- 1 - Accelerated Detection and Diagnosis of Invasive Alien Pests and Pathogens in Imported Plants Steve Woodward 1 , Eleni Siasou 1 , Anna Brown 2 , Benjamin Drake 3 , Rainer Ebel 4 , Paul Gascoine 5 , Jean-Claude Grégoire 6 , Gail Preston 7 , Michael Shaw 8 & Stewart Snape 9 1: University of Aberdeen, Institute of Biological and Environmental Sciences, Department of Plant and Soil Science, Cruickshank Building, St. Machar Drive, Aberdeen AB24 3UU, UK. 2: Forest Research, Alice Holt Lodge, Farnham, Surrey GU10 4LH. 3: ADAS, Pendeford House, Wobaston Road, Wolverhampton WV9 5AP 4: University of Aberdeen, School of Natural and Computing Sciences, Department of Chemistry, Meston Building, Aberdeen AB24 3UE. 5: Smiths Detection plc, Bushey, Watford WD23 2BW. 6: Lutte Biologique et Ecologie Spatiale , CP160/12, av. F.D. Roosevelt 50, 1050 Bruxelles, Belgium. 7: Department of Plant Sciences, University of Oxford, South Parks Road, Oxford, OX1 3RB. 8: University of Reading, School of Agriculture, Policy and Development, Whiteknights, Reading RG6 6AR. 9: Forestry Commission, Silvan House, Corstorphine Road, Edinburgh EH12 7AT. Contacts: [email protected] ; [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

- 1 -
Accelerated Detection and Diagnosis of Invasive Alien Pests and Pathogens in Imported Plants
Steve Woodward1, Eleni Siasou1, Anna Brown2, Benjamin Drake3, Rainer Ebel4, Paul Gascoine5, Jean-Claude Grégoire6, Gail Preston7, Michael Shaw8 & Stewart Snape9
1: University of Aberdeen, Institute of Biological and Environmental Sciences, Department of Plant and Soil Science, Cruickshank Building, St. Machar Drive, Aberdeen AB24 3UU, UK.
2: Forest Research, Alice Holt Lodge, Farnham, Surrey GU10 4LH.
3: ADAS, Pendeford House, Wobaston Road, Wolverhampton WV9 5AP
4: University of Aberdeen, School of Natural and Computing Sciences, Department of Chemistry, Meston Building, Aberdeen AB24 3UE.
5: Smiths Detection plc, Bushey, Watford WD23 2BW.
6: Lutte Biologique et Ecologie Spatiale , CP160/12, av. F.D. Roosevelt 50, 1050 Bruxelles, Belgium.
7: Department of Plant Sciences, University of Oxford, South Parks Road, Oxford, OX1 3RB.
8: University of Reading, School of Agriculture, Policy and Development, Whiteknights, Reading RG6 6AR.
9: Forestry Commission, Silvan House, Corstorphine Road, Edinburgh EH12 7AT.
Contacts: [email protected]; [email protected]

- 2 -
Contents
Summary................................................................................................................................................................. - 3 -
Introduction ............................................................................................................................................................ - 4 -
Volatile Organic Compounds Produced by Plants .................................................................................................. - 6 -
Changes in VOC Signatures in Stressed Plants........................................................................................................ - 7 -
Volatile Organic Compounds from Microorganisms ............................................................................................ - 15 -
The application of VOC profiling in disease detection.......................................................................................... - 20 -
Methods for detecting volatile chemicals ............................................................................................................ - 22 -
Statistical and economic aspects of improved biosecurity .................................................................................. - 26 -
Economic justifications for additional detection technologies ............................................................................ - 30 -
How Can These Methods be Applied in Practice? ................................................................................................ - 32 -
Conclusions ........................................................................................................................................................... - 34 -
References ............................................................................................................................................................ - 35 -

- 3 -
Summary
The possibilities of utilizing state-of-the art chemical detection instrumentation and image analyses to diagnose the presence of alien invasive pests and pathogens in imported plants and plant products are reviewed.
All organisms produce volatile organic compounds (VOCs) which can be detected by a range of instruments already widely in use at ports.
Over 100,000 VOCs produced by plants are known.
VOCs produced by some of the organisms that interact with plants are known. For example:
o Bark beetles are known to both alter organic molecules already present in host tissues, and to produce others themselves. These VOCs have roles in communicating with other individuals of the same species.
o Bacteria and fungi produce a range of relatively simple and easily detected VOCs during different phases of growth.
The suite (profile) of VOCs produced by an organism following stress differs from those produced by the healthy organism.
VOCs produced by healthy versus damaged plants differ significantly, both quantitatively and qualitatively. Moreover, VOCs produced by the same plant may differ depending on the precise cause of the damage/stress.
There is increasing research on the production of VOCs by microorganisms and by many insects, presenting a wide range of specific VOCs which can be used in developing this science for the detection of pests and pathogens.
Given the packing and transport systems used to export plants and plant products, it is likely that VOCs analyses could be utilized in the detection and diagnosis of the presence of alien invasive pests and pathogens in consignments.
Moreover, the widespread presence and use of highly sensitive and discriminatory instrumentation at ports throughout the world means that, with further calibration, existing equipment could be used in this work.

- 4 -
Introduction During importation, the detection and diagnosis of potentially invasive pests and pathogens
on plants at air- and sea-ports of entry into the UK is a major focus of phytosanitary
inspections. The authorities intercept many such organisms in consignments of plants and
plant-based products during the routine surveys undertaken, but when the numbers of units
inspected, approximately 2%, is set against the discovery rate, it is clear that the number of
invasive pests and pathogens of plants coming into the UK may be very high. The number of
recent pest and disease epidemics that have arisen over the last 20-30 years is also clear
evidence of a failure in phytosanitary regulation on an international scale (Brasier 2008;
Santini et al. 2013).
Many of the invasive organisms not detected at ports of entry have or are
establishing in our forests and woodlands, resulting in high rates of damage and threatening
entire ecosystems. The public outcry in late October 2012, following the confirmation that
the ash dieback pathogen Hymenoscyphus pseudoalbidus was already present in the UK ash
woodlands, illustrated how seriously the general public now takes these problems. Yet the
ash dieback outbreak was only one in a series of serious influxes of alien pests and
pathogens that have become established in our forests and woodlands in the last 20 – 30
years (Brasier 2008; Santini et al. 2013). Clearly, if global trade in plants and plant products,
which is worth billions of Euros world-wide, is to continue, our ability to detect potential
problematic organisms in trade must be very much improved.
The problems posed by alien invasive pests and pathogens and the lack of ability to
make wide-ranging inspections of incoming plants and plant materials at ports, illustrate the
urgent need for more stringent techniques for detection and diagnosis of these damaging
organisms. Current methods of detection, relying on visual and/or molecular (DNA)-based
detection techniques, are extremely useful and powerful, but have serious constraints in
terms of their overall effectiveness:
1. Both methods must be highly targeted to be successful
a. Inspectors cannot examine (visually) more than a very small proportion of
imported plants;

- 5 -
b. The molecular approach has similar constraints, as the detection of pests and
pathogen depends on sampling tissues that contain DNA of the pest or
pathogen.
2. Both visual and molecular methods are labour intensive and require highly trained
personnel.
3. Both methods are expensive to implement.
The purpose of this review, initiated under DEFRA contract TH0122, is to examine the
potential for alternative methods for detecting potentially damaging invasive pests and
pathogens in imports of plants and products. It is not the intention to suggest that the
current methods should be replaced with this technology, but to add another level of
detection capacity to the system, enabling a more targeted approach for subsequent visual
and molecular detection and diagnosis techniques.
All organisms produce an array of volatile organic compounds (VOCs), which are in
themselves, or as a profile, apparently characteristic of the emitting organism. Moreover,
the profile of VOCs for a given organism changes when that organism is under stress,
whether from a disease-causing agent, or an abiotic agent. It is not the intention in this
review to list all VOCs produced by plants under non-stressing and stressing conditions, but
it is clear that a huge number of plant VOCs can be used to distinguish between species and
within even cultivars and varieties to determine health status, possibly even the cause of
the stress, whether abiotic or biotic. These quite simple organic molecules can be rather
easily detected by the instrumentation currently available at ports (see below).
A first challenge facing researchers aiming to use plant VOCs to detect attack by
pests or disease-causing organisms, however, is selecting which VOCs to study. VOCs used
for the presence of pests or disease can originate from three main sources: the plant, the
pest/pathogen or the action of the pest/pathogen on plant tissues. VOCs are produced by
both aerial and root tissues, although most disease detection focused studies published to
date have focused on foliar VOCs, which are typically released to the atmosphere in higher
concentrations than are root-associated VOCs.

- 6 -
Volatile Organic Compounds Produced by Plants The production of a wide variety of VOCs by plants is well known and was the subject of a
recent review (Dudareva et al. 2013). Humans have used many such products from species
such as basil, garlic, lavender, rosemary, camphor, damask and other roses, pine (and other
gymnosperms), and many spice plants, in perfumes, food and drink flavourings,
preservatives and solvents for centuries. Many other fragrant plants feature strongly in
human history and are well documented in ancient texts. For example, the aromatic
properties of food plants are of immense importance to feeding choices of animals,
including humans (Goff & Klee 2006).
More than 100,000 metabolites produced by plants are known, many of which are
classified as VOCs (Dicke & Loreto, 2010). For example, over 1700 volatile compounds floral
scent compounds have been identified (Knudsen et al. 2006); a high proportion of the 7000
flavour VOCs known from food and beverages are of plant origin (Nijssen et al. 1997). VOCs
can be regarded as ‘infochemicals’, which are released, and which act in plant-plant, plant-
animal and plant-pathogen interactions (Maffei, 2010).
Plant VOCs have a highly significant contribution to VOC emissions overall, with over
50% of VOC emissions per annum in the USA, and over 67% of emissions world-wide coming
from plants (Guenther 1997). It has been estimated that monoterpene emissions from
plants alone are responsible for the release of between 127 and 480 megatonnes of carbon
into the atmosphere globally per annum (Kesselmeier & Staudt 1999).
Plant VOCs fall into two main categories:
constitutive VOCs, which are frequently stored in plant tissues and volatilised
according to their concentration and physicochemical properties, and
inducible VOCs, which are synthesised in response to environmental stimuli.
The distinction between constitutive and inducible VOCs is somewhat ambiguous, however,
as many of the constitutive VOCs normally released from healthy intact plants become
inducible volatiles (IVOCs) after foliar damage, when larger quantities or different ratios are
produced (Holopainen, 2004). Both have uses in the detection of stressed plants, as novel
IVOCs may be produced in response to infection, while changes in the profile and quantity
of constitutive VOCs can also be used to provide information on plant health and disease.

- 7 -
Typical examples of fragrant VOCs produced by plants are the monoterpenes
produced as significant components in the essential oils of the Pinaceae and Citrus spp.,
amongst many others (Hunter & Brogden 1965; Moshonas & Shaw, 1984; Hannover 1992).
Pine resin is principally composed of compounds derived from isoprene and synthesized via
the mevalonic acid and/or methylerythritol pathways, including hemiterpenes,
monoterpenes, diterpenes, sesquiterpenes and carotenoids (Gershenzon & Croteau 1990).
In addition, VOCs may arise from the lipoxygenase pathway (e.g. jasmonates) or the shikimic
acid pathway (phenolic compounds, phenylpropanoids and benzenoids).
Changes in VOCs during fruit ripening have been studied in some detail, to
determine what humans perceive as attractive scents (e.g. Goff & Klee 2006). Tomato fruit,
for example, as it ripens is associated with the appearance of scents of ‘tomato’; these
scents are based on derivatives of linolenic and linoleic acids (Chen et al. 2004), simple
organic compounds that are easily detected. Other low molecular weight VOCs produced
during ripening of tomato and other fruit (e.g. strawberry, apple), and also present in foods
such as bread, cheese, wine and beer, include 2-methylbutanal, 3-methylbutanal, 3-
methylbutanol, phenylacetaldehde, 2-phenylethanol and methyl salicylate (Figure 1; Goff &
Klee 2006; Buttery & Ling 1993).
Changes in VOC Signatures in Stressed Plants Detection of VOCs is of increasing interest as a means of identifying plants that that have
been infected by disease-causing organisms (Sankaran et al., 2010; Jansen et al., 2011).
Most research related to disease detection through VOCs has been undertaken at the
laboratory scale (Hunter, 2002), and routine measurements of VOCs in air have shown that
average concentrations are very much smaller than those used in experiments designed to
study the effects of VOCs on plants (Maffei, 2010). Studies, of plant-plant communication
using VOCs have also suggested that natural VOC signalling occurs mainly over short
distances (Heil & Adame-Alvarez, 2010). However, it has been demonstrated that the
detection of damage associated VOCs is feasible at the glasshouse scale with 60 plants
grown at a floor area of 42 m2 (Jansen et al., 2009a).

