Associate editor: N. Frossard Acid-Sensing Ion Channels (ASICs): Pharmacology and implication in pain Emmanuel Deval a, ⁎, Xavier Gasull a,b , Jacques Noël a , Miguel Salinas a , Anne Baron a , Sylvie Diochot a , Eric Lingueglia a a Institut de Pharmacologie Moléculaire et Cellulaire (IPMC), UMR 6097 CNRS/Université de Nice-Sophia Antipolis (UNS), 660, route des Lucioles, 06560 Valbonne, France b Lab. Neurophysiology, Facultat de Medicina, Universitat de Barcelona-IDIBAPS. Casanova 143, 08036 Barcelona, Spain abstract article info Keywords: Pain Nociception Acid Ion channels ASIC Neurons Tissue acidosis is a common feature of many painful conditions. Protons are indeed among the first factors released by injured tissues, inducing a local pH fall that depolarizes peripheral free terminals of nociceptors and leads to pain. ASICs are excitatory cation channels directly gated by extracellular protons that are expressed in the nervous system. In sensory neurons, they act as “chemo-electrical” transducers and are involved in somatic and visceral nociception. Two highly specific inhibitory peptides isolated from animal venoms have considerably helped in the understanding of the physiological roles of these channels in pain. At the peripheral level, ASIC3 is important for inflammatory pain. Its expression and its activity are potentiated by several pain mediators present in the “inflammatory soup” that sensitize nociceptors. ASICs have also been involved in some aspects of mechanosensation and mechanonociception, notably in the gastrointestinal tract, but the underlying mechanisms remain to be determined. At the central level, ASIC1a is largely expressed in spinal cord neurons where it has been proposed to participate in the processing of noxious stimuli and in central sensitization. Blocking ASIC1a in the spinal cord also produces a potent analgesia in a broad range of pain conditions through activation of the opiate system. Targeting ASIC channels at different levels of the nervous system could therefore be an interesting strategy for the relief of pain. © 2010 Elsevier Inc. All rights reserved. Contents 1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 549 2. ASICs: from gene to pH-sensor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 550 3. Pharmacology of ASICs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 551 4. Peripheral ASICs in pain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5. Central ASICs in pain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1. Introduction It is well known that tissue acidosis produces pain. For instance, direct application of an acidic solution into the skin induces non- adapting pain (Steen et al., 1992, 1995a,b; Steen & Reeh, 1993; Reeh & Steen, 1996; Steen et al., 1996; Ugawa et al., 2002; Jones et al., 2004). During inflammation, protons are among the first mediators released by damaged cells, and tissue acidosis is a common factor found in several other pain-generating conditions such as ischemia, hemato- ma, fracture, tumor development, skin and muscle incision following surgical procedure, arthritis or urinary tract and digestive diseases. The algogenic effects of acidic pH are caused by the depolarization of Pharmacology & Therapeutics 128 (2010) 549–558 Abbreviations: APETx2, Anthopleura elegantissima Toxin 2; ASIC, Acid Sensing Ion Channel; BNC, Brain Na + Channel; CNS, Central Nervous System; COX, cyclooxygen- ease; DAPI, 4′,6-diamidino-2-phenylindole; DEG, Degenrin; DRG, Dorsal Root Ganglia; EIPA, 5-(N-Ethyl-N-isopropyl)amiloride; ENaC, Epithelial Na + Channel; FaNaC, FMRFa- mide-gated Na + Channel; HSB, hydroxystilbamidine; MDEG, Mammalian Degenerin; NPFF, Neuropetide FF; NPSF, Neuropeptide SF; NSAID, Non Steroidal Anti-Inflammatory Drug; PcTx1, Psalmopoeus cambridgei Toxin 1 (Psalmotoxin 1); PNS, Peripheral Nervous System; TRP, Transient Receptor Potential; TRPV1, Transient Receptor Potential vanilloid 1. ⁎ Corresponding author. E-mail address: [email protected] (E. Deval). 552 554 555 555 555 0163-7258/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.pharmthera.2010.08.006 Contents lists available at ScienceDirect Pharmacology & Therapeutics journal homepage: www.elsevier.com/locate/pharmthera

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pharmacology & Therapeutics 128 (2010) 549–558
Contents lists available at ScienceDirect
Pharmacology & Therapeutics
j ourna l homepage: www.e lsev ie r.com/ locate /pharmthera
Associate editor: N. Frossard
Acid-Sensing Ion Channels (ASICs): Pharmacology and implication in pain
Emmanuel Deval a,⁎, Xavier Gasull a,b, Jacques Noël a, Miguel Salinas a, Anne Baron a,Sylvie Diochot a, Eric Lingueglia a
a Institut de Pharmacologie Moléculaire et Cellulaire (IPMC), UMR 6097 CNRS/Université de Nice-Sophia Antipolis (UNS), 660, route des Lucioles, 06560 Valbonne, Franceb Lab. Neurophysiology, Facultat de Medicina, Universitat de Barcelona-IDIBAPS. Casanova 143, 08036 Barcelona, Spain
Abbreviations: APETx2, Anthopleura elegantissima TChannel; BNC, Brain Na+ Channel; CNS, Central Nervoease; DAPI, 4′,6-diamidino-2-phenylindole; DEG, DegenEIPA, 5-(N-Ethyl-N-isopropyl)amiloride; ENaC, Epitheliamide-gated Na+ Channel; HSB, hydroxystilbamidine; MNPFF, Neuropetide FF; NPSF, Neuropeptide SF; NSAID, NoDrug; PcTx1, Psalmopoeus cambridgei Toxin 1 (PsalmotoxSystem; TRP, Transient Receptor Potential; TRPV1,vanilloid 1.⁎ Corresponding author.
E-mail address: [email protected] (E. Deval).
0163-7258/$ – see front matter © 2010 Elsevier Inc. Aldoi:10.1016/j.pharmthera.2010.08.006
a b s t r a c t
a r t i c l e i n f oKeywords:
PainNociceptionAcidIon channelsASICNeuronsTissue acidosis is a common feature of many painful conditions. Protons are indeed among the first factorsreleased by injured tissues, inducing a local pH fall that depolarizes peripheral free terminals of nociceptors andleads to pain. ASICs are excitatory cation channels directly gated by extracellular protons that are expressed inthe nervous system. In sensory neurons, they act as “chemo-electrical” transducers and are involved in somaticand visceral nociception. Two highly specific inhibitory peptides isolated from animal venoms haveconsiderably helped in the understanding of the physiological roles of these channels in pain. At theperipheral level, ASIC3 is important for inflammatory pain. Its expression and its activity are potentiated byseveral pain mediators present in the “inflammatory soup” that sensitize nociceptors. ASICs have also beeninvolved in some aspects of mechanosensation and mechanonociception, notably in the gastrointestinal tract,but the underlying mechanisms remain to be determined. At the central level, ASIC1a is largely expressed inspinal cord neurons where it has been proposed to participate in the processing of noxious stimuli and incentral sensitization. Blocking ASIC1a in the spinal cord also produces a potent analgesia in a broad range ofpain conditions through activation of the opiate system. Targeting ASIC channels at different levels of thenervous system could therefore be an interesting strategy for the relief of pain.
oxin 2; ASIC, Acid Sensing Ionus System; COX, cyclooxygen-rin; DRG, Dorsal Root Ganglia;l Na+ Channel; FaNaC, FMRFa-DEG, Mammalian Degenerin;n Steroidal Anti-Inflammatoryin 1); PNS, Peripheral NervousTransient Receptor Potential
l rights reserved.
© 2010 Elsevier Inc. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5492. ASICs: from gene to pH-sensor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5503. Pharmacology of ASICs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5514. Peripheral ASICs in pain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5515. Central ASICs in pain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5526. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 552Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 552
552554555555
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 552555
1. Introduction
It is well known that tissue acidosis produces pain. For instance,direct application of an acidic solution into the skin induces non-adapting pain (Steen et al., 1992, 1995a,b; Steen & Reeh, 1993; Reeh &Steen, 1996; Steen et al., 1996; Ugawa et al., 2002; Jones et al., 2004).During inflammation, protons are among the first mediators releasedby damaged cells, and tissue acidosis is a common factor found inseveral other pain-generating conditions such as ischemia, hemato-ma, fracture, tumor development, skin and muscle incision followingsurgical procedure, arthritis or urinary tract and digestive diseases.The algogenic effects of acidic pH are caused by the depolarization of

550 E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
the peripheral free terminals of nociceptive neurons, which detectnoxious stimuli. This depolarization is caused by activation of ionicchannels directly gated by protons. These channels are “chemo-electrical” transducers that belong to two families of ionic channels,Transient Receptor Potential channels (TRPs), with a preeminent rolefor the Transient Receptor Potential Vanilloid receptor type 1(TRPV1), and Acid-Sensing Ion Channels (ASICs). ASICs are particu-larly sensitive to protons, with somemembers able to activate for verysmall acidification (e.g., from physiological pH7.4 to pH7.2) and togenerate a sustained depolarizing current compatible with thedetection of non-adapting pain (Lingueglia et al., 1997; Waldmannet al., 1997a; Yagi et al., 2006; Deval et al., 2008). ASICs are also widelyexpressed in the central nervous system (CNS) where they participatein the transmission and modulation of pain signals at the level of thespinal cord. This review will focus on recent advances in ourunderstanding of the role of ASICs in pain, both at the central and theperipheral level, supporting these channels as new potent therapeutictargets for the relief of pain.
2. ASICs: from gene to pH-sensor
The first ASICs (initially called MDEG, BNC1 or BNaC1-2) have beencloned in the late 90's (Price et al., 1996;Waldmann et al., 1996; Garcia-Anoveros et al., 1997), and thereafter identified as proton-gatedchannels by Michel Lazdunski's group (Waldmann et al., 1997b). Theybelong to the epithelial sodium channel (ENaC)/degenerin (DEG)superfamily of ion channels (Kellenberger & Schild, 2002). In rodents,at least six ASIC subunits (ASIC1a, ASIC1b, ASIC2a, ASIC2b, ASIC3 andASIC4, see Fig. 1) encoded by four different genes (ACCN1-4, note thatACCN1 and ACCN2 encode for ASIC2 and ASIC1, respectively) have beendescribed. ASIC1 and ASIC2 both have two splice variants named ASIC1a(Garcia-Anoveros et al., 1997;Waldmannet al., 1997b), ASIC1b (Chen etal., 1998a; Bassler et al., 2001), ASIC2a (Price et al., 1996;Waldmann etal., 1996; Garcia-Anoveros et al., 1997) and ASIC2b (Lingueglia et al.,1997). In humans, the ASIC1b mRNA has not been described so far, andASIC3 (Waldmann et al., 1997a; de Weille et al., 1998; Ishibashi &Marumo, 1998; Babinski et al., 1999) and ASIC4 (Akopian et al., 2000;Grunder et al., 2000) have three and two splice variants, respectively(NCBI database, accession numbers NM_004769, NM_020321 andNM_020322 for ASIC3, and NM_018674 and NM_182847 for ASIC4),differing in their C-terminal domain (ASIC3), and in the extracellularloop (ASIC4). The physiological relevance and the properties of theseASIC3 and ASIC4 human splice variants remain to be established.
