[CANCER RESEARCH 44, 1857-1862, May 1984] Activation of Cytotoxic Activity of Human Blood Lymphocytes by Tumor- promoting Compounds1 Oscar F. Ramos,2-3 Maria G. Masucci,2 and Eva Klein Department of Tumor Biology, Karolinska Institutet, S-104 01 Stockholm, Sweden ABSTRACT Three categories of tumor promoters and chemically related but inactive substances were tested for their effect on the cytotoxic activity of human blood lymphocytes against K562 and Daudi targets. Lymphocytes incubated overnight in the presence of phorbol esters 12-0-tetradecanoylphorbol-13-acetate and phorbol-12,13-dibutyrate [P(Bu)2] had enhanced function. Incu bation with 4-a-phorbol-12,13-didecanoate was without effect. Enhancing activity was also exerted by the indole alkaloids, teleocidin and lyngbyatoxin A, and the polyacetates, aplysiatoxin and debromoaplysiatoxin, but not by dihydroteleocidin. Only the tumor-promoting compounds activated the cytotoxic potential. The substances acted in a dose-dependent manner with optimal activity at characteristic concentrations. Overnight incubation of lymphocytes at 4°did not change their spontaneous cytotoxicity but abolished the enhancing effect of P(Bu)2. Thus, P(Bu)2-induced activation occurred only on meta- bolically active cells. The activation did not require DNA synthe sis. Similar to controls, the P(Bu)2-treated cells required divalent cations and an intact cytoskeleton in order to perform their lytic function. Experiments with the various metabolic inhibitors indi cate that phorbol ester treatment does not induce an alternative cytotoxic mechanism since, as with untreated lymphocytes, P(Bu)2-activated cells require contact with the target and intact secretory functions. The enhanced cytolytic potential was not due to induction of a-interferon (IFN-a) production, as shown by the fact that the effect was not abolished by addition of anti-IFN antibodies during the P(Bu)2 treatment of lymphocytes or during the cytotoxic assay. However, the presence of antiserum against IFN reduced the cytotoxic potential of control cells, suggesting that endoge nous IFN production contributes to the maintenance of lytic function in cultured cells. If this mechanism is counteracted by addition of anti-IFN serum, the phorbol esters can provide an alternative activation signal. When P(Bu)2-activated lymphocytes were subsequently treated with IFN-a or IFN-y, their lytic capac ity was further increased. These results indicate that P(Bu)2 and IFN activate cytotoxic potential through different pathways. INTRODUCTION Phorbol esters and similar diterpenes act as tumor promoters in mouse skin (8, 40). In order to understand the mechanism of tumor promotion, these compounds have been widely studied in in vitro systems. They were found to alter the growth behavior, 1This investigation was supported by USPHS Grant 5 R01 CA25250-04 awarded by the National Cancer Institute, Department of Health and Human Services, and by the Swedish Cancer Society. 2 Recipients of fellowships from the Cancer Research Institute/Concern Foun dation. 3 To whom requests for reprints should be addressed. Received August 1,1983; accepted February 10,1984. differentiation, and functional characteristics of cells (40). Their effect is initiated by interaction with specific receptors expressed on the plasma membrane (9, 11, 33, 43). These effects were studied extensively with phorbol esters like TPA4 (8, 40). Indole alkaloids and polyacetates derived from Streptomyces and the blue-green alga Lyngbya majuscula are 2 further classes of tumor promoters chemically unrelated to phorbol esters (10,13). They interact with the same surface receptor as does TPA and induce similar biological effects (12, 39). Tumor promoters modify the immune response and the func tional behavior of lymphocytes (for review, see Ref. 24). They act as mitogens and enhance the action of other mitogens (15). They stimulate the production of IFN and lymphokines (27, 33, 44, 45). They activate the cytotoxic potential of polymorphonu- clear leukocytes (7), and under certain conditions both stimula tory and inhibitory effects on the tumoricidal capacity of TPA- treated macrophages were reported (18, 20, 23). Phorbol esters modify the cytotoxic activity of lymphocytes. They inhibit the generation of allospecific cytotoxic cells in mixed- lymphocyte cultures but activate the cytotoxic potential of pre- sensitized effectors (2, 26). Depending on the concentration, the length of their interaction, and the presence of regulatory cells, tumor promoters can either enhance or suppress NK activity (for review, see Ref. 19). We have shown previously that the tumor promoters TPA and P(Bu)2 have a complex modulatory effect on human NK assays because they can influence the behavior of both effector and target cells (30). Long-term incubation with nM concentrations of the compounds resulted in activation of the lytic potential of lymphocytes. This effect was counteracted via activation of the suppressive function of monocytes. The target sensitivity was also modified. In the present study, we have tested the effect of the indole alkaloids teleocidin and lyngbyatoxin A and the polyacetates aplysiatoxin and debromoaplysiatoxin on the human NK system using K562 and Daudi targets. In addition, we analyzed certain characteristics of the P(Bu)2-induced enhancement of the lytic function with special attention to the possible involvement of IFN on the phenomenon. MATERIALS AND METHODS Experimental Design. Buffy coat leukocytes were obtained from healthy blood donors. Mononuclear cells were separated by the method of Boyurn (4) on Ficoll-lsopaque gradients and were subsequently incu bated in a plastic bottle (Falcon) at 37°in 5% CO2:air for 60 min in order to deplete mononuclear adherent cells. Nonadherent cells were subjected to a nylon wool column chromatography at 37°in 5% C02:air for 45 min. The nylon-passed cells were washed twice and counted. Parallel aliquots 'The abbreviations used are: TPA, 12-O-tetradecanoylphorbol-13-acetate; P(Bu>2,phorbol-12,13-dibutyrate; 4a-PDD, 4a-phorbol-12,13-didecanoate; IFN, in- terferon; NK, natural killer (cells); DMSO. dimethyl sulfoxide. MAY 1984 1857 Research. on January 3, 2016. © 1984 American Association for Cancer cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

[CANCER RESEARCH 44, 1857-1862, May 1984]
Activation of Cytotoxic Activity of Human Blood Lymphocytes by Tumor-promoting Compounds1
Oscar F. Ramos,2-3 Maria G. Masucci,2 and Eva Klein
Department of Tumor Biology, Karolinska Institutet, S-104 01 Stockholm, Sweden
ABSTRACT
Three categories of tumor promoters and chemically relatedbut inactive substances were tested for their effect on thecytotoxic activity of human blood lymphocytes against K562 andDaudi targets. Lymphocytes incubated overnight in the presenceof phorbol esters 12-0-tetradecanoylphorbol-13-acetate andphorbol-12,13-dibutyrate [P(Bu)2] had enhanced function. Incubation with 4-a-phorbol-12,13-didecanoate was without effect.
