Thomas Austin Acetylcholine and dopamine systems Page 1 of 21 Acetylcholine and dopamine systems: Outlining the pharmacological issues for motor disorders Completed as part of a BSc Psychology degree. Submission: 7/01/2014 Word Count: 2811

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Thomas Austin Acetylcholine and
dopamine systems
Page 1 of 21
Acetylcholine and dopamine systems: Outlining the
pharmacological issues for motor disorders
Completed as part of a BSc Psychology degree.
Submission: 7/01/2014
Word Count: 2811

Thomas Austin Acetylcholine and
dopamine systems
Page 2 of 21
Acetylcholine and dopamine systems: Outlining the pharmacological issues
for motor disorders
Abstract
An explorative review of literature of pharmacological aspects of acetylcholine
(ACh) and dopamine (DA) neurotransmitter systems on the methods of causation of
motor dysfunction, ranging from dyskinesia to tremulous jaw movements. The
current author comments on the ACh system with reference to nicotinic ACh receptor
(nAChR) blockade, competitive antagonism and tacrine-induced jaw tremor
research. The DA system implication is also reviewed and raises current topical
issues models of neuro-circuitry and ACh-DA interactions. To conclude, the ACh and
DA systems and their interactions require future research to establish concrete
mechanisms and the complexities of these system interactions to be explored in
relation to dysfunctional motor behaviour.

Thomas Austin Acetylcholine and
dopamine systems
Page 3 of 21
1. Introduction
Acetylcholine (ACh) and dopamine (DA) are two neurotransmitters that
inhabit the brain and despite their differences are related in many neurological
structures (Parent & Hazrati, 1995) and behaviours (Nunes, Randall, Podurgiel,
Correa, & Salamone, 2013).
ACh is involved in the autonomic nervous system and can be observed in the
peripheral and central nervous systems. Initially ACh was found in cardiovascular
tissue, in which it is inhibitory but research has progressed the documentation of the
neurotransmitter in the brain and on other motor behaviours (Edwards, Dolezal,
Zemkova, & Vyskocil, 1985).
The ACh system is based upon two main types of receptor; nicotinic
(nAChRs) and muscarinic (mAChRs) receptors. The nAChRs are ligand-gated
channels comprised of five subunits; α1 to α6, β1 to 4, γ, δ, and ε. There are many
nAChR variants as the receptors can be hetero- or homo-pentamers, with each
variant expressing a unique affinity for the possible agonist and antagonist agents
(Papke & Porter Papke, 2002). Muscarinic receptors are metabotropic and therefore
use G proteins as their method of signalling, caused by ACh binding to one of seven
transmembrane areas.(Ishii & Kurachi, 2006). This receptor only has five types (M1-
M5), despite M5 being pharmacologically elusive but genetically identified (Caulfield
& Birdsall, 1998) The role of mAChR antagonists in parkinsonian symptomology is
easily documented as several agents are used in the parkinsonian treatment
(Langmeand, Watson, & Reavill, 2008).

Thomas Austin Acetylcholine and
dopamine systems
Page 4 of 21
DA is a neurotransmitter with complex roles in behaviour, from addiction
(Diana, 2011) to Parkinson’s disease (Merims & Giladi, 2008). Many psychological
disorders are attributed to dopamine-related abnormality such as schizophrenia and
attention deficit hyperactivity disorder (ADHD). DA is also secreted and has purpose
outside the brain.
DA receptors (DARs) are split into two families; D1-like (D1 and D5) and D2-
like (D2, D3 and D4) subtypes. Postulation surrounding a sixth and seventh subtypes
(D6 and D7) was expressed, however there is considerable lack of supporting
research (Contreras et al., 2002).
2. Acetylcholine
2.1. Movement disorders
Research that implicates ACh as playing a major role in movement disorders
uses a wide range of techniques and experiment types. Research with animals,
stemming from agricultural toxicology research into livestock disease, where as other
research utilises a pharmacological and behavioural paradigm. A toxicological
article–regarding the nAChR antagonists (deltaline and methyllycaconitine (MLA))
and agonists (nicotine and anabasine) from Delphinium and Nicotiana species,
respectively– indicates support for AChs involvement in motor disorders (Aiyar et al.,
1979).
The authors studied the agents’ method of mechanism in the body and found
deltaline and MLA in excessive amounts to cause competitive antagonism-induced
paralysis via blockade of nAChRs at post-synaptic neuromuscular sites. However,
the experiment was a descriptive exploration of the method of action, rather than
adhering to a pharmacological paradigm.

