ORIGINAL ARTICLE Morphological and histological characterization of the osmophores and nectaries of four species of Acianthera (Orchidaceae: Pleurothallidinae) Marcos Cabral de Melo • Eduardo Leite Borba • Elder Anto ˆnio Sousa Paiva Received: 18 February 2009 / Accepted: 13 April 2010 / Published online: 15 May 2010 Ó Springer-Verlag 2010 Abstract Nectar and floral odor are frequently associated with the presence and maintenance of specialized pollina- tion systems in Orchidaceae. We studied flowers of four Acianthera species, a genus of myophilous orchids belonging to the largest fly-pollinated orchid group Pleurothallidinae, in order to characterize the secretory structures related to their pollination mechanism. Flowers at anthesis were sampled to detect volatile compounds and nectar; samples were fixed for light microscopy and scan- ning and transmission electron microscopy. The labellum presents epidermal cells and the first mesophyll layer involved with secretory processes. Cellular characteristics of these regions associated with the occurrence of sugars allowed us to recognize them as nectaries. Some portions of the sepals also shown to be involved with secretory processes and the presence of nitrogenated volatile com- pounds characterize them as osmophores. The production of nectar in these species makes the occurrence of sapr- omyophily questionable, even though these flowers present characteristics of this floral syndrome. The presence of osmophores on the sepals reinforces that this localization is common among the Pleurothallidinae, whilst they occur in a different region (labelum) in the other major fly-polli- nated orchid group (Bulbophyllum). Keywords Acianthera Á Cell ultrastructure Á Myophily Á Nectaries Á Orchidaceae Á Osmophores Introduction The subtribe Pleurothallidinae, with approximately 4,100 species, and the unrelated genus Bulbophyllum Thouars, with approximately 2,100 species are two of the largest fly-pollinated orchid groups. They exhibit some of the most interesting examples of floral convergence due to adapta- tion to the same group of pollinators (Dressler 1993). Although the flowers of these myophilous orchids share a number of morphological characters, floral biology studies have demonstrated that there is still a large diversity of pollination mechanisms associated with fly-pollination, involving both biotic and wind-assisted biotic mechanisms (van der Pijl and Dodson 1966; Borba and Semir 1998). Among them, different adaptations to attract these insects stand out; some of them are associated with the insects’ feeding and/or reproductive instincts (Borba and Semir 1998, 2001; Singer and Cocucci 1999). Evolution of these differences in many myophilous orchids seems to be rela- ted to the great specificity of the plant-pollinator relation- ship to a degree that was not initially expected. Odor and nectar are important elements to maintain the plant-pollinator relationship. Floral odor is responsible for long distance attraction while nectar constitutes a principal floral reward (Proctor et al. 1996). Nectar is the principal reward to pollinators among the Orchidaceae (Dressler 1993) and is commonly present in orchids pollinated by anthophilous species of Diptera. The presence of nectar may result in high pollen transfer, as previously demon- strated in epidendroid orchids (Peter and Johnson 2009). Although nectar is not commonly found among sapromy- ophilic plants, as their pollinators are attracted by ovipo- sition instinct (Faegri and van der Pijl 1979; Proctor et al. 1996), it seems to be an important element in the mechanism of deceit-pollination in some wind-assisted M. C. de Melo Á E. L. Borba Á E. A. S. Paiva (&) Departamento de Bota ˆnica, Instituto de Cie ˆncias Biolo ´gicas, Universidade Federal de Minas Gerais, Av. Anto ˆnio Carlos, 6627, Pampulha, Belo Horizonte, Minas Gerais 31270-901, Brazil e-mail: [email protected] 123 Plant Syst Evol (2010) 286:141–151 DOI 10.1007/s00606-010-0294-1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Morphological and histological characterizationof the osmophores and nectaries of four speciesof Acianthera (Orchidaceae: Pleurothallidinae)
Marcos Cabral de Melo • Eduardo Leite Borba •
Elder Antonio Sousa Paiva
Received: 18 February 2009 / Accepted: 13 April 2010 / Published online: 15 May 2010
� Springer-Verlag 2010
Abstract Nectar and floral odor are frequently associated
with the presence and maintenance of specialized pollina-
tion systems in Orchidaceae. We studied flowers of
four Acianthera species, a genus of myophilous orchids
belonging to the largest fly-pollinated orchid group
Pleurothallidinae, in order to characterize the secretory
structures related to their pollination mechanism. Flowers
at anthesis were sampled to detect volatile compounds and
nectar; samples were fixed for light microscopy and scan-
ning and transmission electron microscopy. The labellum
presents epidermal cells and the first mesophyll layer
involved with secretory processes. Cellular characteristics
of these regions associated with the occurrence of sugars
allowed us to recognize them as nectaries. Some portions
of the sepals also shown to be involved with secretory
processes and the presence of nitrogenated volatile com-
pounds characterize them as osmophores. The production
of nectar in these species makes the occurrence of sapr-
omyophily questionable, even though these flowers present
characteristics of this floral syndrome. The presence of
osmophores on the sepals reinforces that this localization is
common among the Pleurothallidinae, whilst they occur in
a different region (labelum) in the other major fly-polli-
nated orchid group (Bulbophyllum).
Keywords Acianthera � Cell ultrastructure � Myophily �Nectaries � Orchidaceae � Osmophores
Introduction
The subtribe Pleurothallidinae, with approximately 4,100
species, and the unrelated genus Bulbophyllum Thouars,
with approximately 2,100 species are two of the largest
fly-pollinated orchid groups. They exhibit some of the most
interesting examples of floral convergence due to adapta-
tion to the same group of pollinators (Dressler 1993).
Although the flowers of these myophilous orchids share a
number of morphological characters, floral biology studies
have demonstrated that there is still a large diversity of
pollination mechanisms associated with fly-pollination,
involving both biotic and wind-assisted biotic mechanisms
(van der Pijl and Dodson 1966; Borba and Semir 1998).
Among them, different adaptations to attract these insects
stand out; some of them are associated with the insects’
feeding and/or reproductive instincts (Borba and Semir
1998, 2001; Singer and Cocucci 1999). Evolution of these
differences in many myophilous orchids seems to be rela-
ted to the great specificity of the plant-pollinator relation-
ship to a degree that was not initially expected.
Odor and nectar are important elements to maintain the
plant-pollinator relationship. Floral odor is responsible for
long distance attraction while nectar constitutes a principal
floral reward (Proctor et al. 1996). Nectar is the principal
reward to pollinators among the Orchidaceae (Dressler
1993) and is commonly present in orchids pollinated by
anthophilous species of Diptera. The presence of nectar
may result in high pollen transfer, as previously demon-
strated in epidendroid orchids (Peter and Johnson 2009).
Although nectar is not commonly found among sapromy-
ophilic plants, as their pollinators are attracted by ovipo-
sition instinct (Faegri and van der Pijl 1979; Proctor
et al. 1996), it seems to be an important element in the
mechanism of deceit-pollination in some wind-assisted
M. C. de Melo � E. L. Borba � E. A. S. Paiva (&)
Departamento de Botanica, Instituto de Ciencias Biologicas,
Universidade Federal de Minas Gerais,
Av. Antonio Carlos, 6627, Pampulha, Belo Horizonte,
Minas Gerais 31270-901, Brazil
e-mail: [email protected]
123
Plant Syst Evol (2010) 286:141–151
DOI 10.1007/s00606-010-0294-1

fly-pollinated Bulbophyllum species (Borba and Semir
1998) or in partially deceitful species of Acianthera (Borba
and Semir 2001).