Fig. 1. A) Schematic representation of an ASIC subunit. B) Trimeric organization
Each ASIC subunit has two hydrophobic transmembrane domainsflanking a large extracellular loop (Fig. 1). Resolution of the crystalstructure of the chicken ASIC1a channel has shown that three subunitsare necessary to form a functional channel (Jasti et al., 2007; Gonzaleset al., 2009). The ASIC2b and ASIC4 subunits do not form functionalhomomeric H+-gated channels (Lingueglia et al., 1997; Akopian et al.,2000; Grunder et al., 2000). ASIC2b can associate with other ASICsubunits to confer important functional properties on the heteromericchannels (Lingueglia et al., 1997; Deval et al., 2004). The role of ASIC4is still not known, although heterologously expressed ASIC4 istargeted to the cell membrane and can associate with ASIC1a andASIC3 to down regulate their expression (Donier et al., 2008). ASICscan also be part of macromolecular complexes containing accessoryproteins, which are important for controlling both functionalproperties and subcellular localization of the channels (Anzai et al.,2002; Baron et al., 2002a; Duggan et al., 2002; Hruska-Hageman et al.,2004, 2002; Deval et al., 2004; Price et al., 2004; Donier et al., 2005;Gao et al., 2005; Deval et al., 2006; Zha et al., 2006; Chai et al., 2007;Wetzel et al., 2007; Donier et al., 2008; Schnizler et al., 2009).
ASICs are voltage-independent, depolarizing cationic channelsmainly permeable to Na+ ions. Homomeric ASIC1a has also beenreported to conduct Ca2+ ions (Waldmann et al., 1997b). However,measures of the relative calcium permeability vary among studies(PNa/PCa between 2.5 and 50) (Waldmann et al., 1997b; Bassler et al.,2001; Yermolaieva et al., 2004) and a recent work strongly argues thatneither the human nor the chick ASIC1a channels are able to conductmeasurable Ca2+ when external Na+ and Ca2+ concentrations arekept within the physiological range (Samways et al., 2009). Theincrease in intracellular free Ca2+ concentration observed in neuronsfollowing activation of ASIC1a channels (Xiong et al., 2004; Yermo-laieva et al., 2004; Herrera et al., 2008; Samways et al., 2009) mightthus be mediated by the secondary activation of voltage-gated Ca2+
channels and Ca2+ release from intracellular stores (Zha et al., 2006).ASIC currents activate transiently upon extracellular acidification
(Fig. 2), the activation threshold, pH sensitivity and current kineticsdepending on the subunit composition of the channel. Such heteroge-neity of ASICs allows for a large functional diversity associated withdifferent types of responses to protons in neurons. Among ASICs, ASIC1and ASIC3 are the most sensitive to protons, being activated by verysmall acidifications. Indeed, they sense ΔpH as small as −0.4 and−0.2units from thephysiological pH7.4, respectively,withpH0.5 for activationaround 6.5 for both ASIC1 and ASIC3 (Yagi et al., 2006; Lingueglia, 2007;Deval et al., 2008). Conversely, ASIC2a currents are activated by morepronounced acidification with activation thresholds close to pH6.0
of a functional ASIC channel (TM1 and 2: transmembrane domains 1 and 2).

A
8 7 6 5
0pH
1.0
0.2
0.4
0.6
0.8
physiologicalphysiologicalpHpH
ASIC1a
ASIC3
I/Imaxactivationinactivation
I/Imax
pH7.6 7 .4 7.2 7 .0 6.8 6 .6
0
0.1
0.2
0.3
control
Arach. acidCB
pHpH 7.07.0
no window current(ASIC1a)
pHpH 7.07.0
window current(ASIC3)
Fig. 2. ASIC3 window current. A) Schematic representation of the pH-dependent activation and inactivation curves of ASIC1a and ASIC3 currents. B) Moderate extracellularacidification (holding potential of −80 mV) only induces a transient current for ASIC1a (no window current) whereas it evokes an additional sustained current that does notinactivate for ASIC3 (window current due to the overlap between the activation and inactivation of the channel as shown in A ; scale bars: 500 pA/2 s). C) The ASIC3 window currentis potentialized by arachidonic acid, the precursor of inflammatory lipids.
551E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
(Lingueglia et al., 1997; Baron et al., 2001; Baron et al., 2002a) and pH0.5
between 4.1 and 5.0 (Lingueglia, 2007; Lingueglia & Lazdunski, 2009). Inrodents, ASICs arewidely present in thenervous system, central neuronsexpressing the ASIC1a and ASIC2 subunits (Price et al., 1996;Waldmannet al., 1996; Garcia-Anoveros et al., 1997; Lingueglia et al., 1997;Waldmann et al., 1997b; Baron et al., 2002b;Wemmie et al., 2003, 2002;Wu et al., 2004; Baron et al., 2008) whereas peripheral sensory neuronsdisplay most of the ASIC subunits (Waldmann et al., 1997a; Chen et al.,1998b; Voilley et al., 2001; Mamet et al., 2002; Poirot et al., 2006). It isinteresting to note that the neuronal expression of ASIC1b andASIC3 hasbeen reported tobe specific to sensoryneurons in rodents (Waldmannetal., 1997a; Chen et al., 1998b).
ASIC3 channels are of particular interest because they generate abiphasic current in response to extracellular acidosis (Fig. 2). Inaddition to the rapidly inactivating peak current, ASIC3 has asustained current that lasts as long as the extracellular pH remainsacid (Waldmann et al., 1997a; Deval et al., 2003; Yagi et al., 2006;Salinas et al., 2009). For modest acidifications of the extracellularmedium (7.4NpHN6.5), the persistent activity of ASIC3 is due to anincomplete inactivation of the current. At these pH values, a windowcurrent is generated by the overlap between the pH-dependentactivation and inactivation curves of the channel (Fig. 2). This windowcurrent is potentiated by pro-inflammatory molecules and canaccount up to 30% of the maximal peak current (Deval et al., 2008).More severe acidifications (below pH6.0) also activate a sustainedASIC3 current but through a different specific mechanism (Salinas etal., 2009). ASIC3 is thus able to elicit a sustained response over a widerange of acidic pH, except in the pH interval 6.0–6.5. This sustainedcurrent produces persistent depolarizations of the neuronal mem-brane that are compatible with the perception of non-adapting painelicited by moderate and strong acidifications (Steen et al., 1995a;Mantyh et al., 2002; Woo et al., 2004). It is interesting to note thatsevere tissue acidosis (below pH6.0) can occur under certainpathological conditions such as gastritis and peptic ulcer (Holzer,2010), cystitis or bone cancer and bone metastases (Mantyh et al.,2002).
3. Pharmacology of ASICs
The pharmacology of ASICs is still relatively poor compared forinstance with voltage-dependent sodium (NaV) channels (Denac et al.,2000). Both positive modulators and inhibitors of ASICs have beenreported. Potentiators include some metal ions and neuropeptidesrelated to FMRFamide and opioid peptides. Inhibitors of ASICs includenon-specific and non-discriminativemolecules aswell as highly specificnatural peptide toxins isolated from animal venoms (Diochot et al.,2007). ASICs are also modulated by temperature. Cold temperaturespotentiate the acid-activated currents by slowing the rate of desensi-tization (Askwith et al., 2001). Temperature also modulates proton-gated currents in sensoryneurons, either by slowing (cold)or increasing(warm) the rate of desensitization (Askwith et al., 2001; Neelands et al.,2010), which could regulate the transduction of sensory stimuli incertain physiological or pathophysiological conditions.
3.1. Neuropeptides
ASICs belong to the same family of channels as the invertebrateFMRFamide-gated Na+ channel (FaNaC) (Lingueglia et al., 1997,2006), and the structural similarity between these channels led to thehypothesis that FMRFamide and structurally related peptides could beagonists of ASICs (Askwith et al., 2000). FMRFamide, neuropeptide FF(NPFF, FLFQPQRFamide) and neuropeptide SF (NPSF, SLAAPQRFa-mide) do not directly activate ASICs but they potentiate the current ofrecombinant ASIC1a and ASIC3 channels (EC50~10–50 μM) (Askwithet al., 2000; Catarsi et al., 2001; Deval et al., 2003; Chen et al., 2006b),the effects being more pronounced on ASIC3 currents (Lingueglia etal., 2006). Indeed, the effect of these peptides on native current isstrongly dependent on the presence of the ASIC3 subunit (Xie et al.,2002, 2003). FMRFamide and related peptides seems to act directly onthe channel (Askwith et al., 2000; Deval et al., 2003) to increase thepeak current and to mainly slow down the inactivation rate, whichcauses a pronounced sustained phase during acidification. Contrary toFMRFamide that is only present in invertebrates, NPFF and NPSF are

552 E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
largely expressed in the CNS of mammals, especially in the spinal cord(Panula et al., 1999; Vilim et al., 1999). NPFF has also been detected insmall and intermediate-sized DRG neurons (Allard et al., 1999).Interestingly, the expression levels of both the NPFF precursor andASIC subunits are increased in inflammatory conditions (Vilim et al.,1999; Voilley et al., 2001; Duan et al., 2007). The positive modulationof ASIC3-mediated sustained depolarization by NPFF could thereforeplay a role in regulating nociceptor excitability under inflammatoryconditions (Deval et al., 2003). More recently, dynorphin A and bigdynorphin, two endogenous opioid neuropeptides, have been shownto directly bind to ASIC1a to enhance channel activity by preventingsteady-state desensitization (Sherwood and Askwith, 2009), andASIC1a could constitute a novel nonopioid receptor target fordynorphin action in the CNS.
3.2. Peptide toxins from animal venoms
Two peptide toxins, PcTx1 and APETx2, isolated from the venomsof the spider Psalmopoeus cambridgei (Escoubas et al., 2000) and thesea anemone Anthopleura elegantissima (Diochot et al., 2004), havebeen shown to specifically inhibit ASIC1a and ASIC3 channels,respectively (Diochot et al., 2007). PcTx1 is a 40-amino acids peptidethat specifically blocks ASIC1a homomeric channels with an IC50 of~1nM (Escoubas et al., 2000), and that has potent analgesic propertieswhen injected intrathecally (i.t.) or intracerebroventricularly (i.c.v.) inrodents (Duan et al., 2007; Mazzuca et al., 2007) (see section 5).APETx2, a 42-amino acid peptide, inhibits ASIC3-containing channels(ASIC3, ASIC3/1a, ASIC3/1b, ASIC3/2b) with an affinity ranging from63 nM to 2 μM depending on the subunit composition of theheteromeric channel (Diochot et al., 2004). APETx2 also has analgesicproperties when applied peripherally (Deval et al., 2008) (seesection 4-1). Although the mechanism of action of PcTx1 on ASIC1ahas been well characterized, the binding site for APETx2 on ASIC3 andthe mechanism of inhibition are not documented yet. PcTx1 binds tothe extracellular loop of homomeric ASIC1a at a site close to the“acidic pocket” (involved in pH sensing) at the interface between twosubunits in the trimeric channel (Qadri et al., 2009; Salinas et al.,2006), and stabilizes the channel (Qadri et al., 2009). The toxin acts asa gating modifier that binds with higher affinity to the inactivatedstate of the channel, robustly increasing its apparent affinity forprotons and shiftingmost channels into the inactivated state at pH 7.4(Chen et al., 2005). PcTx1 also interacts with ASIC1b but instead ofinhibiting the channel, it promotes its opening in response to slightacidification because the toxin binds more efficiently to the open stateof the channel (Chen et al., 2006a).
