1 ACTIVATION OF BACTERIAL THERMOALKALOPHILIC LIPASES IS SPURRED BY DRAMATIC STRUCTURAL REARRANGEMENTS César Carrasco-López ¶ , César Godoy ║ , Blanca de las Rivas ║ , Gloria Fernández-Lorente † , José M. Palomo ║ , José M. Guisán ║ , Roberto Fernández-Lafuente ║ , Martín Martínez-Ripoll ¶ and Juan A. Hermoso ¶ * From the ¶ Grupo de Cristalografía Macromolecular y Biología Estructural, Instituto de Química-Física Rocasolano, CSIC, Serrano 119, 28006-Madrid, Spain, the ║ Departamento de Biocatálisis. Instituto de Catálisis. CSIC. Campus Universidad Autonoma. Cantoblanco. 28049-Madrid. Spain, and the † Departamento de Microbiología. Instituto de Fermentaciones Industrial, CSIC, Juan de la Cierva 3, 28006-Madrid, Spain. Running head: Activation Mechanism of Thermoalkalophilic Lipases Address correspondence to: Juan A. Hermoso, Grupo de Cristalografía Macromolecular y Biología Estructural, Inst. Rocasolano, CSIC, Serrano, 119, 28006 Madrid, SPAIN. Tel: 34-915619400; Fax:34- 915642431; E-mail: [email protected] The bacterial thermoalkalophilic lipases that hydrolyze saturated fatty acids at 60–75 °C and pH 8–10, are grouped as the lipase family I.5. We report here the crystal structure of the lipase from Geobacillus thermocatenulatus, the first structure of a member of the lipase family I.5 showing an open configuration. Unexpectedly, enzyme activation involves large structural rearrangements of around 70 amino acids and the concerted movement of two lids, the α6 and α7 helices, unmasking the active site. Central in the restructuring process of the lids are both the transfer of bulky hydrophobic residues out of the N-terminal end of the α6 helix, and the incorporation of short side chain residues to the α6 C-terminal end. All these structural changes are stabilized by the Zn ++ binding domain, which is characteristic of this family of lipases. Two detergent molecules are placed in the active site, mimicking chains of the triglyceride substrate, demonstrating the position of the oxyanion hole and the three pockets that accommodate the sn-1, sn-2 and sn-3 fatty acids chains. The combination of structural and biochemical studies indicate that the lids opening is not mediated by temperature but triggered by interaction with lipid substrate. Lipases (triacylglycerol lipase EC 3.1.1.3) catalyse the hydrolysis of long-chain triacylglycerides at water/oil interfaces. They are very versatile enzymes because, in vitro, they catalyse both the hydrolysis and the synthesis of a great variety of esters (1,2), the hydrolysis and transesterification of triacylglycerols (3) or the resolution of racemic mixtures (4). Most lipases contain a lid domain controlling access to the active site (5). The interaction of the enzyme with lipid aggregates induces the displacement of the lid, which makes the active site accessible to individual substrate molecules and increases the catalytic activity. This phenomenon is known as interfacial activation (5, 6). Much of the interest in extremophiles stems from their surprising properties. There has been extensive research on the structural proteins and key metabolic enzymes that are responsible for the organisms’ unusual properties. Recent research has focused on the identification of extremozymes relevant for industrial biocatalysis. The bacterial thermoalkalophilic lipases are among the most biocatalytically relevant extremozymes due to their resistance to proteases, detergents and chaotropic agents together with their extreme stability at elevated temperatures and in organic solvents (7). Thermoalkalophilic lipases are found from several of thermophilic aerobic Bacteria recently reclassified as the new genus Geobacillus (8), Geobacillus stearothermophilus (9), Geobacillus thermocatenulatus (10), Geobacillus thermoleovorans (11), and Geobacillus sp. TP10A. (12). Thermoalkalophilic http://www.jbc.org/cgi/doi/10.1074/jbc.M808268200 The latest version is at JBC Papers in Press. Published on December 3, 2008 as Manuscript M808268200 Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on August 28, 2016 http://www.jbc.org/ Downloaded from by guest on August 28, 2016 http://www.jbc.org/ Downloaded from by guest on August 28, 2016 http://www.jbc.org/ Downloaded from by guest on August 28, 2016 http://www.jbc.org/ Downloaded from by guest on August 28, 2016 http://www.jbc.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
ACTIVATION OF BACTERIAL THERMOALKALOPHILIC LIPASES IS SPURRED BY
DRAMATIC STRUCTURAL REARRANGEMENTS
César Carrasco-López¶, César Godoy║, Blanca de las Rivas║, Gloria Fernández-Lorente†, José M.
Palomo║, José M. Guisán║, Roberto Fernández-Lafuente║, Martín Martínez-Ripoll¶ and Juan A.
Hermoso¶*
From the ¶Grupo de Cristalografía Macromolecular y Biología Estructural, Instituto de Química-Física Rocasolano, CSIC, Serrano 119, 28006-Madrid, Spain, the ║ Departamento de Biocatálisis. Instituto de Catálisis. CSIC. Campus Universidad Autonoma. Cantoblanco. 28049-Madrid. Spain, and the †Departamento de Microbiología. Instituto de Fermentaciones Industrial, CSIC, Juan de la Cierva 3, 28006-Madrid, Spain.
Running head: Activation Mechanism of Thermoalkalophilic Lipases Address correspondence to: Juan A. Hermoso, Grupo de Cristalografía Macromolecular y Biología Estructural, Inst. Rocasolano, CSIC, Serrano, 119, 28006 Madrid, SPAIN. Tel: 34-915619400; Fax:34-915642431; E-mail: [email protected] The bacterial thermoalkalophilic lipases that hydrolyze saturated fatty acids at 60–75 °C and pH 8–10, are grouped as the lipase family I.5. We report here the crystal structure of the lipase from Geobacillus thermocatenulatus, the first structure of a member of the lipase family I.5 showing an open configuration. Unexpectedly, enzyme activation involves large structural rearrangements of around 70 amino acids and the concerted movement of two lids, the α6 and α7 helices, unmasking the active site. Central in the restructuring process of the lids are both the transfer of bulky hydrophobic residues out of the N-terminal end of the α6 helix, and the incorporation of short side chain residues to the α6 C-terminal end. All these structural changes are stabilized by the Zn++ binding domain, which is characteristic of this family of lipases. Two detergent molecules are placed in the active site, mimicking chains of the triglyceride substrate, demonstrating the position of the oxyanion hole and the three pockets that accommodate the sn-1, sn-2 and sn-3 fatty acids chains. The combination of structural and biochemical studies indicate that the lids opening is not mediated by temperature but triggered by interaction with lipid substrate.
Lipases (triacylglycerol lipase EC 3.1.1.3) catalyse the hydrolysis of long-chain triacylglycerides at
water/oil interfaces. They are very versatile enzymes because, in vitro, they catalyse both the hydrolysis and the synthesis of a great variety of esters (1,2), the hydrolysis and transesterification of triacylglycerols (3) or the resolution of racemic mixtures (4). Most lipases contain a lid domain controlling access to the active site (5). The interaction of the enzyme with lipid aggregates induces the displacement of the lid, which makes the active site accessible to individual substrate molecules and increases the catalytic activity. This phenomenon is known as interfacial activation (5, 6).
Much of the interest in extremophiles stems from their surprising properties. There has been extensive research on the structural proteins and key metabolic enzymes that are responsible for the organisms’ unusual properties. Recent research has focused on the identification of extremozymes relevant for industrial biocatalysis. The bacterial thermoalkalophilic lipases are among the most biocatalytically relevant extremozymes due to their resistance to proteases, detergents and chaotropic agents together with their extreme stability at elevated temperatures and in organic solvents (7). Thermoalkalophilic lipases are found from several of thermophilic aerobic Bacteria recently reclassified as the new genus Geobacillus (8), Geobacillus stearothermophilus (9), Geobacillus thermocatenulatus (10), Geobacillus thermoleovorans (11), and Geobacillus sp. TP10A. (12). Thermoalkalophilic
http://www.jbc.org/cgi/doi/10.1074/jbc.M808268200The latest version is at JBC Papers in Press. Published on December 3, 2008 as Manuscript M808268200
Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

2
lipases show optimal activity at 60–75 °C and pH 8–10 and share about 95% amino acid sequence identity among them and show a significant similarity of 30–35% with the mature lipases from other Gram-positive bacteria Staphylococcus strains. In contrast to the similarity with staphylococcal lipases, the thermoalkalophilic lipases exhibit no sequence similarity with other microbial lipases (13). They are also characterized by their significantly larger molecular sizes (40–45 kDa), relative to other microbial lipases (usually under 35 kDa). Thus, the thermoalkalophilic lipases and staphylococcal lipases were originally grouped in one lipase family named the lipase family I.5 (14) or Staphylococcus family (13). Later, because of their lower level of sequence similarity to the Geobacillus lipases, the Staphylococcus enzymes were re-assigned to family I.6. (15).