The genus Acianthera (Pleurothallidinae) comprises
approximately 200 species distributed throughout tropical
South America, especially southeastern Brazil (Pridgeon
et al. 2005). As with other members of the subtribe,
Acianthera in humid forests are predominantly epiphytes,
or otherwise, lithophytes on rocky soils exposed to direct
sunlight. Their flowers show features typical of myophilous
flowers such as diurnal anthesis and, frequently, unpleasant
odor. Borba and Semir (2001) studied the pollination
biology of a group of Brazilian Acianthera species and
found that species pollinated by Phoridae flies have nectar
on the labellum, while those pollinated by Chloropidae
flies are nectarless and use deceit-pollination. In all these
species scent emission occurs in the sepals (E. L. Borba,
UFMG, Minas Gerais, Brazil and J. R. Trigo, UNICAMP,
Sao Paulo, Brazil, unpublished data).
Acianthera hamosa (Barb Rodr.) Pridgeon & M. W.
Chase, A. limae (Porto & Brade) Pridgeon & M. W. Chase,
A. modestissima (Rchb. f. & Warm.) Pridgeon & M. W.
Chase, and A. prolifera (Herb. ex Lindl.) Pridgeon &
M. W. Chase constitute a group of species that have very
similar overall flower morphology. However, they have
subtle morphological differences that may be related to the
attraction of distinct groups of pollinators (different fami-
lies of Diptera) and, thus, contribute to their reproductive
isolation (Melo 2008). These species have similar flower
morphology to those studied by Borba and Semir (2001).
Some areas of the sepals and labellum are shiny, and the
base of the labellum is frequently moist, which may indi-
cate that these regions can be involved in the release of
secretions, like those from osmophores or nectaries (Vogel
1990; Pacini et al. 2003). This may suggest that the papillose
regions of the labellum of those species pollinated by
Phoridae flies (A. hamosa, A. limae, and A. modestissima)
and Chloropidae flies (A. prolifera) are nectaries, even
though such structures were believed to be absent in the latter
(Melo 2008).
All four species seem to have osmophores located at the
adaxial face of sepals. The micromorphology of osmo-
phores in fly-pollinated orchids has been examined only in
a few species of Bulbophyllum and Pleurothallidinae before
(Pridgeon and Stern 1985; Vogel 1990; Teixeira et al.
2004). These studies have shown that osmophores are
generally found in the labellum, in species of Bulbophyl-
lum, or the sepals, in the Pleurothallidinae (Masdevallia,
Pleurothallis, Restrepia, and Scaphosepalum) (Vogel
1990; Teixeira et al. 2004). If osmophores occur in dif-
ferent parts of the flowers in other species of the Pleuro-
thallidinae, such as in Acianthera, is unknown. But if it
does, this could reinforce the idea of divergence in
anatomical and functional aspects of the flowers of
Bulbophyllum and Pleurothallidinae species, despite the
apparent morphological convergence of their flowers (van
der Pijl and Dodson 1966).
In this study we investigated the micromorphological
features of the labellum and sepals of four species of
Acianthera pollinated by Chloropidae (A. prolifera) and
Phoridae flies (A. hamosa, A. limae, A. modestissima) using
light microscopy and transmission and scanning electron
microscopy. We aim to (1) determine the occurrence of
nectary glands and osmophores in flowers of Acianthera;
(2) correlate these structures with the pollination mecha-
nism observed in the group; and (3) correlate the
morphology and location of these structures with that
observed in the two largest myophilous groups in the
family, Pleurothallidinae and Bulbophyllum.
Materials and methods
Plant material
Flowers were collected from plants cultivated at the
greenhouse of the Universidade Federal de Minas Gerais.
These plants were collected from wild populations previ-
ously included in reproductive biology studies by our group
(Melo 2008). In the wild these plants are usually found
growing on rock outcrops (A. modestissima and A. prolif-
era) or on the forest floor of gallery forests (A. hamosa and
A. limae) in areas of campos rupestres vegetation in Minas
Gerais State, Brazil. Voucher specimens were deposited in
the herbarium BHCB (A. hamosa, M.C. Melo 08; A. limae,
M.C. Melo 07; A. modestissima, M.C. Melo 06; A. prolif-
era, M.C. Melo 04).
Light microscopy
Flowers in full anthesis of all four species were fixed in
Karnovsky solution (Karnovsky 1965) for 48 h, dehydrated
in an ethanol series, and embedded in 2-hydroxyethyl-
methacrylate resin (Leica). Transverse and longitudinal
sections of 5 lm were made and stained with 0.05%
toluidine blue at pH 4.3 (O’Brien et al. 1964). Cross-
sections of fresh sepals and labellum were done by hand
and used in histochemical tests using Lugol’s solution to
detect starch (Johansen 1940) and Sudan Red B for lipids
(Pearse 1980), with their respective controls. These tests
were performed in flowers that have been in anthesis for up
to 48 h. To detect starch dynamics, the Lugol test was
repeated on flowers close to senescence (approximately
10 days in A. hamosa and A. modestissima and 20–25 days
in A. limae and A. prolifera). Glycerin was applied
to paradermal sections to observe stomatal movements
142 M. C. de Melo et al.
123

(Jernstedt and Clark 1979). Descriptions of structural
characteristics were made, and attention was especially
given to cells and tissue in the sepals and labellum likely to
be involved in secretory processes.
Scanning electron microscopy
Tissue samples of the lateral sepals and labellum of
A. limae, A. modestissima, and A. prolifera were prepared
for observation under scanning electron microscopy. Due
to low availability of flowers, A. hamosa was not included
in this analysis. Tissue was taken from the flowers up to
48 h after the beginning of anthesis, fixed in 2.5% glutar-
aldehyde (0.1 M phosphate buffer, pH 7.2), dehydrated in
an ethanol series, dried to critical point, and subsequently
sputter-coated with ca. 10 nm of gold as described by
Robards (1978). The samples were examined in a scanning
electron microscope, model Quanta 200 (Fei Company,
Hillsboro, OR, USA), and all images were processed
digitally.
Transmission electron microscopy
Overall, the floral anatomy of all the Acianthera species
included in this study is very similar. Therefore, only
flowers of A. prolifera were used for the TEM study. This
similarity was confirmed by observations of the sepals and
labellum under light and scanning electron microscopy and
the chemical nature of the substances they secrete. Tissue
samples from the lateral sepals and labellum were obtained
from flowers up to 48 h after beginning of the anthesis,
fixed in Karnovsky solution (Karnovsky 1965) for 24 h,
post-fixed in 1% osmium tetroxide (in 0.1 M phosphate
buffer, pH 7.2), and processed using standard methodolo-
gies (Roland 1978) for observation under transmission
electron microscope. Ultra-thin sections were stained
with uranyl acetate and lead citrate and examined under
a Philips CM 100 transmission microscope at 60 kV.
Descriptions were focused on cells and tissues that appeared
to be involved in secretory processes.
Preliminary analyses of nectar and volatile compounds
The presence of sugars was detected by thin layer chro-
matography (TLC) following Dafni (1992). Residual floral
secretions on the adaxial surface of the labellum and on the
lateral sepals of about ten flowers of each of the four
species were soaked in situ with distilled water (2 ll
droplet) for 10 min. These samples were then removed and
applied to 10 9 10 cm silica gel TLC plates prepared with
0.02% sodium acetate. It is important to emphasize that
samples from the labellum and sepals were analyzed sep-
arately. Standard marker solutions of fructose, glucose, and
sucrose, as well as distilled water (control) were also pre-
pared and applied to the same plates. These plates were run
with a mobile phase of chloroform:methanol (6:4), dried at
room temperature, sprayed with an orcinol-sulfuric acid
solution, and then heated to 120�C for 5 min. Sugars
stained as dark-purple bands against a yellow background
(Stahl 1988).