Enhancing activity was also exerted by the indole alkaloids,teleocidin and lyngbyatoxin A, and the polyacetates, aplysiatoxinand debromoaplysiatoxin, but not by dihydroteleocidin. Only thetumor-promoting compounds activated the cytotoxic potential.The substances acted in a dose-dependent manner with optimal
activity at characteristic concentrations.Overnight incubation of lymphocytes at 4°did not change their
spontaneous cytotoxicity but abolished the enhancing effect ofP(Bu)2. Thus, P(Bu)2-induced activation occurred only on meta-
bolically active cells. The activation did not require DNA synthesis. Similar to controls, the P(Bu)2-treated cells required divalent
cations and an intact cytoskeleton in order to perform their lyticfunction. Experiments with the various metabolic inhibitors indicate that phorbol ester treatment does not induce an alternativecytotoxic mechanism since, as with untreated lymphocytes,P(Bu)2-activated cells require contact with the target and intact
secretory functions.The enhanced cytolytic potential was not due to induction of
a-interferon (IFN-a) production, as shown by the fact that theeffect was not abolished by addition of anti-IFN antibodies during
the P(Bu)2 treatment of lymphocytes or during the cytotoxicassay. However, the presence of antiserum against IFN reducedthe cytotoxic potential of control cells, suggesting that endogenous IFN production contributes to the maintenance of lyticfunction in cultured cells. If this mechanism is counteracted byaddition of anti-IFN serum, the phorbol esters can provide analternative activation signal. When P(Bu)2-activated lymphocyteswere subsequently treated with IFN-a or IFN-y, their lytic capac
ity was further increased. These results indicate that P(Bu)2 andIFN activate cytotoxic potential through different pathways.
INTRODUCTION
Phorbol esters and similar diterpenes act as tumor promotersin mouse skin (8, 40). In order to understand the mechanism oftumor promotion, these compounds have been widely studied inin vitro systems. They were found to alter the growth behavior,
1This investigation was supported by USPHS Grant 5 R01 CA25250-04
awarded by the National Cancer Institute, Department of Health and HumanServices, and by the Swedish Cancer Society.
2 Recipients of fellowships from the Cancer Research Institute/Concern Foun
dation.3To whom requests for reprints should be addressed.
Received August 1,1983; accepted February 10,1984.
differentiation, and functional characteristics of cells (40). Theireffect is initiated by interaction with specific receptors expressedon the plasma membrane (9, 11, 33, 43). These effects werestudied extensively with phorbol esters like TPA4 (8, 40). Indole
alkaloids and polyacetates derived from Streptomyces and theblue-green alga Lyngbya majuscula are 2 further classes of tumor
promoters chemically unrelated to phorbol esters (10,13). Theyinteract with the same surface receptor as does TPA and inducesimilar biological effects (12, 39).
Tumor promoters modify the immune response and the functional behavior of lymphocytes (for review, see Ref. 24). Theyact as mitogens and enhance the action of other mitogens (15).They stimulate the production of IFN and lymphokines (27, 33,44, 45). They activate the cytotoxic potential of polymorphonu-
clear leukocytes (7), and under certain conditions both stimulatory and inhibitory effects on the tumoricidal capacity of TPA-
treated macrophages were reported (18, 20, 23).Phorbol esters modify the cytotoxic activity of lymphocytes.
They inhibit the generation of allospecific cytotoxic cells in mixed-lymphocyte cultures but activate the cytotoxic potential of pre-sensitized effectors (2, 26). Depending on the concentration, thelength of their interaction, and the presence of regulatory cells,tumor promoters can either enhance or suppress NK activity (forreview, see Ref. 19).
We have shown previously that the tumor promoters TPA andP(Bu)2 have a complex modulatory effect on human NK assaysbecause they can influence the behavior of both effector andtarget cells (30). Long-term incubation with nM concentrations of
the compounds resulted in activation of the lytic potential oflymphocytes. This effect was counteracted via activation of thesuppressive function of monocytes. The target sensitivity wasalso modified.
In the present study, we have tested the effect of the indolealkaloids teleocidin and lyngbyatoxin A and the polyacetatesaplysiatoxin and debromoaplysiatoxin on the human NK systemusing K562 and Daudi targets. In addition, we analyzed certaincharacteristics of the P(Bu)2-induced enhancement of the lytic
function with special attention to the possible involvement of IFNon the phenomenon.
MATERIALS AND METHODS
Experimental Design. Buffy coat leukocytes were obtained fromhealthy blood donors. Mononuclear cells were separated by the methodof Boyurn (4) on Ficoll-lsopaque gradients and were subsequently incubated in a plastic bottle (Falcon) at 37°in 5% CO2:air for 60 min in order
to deplete mononuclear adherent cells. Nonadherent cells were subjectedto a nylon wool column chromatography at 37°in 5% C02:air for 45 min.
The nylon-passed cells were washed twice and counted. Parallel aliquots
'The abbreviations used are: TPA, 12-O-tetradecanoylphorbol-13-acetate;
P(Bu>2,phorbol-12,13-dibutyrate; 4a-PDD, 4a-phorbol-12,13-didecanoate; IFN, in-terferon; NK, natural killer (cells); DMSO. dimethyl sulfoxide.