Thomas Austin Acetylcholine and
dopamine systems
Page 5 of 21
However, the psychopharmacological implications of these nAChR
antagonists and agonists’ methods of action were not commented upon in the
publication. A recent study (Welch, Pfister, Lima, Green, & Gardner, 2013) applied
Aiyar et al.’s work by studying the action of both nAChR antagonists and agonists, of
the Delphinium species, in pharmacological experiments on mice to further define
the chemical actions on the neurology of the subjects.
Welch et al. used non-lethal doses of the two isolated toxins in the
Delphinium species: deltaline and MLA, as well as nicotine and anabasine. However,
Welch et al. uses the substances to specifically evaluate the effect on motor
coordination in the subject using a battery of assorted motor performance tasks; a
balance beam, a grip strength, a rotarod, open field analysis and featured a tremor
monitor.
The results showed significance between the non-lethal dose agonists and
antagonists used and acute movement and coordination dysfunction, and then the
subject undergoes a brief period of inactivity. This illustrates the control of ACh and
the endogenous receptors on movement signalling on a neuronal level, agonist and
antagonists of nAChRs can both cause motor dysfunction. The study holds an
agricultural rationale by explaining the connection between Delphinium toxicity and
the loss of livestock due to poisoning (paralysis). However, the aforementioned study
(Welch et al., 2013) can be criticised due to the lack of generalisation of results to a
human population as the subjects were mice.

Thomas Austin Acetylcholine and
dopamine systems
Page 6 of 21
To elucidate; the methodology features the use of a median lethal dose
(LD50) of the administered agents for mice to find the non-lethal dose. The non-
lethal dose was calculated by finding 75% of the LD50. However, the LD50 relies on
many factors such as metabolic rate, agent-associated cytoarchitectual damage and
many environmental and genetic factors regarding receptor frequency, sensitivity
and physical distribution. Therefore, the results cannot be used to represent a
human population as it may result in an over/under pronounced or otherwise skewed
effect on motor dysfunction and/or nAChR activity.
Acetylcholine (ACh)
nAChR
Agonists
nAChR
Antagonists
Diagram 1. Illustration of the antagonists (deltaline and methyllycaconitine (MLA)) and agonists (nicotine and
anabasine) that were used in research (Welch et al., 2013). The 2D ACh chemical has been added to represent
the ACh system.

Thomas Austin Acetylcholine and
dopamine systems
Page 7 of 21
Furthermore, the pharmacokinetics of deltaline and MLA have not been
evaluated and remains unclear. This information would aid pharmacological
interpretation of results.
Anabasine has been found to cause sustained muscle paralysis in adult and
embryo-stage animals, in-utero, thus causing teratogenic effects by manipulation of
the nACh system triggering motor dysfunction in (Green, Lee, Panter, & Brown,
2012) teratogenic effects are due to the motor dysfunction in critical stages of foetal
development (Weizweig et al., 2008).
Furthermore, a study supporting the postulation of ACh enrolment in motor
dysfunction symptomology used an indirect study design, focusing on nicotine as an
AChR agonist; nicotine initially stimulates nAChRs in the ACh system (Wang & Sun,
2005). However after prolonged and persistence agonism the receptors cause
tachyphalaxis, a sudden reduction in receptor sensitivity. Further evidence of AChs
role in movement disorders comes from a review of tacrine, an acetylcholinesterase
and an Alzheimer’s disease treatment, and links with tremulous jaw movements
(TJMs)(Cousins et al., 1999). The TJM is a type of tremor, a dysfunctional movement
disorder characteristic of parkinsonian syndrome (Keltner, 1994).
The authors accompanied the review with three experiments to determine the
extent, if any, that ventrolateral striatal ACh modulates TJMs in-vivo (rats, n=89).
Several doses of tacrine were used; 2.5mg/kg and 5mg/kg. The first experiment
consisted of subject’s ventrolateral neostraitum micro-injected with an ACh synthesis
inhibitor (hemicholinium-3) in rats exhibiting tacrine-induced TJMs. The result of this
was a reduction in the TJM severity, alleviating the motor dysfunction by reducing
the ACh available for binding in the synapse.