The Whiff test (Amsel et al. 1983) was used to detect
volatile amines, generally responsible for the unpleasant
scent emitted by flowers. The dorsal sepal, lateral sepals
and petals, and the labellum of ten flowers from each of the
four species were immersed in a solution of 10% KOH, in a
closed vessel for 1 min. As control, we used a flask with
just the KOH solution and another flask with the same
solution but with a piece of leaf from one of the species.
Floral parts that gave off fishy odors were considered as
sites where volatile compounds are probably emitted.
Results
The organization and anatomy of the floral parts are very
similar in all four species. The epidermis of the entire
perianth is uniseriate (Fig. 1). The parenchyma mesophyll
is homogenous and with collateral vascular bundles
(Fig. 1a, c), and many idioblasts containing raphides.
Given the great morphological similarity among the flow-
ers of the Acianthera studied here, we present a description
applicable to all four species, highlighting, where appro-
priate, the peculiarities of certain species.
Labellum structure
The labellum has two calli along its median portion; these
are discrete and consist of epidermal projections that are
structurally similar to those observed on the entire adaxial
face (Fig. 2a, g). The epidermis of the labellum is unise-
riate, with juxtaposed cells, and no stomata are present
(Figs. 1b, 2). Papillae are found on the entire adaxial face
(Fig. 2), from the distal region of the labellum to its base
where it joins the column. These papillae are intercalated
with unspecialized epidermal cells. The papillae have
sculptured surfaces with irregular areas delimited by raised
borders, especially in A. limae (Fig. 2c, f, h). Both the
papillae and the other epidermal cells of the adaxial face
have a dense cytoplasm and conspicuous nuclei. Both cell
types are covered by a continuous cuticle that is firmly
joined to the cell walls. On the abaxial face, on the con-
trary, the epidermal cells are flattened with large vacuoles.
The labellum mesophyll is parenchymatous, and the
superficial layer of the adaxial face is formed by cells with
a denser cytoplasm than the internal cells; starch grains are
also frequently observed here. There is considerably less
Osmophores and nectaries in Acianthera (Orchidaceae) 143
123
starch in the mesophyll cells of flowers during the secretory
phase and practically none in the mesophyll of presenes-
cent flowers. Vascular bundles (one dorsal and two laterals)
are present near the abaxial face, but no ramifications were
observed to the adaxial face (Fig. 1a).
Sepal structure
The epidermis of the sepals is uniseriate with juxtaposed
cells and covered by cuticle. The epidermis on the adaxial
face contains regions that are characterized by the presence
of cells with dense cytoplasm and conspicuous nuclei
(Fig. 1a, c, d). The epidermal cells on both sides of the
sepal are projected slightly upwards, giving the sepals a
subtle papillose appearance (Fig. 1c). The mesophyll, and
especially the area next to the adaxial face, is formed by a
two-to-three-cell layer with large numbers of starch grains
(Fig. 1e).
The apex of the dorsal sepal has a region formed by cells
with dense cytoplasm, while in the lateral sepals this region
extends along both sides of the fusion line from the basal
third of the sepals up to the apex. This area is restricted
only to the adaxial face of the sepals, where the stomata
occur exclusively. The stomata have wide pores and
inflated outwardly projecting guard cells (Fig. 3). The
cuticle is smooth, not ornamented, and without pores or
signs of ruptures of any kind (Fig. 3c, f). In some stomata,
especially in A. modestissima and A. limae, the cuticle
covers the stomatic pore, obliterating it (Fig. 3a). The
stomata were permanently open and no movements were
observed.
The underlying parenchymatous mesophyll on the
adaxial face of the sepals, towards the epidermal cells
described above, includes cells with dense cytoplasm,
conspicuous nuclei, and numerous plastids with starch
grains (Fig. 1c–e). The quantity of starch grains decreases
with the age of the flowers. Although starch grains can
be observed throughout the mesophyll and epidermis, they
are mostly concentrated in the cells containing dense
cytoplasm.
Labellum ultrastructure
The cells of the epidermis and the first layer of the meso-
phyll, on the adaxial face of the labellum, have morpho-
logical features that suggest their involvement in secretory
processes, while in other regions, the cells are vacuolated
suggesting low levels of metabolic activity. The epidermal
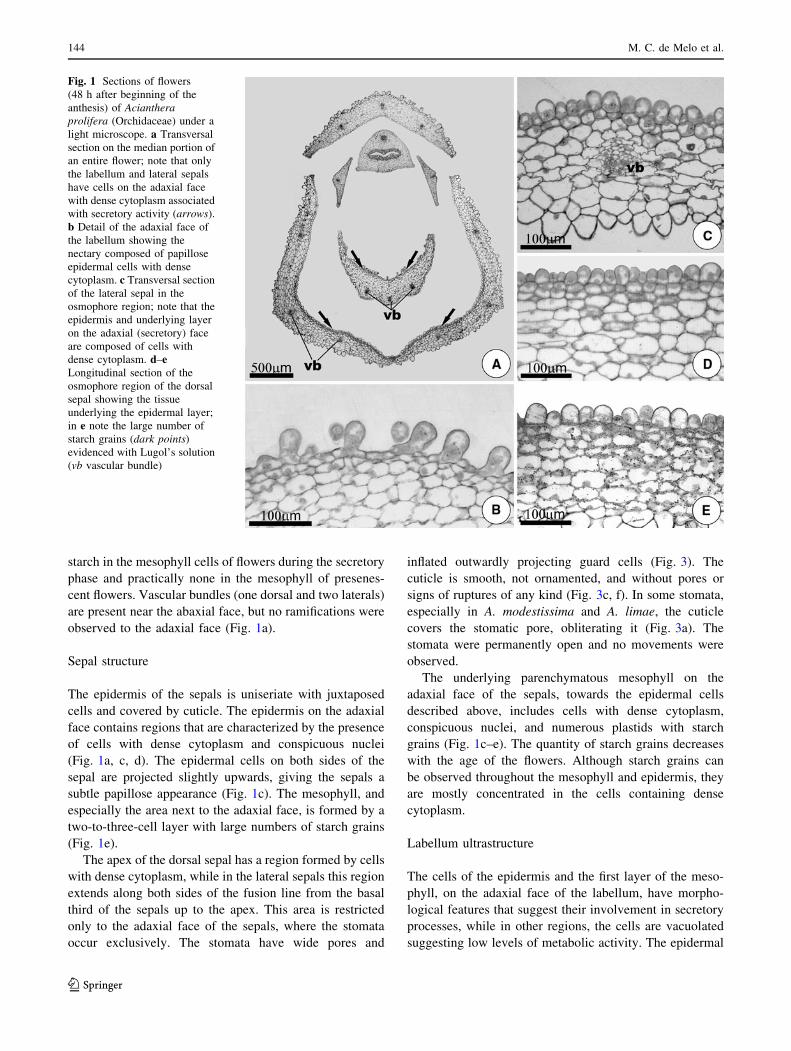
Fig. 1 Sections of flowers
(48 h after beginning of the
anthesis) of Aciantheraprolifera (Orchidaceae) under a
light microscope. a Transversal
section on the median portion of
an entire flower; note that only
the labellum and lateral sepals
have cells on the adaxial face
with dense cytoplasm associated
with secretory activity (arrows).
b Detail of the adaxial face of
the labellum showing the
nectary composed of papillose
epidermal cells with dense
cytoplasm. c Transversal section
of the lateral sepal in the
osmophore region; note that the
epidermis and underlying layer
on the adaxial (secretory) face
are composed of cells with
dense cytoplasm. d–eLongitudinal section of the
osmophore region of the dorsal
sepal showing the tissue
underlying the epidermal layer;
in e note the large number of
starch grains (dark points)
evidenced with Lugol’s solution
(vb vascular bundle)
144 M. C. de Melo et al.
123
cells of the adaxial face have thin walls and a dense and
organelle-rich cytoplasm (Fig. 4). Ordinary epidermal cells
and papillae are similar in their cytoplasmic composition.
