse, 5’-

CIRCULATIONAHA/2012/097238 R3
3
ACGGGACCGAAGAGACAAC-3’; Bax forward, 5’-AGGATGCGTCCACCAAGAAGCT-
3’ and Bax reverse, 5’- TCCGTGTCCACGTCAGCAATCA-3’; Bcl2 forward, 5’-
GGGAGGATTGTGGCCTTCTT-3’ and Bcl2 reverse, 5’-CATCCCAGCCTCCGTTATCCT-
3’; Puma forward, 5’- ACCGCTCCACCTGCCGTCAC-3’ and Puma reverse, 5’-
ACGGGCGACTCTAAGTGCTGC-3’; BTG2 forward, 5’-
GAGCGAGCAGAGACTCAAGGTT-3’ and BTG2 reverse, 5’-
CGATAGCCAGAACCTTTGGATGG-3’. After addition of cDNA to SYBR Green Master
Mix PCR (300 nM of each specific primer, PCR was performed in a total volume of 25 µL in
a 7900HT real-time PCR cycler (Applied Biosystems, Courtaboeuf, France). All cDNA
samples were tested in duplicate and analyzed using ABI Prism Sequence Detection Software
v.1.7 (PE Applied Biosystems). Samples were compared using the relative Ct method. The Ct
value, which is inversely proportional to the initial template copy number, is the calculated
number of cycles with a fluorescence signal significantly above background levels. Fold
induction or repression was measured relative to controls and calculated after adjusting for
18s using 2–[Ct]
, where Ct = Ct tested gene – Ct 18s and Ct = Ct control – Ct
treatment.
Western Blotting
Tissues were sonicated in lysis buffer (Cell Signaling Technology, Beverly, MA). The lysate
was centrifuged for 10 minutes at 2000 rpm at 4°C and the supernatant was kept frozen. The
protein concentration of each sample was determined using the Bradford method. Samples
were subjected to electrophoresis in 10% or 15% polyacrylamide gels under reducing
conditions. After electrophoresis, proteins from the gel were electroblotted onto
polyvinylidene difluoride membranes (Millipore, Molsheim, France) for 2 h. After transfer,
the membranes were saturated with phosphate buffer saline (PBS)/5% milk then probed

CIRCULATIONAHA/2012/097238 R3
4
overnight at 4°C with antibodies. The antibodies used were anti-P-p53 Ser15 (Cell Signaling
Technology, MA, USA) at a final dilution of 1:1000 ; monoclonal anti p53 (Cell Signaling
Technology) at a final dilution of 1:1000; monoclonal anti-p21Waf1/Cip1 antibody (Cell
Signaling Technology) at a final dilution of 1:1000, or polyclonal anti-MDM2 (Abcam,
Cambridge, UK) at a final dilution of 1:1000. After three 5-minute washes in TBS-T,
membranes were incubated for 1 hour at room temperature with horseradish peroxidase-
conjugated secondary antibodies at a final dilution of 1:1000 and washed twice for 5 minutes
in TBS-T. Proteins were visualized by electrochemiluminescence (Amersham, Little Chalfont,
UK) according to the manufacturer’s instructions. Membranes were re-probed with anti--
actin or anti-GAPDH antibody (Sigma, Saint-Quentin-Fallavier, France) at a final dilution of
1:1000 for normalization. Complementary experiments were performed in mouse tissue to
assess phosphorylated-p53 levels by using a co-immunoprecipitation procedure or to
determine the nuclear and cytoplasmic fractions of total p53 protein.
Immunoprecipitation
Immunoprecipitation was carried out at 4°C. Lung tissue was sonicated in lysis buffer (Cell
Signaling Technology, MA, USA). The lysate was centrifuged for 10 minutes at 2000 rpm at
4°C and the supernatant was kept frozen. After assessment of the protein concentration, 1 mg
of protein from each sample was incubated with the p53 (1C12) antibody (Cell
SignalingTechnology) overnight then with protein A/G PLUS-Agarose beads (Santa Cruz
Biotechnology, Santa Cruz, CA, USA) for 3 h. The protein–antibody complexes recovered on
beads were subjected to Western blot analysis after separation by SDS–PAGE. The membrane
was probed with phosphorylated-p53 (Ser15) primary antibody (Cell Signaling Technology)
and with a horseradish-peroxidase-conjugated secondary antibody. Blots were visualized by
electrochemiluminescence.

CIRCULATIONAHA/2012/097238 R3
5
Cytosolic and nuclear fraction isolation
To obtain the nuclear fraction, lung tissue was incubated in hypotonic buffer (20 mM Tris
HCl, pH 7.4; 10 mM NaCl; 3 mM MgCl2) for 15 min at 4°C. Triton (0.3%) was added to the
lysate, which was then centrifuged at 14,000 g for 10 min at 4°C and separated into soluble
and insoluble fractions. The insoluble fraction containing the cytosolic protein was kept at -
80°C, and the pellet containing the nuclei was incubated in lysis buffer (Invitrogen, Carlsbad,
CA, USA) for 30 min. After extraction, the sample was centrifuged at 14,000 g for 10 min at
4°C, and the supernatant constituted the nuclear fraction. Cytosolic and nuclear protein
samples were analyzed by Western blotting. The membranes were incubated with the
appropriate primary antibody, p53 (1C12) antibody (Cell Signaling Technology), lamin A/C
antibody (Santa Cruz), or alpha-tubulin (Cell Signaling Technology).