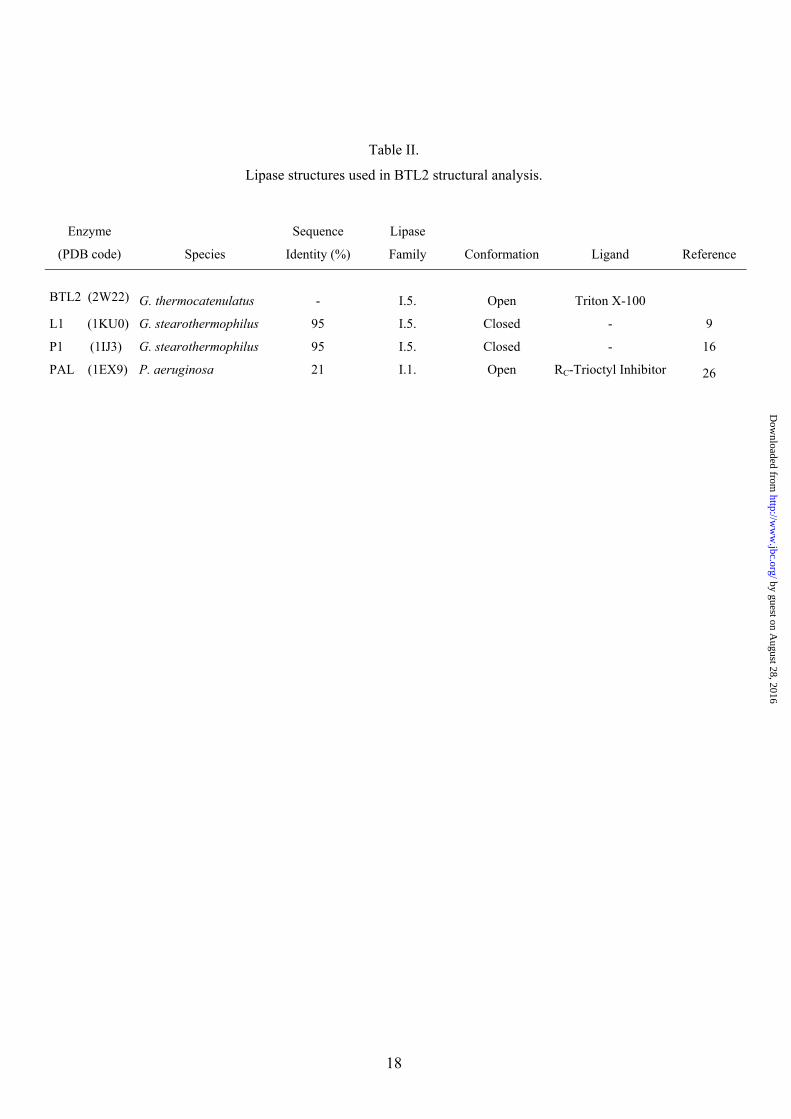
Crystal structures of two isoforms of the Geobacillus stearothermophilus lipase (named L1 and P1), and of the lipase from Geobacillus zaliae (named T1) have been reported (9, 16, 17). In those structures, lipases presented its closed conformation with the active site buried under a long lid helix. The structures exhibited a zinc-binding site in an extra domain accounting for the larger molecular size of the family I.5 enzymes in comparison to other microbial lipases.
The thermophile Geobacillus thermocatenulatus produces two lipases (BTL1 and BTL2) with different sizes (13). BTL2 is a 43 kDa protein (predicted from the DNA sequence) that showed high stability at medium temperatures (50°C), alkaline pH (9.0–11.0) and in organic solvents (2-propanol, acetone, methanol) (18). Furthermore, it has been reported that this lipase presents a high specificity towards very different substrates with a great selectivity in the resolution of key intermediates in the synthesis of drugs (19). Herein, we report the crystal structure of the BTL2 in an open conformation with two molecules of Triton detergent present into the active site. Activation involves dramatic concerted structural rearrangements in which the zinc binding domain plays a critical role. The combination of crystallographic and biochemical studies has allowed us to gain unprecedented insights into the activation events that lead to the catalytic turnover processes. This knowledge is central in understanding how interfacial activation is triggered in thermoalkalophilic lipases and paves the way for engineering lipases with biotechnological purposes.
EXPERIMENTAL PROCEDURES
Cloning, Expression and Purification of BTL2. The gene corresponding to the mature lipase from Geobacillus thermocatenulatus BTL2 was cloning into pT1 expression vector as previously described (18). Cells carrying the recombinant plasmid pT1BTL2 were grown at 30 ºC and overexpression was induced raising the temperature to 42 ºC for 20 hours. The enzyme was purified from E. coli crude extract using a sequential chromatography steps procedure in batch (octyl-sepharose, PEI-agarose and Q-sepharose) as previously described (22). In this purification procedure, a last washed step containing Triton X-100 (10 nm in distilled water) before enzyme elution from Q-sepharose, permitted BTL2 activation for crystallization experiments.
Immobilization of BTL2. BTL2 immobilization on cyanogen bromide agarose (CNBr) was performed as previously described (20). This preparation (prepared in the presence of detergent but later thoroughly washed with distilled water) permitted to immobilize the monomeric form of the lipase, avoiding any intermolecular lipase-lipase interactions that could interfere the properties of soluble lipases (21).
BTL2 Activity Measurements. Lipase activity was measured by absorbance variation at 348 nm of p-nitrophenol due to pNPB hydrolysis (0.4 mM) in buffer M (sodium phosphate 25 mM, pH 7.0) at 25 ºC, using a themostatized cuvette with continuous magnetic stirring.
BTL2 Activation Experiments. To test the detergent effect in BTL2 activation, increasing concentrations of Triton X-100 were added to purified BTL2 (soluble and immobilized) in 25 mM sodium phosphate at pH 7 and 25 ºC at a concentration of 0.1 mg of BTL/ ml.
To study lipase inhibition kinetics, D-pNP (0.6 µM) was added to purified BTL2 (soluble and immobilized) at a concentration of 0.1 mg of BTL/ ml in 25 mM sodium phosphate at pH 7 and 25 ºC in the absence of Triton X-100 and in increasing concentrations of Triton X-100. Enzyme activity was checked at different times using the assay described above.
Crystalization and Data Collection. Native crystals of BTL2 were grown using hanging drop vapor diffusion method as previously reported (22). Good-quality diffracting crystals, showing rice-grain shapes were found in 0.05 M sodium citrate pH 5.6, MPD 13% and ammonium acetate 0.2 M. Native data sets were collected using
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

3
synchrotron radiation source at the ESRF (Grenoble) on BM16 beamline using an ADSC Reverse PHI detector and a wavelength of 0.979234 Å. Collected images were processed and scaled using MOSFLM and SCALA programs, respectively from CCP4 package (Collaborative Computational Project, Number 4, 1994). BTL2 crystals belong to orthorhombic I222 space group (a = 73.07 Å, b = 129.08 Å, c = 127.49 Å.).
Structure Determination and Refinement. The BTL2 structure was solved by the molecular replacement method with MOLREP program from the CCP4 package using P1 lipase structure from Geobacillus stearothermophilus (PDB code 1ji3) as initial model. The model was subjected to successive refinement cycles with the CNS program (23) and intensive manual model building used the software package O (24). Excellent density maps were obtained for BTL2 structure except for the first two residues of polypeptide chain. Two fragments of Triton X-100 used in the purification procedure were found in the catalytic groove of BTL2 structure and three MPD molecules on the protein molecular surface involved in crystal packing interactions. Water molecules were gradually added with waterpick routine of CNS program.