- 8 -
Figure 1. Structures of (a) 2-methylbutanal, (b) 3-methylbutanal, (c) 3-methylbutanol, (d)
phenylacetaldehde, (e) 2-phenylethanol and (f) methyl salicylate.
The profile of VOCs produced by plants varies between tissues, and between stages in the
life cycle of the emitting organism (e.g. Bufler et al. 1990). Abiotic factors , including
temperature, lack or excess of soil moisture, lack or excess of light, lack of oxygen, air
pollution, nutrient deficiencies, mineral toxicities, soil acidity or alkalinity, and
agrochemicals can strongly regulate the emission of VOCs through modulating the
environment around plants (Sharkey and Loreto, 1993; Zhang et al. 1999; Peñuelas & Llusia,
2001; Gouinguene & Turlings, 2002; Wildt et al., 2003; Graus et al., 2004: Ebel et al. 2006;
Copolovici & Niinemets, 2010; Maffei, 2010; Arimura et al. 2004). There are natural diurnal
variations in production, for example (Niinemets et al. 2004; Graus et al. 2004); terpenoid
volatile emissions from Sitka spruce (Picea sitchensis) saplings on treatment with MeJA
showed a diurnal rhythm pattern (Miller et al. 2004). Atmospheric pollution or excessive
temperatures or drought can also modulate emissions (Loreto & Velikova 2001; Peñuelas &
Llusià 2002).
a
c
b
d
f e

- 9 -
VOCs can also be affected by the interaction of multiple biotic and abiotic factors
(Dicke et al. 2009; Maffei, 2010). For example, both fungal infection and ozone treatment of
tomato plants were found to induce similar VOCs (Jansen et al. 2009b), and plants under
simultaneous compatible bacterial and pest attack can produce volatiles in quantities
greater than those produced by plants subject to herbivory alone (Cardoza & Tumlinson,
2006). In contrast, in maize, the emission of herbivore-induced VOCs was reduced by
approximately 50% when plants were infected with a necrotrophic fungal pathogen (Rostas
et al., 2006). VOC emissions also vary according to plant age, genotype and over the course
of infection (Zhang et al., 2008; 2009; Holopainen & Gershenzon, 2010). Disease detection is
therefore likely to be contingent on the identification of a characteristic blend of stress-
associated VOCs, and of compounds that are characteristic of specific plant diseases.
Much work on changes to plant VOCs induced by biotic and abiotic stresses has been
carried out using model plant species. Both quantitative and qualitative changes in the
production of VOCs by infected plants have been reported. However, there are relatively
few reports of VOCs that are exclusively associated with the onset of disease, or with
specific plant diseases. More commonly, a volatile known to be produced in a variety of
contexts may be found to be distinctively present in diseased individuals in a specific plant
species. Specific examples include:
The production of phenylacetaldehyde O-methyloxime by grapefruit leaves
infected with the citrus canker pathogen Xanthomonas axonopodis pv. citri, but
not by leaves infected with the related pathogen X. axonopodis pv. citrumelo
(Zhang & Hartung 2005).
The emission by powdery mildew-infected tomato foliage of (Z)-3-heptanol, 1,6-
anhydro--D-glucopyranose, and 1-fluorododecane, but not by healthy or
herbivore damaged leaves (Laothawornkitkul et al. 2008). Moreover, using
electronic nose technology, it was possible to distinguish between healthy
tomato plants and those infected with powdery mildew (Oidium lycopersicum)
or infested by spider mites (Ghaffari et al. 2010).
The specific detection of 1-pentanol and ethyl boronate produced by mangoes
inoculated with Lasiodiplodia theobromae, whereas thujol was observed only in

- 10 -
Colletotrichum gloeosporioides inoculated mangoes (Moalemiyon et al. 2006;
Moalemiyan et al. 2007)
Similar technology was used to determine differences in VOCs produced by
healthy and diseased fruits of bell pepper (Kim et al. 2007).
More complex interactions involving the production of VOCs have also been noted. Attacks
on tobacco by foliar herbivores changed the emission patterns of cis-3-hexen-1-ol, linalool
and cis--bergamotene (Kessler & Baldwin 2001). These changes attracted parasitoids and
predators of the herbivores, lead to increased predation and lowered herbivore damage to
the host plants. Interestingly, cis-3-hexen-1-ol is available commercially, to add the flavor of
‘new mown hay’ to beer. Moreover, cis--bergamotene is produced in seed and as a flower
scent by a range of plant species, and acts as a kairomone (attractant) for certain types of
insect (Rocca et al. 1992; Nilsson 2009).
Infection by the bacterial pathogen Pseudomonas syringae elicited emission of the
terpenoid (E,E)-4,8,12-trimethyl-1,3,7,11-tridecatetraene in Arabidopsis (Attaran et al.
2008); this same compound occurs as a floral scent component in a variety of plants, but is
produced at negligible levels by healthy, undamaged Arabidopsis plants. It should also be
noted that the quantity of VOCs produced generally varies over the course of an infection.
For example, the quantities of induced VOCs were found to be higher at 7 days than 3 days
post-infection and greater when cereal plants were infected with Fusarium on leaves rather
than through soil (Piesik et al. 2011).
With the possible exception of fruit trees and species in the Pinaceae, woody plants
have received less attention than herbaceous species in terms of both constitutive and
induced VOCs. From the work that is published, however, it is clear that a broad range of
inducible VOCs are produced by woody plants, including alkenes, alkanes, carboxylic acids,
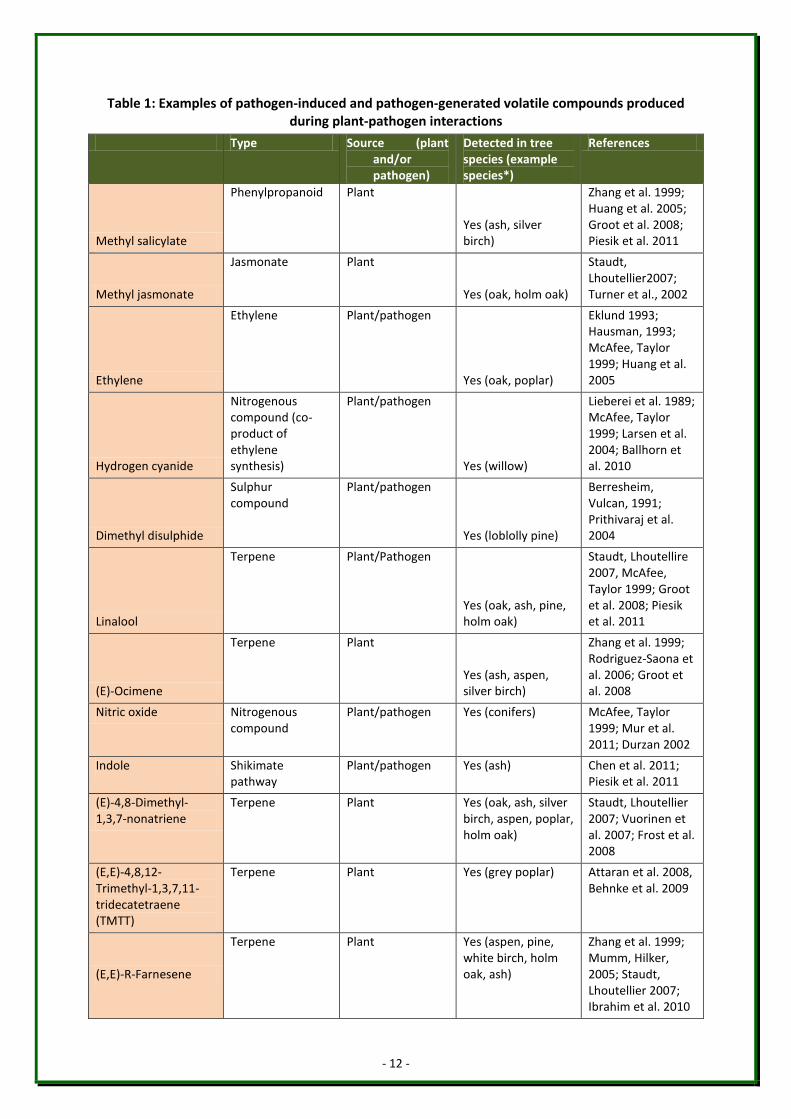
nitrogen-containing compounds and alcohols (Table 1). Major compounds tend to be
isoprene, terpenes and green leaf volatiles (GLVs) (Holopainen & Gershenzon, 2010). Some
of the most commonly detected VOCs produced by diseased plants include (Z)-3-hexenol,
methyl salicylate, (E)-β-ocimene, linalool, (E)-β-farnesene and (E)-4,8-dimethylnona-1,3,7-

- 11 -
triene (DMNT) (Jansen et al. 2011), all of which have been shown to be produced by trees
such as oak, ash, pine and silver birch (Table 1).
The production of terpenoids in the resin of Pinaceae is under strong genetic control
and terpene profiles have been widely used in chemotaxonomic studies to characterize
conifer species, provenances, clones and hybrids (Baradat & Yazdani 1988, Hannover 1992,
Adams et al. 1993, Lang 1994, Baradat et al. 1996, Tognetti et al. 2000). Much work carried
out on resin volatiles produced by the Pinaceae, however, has demonstrated changes in
volatile profiles in damaged or pest/pathogen attacked plants, compared to those in
undamaged plants (e.g. Lewinsohn et al. 1991; Michelozzi et al. 1995; Woodward et al.
2007), and that these changes are specific to the type of damage (wounding or wounding &
inoculation). These results indicate that changes in monoterpene profiles in the resins may
be suitable markers for detecting damage in this family of trees.
Recent work has also demonstrated the detection of infection of Eucalyptus globulus
by the fungal pathogen Teratosphaeria (Mycosphaerella) nubilosa cause of Mycosphaerella
leaf disease (Hantao et al. 2013). Concentrations of over 40 VOCs differed between infected
and uninfected trees. Application of advanced statistical techniques (partial least squares
discriminant analysis) differentiated infected and uninfected trees with an accuracy of
100%.
Infection of willow by rust significantly reduced isoprene emissions, but increased
emission of (Z)-β-ocimene, sesquiterpenes and lipoxygenase (LOX) dependent volatiles
(Toome et al. 2010).
VOCs produced by Citrus are also indicative of infection. For example, Cevallos-
Cevallos et al. (2009) used volatile derivatives of hesperidin, naringenin, and quercetin to
diagnose huanglongbing (citrus vein phloem degeneration) diseases caused by phytoplasma
in orchard-grown citrus trees. The ability to detect diseases caused by these phloem-limited
phytoplasma would be an enormous benefit, not only in terms of the import/export trade,
but also in commercial flows within countries.
Some of the best studied examples of changes in the production of VOCs by plants
are the terpene responses of pines to attack by bark beetles (e.g. Seybold et al. 2006).
Coleoptera are attracted to suitable host trees by the monoterpene profiles, often
associated with declining health in the host (Seybold et al. 2006). The bark beetles utilize the
- 12 -
Table 1: Examples of pathogen-induced and pathogen-generated volatile compounds produced during plant-pathogen interactions
Type Source (plant and/or pathogen)
Detected in tree species (example species*)
References
Methyl salicylate
Phenylpropanoid Plant
Yes (ash, silver birch)
Zhang et al. 1999; Huang et al. 2005; Groot et al. 2008; Piesik et al. 2011
Methyl jasmonate
Jasmonate Plant
Yes (oak, holm oak)
Staudt, Lhoutellier2007; Turner et al., 2002
Ethylene
Ethylene Plant/pathogen
Yes (oak, poplar)
Eklund 1993; Hausman, 1993; McAfee, Taylor 1999; Huang et al. 2005
Hydrogen cyanide
Nitrogenous compound (co-product of ethylene synthesis)
Plant/pathogen
Yes (willow)
Lieberei et al. 1989; McAfee, Taylor 1999; Larsen et al. 2004; Ballhorn et al. 2010
Dimethyl disulphide
Sulphur compound
Plant/pathogen
Yes (loblolly pine)
Berresheim, Vulcan, 1991; Prithivaraj et al. 2004
Linalool
Terpene Plant/Pathogen
Yes (oak, ash, pine, holm oak)
Staudt, Lhoutellire 2007, McAfee, Taylor 1999; Groot et al. 2008; Piesik et al. 2011
(E)-Ocimene
Terpene Plant
Yes (ash, aspen, silver birch)
Zhang et al. 1999; Rodriguez-Saona et al. 2006; Groot et al. 2008
Nitric oxide Nitrogenous compound
Plant/pathogen Yes (conifers) McAfee, Taylor 1999; Mur et al. 2011; Durzan 2002
Indole Shikimate pathway
Plant/pathogen Yes (ash) Chen et al. 2011; Piesik et al. 2011
(E)-4,8-Dimethyl-1,3,7-nonatriene
Terpene Plant Yes (oak, ash, silver birch, aspen, poplar, holm oak)
Staudt, Lhoutellier 2007; Vuorinen et al. 2007; Frost et al. 2008
(E,E)-4,8,12-Trimethyl-1,3,7,11-tridecatetraene (TMTT)
Terpene Plant Yes (grey poplar) Attaran et al. 2008, Behnke et al. 2009
(E,E)-R-Farnesene
Terpene Plant Yes (aspen, pine, white birch, holm oak, ash)
Zhang et al. 1999; Mumm, Hilker, 2005; Staudt, Lhoutellier 2007; Ibrahim et al. 2010

- 13 -
Limonene
Terpene Plant Yes (pine, spruce, holm oak)
Hanover 1992; Barnola et al. 1997; Staudt, Lhoutellire 2007
β-Caryophyllene
Terpene Plant Yes (pine, silver birch, white birch, holm oak)
Barnola et al. 1997, Staudt, Lhoutellier 2007; Mayer et al. 2008ab; Piesik et al. 2011
(Z)-3-Hexen-1-ol
GLV Plant Yes (silver birch, white birch, aspen, ash)
Zhang et al. 1999; Rodriguez-Saona et al. 2006; Piesik et al. 2011
(Z)-3-Hexenal
GLV Plant Yes (silver birch, white birch, aspen, ash)
Croft et al. 1993; Zhang et al. 1999; Prost et al. 2005; Rodriguez-Saona et al. 2006; Chen et al. 2011; Piesik et al. 2011
(Z)-3-Hexenyl acetate
GLV Plant Yes (silver birch, white birch, aspen, poplar, ash)
Zhang et al. 1999; Rodriguez-Saona et al. 2006; Frost et al. 2008; Piesik et al. 2011
(Z)-Ocimene Terpene Plant Yes (ash, aspen,
poplar) Frost et al. 2008; Piesik et al. 2011
(-)-β-Pinene Terpene Plant Yes (elm, pine, spruce, holm oak)
Hanover, 1992; McLeod et al. 2005; Staudt, Lhoutellier 2007; Piesik et al. 2011
(-)--Cubebene Terpene Plant Yes (elm, silver birch)
Zhang et al. 1999; McLeod et al. 2005
(+)--Cadinene Terpene Plant Yes (elm, holm oak) McLeod et al. 2005; Staudt, Lhoutellier 2007
R-Caryophyllene Terpene Plant Yes (oak, ash, poplar, white birch, pine. Holm oak)
Zhang et al. 1999; Staudt, Lhoutellier 2007; Kopke et al. 2008; Frost et al. 2008; Piesik et al. 2011
R-Humulene Terpene Plant Yes (oak, pine, holm oak)
Staudt, Lhoutellier 2007; Kopke et al. 2008
Germacrene-D Terpene Plant Yes (oak, poplar, holm oak)
Arimura et al. 2004; Staudt, Lhoutellier 2007; Frost et al. 2008
(E)-Nerolidol Terpene Plant/Pathogen Yes (oak, ash, holm oak)
McAfee, Taylor 1999; Staudt, Lhoutellier 2007