Transient tranfection of pulmonary-artery smooth muscle cells
Cells were seeded in 24-well plates and transiently transfected the following day with a p53
reporter construct (p53 Cignal Reporter Assay, Qiagen, Courtaboeuf, France), as well as with
positive and negative controls, using Lipofectamin 2000 (Invitrogen) according to the
manufacturer’s instructions. After 18 h of transfection, the medium was changed and replaced
by complete growth medium, and the cells were treated with Nutlin-3a 2.5 µM, 5 µM, 10 µM,
or ethanol alone. A dual-luciferase assay (Promega, Charbonnières les Bains, France) was
performed 24 h after treatment. Cells were lysed with lysis buffer, and luciferase activity was
quantified using a Tristar luminometer (Berthold, Thoiry, France). Values are expressed as
arbitrary units using a construct constitutively expressing Renilla luciferase for internal
normalization. Firefly/Renilla activity ratios were generated from experimental and control
transfections. Experiments were done in triplicate.

CIRCULATIONAHA/2012/097238 R3
6
Immunohistochemistry
Paraffin-embedded sections were deparaffinized using xylene and a graded series of ethanol
dilutions then incubated in citrate buffer (0.01 M, pH 6) at 90°C for 20 minutes. Tissue was
permeabilized with 0.1% Triton X-100 in PBS for 10 minutes. Endogenous peroxidase
activity was blocked with 3% H2O2 and 10% methanol in PBS for 10 minutes. Slides were
incubated for 60 minutes in 1% bovine serum albumin and 5% goat serum in PBS then
incubated overnight with anti-p21 mouse antibody (1:200, Santa Cruz Biotechnology) and
anti-Ki67 rabbit antibody (1:300, Abcam). We used the ABC Vectastain kit (Vector Labs,
Burlingame, CA, USA) to mark the primary antibodies according to the user’s guide. The
staining substrate was diaminobenzidine (FastDAB, Sigma-Aldrich, St Louis, MO, USA) and
the sections were counterstained with methyl green.
For terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL), epitopes were
retrieved by treatment with DNase1 digestion for 10 min at room temperature. After
endogenous peroxidase blockade with H2O2, tissues were incubated in equilibration buffer
and treated with terminal deoxynucleotidyl transferase (TdT) to detect TUNEL-positive nuclei
as suggested by the manufacturer (Roche, Meylan, France). Tissues were then incubated with
peroxidase-conjugated anti-digoxigenin antibodies and color was developed with DAB. After
counterstaining with methyl green, sections were protected with coverslips secured with
mounting medium. For immunofluorescence, slides were incubated overnight at 4°C with
anti-MDM2 SMP-14 mouse antibody (1:50, Santa Cruz Biotechnology) and anti-SMA
rabbit antibody (1:200, Abcam) then exposed to anti-mouse Alexa Fluor (1:1000, Cell
Signaling technology) and anti-rabbit Alexa Fluor (1:1000, Invitrogen) antibodies. Nuclei
were stained with Hoechst.

CIRCULATIONAHA/2012/097238 R3
7
Online Table I
Normoxia
Vehicle
Normoxia
Nutlin-3a
Hypoxia
Vehicle
Hypoxia
Nutlin-3a
P value
HR, bpm 658±4 665±13 620±9 653±5 * 0.03
LVEF, % 87±4 83±2 83±3 82±2 0.57
CO, mL/min 33.4±1.8 35.8±2.4 36.8±3.5 33.7±1.9 0.64
Pulmonary TVI, cm 3.3±0.1 3.2±0.1 2.5±0.1† 2.3±0.1 0.001
PAT, ms 19.6±0.7 20.8±0.9 11.8±0.6† 14.1±0.8 * 0.001
ET, ms 52.8±1.9 51.8±0.8 49.4±1.3 47.0±0.8 0.02
PAT/ET, % 0.37±0.02 0.40±0.02 0.24±0.02† 0.30±0.02 * 0.001
Echocardiographic parameters in nonsedated mice. Data are mean±SEM. *P<0.05 for
comparison between hypoxic mice treated with Nutlin-3a and control untreated hypoxic mice.
† p<0.05 for comparison between vehicle-treated hypoxic and normoxic mice. HR, heart rate;
LVEF, left ventricular ejection fraction; CO, cardiac output; TVI, systolic time-velocity
integral of pulmonary flow; PAT, pulmonary acceleration time; ET, ejection time.