RESULTS AND DISCUSSION
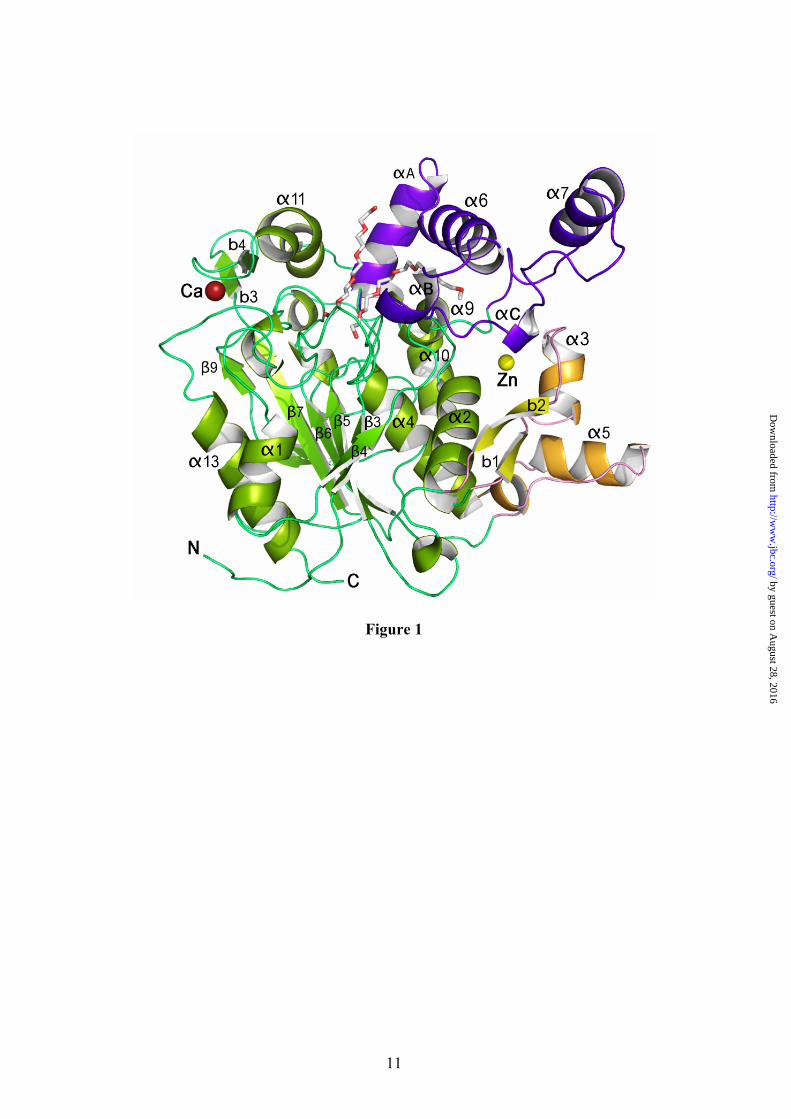
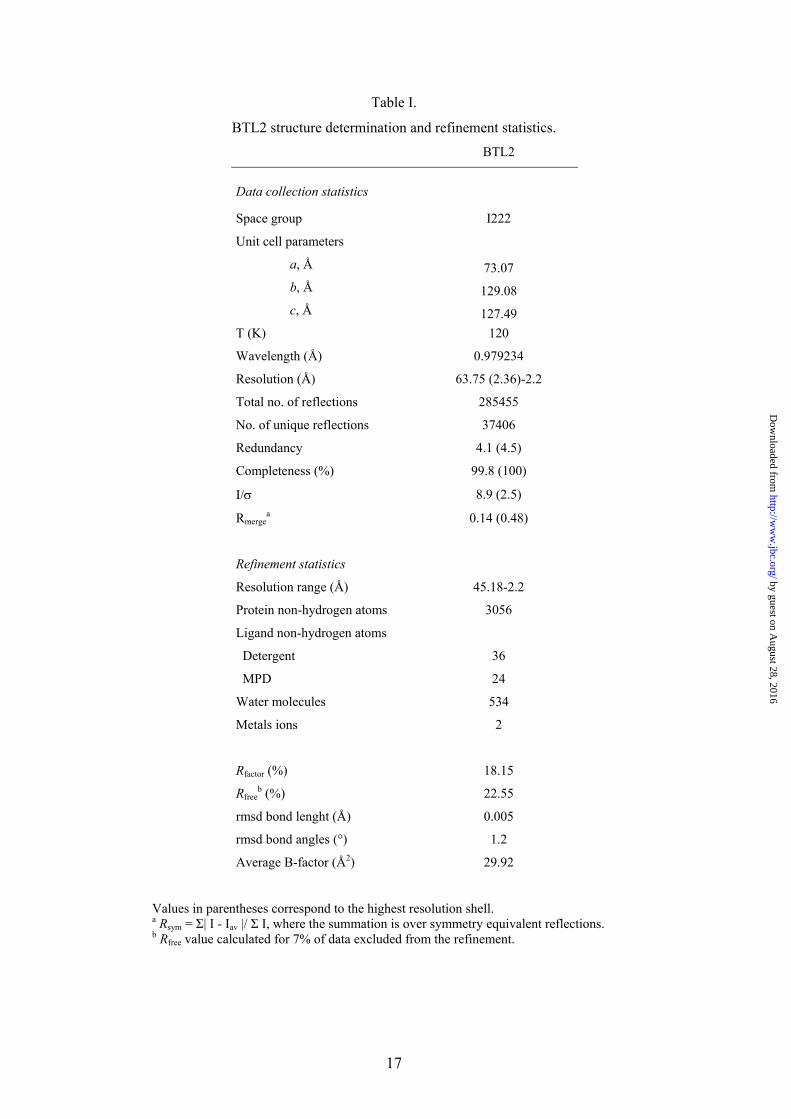
Overall Structure. The crystal structure of BTL2 has been solved at 2.2 Å resolution (structural determination parameters and refinement statistics are summarized in Table I). The three-dimensional structure of BTL2 (Fig. 1) comprises 389 residues and consists of an irregular (α/β) hydrolase fold formed by a central β-sheet of seven strands (β3-β9) surrounded by α-helices (α1 and α13 on one side and α2, α4 and α10 on the other side). An extra domain involved in Zn coordination is present in this family of lipases consisting of helices α3 and α5 and strands b1 and b2, as previously described (9). No significant variations are observed in the Zn coordination in BTL2 versus B. stearothermophilus lipases (lipases used in BTL2 structural analysis are summarized in Table II). Zn coordination bonds are formed with His82 and His88 (2.29 Å and 2.21 Å respectively) from the extra domain, and Asp62 (2.11 Å and 2.96 Å) and Asp239 (2.13 Å and 2.71 Å) from the (α/β) hydrolase core domain. A Ca++ ion is also found to form hydrogen bonds with regions comprised between b4 and α11, and β9
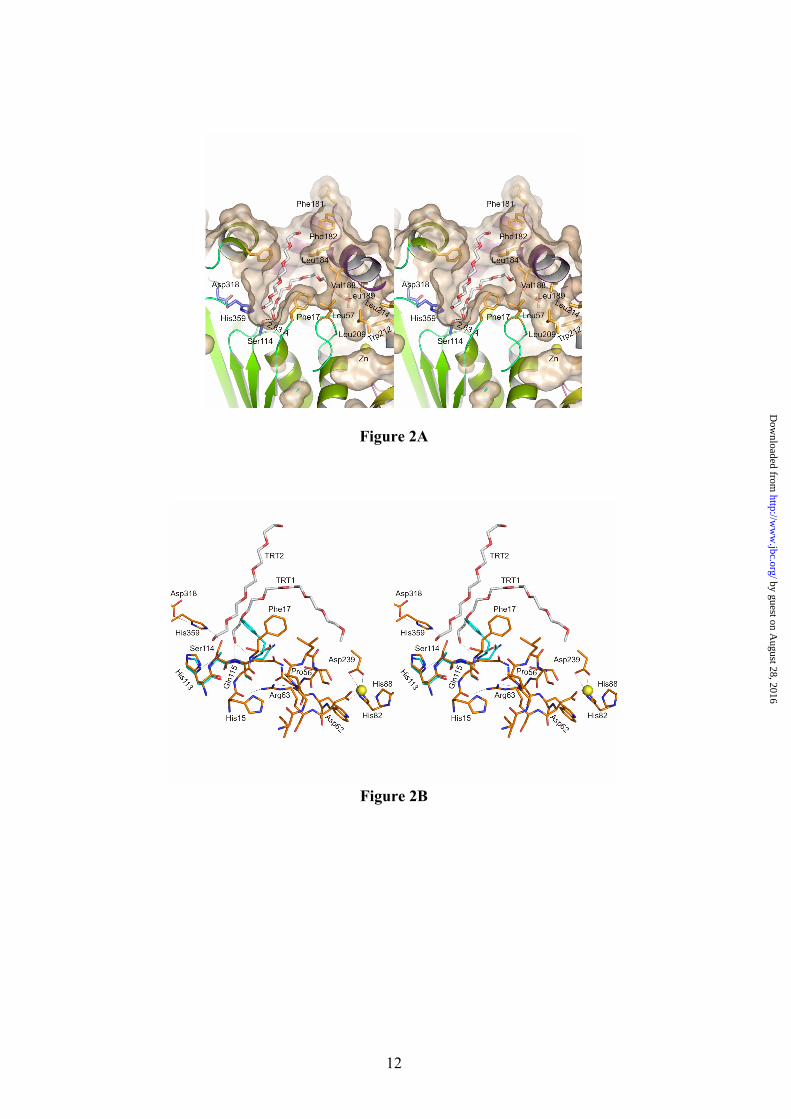
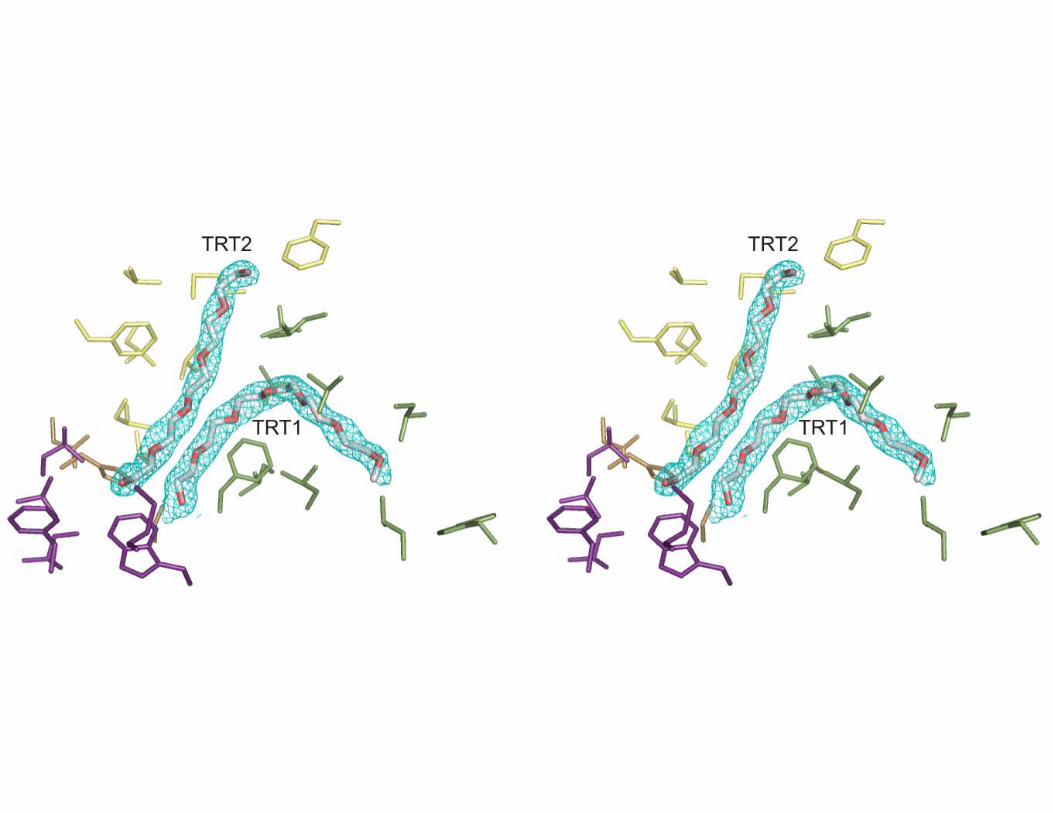
and α13 (Figure 1). The coordination includes interactions with carboxyl oxygen atoms of Glu361 (2.96 Å) and Asp366 (2.89 Å), and with two main chain carbonyl oxygen atoms of Gly287 (2.61 Å) and Pro367 (2.58 Å). BTL2 structure has been solved in the open (active) conformation with two Triton X-100 detergent molecules placed at the active site (Figure 2 and Fig.1 Supp. Mat).