- 14 -
Terpinolene Terpene Plant Yes (pine, spruce) Hanover, 1992
Sabinene
Terpene Plant
Yes (pine, red cedar, spruce, holm oak)
Hanover, 1992; Zhang et al. 1999; Staudt, Lhoutellier 2007; Foster et al. 2013
Benzyl acetate Shikimate pathway
Plant Yes (laurel) Piesik et al. 2011; Politeo et al. 2007
*Please note that this column lists examples of tree species in which specific volatile chemicals have been detected. This list is not exhaustive: the absence of a species does not necessarily indicate that a specific chemical is not produced by this species.
monoterpenes to attract further beetles to the tree, most commonly through oxygenation
of specific monoterpenes produced by the host, although in certain cases the beetles may
produce the oxygenated monoterpenes by de novo synthesis (Seybold & Tittiger 2003). This
kairomone (attractant) effect is well known in several species of Dendroctonus and Ips, for
example (Wood 1970; Silverstein 1977; Seybold et al. 2006). The fact that the oxygenated
monoterpenes are produced specifically by the insect pests gives a ready target for
detection using high resolution chemical techniques.
Changes in plant host VOCs following attack by herbivorous insects are known in
several other host-insect systems (Pare & Tumlinson 1999; Kessler & Baldwin 2001;
Gouinguene et al. 2003; Zhang & Wang 2007; Staudt & Lhoutellier 2007). In an experimental
system examining responses of silver birch (Betula pendula) to infestation with larvae of
Epirrita autmunata and the leaf spot fungal pathogen Marssonina betulae, it was shown
that changes in host-produced VOCs differed substantially between plants damaged by the
insect and by the fungus (Vuorinen et al. 2007). Production of both methyl salicylate and (E)-
4,8-dimethyl-1,3,7-nonatriene was stimulated by insect attack, but not by the pathogen,
whereas infection by M. betulae caused significantly increased concentrations of (Z)-
ocimene and (E)-β-ocimene. Clearly, the system demonstrates differential responses in birch
which could be used to determine the possible cause(s) of alterations in VOC production. In
holm oak (Quercus ilex), attack by gypsy moth (Lymantria dispar) caterpillars also lead to a
significant increase in VOC release from the foliage of attacked trees (Staudt & Lhoutellier
2007).
Experimental systems have already been developed to enable detection of certain
insect pests in glasshouse crops. For example, Miresmailli et al. (2010) showed that

- 15 -
caterpillars of the cabbage looper moth, Trichoplusia ni, could be detected in a tomato crop
using GC-MS analyses of VOCs produced from affected plants. A zNose™ system was also
calibrate to detect (Z)-3-hexenyl acetate, (E)-β-ocimene and ϐ-caryophyllene, three VOCs
associated with the insect, and not with tomato plants. These methods were useable in the
glasshouse, demonstrating that, with further development, such systems may be utilized to
detect infestations early in the development of the pest. Different levels of infestation by
one of the most damaging rice pests, the brown plant hopper (Nilaparvata lugens), could be
detected on rice plants grown in a glasshouse using an E-nose system calibrated to sense 10
compounds in the VOCs from the affected plants (Zou & Wang 2011), although the system
relied on glasshouse grown plants enclosed in chambers in order to sample the headspace
volatiles.
Changes in VOC production following damage by phytophagous insects is a well-
known phenomenon, occurring in many, if not all, insect herbivore-plant host interactions
(Pare & Tumlinson 1999; Arimura et al. 2005), and presents an excellent opportunity for the
detection of pest problems inside plants and plant products during shipment. A further
intriguing possibility here is the use of excised insect sensor organs (usually antennae) to
detect tiny quantities of kairomones/pheromones released by insects attacking plants or
plant products (Sevonkaev & Katz 2008). Antennae are attached to electronic amplifiers to
determine changes in signal potentials (Cooper & Cass 2004). As insect antennae are
naturally calibrated to detect highly specific chemical cues in tiny quantities, the method
could be used in the detection of known problematic insect pests.
Volatile Organic Compounds from Microorganisms Bacteria: The production of VOCs by prokaryotes is a well-known phenomenon, which may
be involved naturally in cell-to-cell signaling and in the inhibition of potential competitive
microorganisms (Schultz et al. 2010). The range of VOCs known from bacteria is extensive,
and includes many small molecular weight compounds, some of which are identical to those
produced by plants. Soil-borne Pseudomonas spp., for example, produce compounds
including benzothiazole, cyclohexanol, nonanal, decanal, and dimethyl trisulphide (Fernando
et al. 2005; Sauer & Kliem 2010), each of which is relatively easily detected using routinely
available chemical methods (see below). Dimethyl sulphide and other related compounds

- 16 -
are further easily detected compounds, produced by bacteria in many substrates, including
cheese (Seefeldt & Weimer 2000).
Although many bacterial VOCs appear to be produced by a wide range of
actinobacteria, myxobacteria and cyanobacteria, other compounds are strain specific
(Schultz & Dickschat 2007) and could prove useful in diagnosing the presence of bacterial
pathogens in plants.
Fungi: The fungi also produce many volatile compounds (Splivallo et al., 2011; Kramer and
Abraham, 2012) in a number of chemical classes including monoterpenes, sesquiterpenes,
alcohols, aldehydes, aromatic compounds, esters, furans, hydrocarbons, ketones, and
nitrogen- and sulphur-containing compounds (Splivallo et al., 2007a; Campos et al., 2010;
Kramer and Abraham, 2012). As with bacteria, it is obvious in mycology laboratories or in
damp buildings that fungi produce characteristic odours. The odour of the common
mushroom, Agaricus bisporus, results partly from the production of 1-octen-3ol, for
example (Wąsowicz 1974). Recently, it was proposed that a combination of morphological,
molecular and metabolite data be used in the identification and classification of fungi
(Frisvad 2011), and some published work confirms the utility of this approach (Zhang et al.
2010).
A great deal of research carried out to date has focused on the volatiles produced by the
genera:
Penicillium, which includes many spoilage organisms, plant pathogens, and species
such as P. roqueforti, P. camemberti and P. chrysogenum, of use in the food and
pharmaceutical industries (Whilborg et al. 2008) and P. expansum, P. italicum and
P. digitatum attacking fruits (Chitzandis 1988a,b; Archer, 1988). It is pertinent to
note here that VOCs produced by some strains of P. expansum may have
exploitable repellent activity against the pine weevil, Hylobius abietis (Azeem et al.
2013).
Aspergillus: again, spoilage organisms that, in this genus, produce aflatoxins which,
being carcinogenic and mutagenic, are regulated in human foodstuffs. Aspergillus
spp., particularly A. flavus and A. parasticus, commonly contaminate food crops,
such as maize, peanuts, true nuts, dried fruits and vegetables. A range of VOCs is
produced by Aspergillus spp. (Kaminski et al. 1972; Zeringue et al. 1993; Roze et al.

- 17 -
2007). Recent work suggests that VOCs from Aspergillus play a major role in the
production of the toxins (Roze et al. 2007). Hence, the production of the toxins can
be accurately estimated through analyses of VOCs.
Fusarium: targeted to enable rapid determination of mycotoxins or mycotoxigenic
fungi in stored food, particularly cereals (Magan & Evans, 2000; Demyttenaere et
al. 2004; Zhang et al. 2007). The availability of electronic detection methods
(‘electronic noses’; ‘e-noses’) enables rapid detection of toxins from fungi such as
Fusarium in grain stores, reducing public health risks (Schnürer et al. 1999; Magan
& Evans, 2000). There are many Fusarium species that are pathogenic on growing
plants, however, including the highly damaging alien invasive species, F. circinatum,
cause of pitch canker on pines.
Trichoderma, a genus of very common fungi which appear to be ubiquitous. Several
species in this genus are exploited for the control of a range of plant diseases,
particularly those caused by soil-borne fungi (Elad et al. 1982; Papavizas 1985).
There are several mechanisms proposed for the inhibition of other fungi by
Trichoderma spp., including production of lytic enzymes, competition for infection
sites on roots or for nutrients and induction of host resistance (Benitez et al. 2004).
VOCs, however, are also long-known components in the arsenal used by
Trichoderma to protect the occupied niche (Rathore et al. 1992; Pandey &
Uapadhay 1997; De Respinis et al. 2010; Patil et al. 2012).
Tuber: the truffles. The drive in this research has been the development of
portable, field-useable instrumentation to detect truffles in forests and to
determine the presence of mixed truffle species in goods sold as a single species
(Bellesia et al. 1996; 2001; Diaz et al. 2002). Considerable success has been
achieved in this application, suggesting that the approach could be modified for use
with other fungi.
There is, however, an increasing body of work published on VOCs from other fungal genera.
Interesting recent work of direct relevance to the present review is the development
of VOC methods to determine fungi growing in water-damaged buildings (Polizzi et al.
2012). The method could be used to rapidly discriminate between strains of Aspergillus,
Chaetomium and Epicoccum isolated from inside damp buildings. The fungi were first
cultured in the laboratory, before analysis. Discrimination based on VOCs profiles was

- 18 -
obtained through Although it was recognised by the authors that low concentrations of the
VOCs would occur in the buildings (cf. Pasenen et al. 1998; Kim, Elfman et al. 2007;
Schleibinger et al. 2002, 2008), and that different VOCs may be produced by the same
fungus at different points in the life cycle (Matysik et al. 2008), it was concluded that VOCs
analyses could be used to complement classical and molecular identification techniques.
Discrimination between various fungal groups was also achieved in recent work
using mycorrhizal, pathogenic and saprotrophic species (Műller et al. 2013); VOCs produced
by the three groups were analyzed using GC-MS and differences determined using principal
component analysis (PCA). In this type of work, PCA uses data on the occurrence of all
compounds detected to discriminate species or strains, and is an essential tool when single
species-specific compounds are not present.
Further work on three species of Penicillium, along with other true fungi and a
species of the Oomycota genus Saprolegnia, demonstrated that different groups of fungi
and fungus-like organisms could be distinguished based on VOCs analyses (Wihlborg et al.
2008). Distinguishing between different fungi using these methods strengthens the case for
extending research into the volatile compounds produced by plant pathogenic fungi, to
determine if the method discriminates between these organisms. VOCs can be highly
discriminatory in Penicillium: P. roqueforti is a species complex, yet the different biological
species within the complex can be distinguished by VOCs analysis (Karlshøj & Larsen 2005).
Aromas of wine may sometimes be affected by the presence of fungi such as Botrytis
cinerea or Penicillium expansum in the must obtained from pressed grapes. This problem
lead to the development of GC-MS methods to detect VOCs produced by the two fungi in
must (Morales-Valle et al. 2011). Geosmin, a VOC produced by P. expansum when
competing with B. cinerea (La Guerche et al. 2007), was easily detected in contaminated
must, giving a marker metabolite for the presence of both fungi in this particular substrate
(Morales-Valle et al. 2011).
It is known from work in various laboratories, that many species of fungi have
characteristic smells when in culture. Species in the genus Ceratocystis, for example,
produce fruity odours in culture (Christen et al. 1997); as C. platani and C. fagacearum, both
formerly placed in the ‘C. fimbriata group’, cause highly damaging tree diseases, the
discrimination of these species based on human sensing of volatiles suggests that the highly
- 19 -
sensitive instrumentation available for VOCs analysis may be able to detect the presence of
these pathogens in plants. Some unsubstantiated reports also suggest that it is possible to
distinguish between different species of Phytophthora based on the human sense of smell
too; unfortunately, there is little information in the literature on production of VOCs by
Phytophthora spp., with reports confined to aromas produced following infection of potato
by P. infestans (de Lacey Costello et al. 2001; Laothawornkitkul et al. 2010).
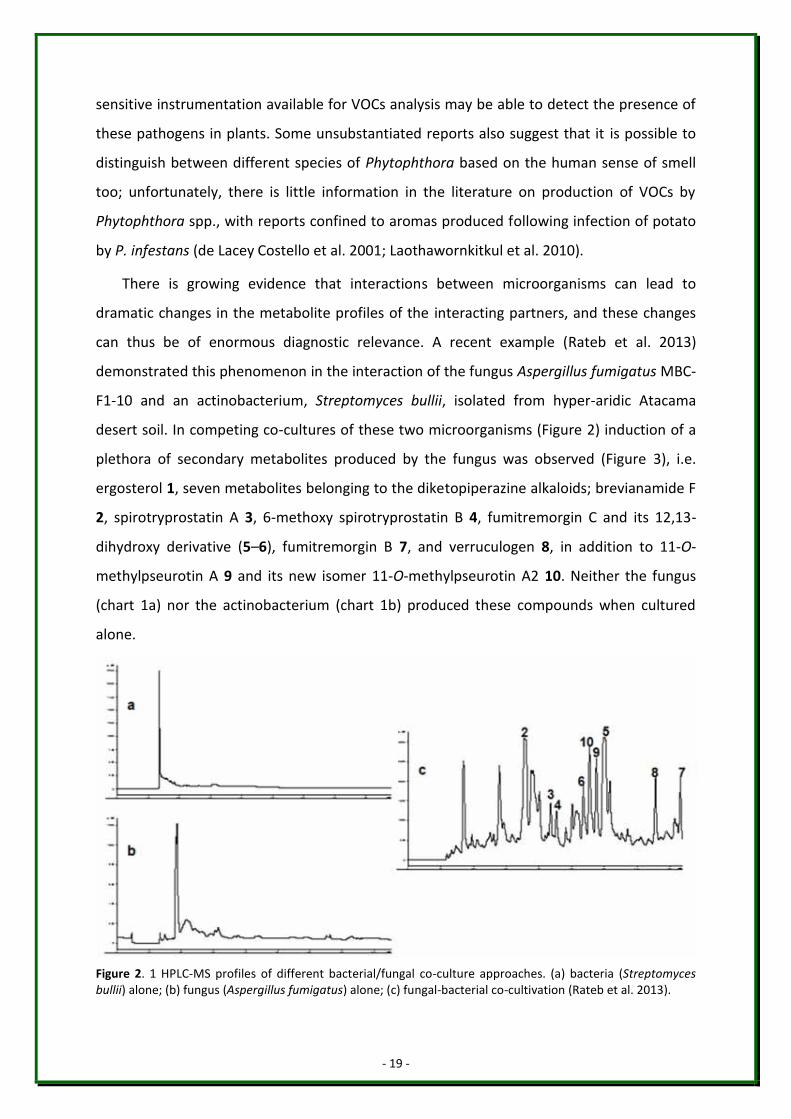
There is growing evidence that interactions between microorganisms can lead to
dramatic changes in the metabolite profiles of the interacting partners, and these changes
can thus be of enormous diagnostic relevance. A recent example (Rateb et al. 2013)
demonstrated this phenomenon in the interaction of the fungus Aspergillus fumigatus MBC-
F1-10 and an actinobacterium, Streptomyces bullii, isolated from hyper-aridic Atacama
desert soil. In competing co-cultures of these two microorganisms (Figure 2) induction of a
plethora of secondary metabolites produced by the fungus was observed (Figure 3), i.e.
ergosterol 1, seven metabolites belonging to the diketopiperazine alkaloids; brevianamide F
2, spirotryprostatin A 3, 6-methoxy spirotryprostatin B 4, fumitremorgin C and its 12,13-
dihydroxy derivative (5–6), fumitremorgin B 7, and verruculogen 8, in addition to 11-O-
methylpseurotin A 9 and its new isomer 11-O-methylpseurotin A2 10. Neither the fungus
(chart 1a) nor the actinobacterium (chart 1b) produced these compounds when cultured
alone.
Figure 2. 1 HPLC-MS profiles of different bacterial/fungal co-culture approaches. (a) bacteria (Streptomyces bullii) alone; (b) fungus (Aspergillus fumigatus) alone; (c) fungal-bacterial co-cultivation (Rateb et al. 2013).