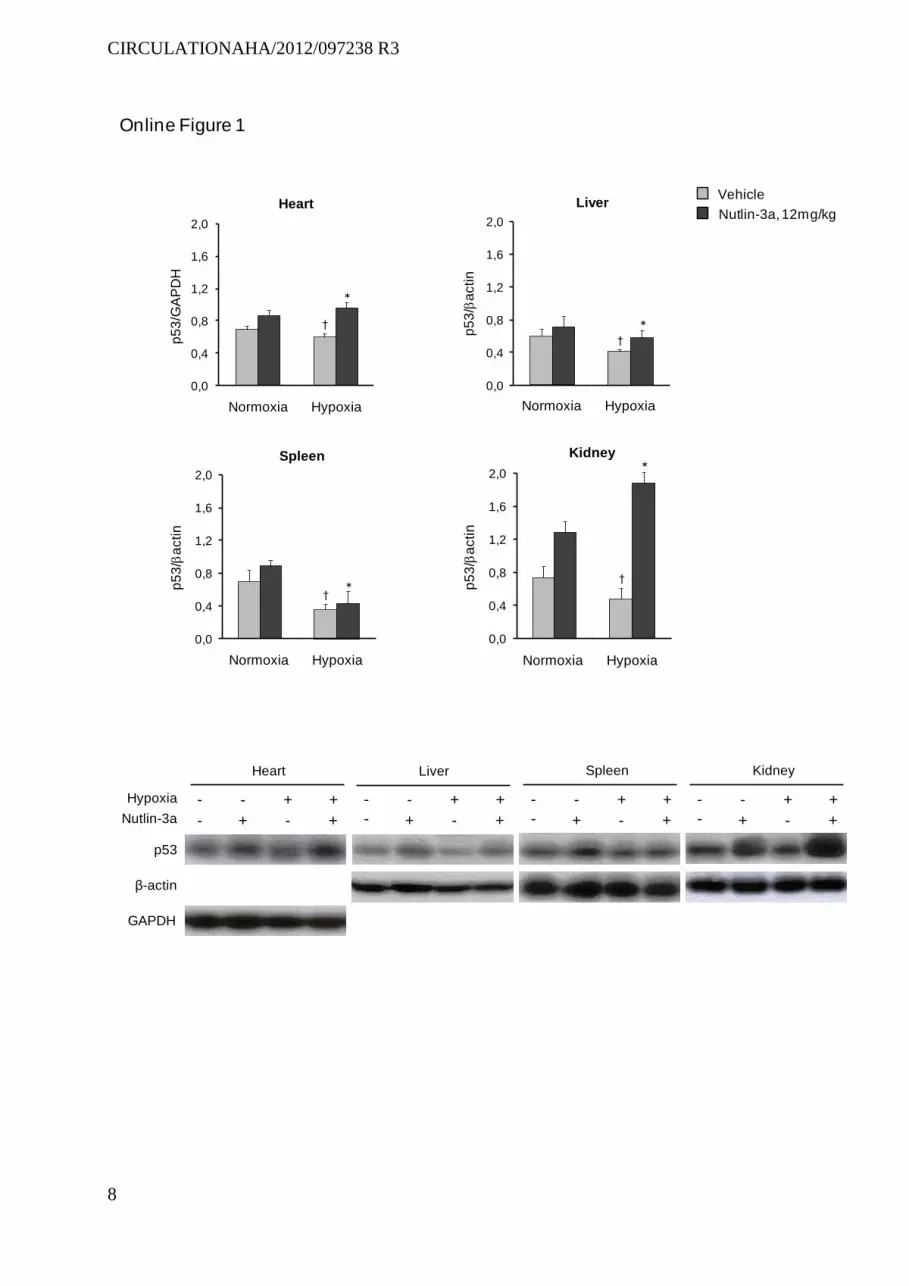
CIRCULATIONAHA/2012/097238 R3
8
Nutlin-3a
p53
β-actin
Hypoxia
Heart
- +
-
-
+
+
+
Liver Spleen Kidney
GAPDH
-
+
-
-
+
+
+-
- +
-
-
+
+
+-
- +
-
-
+
+
+-
-
0,0
0,4
0,8
1,2
1,6
2,0
p5
3/G
AP
DH
Normoxia Hypoxia
†
Heart
Online Figure 1
Vehicle
Nutlin-3a, 12mg/kg
0,0
0,4
0,8
1,2
1,6
2,0
p5
3/
actin
Normoxia Hypoxia
†
Liver
0,0
0,4
0,8
1,2
1,6
2,0
p5
3/
actin
Normoxia Hypoxia
†
Spleen
0,0
0,4
0,8
1,2
1,6
2,0p
53
/a
ctin
Normoxia Hypoxia
†
Kidney
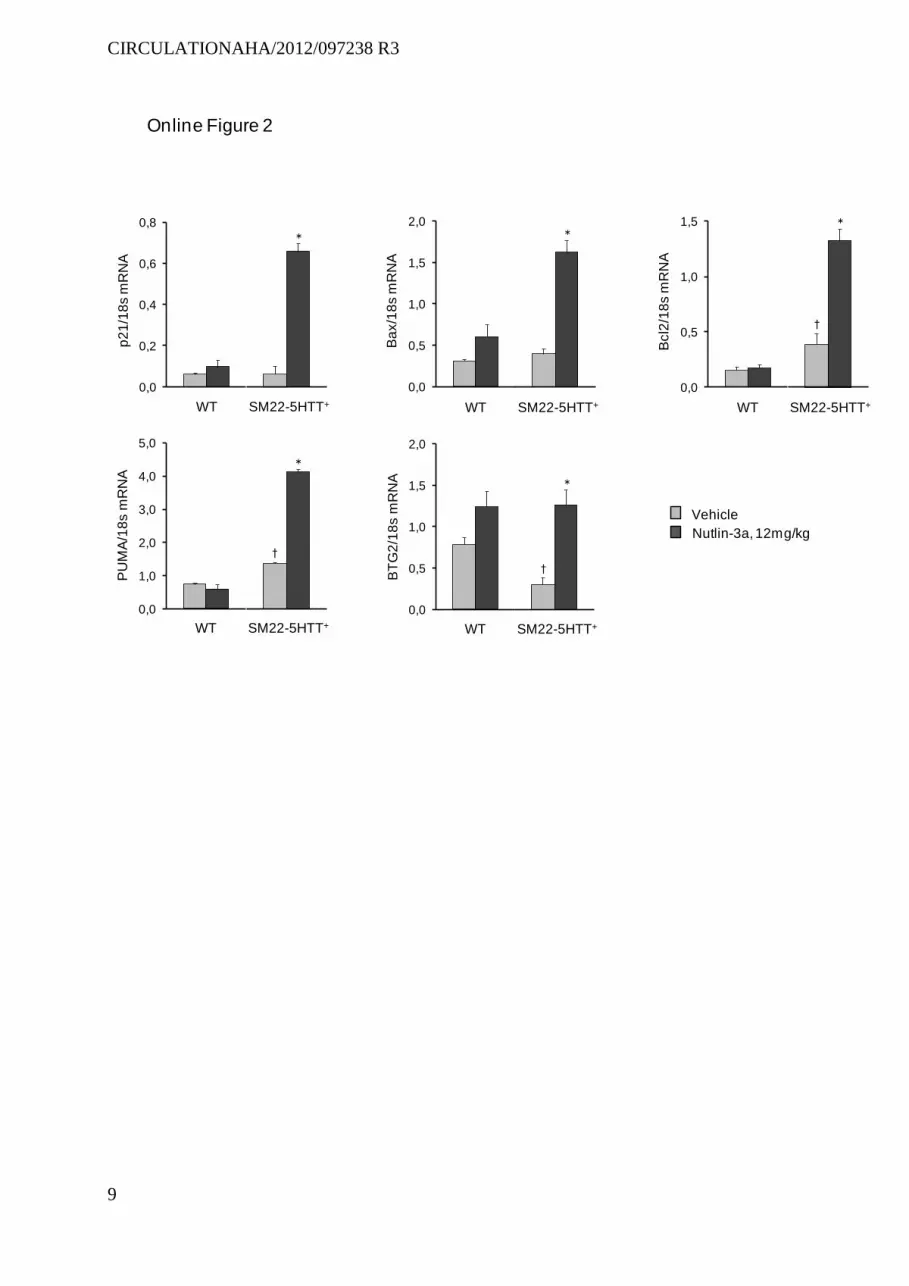
CIRCULATIONAHA/2012/097238 R3
9
Online Figure 2
0,0
1,0
2,0
3,0
4,0
5,0
PU
MA
/18
sm
RN
A
WT SM22-5HTT+