The nuclei of these cells are large, slightly lobed, and
nucleoli are evident (Fig. 4c). Vacuoles are numerous, with
one or more central vacuoles and numerous small ones in
the periphery (Fig. 4a, b). Also the fusion of vacuoles and
the presence of intravacuolar membranes are observed
here.
Numerous mitochondria, ranging from globose to elon-
gated and with well-developed cristae, were observed in the
epidermal cells on the adaxial face of the labellum (Fig. 4b).
Rough endoplasmic reticulum was associated with the
smaller vacuoles, often surrounding them (Fig. 4b). The
plastids were globe-shaped, had poorly developed mem-
brane systems and dense stroma, and contained starch grains
(Fig. 4c). Free ribosomes and dictyosomes were also
observed here, although they were not frequent (Fig. 4b).
The epidermal cells have ample periplasmatic spaces and
present vesicles and flocculated material (Fig. 4b), which
suggests secretory activity taking place here.
The parenchymatous cells of the mesophyll have a
single large vacuole (Fig. 4d). Mitochondria and plastids
are also seen in these cells, the latter containing starch
grains, especially in young flowers.
Sepal ultrastructure
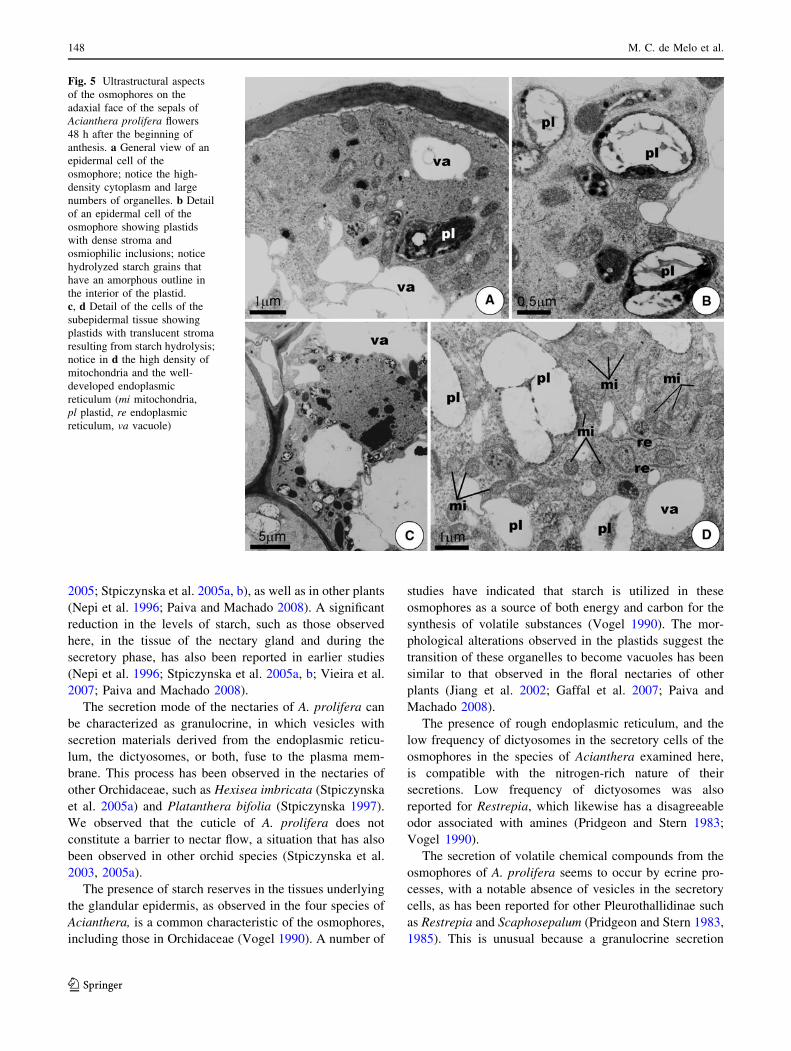
The cytoplasm of the epidermal cells, on the adaxial face of
the sepals, is organelle-rich (Fig. 5) and has lobed nuclei
and evident nucleoli. The cell membrane is sinuous,
forming a conspicuous periplasmatic space, with no signs
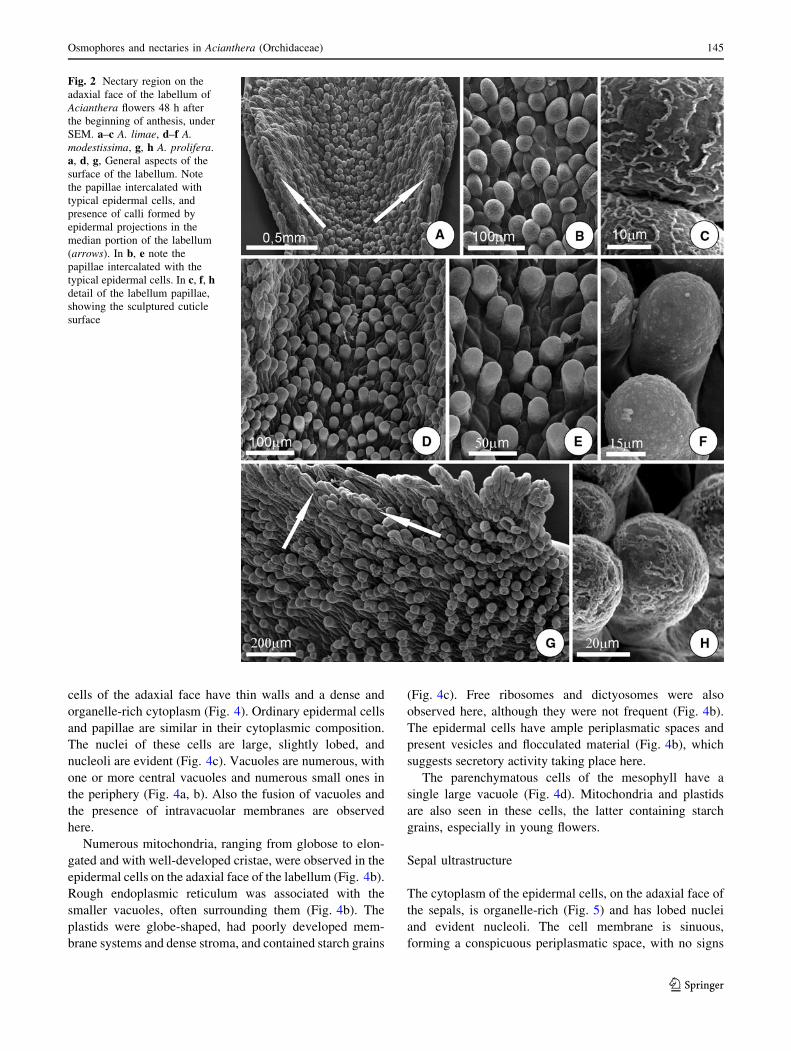
Fig. 2 Nectary region on the
adaxial face of the labellum of
Acianthera flowers 48 h after
the beginning of anthesis, under
SEM. a–c A. limae, d–f A.modestissima, g, h A. prolifera.
a, d, g, General aspects of the
surface of the labellum. Note
the papillae intercalated with
typical epidermal cells, and
presence of calli formed by
epidermal projections in the
median portion of the labellum
(arrows). In b, e note the
papillae intercalated with the
typical epidermal cells. In c, f, hdetail of the labellum papillae,
showing the sculptured cuticle
surface
Osmophores and nectaries in Acianthera (Orchidaceae) 145
123
of secretions accumulating here (Fig. 5a). The nucleus is
usually found in the lower third of the cells, while the
central portion of the cell contains numerous vacuoles.