The Catalytic Machinery of BTL2. The catalytic machinery of termoalkalophilic lipases is formed by the catalytic triad and the oxyanion hole. The active site of BTL2 contains the catalytic triad residues (Ser114, His359 and Asp318) located at their canonical positions in the α/β hydrolase fold (Figure 2A). The catalytic serine is in the Ala-X-Ser-X-Gly motif characteristic of the termoalkalophilic lipases (versus the Gly-X-Ser-X-Gly motif of most lipases and serine proteases) (25). The catalytic serine is exposed to solvent and substrate. In the closed state of L1 lipase it has been observed that catalytic serine is in tight side-chain packing with some residues of the active site (His113, Phe17, Ile320, Thr270 and Met326) resulting in the stabilization of the serine loop and contributing to the lipase thermostability (9). In the BTL2 open structure this tight packing is also maintained except for the Phe17 (Figure 2B). This residue changes its conformation allowing the access of the substrate to the catalytic Ser that changes its conformation reorienting its Oγ group to the substrate. The movement of Phe17 involves a change in its χ1 side-chain torsion angle (∆χ1= 100º) from the closed to the open state. The conformer change of catalytic serine from the closed to the open forms, also involves changes in the hydrogen-bonding pattern. In closed state, the serine side chain Oγ atom points to His113 Nδ1 atom (2.89 Å) and is 3.36 Å from the catalytic histidine Nε2 atom. However, in the active state, the serine side chain Oγ atom is 2.96 Å from the catalytic histidine Nε2 atom and 5.08 Å from the His113 Nδ1 atom. No changes are observed in the hydrogen bonds between catalytic residues His359 (Nδ1 atom) and Asp318 (Oδ2 atom) in both closed and open states (2.68 Å and 2.81 Å respectively) (Fig. 2B).
The oxyanion produced in the tetrahedral intermediate is stabilized by interactions in the oxyanion hole, which in families I.1 and I.2 of bacterial lipases has been identified to be formed by the main chain amide groups of a couple of residues (Met16 and His83 in I.1 family, and Leu17 and Gln88 in I.2 family (26). The
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

4
equivalent atoms in BTL2 are Phe17 and Gln115 that occupy virtually identical positions. The Phe17 and Gln115 backbone nitrogen atoms localize the oxyanion binding pocket in a position that is very well conserved within the α/β hydrolase-fold enzymes (27-29). Phe17 is the second residue of the tetrapeptide motif Gly-Hyd-X-Gly (Hyd= Phe, Leu or Ile) located between strand β3 and helix α4 and highly conserved in bacterial lipases from I.5 family. No relevant differences are observed in the backbone of this region between the closed and open forms, and therefore it seems that formation of the oxyanion binding pocket only requires the movement of the side chain of Phe17 residue.
In families I.1 and I.2 of bacterial lipases a totally buried Arg residue has been found connecting, via hydrogen bonds, the oxyanion hole tetrapeptide motif to the loop between strand β3 and helix α2 (26). The variation of the hydrogen-bonding pattern of this Arg between the closed and open forms of I.1 and I.2 lipases has been predicted to play an important stabilizing role during the opening of the lid of these lipases (26). An Arg residue is also found in the equivalent positions of BTL2 (Arg63) connecting His15 (through a hydrogen bond between its carbonyl oxygen atom and the Arg63 NH2 atom) to Gly55 (through a hydrogen bond between its carbonyl oxygen atom and the Arg63 NH1 atom) (Figure 2B). Comparison with the closed form of L1 lipase reveals that there is no variation in the hydrogen bonding pattern of this Arg residue in agreement with the preformed oxyanion hole found in BTL2. Interestingly, Arg63 is placed just after Asp62 that is directly involved in Zn coordination. Therefore in I.5 family of bacterial lipases the Arg63 might play a stabilizing role of the oxyanion binding pocket, a role that should be reinforced by the Zn binding domain which is characteristic of this family of lipases.
The active site cleft and binding interactions with the substrate. Two molecules of Triton detergent have been identified occupying the hydrophobic active site cleft (Fig. 2A). This cleft is 14 Å deep and has an ovoid shape with approximate dimensions of 18 x 25 Å, with an area of 847.9 Å2 and a volume of 1183.1 Å3 (as calculated by CASTp, http://sts.bioengr.uic.edu/castp/index.php). Its walls are lined mostly with hydrophobic/aromatic side chains (Ala241, Ile320, Ile363, Leu171, Leu184, Leu189, Leu209, Leu245, Leu360, Leu57, Met174, Phe17, Phe182, Phe291, Pro165, Tyr30, Val172, Val175, Val188, Val234, Val295,
Val321 and Val365) in order to achieve a perfect stabilization of the lipid substrate. At the base of the cleft, the Phe17 side chain divides it in two parts, thus giving a boomerang-like shape to the active site as also described for other lipases (Fig. 2A), with four binding pockets, an oxyanion hole and three pockets for the different branches of the triacylglycerol substrate.
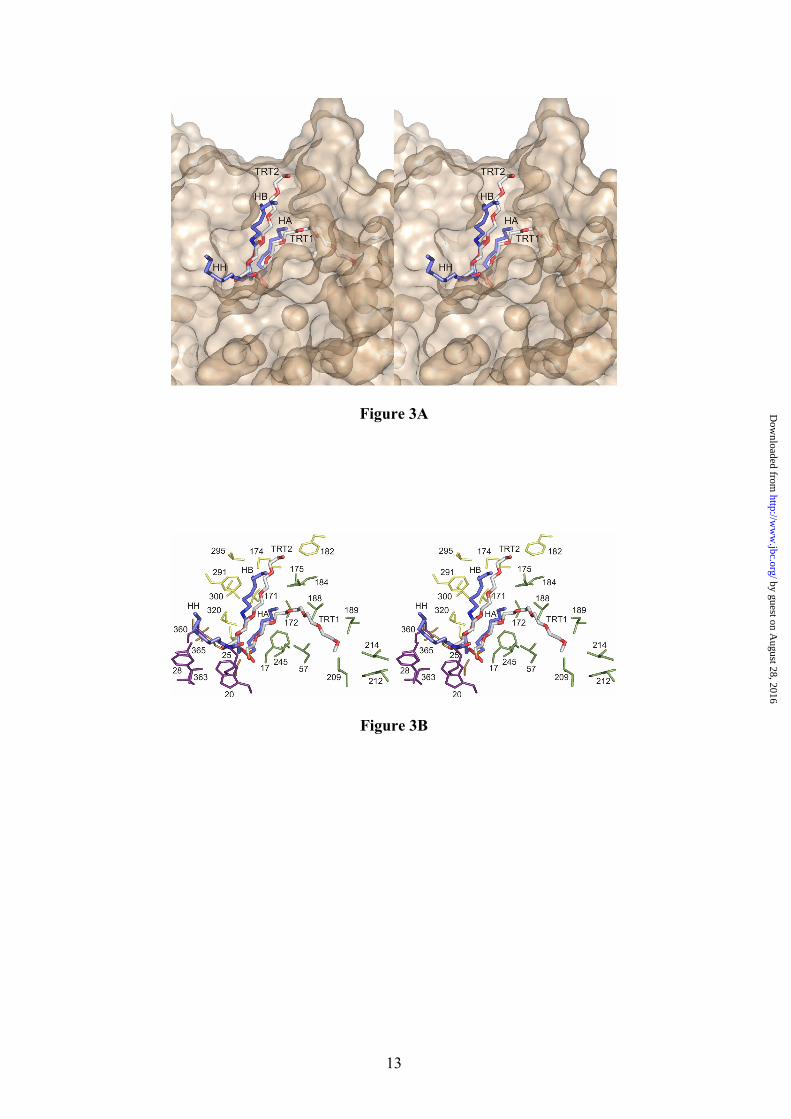
The Pseudomonas aeruginosa lipase (PAL) belonging to I.1 family of bacterial lipases has been crystallized in complex with a triglyceride-like inhibitor (26) (Table II). The complex has demonstrated, for the I.1 family, the position of the oxyanion hole and of the three pockets that accommodate the sn-1, sn-2 and sn-3 fatty acid chains. Despite the structural differences between PAL and BTL2, superimposition of the (α/β) hydrolase core of both enzymes yields a well fitted triglyceride analog into the active site cleft of the BTL2 lipase (Fig. 3A). Remarkably, superimposition reveals that Triton X-100 moieties found in the BTL2 structure are coincident with two acyl chains (sn-1 and sn-3) of the triacylglycerol substrate in PAL. Therefore, structural analysis of PAL and BTL2 offers the opportunity of direct identification of the three acyl-binding pockets of the tryglyceride substrate in the I.5 family of bacterial lipases. The Triton X-100 molecule TRT1 found in BTL2 is accommodated in a large groove formed by the side chains of Phe17, Leu184, Val188, Leu189, Leu57, Leu209, Leu214 and Trp212 (Fig. 3). The long aliphatic chain of 20 atoms of the detergent perfectly fits in this cleft and is bound via van der Waals interactions. This cavity in PAL corresponds to the the acyl chain pocket A (HA). As in the PAL structure, the alcohol binding pocket is separated from the acyl pocket by the side chain of Phe17 (Met16 in PAL), however the acyl pocket groove is larger in BTL2 (about 9.0 Å x 22 Å) than in PAL (about 8.5 Å x 9.0 Å) pointing to longer chain substrates in I.5 family than in I.1 family of bacterial lipases.