†
0,0
0,5
1,0
1,5
2,0
BT
G2
/18
sm
RN
A
WT SM22-5HTT+
†
0,0
0,5
1,0
1,5
Bcl2
/18
s m
RN
A
WT SM22-5HTT+
†
0,0
0,5
1,0
1,5
2,0
Ba
x/1
8s
mR
NA
WT SM22-5HTT+
0,0
0,2
0,4
0,6
0,8
p2
1/1
8s
mR
NA
WT SM22-5HTT+
Vehicle
Nutlin-3a, 12mg/kg
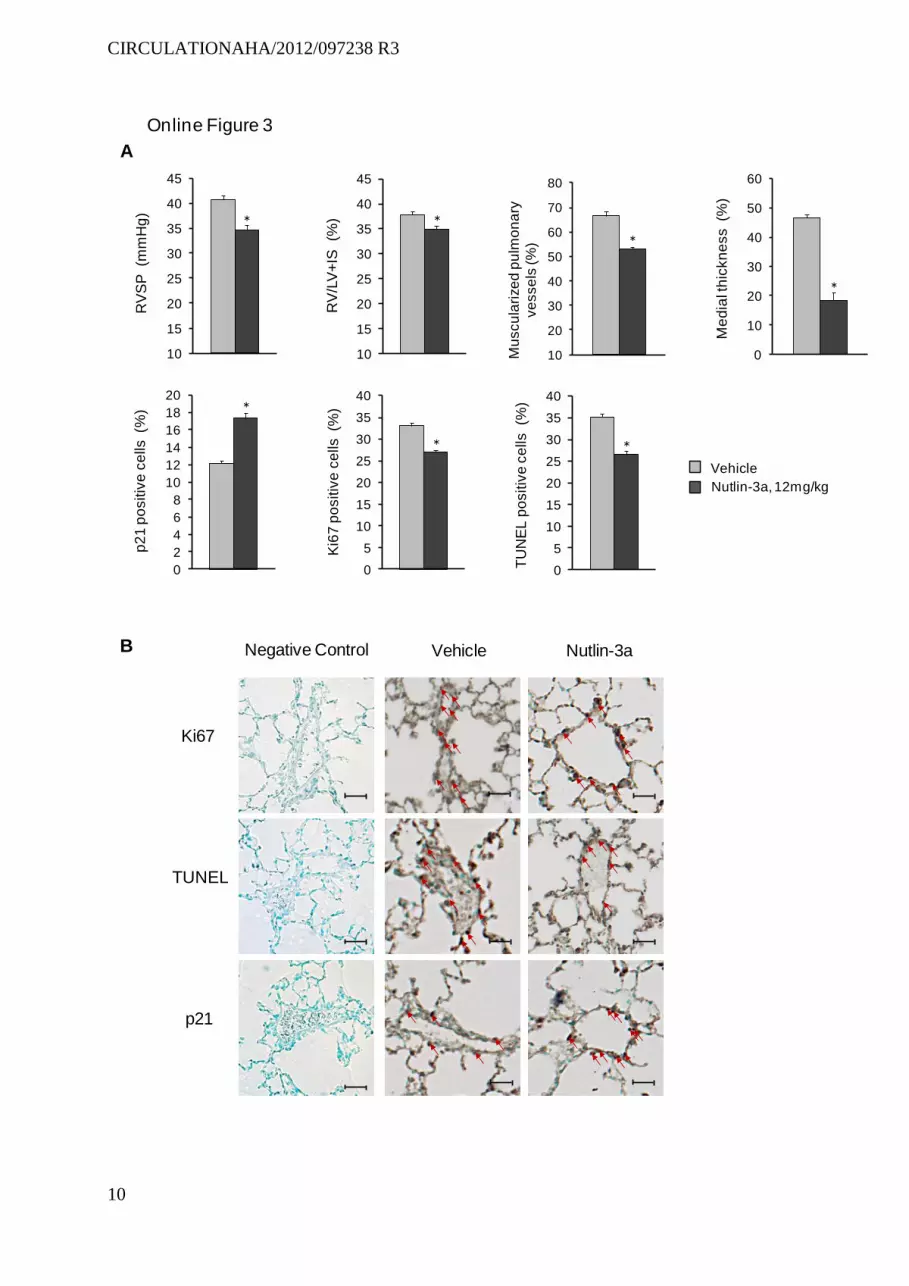
CIRCULATIONAHA/2012/097238 R3
10
10
20
30
40
50
60
70
80
Mu
scu
lari
ze
dp
ulm
on
ary
ve
sse
ls (%
)
10
15
20
25
30
35
40
45
RV
/LV
+IS
(%
)
10
15
20
25
30
35
40
45
RV
SP
(m
mH
g)
Online Figure 3
B
Vehicle
Nutlin-3a, 12mg/kg
0
2
4
6
8
10
12
14
16
18
20
p2
1 p
ositiv
e c
ells (%
)
0
5
10
15
20
25
30
35
40
Ki6
7 p
ositiv
e c
ells
(%)
0
5
10
15