3.3. Organic compounds
Amiloride, a potassium-sparing diuretic, was the first blocker ofASICs to be described (Waldmann et al., 1996, 1997a,b). It is however aweak and non-selective ASIC inhibitor that also blocks other ionicchannels and exchangers (Frelin et al., 1988; Kleyman & Cragoe, 1988).Amiloride acts as a non-discriminative pore blocker of ASICs at themicromolar range of concentrations (IC50 ranging between 5 and100 μM). Amiloride derivatives benzamil and EIPA are also poorly-selective, reversible blockers of ASICs. The A-317567 compound appearsas amore selective but still non-discriminative, non-amiloride inhibitorof native and recombinant ASICs (IC50 ranging between 2 and 30 μM)(Dube et al., 2005). This blocker of ASICs reproduces withmore potencythe analgesic effects of amiloride in rat models of inflammatory andpost-operative pain (Dube et al., 2005). The amidine moiety of A-317567 is critical for the inhibitory effect on ASIC3 current, and anevaluation of a series of indole amidines modified at the 2-position ofthe indole ring led to the discovery of a more potent ASIC3 blocker(IC50=133 nM) (Kuduk et al., 2009). Very recently, four anti-protozoaldiarylamidines (DAPI, diminazene, HSB and pentamidine) have been
reported to potently inhibit ASIC currents in primary cultures ofhippocampal neurons with apparent affinities ranging from 300 nM to38 μM (Chen et al., 2010), thus constituting a novel class of ASICblockers.
Besides their well-known ability to inhibit cyclooxygeneases(COX), non-steroid anti-inflammatory drugs (NSAIDs) also inhibitthe activity of both ASIC1a and ASIC3 channels (Voilley et al., 2001).Whereas ASIC1a is inhibited by flurbiprofen and ibuprofen, thesustained current of ASIC3 is blocked by salicylic acid, aspirin anddiclofenac. Inhibition occurs with relatively low affinity but, never-theless, in the range of therapeutic concentrations of NSAIDs (IC50ranging from 92 to 249 μM) (Voilley et al., 2001). A proposedmechanism of action is allosteric inhibition of ASICs by slowing downthe recovery after inactivation (Dorofeeva et al., 2008).
3.4. Bi- and trivalent cations
ASICs are inhibitedbyavarietyof heavymetal ions (Gd3+, Pb2+,Ni2+,Cd2+, Cu2+) (Babinski et al., 2000; Allen & Attwell, 2002; Wang et al.,2006, 2007; Staruschenko et al., 2007) and bivalent cations (Ca2+,Mg2+
and Zn2+) (Waldmann et al., 1997b; Baron et al., 2001; de Weille &Bassilana, 2001; Babini et al., 2002; Immke&McCleskey, 2003; Chu et al.,2004; Paukert et al., 2004). Both the ASIC1a and the ASIC2a single-channel conductances are reduced by elevated levels of extracellularcalcium (de Weille & Bassilana, 2001). It has been proposed that thebinding of calcium ions to a high affinity site on the extracellular side oftheporeofASIC3blockspermeationof thechannel (Immke&McCleskey,2003). Thesite thatmediates this blockhas been identified inASIC1a, andcalcium inhibits the channel in the open state (IC50=3.9 mM at pH 5.5)with a better affinity than for ASIC1b (IC50=10 mM at pH 4.7) (Paukertet al., 2004).
Interestingly, Zn2+ has a dual effect on ASICs with a potentiatingeffect at micromolar concentrations (EC50~100 μM) and an inhibitoryeffect at nanomolar concentrations (Baron et al., 2001; Chu et al.,2004). The low-affinity potentiating effect of Zn2+ affects ASIC2a-containing channels (Baron et al., 2001) whereas the high-affinityinhibitory effect is observed on both homomeric ASIC1a andheteromeric ASIC1a/2a channels (Chu et al., 2004). High concentra-tion of Zn2+ (i.e., 300 μM) has also been reported to inhibit ASIC3current (Poirot et al., 2006). Synaptically released Zn2+ could thusmodulate ASICs and the neuronal excitability during the acidificationcaused by synaptic activity (Miesenbock et al., 1998).
4. Peripheral ASICs in pain
One of the first experimental evidence for the existence of acid-activated cation channels was provided in the early 80's by studiesperformed on sensory neurons isolated from dorsal root andtrigeminal ganglia (DRG and TG) (Krishtal & Pidoplichko, 1980,1981a,b). These studies have raised the question of a possible role forthese proton-sensitive channels in nociception (Krishtal & Pido-plichko, 1981b). It is now clear that ASICs support the proton-activated currents identified by Krishtal and colleagues in DRG and TGneurons. Almost all ASIC subunits are expressed in sensory neurons ofthe trigeminal, vagal and dorsal root ganglia, including small andmedium-diameter neurons involved in nociception, consistent with arole for these channels in pain detection (Waldmann et al., 1997a,b;Chen et al., 1998b; Liu & Simon, 2000; Bassler et al., 2001; Garcia-Anoveros et al., 2001; Voilley et al., 2001; Alvarez de la Rosa et al.,2002; Chen et al., 2002; Page et al., 2004, 2005; Fukuda et al., 2006;Poirot et al., 2006). The channels have been detected in the soma andin the peripheral terminals of DRG neurons (Price et al., 2000, 2001;Garcia-Anoveros et al., 2001; Alvarez de la Rosa et al., 2002), but seemto be absent (at least for ASIC1a and ASIC2a) from the centralterminals in the dorsal horn of the spinal cord (Garcia-Anoveros et al.,2001; Duan et al., 2007).

553E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
4.1. Cutaneous somatic pain
Despite their properties and their localization in nociceptors, therole of ASICs in sensing cutaneous pain has remained unclear up torecently. This is in large part due to the absence of a clear painphenotype in knockout mice for ASIC1a, ASIC2 and ASIC3 (Price et al.,2000, 2001; Chen et al., 2002; Page et al., 2004). Although a bluntedresponse of C-fibers to pH5.0 (Price et al., 2001), ASIC3 knockout micehave no modification of their response to injection of acidic solutions(acetic acid pH 3.0) into the skin of the hind-paw and no modificationof the inflammatory thermal hyperalgesia after carrageenan injectioninto the hind paw (Price et al., 2001; Chen et al., 2002). Moreover,ASIC3 knockout mice display a reduced latency to the onset of painresponses, or more pain-related behaviors, when stimuli of moderateto high intensity are used (Chen et al., 2002). Similarly, the transgenicexpression of an ASIC3 dominant-negative subunit in mice leads to anincreased sensitivity to acidic, mechanical and inflammatory stimuli(Mogil et al., 2005). In ASIC2 null mice, both the acid-evoked currentsin cultured sensory neurons and the response of nociceptors to acidare normal (Price et al., 2000). Finally, the deletion of ASIC1a or ASIC3neither alters C-fiber responses to mechanical stimuli, nor does itchange mice paw withdrawal responses to mechanical and thermalstimuli (Price et al., 2001; Page et al., 2004). Mechanosensitivity of theAβ- and Aδ-fibers are, however, altered in ASIC3 knockout mice whencompared to wild type littermates, Aβ-fibers being more sensitivewhereas the sensitivity of Aδ-fibers is reduced (Price et al., 2001).Altogether, these data were not supporting a clear role for ASICs inmediating cutaneous pain associated with tissue acidosis in mice.
On the other hand, data obtained in human and rat usingpharmacological inhibitors of ASICs clearly involve these channels insensing cutaneous acidic pain (Ugawa et al., 2002; Jones et al., 2004;Dube et al., 2005; Deval et al., 2008). Acid-induced cutaneous pain inhealthy human volunteers is blocked by amiloride and/or NSAIDs(Ugawa et al., 2002; Jones et al., 2004). The specific ASIC blocker A-317567 (see section 3-3) inhibits inflammatory thermal hyperalgesiaand post-operative pain in rats (Dube et al., 2005). Supporting theseexperiments, local peripheral inhibition of ASICs by the non selectiveblockers amiloride and benzamil reduces cutaneous inflammatory painin rats (Rocha-Gonzalez et al., 2009). Selective peripheral inhibition ofASIC3 by the specific peptide toxin APETx2 and in vivo knockdownof thechannelwith a siRNA (Deval et al., 2008) lead to a clear demonstration ofthe role of ASIC3 in cutaneous pain in rats. ASIC3 is largely expressed inrat skin nociceptors where it acts as a sensor of acid-induced cutaneouspain and integrates multiple molecular signals produced duringinflammation to contribute to primary thermal inflammatory hyper-algesia (i.e., the increased response to thermal stimuli at the site ofinjury) (Deval et al., 2008). This role is fully consistentwith the increasedexpression of ASICs measured in rat DRG following hindpawinflammation (Voilley et al., 2001; Mamet et al., 2002, 2003), and thepost-translational up-regulation of ASIC3 channel activity by severalcomponents of the “inflammatory soup”, such as bradykinin, 5-hydroxytryptamine (Deval et al., 2004), hypertonicity (Deval et al.,2008), arachidonic acid (Allen & Attwell, 2002; Smith et al., 2007; Devalet al., 2008) and nitric oxide (Cadiou et al., 2007). ASIC3 channels havetherefore been proposed to participate in the peripheral sensitization ofcutaneous nociceptors during inflammation (Mamet et al., 2002, 2003;Deval et al., 2008). Among theASIC, ASIC3-containing channels appear tohave a preponderant role in sensing acidic cutaneous pain in rodentssince inhibition of ASIC1a homomeric channels, by subcutaneousinjections of the specific toxin blocker PcTx1, has no effect in mousemodels of acutepain (Mazzucaet al., 2007)or in amodel of inflammatorypain in rat (Duan et al., 2007; Deval et al., 2008). However, one cannotexclude a role of the ASIC1a subunit at the periphery when associatedwith ASIC3 in heteromeric channels that are not blocked by PcTx1.
It is interesting to note the discrepancy between the rat andmousedata about the role of ASIC3 in cutaneous pain. This could be explained
by different parameters including compensation in knockout animalsor species differences in the basal expression level of ASICs. Forinstance, it has been reported that transient proton-induced currents(i.e., ASIC currents) are less frequent and have smaller amplitudes inmouse than in rat sensory neurons and nociceptors (Leffler et al.,2006; Deval et al., 2008). Differences between rats andmice could alsobe due to differences in pain behaviours or in the characteristics of theexperimental pain models in these two species (Wilson & Mogil,2001; Mogil, 2009). For instance, unlike in rats, no role has beendemonstrated for ASIC3 in the acute phase (i.e., after a few hours) ofinflammation in knockout mice (Price et al., 2001; Chen et al., 2002;Mogil et al., 2005; Yen et al., 2009), but analysis of the sub-acute phase(i.e., after 1–2 days) reveals an implication of the channel in themaintenance of primary hyperalgesia (Yen et al., 2009).