Among the organelles in these epidermal cells, plastids and
mitochondria with well-developed cristae are the most
abundant (Fig. 5a, b). Many of the plastids are polymor-
phic, although globose forms predominate; the stroma is
dense and there are many osmiophilic inclusions (Fig. 5a,
b). A large part of the plastid stroma is occupied by starch
that appears to be partially hydrolyzed; it has an amorphous
aspect (Fig. 5b). Free ribosomes and rough endoplasmic
reticulum can be seen in the cytoplasm of the epidermal
cells (Fig. 5d). Dictyosomes are rare in both the epidermal
and the secretory cells of the mesophyll.
Mesophyll cells of the adaxial faces of the sepals show
ultrastructural characteristics similar to those seen in the
epidermal layer (Fig. 5c, d). These cells are connected to
each other and with the epidermis by plasmodesmata.
Some plastids contained no starch reserves at this stage,
and this was more evident in the cells of the internal layers
of the mesophyll. Frequent fusion of plastids or fusion of
plastids and vacuoles was also observed at this phase.
Analyses of nectar and volatile compounds
The TLC analyses indicated the presence of sugars on the
surface of the labellum of the four species examined but
were negative for the sepals. Samples of the labellum
yielded bands with Rfs corresponding to sucrose (0.46) and
to glucose and/or fructose (0.55). Two bands were observed
for A. hamosa and A. modestissima, one intense band for
sucrose and one weak band indicating a monosaccharide.
Analysis of the samples of A. limae and A. prolifera, on the
other hand, showed two bands with similar intensity.
The Whiff test was positive for all four species and
strong fishy odor was perceived from the dorsal and lateral
sepals. However, odor intensity was stronger in the lateral
sepals. No odor was perceived from the labellum or any
vegetative organ soaked in KOH for any of the species.
Discussion
The structural and chemical analyses of the four species
provide strong evidence to conclude that some regions of
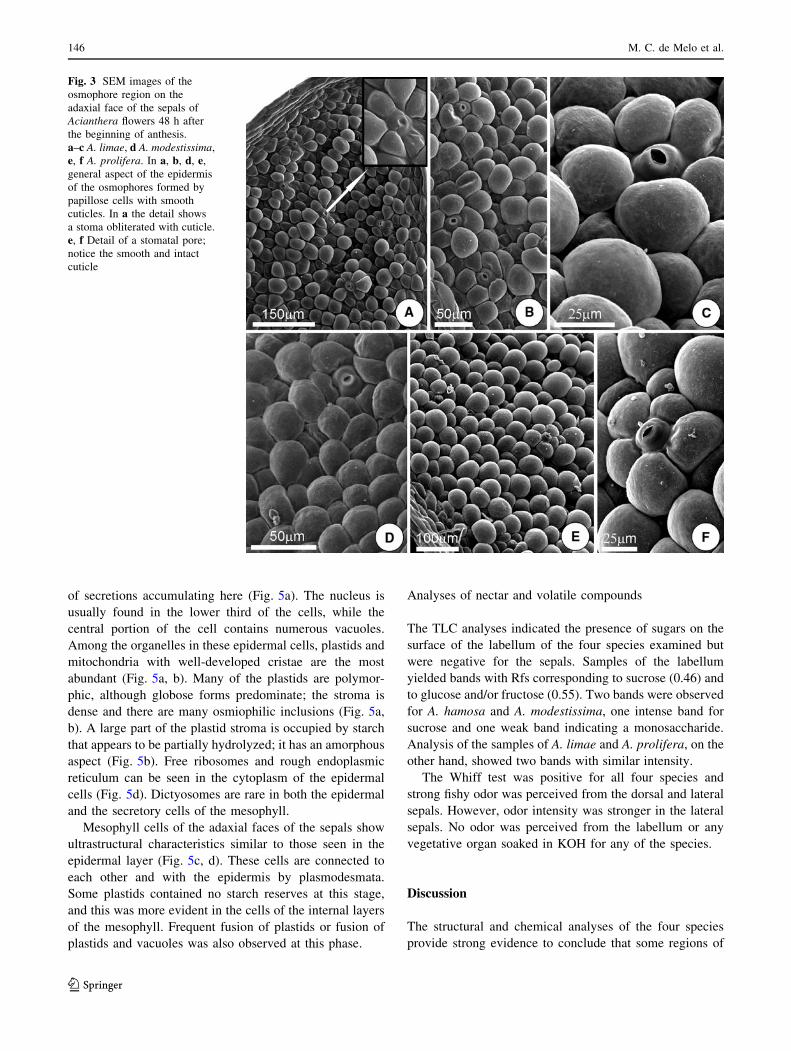
Fig. 3 SEM images of the
osmophore region on the
adaxial face of the sepals of
Acianthera flowers 48 h after
the beginning of anthesis.
a–c A. limae, d A. modestissima,
e, f A. prolifera. In a, b, d, e,
general aspect of the epidermis
of the osmophores formed by
papillose cells with smooth
cuticles. In a the detail shows
a stoma obliterated with cuticle.
e, f Detail of a stomatal pore;
notice the smooth and intact
cuticle
146 M. C. de Melo et al.
123
the labellum and the calyx are secretory. These are easily
identified by their high cytoplasmic density and starch
presence. Also, the ultrastructural analysis of A. prolifera
showed an abundance of mitochondria, which are usually
found in areas of elevated energetic demands due to
secretory processes. The abundance of mitochondria in the
tissue-forming part of osmophores and nectary glands in
the Orchidaceae has been previously reported (Pridgeon
and Stern 1983; Stpiczynska et al. 2005b).
The presence of sugars on the outer surface of the
labellum of the four species of Acianthera allowed us to
delimit the nectary gland. This was corroborated by the
ultrastructural characteristics of the epidermal cells of the
labellum in A. prolifera studied under TEM. Meanwhile,
the presence of volatile nitrogenized compounds, high
cytoplasmic density of the epidermal cells, and the pres-
ence of starch in the subepidermal layers allowed us to
identify the location of osmophores in the papillose
regions of the adaxial face of the sepals, which are larger
in the lateral sepals than in the dorsal sepal. These his-
tological and chemical characteristics have been observed
before in other studies and seem to be typical of the
osmophores in the Orchidaceae and other plant families
(Pridgeon and Stern 1983, 1985; Vogel 1990; Garcia et al.
2007).
The overall morphology of the nectary glands examined
here is similar to that previously described in the Orchid-
aceae, and it is formed by a secretory epidermis associated
with layers of parenchymatic tissue (Stpiczynska and
Matusiewicz 2001; Teixeira et al. 2004; Stpiczynska et al.
2005b; Vieira et al. 2007). The presence of papillae, like
the ones observed in this study, or unicellular trichomes
that increase the secretion surface has also been described in
the nectary glands of other Orchidaceae species (Stpiczynska
1997; Stpiczynska and Matusiewicz 2001; Teixeira et al.
2004; Stpiczynska et al. 2005b).