- 20 -
The application of VOC profiling in disease detection Despite the possible negative factors associated with diagnosing problems in plants
discussed above, there are multiple reasons to believe that VOCs provide a valuable tool in
the development of advanced technologies for disease detection. One particularly strong
argument supporting this approach for distinguishing plant-pest/pathogen interactions from
other stressors comes from molecular studies of plant signal transduction during disease.
Three major defense signaling pathways have been recognized as important for plant biotic
interactions and for the production of defence associated VOCs: the jasmonic acid, salicylic
HO
HN HN
N
O
O
H
H
N
N
HN
O
O
H
H
O
MeO
N
N
HN
O
O H
O
MeO
NH
N
N
MeO
O
O
H
RR
5 R = H6 R = OH
NN
N
OHMeO
O
O H
OH
NN
N
O O
O
O H
OHOH
NHO
O
OHO
OMe R
OMe
O
172
3
5
6
8
9
21
11 10
15
16
1
2
3
4
7
8
9 R = -10R =-
MeO
Figure 3. Structure of fungal metabolites induced by competing co-culture between Aspergillus fumigatus MBC-F1-10 and Streptomyces bullii (Rateb et al. 2013).
acid, and ethylene pathways (Koornneef & Pieterse, 2008). Generally, salicylic acid–
dependent defences are activated more strongly in response to biotrophic pathogens and
jasmonic acid– and ethylene-dependent defences are activated to a higher extent in
response to necrotrophic pathogens and herbivorous insects. There is extensive crosstalk
between these different signal transduction pathways, leading to antagonistic and

- 21 -
synergistic interactions that may be diagnostic for specific stressors, or the outcome of
plant-pathogen interactions. For example, VOC profiles are differentially induced by
compatible and incompatible bacterial infection and pest damage when pathogens or pests
are applied alone or in combination to the same host (Cardoza & Tumlinson, 2006). VOCs
may also have direct antimicrobial activity against certain pathogens (Prost et al. 2005;
Wright et al. 2000), which may have imposed further selection for pathogen modulation and
metabolism of plant VOCs and VOC precursors, which may be diagnostic of infection (Voigt
et al. 2005).
A second argument comes from the proposal that plant VOCs can be combined with
pathogen VOCs to generate a ‘signature’ profile characteristic of bacterial or fungal
infections of plants. Although there can be significant overlap between plant and microbial
VOCs, microbial VOCs (MVOCs) often represent distinctive components in VOC profiles.
Plant pathogenic fungi, oomycetes and bacteria have been found to produce a variety of
VOCs, including cyanide, ethylene, dimethyl disulfide, nitric oxide, ethanol, 2,3-butanediol,
2-butanone, 2-methyl propanoic acid, 3-methyl butanoate, 2-methyl butanoate, 2-hydroxy-
2-butanone, 3-methyl-butanol, 1,5-dimethylpyrazine and benzaldehyde (McAfee and Taylor,
1999; Effantin et al. 2011; Spinelli et al. 2012), while Demyttenaere et al. (2004) showed
that it was possible to distinguish toxigenic Fusarium species on the basis of their volatile
profiles. Micro-organisms may also transform naturally occurring plant compounds into
compounds that are not released by healthy plants. For example, a P450 present in the
plant-associated bacterium Pseudomonas putida transforms cis- and trans-nerolidol to the
9-hydroxy product (Bell et al. 2013). Some metabolic pathways associated with the
production of pathogen volatiles, such as the 3-hydroxy-2-butanone pathway of
Pectobacterium carotovorum, are known to play an important role in pathogenesis
(Marquez-Villavicencio et al. 2011). A potential complicating factor in the use of MVOCs to
detect infection is the presence of non-pathogenic fungal and bacterial endophytes, which
can be abundant sources of VOCs (Zhi-Lin et al. 2012), and which may also act alongside
pathogens to modulate plant VOC profiles (Jallow et al. 2008; Lopez-Raez et al. 2010).
Thirdly, there is increasing evidence that specific VOC combinations are produced as
a means of communication between plants and other organisms in their environment, and
that natural selection has promoted the evolution of distinctive VOC profiles that signal

- 22 -
herbivore damage or pathogen attack both within plants and between plants and other
organisms. In this context, detection of plant VOCs serves as a means of ‘eavesdropping’ on
naturally occurring VOC signaling mechanisms (Baldwin et al. 2006; Heil & Bueno 2007;
Dicke & Baldwin 2010; Hiltpold et al. 2011). For example, predators (natural enemies) of
herbivores can be attracted to plants by the specific VOCs associated by herbivore damaged
plants, responding to subtle, but consistent differences in VOC composition (Kessler &
Baldwin 2001; Erb et al., 2010), and pathogens may themselves generate VOCs or modulate
plant VOCs to attract insect vectors (McLeod et al. 2005; Srinivasan et al., 2006; Medina-
Ortega et al., 2009; Mauck et al., 2010; Jansen et al. 2011; Mann et al. 2012). Multivariate
statistical methods, artificial neural networks and discriminant analysis models have been
used successfully to detect differences in VOCs characteristic of specific plant genotypes or
stressors (Tikunov et al., 2005; Vikram et al., 2006; Markom et al., 2009; Jansen et al., 2011),
and some progress has been made in the automation of detection methods (Jansen et al.,
2010).
Finally, it is useful that plants gain inherent benefits from the production of VOCs as
mixtures, rather than individual compounds, which can be exploited in identifying profiles
linked to plant disease. At the molecular level, the prevalence of VOC mixtures may be a
consequence of the properties of the biosynthetic pathways that produce them. However,
at the organismal level, the production of mixtures may be considered a direct way to
enhance VOC function. Where VOC s are used as infochemicals, or released as a
consequence of plant signal transduction mechanisms, the release of mixtures may result in,
or reflect the transmission of messages with greater specificity and higher information
content. In a defensive context, a diverse combination of chemical defenses may help
achieve simultaneous protection against numerous predators, parasites and competitors,
enable synergistic interactions between components, and impede the ability of enemies to
evolve resistance (Gershenzon & Dudereva, 2007).
Methods for detecting volatile chemicals Chemical methods are available, based on instrumentation already in use by the military
and at civilian air- and sea ports, that can detect specified volatile or ionized compounds
including narcotics, toxins or dangerous explosives, in tiny quantities, sometimes less than

- 23 -
one part per billion (<nanogram level). These techniques are widely deployed for security
measures at ports of entry into countries for people and goods; they are also being
developed to detect medically important pathogens, certain types of diseases (e.g. Garner
et al. 2007; van Veen et al. 2010; Qin et al. 2010) and the use of narcotics (e.g. Miekisch et
al. 2008). The routinely-used machines are based on several technologies, with different
methods appropriate for different types of substances or situations:
1. Ion mobility spectrometry (IMS), which utilizes ionization of target molecules, e.g. by
static charge, and identification of the ionized products following separation in an
electric field. The method is sensitive to parts per billion concentrations. Both table-
top and hand-held IMS instruments are available.
2. Gas chromatography / mass spectrometry (GC-MS), a highly sensitive technique, in
which suitable samples are injected into a coated column, the coating of which retains
compounds based on polarity, volatility and other attributes, leading to separation of
the molecules. The pure compounds are then passed directly into a mass
spectrometer, which enables identification based on total mass and characteristic
fragmentation patterns. Computer databases can then give most likely identities for
the molecules present in the sample.
3. Fourier-Transformed Infra Red Spectroscopy (FTIR). Chemical compounds have
characteristic spectra in the infra red region. FTIR enables the detection of compounds
in mixtures through the presence of certain signature peaks in these spectra.
The main company in the UK manufacturing instruments based on the methods listed
above, along with other types of detection and surveillance instrumentation, is Smiths
Detection plc, with head offices and laboratory facilities in Watford, England. This company
is currently moving away from large and expensive hardware to simpler, portable
instruments. These smaller instruments will be usable under ‘field’ conditions, and, being
simple to use and robust, should be ideal for the applications proposed here.
Smiths Detection is involved in many areas of security and surveillance using
advanced detection technologies. These include X-ray imaging, millimetre wave imaging,
Raman spectroscopy and the detection of biological and chemical warfare agents (both
explosives and toxins) using ion mobility spectroscopy (IMS) and gas chromatography / mass
spectrometry (GC–MS). Smiths Detection is also involved with instrumentation for the

- 24 -
detection of illicit drugs and contraband material using a range of technologies. Smiths
Detection has a long track record of providing high quality instruments for both UK and
foreign governments.
There are several key needs for field based instrumentation:
it needs to handle impure, possibly contaminated samples or the target mixed with
common chemicals.
It should be robust for ‘real world’ operation.
It should be simple to operate by non expert staff; and
Inexpensive to run and maintain.
A challenge for laboratory based sensors is the requirement to respond to the target
molecules amongst a highly diverse background of confounding material, including common
chemicals such as cleaning materials, pollen, benign organisms and other particulate matter
such as diesel elements and dust.
The VOCs described above and listed in Table 1 could be detected and identified by
GC-MS or possibly FTIR (Fourier transformed infra red spectroscopy) although much
depends on the location of the plant material and the sampling method used. For example a
container full of plant material, under inspection after a few days in transit would
accumulate VOCs, thereby assisting in the detection process. Also some of the Smiths
equipment uses a solid phase micro extraction (SPME) cartridge, to which the molecules are
absorbed, which can be exposed to the atmosphere under test for some time, or even
included in standardized packaging for plants and plant products in transit. Simple heating
of the probe when inside the analytical equipment releases the concentrated sample for
testing and identification. Lower concentration molecules (for example in a non-sealed
container) may require longer absorbtion times (a few minutes), but could still be analysed
rapidly.
Current sensitivities for the Smiths GC-MS are around the parts per million level, well
within range of the published data; some cannabinoids, which are plant-dervied
compounds, are detectable at the parts per billion level. For this particular project, it is still
undecided as to which molecules are of interest and more importantly the detection levels
required. Equally important would be some indication of the biological background involved

- 25 -
and what influence this has on the final analysis. It may be that some simple sample
preparation work is required to remove interfering material and concentrate the target
molecules before identification. The Smiths instruments are designed to look for ‘the needle
in a haystack’ scenario whereby the rare molecules of interest are in a much smaller
concentration than the common components of the background. Normally, the difficulty
comes when the surrounding biological background is very similar to the target and
selectivity becomes crucial. It remains to be seen as to the targets for this project, but the
advantage of a range of compounds, if that is the final concept, is that a consensus of
indicative results is all that is needed rather than the one ‘magic bullet’ target approach.
For the most accurate work the Smiths Guardion GC –MS would be the most suitable
instrument that is currently available, since the mixture of VOCs would first be separated by
the gas chromatography column and then identified using the mass spectrometry capability.
This device could test 10-15 samples per hour and is battery powered and field portable.
Orthogonal testing using, for example, FTIR or IMS, could assist in the overall analysis and
provide more data for confirmation. Each of these devices is simple to use by non-experts
(after some short training) and can retain the analytical information for further inspection
by scientific staff at a later date.
From a preliminary assessment of the requirements for the detection of potential
pests and pathogens in plant consignments, a range of indicative molecules would be the
most useful route to follow, but this approach needs some careful data handling. It could be
most beneficial to use a matrix of target molecules from the various pest organisms and
plant species, and some Bayesian analyses to determine the status of the shipment under
discussion. These initial screening tests would then indicate which consignments need
further, possibly manual inspection to finally decide the outcome. Analysis of the volatile
components using GC-MS or other approaches would serve to identify ‘healthy’ or
acceptable shipments, freeing the valuable human experts for more time on the more
questionable or indeterminate consignments.

- 26 -
Statistical and economic aspects of improved biosecurity In order to determine the overall utility of including a VOCs methods for detecting potential
invasive pests and pathogens in trade, both the statistical approach to sampling and the
economic gains (or losses) resulting from such inclusion must be assessed.
Overall concept: The routes through which pests or pathogens might arrive can be modelled
as a list, each member of which has a rate of establishment per import and an associated
volume of trade. The nature of the risk depends on the origin and type of the trade or
import. The overall rate of invasion associated with a route will depend on the product of
the intrinsic riskiness of the import and the volume. So, for example, the risks of containers
of bare-root Poinsettia from Germany via Portugal arriving in Southampton differ from
cuttings returned in the luggage of adventurous tourists to Nepal arriving in Heathrow,
which in turn differ from air-freight containers of cut roses from Kenya arriving into
Sleaford. The total rate of establishment of alien pests and diseases is then the sum of the
individual routes. This will be lower than the rate of arrival, but not necessarily much lower.
The aim of border biosecurity is to reduce the rate of arrival to the point where affordable
action can reduce the rate of establishment to some level subjectively deemed acceptable.
Current UK biosecurity depends on European Union policy. As a fundamental principle of
the EU is to facilitate trade among the partners, internal barriers to movement of living
plants and the soils they are grown in are based on the presumption that impeding trade
can only be justified in cases where a clear and defined threat will be reduced by the barrier.
Allocation of effort and required effort: Horticultural trade in and out of the UK has
increased hugely in the last 30 years. Average transport times have also decreased very
substantially. Passenger flight growth and accompanying baggage has also grown hugely.
Suppose that each passenger journey and each imported plant carries a (tiny) risk of
bringing in a pathogen not yet established in the UK. These risks will vary, of course, but we
could in principle weight sources and form an average risk ψ, the probability that a single
border movement will lead to establishment of a novel invasive disease. Now suppose
there are n such movements per year, and perfectly effective quarantine measures are
applied to a proportion θ of these movements. Quite how these hypothetical measures
capable of detecting previously unknown pathogens relate to actual measures which are
non-invasive and seek to ensure minimal delay to shipments need not be spelt out, but we

- 27 -
can use the percentage of shipments currently examined as an extreme upper limit. The
average rate of establishment of novel diseases is therefore per year.
Assuming establishments are close to independent, the realised number of establishments
in a given time will follow a Poisson distribution:
(1)
and
In aggregate, n has increased by a large factor (agricultural trade as a whole has
increased roughly 10 fold) over the last 40 years. Using the subscripts 2 and 1 to denote
current and past figures, we can ask how much improved quarantine would have to be to
keep the average number of invasions constant:
so
(2)
and therefore
(3)
Suppose that in 1970, 80% of cargoes were inspected, and transit times, pre-
containerisation and air freight, must have been greater. Taking
and inserting in equation (3) we see that θ2 needs to be 0.98 in order to keep the rate of
invasions the same. That is, 98% of ten times as many cargoes need to be inspected. It
would be of interest to see how the PHSI budget has evolved in real terms over the last 50
years.
The calculation above takes no account of the relative risk from different shipments.
Unless high risk shipments have become proportionately less common, the general thread
of the argument above applies, but it is no longer possible to express a single required
improvement.