The acyl chain binding pocket corresponding to the sn-2 moiety of the triglyceride analog (HH pocket) in PAL corresponds in BTL2 to a hydrophobic pocket lined by Ile363, Trp20, Phe28, Met25, Leu360 and Val365 (Figure 3). The latter residues are part of the calcium-binding loop (residues 358 to 367). The HH pocket is similar in PAL than in BTL2, and there is space for about 8-10 fatty acid carbon atoms. The acyl chain binding pocket the sn-1 moiety of the triglyceride analog (HB pocket) in PAL corresponds in BTL2 to a hydrophobic pocket lined by Ile320, Val321,
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

5
Leu171, Val175, Leu184, Met174, Phe291 and Val295 (Fig. 3). The hydrophobic nature of HB pocket is similar in both BTL2 and PAL lipases, however in BTL2 the HB pocket tightly interacts with TRT2 Triton X-100 molecule, while in PAL this pocket is more exposed to the solvent. The three pockets (HA, HH and HB) identified in BTL2 should also be present in the rest of the members of the I.5 family of bacterial lipases considering their high degree of sequence similarity.
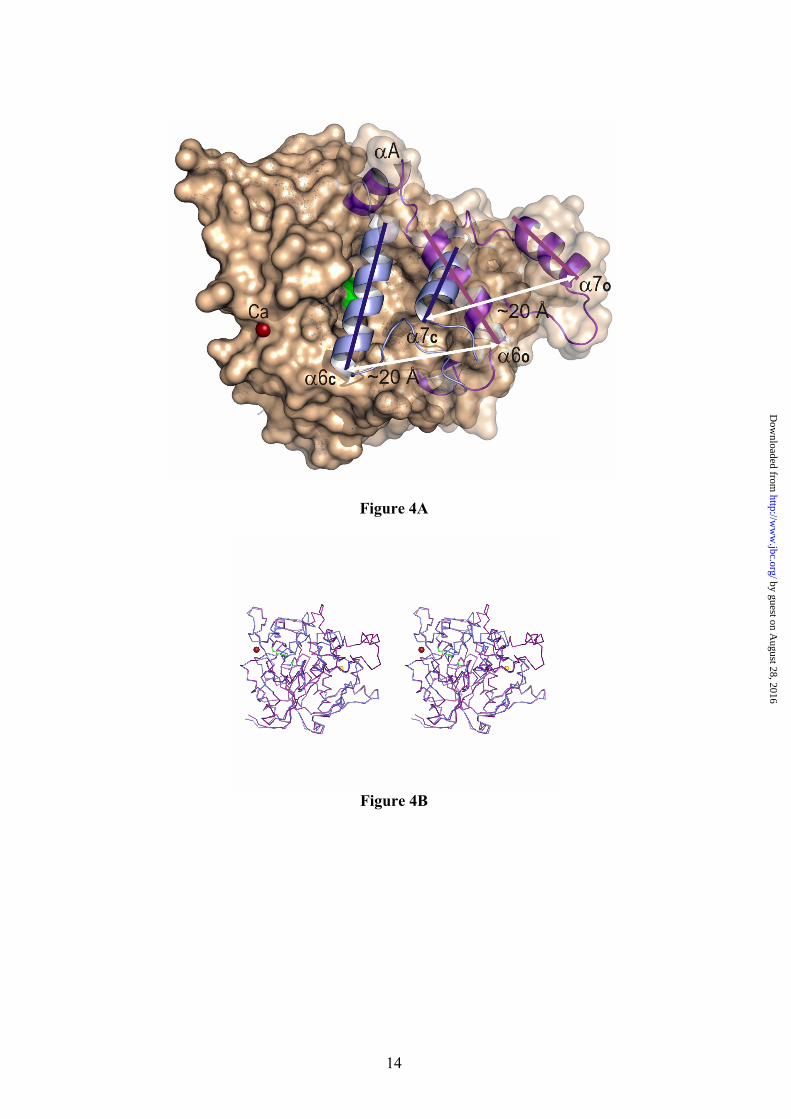
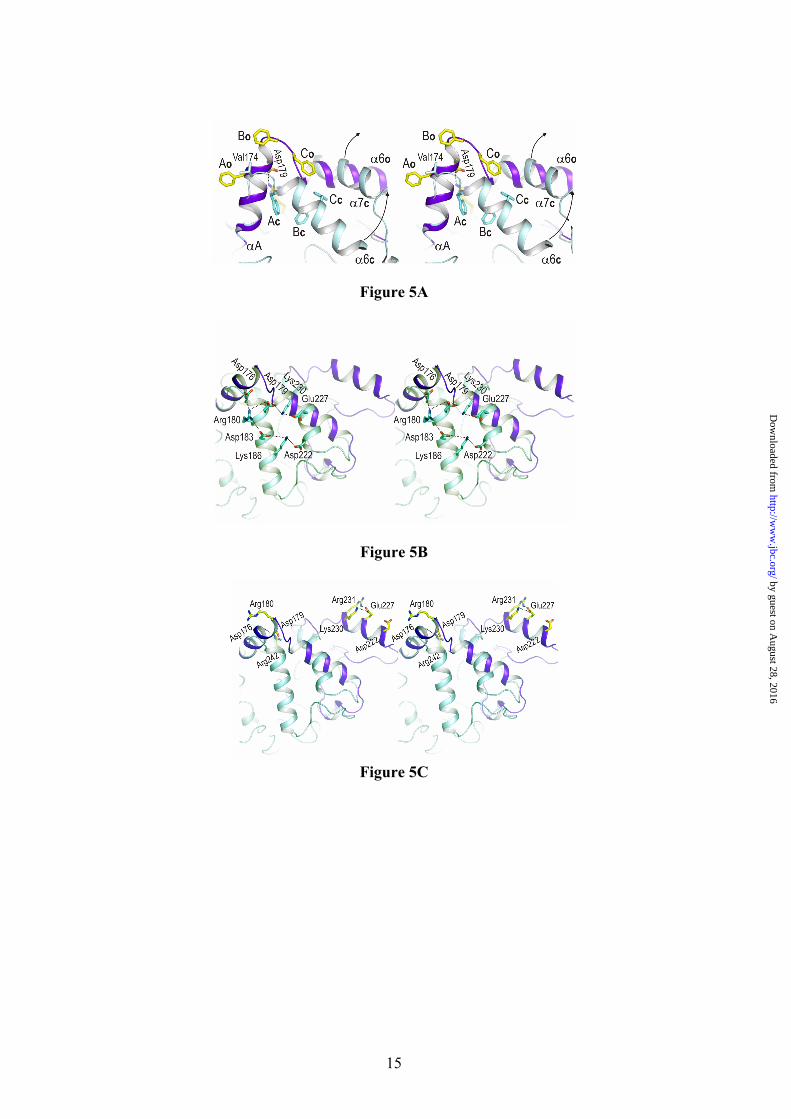
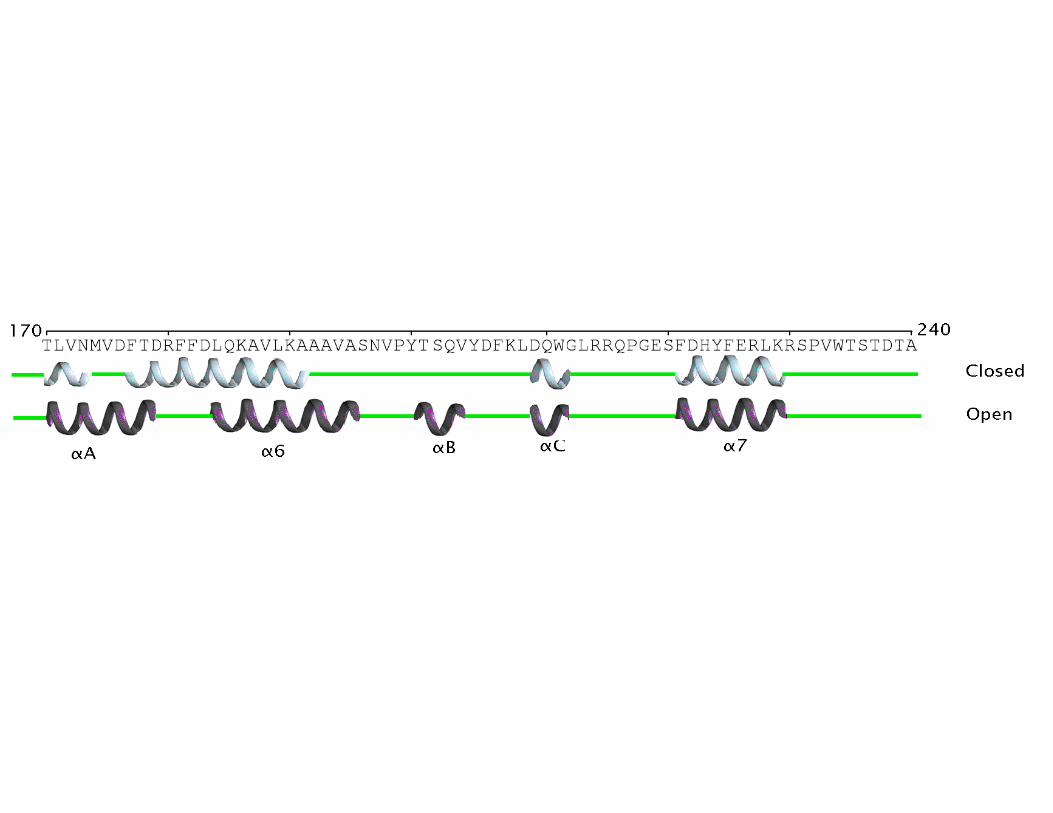
Activation Mechanism. The existence of an adjustable loop at only one end of the α6 helix and the bipartite distribution of hydrophobic resides in that α6 helix, with bulky side chains at the N-terminal half and short side chains residues at the C-terminal half, led to the proposed model of unidirectional opening mechanism of the lid (9). In this model, lid opening should involve only the C-terminal half of the α6 helix (around eight amino acids). Unexpectedly, enzyme activation involves both dramatic conformational rearrangements of two lids covering the active site (associated to α6 and α7 helices respectively), that upon activation move more than 20 Å away (Fig. 4A), and produces new secondary structure elements in the enzyme (αA, αB and αC) (Figure 1). Overall, enzyme activation results in changes around 70 amino acids in the region comprised between Thr169 to Asp239 (coloured magenta in Fig. 1) and the creation of a large hydrophobic cavity. Moreover, activation causes the hyper-exposure of ten aromatic side chains (Phe28, Phe177, Phe181, Phe182, Tyr200, Tyr205, Phe207, Phe222, Phe226 and Phe299).