MAY 1984 1857
Research. on January 3, 2016. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

O. F. Ramos et al.
of effector cells were incubated overnight in medium (control) or in thepresence of different tumor-promoting agents as described previously(30). The effects of various metabolic inhibitors on the P(Bu)a-induced
NK activation were tested. The drugs were added to the cell suspensions30 min before the addition of PfBufe and left during the incubation period(18 hr) with the phorbol ester. The cells were then washed twice toremove P(Bu)2 and resuspended in a fresh culture medium containingthe respective drugs. The lytic potential was measured in a standard51Cr release assay. The viability of the effector cells before the cytotoxic
assay was more than 90%.Medium. RPMI 1640 was supplemented with 10% heat-inactivated
newborn calf serum (Gibco Europe U.K.), streptomycin (10 ¿ig/ml),penicillin (120 Mg/ml), and glutamine (2 mw).
Reagents. P(Bu>2,4«-PDD, and TPA were dissolved in DMSO andstored at -20° (0.2 mw). Teleocidin, lyngbyatoxin A, aplysiatoxin, and
debromoaplysiatoxin (10) were donated by Dr. Richard E. Moore, Department of Chemistry, University of Hawaii. The compounds weredissolved in DMSO and stored at -20°. The final concentration of DMSO
was always less than 0.05%. NaN3 (Merck, Darmstadt, Germany), 2-deoxyglucose (Sigma Chemical Co., St. Louis, MO), and 1-/3-o-arabino-
furanosylcytosine (Sigma) were diluted in medium and used at finalconcentrations of 10 mwi, 50 HIM, and 25 //g/ml, respectively. Cytochal-
asin B (Sigma), colchicine (Sigma), and monensin (gift of Dr. Hans Wigzell,Karolinska Institutet, Stockholm, Sweden) were dissolved in DMSO andused at final concentrations of 20 ^M, 10 MM,and 1 Mg/ml, respectively.EDTA (KEBO, Stockholm, Sweden) was diluted in H2O and used at 10ITÃŒM.
IFN and Anti-IFN Treatment. Parallel aliquots of 106 lymphocytes
were incubated for the indicated lengths of time in control medium or inthe presence of 30 nw P(Bu>2. To one aliquot, 103 units IFN-a or IFN-7
per ml were added during the last 3 hr before the cytotoxic assay. Thecells were washed before their use as effectors [IFN-a (specific activity,106 units/mg protein) was a kind gift of Dr. Kari Cantell, Helsinki, Finland;IFN-7 (specific activity, 5 x 105 units/mg protein) was a gift of Dr. E.
Falcof, Paris, France).Sheep anti-IFN-« serum (4.5 x 105 neutralizing units/ml) was a gift of
Dr. Kari Cantell, Helsinki, Finland. Different concentrations of the anti-
serum were added to the cell suspension 15 min before the addition ofP(Bu)2 or IFN.
Cytotoxic Assay. Techniques for the target radiolabeling and cytotoxic assays have been described previously (25, 30). K562 and Daudicells were incubated with sodium [51Cr]chromate (100 ¿iCi/ml;NewEngland Nuclear, Boston, MA) for 1 hr at 37°in 5% CO2:air. Labeledtargets (4 x 103) in 20 p\ were mixed with 100 M!of triplicated dilutions
of effector lymphocytes to obtain effector:target ratios of 25:1, 8:1, and
3:1. After 4 hr of incubation at 37°,in 5% CO2:air, 60 n\ of the superna-
tants were collected, and their radioactivity was determined in a -y-
counter. The lytic effect was determined by the formula:
% of specific 51Cr release
_ Experimental release - spontaneous release
Total incorporation - spontaneous release
The results are expressed as lytic units/106 cells or percentage of specific51Cr release. The number of lytic units from each population was calcu
lated from the dose-response curve and represents the number of killer
cells for 15% lysis against K562 and Daudi. The significance of differences between the mean values of triplicate tests was analyzed by theStudent f test.
RESULTS
Effect of Tumor Promoters on the Cytotoxic Activity ofLymphocytes
Tumor promoters and structurally related nonactive compounds were tested (Table 1). In accordance with our previousresults (30), overnight pretreatment with the phorbol esters TPAand P(Bu)2elevated the cytotoxic potential of lymphocytes testedagainst K562 and Daudi. The nonpromoter phorbol ester 4a-
PDD did not enhance the cytotoxicity.Pretreatment with the tumor-promoting alkaloids teleocidin
and lyngbyatoxin A also activated the cytotoxic function. Dihy-
droteleocidin, a weak tumor promoter, was not active (10). Athird class of tumor promoters, the polyacetates, aplysiatoxinand debromaplysiatoxin, also showed an enhancing effect. Eachcompound had a characteristic dose response (Chart 1).
Effect of Metabolic Inhibitors on the Cytotoxic Activity ofLymphocytes
Overnight incubation of the effector cells at 4°did not affect
the subsequently tested lytic activity (Chart 2). However, at lowtemperatures, the P(Bu)2-induced activation did not occur. Since
binding of P(Bu)2 to membrane receptors is shown to occur at4°,it is likely that cellular metabolism is required for the P(Bu)2-
enhancing effect (9, 33, 43).Inhibition of DNA synthesis. The presence of 1-0-o-arabino-
TaWelEffect of different tumor-promotingcompounds on the cytotoxic activity of human lymphocytes againstK562 and Daudi targets
Aliquots (108/ml)of lymphocytes were incubated overnight in medium (control) or in the presenceof the indicated compounds.