- 28 -
To see this, write:
(4)
The probability of one or more establishments is the complement of this. The condition for
a constant rate of ingress, S, is
(5)
in which we allow for the possibility that the inspection rate for trades differs.
Some increased insight into the problem can be obtained by examining (5) under
simplifying assumptions. If we wished to know how far to intensify inspection so as to keep
a constant rate of incursion all inspection rates would need to be increased in accord with
equation (3); the basic insight that the uninspected number (not proportion) of trades has
to remain constant is not changed. However, the possibility of optimising allocation of
effort so as to minimise ingress subject to a given total spend does arise.
There is unlikely to be a unique minimum to the right-hand side of (5) constrained by
a fixed total cost (which would be a function of the θi2), but it may be possible to obtain
management guidance by looking at the effect of altering a particular θi2 . As an example,
suppose a monitoring device is developed which greatly increases the inspection efficiency
of a high-volume, low-risk trade, but the cost has to be recovered from elsewhere in the
system. It is unlikely that a formal model would be the best way to proceed, but a list-
based, systematic detailing might well improve decision-making about deployment of
resources.
An alternative way of framing the management problem is to ask where increments
of effort should be placed as they become available, without altering what is already in
place. We are trying to minimise S, and the partial derivatives with respect to inspection
efficiency are:

- 29 -
(6)
So it will always be best to allocate a little extra effort to the trades with the highest
volume-weighted risk – even if some other trade is unregulated. This is not intuitively a
good idea, but does suggest that the best policy may not be easy to specify and more
detailed modelling might be useful; more generally, it suggests that this area of research
could yield genuinely new understanding.
Optimal sampling therefore requires a reasonably detailed model of the entire
system of pathways of entry into the UK, a specification of what we are trying to achieve,
and information about the relative costs and efficiency of inspection of different pathways
of entry. The dual aim of minimising interference with individual freedom and trade, while
minimising establishment of exotic pathogens also requires consideration of the methods
used to detect pathogens.
Realistic sampling and operator-receiver characteristics: Any inspection method will
actually be imperfect; it will miss some infected consignments – for reasons including
procedural error, intrinsic imperfection in the detection method due to latency and other
inevitable technical problems, and lack of capacity – and will wrongly condemn some
healthy consignments through procedural error, faulty identification and so on. This results
in a table of probabilities, which can also be associated with (not necessarily monetary)
costs.
Inspection result Actual state
Infected healthy Totals
Positive (ie stated infected) Pr{+|+} Pr{+|-} Intercepted proportion Pr{+}
Negative (ie stated healthy) Pr{-|+} Pr{-|-}
Totals i h
In evaluating any novel detection method, this table is key. In any plausible case h >> i. The
large costs of an incursion or eradication are associated with Pr{-|+}. Because h >> i, the
false positive proportion of intercepts is likely to be high. This is minimised by instituting a
series of further checks, including visiting the importer after internal transit. But minimising
Pr{+|-} will increase Pr{-|+}. Reducing Pr{+|-} + Pr{-|+} as far as possible will increase

- 30 -
complexity and time, and therefore reduce θi . Any new technology improving detection
therefore needs Table 1 to be evaluated in the context not just of the immediate
equipment, but also the effect on follow-up and trace-forward operations.
Sampling strategy at the level of individual consignments: This is a question about how to
best use established (primarily visual and olefactory) and novel (auditory, expanded
spectrum visual and volatile detection) methods to detect diseased plants in a large
consignment in limited time. Methods depending on altered volatile emissions (whether
detected by animals or machinery) will have the best chance of positive detection if the
volatiles have been emitted into a closed space and thereby concentrated. Containers with
fan recirculation systems are ideal in this way. There will be conventional statistical issues
to do with the variation in concentration within a consignment and the variation in
concentration in similarly infected consignments. These need solution, but cannot be
further specified without the detail of the technology to be deployed. Similarly, there is
both an optimisation and a pathology question about how to use a positive result which
cannot immediately be linked to visible symptoms.
Measuring the effectiveness of interceptions: The most robust way to measure the risks run
would be to “spike” trades at their origin with differing proportions of plants inoculated
with known pathogens already established in the UK and therefore not posing additional
risk. This would allow formal comparisons of novel detection techniques with visual
inspections, and provide a means of evaluating problems caused by the need to move
packaging in order to see the hosts, latency, and other factors. Again the cost of this
approach would make a formal statistical approach important.
Economic justifications for additional detection technologies IAS are estimated to cost the EU at least €12 billion per annum, based on data from the past
20 years; as increasing numbers of IAS enter and establish in Europe, the costs are
increasing (Kettunen et al. 2009). Impacts in the UK of all invasive species were estimated by
Williams et al. (2010) to cost approximately £1.7 billion per annum. The financial impact of
invasive alien pests and pathogens to forestry are difficult to quantify in absolute terms
(Price 2010), but it was predicted that the annual costs to the forest environment of a
restricted number of tree diseases alone (Dothistroma septosporum on Pinus nigra var.

- 31 -
laricio, Phytophthora ramorum, acute oak decline and a ‘nasty surprise on Sitka spruce’ [sic])
could be in the region of £ 172 million (Price 2010). Despite the apparently dramatic figures
for losses and other costs following establishment of an invasive pest or pathogen, it is
essential that any additional biosecurity systems put in place at borders or elsewhere are
adequately predicted, to be certain that there is an economic benefit in their
implementation.
There are a number of publications in which the costs and benefits of biosecurity protocols
at national and state borders are modelled (e.g. Hinchey and Fisher 1991; Cook et al. 2007;
Cook and Matheson 2008; Sonigo et al. 2011).
A model for the development of a system to determine the reductions in economic losses
resulting from preventing the entry of a given invasive and damaging pest or pathogens is
given in Figure 4, using the agent causing pine pitch canker, Fusarium circinatum, as an
example, and based on Cook and Matheson (2008). The model could be adapted for a range
of pests and pathogens, using appropriate data.
In order to develop an understanding of the economic viability of any proposed changes to
phytosanitary procedures, the model below (Figure 4), prepared based on the threat posed
by Fusarium circinatum, cause of pitch canker of pines, could be followed. The sequence of
events to be followed is:
1. Run the model (Figure 4) to obtain probability weighted losses from damage (pest or
infection) under the status-quo scenario (current standard government response to
spread, existing probability of entry).
2. Modify the probability of entry to reflect the changes from the introduction and
adoption of the new detection technologies.
3. Re-run the model. This repetition should show a reduction in the probability
weighted losses incurred by the forestry or horticulture sector.
4. The reduction in probability weighted losses will also entail development and
adoption costs associated with the new detection technologies. Clearly, for an
efficiency gain, the reduction in losses must be greater than the costs of
development and adoption of the novel technology.
A sensitivity analysis using different discount rates is also required, as the choice of discount
rate may heavily influence the probability weighted losses: higher discount rates will

- 32 -
diminish the benefits of reducing future spread much more rapidly compared to lower
discount rates. It is also pertinent to consider the non-market benefits of forests and
landscapes, particularly when the affected plant species at risk are highly visible to the
general public, i.e. outside of commercial timber plantations.
A highly relevant recent publication in which the economic impacts of an invasive tree
pathogen are considered in detail is that of Cook and Matheson (2008), where the economic
implications of the establishment of F. circinatum in the pine plantations of Australia are
modelled. It was suggested that large economic benefits would accrue from specifically
targeting the likelihood of pathogen arrival and the speed of spread:
“Using a hypothetical example to illustrate this point, let us assume the probability
of arrival can be lowered by 25% as a result of $0.25 million per year being
invested in pre-border and border security measures to avoid F. circinatum
incursions over 30 y. If we consider our distribution of entry and establishment
probabilities, Uniform (0.035, 0.3), and think in terms of the median (0.13), this
change would be equivalent to decreasing the likelihood of an arrival event
occurring once every 7.5 y to once every 10 y. If this were achieved we predict a
benefit/cost ratio of 1.7:1 would result with about $13 million prevented damages
over time” (Cook and Matheson 2008).
Clearly, preventing the ingress of alien pests and pathogens into a given state (or even at
the continental level) will have immense economic advantages, supporting the introduction
of further, highly sensitive and accurate detection methods into the plant and plant
products trade routes.
How Can These Methods be Applied in Practice? Two approaches can be used during the technological development phase:
1. Detection of volatile metabolites of bacterial and fungal pathogens, and of pest
invertebrates characteristic of the organisms (to family/genus level, or species where
possible);

- 33 -
Figure 4. Model for a system to determine reductions in economic losses resulting from reducing the
probability of entry of a given invasive and damaging pest or pathogens. The example used here is Fusarium
circinatum.
2. Detection of characteristic changes in the volatile components produced by affected
plants following attack by pests and/or pathogens, including transformation of plant
metabolites by pest and pathogen activities.
It may also be possible to utilize a combination of these two approaches to detect the
presence of threatening organisms. As plants are shipped both inter and intra-continentally
in containers, sometimes heavily wrapped and/or sealed, concentrations of volatiles will be
higher in transit compared with ambient conditions.
Calibration of the currently available instruments enables the detectors to respond
to the target molecules amongst a very diverse background of confounding material,
including common chemicals such as cleaning materials, pollen, benign organisms and other
particulate matter such as diesel elements and dust. The literature, coupled with the

- 34 -
experience of the chemists and engineers at Smiths Detection, strongly suggest that the
approach is feasible for the detection of organisms producing defined VOCs.
The data already published strongly suggest that a range of indicative and potentially
diagnostic molecules are produced by different micro-organisms and insect pests. The
information generated in the development phase, however, will require careful handling
and analysis. It may be most effective to target a matrix of molecules from some species
along with Bayesian analyses to determine what potential problems are being transported
in the shipment examined. These initial screening tests could then indicate which
consignments need further, possibly visual inspection and molecular analysis, as suggested
above. Analysis of the volatile components using GC-MS, IMS or other approaches would
serve to identify ‘healthy’ or acceptable shipments, freeing up the time of the phytosanitary
inspectors to examine the more questionable consignments.
Conclusions The further development of chemical analytical instrumentation to detect pests and
pathogens in plant consignments requires an innovative, forward-thinking and
interdisciplinary approach, utilizing the expertise of pathologists, entomologist, chemists,
engineers, IT specialists and end-users. In addition, economic aspects of taking this
approach rather than one focused solely on molecular methods must be considered.
It is evident that the currently available detection instrumentation, following
appropriate parameterization, could be used in the detection of VOCs produced by
microorganisms or by infected plants. The feasibility for this approach requires in depth
investigation, including determination of the costs and benefits of applying these
technologies over the current systems based on visual assessments and, when indicated,
molecular techniques. It is not envisaged that VOC detection techniques would replace the
current methods, but they will add a further technique to the arsenal utilized by
phytosanitary officials when dealing with large numbers of plants and plant product
consignments at ports of entry into the EU and the UK. Moreover, a positive result in
chemical detection will indicate to the current inspectors where more detailed
investigations could be made for example, taking tissues samples for molecular analyses.

- 35 -
The instrumentation to detect VOCs produced by pests and pathogens, or by
attacked plants, is already available. Moreover, the machinery is highly sensitive to a wide
range of small molecular weight compounds; currently, the instrumentation is calibrated for
the detection of several compounds that are derived from plants, and are chemically related
(through metabolic pathways) to known VOCs released by both plants and microorganisms.
Hence, with suitable parameterization based on further knowledge of VOCs produced in
plant-pest and plant-pathogen interactions, it will be possible within a small time scale, to
use these machines in the detection of potentially invasive pests and pathogens of plants.
It could be argued that systems relying on the detection of VOCs in plant and plant
product consignments would not be of use in diagnosing the presence of soil-borne pests
and pathogens, but emerging research indicates that soil sampling for VOCs is also highly
likely to be feasible in the near future (Insam & Seewald 2010). Such adaptations of the VOC
detection technologies could prove invaluable in detecting the presence of pathogens such
as the Oomycota (Phytophthora, Pythium) and true fungi, including many species of
Fusarium or Rhizoctonia, or root-feeding insects, such as vine weevils, cut worms and wire
worms.
As there are likely to be large numbers of currently unknown invasive agents
threatening forest ecosystems, it is important that detection methods recognize signature
metabolites of groups of pests or pathogens (e.g. by genus or family), rather than
discriminating to the species level. This requirement does not preclude using the technique
to detect to the specific level, of course. The IMS and/or GC/MS or GasID instruments will
provide, initially, a technique to focus sampling for other diagnostic methods, such as highly
discriminatory molecular tools.
With such large numbers of plants imported into European countries, it will remain
true that sampling will be imperfect; the potential for increased detection rates using state-
of-the-art chemical methods, however, will greatly enhance the efficacy of phytosanitary
control at borders.
References Adams, R.P., T. Demeke and H.A. Abulfatith. (1993). RAPD DNA fingerprints and terpenoids:
clues to past migrations of Juniperus in Arabia and east Africa. Theoretical and Applied Genetics 87:22-26.