Structural superimposition of BTL2 open lipase with the L1 closed lipase from Geobacillus stearothermophilus yields a rmsd (root mean square deviation) of 0.55 Å for 384 backbone Cα atoms. While the (α/β) hydrolase core is mainly conserved between both structures, strong differences are observed in the 169-239 region (rmsd of 7.49 Å for the backbone Cα) (Figure 4B and Fig1-Supplementary mat.). Pivotal in this rearrangement are the movements of α6 and α7 helices. While movement of α7 helix (residues 222-236) can be described as a “typical” lid opening around a molecular hinge (placed at Gln211 and Asp239), the α6 helix movement involves both a lateral displacement and a structural reorganization of their residues. In the closed conformation the α6 helix (α6c) comprises residues 177-192 (FTDRFFDLQKAVLEAA) (BTL2 residue numbering) and upon activation the
α6 helix (α6o) comprises residues 184-197 (LQKAVLEAAAVASN). The seven first residues at the N-terminal half of α6c, having bulky side chains, upon activation are incorporated to the αA helix and the loop connecting both helices (Fig. 5A). Thus the αA helix, formed by only four residues in the closed form (TLVN), is formed by eleven residues (TLVNMVDFTDR) in the open form. It is worth noting that the size of α6 helix remains similar in both closed and open forms. This is possible thanks to the incorporation to the α6 helix of five short side chains residues from the adjustable loop previously described (9).
How these large structural rearrangements are produced? Structural analysis of closed and open forms provides relevant insights about the molecular events triggering restructuring of the lids and stabilization of the active conformation. In the closed state, a short hydrophobic side chain (Val or Leu in the I.5 family) placed in the loop connecting αA to α6 is protruding to the solvent (Fig. 5A). Activation causes the first Phe residue from the N-terminal half of α6 (Phe177 in BTL2) to occupy the position of the Val/Leu residue of the closed form restructuring the loop to increase the length of the αA helix. This movement of Phe177 could be triggered by the interaction with the lipid interface. Other residues from the N-terminal half of α6c (Thr178, Asp179 and Arg180) are successively incorporated to the αA helix. Interestingly, incorporation of Asp179 to αA helix allows a strong salt bridge interaction with Arg242 (2.76 Å) anchoring the open lid to the (α/β) hydrolase core. The restructuring process of αA helix is stopped by two consecutive Phe residues (Phe181 and Phe182) that go out, from the highly hydrophobic core of the closed state, to a completely exposed position on the BTL2 surface, presumably to interact with the lipid interface. As a collateral effect, the αA helix refolding provokes a lateral displacement of α6 helix, unmasking the active site. The transfer of amino acids from α6 to αA helices is accompanied by refolding of loop after α6, in such a way that five residues from the loop are incorporated to the α6o helix. A general view of the structural rearrangements involved in the activation process can be obtained from the movie 1 (supplementary material).
As here described, the activation of thermoalkalophilic lipases involves concerted movements of lids α6 and α7. A strong network of salt bridge interactions is observed in this region for the closed state and specifically two
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

6
interactions (Asp179:Lys230 and Lys186:Asp222) directly connect both lids (Fig. 5B). In the active state fewer of salt bridge interactions are observed and none of them linking α6 and α7 lids (Fig. 5C). Structure of BTL2 reveals that Zn binding domain, characteristic from the I.5 family of lipases, plays a pivotal role in the stabilization of the large structural rearrangements involved in the activation process. Molecular hinge of α7 lid is located at residues Asp239 and Gln211, the former being directly involved in the Zn++ coordination and Gln211 making a hydrogen bond with Asp62 (3.08 Å) that directly coordinates the Zn cation. Besides, the same region (residues 208-211) acts as molecular hinge of the α6 lid. In this region two salt bridge interactions (Lys208:Glu24 2.79 Å and Asp210:Arg93 3.08 Å) connect lid regions with the inner core. All these interactions are preserved in the closed and open forms. It is worth noting that mutagenesis experiments on the zinc-ligand residues (30) results in a notable decrease in the optimal temperature for maximal activity (from 60 to 45–50 ºC) and in the abolition of the Zn++-induced thermal stabilization. Therefore the role of the Zn++ binding domain seems to be critical in stabilizing the structural refolding associated to the activation process and in the thermal stabilization of the active form at high temperatures.
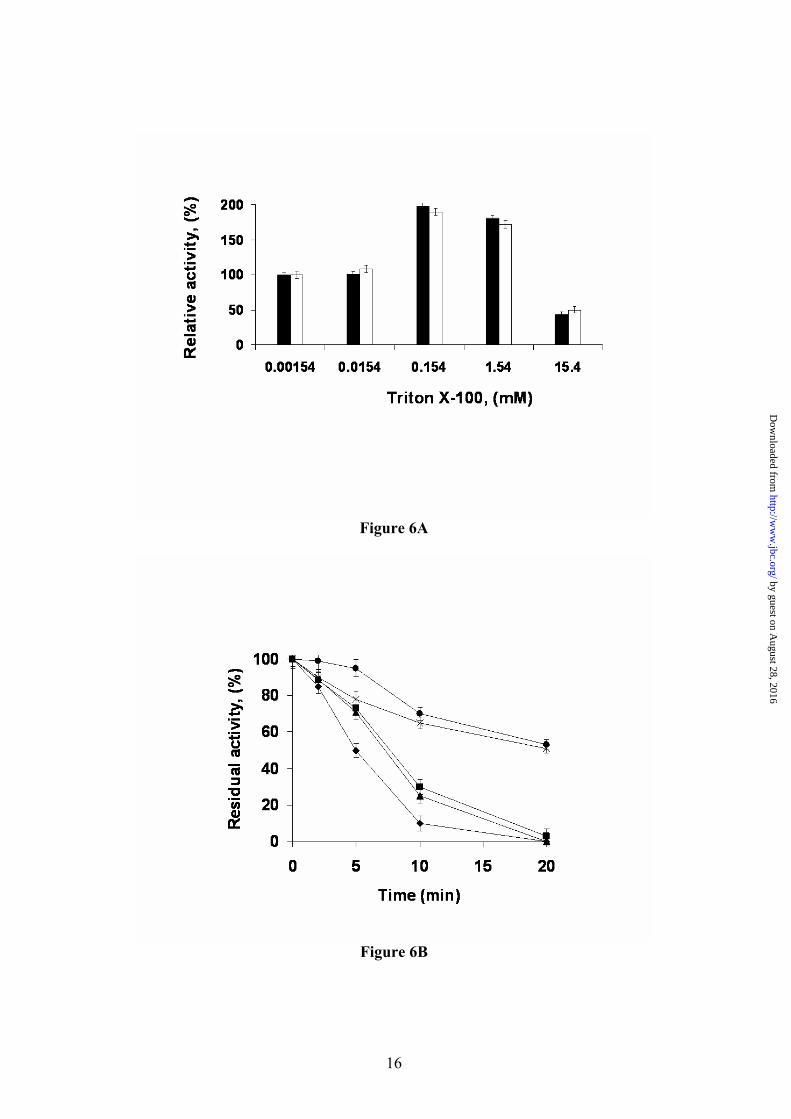
Is temperature the switch of the bacterial thermoalkalophilic lipases? Based on the strong hydrophobic interactions observed for the α6 lid helix with the catalytic triad region in the structure of the closed L1 lipase, it has been proposed that the lid may not be opened at low temperatures even in the presence of lipid substrates (9,30). Therefore, only the increased kinetic energy at high temperatures should allow the movement of the lid by effective solvation of the residues involved in hydrophobic interactions. Considering that the crystal structure of BTL2 has been solved in an open conformation in absence of high temperatures, another mechanism should be proposed. The effect of Triton X-100 in BTL2 activation at low temperatures was explored by an enzymatic kinetic at 25 ºC (Fig. 6A). The experiment was performed using both soluble and monomer-immobilized enzyme to discard any possible artefact due to lipase-lipase interactions breaking (31). Both curves were almost identical and showed an increase in the BTL2 activity by a 2-fold factor at 0.15 mM of detergent concentration. At higher detergent concentrations (15 mM) the activity decreased to 40-50 % compared to control (without detergent). The
enzyme activity increment could be related to the stabilization of the lipase open form stimulated by detergent molecules interaction with hydrophobic catalytic groove as in the BTL2 open form with TRT1 and TRT2; this is a known effect in soluble and immobilized lipases (32). BTL2 activity decrease may be related to the inhibitory effect of Triton X-100 at concentrations higher than 1 mM in which, a competitive inhibitory effect could prevent the substrate catalysis, as also observed in other lipases (33).