Thomas Austin Acetylcholine and
dopamine systems
Page 8 of 21
A repeat of the initial experiment was conducted, but was altered and used as
a control against the natural diffusion of hemicholinium-3 to other cortices in the
surrounding area and causing unwanted effects. The importance of the ventrolateral
neostriatum was supported by a previous study, using scopolamine and the
methylated derivative to reduce TJMs (Mayorga, Carriero, Cousins, Gianutsos, &
Salamone, 1997).
Fig1. A scatterplot illustrating the correlational relationship between ACh blood levels and number
of TJMs spotted. Circles mark each control group subjects; squares mark the 2.5mg/kg tacrine group
subjects; triangles mark 5.0mg/kg tacrine group subjects (Cousins et al., 1999, p.445).

Thomas Austin Acetylcholine and
dopamine systems
Page 9 of 21
Cousins et al. then conducted a final experiment, tacrine-induced TJM
extracellular ACh levels were assessed using, a surgically placed, ventrolateral
striatum dialysis module (1999). The authors found a 5mg/kg tacrine dose to cause
acute increase in ventrolateral ACh levels, 324-fold of baseline. This increase of ACh
was statistically significant when correlated with TJMs (r =+.56, p< .0001). The
authors’ concluded that ventrolateral ACh system plays a substantial role in
tremulous movements and therefore in parkinsonian symptoms.
Despite Cousins et al.’s high standard of research methods, from meticulous
record keeping to an effective review of past literature, the study can be criticised on
several key points. Primarily, despite obtaining a significant overall effect, the groups
that received 2.5mg/kg of tacrine showed no significant relationship between
ventrolateral neostriatal ACh and TJMs. This highlights the possible reductionism of
the implied overall results as the spread of the significant correlations is not even.
The authors correlations are as follows; 5mg/kg (r= +.9, p< .001), 2.5mg/kg (r= + .45,
no significance), combination of 5mg/kg and 2.5mg/kg (r= + .56, p< .05). However,
the statistics conducted between the ventrolateral neostriatal ACh levels and TJMs in
the 2.5mg/kg and 5mg/kg tacrine-dosed groups were found to be significant.
Secondly, the authors’ comment on the possibility that tacrine’s alteration of ACh
levels may not be as presumed –as an acetylcholinesterase inhibitor— but from
several other methods of mechanism such as an increase of receptor sensitivity or
an encouragement of ACh release. The lack of generalisation of the results to a
human population hinders the results; however, Cousins et al. included and
discussed research regarding human in-vivo tacrine symptoms in Alzheimer patients.
This final issue can be overlooked as in-vivo human testing would be unethical and
rats are better than many other alternative populations due to brain similarities.

Thomas Austin Acetylcholine and
dopamine systems
Page 10 of 21
In addition, an α4 nAChR mutation in mice was found to elicit akinesia,
muscle rigidity and tremor symptomology. This symptomology is recognised in
parkinsonian syndrome and other movement disorders. The pharmacological
blockade and genetic deletion of nAChRs does not affect movement. However,
genetic mutation on the coding gene for α4 nAChRs (Leu9’Ala) in dopaminergic
neurones in the dorsal striatum can result in hypersensitivity of α4 receptors when
agonised (Soll, Grady, Salminen, Marks, & Tapper, 2013). The authors compared
wild-type (WT) and Leu9’Ala genes in mice and used both groups with an α4β2
nAChR antagonist (dihydro-β-erythroidine; DHβE). In WT mice the DhβE cause no
motor behaviour affect, however when the α4 nAChR antagonism occurs in Leu9’Ala
mice reversible motor dysfunction is exhibited resulting in increased movement
speeds, akinesia, catalepsy and tremors. This outlines the relationship between
nAChRs and their role in mediation of parkinsonian-type symptoms and suggests
that genetic disposition can place vulnerabilities within a component of the ACh
system (genetics for one specific receptor subunit) instead of the entire system itself.
The proposed phenomenon uses the desensitivity of the genetically altered α4
nAChR as a basis for motor dysfunction but the authors have linked this, indirectly,
to the dopaminergic motor pathway circuit (i.e. D1-5 agonism) (Soll et al., 2013).