Cytotoxic activity (lytic units'/I 0" cells) in 4-hr 51Crreleaseassay
Phorbolesters Indolealkaloids Polyacetates
Experi-TargetmentK562
12345Daudi
12345Control90
±10°155
±5108±1072±890
±1314±
612±223±116±
412± 24a-PDD(30
HM)68
±4(NS)96±10(IMS)18
±2(NS)10± 5(NS)TPA
(30nw)310±10(S)C625
±25(S)600±30(S)238±7(S)403±27(S)63
±7(S)49±10(S)147±10(S)120
±10(S)312±28(S)P(Bu>2
(30nw)166
±250±300±171±166±46
±30±138±100±50
±14(S)10(S)5(S)4(S)4(S)4(S)4(S)7(S)8(S)6(S)Dihydro-
teleocidin(15ng/ml)75
±5(NS)83±17(NS)17
±3(NS)10± 3(NS)Teleocidin
(15ng/ml)313
±17(S)278±2(S)300±10(S)186±14(S)294
±12(S)59
±3(S)41± 4(S)93±7(S)114±6(S)58± 3(S)Lyngbyatoxin
A (15ng/ml)192
±5(S)417±8(S)416±24(S)155
±5(S)390±30(S)43
±4(S)39±5(5)98±2(S)90.t8(S)74± 6(S)Aplysiatoxin
(15ng/ml)190
±10(S)227±3(S)834±36(S)157±7(S)208±20(S)62
±8(S)47±3(S)90±5(S)50±4(S)66± 6(S)Debromo
aplysiatoxin(250ng/ml)150
±362±714±134±166
±36
±30±50±60±53
±5(S)8(S)36
(S)6(S)4(S)3(S)2(S)2(S)5(S)7(S)
aOne lytic unit corresponds to the number of effector cells required for 15% lysis of target.6 Mean ±S.D. of triplicate samples.c S, significantlydifferent from control (p < 0.01); NS, nonsignificant.
1858 CANCER RESEARCH VOL. 44
Research. on January 3, 2016. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

Tumor Promoter Induction of NK Activation
TO
50
30
S 100)
«C
O
50
30
10
Aplysiatoxin Debromoaplysiatoxin P(Bu)2
O 1 2.5 5 10 20 40 80 120 O 51 62 125 250 5O) 1000 O 15 30 60 120 240 Dose ng/ml
Te i eoe id i n Lyngbyatoxm A TRA
O 1 2.5 5 10 20 40 80 120 O 1 25 5 10 20 40 80 120 O 15 30 60 120 240 Dose ng/ml
Chart 1. Dose-response curves of the enhancing effect of different tumor promoting agents on the cytotoxic activity of blood lymphocytes. Effector cells wereincubated with the compounds for 18 hr. Effectortarget ratio, 25:1, Target, K562. Bars, S.D. of triplicate samples.
200
160-
120
80
40
-L.3_L7*r-ill4*JL_I_
3 1
Experiments
Chart 2. Effect of temperature on the potentiation of cytotoxicity by PfBufe. Thelymphocytes were incubated for 18 hr in medium (D) or in the presence of 30 nwP(Bu)j (B). Preincubation was performed at 37°or 4°.Target, K562. The resultsare expressed in lytic units (LU)/108 cells. Bars, S.D. of triplicate samples.
furanosylcytosine (25 /¿g/ml)during the preincubation of lymphocytes did not alter their cytotoxic function. Thus, DNA synthesisis not needed either for the cytotoxic process or for the activationby phorbol (Chart 3).
Inhibitors of Cell Respiration and Glucose Metabolism.Inhibition of cytochrome function by 10 mw NaN3 abolished
Control
Ara. C
LU/106
4.0 80 120 160 2OO
t"
Chart 3. Effect of inhibition of DNA synthesis on the potentiation of cytotoxicityby P(Bufe. Control (O) and PfBu^treated (•)cells were incubated in the presence
J^ of 1-/3-D-arabinofuranosylcytosine (X>ra.C). LU, lytic units.
completely the lytic capacity of control and phorbol-treated effectors (Chart 4). Inhibition of glucose metabolism by 50 HIM 2-
deoxyglucose gave partial reduction.Inhibitors of Microtubules, Microfilaments, and the Secre
tory Apparatus. Complete inhibitionof cytotoxicity was obtainedby addition of 20 ¿¿Mcytochalasin B (Chart 5). At this concentration, the drug disrupts microfilament structures (16). Chelatingagents also inhibited the lysis. Inhibition of microtubule assemblyachieved by 10 UMcolchicine reduced the lytic activity of controland P(Bu)2-activated lymphocytes. The addition of monensin, a
carboxylic ionophore which prevents release of lysosomal enzymes, also abrogated the cytotoxicity (Table 2) (5,16).
Role of IFN in P(Bu)2-induced Activation of Lytic Potential
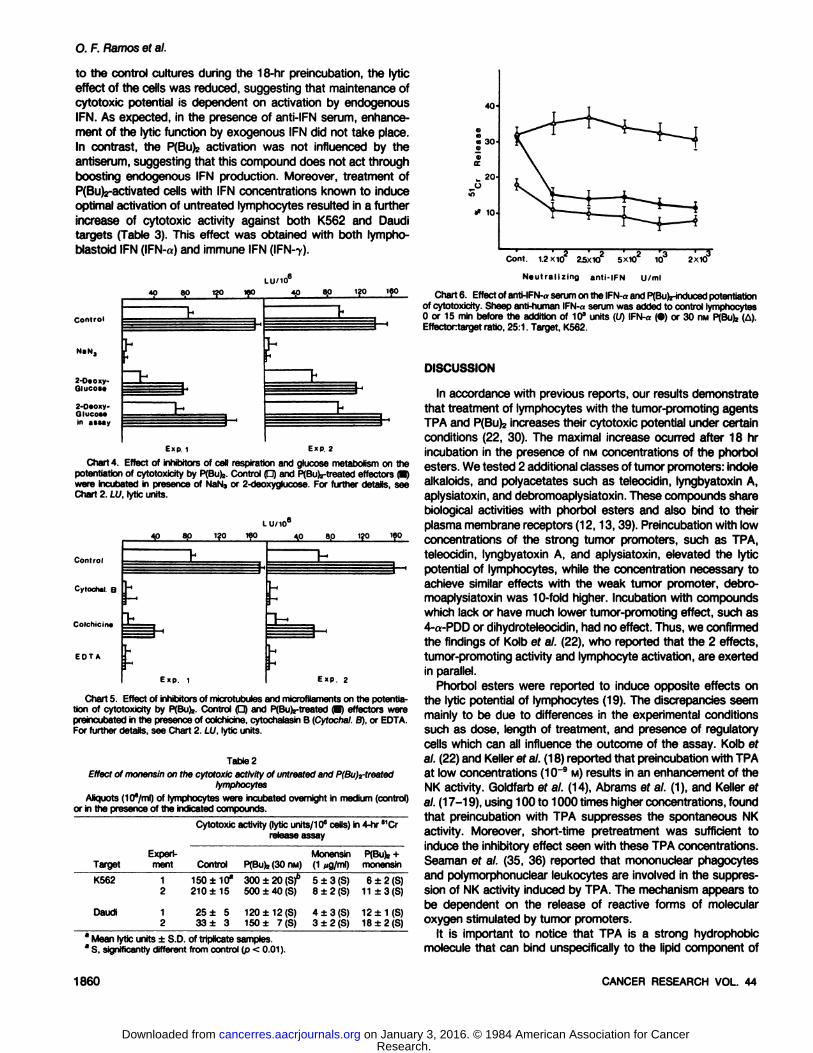
The effector cells were pretreated with P(Bu)2 in the presenceof anti-IFN-a serum (Chart 6). When anti-IFN serum was added
MAY 1984 1859
Research. on January 3, 2016. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