4.2. Deep somatic pain
More than 50% of small skeletal muscle afferents in rat (Molliver etal., 2005) and more than 30% of DRG neurons innervating the kneejoint in mouse (Ikeuchi et al., 2009) express ASIC3 subunit.Furthermore, ASIC2 and/or ASIC3 expression in DRG neurons isincreased in mouse models of muscle inflammation (Walder et al.,2010) and acute arthritis (Ikeuchi et al., 2009). ASIC3 null mice haveclearly demonstrated a role for this channel in pain arising from thedeeper tissues such as muscles or joints (Price et al., 2001; Sluka et al.,2003, 2007; Ikeuchi et al., 2008, 2009;Walder et al., 2010). ASIC3 (butnot ASIC1a) knockout mice do not develop secondary mechanicalhyperalgesia (i.e., an increased mechanical response outside the siteof injury) caused by repeated intramuscular acid injection (Sluka etal., 2003) or muscle inflammation (Sluka et al., 2007). Thus, ASIC3 inprimary afferent fibers innervating muscle is critical for thedevelopment of secondary hyperalgesia that results from muscleinsult (Sluka et al., 2007). Peripherally located ASIC1a channels arenot involved in secondary mechanical hyperalgesia but contribute toprimary hyperalgesia induced by muscle inflammation in mice(Walder et al., 2010), suggesting differential roles for ASIC1a andASIC3 in muscle pain. Experiments performed in mouse models ofarthritis and joint pain have shown a similar involvement of ASIC3 inthe secondary, but not the primary, mechanical hyperalgesiaproduced by inflammation (Ikeuchi et al., 2008). These resultsconfirm the important role of ASIC3 located on the primary afferentfibers innervating deep tissues in sensing pain caused by acidosis, thusincreasing the nociceptive input to the spinal cord resulting in centralsensitization and secondary hyperalgesia (Ikeuchi et al., 2008). Ingood agreement with this, activation of rat peripheral nociceptors byacid stimuli has been reported to induce central sensitization at thelevel of the dorsal spinal cord (Carpenter et al., 2001).
4.3. Visceral pain
4.3.1. Gastrointestinal pain and mechanoperceptionGastric acid is a noxious stimulus that contributes to pain arising
from the esophagus, stomach and upper small intestine (Wultsch etal., 2008). In rats, all gastric DRG neurons and ~55% of nodose ganglia(NG) neurons exhibit transient, amiloride-sensitive, ASIC-like cur-rents (Sugiura et al., 2005). The properties of theses currents arealtered in case of gastric ulcers with increased pH sensitivity, currentdensities and changes in current kinetics. Conversely to ASIC2, ASIC3plays a major role in the gastric acid hyperresponsiveness associatedwith experimental gastritis (Wultsch et al., 2008).
A study performed in human tissues has revealed the expression ofASIC1, ASIC2 andASIC3 subunits in intestine, sensory ganglia andnerves(Yiangou et al., 2001). Moreover, it shows both an increasedimmunoreactive signal and increased number of ASIC3-positiveneurons in the myenteric and sub-mucous plexus of inflamed intestinefrom patients with Crohn's disease (an inflammatory bowel disease;

554 E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
(Yiangou et al., 2001)). qPCR analysis of neurons isolated by laser-capturemicrodissection has identified ASIC3 as themost abundant ASICtranscript in mouse colonic DRG neurons (Hughes et al., 2007), in goodagreement with its contribution to the detection of noxiousmechanicalstimuli in mice colon (Jones et al., 2005). Interestingly, ASIC3 has beeninvolved in inflammation-induced sensitization of colon afferent fibersto stretch (Jones et al., 2005), and in the development of non-inflammatory visceral hypersensitivity (Jones et al., 2007).
Visceral mechanoreceptors are critical for perceived sensations andautonomic reflex control of gastrointestinal function, and reducingcolonic mechanical hypersensitivity is an important potential strategyfor reducing visceral pain (Hughes et al., 2007). Mice lacking ASIC1a,ASIC2 and ASIC3 subunits show altered colonic mechanosensoryfunction, implicating ASICs in the mechanotransduction process (Pageet al., 2005). Loss of ASIC1a increases mechanosensitivity in all colonicand gastroesophageal mechanoreceptor subtypes, and it prolongs thegastric emptying time (Page et al., 2004). ASIC3 makes a positivecontribution tomechanosensitivity in three out of four classes of visceralafferents (Page et al., 2005), whereas the role of ASIC2 widely differsacross subclasses of afferents and regions of the gut, with a negativeinfluence in lower gut afferents and a positive influence in upper gutafferents (Page et al., 2005). The role of ASIC2 in visceral mechan-osensitivity is, however, not entirely clear, and colonic mechanonoci-ception measured by stretch-induced CGRP release is not affected inASIC2 knock-out mice (Roza et al., 2004). Interestingly, the generalcontribution of ASICs to mechanoreceptor function in visceral fibers ismuch larger and significantly different than the one observed in skin,where ASIC3 and possibly ASIC2 have some contribution to normaltouch sensation (Price et al., 2000, 2001; Chen et al., 2002). However,available data in colon and skin do not favor a direct role for ASICs inmechanotransduction (Drew et al., 2004; Roza et al., 2004).
4.3.2. Heart and lung nociceptionSensory neurons located in the nodose and dorsal root ganglia
innervating the heart sense ischemia and mediate angina. EdwinMcCleskey's group has reported a high level of ASIC currents in theseneurons and has proposed a critical role for acidity in the sensationassociated with myocardial ischemia (Benson et al., 1999). They haveshown that recombinant ASIC3 and ASIC3/2b channels reproduce thefunctional features of the native channel that underlie the large acid-evoked current in cardiac afferents (Sutherland et al., 2001). Forinstance, both recombinant ASIC3 and native channels are opened atpH 7.0, a value reached in the first few minutes of a heart attack.Moreover, cardiac sensory neurons from rat DRG exhibit a sustainedcurrent when pH decreases from 7.4 to 7.0 that is characteristic of thewindow current observed with the recombinant homomeric ASIC3(Fig. 2) and heteromeric ASIC3/2b channels (Yagi et al., 2006). Inmouse, the current properties of cardiac dorsal root ganglia neuronshave been recently shown to more closely match the properties ofheteromeric ASIC3/2a channels (Hattori et al., 2009). Interestingly,lactic acid that is produced during cardiac ischemia and has beensuggested to trigger angina chest pain, dramatically increases ASIC3current (Immke &McCleskey, 2001a,b). ASIC3 has thus been proposedto be the sensor of myocardial acidity that triggers cardiac pain(Sutherland et al., 2001).
Functional ASIC-like currents have been recorded on rat vagalpulmonary sensory neurons (Gu & Lee, 2006), and ASIC3 is expressedin rat vagal and glossopharyngeal sensory ganglia (Fukuda et al.,2006), as well as in spinal afferent neurons projecting to the rat lungand pleura (Groth et al., 2006). Cough reflex is caused by actionpotential discharge in vagal sensory nerves, and the most commontussive stimuli include mechanical perturbations, anosmotic solu-tions, various chemical agents and acidic solutions (Kollarik & Undem,2002). Citric acid-evoked coughing in guinea pigs may involvesequential activation of ASICs and chloride channels (Canning et al.,2006). ASICs could therefore participate in the response to airway
acidification, such as cough and bronchoconstriction (Kollarik &Undem, 2002; Canning et al., 2006) that are associated with bothphysiological (e.g., exercise) and pathophysiological (e.g., chronicobstructive pulmonary disease) conditions.
4.4. ASICs and TRPs
As previously mentioned, ASICs are not the only proton-activatedcation channels expressed by nociceptors, and a role for TRPV1 in theresponse of sensory neurons to protonshas also been reported (Caterinaet al., 2000). Some evidence suggests that ASICs and TRPV1 servecomplementary roles in proton sensitivity of sensory neurons. First, thedistribution of both types of channels is significantly different, with lessthan 50% overlap in rat DRG (Molliver et al., 2005; Ugawa et al., 2005).Second, the two channels are not activated at the same range of pH,TRPV1 being activated by more severe acidifications (pH0.5 of 5.4,(Tominaga et al., 1998)) as compared with most ASICs. In goodagreement with this point, the pain provoked by skin acidifications inhealthy human volunteers is relieved by pharmacological blockers ofASICs (i.e., amiloride) andTRPV1 (i.e., capsazepine) (Ugawa et al., 2002),but ASICs are the main mediators of pain caused by moderate acidity(NpH6.0) while TRPV1 contributes, together with ASICs, to pain elicitedby more severe acidification (bpH6.0). In addition, the extent of therespective roles of ASICs and TRPV1 seems to vary between species(Leffler et al., 2006), and also probably between tissues, within tissues,or under different pathophysiological states. Other TRPs like TRPA1 andTRPV4 also contribute to peripheral pain in response to inflammation(White et al., 2010). Itwill be therefore interesting to further investigatethe functional relationships and the respective contributionof ASICs andTRPs, regarding the increasing role of these two types of ion channels insensory perception and pain.
5. Central ASICs in pain
Central neurons, including those involved in pain signal transmis-sion (i.e., second-order neurons located in the spinal dorsal horn),largely display ASIC1a and ASIC1a/2a currents (Baron et al., 2002b;Wemmie et al., 2002; Wemmie et al., 2003; Wu et al., 2004; Duan etal., 2007; Baron et al., 2008). Neurons in the spinal dorsal horn areintegrators of peripheral sensory-nociceptive inputs and transmit thenociceptive message to upper central levels and brain (Fig. 3). Inaddition to the high expression level of ASIC1a, ASIC2a and ASIC2bsubunits in rat dorsal horn neurons (Baron et al., 2008), theexpression of both ASIC1a and ASIC2a is increased after peripheralinflammation (Wu et al., 2004). The particular biophysical propertiesof ASIC1a and ASIC1a/2a currents (i.e., pH-dependent reactivation)have been proposed to modulate the excitability of spinal neuronsduring intense synaptic activity (Baron et al., 2008) due to chronicpain. Together, these data strongly argue for a participation of ASICs incentral nociceptive transmission and/or modulation, both underphysiological and pathophysiological conditions (Wu et al., 2004;Duan et al., 2007; Baron et al., 2008). ASIC1a and ASIC2 subunits arepresent in the postsynaptic membrane of hippocampal neurons(Hruska-Hageman et al., 2002; Wemmie et al., 2002; Zha et al.,2009), where ASIC1a contributes to synaptic plasticity (Wemmie etal., 2002). It is thus conceivable to speculate a similar synapticlocalization of ASICs in the spinal cord, where synaptic plasticityprocesses are relevant to central sensitization (Basbaum et al., 2009).Indeed, ASIC1a has been involved in two forms of central sensitizationat the level of the spinal dorsal horn (Duan et al., 2007), and thepharmacological block of ASIC1a activity by the specific toxin PcTx1injected intrathecally produces strong analgesic effects in severalrodent models of acute and chronic pain (Duan et al., 2007; Mazzucaet al., 2007). The genetic knockdown of ASIC1a expression byintrathecal delivery of antisense oligonucleotides produces similaranalgesic effects (Duan et al., 2007; Mazzuca et al., 2007). These
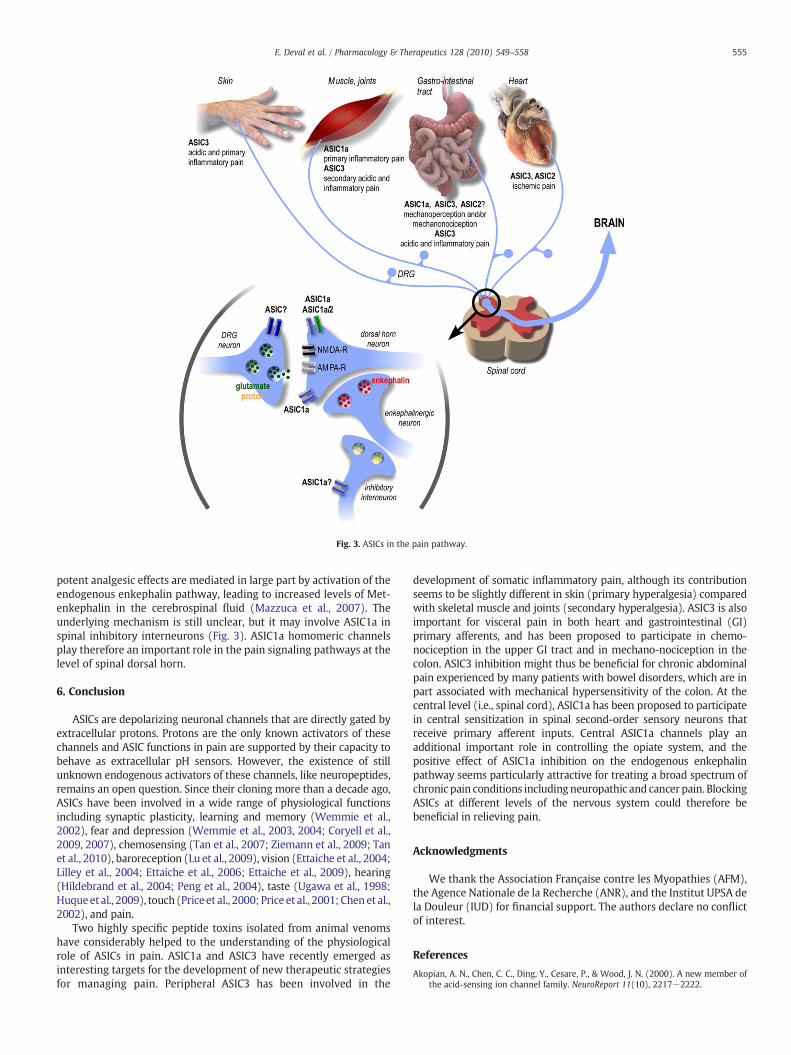
Fig. 3. ASICs in the pain pathway.