Thomas Austin Acetylcholine and
dopamine systems
Page 11 of 21
2.3. Conclusion
In conclusion, research regarding nAChR antagonism (Aiyar et al., 1979;
Welch et al., 2013) and agonism (Green et al., 2012; Wang & Sun, 2005) shows that
either activity at the nAChR site can cause motor dysfunction such as paralysis.
However, the method of action is complex and needs to be focused on: The ACh
system is not directly in control of movement dysfunction observed but is a result of
ACh mediation in brain areas enrolled in motor capabilities (Cousins et al., 1999;
Keltner, 1994; Mayorga et al., 1997). Some research supports the postulation that
genetics can play a role in nAChR-related motor dysfunction (Soll et al., 2013).

Thomas Austin Acetylcholine and
dopamine systems
Page 12 of 21
3. Dopamine
3.1. Movement disorders
In a recent review of the pharmacology (2007) of L-DOPA, a parkinsonian
motor dysfunction treatment drug, and the neural circuitry involved in prolonged L-
DOPA-induced dyskinesia symptomology (Cenci). The motor dysfunction is
mediated by extracellular DA in the basal ganglia and striatal areas and causes a
cascade of alterations resulting in the expression of genes that cause predisposition
for long-term dyskinesia. This occurs primarily in the D1 DA subunit receptors in the
striatal neurones, which encourage inhibitory neurotransmitters (prodynorphin) and
inhibitory enzyme secretion, such as glutamic acid decarboxylase. As a result the
globus pallidus’ internal segment’s (GPi) and substantia nigra pars reticulate’s
inhibition causes dysfunctional neocortical activity produces dysfunctional movement
(Cenci, 2007). The researcher concluded that this cascade affects the GABA signal
projection into the basal ganglia, further producing abnormal activation in motor
structures. Cenci (2007) uses an extensive literature review to support the neuro-
circuitry model proposed. In summary; DA’s modulation of movement dysfunction in
L-DOPA-induced parkinsonian-type symptoms.
Moreover, Cenci’s review is based upon a paper (Parent & Hazrati, 1995) that
reviewed research on the cortical basal ganglia and cortico-thalamic loop. This
article further documented and commented upon the importance of basal ganglical
modulation of premotor neurones and psychomotor behaviours.
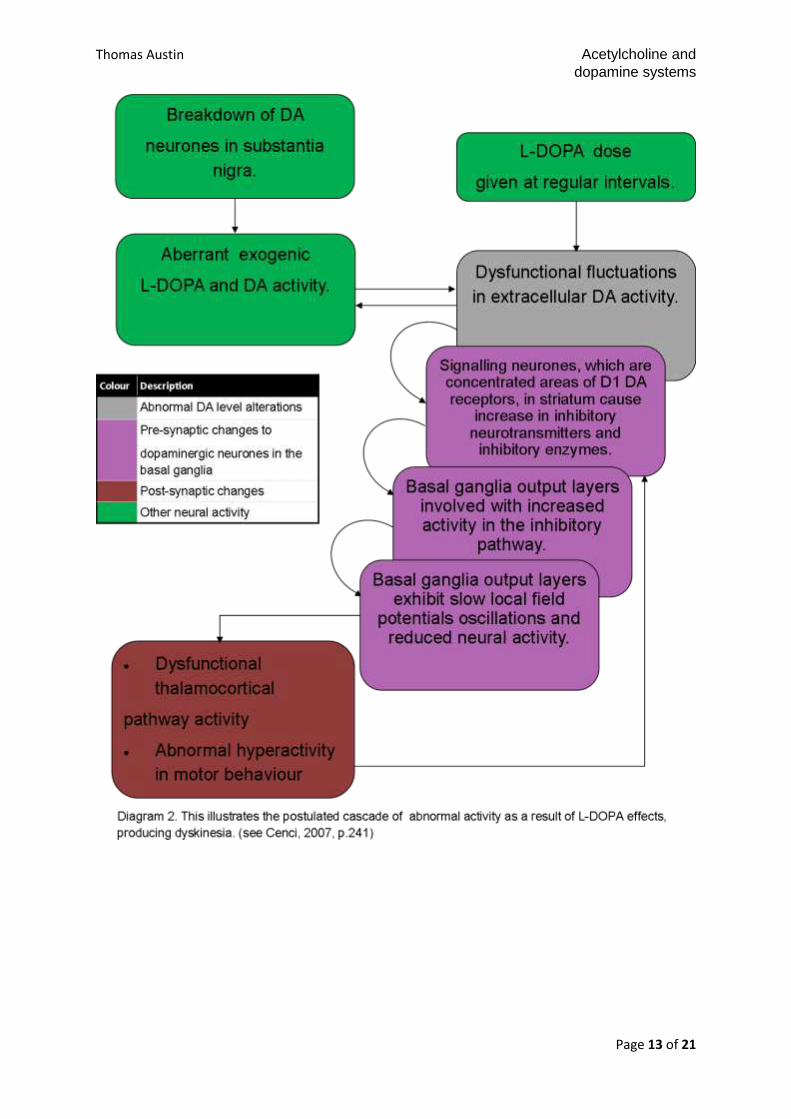
Thomas Austin Acetylcholine and
dopamine systems
Page 13 of 21