Idioblasts containing raphides of calcium oxalate have
been observed in the sepals and petals of other species of
orchids (Stpiczynska et al. 2003, 2005a), and their presence
near plant secretory structures has often been noted in
diverse taxonomic groups (Schnell et al. 1963). According
to Paiva and Machado (2005), the presence of these crys-
tals may be related to the elimination of excess calcium
from the cytosol.
The nectaries of the four species of Acianthera exam-
ined here are not vascularized, but they are associated with
large starch reserves that can be used as energy sources for
secretory cells. Starch accumulation in the presecretory
phase is commonly seen in the nectaries of orchid species
(Figueiredo and Pais 1992; Stpiczynska 1997; Davies et al.
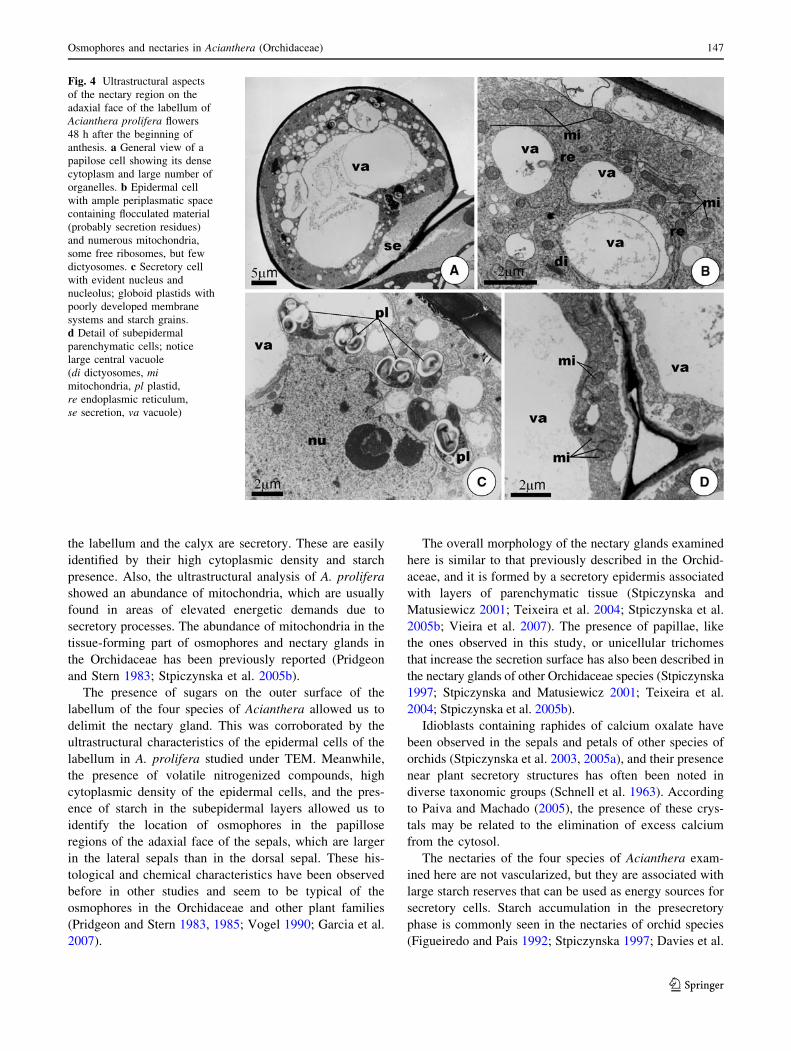
Fig. 4 Ultrastructural aspects
of the nectary region on the
adaxial face of the labellum of
Acianthera prolifera flowers
48 h after the beginning of
anthesis. a General view of a
papilose cell showing its dense
cytoplasm and large number of
organelles. b Epidermal cell
with ample periplasmatic space
containing flocculated material
(probably secretion residues)
and numerous mitochondria,
some free ribosomes, but few
dictyosomes. c Secretory cell
with evident nucleus and
nucleolus; globoid plastids with
poorly developed membrane
systems and starch grains.
d Detail of subepidermal
parenchymatic cells; notice
large central vacuole
(di dictyosomes, mimitochondria, pl plastid,
re endoplasmic reticulum,
se secretion, va vacuole)
Osmophores and nectaries in Acianthera (Orchidaceae) 147
123
2005; Stpiczynska et al. 2005a, b), as well as in other plants
(Nepi et al. 1996; Paiva and Machado 2008). A significant
reduction in the levels of starch, such as those observed
here, in the tissue of the nectary gland and during the
secretory phase, has also been reported in earlier studies
(Nepi et al. 1996; Stpiczynska et al. 2005a, b; Vieira et al.
2007; Paiva and Machado 2008).
The secretion mode of the nectaries of A. prolifera can
be characterized as granulocrine, in which vesicles with
secretion materials derived from the endoplasmic reticu-
lum, the dictyosomes, or both, fuse to the plasma mem-
brane. This process has been observed in the nectaries of
other Orchidaceae, such as Hexisea imbricata (Stpiczynska
et al. 2005a) and Platanthera bifolia (Stpiczynska 1997).
We observed that the cuticle of A. prolifera does not
constitute a barrier to nectar flow, a situation that has also
been observed in other orchid species (Stpiczynska et al.
2003, 2005a).
The presence of starch reserves in the tissues underlying
the glandular epidermis, as observed in the four species of
Acianthera, is a common characteristic of the osmophores,
including those in Orchidaceae (Vogel 1990). A number of
studies have indicated that starch is utilized in these
osmophores as a source of both energy and carbon for the
synthesis of volatile substances (Vogel 1990). The mor-
phological alterations observed in the plastids suggest the
transition of these organelles to become vacuoles has been
similar to that observed in the floral nectaries of other
plants (Jiang et al. 2002; Gaffal et al. 2007; Paiva and
Machado 2008).
The presence of rough endoplasmic reticulum, and the
low frequency of dictyosomes in the secretory cells of the
osmophores in the species of Acianthera examined here,
is compatible with the nitrogen-rich nature of their
secretions. Low frequency of dictyosomes was also
reported for Restrepia, which likewise has a disagreeable
odor associated with amines (Pridgeon and Stern 1983;
Vogel 1990).
The secretion of volatile chemical compounds from the
osmophores of A. prolifera seems to occur by ecrine pro-
cesses, with a notable absence of vesicles in the secretory
cells, as has been reported for other Pleurothallidinae such
as Restrepia and Scaphosepalum (Pridgeon and Stern 1983,
1985). This is unusual because a granulocrine secretion
Fig. 5 Ultrastructural aspects
of the osmophores on the
adaxial face of the sepals of
Acianthera prolifera flowers
48 h after the beginning of
anthesis. a General view of an
epidermal cell of the
osmophore; notice the high-
density cytoplasm and large
numbers of organelles. b Detail
of an epidermal cell of the
osmophore showing plastids
with dense stroma and
osmiophilic inclusions; notice
hydrolyzed starch grains that
have an amorphous outline in
the interior of the plastid.
c, d Detail of the cells of the
subepidermal tissue showing
plastids with translucent stroma
resulting from starch hydrolysis;
notice in d the high density of
mitochondria and the well-
developed endoplasmic
reticulum (mi mitochondria,
pl plastid, re endoplasmic
reticulum, va vacuole)
148 M. C. de Melo et al.
123

mode seems to be common in other groups of orchids
(Vogel 1990; Stpiczynska 1993, 2001).
Emission of volatile compounds by osmophores by
cuticular diffusion processes has been observed in
Orchidaceae before, such as in species of Scaphosepalum
(Pridgeon and Stern 1985) and Stanhopea (Stern et al.