20
25
30
35
40
TU
NE
L p
ositiv
e c
ells (%
)
0
10
20
30
40
50
60
Me
dia
l th
ickn
ess (%
)
A
Ki67
TUNEL
p21
Vehicle Nutlin-3aNegative Control
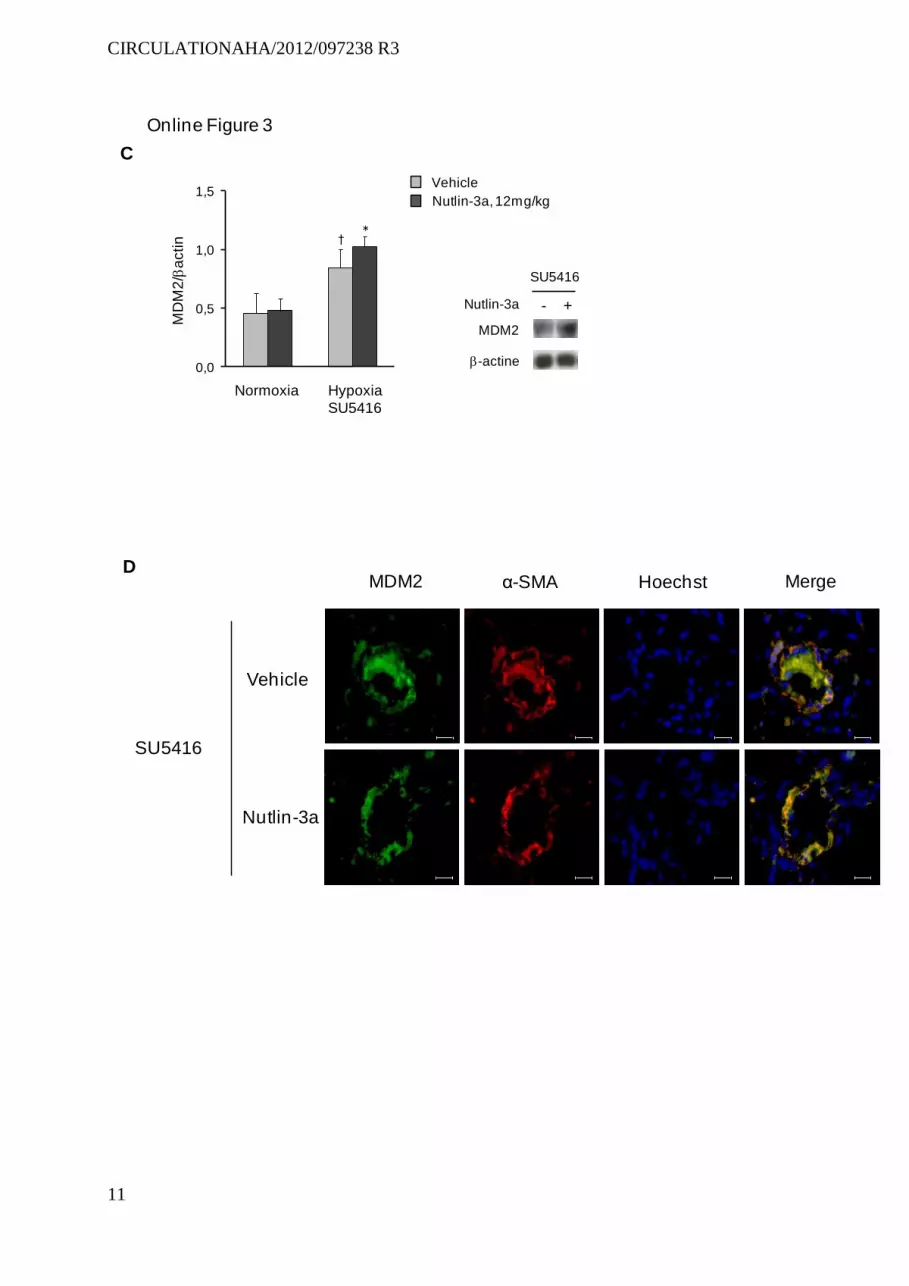
CIRCULATIONAHA/2012/097238 R3
11
Nutlin-3a
SU5416
- +
MDM2
-actine0,0
0,5
1,0
1,5M
DM
2/
actin
†
Normoxia Hypoxia
SU5416
Vehicle
Nutlin-3a
SU5416
MDM2 α-SMA Hoechst Merge
C
D
Vehicle
Nutlin-3a, 12mg/kg
Online Figure 3
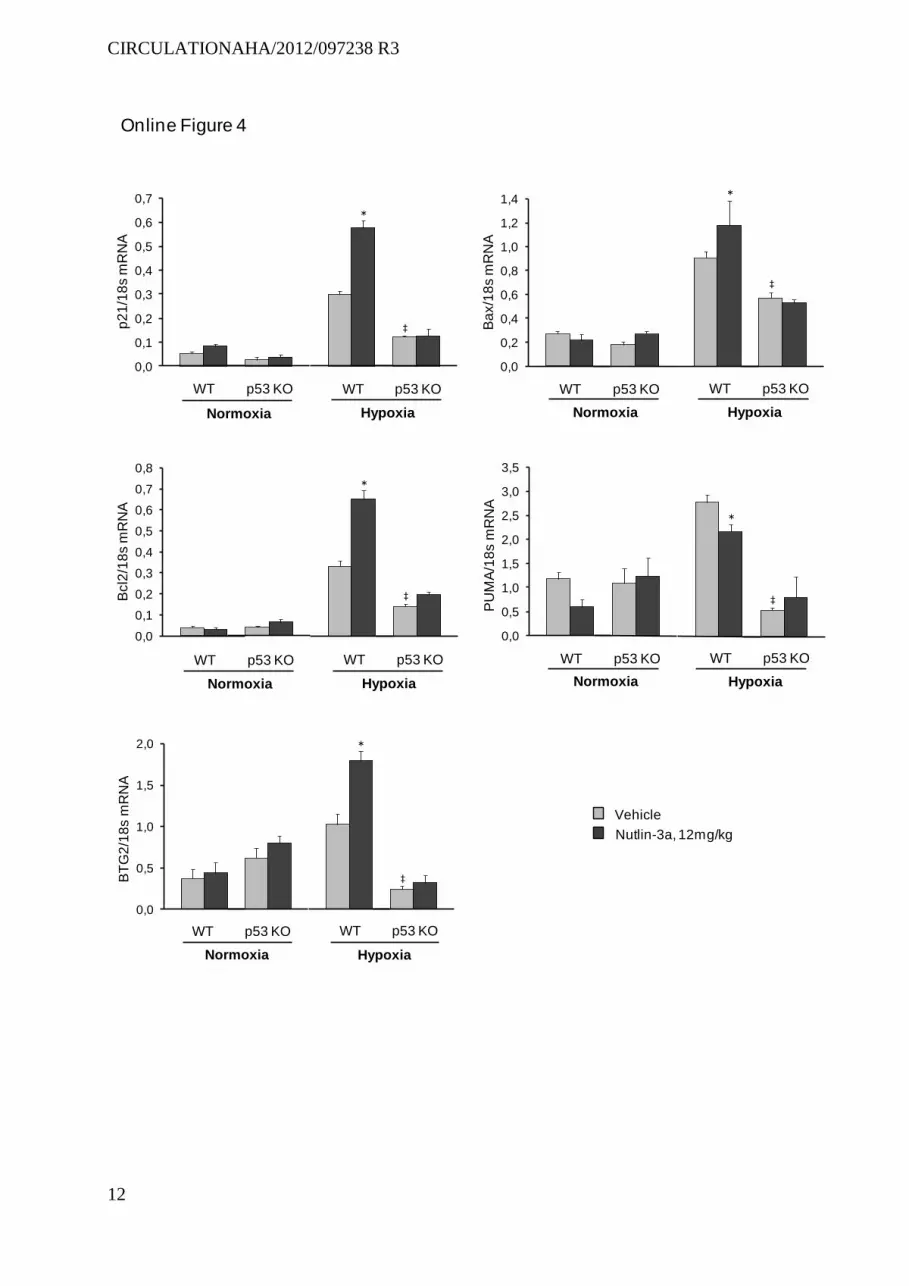
CIRCULATIONAHA/2012/097238 R3
12
0,0
0,5
1,0
1,5
2,0
BT
G2
/18
sm
RN
A
WT p53 KO WT p53 KO
Normoxia Hypoxia
‡
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
WT p53 KO WT p53 KO