Thomas Austin Acetylcholine and
dopamine systems
Page 14 of 21
Furthermore, in 2010, hindbrain pedunculopontine and laterodorsal tegmental
nuclei project into the midbrain DA neurons causing nACh and mACh receptor
neurone excitation within the substantia nigra and ventral tegmental structure
(Lester, Rogers, & Blaha); thus controlling the flow of dopamine into the dorso-
ventral striatum and prefrontal cortex areas. Forebrain DA modulation has been
recorded to stem from ACh neurones in the striatum, further implementing the area
in ACh and DA-related dysfunctional motor behaviour.
Hoebel, Avena, & Rada supported the link between ACh and DA activity
modulation of dysfunctional parkinsonian motor symptoms (2007).
An experiment in 1975 investigated the behavioural products of DA,
haloperidol (an inverse DA agonist), carbachol (AChR agonist) and atropine (mAChR
antagonist) injection in rhesus monkeys, individually and in combinations(Cools,
Hendricks, & Korten) . DA injections caused a dynamic phase (P1; non-dysfunctional
increase in manipulative movement), the expression of spasmodic torticollis –related
to dystonic movements) in the dystonic phase (P2) and the dyskinetic phase (P3)
where speech and other limb dyskinesia was produced. DA-induced P1 and P2 were
counteracted by haloperidol, but there was no effect on DA-induced P3. Carbachol-
induced P2 was enhanced by high DA dose, but P4 was eliminated in subjects.
Carbachol induced P1, P2 and P3 as well as an additional epileptic phase (P4),
characterised by generalised epileptic seizures. Finally, atropine countered P1, P2
and P4 but not P3. These finding suggest that the striatal ACh-DA interactions are
more complicated than the two-dimensional balance model but propose that the
model is involved in the mediation of generalised epileptoid seizures. The study also
highlighted a secondary method of action relating to speech dyskinesia behaviour;
implementing neurotransmitters other than DA.

Thomas Austin Acetylcholine and
dopamine systems
Page 15 of 21
However, again the generalizability of studies that use animal subjects is an
issue with Cools, Hendricks and Korten’s study. This is because, despite numerous
similarities in neurology as well as behaviour, animals such as rhesus monkeys are
ultimately not human and may have different within-species variation of receptor
variables. Nevertheless, the results are link to human behavioural phenomena such
as parkinsonian-type motor dysfunction.
Despite this limitation, Cools, Hendricks and Korten have efficiently utilised
pharmacological methodology to support a less reductionist model of ACh-DA
interactions and effects on motor behaviour. Replications of this study should use a
wider range of pharmacological agents to test for further underlying mechanisms of
the brains motor areas.
The basal ganglia have been implicated as the main structure involved in the
interactions between ACh and DA systems in the brain. Lehmann and Langer
demonstrated the need for a balance between the two neurotransmitters. An
increase in ACh can be DA-induced and is associated with motor dysfunction, as
well as high levels of DA being attributed to hyperkinesia symptoms (Lehmann &
Langer, 1983).
Parkinsonian symptom progression has been correlated with DA neurone
terminal degradation, located in the basal ganglia, using a [18F]dopa-PET scan
technique and is used to monitor disease progression (Morrish, Sawle, & Brooks,
1996). However, this study has several main criticisms. Primarily, the issue of
reductionism, as ACh and other neurotransmitters are left out of the model used.
Secondly, a criticism regarding the sample being too small (n=10) for reliability is a
marker for future replication despite using clinical and PET scan methods.