- 36 -
Archer, S.A. (1988). Penicillium expansum Link. pp. 253-255 in: Smith, I.M., Dunez, J., Lelliott, R.A., Phillips, D.H., Archer, S.A. (eds). European handbook of Plant Diseases. Blackwell Scientific Publications, Oxford, London.
Arimura, G., Huber, D.P.W., Bohlmann, J. (2004). Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides): cDNA cloning, functional characterization, and patterns of gene expression of (–)-germacrene D synthase, PtdTPS1. Plant Journal 37:603–616.
Arimura, G., Kost, C., Boland, W. (2005). Herbivore-induced, indirect plant defences. Biochimica et Biophysica Acta 1734:91–111.
Attaran, E., Rostás, M., Zeier, J. (2008). Pseudomonas syringae elicits emission of the terpenoid (E,E)-4,8,12-trimethyl-1,3,7,11-tridecatetraene in Arabidopsis leaves via jasmonate signaling and expression of the terpene synthase TPS4. Molecular Plant-Microbe Interactions 21: 1482-1497.
Azeem, M., Rajarao, G.K., Nordenhem, H., Nordlander, G., Borg-Karlson, A.K. (2013). Volatiles reduce pine weevil attraction to host plants. Journal of Chemical Ecology 39:120-128.
Baldwin, I.T., Halitschke, R., Paschold, A., von Dahl, C.C., Preston, C.A. (2006). Volatile
signaling in plant-plant interactions: "talking trees" in the genomics era. Science
311:812-815.
Ballhorn, D.J., Pietrowski, A., Lieberei, R. (2010). Direct trade-off between cyanogenesis and
resistance to a fungal pathogen in lima bean (Phaseolus lunatus L.). Journal of Ecology
98:226-236.
Baradat, Ph., Yazdani, R. (1988). Genetic expression for monoterpenes in clones of Pinus sylvestris grown on different sites. Scandinavian Journal of Forest Research 3:25-36.
Baradat, Ph., Michelozzi, M., Tognetti, R., Khouja, M.L., Khaldi, A.. (1996). Geographical variation of Pinus halepensis Mill. Pp 141-158 in: Population Genetics and Genetic Conservation of Forest Trees. Baradat, Ph., Adams, W.T., Müller-Starck, G. (Eds). SPB Academic Publishing, Amsterdam, pp. 141-158.
Barnola, L.F., Cedeno, A., Hasegawa, M. (1997). Intraindividual variations of volatile terpene
contents in Pinus caribaea needles and its possible relationship to Atta laevigata
herbivory. Biochemical Systematics and Ecology 25:707–716.
Behnke K., Kleist, E., Uerlings, R., Wildt, J., Rennenberg, H., Schnitzler, J-P. (2009). RNAi-
mediated suppression of isoprene biosynthesis in hybrid poplar impacts ozone
tolerance. Tree Physiology 29: 725-736.
Bell, S.G., French, L., Rees, N.H., Cheng, S.S., Preston, G., Wong, L-L. (2013). A phthalate
family oxygenase reductase supports terpene alcohol oxidation by CYP238A1 from
Pseudomonas putida KT2440. Biotechnology and Applied Biochemistry 60: 9-17.
Bellesia, F.; Pinetti, A.; Bianchi, A.; Tirillini, B. (1996). Volatile compounds of the white truffle (Tuber magnatum Pico) from Middle Italy. Flavour and Fragrance Journal 11:239-243.
Bellesia, F.; Pinetti, A.; Tirillini, B.; Bianchi, A. (2001). Temperature dependent evolution of volatile organic compounds in Tuber borchii from Italy. Flavour and Fragrance Journal 16:1-6.

- 37 -
Benitez, T., Rincon, A.M., Limon, M.C., Codon, A.C. (2004). Biocontrol mechanisms of Trichoderma strains. International Microbiology 7: 249-260.
Berresheim, H., Vulcan, V.D. (1992). Vertical distributions of COS, CS2, DMS and other sulfur
compounds in a loblolly pine forest. Atmospheric Environment. Part A. General Topics
26:2031-2036.
Brasier, C.M. (2008). The biosecurity threat to the UK and global environment from nternational trade in plants. Plant Pathology 57:792-808.
Bufler, U., Seufert, G., Juttner, F. (1990). Monoterpene patterns of different tissues and plant parts of Norway spruce (Picea abies L. Karst.). Environmental Pollution 68:367-375.
Buttery, R.G., Ling, L. (1993). Volatile components of tomato fruit and plant parts. p. 23-34 in Bioactive Volatiles from Plants. Teranishi, R., Buttery, R.G., Sugisawa, H. (Eds). ACS Symposium Series 525, American Chemical Society, Washington, DC.
Campos, V.P., de Pinho, R.S.C., Freire, E.S. (2010). Volatiles produced by interacting microorganisms potentially useful for the control of plant pathogens. Ciencia e Agrotecnologia 34:525–535.
Cardoza, Y., Tumlinson, J. (2006). Compatible and incompatible Xanthomonas infections
differentially affect herbivore-induced volatile emission by pepper plants. Journal of
Chemical Ecology 32: 1755-1768.
Cevallos-Cevallos, J.M., Rouseff, R., Reyes-De-Corcuera, J.I. (2009). Untargeted metabolite
analysis of healthy and Huanglongbing-infected orange leaves by CE-DAD.
Electrophoresis 30: 1–8.
Chen, G., Walker, D., Taylor, A., Lin, Z., Grierson, D. (2004). Identification of a specific isoform of tomato lipoxygenase (TomloxC) involved in the generation of fatty acid-derivedflvor compounds. Plant Physiology 136, 2641-2651.
Chen, Y., Whitehill, J.A., Bonello, P., Poland, T.. (2011). Differential response in foliar
chemistry of three ash species to emerald ash borer adult feeding. Journal of Chemical
Ecology 37:29-39.
Chitzandis, A. (1988a). Penicillium digitatum Sacc. pp. 252-253 in: Smith, I.M., Dunez, J.,
Lelliott, R.A., Phillips, D.H., Archer, S.A. (eds). European handbook of Plant Diseases.
Blackwell Scientific Publications, Oxford, London.
Chitzandis, A. (1988b). Penicillium italicum Wehmer. pp. 253 in: Smith, I.M., Dunez, J.,
Lelliott, R.A., Phillips, D.H., Archer, S.A. (eds). European Handbook of Plant Diseases.
Blackwell Scientific Publications, Oxford, London.
Christen, P., Meza, J.C., Revah, S. (1997). Fruity aroma production in solid state fermentation by Ceratocystis fimbriata: influence of the substrate type and the presence of precursors. Mycological Research 101:911-919.
Cook, D.C., Thomas, M.B., Cunningham, S.A., Anderson, D.L., De Barro, P.J. (2007). Predicting the economic impact of an invasive species on an ecosystem service. Ecological Applications 17:1832-1840.
Cook, D.C., Matheson, A.C. (2008). An estimate of the potential economic impact of pine pitch canker in Australia. Australian Forestry 71: 107-112.

- 38 -
Cooper, J., Cass, J. (2004). Biosensors: A Practical Approach. 2nd edition. Oxford University Press, Oxford, London, New York.
Copolovici, L., Niinemets, U. (2010). Flooding induced emissions of volatile signalling compounds in three tree species with differing waterlogging tolerance. Plant Cell and Environment 33:1582–94.
Croft, K., F Juttner, F., Slusarenko, A.J. (1993). Volatile products of the lipoxygenase pathway
evolved from Phaseolus vulgaris (L.) leaves inoculated with Pseudomonas syringae pv
phaseolicola. Plant Physiology 101:13-24.
De Lacey Costello, B.P.., Evans, P., Ewen, R.J., Gunson, H.E., Jones, P.R.H., Ratcliffe, N.M., Spencer-Phillips, P.T.N. (2001). Gas chromatography-mass spectrometry analyses of volatile organic compounds from potato tubers inoculated with Phytophthora infestans or Fusarium coeruleum. Plant Pathology 50:489-496.
De Respinis, S.; Vogel, G.; Benagli, C,; Tonolla, M.; Petrini, O.; Samuels, G. J., 2010: MALDI-TOF MS of Trichoderma: a model system for the identification of microfungi. Mycological Progress 9:79-100.
Demyttenaere, J.C.R., Moriña, R.M., De Kimpe, N., Sandra, P. (2004). Use of headspace solid-phase microextraction and headspace sorptive extraction for the detection of the volatile metabolites produced by toxigenic Fusarium species. Journal of Chromatography A 1027:147–154.
Diaz, P., Senorans, F.J., Reglero, G., Ianez, E. (2002). Truffle aroma analysis by headspace solid phase microextraction. Journal of Agricultural Food Chemistry 50:6468-6472.
Dicke, M., van Loon, J.J.A., Soler, R. (2009). Chemical complexity of volatiles from plants
induced by multiple attack. Natural Chemistry and Biology 5: 317-324.
Dicke, M., Baldwin, I.T. (2010). The evolutionary context for herbivore-induced plant
volatiles: beyond the ‘cry for help’. Trends in Plant Science 15:167-175.
Dicke, M. & Loreto, F (2010). Induced plant volatiles: from genes to climate change. Trends
in Plant Sciences 15:115-117.
Dudareva, N., Klempien, A., Muhlemann, J.K., Kaplan, I. (2013). Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytologist doi: 10.1111/nph.12145.
Durzan, D.J. (2002). Stress-Induced Nitric Oxide and Adaptive Plasticity in Conifers. Journal
of Forest Science 48: 281–291.
Ebel, R.C., Mattheis, J.P., Buchanan, D.A. (2006). Drought stress of apple trees alters leaf emissions of volatile compounds. Physiologia Plantarum 93:709–12.
Effantin. G., Rivasseau, C., Gromova, M., Bligny, R., Hugouvieux-Cotte-Pattat, N. (2011).
Massive production of butanediol during plant infection by phytopathogenic bacteria of
the genera Dickeya and Pectobacterium. Molecular Microbiology 82: 988-997.
Eklund, L. (1993). Seasonal variations of O2, CO2, and ethylene in oak and maple stems.
Canadian Journal of Forest Research 23:2608-2610.
Elad, Y., Chet, I., Henis, Y. (1982). Degradation of plant pathogenic fungi by Trichoderma harzianum. Canadian Journal of Microbiology 28:719-725.

- 39 -
Erb, M., Foresti, N., Turlings, T. (2010). A tritrophic signal that attracts parasitoids to host-damaged plants withstands disruption by non-host herbivores. BMC Plant Biology 10:247.
Fernando, W.G.D., Ramaraathnam, R., Krishnamoorthy, A.S., Savachuk, S.C. (2005). Identification and use of potential bacterial organic volatiles in biocontrol. Soil Biology and Biochemistry 37:955-964.
Frisvad, J.C. (2011). Rationale for a polyphasic approach in the identification of mycotoxigenic fungi. In: De Saeger, S. (ed): Determining Mycotoxins and Mycotoxigenic Fungi in Food and Feed. Woodhead Publishing Ltd., Cambridge.
Frost, C.J., Mescher, M.C., Dervinis, C., Davis, J.M., Carlson, J.E., De Moraes, C.D. (2008). Priming defense genes and metabolites in hybrid poplar by the green leaf volatile cis-3-hexenyl acetate. New Phytologist 180:722-734.
Gadgil, P., Dick, M., Simpson, J., Bejakovich, D., Ross, M., Bain, J., Horgan, G. Wylie, R. (2003) Management Plan Response to an Incursion of Pine Pitch Canker in Australia or New Zealand. Commissioned and published by the Forest Health Committee on behalf of the Forestry and Forest Products Committee, Canberra.
Garner, C.E., Smith, S., de Lacy Costello, B., White, P., Spencer, R., Probert, C.S.J., Ratcliffe, N.M. (2007). Volatile organic compounds from feces and their potential for diagnosis of gastrointestinal disease. The FASEB Journal 21: 1675-1688.
Gershenzon, J., Croteau, R. (1990). Regulation of monoterpene biosynthesis in higher plants. pp. 99-160 in Towers, G.H.N. & Stafford, H.A. (eds). Biochemistry of the Mevalonic Acid Pathway of Terpenoids. Plenum Press, New York.
Gershenzon, J., Dudareva, N. (2007). The function of terpene natural products in the natural
world. Natural Chemistry and Biology 3: 408-414.
Ghaffari, R., Zhang, F., Iliescu, D., Hines, E., Leeson, M., Napier, R., Clarkson, J. (2010). Early detection of disease in tomato crops: an electronic nose and intelligent systems approach. pp. 1-6 in: Proceedings of the 2010 International Joint Conference on Neural Networks, Barcelona, Spain, 18-23 July 2010.
Goff, S.A. Klee, H.J. (2006). Plant volatile compounds: sensory cues for health and nutritional value? Science 311, 815 – 819.
Gouinguene, S.P., Turlings, T.C.J. (2002). The effects of abiotic factors on induced volatile
emissions in corn plants. Plant Physiology 129:1296–307.
Gouinguene, S., Alborn, H., Turlings, T.C.J. (2003). Induction of volatile emissions in maize by different larval instars of Spodoptera littoralis. Journal of Chemical Ecology 29:145–162.
Graus, M., Schnitzler, J.P., Hansel, A., Cojacariu, C., Rennenberg, H., et al. (2004). Transient
release of oxygenated volatile organic compounds during light-dark transitions in grey
poplar leaves. Plant Physiology 125:1967–75.
Groot, P., Grant, G., Poland, T., Scharbach, R., Buchan, L., Nott, R. et al. (2008).
Electrophysiological response and attraction of emerald ash borer to green leaf volatiles
(GLVs) emitted by host foliage. Journal of Chemical Ecology 34: 1170-1179.
Guenther, A. (1997). Seasonal and spatial variations in natural volatile organic compound
emissions. Ecological Applications 7:34–45.