O. F. Ramos et al.
to the control cultures during the 18-hr preincubation, the lytic
effect of the cells was reduced, suggesting that maintenance ofcytotoxic potential is dependent on activation by endogenousIFN. As expected, in the presence of anti-IFN serum, enhance
ment of the lytic function by exogenous IFN did not take place.In contrast, the P(Bu)2 activation was not influenced by theantiserum, suggesting that this compound does not act throughboosting endogenous IFN production. Moreover, treatment ofP(Bu)2-activated cells with IFN concentrations known to induce
optimal activation of untreated lymphocytes resulted in a furtherincrease of cytotoxic activity against both K562 and Dauditargets (Table 3). This effect was obtained with both lympho-blastoid IFN (IFN-a) and immune IFN (IFN-T).
Control
4080H1fOLU/10 ICO 4,08.0 1?0t160
Exp 1 E«P 2
Chart 4. Effect of inhibitors of cell respiration and glucose metabolism on thepotentiation of cytotoxicity by PfBufc.Control (D) and PfBufe-treatedeffectors P)were incubated in presence of NaN3or 2-deoxyglucose. For further details, seeChart 2. LU, lytic units.
LU/10°
ControlCytochal
B
ColchicineEDTA4.0
8.0 120160HI:Exp.
14,0
8.0 120160HtExp.
2
Chart 5. Effect of inhibitors of microtubules and microfilamentson the potentiation of cytotoxicity by P(Bu)j. Control P) and P(Bu)z-treated(P effectors werepreincubated in the presenceof colchicine, cytochalasinB (Cytochal.B), or EDTA.For further details, see Chart 2. LU, lytic units.
Table 2Effect of monensin on the cytotoxic activity of untreatedand P(Bu)2-treated
lymphocytesAliquots (10°/ml)of lymphocytes were incubated overnight in medium (control)
or in the presenceof the indicated compounds.Cytotoxic activity (lytic units/106cells) in 4-hr '
releaseassay'Cr
TargetK562DaudiExperiment121
2Control150
±10*
210±1525
± 533 ± 3P(Bu),
(30nM)300±20 (S)6
500 ±40(S)120
±12 (S)150 ± 7(S)Monensin
(1„g/ml)5
±3 (S)8 + 2(S)4
±3 (S)3 ±2 (S)P(Bufe
+monensin6
±2 (S)11±3(S)12
±1(S)18±2(S)
40
•Ma 30«O
1C
w 20JJZ
# 10-
8 Mean lytic units ±S.D. of triplicate samples.a S, significantlydifferent from control (p < 0.01).
Com. 1.2X10 25X102 5X102 103 2X10
Neutralizing anti-IFN U/ml
Chart 6. Effect of anti-IFN-aserumon the IFN-aand PfBufe-inducedpotentiationof cytotoxicity. Sheep anti-human IFN-«serum was added to control lymphocytes0 or 15 min before the addition of 10" units (U) IFN-a (•)or 30 nw PfBufe(A).Effectortarget ratio, 25:1. Target, K562.
DISCUSSION
In accordance with previous reports, our results demonstratethat treatment of lymphocytes with the tumor-promoting agents
TPA and P(Bu)2 increases their cytotoxic potential under certainconditions (22, 30). The maximal increase ocurred after 18 hrincubation in the presence of nM concentrations of the phorbolesters. We tested 2 additional classes of tumor promoters: indolealkaloids, and polyacetates such as teleocidin, lyngbyatoxin A,aplysiatoxin, and debromoaplysiatoxin. These compounds sharebiological activities with phorbol esters and also bind to theirplasma membrane receptors (12,13,39). Preincubation with lowconcentrations of the strong tumor promoters, such as TPA,teleocidin, lyngbyatoxin A, and aplysiatoxin, elevated the lyticpotential of lymphocytes, while the concentration necessary toachieve similar effects with the weak tumor promoter, debromoaplysiatoxin was 10-fold higher. Incubation with compoundswhich lack or have much lower tumor-promoting effect, such as4-a-PDD or dihydroteleocidin, had no effect. Thus, we confirmed
the findings of Kolb ef a/. (22), who reported that the 2 effects,tumor-promoting activity and lymphocyte activation, are exerted
in parallel.Phorbol esters were reported to induce opposite effects on
the lytic potential of lymphocytes (19). The discrepancies seemmainly to be due to differences in the experimental conditionssuch as dose, length of treatment, and presence of regulatorycells which can all influence the outcome of the assay. Kolb etal. (22) and Keller ef al. (18) reported that preincubation with TPAat low concentrations (10~9 M) results in an enhancement of the
NK activity. Goldfarb ef al. (14), Abrams ef al. (1), and Keller efal. (17-19), using 100 to 1000 times higher concentrations, found
that preincubation with TPA suppresses the spontaneous NKactivity. Moreover, short-time pretreatment was sufficient to
induce the inhibitory effect seen with these TPA concentrations.Seaman ef al. (35, 36) reported that mononuclear phagocytesand polymorphonuclear leukocytes are involved in the suppression of NK activity induced by TPA. The mechanism appears tobe dependent on the release of reactive forms of molecularoxygen stimulated by tumor promoters.