Thomas Austin Acetylcholine and
dopamine systems
Page 16 of 21
3.3 Conclusion
In conclusion, the link between DA systems and motor dysfunction is
thoroughly documented (Cenci, 2007; Cools et al., 1975; Lester et al., 2010) and
research has clarified the link between the DA-related dysfunction and modern day
motor disorders such as Parkinson’s disease (Morrish et al., 1996). However, Cools,
Hendricks and Korten (1975) have commented and refuted a reductionist
pharmacological mechanism of action model of DA and introduced a new, multi-
neurotransmitter model of motor dysfunction. A [18F]dopa-PET scan study (Morrish
et al., 1996) highlights the need for large sample size to avoid criticism of reliability.
However, the research methods employed are time demanding and restrictive,
hindering replicability but enhancing methodological precision.

Thomas Austin Acetylcholine and
dopamine systems
Page 17 of 21
4. Summary
ACh and DA system models of mechanism should not be reduced and should
be fully explored pharmacologically. A range of neural structures are related to these
two neurotransmitters and should be link using future studies to parkinsonisan and
other motor dysfunction disorders. The applications of research into parkinsonian
and other motor dysfunction disorders can have great impact for design of
pharmacological treatments and would therefore increase the quality of life for those
who have Parkinson’s disease or tremor movements. Disorders that are motor-
related can cause immense impacts to individuals and their relations, but these
disorders can be induced by medication for already severe disorders. As seen in
Alzheimer patients who use tacrine (Keltner, 1994; Mayorga et al., 1997).
Future research should include a wide variety of experimental techniques to
further characterise the neural correlates of motor dysfunction structures using brain
mapping. However, the practicality of conducting neuro-imaging on a subject that
may have a lack of control of movement any result in unusable data. More research
using a wide range of ACh and DA agents should be combined to determine if the
differing receptor subtypes have different effects on behaviour. Subtype selective
agonists and antagonists should also be utilised to pharmacologically measure the
differences between each receptor type and elicited behaviour; the combination of
nACh inhibitors and one selective nAChR agonist could be used to investigate this.

Thomas Austin Acetylcholine and
dopamine systems
Page 18 of 21
References;
Aiyar, V. N., Benn, M. H. M., Hanna, T., Jacyno, J., Roth, S. H., & Wilkens, J.
K. (1979). The principle toxin of Delphinium brownii rydb. and its mode of action.
Experentia, 35(10), 1367–1368.
Caulfield, M. P., & Birdsall, N. J. (1998). International union of pharmacology.
XVII. Classification of muscarinic acetylcholine receptors. Pharmacological Reviews,
50(2), 279–290.
Cenci, M. A. (2007). Dopamine dysregulation of movement control in L-
DOPA-induced dyskinesia. Trends in Neurosciences, 30(5), 236–243.
Contreras, F., Foullllioux, C., Bolivar, A., Simonovis, N., Hernandez-
Hernandez, R., Armas-Hernandez, M., & Velasco, M. (2002). Dopamine,
hypertension and obesity. Journal of Human Hypertension, 16(Suppliment 1), S13–
17.
Cools, A. R., Hendricks, G., & Korten, J. (1975). The acetylcholine-dopamine
balance in the basal ganglia of rhesus monkeys and its role in dynamic, dystonic,
dyskinetic, and epileptoid motor activities. Journal of Neural Transmission, 36(2),
91–105.
Cousins, M. S., Finn, M., Trevitt, J., Carriero, D. L., Conlan, A., & Salamone,
J. D. (1999). The role of ventrolateral striatal acetylcholine in the production of
tacrine-induced jaw movements. Pharmacology, biochemistry, and behavior, 62(3),
439–447.
Diana, M. (2011). The dopamine hypothesis of drug addiction and its potential
therapeutic value. Frontiers in Psychiatry, 2(64).