555E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
potent analgesic effects are mediated in large part by activation of theendogenous enkephalin pathway, leading to increased levels of Met-enkephalin in the cerebrospinal fluid (Mazzuca et al., 2007). Theunderlying mechanism is still unclear, but it may involve ASIC1a inspinal inhibitory interneurons (Fig. 3). ASIC1a homomeric channelsplay therefore an important role in the pain signaling pathways at thelevel of spinal dorsal horn.
6. Conclusion
ASICs are depolarizing neuronal channels that are directly gated byextracellular protons. Protons are the only known activators of thesechannels and ASIC functions in pain are supported by their capacity tobehave as extracellular pH sensors. However, the existence of stillunknown endogenous activators of these channels, like neuropeptides,remains an open question. Since their cloning more than a decade ago,ASICs have been involved in a wide range of physiological functionsincluding synaptic plasticity, learning and memory (Wemmie et al.,2002), fear and depression (Wemmie et al., 2003, 2004; Coryell et al.,2009, 2007), chemosensing (Tan et al., 2007; Ziemann et al., 2009; Tanet al., 2010), baroreception (Lu et al., 2009), vision (Ettaiche et al., 2004;Lilley et al., 2004; Ettaiche et al., 2006; Ettaiche et al., 2009), hearing(Hildebrand et al., 2004; Peng et al., 2004), taste (Ugawa et al., 1998;Huqueet al., 2009), touch (Price et al., 2000; Price et al., 2001; Chen et al.,2002), and pain.
Two highly specific peptide toxins isolated from animal venomshave considerably helped to the understanding of the physiologicalrole of ASICs in pain. ASIC1a and ASIC3 have recently emerged asinteresting targets for the development of new therapeutic strategiesfor managing pain. Peripheral ASIC3 has been involved in the
development of somatic inflammatory pain, although its contributionseems to be slightly different in skin (primary hyperalgesia) comparedwith skeletal muscle and joints (secondary hyperalgesia). ASIC3 is alsoimportant for visceral pain in both heart and gastrointestinal (GI)primary afferents, and has been proposed to participate in chemo-nociception in the upper GI tract and in mechano-nociception in thecolon. ASIC3 inhibition might thus be beneficial for chronic abdominalpain experienced by many patients with bowel disorders, which are inpart associated with mechanical hypersensitivity of the colon. At thecentral level (i.e., spinal cord), ASIC1a has been proposed to participatein central sensitization in spinal second-order sensory neurons thatreceive primary afferent inputs. Central ASIC1a channels play anadditional important role in controlling the opiate system, and thepositive effect of ASIC1a inhibition on the endogenous enkephalinpathway seems particularly attractive for treating a broad spectrum ofchronic pain conditions including neuropathic and cancer pain. BlockingASICs at different levels of the nervous system could therefore bebeneficial in relieving pain.
Acknowledgments
We thank the Association Française contre les Myopathies (AFM),the Agence Nationale de la Recherche (ANR), and the Institut UPSA dela Douleur (IUD) for financial support. The authors declare no conflictof interest.
References
Akopian, A. N., Chen, C. C., Ding, Y., Cesare, P., & Wood, J. N. (2000). A new member ofthe acid-sensing ion channel family. NeuroReport 11(10), 2217−2222.

556 E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
Allard, M., Rousselot, P., Lombard, M. C., & Theodosis, D. T. (1999). Evidence forneuropeptide FF (FLFQRFamide) in rat dorsal root ganglia. Peptides 20(3), 327−333.
Allen, N. J., & Attwell, D. (2002). Modulation of ASIC channels in rat cerebellar Purkinjeneurons by ischaemia-related signals. J Physiol 543(Pt 2), 521−529.
Alvarez de la Rosa, D., Zhang, P., Shao, D., White, F., & Canessa, C. M. (2002). Functionalimplications of the localization and activity of acid-sensitive channels in ratperipheral nervous system. Proc Natl Acad Sci USA 99(4), 2326−2331.
Anzai, N., Deval, E., Schaefer, L., Friend, V., Lazdunski, M., & Lingueglia, E. (2002). Themultivalent PDZ domain-containing protein CIPP is a partner of acid-sensing ionchannel 3 in sensory neurons. J Biol Chem 277(19), 16655−16661.
Askwith, C. C., Benson, C. J., Welsh, M. J., & Snyder, P. M. (2001). DEG/ENaC ion channelsinvolved in sensory transduction are modulated by cold temperature. Proc NatlAcad Sci USA 98(11), 6459−6463.
Askwith, C. C., Cheng, C., Ikuma, M., Benson, C., Price, M. P., & Welsh, M. J. (2000).Neuropeptide FF and FMRFamide potentiate acid-evoked currents from sensoryneurons and proton-gated DEG/ENaC channels. Neuron 26(1), 133−141.
Babini, E., Paukert, M., Geisler, H. S., & Grunder, S. (2002). Alternative splicing andinteraction with di- and polyvalent cations control the dynamic range of acid-sensing ion channel 1 (ASIC1). J Biol Chem 277(44), 41597−41603.
Babinski, K., Catarsi, S., Biagini, G., & Seguela, P. (2000). Mammalian ASIC2a and ASIC3subunits co-assemble into heteromeric proton- gated channels sensitive to Gd3+. JBiol Chem 275(37), 28519−28525.
Babinski, K., Le, K. T., & Seguela, P. (1999). Molecular cloning and regional distributionof a human proton receptor subunit with biphasic functional properties. JNeurochem 72(1), 51−57.
Baron, A., Deval, E., Salinas, M., Lingueglia, E., Voilley, N., & Lazdunski, M. (2002a).Protein kinase C stimulates the acid-sensing ion channel ASIC2a via the PDZdomain-containing protein PICK1. J Biol Chem 277(52), 50463−50468.
Baron, A., Schaefer, L., Lingueglia, E., Champigny, G., & Lazdunski, M. (2001). Zn2+ and H+ are coactivators of acid-sensing ion channels. J Biol Chem 276(38), 35361−35367.
Baron, A., Voilley, N., Lazdunski, M., & Lingueglia, E. (2008). Acid sensing ion channels indorsal spinal cord neurons. J Neurosci 28(6), 1498−1508.
Baron, A., Waldmann, R., & Lazdunski, M. (2002b). ASIC-like, proton-activated currentsin rat hippocampal neurons. J Physiol 539(Pt 2), 485−494.
Basbaum, A. I., Bautista, D. M., Scherrer, G., & Julius, D. (2009). Cellular and molecularmechanisms of pain. Cell 139(2), 267−284.
Bassler, E. L., Ngo-Anh, T. J., Geisler, H. S., Ruppersberg, J. P., & Grunder, S. (2001).Molecular and functional characterization of acid-sensing ion channel (ASIC) 1b. JBiol Chem 276(36), 33782−33787.
Benson, C. J., Eckert, S. P., &McCleskey, E.W. (1999). Acid-evoked currents in cardiac sensoryneurons: a possiblemediator ofmyocardial ischemic sensation.Circ Res 84(8), 921−928.
Cadiou, H., Studer, M., Jones, N. G., Smith, E. S., Ballard, A., McMahon, S. B., et al. (2007).Modulation of acid-sensing ion channel activity by nitric oxide. J Neurosci 27(48),13251−13260.
Canning, B. J., Farmer, D. G., & Mori, N. (2006). Mechanistic studies of acid-evokedcoughing in anesthetized guinea pigs. Am J Physiol Regul Integr Comp Physiol 291(2),R454−R463.
Carpenter, K. J., Nandi, M., & Dickenson, A. H. (2001). Peripheral administration of lowpH solutions causes activation and sensitisation of convergent dorsal hornneurones in the anaesthetised rat. Neurosci Lett 298(3), 179−182.
Catarsi, S., Babinski, K., & Seguela, P. (2001). Selective modulation of heteromeric ASICproton-gated channels by neuropeptide FF. Neuropharmacology 41(5), 592−600.
Caterina, M. J., Leffler, A., Malmberg, A. B., Martin, W. J., Trafton, J., Petersen-Zeitz, K. R.,et al. (2000). Impaired nociception and pain sensation in mice lacking the capsaicinreceptor. Science 288(5464), 306−313.
Chai, S., Li, M., Lan, J., Xiong, Z. G., Saugstad, J. A., & Simon, R. P. (2007). A kinase-anchoring protein 150 and calcineurin are involved in regulation of acid-sensingion channels ASIC1a and ASIC2a. J Biol Chem 282(31), 22668−22677.
Chen, C. C., England, S., Akopian, A. N., &Wood, J. N. (1998a). A sensory neuron-specific,proton-gated ion channel. Proc Natl Acad Sci USA 95(17), 10240−10245.
Chen, C. C., Zimmer, A., Sun,W.H., Hall, J., & Brownstein,M. J. (2002). A role for ASIC3 in themodulation of high-intensity pain stimuli. Proc Natl Acad Sci USA 99(13), 8992−8997.
Chen, J. J., Vasko, M. R., Wu, X., Staeva, T. P., Baez, M., Zgombick, J. M., et al. (1998b).Multiple subtypes of serotonin receptors are expressed in rat sensory neurons inculture. J Pharmacol Exp Ther 287(3), 1119−1127.
Chen, X., Kalbacher, H., & Grunder, S. (2005). The tarantula toxin psalmotoxin 1 inhibitsacid-sensing ion channel (ASIC) 1a by increasing its apparent H+ affinity. J GenPhysiol 126(1), 71−79.
Chen, X., Kalbacher, H., & Grunder, S. (2006a). Interaction of acid-sensing ion channel(ASIC) 1 with the tarantula toxin psalmotoxin 1 is state dependent. J Gen Physiol127(3), 267−276.
Chen, X., Paukert, M., Kadurin, I., Pusch, M., & Grunder, S. (2006b). Strong modulationby RFamide neuropeptides of the ASIC1b/3 heteromer in competition withextracellular calcium. Neuropharmacology 50(8), 964−974.