To further clarify the effect of Triton X-100 on BTL2 lipase activation and stabilization of the open form, we have studied the reactivity of the catalytic serine residue to D-pNP, a serine hydrolase specific covalent inhibitor. Soluble enzyme was incubated with the inhibitor at different concentrations of Triton X-100. In detergent absence, enzyme inhibition was quite slow (50 % after 20 minutes) and Triton X-100 1.5 µM did not change the inhibition rate (neither this concentration was able to increase the enzyme activity). However, detergent concentrations from 0.015 mM to 1.5 mM accelerated the inhibition rate with a lost of 100 % enzyme activity in a shortest time period (Fig 6B). These results were almost identical when immobilized enzyme was used (data not shown). D-pNP inhibits BTL2 lipase irreversibly in presences of submicellar concentrations of Triton X-100 by reacting with the serine residue at the active site. This result indicates that the effect of Triton X-100 detergent is to open the lipase lid, thereby making the active site serine residue accessible to inhibition.
The increases of BTL2 activity in the presence of Triton X-100 and the irreversible inhibition by D-pNP inhibition reached following Triton X-100 treatment, both at 25 ºC, points to the interaction with the lipid interface and/or the nature of the substrate as the main causes of the activation mechanism. Interaction of Phe177 with the lipid interface could trigger the restructuring process leading to the hyper-exposure of ten aromatic residues. Interaction with the hydrophobic interface should also destabilize the charged form of Lys residues in this region; especially Lys186 and Lys230 that connect closed lids through salt bridge interactions. It has been reported that a burial of a lysine residue fully buried in a hydrophobic core environment dramatically decreases its pKa value, the ε-amine group being deprotonated (34). Loss of the inter-lid salt-bridge interactions together with the exposure of aromatic residues, should favor the open structural state observed in BTL2.
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

7
Conclusion. The crystal structure of BTL2 shows, for the first time, the structural rearrangements required for the activation of the I.5 family of bacterial lipases. Structural analysis revealed the different pockets that, upon activation, are responsible of stabilization of the acyl chains of the substrate. These rearrangements are stabilized by the Zn-binding domain that is also critical in their stabilization under high temperatures. The asymmetric amino acid composition of the α6 helix and the presence of the adjustable loop seem to be central in the activation mechanism of the thermoalkalophilic lipases. Structural and biochemical results here reported, point to the enzyme-substrate interaction as the driving force of the activation mechanism. This structure and the novel activation mechanism we have documented for BTL2 open future avenues of research on the engineering lipases
with biotechnological purposes and provides an exquisite example of the determinants involved in large structural rearrangements triggered by the interaction of proteins with lipids. The atomic coordinates and structure factors (PDB code 2w22) have been deposited in the Protein Data Bank, Research Collaboratory for Structural Bioinformatics, Rutgers University, New Brunswick, NJ (http://www.pdb.org/).
Acknowledgements- We thank Douglas Laurents for critically reading the manuscript. César Carrasco-López was a fellow of the Fundayacucho Foundation (Venezuela). This work was supported by grant BFU2005-01645 from Dirección General de Investigación and by “Factoria de Cristalización”, CONSOLIDER INGENIO-2010.
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

8
REFERENCES 1. Björkling, F., Godtfredsen, S. E., Kirk, O. (1991) Trends Biotechnology 9, 360-363 2. Bornscheuer, U. T. (1995) Enzyme Microb 17, 578-586 3. Mukherjee, K. D. (1990) Biocatalysis 3, 277-293 4. Jaeger, K. E., and Reetz, M. T. (1998) Trends Biotechnol 16, 396-403 5. Derewenda, Z. S., and Sharp, A. M. (1993) Trends Biochem Sci 18, 20-25 6. Sarda, L., Desnuelle, P. (1958) Biochim. Biophys. Acta 30, 513-521 7. Demirjian, D. C., Moris-Varas, F., and Cassidy, C. S. (2001) Curr Opin Chem Biol 5, 144-151 8. Nazina, T. N., Tourova, T. P., Poltaraus, A. B., Novikova, E. V., Grigoryan, A. A., Ivanova, A.
E., Lysenko, A. M., Petrunyaka, V. V., Osipov, G. A., Belyaev, S. S., and Ivanov, M. V. (2001) Int J Syst Evol Microbiol 51, 433-446
9. Jeong, S. T., Kim, H. K., Kim, S. J., Chi, S. W., Pan, J. G., Oh, T. K., and Ryu, S. E. (2002) J Biol Chem 277, 17041-17047
10. Schmidt-Dannert, C., Sztajer, H., Stocklein, W., Menge, U., and Schmid, R. D. (1994) Biochim Biophys Acta 1214, 43-53
11. Cho, A. R., Yoo, S. K., and Kim, E. J. (2000) FEMS Microbiol Lett 186, 235-238 12. Bell, P. J. L., Nevalainen, H., Morgan, H. W. and Bergquist, P. L. (1999) Biotechnol. Lett. 21,
1003-1006 13. Schmidt-Dannert, C., Rua, M. L., and Schmid, R. D. (1997) Methods Enzymol 284, 194-220 14. Jaeger, K. E., Dijkstra, B. W., and Reetz, M. T. (1999) Annu Rev Microbiol 53, 315-351 15. Jaeger, K. E., and Eggert, T. (2002) Curr Opin Biotechnol 13, 390-397 16. Tyndall, J. D., Sinchaikul, S., Fothergill-Gilmore, L. A., Taylor, P., and Walkinshaw, M. D.
(2002) J Mol Biol 323, 859-869 17. Matsumura, H., Yamamoto, T., Leow, T. C., Mori, T., Salleh, A. B., Basri, M., Inoue, T., Kai, Y.,
and Rahman, R. N. (2008) Proteins 70, 592-598 18. Schmidt-Dannert, C., Rua, M. L., Atomi, H., and Schmid, R. D. (1996) Biochim Biophys Acta
1301, 105-114 19. Palomo, J. M., Penas, M. M., Fernandez-Lorente, G., Mateo, C., Pisabarro, A. G., Fernandez-
Lafuente, R., Ramirez, L., and Guisan, J. M. (2003) Biomacromolecules 4, 204-210 20. Palomo, J. M., Ortiz, C., Fuentes, M., Fernandez-Lorente, G., Guisan, J. M., and Fernandez-
Lafuente, R. (2004) J Chromatogr A 1038, 267-273 21. Fernandez-Lorente, G., Godoy, C. A., Mendes, A. A., Lopez-Gallego, F., Grazu, V., de Las
Rivas, B., Palomo, J. M., Hermoso, J., Fernandez-Lafuente, R., and Guisan, J. M. (2008) Biomacromolecules 9, 2553-2561
22. Carrasco-López, C., Godoy, C. de las Rivas, B., Fernández-Lorente, G., Palomo, J. M., Guisán, J. M., Fernández-Lafuente, R., Martínez-Ripoll, M., and Hermoso, J. A. (2008) Acta Crystallogr F . 64, 1043-1045.