Normoxia Hypoxia
PU
MA
/18
sm
RN
A
‡
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
WT p53 KO WT p53 KO
Normoxia Hypoxia
Bcl2
/18
sm
RN
A
‡
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Ba
x/1
8s
mR
NA
WT p53 KO WT p53 KO
Normoxia Hypoxia
‡
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
p2
1/1
8s m
RN
A
WT p53 KO WT p53 KO
Normoxia Hypoxia
‡
Online Figure 4
Vehicle
Nutlin-3a, 12mg/kg
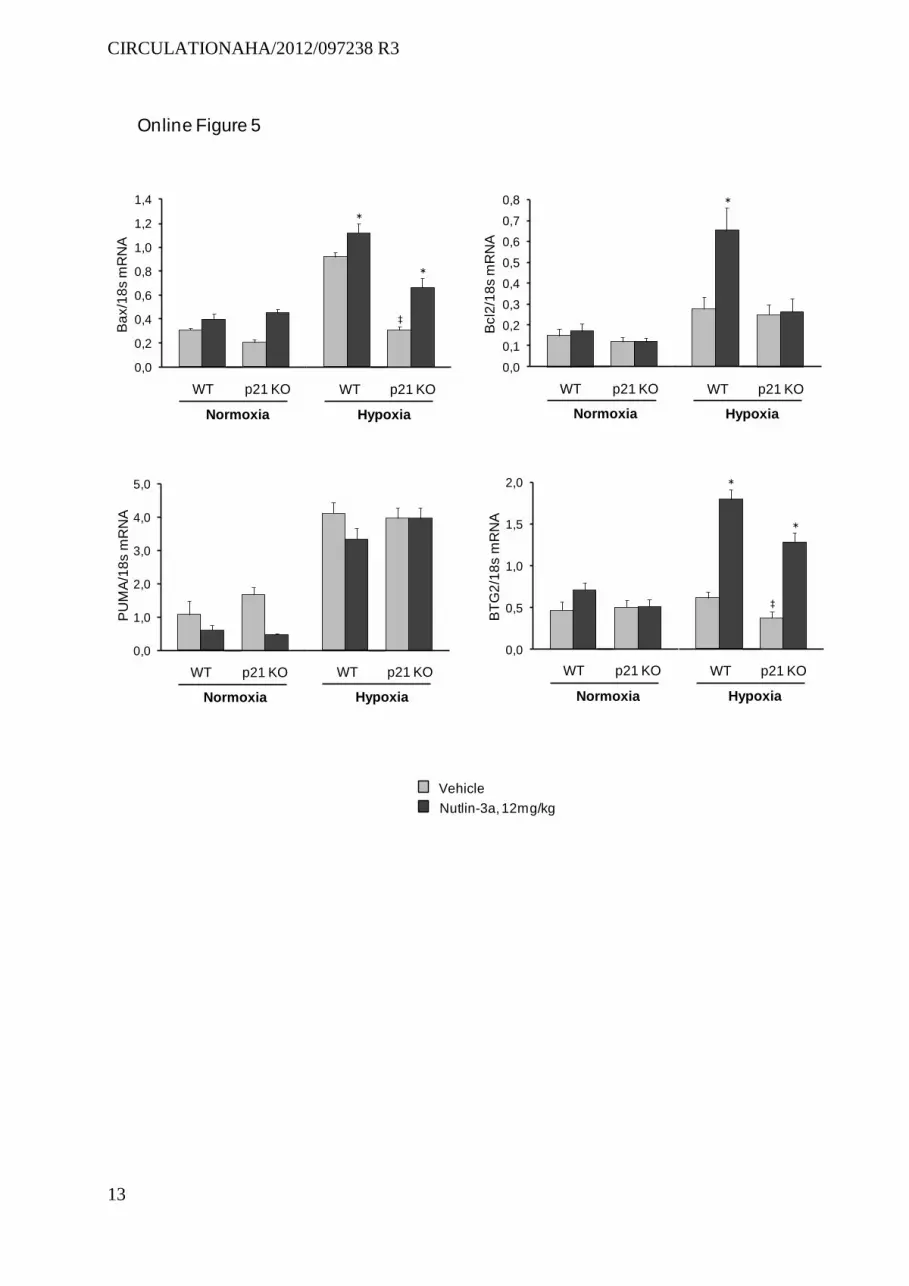
CIRCULATIONAHA/2012/097238 R3
13
Vehicle
Nutlin-3a, 12mg/kg
0,0
1,0
2,0
3,0
4,0
5,0
PU
MA
/18
sm
RN
A
WT p21 KO WT p21 KO
Normoxia Hypoxia
0,0
0,5
1,0
1,5
2,0B
TG
2/1
8s
mR
NA
WT p21 KO WT p21 KO
Normoxia Hypoxia
‡
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
WT p21 KO WT p21 KO
Normoxia Hypoxia
Bcl2
/18
s m
RN
A
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Ba
x/1
8s
mR
NA
WT p21 KO WT p21 KO
Normoxia Hypoxia
‡
Online Figure 5

CIRCULATIONAHA/2012/097238 R3
14
Online figure legends
Online Figure 1
Protein p53 levels measured by Western blot in the heart, liver, spleen, and kidneys in mice
studied on day 21 after exposure to hypoxia or normoxia and treated with daily i.p. injections
of 25 mg/Kg Nutlin-3a. Mice were sacrificed exactly 3 hours after the last Nutlin-3a dose.