- 40 -
Hannover, J.W. (1992). Applications of terpene analysis in forest genetics. New Forests 6: 159-178.
Hantao, L.W., Aleme, H.G., Passador, M.M., Furtado, E.L., de Lima Ribeiro, F.A., Poppi, R.J., Augusto, F. (2013). Determination of disease biomarkers in Eucalyptus by comprehensive two-dimensional gas chromatography and multivariate data analysis. Journal of Chromatography A 1279:86-91.
Hausman, J.F. (1993). Changes in peroxidase activity, auxin level and ethylene production
during root formation by poplar shoots raised in vitro. Plant Growth Regulation 13:263-
268.
Heil, M., Silva Bueno, J.C. (2007). Within-plant signaling by volatiles leads to induction and
priming of an indirect plant defense in nature. Proceedings of the National Academy of
Sciences USA 104:5467-5472.
Heil M, Adame-Álvarez RM (2010). Short signalling distances make plant communication a
soliloquy. Biology Letters 6: 843-845.
Hiltpold, I., Erb, M., Robert, C.A.M., Turlings, T.C.J. (2011). Systemic root signalling in a
belowground, volatile-mediated tritrophic interaction. Plant, Cell & Environment
34:1267-1275.
Hinchy, M.D. and Fisher, B.S. (1991) A Cost-Benefit Analysis of Quarantine. Technical Paper 913, Australian Bureau of Agricultural and Resource Economics, Canberra.
Holopainen, J.K., Gershenzon, J. (2010). Multiple stress factors and the emission of plant
VOCs. Trends in Plant Science 15:176-184.
Hunter,G.L.K., Brogden, W.B. (1965). Analysis of the terpene and sesquiterpene hydrocarbons in some citrus oils. Journal of Food Science 30:383-387.
Huang, J., Schmelz, E., Alborn, H., Engelberth, J., Tumlinson, J. (2005). Phytohormones
mediate volatile emissions during the interaction of compatible and incompatible
pathogens: the role of ethylene in Pseudomonas syringae infected tobacco. Journal of
Chemical Ecology 31: 439-459.
Hunter, M.D. (2002). A breath of fresh air: beyond laboratory studies of plant volatile–
natural enemy interactions. Agricultural and Forest Entomology 4:81-86.
Ibrahim, M.A., Mäenpää, M., Hassinen, V., Kontunen-Soppela, S., Malec, L., Rousi, M. et al
(2010). Elevation of night-time temperature increases terpenoid emissions from Betula
pendula and Populus tremula. Journal of Experimental Botany 61:1583-1595.
Insam, H., Seewald, M.S.A. (2010). Volatile organic compounds (VOCs) in soil. Biology and Fertility of Soils 46:199-213.
Jallow, M.A., Dugassa-Gobena, D., Vidal, S. (2008). Influence of an endophytic fungus on
host plant selection by a polyphagous moth via volatile spectrum changes. Arthropod-
Plant Interactions 2:53-62.
Jansen, R.M.C., Hofstee, J.W., Wildt, J., Verstappen, F.W.A., Bouwmeester, H.J., van Henten
E.J. (2009a). Induced plant volatiles allow sensitive monitoring of plant health status in
greenhouses. Plant Signaling Behaviour 4:824–29.

- 41 -
Jansen, R.M.C., Hofstee, J.W., Wildt, J., Verstappen, F.W.A., Bouwmeester, H.J., et al.
(2009b). Health monitoring of plants by their emitted volatiles: trichome damage and
cell membrane damage are detectable at greenhouse scale. Annals of Applied Biology
154:441–52.
Jansen, R.M.C., Hofstee, J.W., Bouwmeester, H.J., van Henten, E.J. (2010). Automated signal
processing applied to volatile-based inspection of greenhouse crops. Sensors 10:7122–
33.
Jansen, R.M.C., Wildt, J., Kappers, I.F., Bouwmeester, H.J. Hofstee, J.W., van Henten, E.J..
(2011). Detection of diseased plants by analysis of volatile organic compound emission.
Annual Reviews of Phytopathology 49:157-174.
Kaminski, E., Libbey, L.M., Stawicki, S., Wasowicz, E. (1972). Identification of the predominant volatile compounds produced by Aspergillus flavus. Applied Microbiology 24:721-726.
Karlshøj, K., Larsen, T.O. (2005). Differentiation of species from the Penicillium roqueforti group by volatile metabolite profiling. Journal of Agriculture and Food Chemistry 53:708-715.
Kesselmeier, J., Staudt, M. (1999). Biogenic volatile organic compounds (VOC): An overview on emission, physiology and ecology. Journal of Atmospheric Chemistry 33:23–88.
Kessler, A., Baldwin, I.T. (2001). Defensive function of herbivore-induced plant volatile emissions in nature. Science 291:2141-2144.
Kettunen, M., Genovesi, P., Gollasch, S., Pagad, S., Starfinger, U., Brink, P., Shine, C. (2009). Technical Support to EU Strategy on Invasive Alien Species (IAS) - Assessment of the Impacts of IAS in Europe and the EU, Institute for European Environmental Policy (IEEP), Brussels, Belgium.
Kim, I.K., Abd El-Aty, A.M., Shin, H.C., Lee, H.B., Kim, I.S., Shim, J.H. (2007). Analysis of volatile compounds in fresh health and diseased peppers (Capsicum annuum L.) using solvent-free solid injection coupled with gas chromatography-flame ionisation detector and confirmation with mass spectrometry. Journal of Pharmaceutical and Biomedical Analysis 45:487-494.
Kim, J.L., Elfman, L., Mi, Y., Wieslander, G., Smedje, G, Norbäck, D. (2007). Indoor molds, bacteria, microbial volatile organic compounds’ and plasticizers in schools – associations with asthma and respiratory symptoms in pupils. Indoor Air 17:153-163.
Knudsen, J.T., Eriksson, R., Gershenzon, J., Ståhl, B. (2006). Diversity and distribution of floral scent. The Botanical Review 72:1-120.
Koornneef, A., Pieterse, C.M.J. (2008). Cross talk in defense signaling. Plant Physiology 146:839–44.
Köpke, D., Schröder, R., Fischer, H., Gershenzon, J., Hilker, M., Schmidt, A. (2008). Does egg
deposition by herbivorous pine sawflies affect transcription of sesquiterpene synthases
in pine? Planta 228:427-438.
Kramer, R., Abraham, W.R. (2012). Volatile sesquiterpenes from fungi: what are they good for? Phytochemistry Reviews 11: 15–37.

- 42 -
La Guerche, S., De Senneville, L., Blancard, D., Darriet, P. (2007). Impact of the Botrytis cinerea strain and metabolism on (-)-geosmin production by Penicillium expansum in grape juice. Antoine Van Leeuwenhoek 92:331-341.
Lang, K.J. (1994). Abies alba Mill.: differentiation of provenances and provenance groups by the monoterpene patterns in the cortex resin of twigs. Biochemical and Systematic Ecology 22:53-63.
Laothawornkitkul, J., Moore, J.P., Taylor, J.E., Possell, M., Gibson, T.D., Hewitt, C.N., Paul,
N.D.. (2008). Discrimination of plant volatile signatures by an electronic nose: a
potential technology for plant pest and disease monitoring. Environmental Science and
Technology 42:8433-8439.
Laothawornkitkul, J., Jansen, R.M.C., Smid, H.M., Bouwmeester, H.J., Muller, J., van Bruggen, A.H.C. (2010). Volatile organic compounds as a diagnostic marker of late blight infected potato plants: A pilot study. Crop Protection 29: 872-878.
Larsen, M., Trapp, S., Pirandello, A. (2004). Removal of cyanide by woody plants.
Chemosphere 54:325-333.
Lewisohn, E., Gijzen, M., Croteau, R. (1991). Defence mechanisms of conifers: differences in constitutive and wound-induced monoterpene biosynthesis between species. Plant Physiology 96:44-49.
Lieberei, R., Biehl, B., Giesemann, A., Junqueira, N.T.V. (1989). Cyanogenesis Inhibits Active
Defense Reactions in Plants. Plant Physiology 90:33-36.
López-Ráez, J.A., Verhage, A., Fernández, I., García, J.M., Azcón-Aguilar, C., Flors, V., Pozo,
M.J. (2010). Hormonal and transcriptional profiles highlight common and differential
host responses to arbuscular mycorrhizal fungi and the regulation of the oxylipin
pathway. Journal of Experimental Botany 61:2589-2601.
Loreto, F., Velikova, V. (2001). Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiology 127:1781-1787.
McAfee, B.J., Taylor, A. (1999). A review of the volatile metabolites of fungi found on wood
substrates. Natural Toxins 7:283-303.
McLeod, G., Gries, R., von Reuß, S.H., Rahe, J.E., McIntosh, R., König, W.A. et al. (2005). The
pathogen causing Dutch elm disease makes host trees attract insect vectors.
Proceedings of the Royal Society B: Biological Sciences 272: 2499-2503.
Maffei, M.E. (2010). Sites of synthesis, biochemistry and functional role of plant volatiles.
South African Journal of Botany 76: 612-631.
Magan, N., Evans, P. (2000). Volatiles as an indicator of fungal activity and differentiation between species, and the potential use of electronic nose technology for early detection of grain spoilage. Journal of Stored Products Research 36:319-340.
Mann, R.S., Tiwari, S., Smoot, J.M., Rouseff, R.L. Stelinski, L.L. (2012). Repellency and toxicity of plant-based essential oils and their constituents against Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Journal of Applied Entomology 136:87–96

- 43 -
Markom, M.A., Shakaff, A.Y.M., Adom, A.H., Ahmad, M.N., Hidayat, W., Abdullah, A.H., Fikri,
N.A. (2009). Intelligent electronic nose system for basal stem rot disease detection.
Computers and Electronics in Agriculture 66:140-146.
Marquez-Villavicencio, Md.P., Weber, B., Witherell, R.A., Willis, D.K., Charkowski, A.O.
(2011). The 3-hydroxy-2-butanone pathway Is required for Pectobacterium carotovorum
pathogenesis. PLoS ONE 6: e22974.
Matysik, S., Herbarth, O., Mueller, A. (2008). Determination of volatile metabolites originating from mould growth on wall paper and synthetic media. Journal of Microbiological Methods 75:182-187.
Mauck, K.E., De Moraes, C.M., Mescher, M.C. (2010). Deceptive chemical signals induced by
a plant virus attract insect vectors to inferior hosts. Proceedings of the National
Academy of Sciences USA 107: 3600-3605.
Mayer, C.J., Vilcinskas, A., Gross, J. (2008a). Pathogen-induced release of plant allomone
manipulates vector insect behavior. Journal of Chemical Ecology 34:1518–1522.
Mayer, C.J., Vilcinskas, A., Gross, J. (2008b). Phytopathogen lures its insect vector by altering
host plant odor. Journal of Chemical Ecology 34:1045–1049.
Medina-Ortega, K.J., Bosque-Pérez, N.A., Ngumbi, E., Jiménez-Martínez, E.S., Eigenbrode, S.D. (2009). Rhopalosiphum padi (Hemiptera: Aphididae) responses to volatile cues from barley yellow dwarf virus-infected wheat. Environmental Entomology 38: 836-845.
Michelozzi, M., White, T.L., Squillace, A.E., Lowe, W.J. (1995). Monoterpene composition and fusiform rust resistance in slash and loblolly pines. Canadian Journal of Forest Research 25:193-197.
Miekisch, W., Fuchs, P., Kamysek, S., Neumann, C., Schubert, J.K. (2008). Assessment of propofol concentrations in human breath and blood by means of HS-SPME–GC–MS. Clinica Chimica Acta 395:32-37.
Miller, B., Madilao, L.L., Ralph, S., Bohlmann, J. (2005). Insect-induced conifer defense:
White pine weevil and methyl jasmonate induce traumatic resinosis, de novo formed
volatile emissions, and accumulation of terpenoid synthase and putative octadecanoid
pathway transcript in Sitka spruce. Plant Physiology 137: 369–382.
Miresmailli, S., Gries, R., Gries, G., Zamar, R.H., Isman, M.B. (2010). Herbivore-induced plant volatiles allow detection of Trichoplusia ni (Lepidoptera: Noctuidae) infestation on greenhouse tomato plants. Pest Management Science 66:916-924.
Moalemiyon, M., Vikram, A., Kushalappa, A.C. Yaylayan, V. (2006). Volatile metabolite profiling to detect and discriminate stem-end rot and anthracnose diseases of mango fruits. Plant Pathology 55:792-804.
Moalemiyan, M., Vikram, A., Kushalappa, A.C. (2007). Detection and discrimination of two fungal diseases of mango (cv. Keitt) fruits based on volatile metabolite profiling using GC/MS. Postharvest Biology and Technology 45:117-125.
Morales-Valle, H., Silva, L.C., Paterson, R.R.M., Venacio, A., Lima, N. (2011). Effects of the origin of Botrytis cinerea on earthy aromas from grape broth media further inoculated with Penicillium expansum. Food Microbiology 28:1048-1053.

- 44 -
Moshonas, M.G., Shaw, P.E. (1971). Analysis of volatile flavor constituents from grapefruit essence. Journal of Agricultural Food Chemistry 19:119-120.
Műller, A., Faubert, P., Hagen, M., zu Castell, W., Polle, A., Shnitzler, J-P., Rosenkrantz, M. (2013). Volatile profiles of fungi – Chemotyping of species and ecological functions. Fungal Genet. Biol. http://dx.doi.org/10.1016/j.fgb.2013.02.005
Mumm, R., Hilker, M. (2005) The significance of background odour for an egg parasitoid to
detect plants with host eggs. Chemical Senses 30:337–343.
Mur, L.A.J., Mandon, J., Cristescu, S.M., Harren, F.J.M., Prats, E. (2011). Methods of nitric
oxide detection in plants: A commentary. Plant Science 181: 509-519.
Niinemets, U., Loreto, F., Reichstein, M. (2004). Physiological and physicochemical controls on foliar volatile organic compound emissions. Trends in Plant Science 9:180.
Nijssen, L., Visscher, C., Maarse, H., Willemsens, L., Boelens, M. (1997). Volatile Organic Compounds in Food. TNO Nutrition and Food Research Institute, zeist, The Netherlands. Vol. 7, suppl. 1.
Nilsson, A. (2009). Identification and Syntheses of Semiochemicals Affecting Mnesampela privata and Trioza apicalis. Licentiate Thesis, Department of Natural Sciences, Engineering and Mathematics, Mid Sweden University, SE-851 70 Sundsvall, Sweden.
Pandey, K.K. and J.P. Uapadhyay. (1997). Selection of potential biocontrol agents based on
production of volatile and non volatile antibiotics. Vegetation Science 24:140- 143.
Papavizas, G.C. (1985). Trichoderma and Gliocladium: Biology, ecology, and potential for
biocontrol. Annual Reviews of Phytopathology 23:23-54.
Pare, P.W., Tumlinson, J.H. (1999). Plant volatiles as a defense against insect herbivores. Plant Physiology 121:325–331.
Pasanen A, Korpi A, Kasanen JP, Pasanen P, 1998. Critical aspects on the significance of microbial volatile metabolites as indoor air pollutants. Environment International 24: 703-712.
Patil, A., Laddha, A., Lunge, A., Paikrao, H., Mahure, S. (2012). In vitro antagonistic properties of selected Trichoderma species against tomato root rot-causing Pythium species. International Journal of Science, Environment and Technology 1: 302–315.
Peñuelas, J., Llusià, J. (2002). Linking photorespiration, monoterpenes and thermotolerance in Quercus. New Phytologist 155:227-237.
Piesik, D., Lemńczyk, G., Skoczek, A., Lamparski, R., Bocianowski, J., Kotwica, K., Delaney, K.J.
(2011). Fusarium infection in maize: Volatile induction of infected and neighboring
uninfected plants has the potential to attract a pest cereal leaf beetle, Oulema
melanopus. Journal of Plant Physiology 168:1534-1542.
Politeo, O., Jukić, M., Miloš, M. (2007). Chemical composition and antioxidant activity of free
volatile aglycones from laurel (Laurus nobilis L.) compared to its essential oil. Croatica
Chemica acta 80:121-126.
Polizzi, V., Adams, A., Malysheva, S.V., De Sager, S., Van Peteghem, C., Moretti, A., Picco, A.M., De Kimpe, N. (2012). Identification of volatile markers for indoor fungal growth and chemotaxonomic classification of Aspergillus species. Fungal Biology 116: 941–953.