Thomas Austin Acetylcholine and
dopamine systems
Page 19 of 21
Edwards, C., Dolezal, V., Zemkova, H., & Vyskocil, F. (1985). Is an
acetylcholine transport system responsible for non-quantal release of acetylcholine
at the rodent myoneural junction? Proceedings of the National Academy of Sciences
of the United States of America, 82(10), 3514–3518.
Green, B. T., Lee, S. T., Panter, K. E., & Brown, D. R. (2012). Piperidine
alkaloids: Human and food animal teratogens. Food and Chemical Toxicology, 50(6),
2049–2055.
Hoebel, B. G., Avena, N. M., & Rada, P. (2007). Accumbens dopamine-
acetylcholine balance in approach and avoidance. Current opinion in pharmacology,
7(6), 617–627. doi:10.1016/j.coph.2007.10.014
Ishii, M., & Kurachi, Y. (2006). Muscarinic acetylchonline receptors. Current
Pharmaceutical Design, 12(28), 3573–3581.
Keltner, N. L. (1994). Tacrine: A pharmacological approach to Alzheimer’s
disease. Journal of Psychosocial Nursing and Mental Health Services, 32(3), 37–39.
Langmeand, C. J., Watson, J., & Reavill, C. (2008). Muscarinic acetylcholine
receptors as CNS drug targets. Pharmacology & Therapeutics, 117(2), 232–243.
Lehmann, J., & Langer, S. Z. (1983). The striatal cholinergic interneuron:
Synaptic tartget of dopaminergic terminals? Neuroscience, 10, 1105–1120.
Lester, D. B., Rogers, T. D., & Blaha, C. D. (2010). Acetylcholine-dopamine
interactions in the pathophysiology and treatment of CNS disorders. CNS
Neuroscience & Therapeutics, 16(3), 137–162. doi:10.1111/j.1755-
5949.2010.00142.x

Thomas Austin Acetylcholine and
dopamine systems
Page 20 of 21
Mayorga, A. J., Carriero, D. L., Cousins, M. S., Gianutsos, G., & Salamone, J.
D. (1997). Tremulous jaw movements produced by acute tacrine administration:
possible relation to parkinsonian side effects. Pharmacology biochem behaviour, 56,
273–279.
Merims, D., & Giladi, N. (2008). Dopamine dysregulation syndrome, addiction
and behavioral changes in Parkinson’s disease. Parkinsonism & Related Disorders,
14(4), 273–280.
Morrish, P., Sawle, G., & Brooks, D. (1996). An [18F]dopa–PET and clinical
study of the rate of progression in Parkinson’s disease. Brain, 119(2), 585–591.
Retrieved from http://brain.oxfordjournals.org/content/119/2/585.short
Nunes, E. J., Randall, P. a, Podurgiel, S., Correa, M., & Salamone, J. D.
(2013). Nucleus accumbens neurotransmission and effort-related choice behavior in
food motivation: Effects of drugs acting on dopamine, adenosine, and muscarinic
acetylcholine receptors. Neuroscience and biobehavioral reviews, 37(9), 1–11.
doi:10.1016/j.neubiorev.2013.04.002
Papke, R. L., & Porter Papke, J. K. (2002). Comparative pharmacology of rat
and human alpha7 nAChR conducted with net charge analysis. British Journal of
Pharmacology, 137(1), 49–61.
Parent, A., & Hazrati, L. (1995). Functional anatomy of the basal ganglia .
Brain Research Reviews, 20(1), 91–127.
Soll, L. G., Grady, S. R., Salminen, O., Marks, M. J., & Tapper, A. R. (2013).
A role for α4(non-α6)* nicotinic acetylcholine receptors in motor behavior.
Neuropharmacology, 73C, 19–30. doi:10.1016/j.neuropharm.2013.05.001
Wang, H., & Sun, X. (2005). Desensitized nicotinic receptors in brain. Brain
Research Reviews, 48(3), 420–437.

Thomas Austin Acetylcholine and
dopamine systems
Page 21 of 21
Weizweig, J., Panter, K. E., Patel, J., Smith, D. M., Spangenberger, A., &
Freeman, M. B. (2008). The fetal cleft palate: v. elucidation of the mechanism of
palatal clefting in congenital caprine model. Plastic Reconstructive Surgery, 121(4),
1328–1334.
Welch, K. D., Pfister, J. a, Lima, F. G., Green, B. T., & Gardner, D. R. (2013).
Effect of α7 nicotinic acetylcholine receptor agonists and antagonists on motor
function in mice. Toxicology and applied pharmacology, 266(3), 366–74.
doi:10.1016/j.taap.2012.11.024
Related Documents