1987); or by cuticular pores in species of Restrepia and
Restrepiella (Pridgeon and Stern 1983) and Gymnadenia
conopsea (Stpiczynska 2001). The emission of volatile
compounds in these species of Acianthera, however, seems
to be associated with the presence of stomata. Stomatal
pores were frequently observed on the surface of the nec-
taries that are involved in exogenous secretion, and Vogel
(1990) suggested they could work as possible routes for
volatile secretions. We found evidence here that the
secretion products of species of Acianthera are liberated by
the cells of the osmophores and accumulated in the peri-
plasmatic and intercellular spaces. These compounds are
probably volatilized by daylight temperatures and finally
reach the outside environment through the stomatal pores.
This hypothesis can be further supported by the liberation
of these odors only during the hottest hours of the day. This
has also been reported for other species of Acianthera
(Borba and Semir 2001) and Bulbophyllum (Borba and
Semir 1998).
The diversity of nectaries in Orchidaceae is intimately
related to their adaptations to diverse groups of pollinators
(Baker and Baker 1983; Pacini et al. 2003). Nectaries in
most orchids are formed by protuberances (calli) or
papillose regions located on the column, labellum, or
between the two (Dressler 1993). The presence of exposed
nectaries on the labellum, as in the species of Acianthera
studied here, seems to be common in myophilous orchids,
and it has been found in several species of Acianthera,
Anathallis, Bulbophyllum, Octomeria, and Stelis (van der
Pijl and Dodson 1966; Dressler 1993; Borba and Semir
1998, 2001; Singer and Cocucci 1999).
Nectar is usually offered as a reward to anthophilous
species of Diptera (Singer and Cocucci 1999; Albores-
Ortiz and Sosa 2006) or in mechanisms of partial deceit-
pollination by saprophytic flies (Borba and Semir 1998,
2001). The species in the present study have purple flowers
with disagreeable odors that are generally associated with
sapromyophily (Proctor et al. 1996), but the presence of
nectar makes the occurrence of strict deceit-pollination
doubtful (van der Pijl and Dodson 1966; Faegri and
van der Pijl 1979). Additionally, no oviposition events
were observed in the species pollinated by Chloropidae
(A. prolifera) or by Phoridae (A. hamosa, A. limae, and
A. modestissima) in this group (Melo 2008). Some authors,
however, argue that the absence of nectar is a feature of
sapromyophily s.s., since the insects would be attracted by
oviposition instinct rather than food instinct (van der Pijl
and Dodson 1966; Faegri and van der Pijl 1979). At least in
Orchidaceae, absence of nectar in sapromyophilous sys-
tems can be effective especially in trap-like mechanisms,
as observed in some Bulbophyllum species (Teixeira et al.
2004). However, in systems where it is necessary to keep
the insect in the flower for a longer period of time, the
presence of nectar may be crucial for pollination to occur
successfully, even if the animal has been attracted by
oviposition instinct (Borba and Semir 1998). Thus my-
ophily s.s. and sapromyophily s.s. may represent only the
extremes of a continuum in which the pollination mecha-
nisms of these species cannot be precisely placed.
The four orchids studied here show pollination mecha-
nisms similar to those reported by Borba and Semir (2001)
in another group of Acianthera. In the study by Borba and
Semir (2001), the two Phoridae-pollinated species offered
nectar while the three species pollinated by female flies of
Chloropidae were deceitful. Female flies were only
observed laying their eggs in the flowers of two of them.
Micromorphological analyses of the flowers of these latter
species, similar to the study we have reported here, could
help to clarify how these two groups of species, with
apparently considerable macromorphological convergence,
exploit a similar pollinator group but possess different
pollination mechanisms.
Osmophores may be located in different parts of the
perianth in orchids (Pridgeon and Stern 1983, 1985; Stern
et al. 1987). In the Pleurothallidinae, including the species
studied here, the osmophores seem to be only in the sepals
or on appendages that originate from them, while in
Bulbophyllum osmophores are commonly found on the
labellum. These two groups have flowers with very similar
external morphologies and share many adaptations to my-
ophily (van der Pijl and Dodson 1966; Dressler 1993), but
there are notable differences in floral micromorphology
associated with each group, as mentioned above. At least
initially, these differences may not play an important role
in attracting specific groups of pollinators or determining
different pollination mechanisms, as Bulbophyllum and
Pleurothallidinae present a wide variation in both param-
eters (Borba and Semir 1998; Singer and Cocucci 1999;
Borba and Semir 2001; Blanco and Barboza 2005; Albores-
Ortiz and Sosa 2006). Rather, they may represent only
phylogenetic constraints in both groups. However both
studies on floral biology and morphology are still very
limited for this to be determined in such large groups.
Flowers pollinated by saprophilous flies generally emit
odors similar to proteinaceous material in decomposition,
mainly nitrogen-containing compounds such as amines,
ammonia, and indols (Proctor et al. 1996). These sub-
stances are associated with the odors of various typically
sapromyophilous species belonging to Pleurothallidinae
and Bulbophyllum (Vogel 1990). However, other studies in
Osmophores and nectaries in Acianthera (Orchidaceae) 149
123

species of these groups have not always confirmed their
presence (Kaiser 1993). The latter could be due to the
sampling method, which used polymers associated with the
technique of head-space and used solvents for the polymer
elution (Kaiser 1993). This technique may be inappropriate
when dealing with nitrogenous substances of low molec-
ular weight, such as ethylamine and dimethylamine, which
are apparently common in Acianthera and other fly-polli-
nated species (E. L. Borba, UFMG, Minas Gerais, Brazil,
and J. R. Trigo, UNICAMP, Sao Paulo, Brazil, unpublished
data), where the technique of solid injection seems more
appropriate (Silva et al. 1999). Also, due to the use of very
small amounts of plant tissue in the solid injection tech-
nique, identifying the precise location of the osmophores in
the flower is necessary. Thus, the Whiff test may be an
important indicator as to which technique is best suited to
be employed in these myophilous groups, with the solid
injection being indicated when nitrogenous substances are
present, such as the Neotropical Pleurothallidinae, in con-
trast to the Neotropical Bulbophyllum species, where these
substances are apparently rarer.
Acknowledgments We thank the technical staff of the Centro de
Microscopia Eletronica, Instituto de Biociencias, UNESP Botucatu,
for their help in preparing the samples, and an anonymous reviewer
for improvements to the manuscript. This work was supported by
grants from Conselho Nacional de Desenvolvimento Cientıfico e
Tecnologico (CNPq) and from Pro-Reitoria de Pesquisa/UFMG to E.
Borba. M. Melo received a fellowship from Fundacao de Amparo
a Pesquisa do Estado de Minas Gerais (FAPEMIG). E. Borba and
E. Paiva are supported by a productivity grant (PQ2) from CNPq.
References
Albores-Ortiz O, Sosa V (2006) Polinizacion de dos especies
simpatricas de Stelis (Pleurothallidinae, Orchidaceae). Acta
Bot Mex 74:155–168
Amsel R, Totten PA, Spiegel CA, Chen KCS, Eschenbach DA (1983)
Nonspecific vaginitis: diagnostic criteria and microbial and
epidemiologic associations. Am J Med 74:14–22
Baker HG, Baker I (1983) Floral nectar constituents in relation to
pollinator type. In: Jones CE, Little RJ (eds) Handbook of
experimental pollination biology. Scientific and Academic
Editions, New York, pp 117–141
Blanco MA, Barboza G (2005) Pseudocopulatory pollination in
Lepanthes (Orchidaceae: Pleurothallidinae) by fungus gnats.