Chen, X., Qiu, L., Li, M., Durrnagel, S., Orser, B. A., Xiong, Z. G., et al. (2010). Diarylamidines:high potency inhibitors of acid-sensing ion channels. Neuropharmacology 58(7),1045−1053.
Chu, X. P., Wemmie, J. A., Wang, W. Z., Zhu, X. M., Saugstad, J. A., Price, M. P., et al. (2004).Subunit-dependent high-affinity zinc inhibition of acid-sensing ion channels. JNeurosci 24(40), 8678−8689.
Coryell, M. W., Wunsch, A. M., Haenfler, J. M., Allen, J. E., Schnizler, M., Ziemann, A. E.,et al. (2009). Acid-sensing ion channel-1a in the amygdala, a novel therapeutictarget in depression-related behavior. J Neurosci 29(17), 5381−5388.
Coryell, M. W., Ziemann, A. E., Westmoreland, P. J., Haenfler, J. M., Kurjakovic, Z., Zha,X. M., et al. (2007). Targeting ASIC1a reduces innate fear and alters neuronalactivity in the fear circuit. Biol Psychiatry 62(10), 1140−1148.
de Weille, J., & Bassilana, F. (2001). Dependence of the acid-sensitive ion channel,ASIC1a, on extracellular Ca(2+) ions. Brain Res 900(2), 277−281.
de Weille, J. R., Bassilana, F., Lazdunski, M., & Waldmann, R. (1998). Identification,functional expression and chromosomal localisation of a sustained human proton-gated cation channel. FEBS Lett 433(3), 257−260.
Denac, H., Mevissen, M., & Scholtysik, G. (2000). Structure, function and pharmacology ofvoltage-gated sodiumchannelsNaunyn-Schmiedeberg's.Arch Pharmacol 362, 453−479.
Deval, E., Baron, A., Lingueglia, E., Mazarguil, H., Zajac, J. M., & Lazdunski, M. (2003).Effects of neuropeptide SF and related peptides on acid sensing ion channel 3 andsensory neuron excitability. Neuropharmacology 44(5), 662−671.
Deval, E., Friend, V., Thirant, C., Salinas, M., Jodar, M., Lazdunski, M., et al. (2006).Regulation of sensory neuron-specific acid-sensing ion channel 3 by the adaptorprotein Na+/H+ exchanger regulatory factor-1. J Biol Chem 281(3), 1796−1807.
Deval, E., Noel, J., Lay, N., Alloui, A., Diochot, S., Friend, V., et al. (2008). ASIC3, a sensor ofacidic and primary inflammatory pain. EMBO J 27(22), 3047−3055.
Deval, E., Salinas, M., Baron, A., Lingueglia, E., & Lazdunski, M. (2004). ASIC2b-dependent regulation of ASIC3, an essential acid-sensing ion channel subunit insensory neurons via the partner protein PICK-1. J Biol Chem 279(19),19531−19539.
Diochot, S., Baron, A., Rash, L. D., Deval, E., Escoubas, P., Scarzello, S., et al. (2004). A newsea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel insensory neurons. EMBO J 23(7), 1516−1525.
Diochot, S., Salinas, M., Baron, A., Escoubas, P., & Lazdunski, M. (2007). Peptidesinhibitors of acid-sensing ion channels. Toxicon 49(2), 271−284.
Donier, E., Rugiero, F., Jacob, C., & Wood, J. N. (2008). Regulation of ASIC activity byASIC4–new insights into ASIC channel function revealed by a yeast two-hybridassay. Eur J Neurosci 28(1), 74−86.
Donier, E., Rugiero, F., Okuse, K., & Wood, J. N. (2005). Annexin II light chain p11promotes functional expression of acid-sensing ion channel ASIC1a. J Biol Chem 280(46), 38666−38672.
Dorofeeva, N. A., Barygin, O. I., Staruschenko, A., Bolshakov, K. V., & Magazanik, L. G.(2008). Mechanisms of non-steroid anti-inflammatory drugs action on ASICsexpressed in hippocampal interneurons. J Neurochem 106(1), 429−441.
Drew, L. J., Rohrer, D. K., Price, M. P., Blaver, K. E., Cockayne, D. A., Cesare, P., et al. (2004).Acid-sensing ion channels ASIC2 and ASIC3 do not contribute to mechanicallyactivated currents in mammalian sensory neurones. J Physiol 556(Pt 3), 691−710.
Duan, B., Wu, L. J., Yu, Y. Q., Ding, Y., Jing, L., Xu, L., et al. (2007). Upregulation of acid-sensing ion channel ASIC1a in spinal dorsal horn neurons contributes toinflammatory pain hypersensitivity. J Neurosci 27(41), 11139−11148.
Dube, G. R., Lehto, S. G., Breese, N. M., Baker, S. J., Wang, X., Matulenko, M. A., et al.(2005). Electrophysiological and in vivo characterization of A-317567, a novelblocker of acid sensing ion channels. Pain 117(1–2), 88−96.
Duggan, A., Garcia-Anoveros, J., & Corey, D. P. (2002). The PDZ domain protein PICK1and the sodium channel BNaC1 interact and localize at mechanosensory terminalsof dorsal root ganglion neurons and dendrites of central neurons. J Biol Chem 277(7), 5203−5208.
Escoubas, P., De Weille, J. R., Lecoq, A., Diochot, S., Waldmann, R., Champigny, G., et al.(2000). Isolation of a tarantula toxin specific for a class of proton-gated Na+
channels. J Biol Chem 275(33), 25116−25121.Ettaiche, M., Deval, E., Cougnon, M., Lazdunski, M., & Voilley, N. (2006). Silencing acid-
sensing ion channel 1a alters cone-mediated retinal function. J Neurosci 26(21),5800−5809.
Ettaiche, M., Deval, E., Pagnotta, S., Lazdunski, M., & Lingueglia, E. (2009). Acid-sensingion channel 3 in retinal function and survival. Invest Ophthalmol Vis Sci 50(5),2417−2426.
Ettaiche, M., Guy, N., Hofman, P., Lazdunski, M., & Waldmann, R. (2004). Acid-sensingion channel 2 is important for retinal function and protects against light-inducedretinal degeneration. J Neurosci 24(5), 1005−1012.
Frelin, C., Barbry, P., Vigne, P., Chassande, O., Cragoe, E. J., Jr., & Lazdunski, M. (1988).Amiloride and its analogs as tools to inhibit Na+ transport via the Na+ channel, theNa+/H+ antiport and the Na+/Ca2+ exchanger. Biochimie 70(9), 1285−1290.
Fukuda, T., Ichikawa, H., Terayama, R., Yamaai, T., Kuboki, T., & Sugimoto, T. (2006).ASIC3-immunoreactive neurons in the rat vagal and glossopharyngeal sensoryganglia. Brain Res 1081(1), 150−155.
Gao, J., Duan, B., Wang, D. G., Deng, X. H., Zhang, G. Y., Xu, L., et al. (2005). Couplingbetween NMDA receptor and acid-sensing ion channel contributes to ischemicneuronal death. Neuron 48(4), 635−646.
Garcia-Anoveros, J., Derfler, B., Neville-Golden, J., Hyman, B. T., & Corey, D. P. (1997).BNaC1 and BNaC2 constitute a new family of human neuronal sodium channelsrelated to degenerins and epithelial sodium channels. Proc Natl Acad Sci USA 94(4),1459−1464.
Garcia-Anoveros, J., Samad, T. A., Zuvela-Jelaska, L., Woolf, C. J., & Corey, D. P. (2001).Transport and localization of the DEG/ENaC ion channel BNaC1alpha to peripheralmechanosensory terminals ofdorsal root ganglia neurons. J Neurosci 21(8), 2678−2686.
Gonzales, E. B., Kawate, T., & Gouaux, E. (2009). Pore architecture and ion sites in acid-sensing ion channels and P2X receptors. Nature 460(7255), 599−604.
Groth, M., Helbig, T., Grau, V., Kummer, W., & Haberberger, R. V. (2006). Spinal afferentneurons projecting to the rat lung and pleura express acid sensitive channels. RespirRes 7, 96.
Grunder, S., Geissler, H. S., Bassler, E. L., & Ruppersberg, J. P. (2000). A new member ofacid-sensing ion channels from pituitary gland. NeuroReport 11(8), 1607−1611.
Gu, Q., & Lee, L. Y. (2006). Characterization of acid signaling in rat vagal pulmonarysensory neurons. Am J Physiol Lung Cell Mol Physiol 291(1), L58−L65.
Hattori, T., Chen, J., Harding, A. M., Price, M. P., Lu, Y., Abboud, F. M., et al. (2009). ASIC2aand ASIC3 heteromultimerize to form pH-sensitive channels in mouse cardiacdorsal root ganglia neurons. Circ Res 105(3), 279−286.

557E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
Herrera, Y., Katnik, C., Rodriguez, J. D., Hall, A. A., Willing, A., Pennypacker, K. R., et al.(2008). Sigma-1 receptor modulation of acid-sensing ion channel a (ASIC1a) andASIC1a-induced Ca2+ influx in rat cortical neurons. J Pharmacol Exp Ther 327(2),491−502.
Hildebrand, M. S., de Silva, M. G., Klockars, T., Rose, E., Price, M., Smith, R. J., et al. (2004).Characterisation of DRASIC in the mouse inner ear. Hear Res 190(1–2), 149−160.
Holzer, P. (2010). Acid sensing by visceral afferent neurons. Acta Physiol, doi:10.1111/j.1748-1716.2010.02143.x.
Hruska-Hageman, A. M., Benson, C. J., Leonard, A. S., Price, M. P., & Welsh, M. J. (2004).PSD-95 and Lin-7b interact with acid-sensing ion channel-3 and have oppositeeffects on H+-gated current. J Biol Chem 279(45), 46962−46968.
Hruska-Hageman, A. M., Wemmie, J. A., Price, M. P., &Welsh, M. J. (2002). Interaction ofthe synaptic protein PICK1 (protein interacting with C kinase 1) with the non-voltage gated sodium channels BNC1 (brain Na+ channel 1) and ASIC (acid-sensingion channel). Biochem J 361(Pt 3), 443−450.
Hughes, P. A., Brierley, S. M., Young, R. L., & Blackshaw, L. A. (2007). Localization andcomparative analysis of acid-sensing ion channel (ASIC1, 2, and 3) mRNAexpression in mouse colonic sensory neurons within thoracolumbar dorsal rootganglia. J Comp Neurol 500(5), 863−875.
Huque, T., Cowart, B. J., Dankulich-Nagrudny, L., Pribitkin, E. A., Bayley, D. L., Spielman,A. I., et al. (2009). Sour ageusia in two individuals implicates ion channels of theASIC and PKD families in human sour taste perception at the anterior tongue. PLoSONE 4(10), e7347.
Ikeuchi, M., Kolker, S. J., Burnes, L. A., Walder, R. Y., & Sluka, K. A. (2008). Role of ASIC3 inthe primary and secondary hyperalgesia produced by joint inflammation in mice.Pain 137(3), 662−669.
Ikeuchi, M., Kolker, S. J., & Sluka, K. A. (2009). Acid-sensing ion channel 3 expression inmouse knee joint afferents and effects of carrageenan-induced arthritis. J Pain 10(3), 336−342.
Immke, D. C., &McCleskey, E. W. (2001a). ASIC3: a lactic acid sensor for cardiac pain. SciWorld J 1, 510−512.