It is important to notice that TPA is a strong hydrophobicmolecule that can bind unspecifically to the lipid component of
1860 CANCER RESEARCH VOL. 44
Research. on January 3, 2016. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

Tumor Promoter Induction of NK Activation
TableaAdditive effect of IFN and P(Bu)¡pretreatment on the cytotoxic activity of blood lymphocytes
Parallel aliquota (10"/ml) of lymphocytes were incubated for 18 hr in control medium, 30 nw PfBufe, or IFN (103 units/
ml) alone or in combination.Cytotoxic activity (lytic units/10*cells)TargetK562DaudiExperi
ment12341234Control120±8a84
±4257±10100
±914
±511±315±
520± 6PfBufc240
±5(Sf168±8(S)750±10(S)300±15(S)80
±8(S)65±5(S)85±5(S)92± 8(S)IFN-a180±10(S)140
±7(S)700±6(S)250±12(S)76
±6(S)61± 3(S)115±7(S)150±10(S)IFN-a31
2 ±8(S)C220±10(S)C840±15(S)C920
±6(S)C202+
4(S)C172±8(S)C183+8(S)C400±15(S)CIFN-7168+
7(S)120±7(S)660±16(S)450±14(S)91
± 8(S)86±4(S)90±7(S)170±10(S)IFN-y342
±8(S)C250±9(S)C900±12(S)C830
±8(S)C150±10(S)C140+
9(S)C146±4(S)C450
±11(S)°
3 Mean lytic units ±S.D. of triplicate samples.6 S, significantly different from controls (p < 0.01).c Significantly different from P(Bu)j- or IFN-treated samples (p < 0.01).
the cell membrane (6,9, 33). It is therefore possible that inhibitionof NK activity observed with high TPA doses may be due tononspecific membrane changes (6, 9). P(Bu)2, the compoundthat we used for characterization of the effect on cytotoxicactivity, is less hydrophobic than TPA. Using P(Bu)2, Driedger efa/. (9) and Sando ef al. (33) could demonstrate specific phorbolester receptors on the cell membrane. Saturation of P(Bu)2-
specific binding occurred at 30 MMand was reversible in mediafree of phorbol esters. This dose was found to be optimal foractivation of lytic potential (30).
The rapidity of the enhancement of lytic potential suggeststhat it is not due to alteration of the cell population with subsequent enrichment of certain cell types (22, 30). In accordance,the present experiments show that the DMA synthesis inhibitor,1-0-D-arabinofuranosylcytosine did not abrogate it. The enhancement of lytic potential induced by P(Bu>2seems to be an activemetabolic process since it occurs only in cells incubated at 37°
and is blocked by inhibitors of cell respiration and glucosemetabolism. The information available on the metabolism of TPAindicates that metabolic degradation of the compound is notrequired for its biological activity (3). Its effects are mediated bybinding to specific membrane receptors (9, 12). The metabolicprocesses activated by phorbol receptor binding are, however,still unknown.
The phenomenon of target lysis can be separated in differentsteps (32). The first involves establishment of contact betweeneffectors and targets via the plasma membranes. Relevant tothe elevation of cytotoxic potential is the finding that phorbolesters and other promoters alter the cell membrane of lymphocytes with conspicuous morphological effect (29). Moreover, thetreated lymphocytes aggregate and attach to cells derived fromthe same species (28, 29). The sialic acid content of the treatedcells decreases, which leads to a lower negative surface charge(28, 29, 40). This charge alteration facilitates intercellular contactformation.
Contact between effectors and targets requires divalent cations and intact microfilaments (16, 31, 41). Similar to that in thecontrols, the cytotoxic effect of P(Bu)2-treated cells was inhibited
by EDTA and cytochalasin B. Microtubules are necessary forcytoskeleton organization and cellular secretion, and they wereshown to be involved in the mechanism of cell-mediated cytotox-
icity (5,16, 31, 42). Intact secretory apparatus is also necessaryfor the cytotoxic function of P(Bu)2-treated effector cells as
shown by the inhibitory effect of monensin. The enhancement ofcytotoxicity may occur through action on secretory organellessince phorbol esters were shown to induce the release of lyso-
somal enzymes (34).The experiments with anti-IFN-a serum suggest that the lytic
capacity of cultured lymphocytes requires endogenous IFN production. Since the P(Bu)2-treated cells did not change their
behavior when they were exposed to this antiserum, the phorbolesters do not appear to activate cytotoxicity through inductionof IFN production. Our results are therefore in accordance withthe finding that IFN was not detected in the supernatant ofphorbol ester-treated mouse lymphocytes (22). It remains to beseen whether other lymphokines, such as IL-2, contribute to theenhancement of lytic function. IL-2 was shown to activate lym
phocytes for cytotoxicity and to be induced by P(Bu)2 (22, 24,38,45).
Combined treatment of lymphocytes with phorbol esters andIFN resulted in additional effects. Both compounds were shownto enhance the lytic potential of low-density lymphocytes, which
contain high proportions of NK cells (14, 21, 22, 30). It is stillpossible that within this subset different cells respond to the 2stimuli. It is more likely, however, that the same cells are influenced but potentiations occur through different pathways. Thiswas suggested by Kolb ef al. (22) and Senik ef al. (37) on thebasis of the need of RNA and protein synthesis for the IFN butnot for phorbol ester activation of cytotoxicity.