*P<0.05 compared with values in vehicle-treated mice after exposure to hypoxia or normoxia.
Data are mean±SEM of 6 to 10 animals. †P<0.05 compared with values in vehicle-treated
mice after exposure to hypoxia.
Online Figure 2
Lung mRNA levels of p21, bax, Bcl2, BTG2, and PUMA measured by RT-PCR in SM22-5-
HTT+ mice and control mice studied after 21 days of treatment with vehicle or 12 mg/Kg of
Nutlin-3a. Mice were sacrificed exactly 3 hours after the last Nutlin-3a dose. Data are
mean±SEM of 6 to 10 animals. *P<0.05 compared with values in vehicle-treated mice.
†P<0.05 for SM22-5HTT+ versus vehicle-treated control mice.
Online Figure 3
A, right ventricular systolic pressure (RVSP); right ventricular hypertrophy index
(RV/[LV+S] weight ratio); pulmonary vessel muscularization (percentage of partially and
fully muscularized pulmonary vessels); medial wall thickness of fully muscularized
intraacinar arteries; and percentages of Ki67-positive dividing cells, p21-stained cells, and
TUNEL-positive cells in mice studied on day 21 after exposure to hypoxia and
simultaneously given a weekly intraperitoneal injection of 20 mg/Kg SU5416, and treated
daily by i.p. injections of vehicle or Nutlin-3a. B, representative micrographs of pulmonary

CIRCULATIONAHA/2012/097238 R3
15
vessels stained for Ki67, TUNEL, or p21. No immunoreactivity was detected in sections
incubated with secondary anti-rabbit and anti-mouse antibody but no primary antibody
(Negative controls). C, Lung protein levels of MDM2 measured by Western blot. D,
representative photographs of immunofluorescence staining for MDM2 in mice subjected to
SUGEN/hypoxia and treated with Nutlin-3a or vehicle for 21 days. MDM2 was
predominantly expressed in pulmonary vessels and co-localized with -SMA in smooth
muscle cells (SMCs). Data are mean±SEM of 6 to 10 animals.*P<0.05 compared with values
in vehicle-treated mice.
Online Figure 4
Lung mRNA levels of p21, Bax, Bcl2, PUMA, and BTG2 in wild-type and p53 knockout
(KO) mice studied on day 21 after exposure to hypoxia or normoxia and treated with daily i.p.
injections of 12 mg/Kg Nutlin-3a. Data are mean±SEM of 10 animals. *P<0.05 compared
with values in vehicle-treated mice. ‡P<0.05 compared with values in vehicle-treated control
mice.
Online Figure 5
Lung mRNA levels of Bax, Bcl2, PUMA, and BTG2 in wild-type and p21 knockout (KO)
mice studied on day 21 after exposure to hypoxia or normoxia and treated with daily i.p.
injections of 12 mg/Kg Nutlin-3a. Data are mean±SEM of 6 to 10 animals. *P<0.05 compared
with values in vehicle-treated mice. ‡P<0.05 compared with values in vehicle-treated control
mice.

CIRCULATIONAHA/2012/097238 R3
16
References
1. Thibault HB, Kurtz B, Raher MJ, Shaik RS, Waxman A, Derumeaux G, Halpern EF,
Bloch KD, Scherrer-Crosbie M. Noninvasive assessment of murine pulmonary arterial
pressure: validation and application to models of pulmonary hypertension. Circ Cardiovasc
Imaging.3(2):157-163.
2. Tournoux F, Petersen B, Thibault H, Zou L, Raher MJ, Kurtz B, Halpern EF, Chaput
M, Chao W, Picard MH, Scherrer-Crosbie M. Validation of noninvasive measurements of
cardiac output in mice using echocardiography. J Am Soc Echocardiogr.24(4):465-470.
Related Documents