Immke, D. C., & McCleskey, E. W. (2001b). Lactate enhances the acid-sensing Na+
channel on ischemia-sensing neurons. Nat Neurosci 4(9), 869−870.Immke, D. C., & McCleskey, E. W. (2003). Protons open acid-sensing ion channels by
catalyzing relief of Ca2+ blockade. Neuron 37(1), 75−84.Ishibashi, K., & Marumo, F. (1998). Molecular cloning of a DEG/ENaC sodium channel
cDNA from human testis. Biochem Biophys Res Commun 245(2), 589−593.Jasti, J., Furukawa, H., Gonzales, E. B., & Gouaux, E. (2007). Structure of acid-sensing ion
channel 1 at 1.9 A resolution and low pH. Nature 449(7160), 316−323.Jones, N. G., Slater, R., Cadiou, H., McNaughton, P., & McMahon, S. B. (2004). Acid-
induced pain and its modulation in humans. J Neurosci 24(48), 10974−10979.Jones, R. C., III, Otsuka, E., Wagstrom, E., Jensen, C. S., Price, M. P., & Gebhart, G. F. (2007).
Short-term sensitization of colon mechanoreceptors is associated with long-termhypersensitivity to colon distention in the mouse. Gastroenterology 133(1),184−194.
Jones, R. C., III, Xu, L., & Gebhart, G. F. (2005). The mechanosensitivity of mouse colonafferent fibers and their sensitization by inflammatory mediators require transientreceptor potential vanilloid 1 and acid-sensing ion channel 3. J Neurosci 25(47),10981−10989.
Kellenberger, S., & Schild, L. (2002). Epithelial sodium channel/degenerin family of ionchannels: a variety of functions for a shared structure. Physiol Rev 82(3), 735−767.
Kleyman, T. R., & Cragoe, E. J., Jr. (1988). Amiloride and its analogs as tools in the studyof ion transport. J Membr Biol 105(1), 1−21.
Kollarik, M., & Undem, B. J. (2002). Mechanisms of acid-induced activation of airwayafferent nerve fibres in guinea-pig. J Physiol 543(Pt 2), 591−600.
Krishtal, O. A., & Pidoplichko, V. I. (1980). A receptor for protons in the nerve cellmembrane. Neuroscience 5(12), 2325−2327.
Krishtal, O. A., & Pidoplichko, V. I. (1981a). Receptor for protons in the membrane ofsensory neurons. Brain Res 214(1), 150−154.
Krishtal, O. A., & Pidoplichko, V. I. (1981b). A “receptor” for protons in small neurons oftrigeminal ganglia: possible role in nociception. Neurosci Lett 24(3), 243−246.
Kuduk, S. D., Chang, R. K., Wai, J. M., Di Marco, C. N., Cofre, V., DiPardo, R. M., et al.(2009). Amidine derived inhibitors of acid-sensing ion channel-3 (ASIC3). BioorgMed Chem Lett 19(15), 4059−4063.
Leffler, A., Monter, B., & Koltzenburg, M. (2006). The role of the capsaicin receptorTRPV1 and acid-sensing ion channels (ASICS) in proton sensitivity of subpopula-tions of primary nociceptive neurons in rats and mice. Neuroscience 139(2),699−709.
Lilley, S., LeTissier, P., & Robbins, J. (2004). The discovery and characterization of a proton-gated sodium current in rat retinal ganglion cells. J Neurosci 24(5), 1013−1022.
Lingueglia, E. (2007). Acid-sensing ion channels in sensory perception. J Biol Chem 282(24), 17325−17329.
Lingueglia, E., de Weille, J. R., Bassilana, F., Heurteaux, C., Sakai, H., Waldmann, R., et al.(1997). A modulatory subunit of acid sensing ion channels in brain and dorsal rootganglion cells. J Biol Chem 272(47), 29778−29783.
Lingueglia, E., Deval, E., & Lazdunski, M. (2006). FMRFamide-gated sodium channel andASIC channels: a new class of ionotropic receptors for FMRFamide and relatedpeptides. Peptides 27(5), 1138−1152.
Lingueglia, E., & Lazdunski, M. (2009). Acid-Sensing Ion Channels. In: J. Kew, & C. Davies(Eds.), Ion Channels, From Structure to Function. Oxford University Press.
Liu, L., & Simon, S. A. (2000). Capsaicin, acid and heat-evoked currents in rat trigeminalganglion neurons: relationship to functional VR1 receptors. Physiol Behav 69(3),363−378.
Lu, Y., Ma, X., Sabharwal, R., Snitsarev, V., Morgan, D., Rahmouni, K., et al. (2009). Theion channel ASIC2 is required for baroreceptor and autonomic control of thecirculation. Neuron 64(6), 885−897.
Mamet, J., Baron, A., Lazdunski, M., & Voilley, N. (2002). Proinflammatory mediators,stimulators of sensory neuron excitability via the expression of acid-sensing ionchannels. J Neurosci 22(24), 10662−10670.
Mamet, J., Lazdunski, M., & Voilley, N. (2003). How nerve growth factor drivesphysiological and inflammatory expressions of acid-sensing ion channel 3 insensory neurons. J Biol Chem 278(49), 48907−48913.
Mantyh, P. W., Clohisy, D. R., Koltzenburg, M., & Hunt, S. P. (2002). Molecularmechanisms of cancer pain. Nat Rev Cancer 2(3), 201−209.
Mazzuca, M., Heurteaux, C., Alloui, A., Diochot, S., Baron, A., Voilley, N., et al. (2007). Atarantula peptide against pain via ASIC1a channels and opioid mechanisms. NatNeurosci 10(8), 943−945.
Miesenbock, G., De Angelis, D. A., & Rothman, J. E. (1998). Visualizing secretion andsynaptic transmission with pH-sensitive green fluorescent proteins. Nature 394(6689), 192−195.
Mogil, J. S. (2009). Animal models of pain: progress and challenges. Nat Rev Neurosci 10(4), 283−294.
Mogil, J. S., Breese, N. M., Witty, M. F., Ritchie, J., Rainville, M. L., Ase, A., et al. (2005).Transgenic expression of a dominant-negative ASIC3 subunit leads to increasedsensitivity to mechanical and inflammatory stimuli. J Neurosci 25(43), 9893−9901.
Molliver, D. C., Immke, D. C., Fierro, L., Pare, M., Rice, F. L., & McCleskey, E. W. (2005).ASIC3, an acid-sensing ion channel, is expressed in metaboreceptive sensoryneurons. Mol Pain 1, 35.
Neelands, T. R., Zhang, X. F., McDonald, H., & Puttfarcken, P. (2010). Differential effectsof temperature on acid-activated currents mediated by TRPV1 and ASIC channels inrat dorsal root ganglion neurons. Brain Res 1329, 55−56.
Page, A. J., Brierley, S. M., Martin, C. M., Martinez-Salgado, C., Wemmie, J. A., Brennan,T. J., et al. (2004). The ion channel ASIC1 contributes to visceral but not cutaneousmechanoreceptor function. Gastroenterology 127(6), 1739−1747.
Page, A. J., Brierley, S. M., Martin, C. M., Price, M. P., Symonds, E., Butler, R., et al. (2005).Different contributions of ASIC channels 1a, 2, and 3 in gastrointestinalmechanosensory function. Gut 54(10), 1408−1415.
Panula, P., Kalso, E., Nieminen, M., Kontinen, V. K., Brandt, A., & Pertovaara, A. (1999).Neuropeptide FF and modulation of pain. Brain Res 848(1–2), 191−196.
Paukert, M., Babini, E., Pusch, M., & Grunder, S. (2004). Identification of the Ca2+
blocking site of acid-sensing ion channel (ASIC) 1: implications for channel gating. JGen Physiol 124(4), 383−394.
Peng, B. G., Ahmad, S., Chen, S., Chen, P., Price, M. P., & Lin, X. (2004). Acid-sensing ionchannel 2 contributes a major component to acid-evoked excitatory responses inspiral ganglion neurons and plays a role in noise susceptibility of mice. J Neurosci 24(45), 10167−10175.
Poirot, O., Berta, T., Decosterd, I., & Kellenberger, S. (2006). Distinct ASIC currents areexpressed in rat putative nociceptors and are modulated by nerve injury. J Physiol576(Pt 1), 215−234.
Price, M. P., Lewin, G. R., McIlwrath, S. L., Cheng, C., Xie, J., Heppenstall, P. A., et al.(2000). The mammalian sodium channel BNC1 is required for normal touchsensation. Nature 407(6807), 1007−1011.
Price, M. P., McIlwrath, S. L., Xie, J., Cheng, C., Qiao, J., Tarr, D. E., et al. (2001). The DRASICcation channel contributes to the detection of cutaneous touch and acid stimuli inmice. Neuron 32(6), 1071−1083.
Price, M. P., Snyder, P. M., & Welsh, M. J. (1996). Cloning and expression of a novelhuman brain Na+ channel. J Biol Chem 271(14), 7879−7882.
Price, M. P., Thompson, R. J., Eshcol, J. O., Wemmie, J. A., & Benson, C. J. (2004). Stomatinmodulates gating of acid-sensing ion channels. J Biol Chem 279(51), 53886−53891.
Qadri, Y. J., Berdiev, B. K., Song, Y., Lippton, H. L., Fuller, C. M., & Benos, D. J. (2009).Psalmotoxin-1 docking to human acid-sensing ion channel-1. J Biol Chem 284(26),17625−17633.
Reeh, P. W., & Steen, K. H. (1996). Tissue acidosis in nociception and pain. Prog Brain Res113, 143−151.
Rocha-Gonzalez, H. I., Herrejon-Abreu, E. B., Lopez-Santillan, F. J., Garcia-Lopez, B. E.,Murbartian, J., & Granados-Soto, V. (2009). Acid increases inflammatory pain inrats: effect of local peripheral ASICs inhibitors. Eur J Pharmacol 603(1–3), 56−61.
Roza, C., Puel, J. L., Kress, M., Baron, A., Diochot, S., Lazdunski, M., et al. (2004). Knockoutof the ASIC2 channel inmice does not impair cutaneousmechanosensation, visceralmechanonociception and hearing. J Physiol 558(Pt 2), 659−669.
Salinas, M., Lazdunski, M., & Lingueglia, E. (2009). Structural elements for thegeneration of sustained currents by the acid pain sensor ASIC3. J Biol Chem 284(46), 31851−31859.
Salinas, M., Rash, L. D., Baron, A., Lambeau, G., Escoubas, P., & Lazdunski, M. (2006). Thereceptor site of the spider toxin PcTx1 on the proton-gated cation channel ASIC1a. JPhysiol 570(Pt 2), 339−354.
Samways, D. S., Harkins, A. B., & Egan, T. M. (2009). Native and recombinant ASIC1areceptors conduct negligible Ca2+ entry. Cell Calcium 45(4), 319−325.
Schnizler, M. K., Schnizler, K., Zha, X. M., Hall, D. D., Wemmie, J. A., Hell, J. W., et al.(2009). The cytoskeletal protein alpha-actinin regulates acid-sensing ion channel1a through a C-terminal interaction. J Biol Chem 284(5), 2697−2705.
Sherwood, T. W., & Askwith, C. C. (2009). Dynorphin opioid peptides enhance acid-sensing ion channel 1a activity and acidosis-induced neuronal death. J Neurosci 29(45), 14371−14380.