In conclusion, PCBufeand other tumor promoters can, undercertain conditions, increase the cytolytic activity of human bloodlympohcytes. The activation does not occur through IFN production. The cytolytic function of activated cells requires contactwith the target and an intact secretory mechanism. The experiments suggest that an alternative lytic mechanism does notseem to be generated by phorbol ester treatment. On the otherhand, the pathway of activation seems to be different for IFNand phorbol esters.
REFERENCES
1. Abrams, S. I., Bray, R. A., and Brahmi, Z. Mechanism of action of phorbolmyristate acetate on humannatural killer cell activity. Cell. Immunol.,80:230-240, 1983.
2. Andreotii, P. E. Phorbolester tumor promoter modulationof altoantigenspecificT lymphocyte responses.J. Immunol., 729: 91-96,1982.
3. Berry, D. L, Bracken, W. M., Fischer,S. M., Viaje,A., and Slaga,T. Metabolicconversion of 12-O-tetradecanoylphorbol-13-acetatein adult and newborn
MAY 1984 1861
Research. on January 3, 2016. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

0. F. Ramos et al.
mouse skin and mouse liver microsomes. Cancer Res., 38: 2301-2306,1978.4. Boyurn, A. Isolationof mononuclearcells and granulocytes from human blood.
J. Clin. Lab. Invest. Suppl., 21: 77-81, 1968.5. Carpen, 0., Virtanen, I., and Saksela, E. Ultrastructure of human natural killer
cells: nature of the cytolytic contacts in relation to cellular secretion. J.Immunol., 128: 2691-2697,1982.
6. Cooper, R. A., Braunwald, A. D., and Kuo, A. L. Phorbol esters induction ofleukemiacell differentiationis a membranemediatedprocess. Proc. Nati.Acad.Sci. USA, 79: 2865-2869,1982.
7. Dallegri,F., Patrone, F., Bonvini, E., Gahrton, G., Holm, G., and Sacchetti, C.Ox erythrocyte cytotoxicity by phorbol myristate acetate-activated humanneutrophils.Scand. J. Immunol., 17: 109-114,1983.
8. Diamond, L., O'Brien, T. G., and Baird, W. M. Tumor promoters and themechanismof tumor promotion. Adv. Cancer Res., 32: 1-75, 1980.
9. Driedger, P. E., and Blumberg, P. M. Specific binding of phorbol ester tumorpromoters. Proc. Nati. Acad. Sci. USA, 77: 567-571, 1980.
10. Eliasson, L., Kallin, B., Patarroyo, M., Klein, G., Fujiki, H., and Sugimura, T.Activation of the EBV-cycle and aggregation of human blood lymphocytes bythe tumor promoters teleocidin. lyngbyatoxin A, aplysiatoxin and debromoa-plysiatoxin. Int. J. Cancer,37: 7-11, 1983.
11. Estensen, R. D., Dehood, D. K., and Cole, C. Binding of (3H)12-O-tetradeca-noylphorbol-13-acetateto intact human peripheralblood lymphocytes. CancerRes., 40: 1119-1124,1980.
12. Esumi, M., and Fujiki, H. Spingomyelin is a possible constituent of bindingsites for the tumor promoter phorbol ester, indole alkaloids and polyacetates.Biochem. Biophys. Res. Commun., 772: 709-716,1983.
13. Fujiki, H., Mori, M., Nakayasu, M., Terada, M., Sigimura, T., and Moore, R.IndolealkaloidsdehydroteleocidinB, teleocidinand lyngbyatoxinA as membersof a new class of tumor promoters. Proc. Nati. Acad. Sci. USA, 78: 3872-3876, 1981.
14. Goldfarb, R. H., and Herberman, R. B. Natural killer cell reactivity: regulatoryinteractions among phorbol esters, Interferon, cholera toxin and retinoid acid.J. Immunol., 726: 2129-2135,1981.
15. Kabelitz, D., Tötterman,T. H., Gidlund, M., Nilsson, K., and Wigzell, H.Activation of humanT lymphocytes by ^-O-tetradecanoylphorbol-IS-acetate:role of accessory cells and interaction with lectins and allogeneic cells. Cell.Immunol., 70: 277-286, 1982.
16. Katz, P., Zaytoun, A. M., and Lee, L. H. Mechanism of human cell mediatedcytotoxicity: III. Dependenceof natural killingon microtubuleand microfilamentintegrity. J. Immunol., 729: 2816-2825, 1982.
17. Keller, R. Suppression of natural antitumor defense mechanism by phorbolesters. Nature (London.),282: 729-730,1979.
18. Keller,R. Tumor promoting diterpeneesters inducemacrophagedifferentiationbut prevent activation for tumoricidal activity of macrophagesand NK cells. In:R. B. Herberman (ed.), NK Cells and other Natural Effector Cells, pp. 601-604. New York: Academic Press, Inc., 1982.
19. Keller, R. Host defense mechanism against tumor as the principal targets oftumor promoters. J. Cancer Res. Clin. Oncol., 705: 203-211,1983.
20. Keller, R., Keist, R., Adolf, W., Opferkuch, H. J., Schmidt, R., and Hecker, E.Tumor promoting diterpene esters prevent macrophage activation and suppress macrophage tumoricidal capacity. Exp. Cell. Biol., 50: 121-134,1982.
21. Klein, E. Natural and activated cytotoxic T lymphocytes. Immunol. Today, 7:IV-VI, 1980.
22. Kolb, J. D., Senik, A., and Castagna, M. In vitro stimulation of mouse NK cellactivity by phorbol esters: a pathway different from interferon induced activation. Cell. Immunol.,65: 258-271,1981.
23. Laskin, D. L., Laskin, J. D., Kessler, F. K., Weinstein, I. B., and Charchman,R. A. Enhancement of macrophage induced cytotoxicity by phorbol estertumor promoters. Cancer Res., 41: 4523-4528,1981.
24. Mastro, A. M. Phorbolesters: tumor promotion, cell regulationand the immuneresponse. In: S. B. Mizel (éd.),Lymphokines,Vol. 6, pp. 263-313. New York:Academic Press, Inc., 1982.