23. Brunger, A. T., Adams, P. D., Clore, G. M., DeLano, W. L., Gros, P., Grosse-Kunstleve, R. W., Jiang, J. S., Kuszewski, J., Nilges, M., Pannu, N. S., Read, R. J., Rice, L. M., Simonson, T., and Warren, G. L. (1998) Acta Crystallogr D Biol Crystallogr 54, 905-921
24. Jones, T. A., Zou, J. Y., Cowan, S. W., and Kjeldgaard, M. (1991) Acta Crystallogr A 47 ( Pt 2), 110-119
25. Kim, H. K., Park, S. Y., Lee, J. K., and Oh, T. K. (1998) Biosci Biotechnol Biochem 62, 66-71 26. Nardini, M., Lang, D. A., Liebeton, K., Jaeger, K. E., and Dijkstra, B. W. (2000) J Biol Chem
275, 31219-31225 27. Ollis, D. L., Cheah, E., Cygler, M., Dijkstra, B., Frolow, F., Franken, S. M., Harel, M.,
Remington, S. J., Silman, I., Schrag, J., and et al. (1992) Protein Eng 5, 197-211 28. Nardini, M., and Dijkstra, B. W. (1999) Curr Opin Struct Biol 9, 732-737 29. Cygler, M., and Schrag, J. D. (1997) Methods Enzymol 284, 3-27 30. Choi, W. C., Kim, M. H., Ro, H. S., Ryu, S. R., Oh, T. K., and Lee, J. K. (2005) FEBS Lett 579,
3461-3466 31. Palomo, J. M., Fuentes, M., Fernandez-Lorente, G., Mateo, C., Guisan, J. M., and Fernandez-
Lafuente, R. (2003) Biomacromolecules 4, 1-6
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

9
32. Fernandez-Lorente, G., Palomo, J. M., Mateo, C., Munilla, R., Ortiz, C., Cabrera, Z., Guisan, J. M., and Fernandez-Lafuente, R. (2006) Biomacromolecules 7, 2610-2615
33. Fernandez-Lorente, G., Palomo, J. M., Cabrera, Z., Fernandez-Lafuente, R., and Guisan, J. M. (2007) Biotechnol Bioeng 97, 242-250
34. Stites, W. E., Gittis, A. G., Lattman, E. E., and Shortle, D. (1991) J Mol Biol 221, 7-14
FOOTNOTES
The abbreviations used are: pNPB, p-nitrophenylphosphate butyrate; D-pNP, diethyl p-nitrophenylphosphate; MPD, 2-methyl-2,4-pentanediol.
FIGURE LEGENDS
Fig. 1. Structure of the open conformation of BTL2. View of the BTL2 structure with domains colored differently: (α/β) hydrolase core, green; cap domain (residues 169-239), purple; and the Zinc ion extra domain, yellow. Secondary structure elements are labelled. Metal ions (Zn++, yellow; Ca++ dark red) are spheres. The two molecules of Triton X-100 detergent at the active site are in sticks. Fig. 2. The active site (cleft) of BTL2. (A) Stereo view of the catalytic crevice with two chains of Triton X-100 (white sticks). Catalytic triad (Ser114, His359 and Asp318) is colored in blue. Residues lining the hydrophobic groove are drawn as yellow sticks. (B) Residues of BTL2 involved in the catalytic machinery and oxyanion stabilization are colored in orange; main differences with the closed structure (L1 lipase from B. stearothermophilus) are highlighted in blue. Residues coordinating Zn cation (yellow sphere) are also labelled. Broken lines indicate main interactions. Fig. 3. Putative binding of a triacylglycerol molecule to I.5 lipases. (A) Stereo view of the surface representation of the substrate binding site of BTL2. The two detergent moieties (TRT1 and TRT2) are represented as white sticks. The structure of the triglyceride-like inhibitor (blue sticks) of the P. aeruginosa lipase complex (PDB code 1EX9) is represented as obtained by direct superimposition of the (α/β) hydrolase core of both enzymes. The identified three acyl chain binding pockets (HA, HB and HH) are labelled. (B) Stereo view of the substrate binding mode in BTL2. The tryglyceride-like inhibitor (blue) and Triton X-100 molecules (white), the catalytic triad (brown) and the residues that line the three acyl chain binding pockets, HH (purple), HA (green) and HB (yellow) are labelled. Fig. 4. BTL2 Structural rearrangements upon activation. (A) Molecular surface of BTL2 (brown) with the catalytic Ser residue highlighted in green. Lids changes from the closed (inactive state) to open (active state) structures are represented; α6 and α7 in the closed state (α6c and α7c) are colored in blue; αA, α6 and α7 in the open state (α6o, and α7o) in purple. Upon activation structural rearrangements implies a 20 Å displacement of each lid to unmask the catalytic site. (B) Stereo view of the backbone superimposition of the open conformation of BTL2 (purple) and the closed conformation of B. stearothermophilus L1 lipase (blue). Fig. 5. The activation mechanism. (A) Stereo view showing the differences in the cap domain from closed state (cyan) to the open state (purple). Activation exposes various aromatic residues (yellow, Ao, Bo, Co, corresponding to Phe177, Phe181 and Phe182 respectively) that are buried, and tightly packed, in the closed state (blue, Ac, Bc, and Cc). Upon activation Phe177 interchanges its position with Val174. The successive incorporation of residues from the N-terminal end of α6 helix to αA results in a movement of the lids (indicated by arrows). The active conformation of the lids is stabilized by a salt
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

10
bridge interaction with the core (Asp 179- Arg242). (B) Salt bridge interaction network in the closed L1 structure. (C) Salt bridge interaction network in the open BTL2 structure. Fig. 6. Effect of Triton X-100 in the activity (A) and irreversible inhibition caused by D-pNP (B) of different preparations of BTL2. (A) Activity determinations were carried out as described in methods using different concentrations of Triton X-100. Black bars, results for BrCN immobilized BTL2, white bars results for soluble BTL2. (B) Figure shows the decrease in enzyme activity caused by the irreversible inhibition of BTL2 produced by incubation in the presence of D-pNP. Experiments were carried out as described in methods using different concentration of Triton X-100. Without Triton X-100 (circles) and with various concentrations of Trinton X-100: 0.00154 mM (asterisk); 0.0154 mM (squares); 0.154 mM (triangles) and 1.54 mM (rhombus).
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
12
Figure 2A
Figure 2B
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
13
Figure 3A
Figure 3B
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
14
Figure 4A
Figure 4B
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
15
Figure 5A
Figure 5B
Figure 5C
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
16
Figure 6A
Figure 6B
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
17
Table I.
BTL2 structure determination and refinement statistics. BTL2
Data collection statistics
Space group I222
Unit cell parameters
a, Å 73.07 b, Å 129.08 c, Å 127.49
T (K) 120
Wavelength (Å) 0.979234
Resolution (Å) 63.75 (2.36)-2.2
Total no. of reflections 285455
No. of unique reflections 37406
Redundancy 4.1 (4.5)
Completeness (%) 99.8 (100)
I/σ 8.9 (2.5)
Rmergea 0.14 (0.48)
Refinement statistics
Resolution range (Å) 45.18-2.2
Protein non-hydrogen atoms 3056
Ligand non-hydrogen atoms
Detergent 36
MPD 24
Water molecules 534
Metals ions 2
Rfactor (%) 18.15
Rfreeb (%) 22.55
rmsd bond lenght (Å) 0.005
rmsd bond angles (°) 1.2
Average B-factor (Å2) 29.92
Values in parentheses correspond to the highest resolution shell. a Rsym = Σ| I - Iav |/ Σ I, where the summation is over symmetry equivalent reflections. b Rfree value calculated for 7% of data excluded from the refinement.
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
18
Table II.
Lipase structures used in BTL2 structural analysis.
Enzyme
(PDB code)
Species
Sequence
Identity (%)
Lipase
Family
Conformation
Ligand
Reference
BTL2 (2W22)
G. thermocatenulatus
-
I.5.
Open
Triton X-100
L1 (1KU0) G. stearothermophilus 95 I.5. Closed - 9
P1 (1IJ3) G. stearothermophilus 95 I.5. Closed - 16
PAL (1EX9) P. aeruginosa 21 I.1. Open RC-Trioctyl Inhibitor 26
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Supplementary Material
Figure Legends
Fig. 1. Modification of the secondary structure elements from the closed (L1 lipase from B. stearothermophilus) to the open conformations (BTL2). Only region rearranged upon activation is drawn. Cartoons represent secondary structure elements, closed state (cyan) and open state (magenta).
Fig. 2. Electron density for Triton X-100 detergent molecules. Stereo view of the 2Fo-Fc electron density map of Triton X-100 molecules contoured at 1.0 σ level. Color codes for the amino acids are the same as in Figure 3B. Movie 1: Structural rearrangements between closed and open structures of BTL2. Animation was done using the Yale Morph Server (http://molmovdb.org/morph) in the single-chain morph option with 30 frames. An energy minimization is performed for each single frame between initial (closed BTL2) and final state (open BTL2). BTL2 closed structure was modelled by in silico mutations with the O graphic program (24) based on the closed conformation of the P1 lipase from B. stearothermophilus (PDB code 1ji3) sharing 95% of sequence identity. Reference: Krebs, W.G. and Gerstein, M. (2000). The morph server: a standardized system for analyzing and visualizing macromolecular motions in a database framework. Nucleic Acids Res. 1665-75.

and Juan A. HermosoJosé M. Palomo, José M. Guisán, Roberto Fernández-Lafuente, Martín Martínez-Ripoll César Carrasco-López, César Godoy, Blanca de las Rivas, Gloria Fernández-Lorente,
rearrangementsActivation of bacterial thermoalkalophilic lipases is spurred by dramatic structural
published online December 3, 2008J. Biol. Chem.
10.1074/jbc.M808268200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2008/12/09/M808268200.DC1.html
http://www.jbc.org/content/early/2008/12/03/jbc.M808268200.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on August 28, 2016
http://ww
w.jbc.org/
Dow
nloaded from
Related Documents








![35 [2,3]-sigmatropic rearrangements](https://static.cupdf.com/doc/110x72/55504042b4c905b2788b48e9/35-23-sigmatropic-rearrangements.jpg)