Ann Bot 95:763–772
Borba EL, Semir J (1998) Wind-assisted fly pollination in three
Bulbophyllum (Orchidaceae) species occurring in the Brazilian
‘‘campos rupestres’’. Lindleyana 13:203–218
Borba EL, Semir J (2001) Pollinator specificity and convergence in
fly-pollinated Pleurothallis (Orchidaceae) species: a multiple
population approach. Ann Bot 88:75–88
Dafni A (1992) Pollination ecology—a practical approach. Oxford
University Press, New York
Davies KL, Stpiczynska M, Gregg A (2005) Nectar-secreting floral
stomata in Maxillaria anceps Ames & C. Schweinf. (Orchida-
ceae). Ann Bot 96:217–227
Dressler RL (1993) Phylogeny and classification of the orchid family.
Dioscorides, Portland
Faegri K, van der Pijl L (1979) The principles of the pollination
ecology, 3th edn. Pergamon, Oxford
Figueiredo ACS, Pais MS (1992) Ultrastructural aspects of the
nectary spur of Limodorum abortivum (L.) Sw. (Orchidaceae).
Ann Bot 70:325–331
Gaffal KP, Friedrichs GJ, El-Gammal S (2007) Ultrastructural
evidence for a dual function of the phloem and programmed
cell death in the floral nectary of Digitalis purpurea. Ann Bot
99:593–607
Garcia MTA, Galati BG, Hoc PS (2007) Ultrastructure of the corona
of scented and scentless flowers of Passiflora spp. (Passiflora-
ceae). Flora 202:302–315
Jernstedt JA, Clark C (1979) Stomata on the fruits and seeds of
Eschscholzia (Papaveraceae). Am J Bot 66:586–590
Jiang L, Erickson AH, Rogers JC (2002) Multivesicular bodies: a
mechanism to package lytic and storage functions in one
organelle? Trends Cell Biol 12:362–367
Johansen DA (1940) Plant microtechnique. McGraw, New York
Kaiser R (1993) The scents of orchids: olfactory and chemical
investigations. Elsevier, Amsterdam
Karnovsky MJ (1965) A formaldehyde-glutaraldehyde fixative of
light osmolality for use in electron microscopy. J Cell Biol
27:137–138
Melo MC (2008) Biologia reprodutiva, variabilidade morfologica e
micromorfologia floral de quatro especies de Acianthera(Orchidaceae) ocorrentes em campos rupestres. MSc Thesis,
Universidade Federal de Minas Gerais, Brazil
Nepi M, Ciampolini F, Pacini E (1996) Development and ultrastruc-
ture of Cucurbita pepo nectaries of male flowers. Ann Bot
78:95–104
O’Brien TP, Feder N, McCully ME (1964) Polychromatic staining of
plant cell walls by toluidine blue. Protoplasma 59:368–373
Pacini E, Nepi M, Vesprini J (2003) Nectar biodiversity: a short
review. Plant Syst Evol 238:7–21
Paiva EAS, Machado SR (2005) Role of intermediary cells in
Peltodon radicans (Lamiaceae) in the transfer of calcium and
formation of calcium oxalate crystals. Braz Arch Biol Tech
48:147–153
Paiva EAS, Machado SR (2008) The floral nectary of Hymenaeastigonocarpa (Fabaceae, Caesalpinioideae): structural aspects
during floral development. Ann Bot 101:125–133
Pearse AGE (1980) Histochemistry theoretical and applied, vol 2, 4th
edn. Longman, London
Peter CI, Johnson SD (2009) Reproductive biology of Acrolophiacochlearis (Orchidaceae): estimating rates of cross-pollination in
epidendroid orchids. Ann Bot 104:573–581. doi:10.1093/aob/
mcn218
Pridgeon AM, Stern WL (1983) Ultrastructure of osmophores in
Restrepia (Orchidaceae). Am J Bot 70:1233–1243
Pridgeon AM, Stern WL (1985) Osmophores of Scaphosepalum(Orchidaceae). Bot Gaz 146:115–123
Pridgeon AM, Cribb PJ, Chase MW, Rasmussen FN (2005) Genera
Orchidacearum. Epidendroideae vol 4, part 1. Oxford University
Press, Oxford
Proctor M, Yeo P, Lack A (1996) The natural history of pollination.
Harper Collins, London
Robards AW (1978) An introduction to techniques for scanning
electron microscopy of plant cells. In: Hall JL (ed) Electron
microscopy and cytochemistry of plant cells. Elsevier, New
York, pp 343–403
Roland AM (1978) General preparations and staining of thin sections
In: Hall JL (ed) Electron microscopy and cytochemistry of plant
cells. Elsevier, New York, pp 1–62
150 M. C. de Melo et al.
123

Schnell R, Cusset G, Quenum M (1963) Contribution a l’etude des
glandes extra-florales chez quelques groupes de plantes tropi-
cales. Rev Gen Bot 70:269–341
Silva UF, Borba EL, Semir J, Marsaioli AJ (1999) A simple solid
injection device for the analyses of Bulbophyllum (Orchidaceae)
volatiles. Phytochemistry 50:31–34
Singer RB, Cocucci AA (1999) Pollination mechanism in four
sympatric southern Brazilian Epidendroideae orchids. Lindleya-
na 14:47–56
Stahl E (1988) Thin-layer chromatography: a laboratory handbook,
2nd edn. Springer, Berlin
Stern WL, Curry KJ, Pridgeon AM (1987) Osmophores of Stanhopea(Orchidaceae). Am J Bot 74:1323–1331
Stpiczynska M (1993) Anatomy and ultrastructure of osmophores of
Cymbidium tracyanum Rolfe (Orchidaceae). Acta Soc Bot Pol
62:5–9
Stpiczynska M (1997) The structure of the nectary of Platantherabifolia L. (Orchidaceae). Acta Soc Bot Pol 66:5–11
Stpiczynska M (2001) Osmophores of the fragrant orchid Gymnade-nia conopsea L. (Orchidaceae). Acta Soc Bot Pol 70:91–96
Stpiczynska M, Matusiewicz J (2001) Anatomy and ultrastructure of
the spur nectary of Gymnadenia conopsea L. (Orchidaceae).
Acta Soc Bot Pol 70:267–272
Stpiczynska M, Davies KL, Gregg A (2003) Nectary structure and
nectar secretion in Maxillaria coccinea (Jacq.) L.O. Williams ex
Hodge (Orchidaceae). Ann Bot 93:87–95
Stpiczynska M, Davies KL, Gregg A (2005a) Comparative account of
nectary structure in Hexisea imbricata (Lindl.) Rchb.f. (Orchid-
aceae). Ann Bot 95:749–756
Stpiczynska M, Milanesi C, Faleri C, Cresti M (2005b) Ultrastructure
of the nectary spur of Platanthera chlorantha (Custer) Rchb.
(Orchidaceae) during successive stages of nectar secretion. Acta
Biol Crac 47:111–119
Teixeira SP, Borba EL, Semir J (2004) Lip anatomy and its
implications for the pollination mechanism of Bulbophyllumspecies (Orchidaceae). Ann Bot 93:499–505
van der Pijl L, Dodson CH (1966) Orchid flowers: their pollination
and evolution. University of Miami Press, Coral Gables
Vieira MF, Andrade MRS, Bittencourt NS, Carvalho-Okano RM
(2007) Flowering phenology, nectary structure and breeding
system in Corymborkis flava (Spiranthoideae: Tropidieae), a
terrestrial orchid from a Neotropical forest. Aust J Bot 55:635–
642
Vogel S (1990) The role of scent glands in pollination (transl. by
Bhatti JS). Smithsonian Institute, Washington, DC
Osmophores and nectaries in Acianthera (Orchidaceae) 151
123
Related Documents