25. Masucci, M. G., Klein, E., and Argov, S. Disappearanceof NK effect after
explanation of lymphocytes and generationof similar non-specificcytotoxicitycorrelates to the level of blastogenesis in activated cultures. J. Immunol., 724:2458-2563, 1980.
26. Munger, W. E., and Lindquist, R. R. Effect of phorbol esters on alloimmunecytolysis. Cancer Res., 42: 5023-5029, 1982.
27. Mizel,Z.B., Rosenstreich,D. L., and Oppenheim,J. J. Phorbolmyristic acetatestimulates LAPproduction by the macrophagescell line P388D.Cell. Immunol.,40:230-237,1978.
28. Patarroyo, M., Biberfeld, P., Klein, E., and Kelin, G. Phorbol 12,13-dibutyrate(P(Bu>2)treted human blood mononuclear cells bind to each other. Cell.Immunol., 75:144-150, 1983.
29. Patarroyo, M., Yogeeswaran, G., Biberfeld, P., Klein, E., and Klein,G. Effectsof phorbol-12-13-dibutyrateP(Bu)j on human blood lymphocytes: induction ofmorphologicalchanges, cell to cell aggregation and biochemicalalterations ofthe cell membranecomposition. Int. J. Cancer,30: 707-714,1982.
30. Ramos, O. F., Masucci, M. G., Bejarano, M. T., and Klein, E. The tumorpromoter phorbol-12-13-dibutyrate (P(Bu)z)stimulates cytotoxic activity ofhuman blood lymphocytes. Immunobiology, 765:403-414, 1983.
31. Roder, J. C., Argov, S., Klein, M., Petersson, C., Kiessling, R, and Hansson,M. Target effector interaction in the natural killer cell system. V. Energyrequirements, membrane integrity and the possible involvementof lysosomalenzymes. Immunology,40: 107-115,1980.
32. Roder, J. C., Kiessling, R., Biberfeld, P., and Andersson, B. Target effectorinteraction in the natural killer system. II. The isolation of NK cells and studieson the mechanismof killing. J. Immunol.,75: 144-152, 1983.
33. Sando, J. J., Hilfiker, M. L., Salomon, D. S., and Parrar,J. J. Specific receptorfor phorbol ester in lymphoid cell population: role in enhanced production ofcell growth factor. Proc. Nati. Acad. Sci. USA, 78. 1189-1193, 1981.
34. Schleimer, R. P., Gillespie,E., Daiuta, R., and Lichtenstein, L. M. Releaseofhistaminefrom human leukocytes stimulatedwith the tumor promotingphorboldiesters. II. Interaction with other stimuli. J. Immunol., 728: 136-141,1982.
35. Seaman, W. E., Gindhart, T. D., Blackman, M. A., Dalai, B., Talal, N., andWerb, Z. Natural killing of tumor cell by human peripheralblood cells. J. Clin.Invest., 67: 1324-1333,1981.
36. Seaman, W. E., Gindhart, T. D. Blackman, M. A., Dalai, B., Talal, N., andWerb, Z. Suppressionof natural killing in vitro by monocytes and polymorpho-nuclear leukocytes. J. Clin. Invest., 69: 876-888, 1982.
37. Senik, A., Kolb, J. P., Om, A., and Gidlund. M. Study of the mechanismfor invitro activation of mouse NK cells by IFN. Scand. J. Immunol., 72: 51-60,1980.
38. Suzuki, R., Handa, K., Itoh, K., and Kumagai, K. Natural killer cells (NK) asresponder to IL-2.1. Proliferativeresponse and establishment of cloned cells.J. Immunol., 730: 981-987, 1983.
39. Umezawa, K., Weinstein, B., Horowitz, A., Fujiki, H., Matsushima, T., andSigimura, T. Similarity of teleocidin B and phorbol esters tumor promoter ineffect on membrane receptors. Nature (Lond.),290: 411-413, 1981.
40. Weinstein, I. B., Lee, L. S., Fischer,P. B., Mufson, A., and Yamasaki,K. Actionof phorbol ester in cultures: mimicry of transformation, altered differentiationand effects on cell membranes.J. Supramol. Struct., 72: 195-205, 1979.
41. Wessels, N. K., Spooner, B. S., Ash, J. F., Bradley, M. O., Luduena, M. A.,Taylor, L., Wrenn, J. J., and Yamada, K. M. Microfilaments in cellular anddevelopmentalprocesses. Science (Wash. DC), 777: 135-144, 1971.
42. Wilson, L., and Bryan, J. Biochemicaland pharmacology properties of micro-tubules. Adv. Ce.. Mol. Biol., 3: 21-50, 1974.
43. Yamasaki, H., Drevon, C., and Martel, N. Specific bindingof phorbol esters toFriend erythroleukemia cells—generalproperties, down regulation and relationship to cell differentiation. Carcinogenesis(Lond.),3: 905-910, 1982.
44. Yip, Y. K., Barrowclough, B. S., Urban, C., and Vilcek, J. Purification of twosubspecies f human gamma (immune)¡nterferon.Proc. Nati. Acad. Sci. USA,79: 1820-1824, 1982.
45. Yip, Y. K., Kelker, H. C., Stone-Wolff, D. S.. Pearistein, K. Urban, C., andVilcek, J. Stimulationof lymphokineproduction by teleocidin,asplysiatoxinanddebromoaplysiatoxin.Cell. Immunol.,79: 389-395. 1983.
1862 CANCER RESEARCH VOL. 44
Research. on January 3, 2016. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

1984;44:1857-1862. Cancer Res Oscar F. Ramos, Maria G. Masucci and Eva Klein by Tumor-promoting CompoundsActivation of Cytotoxic Activity of Human Blood Lymphocytes
Updated version
http://cancerres.aacrjournals.org/content/44/5/1857
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on January 3, 2016. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
Related Documents