Sluka, K. A., Price, M. P., Breese, N. M., Stucky, C. L., Wemmie, J. A., &Welsh, M. J. (2003).Chronic hyperalgesia induced by repeated acid injections in muscle is abolished bythe loss of ASIC3, but not ASIC1. Pain 106(3), 229−239.
Sluka, K. A., Radhakrishnan, R., Benson, C. J., Eshcol, J. O., Price, M. P., Babinski, K., et al.(2007). ASIC3 in muscle mediates mechanical, but not heat, hyperalgesiaassociated with muscle inflammation. Pain 129(1–2), 102−112.
Smith, E. S., Cadiou,H., &McNaughton, P. A. (2007). Arachidonic acid potentiates acid-sensingion channels in rat sensory neurons by a direct action. Neuroscience 145(2), 686−698.

558 E. Deval et al. / Pharmacology & Therapeutics 128 (2010) 549–558
Staruschenko, A., Dorofeeva, N. A., Bolshakov, K. V., & Stockand, J. D. (2007). Subunit-dependent cadmium and nickel inhibition of acid-sensing ion channels. DevNeurobiol 67(1), 97−107.
Steen, K. H., Issberner, U., & Reeh, P. W. (1995a). Pain due to experimental acidosis inhuman skin: evidence for non-adapting nociceptor excitation. Neurosci Lett 199(1),29−32.
Steen, K. H., & Reeh, P. W. (1993). Sustained graded pain and hyperalgesia fromharmless experimental tissue acidosis in human skin. Neurosci Lett 154(1–2),113−116.
Steen, K. H., Reeh, P. W., Anton, F., & Handwerker, H. O. (1992). Protons selectivelyinduce lasting excitation and sensitization tomechanical stimulation of nociceptorsin rat skin, in vitro. J Neurosci 12(1), 86−95.
Steen, K. H., Reeh, P. W., & Kreysel, H. W. (1995b). Topical acetylsalicylic, salicylic acidand indomethacin suppress pain from experimental tissue acidosis in human skin.Pain 62(3), 339−347.
Steen, K. H., Reeh, P. W., & Kreysel, H. W. (1996). Dose-dependent competitive block bytopical acetylsalicylic and salicylic acid of low pH-induced cutaneous pain. Pain 64(1), 71−82.
Sugiura, T., Dang, K., Lamb, K., Bielefeldt, K., & Gebhart, G. F. (2005). Acid-sensingproperties in rat gastric sensory neurons from normal and ulcerated stomach. JNeurosci 25(10), 2617−2627.
Sutherland, S. P., Benson, C. J., Adelman, J. P., & McCleskey, E. W. (2001). Acid-sensingion channel 3 matches the acid-gated current in cardiac ischemia-sensing neurons.Proc Natl Acad Sci USA 98(2), 711−716.
Tan, Z. Y., Lu, Y., Whiteis, C. A., Benson, C. J., Chapleau, M. W., & Abboud, F. M. (2007).Acid-sensing ion channels contribute to transduction of extracellular acidosis in ratcarotid body glomus cells. Circ Res 101(10), 1009−1019.
Tan, Z. Y., Lu, Y., Whiteis, C. A., Simms, A. E., Paton, J. F., Chapleau, M. W., et al. (2010).Chemoreceptor hypersensitivity, sympathetic excitation, and overexpression ofASIC and TASK channels before the onset of hypertension in SHR. Circ Res 106(3),536−545.
Tominaga, M., Caterina, M. J., Malmberg, A. B., Rosen, T. A., Gilbert, H., Skinner, K., et al.(1998). The cloned capsaicin receptor integrates multiple pain-producing stimuli.Neuron 21(3), 531−543.
Ugawa, S., Minami, Y., Guo, W., Saishin, Y., Takatsuji, K., Yamamoto, T., et al. (1998).Receptor that leaves a sour taste in the mouth. Nature 395(6702), 555−556.
Ugawa, S., Ueda, T., Ishida, Y., Nishigaki, M., Shibata, Y., & Shimada, S. (2002). Amiloride-blockable acid-sensing ion channels are leading acid sensors expressed in humannociceptors. J Clin Invest 110(8), 1185−1190.
Ugawa, S., Ueda, T., Yamamura, H., & Shimada, S. (2005). In situ hybridization evidencefor the coexistence of ASIC and TRPV1 within rat single sensory neurons. Brain ResMol Brain Res 136(1–2), 125−133.
Vilim, F. S., Aarnisalo, A. A., Nieminen, M. L., Lintunen, M., Karlstedt, K., Kontinen, V. K.,et al. (1999). Gene for pain modulatory neuropeptide NPFF: induction in spinalcord by noxious stimuli. Mol Pharmacol 55(5), 804−811.
Voilley, N., de Weille, J., Mamet, J., & Lazdunski, M. (2001). Nonsteroid anti-inflammatory drugs inhibit both the activity and the inflammation-inducedexpression of acid-sensing ion channels in nociceptors. J Neurosci 21(20),8026−8033.
Walder, R. Y., Rasmussen, L. A., Rainier, J. D., Light, A. R., Wemmie, J. A., & Sluka, K. A.(2010). ASIC1 and ASIC3 play different roles in the development of hyperalgesiaafter inflammatory muscle injury. J Pain 11(3), 210−218.
Waldmann, R., Bassilana, F., de Weille, J., Champigny, G., Heurteaux, C., & Lazdunski, M.(1997a). Molecular cloning of a non-inactivating proton-gated Na+ channelspecific for sensory neurons. J Biol Chem 272(34), 20975−20978.
Waldmann, R., Champigny, G., Bassilana, F., Heurteaux, C., & Lazdunski, M. (1997b). Aproton-gated cation channel involved in acid-sensing. Nature 386(6621),173−177.
Waldmann, R., Champigny, G., Voilley, N., Lauritzen, I., & Lazdunski, M. (1996). Themammalian degenerin MDEG, an amiloride-sensitive cation channel activated bymutations causing neurodegeneration in Caenorhabditis elegans. J Biol Chem 271(18), 10433−10436.
Wang, W., Duan, B., Xu, H., Xu, L., & Xu, T. L. (2006). Calcium-permeable acid-sensingion channel is a molecular target of the neurotoxic metal ion lead. J Biol Chem 281(5), 2497−2505.
Wang,W., Yu, Y., & Xu, T. L. (2007). Modulation of acid-sensing ion channels by Cu(2+)in cultured hypothalamic neurons of the rat. Neuroscience 145(2), 631−641.
Wemmie, J. A., Askwith, C. C., Lamani, E., Cassell, M. D., Freeman, J. H., Jr., & Welsh, M. J.(2003). Acid-sensing ion channel 1 is localized in brain regions with high synapticdensity and contributes to fear conditioning. J Neurosci 23(13), 5496−5502.
Wemmie, J. A., Chen, J., Askwith, C. C., Hruska-Hageman, A. M., Price, M. P., Nolan, B. C.,et al. (2002). The acid-activated ion channel ASIC contributes to synaptic plasticity,learning, and memory. Neuron 34(3), 463−477.
Wemmie, J. A., Coryell, M. W., Askwith, C. C., Lamani, E., Leonard, A. S., Sigmund, C. D.,et al. (2004). Overexpression of acid-sensing ion channel 1a in transgenic miceincreases acquired fear-related behavior. Proc Natl Acad Sci USA 101(10),3621−3626.
Wetzel, C., Hu, J., Riethmacher, D., Benckendorff, A., Harder, L., Eilers, A., et al. (2007). Astomatin-domain protein essential for touch sensation in the mouse. Nature 445(7124), 206−209.
White, J. P., Cibelli, M., Rei Fidalgo, A., Paule, C. C., Noormohamed, F., Urban, L., et al.(2010). Role of transient receptor potential and acid-sensing ion channels inperipheral inflammatory pain. Anesthesiology 112(3), 729−741.
Wilson, S. G., & Mogil, J. S. (2001). Measuring pain in the (knockout) mouse: bigchallenges in a small mammal. Behav Brain Res 125(1–2), 65−73.
Woo, Y. C., Park, S. S., Subieta, A. R., & Brennan, T. J. (2004). Changes in tissue pH andtemperature after incision indicate acidosis may contribute to postoperative pain.Anesthesiology 101(2), 468−475.
Wu, L. J., Duan, B., Mei, Y. D., Gao, J., Chen, J. G., Zhuo, M., et al. (2004). Characterizationof acid-sensing ion channels in dorsal horn neurons of rat spinal cord. J Biol Chem279(42), 43716−43724.
Wultsch, T., Painsipp, E., Shahbazian, A., Mitrovic, M., Edelsbrunner, M., Lazdunski, M.,et al. (2008). Deletion of the acid-sensing ion channel ASIC3 prevents gastritis-induced acid hyperresponsiveness of the stomach-brainstem axis. Pain 134(3),245−253.
Xie, J., Price, M. P., Berger, A. L., & Welsh, M. J. (2002). DRASIC contributes to pH-gatedcurrents in large dorsal root ganglion sensory neurons by forming heteromulti-meric channels. J Neurophysiol 87(6), 2835−2843.
Xie, J., Price, M. P., Wemmie, J. A., Askwith, C. C., & Welsh, M. J. (2003). ASIC3 and ASIC1mediate FMRFamide-related peptide enhancement of H+-gated currents incultured dorsal root ganglion neurons. J Neurophysiol 89(5), 2459−2465.
Xiong, Z. G., Zhu, X. M., Chu, X. P., Minami, M., Hey, J., Wei, W. L., et al. (2004).Neuroprotection in ischemia: blocking calcium-permeable acid-sensing ionchannels. Cell 118(6), 687−698.
Yagi, J., Wenk, H. N., Naves, L. A., & McCleskey, E. W. (2006). Sustained currents throughASIC3 ion channels at the modest pH changes that occur during myocardialischemia. Circ Res 99(5), 501−509.
Yen, Y. T., Tu, P. H., Chen, C. J., Lin, Y. W., Hsieh, S. T., & Chen, C. C. (2009). Role of acid-sensing ion channel 3 in sub-acute-phase inflammation. Mol Pain 5, 1.
Yermolaieva, O., Leonard, A. S., Schnizler, M. K., Abboud, F. M., & Welsh, M. J. (2004).Extracellular acidosis increases neuronal cell calcium by activating acid-sensing ionchannel 1a. Proc Natl Acad Sci USA 101(17), 6752−6757.
Yiangou, Y., Facer, P., Smith, J. A., Sangameswaran, L., Eglen, R., Birch, R., et al. (2001).Increased acid-sensing ion channel ASIC-3 in inflamed human intestine. Eur JGastroenterol Hepatol 13(8), 891−896.
Zha, X. M., Costa, V., Harding, A. M., Reznikov, L., Benson, C. J., & Welsh, M. J. (2009).ASIC2 subunits target acid-sensing ion channels to the synapse via an associationwith PSD-95. J Neurosci 29(26), 8438−8446.
Zha, X. M., Wemmie, J. A., Green, S. H., & Welsh, M. J. (2006). Acid-sensing ion channel1a is a postsynaptic proton receptor that affects the density of dendritic spines. ProcNatl Acad Sci USA 103(44), 16556−16561.
Ziemann, A. E., Allen, J. E., Dahdaleh, N. S., Drebot, I. I., Coryell, M. W., Wunsch, A. M.,et al. (2009). The amygdala is a chemosensor that detects carbon dioxide andacidosis to elicit fear behavior. Cell 139(5), 1012−1021.
Related Documents