- 45 -
Price, C. (2010). Tree disease – the economic impacts of it all. Quarterly Journal of Forestry, 104: 224-230.
Prithiviraj, B., Vikram, A., Kushalappa, A.C., Yaylayan, V. (2004). Volatile metabolite profiling for the discrimination of onion bulbs infected by Erwinia carotovora ssp. carotovora, Fusarium oxysporum and Botrytis allii. European Journal of Plant Pathology 110:371-377.
Prost, I., Dhondt, S., Rothe, G., Vicente, J., Rodriguez, J.M., Kift, N., Carbonne, F., Griffiths, G., Esquerre-Tugaye, M.T., Rosahi, S., Castresana, C., Humberg, M., Fournier, J. (2005). Evaluation of the antimicrobial activities of plant oxylipins supports their involvement in defense against pathogens. Plant Physiology 139: 1902–1913.
Qin, T., Liu, H., Song, Q., Song, G., Wang, H-Z., Pan, Y-Y., Xiong, F-X., Gu, K-S., Sun, G-P., Chen, Z-D. (2010). The screening of volatile markers for hepatocellular carcinoma. Cancer Epidemiology and Biomarkers 19:2247-2253.
Rateb, M.E., Halluburton, I., Houssen, W., Bull, A., Goodfellow, M., Santhanam, R., Jaspars, M., Ebel, R. (2013). Induction of diverse secondary metabolites in Aspergillus fumigatus by microbial co-culture. RSC Advances. Doi: 10.139/C3RA42378F
Rathore, V.R.S., Mathur, K., Hodha, B.C., Mathur, K. (1992). Activity of volatile and non-
volatile substances produced by Trichoderma viride on ginger rhizome rot pathogens. Indian Phytopathology 45, 253-254.
Rocca, J.R., Nation, J.L., Strekowski, L., Battiste, M.A. (1992) Comparison of volatiles emitted by male Caribbean and Mexican fruit flies. Journal of Chemical Ecology 18:223-244.
Rodriguez-Saona, C., Poland, T.M., Miller, J.R., Stelinski, L.L., Grant, G.G., de Groot, P.,
Buchan, L., MacDonald, L. (2006). Behavioral and electrophysiological responses of the
emerald ash borer, Agrilus planipennis, to induced volatiles of Manchurian ash, Fraxinus
mandshurica. Chemoecology 16:75-86.
Roze, L.V., Beaudry, R.M., Arthur, A.E., Calvo, A.M., Linz, J.E. (2007). Aspergillus volatiles regulate aflatoxin synthesis and asexual sporulation in Aspergillus parasiticus. Applied and Environmental Microbiology 73:7268-7276.
Sankaran, S., Mishra, A., Ehsani, R., Davis, C. (2010). A review of advanced techniques for
detecting plant diseases. Computers and Electronics in Agriculture 72:1-13.
Santini, A., Ghelardini, L., de Pace, C., Desprez-Loustau, M-L., Capretti, P., Chandelier, A., Cech, T., Chira, D., Diamandis, S., Gaitniekis, T., Hantula, J., Holdenrieder, O., Jankovsky, L., Jung, T., Jurc, D., Kirisits, T., Kunca, A., Lygis, V., Malecka, M., Marçais, B., Schmitz, S., Schumacher, J., Solheim, H., Solla, A., Szabò, I., Tsopelas, P., Vannini, A., Vettraino, A.M., Webber, J., Woodward, S., & Stenlid, J. (2013). Biogeographic patterns and determinants of invasion by alien forest pathogenic fungi in Europe. New Phytologist 197: 238–250.
Sauer, S., Kliem, M., 2010: Mass spectrometry tools for the classification and identification of bacteria. Nature Reviews of Microbiology 8: 74-82.
Schleibinger, H., Brattig, C., Mangler, M. (2002). Microbial volatile organic compounds (MVOC) as indicators for fungal damage. Indoor Air 4:707-712.
Schleibinger, H., Laussmann, D. Bornehag, C., Eis, D., Rueden, H. (2008). Microbial volatile organic compounds in the air of moldy and mold-free indoor environments. Indoor Air 18:113-124.

- 46 -
Schnürer, J., Olsson, J., Borjesson, T. (1999). Fungal volatiles as indicators of food and feeds spoilage. Fungal Genetics and Biology 27:209-217.
Schultz, S., Dickschat, J.S. (2007). Bacterial volatiles: the smell of small organisms. Natural Products Reports 24:814-842.
Schultz, S., Dickschat, J.S., Kunze, B., Wagner-Dobler, I., Diestel, R., Sasse, F. (2010). Biological activity of volatiles from marine and terrestrial bacteria. Marine Drugs 8:2976-2987.
Seefeldt, K.E., Weimer, B.C. (2000). Diversity of sulphur compound production in lactic acid bacteria. Journal of Dairy Science 83:2740-2746.
Sevonkaev, I.V., Katz, E. (2008).Biosensors based on immobilized insects fragments. Journal of Solid State Electrochemistry 12:7-14.
Seybold, S.J., Tittiger, C. (2003). Biochemical and molecular biology of de novo isoprenoid pheromone production in the Scolytidae. Annual Reviews of Entomology 48:425-453.
Seybold, S.J., Huber, D.P.W., Lee, J.C., Graves, A.D., Bohlmann, J. (2006). Pine monoterpenes and pine bark beetles: a marriage of convenience for defense and chemical communication. Phytochemistry Reviews 5:143-178.
Sharkey, T., Loreto, F. (1993). Water stress, temperature and light effect on the capacity for
isoprene emission and photosynthesis of kudzu leaves. Oecologia 95:328–33.
Silverstein, R.M. (1977). Complexity, diversity, and specificity of behavior-modifying chemicals: examples mainly from Coleoptera and Hymenoptera. pp. 231-251 in: Shorey, H.H. & McKelvey, J.J. (eds). Chemical Control of Insect Behavior: Theory and Application. John Wiley & Sons, New York.
Sonigo, P., Turbe, A., Berman, S., Reilly, Nyegaard Hvid, H. (2011). A comparative assessment of existing policies on ivasive species in the EU member states and in selected OECD countries. Report to European Commission DG Environment. 258 pp.
Spinelli, F., Cellini, A., Vanneste, J., Rodriguez-Estrada, M., Costa, G., Savioli, S. et al. (2012).
Emission of volatile compounds by Erwinia amylovora: biological activity in vitro and
possible exploitation for bacterial identification. Trees 26:141-152.
Splivallo, R., Bossi, S., Maffei, M., Bonfante, P. (2007). Discrimination of truffle fruiting body versus mycelial aromas by stir bar sorptive extraction. Phytochemistry 68: 2584–2598.
Splivallo, R., Ottonello, S., Mello, A., Karlovsky, P. (2011). Truffle volatiles: from chemical ecology to aroma biosynthesis. New Phytologist 189: 688–699.
Srinivasan, R., Alvarez, J.M., Eigenbrode, S.D., Bosque-Pérez, N.A. (2006) Influence of hairy
nightshade Solanum sarrachoides (Sendtner) and Potato leafroll virus
(Luteoviridae:Polerovirus) on the host preference of Myzus periscae (Sulzer)
(Homoptera: Aphididae). Environmental Entomology 35:546–553.
Staudt, M., Lhoutellier, L. (2007). Volatile organic compound emission from holm oak infested by gypsy moth larvae: evidence for distinct responses in damaged and undamaged leaves. Tree Physiology 27: 1433–1440.
Tikunov, Y., Lommen, A., de Vos, C.H.R., Verhoeven, H.A., Bino, R.J., Hall, R.D., Bovy, A.G.
(2005). A novel approach for nontargeted data analysis for metabolomics. Large-scale
profiling of tomato fruit volatiles. Plant Physiology 139:1125-1137.

- 47 -
Tognetti R., Michelozzi, M., Lauteri, M., Brugnoli, E., Giannini, R. (2000). Geographic variation in growth, carbon isotope discrimination, and monoterpene composition in Pinus pinaster Ait. provenances. Canadian Journal of Forest Research 30:1682-1690.
Toome, M., Randjarv, P., Copolovici, L., Niinemets, U., Heinsoo, K., et al. (2010). Leaf rust
induced volatile organic compounds signalling in willow during the infection. Planta
232:235–243.
Turner, J.G., Ellis, C., Devoto, A. (2002). The jasmonate signal pathway. Plant Cell 14:S153–
S164.
van Veen, S. Q.; Claas, E. C.; Kuijper, E. J., 2010: High-throughput identification of bacteria and yeast by Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry in conventional medical microbiology laboratories. Journal of Clinical Microbiology 48: 900-907.
Vikram, A., Lui, L.H., Hossain, A., Kushalappa, A.C.. (2006). Metabolic fingerprinting to discriminate diseases of stored carrots. Annals of Applied Biology 148:17-26.
Voigt, C.A., Schafer, W., Salomon, S. (2005). A secreted lipase of Fusarium graminearum is a virulence factor required for infection of cereals. Plant Journal 42: 364–375.
Vuorinen, T., Nerg, A.M., Syrjala, L., Peltonen, P., Holopainen, A.M. (2007). Epirrita autumnata induced VOC emission of silver birch differ from emission induced by leaf fungal pathogen. Arthropod-Plant Interactions 1:159–165.
Wąsowicz, E. (1974). Identification of the volatile flavor compounds in mushroom (Agaricus bisporus). Bulletin of the Academy of Polish Sciences, Series: Science and Biology 22:143-151.
Wihlborg, R., Pippitt, D., Marsili, R. (2008). Headspace sorptive extraction and GC-TOFMS for the identification of volatile fungal metabolites. Journal of Microbiological methods 75:244-250.
Wildt, J., Kobel, K., Schuh-Thomas, G., Heiden, A.C. (2003). Emissions of oxygenated volatile organic compounds from plants part II: emissions of saturated aldehydes. Journal of Atmospheric Chemistry 45:173–96.
Williams, F., Eschen, R., Harris, A., Djeddour, D., Pratt, C., Shaw, R.S., Varia, S., Lamontagne-Godwin, J., Thomas, S.E., Murphy, S.T. (2010). The economic cost of invasive non-native species on Great Britain. CABI, Wallingford, Egham. 199 pp.
Wright, M.S., Greene-McDowelle, D.M., Zeringue, H.J., Bhatnagar, D., Cleveland, T.E. (2000). Effects of volatile aldehydes from Aspergillus-resistant varieties of corn on Aspergillus parasiticus growth and aflatoxin biosynthesis. Toxicon 38: 1215–1223.
Wood, D.L. (1970). Pheromones of bark beetles. pp 301–316 in: Wood, D.L., Silverstein, R.M. & Nakajima, M. (eds): Control of Insect Behavior by Natural Products. Academic Press, New York.
Woodward, S., Bianchi, S., Bodles, W.J.A., Beckett, E., Michelozzi, M. (2007). Physical and chemical responses of Sitka spruce (Picea sitchensis) clones to colonization by Heterobasidion annosum as potential markers for relative host susceptibility. Tree Physiology 27:1701-1710.
Zeringue, H.J., Batnagar, D., Cleveland, T.E. (1993). C15H24 volatile compounds unique to aflatoxigenic strains of Aspergillus flavus. Applied and Environmental Microbiology 59:2264-2270.

- 48 -
Zhang, A., Hartung, J.S. (2005). Phenylacetaldehyde O-methyloxime: A volatile compound
produced by grapefruit leaves infected with the citrus canker pathogen, Xanthomonas
axonopodis pv. citri. Journal of Agricultural and Food Chemistry 53: 5134-5137.
Zhang, C.L., Wang, G.P., Mao, L.J., Komon-Zelazowska, M., Yuan, Z.L., Lin, F.C., Druzhinina, I.S., Kubicek, C.P. (2010). Muscadro fenguangensis sp. nov. from southeast China. Morphology, physiology and production of volatile compounds. Fungal Biology 114:797-808.
Zhang, H.M., Wang, J. (2007). Detection of age and insect damage incurred by wheat, with an electronic nose. Journal of Stored Products Research 43:489–495.
Zhang, H.M., Wang, J., Tian, X., Yu, H., Yu, Y. (2007). Optimization of sensor array and detection of stored duration of wheat by electronic nose. Journal of Food Engineering 82:403-408.
Zhang, P-Y., Chen, K-S., He, P-Q., Liu, S-H., Jiang, W-F. (2008). Effects of crop development
on the emission of volatiles in leaves of Lycopersicon esculentum and its inhibitory
activity to Botrytis cinerea and Fusarium oxysporum. Journal of Integrative Plant Biology
50: 84-91.
Zhang, P., Chen, K. (2009). Age-dependent variations of volatile emissions and inhibitory
activity toward Botrytis cinerea and Fusarium oxysporum in tomato leaves treated with
chitosan oligosaccharide. Journal of Plant Biology 52: 332-339.
Zhang, Q.-H., G. Birgersson, J Zhu, C Löfstedt, J Löfqvist, and F Schlyter. (1999). Leaf volatiles from nonhost deciduous trees: Variation by tree species, season and temperature, and electrophysiological activity in Ips typographus. Journal of Chemical Ecology 25:1923-1943.
Zhi-Lin, Y., C. Yi-Cun, X. Bai-Ge, and Z. Chu-Long. (2012). Current perspectives on the volatile-producing fungal endophytes. Critical Reviews in Biotechnology 32:363-373.
Zou, B., Wang, J. (2011). Use of electronic nose technology for identifying rice infestation by Nilaparvata lugens. Sensors and Actuators B 160:15-21.
Related Documents











