ISSN: 1524-4571 Copyright © 2007 American Heart Association. All rights reserved. Print ISSN: 0009-7330. Online TX 72514 Circulation Research is published by the American Heart Association. 7272 Greenville Avenue, Dallas, DOI: 10.1161/01.RES.0000258877.57836.d2 published online Jan 25, 2007; Circ. Res. Catherine C. Hedrick David T. Bolick, Angela M. Whetzel, Marcus Skaflen, Tracy Deem, Jianyi Lee and Monocyte/Endothelial Interactions in Aorta Absence of the G Protein-Coupled Receptor G2A In Mice Promotes http://circres.ahajournals.org/cgi/content/full/01.RES.0000258877.57836.d2/DC1 Data Supplement (unedited) at: http://circres.ahajournals.org located on the World Wide Web at: The online version of this article, along with updated information and services, is http://www.lww.com/reprints Reprints: Information about reprints can be found online at [email protected] 410-528-8550. E-mail: Fax: Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters http://circres.ahajournals.org/subscriptions/ Subscriptions: Information about subscribing to Circulation Research is online at by on May 18, 2011 circres.ahajournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ISSN: 1524-4571 Copyright © 2007 American Heart Association. All rights reserved. Print ISSN: 0009-7330. Online
TX 72514Circulation Research is published by the American Heart Association. 7272 Greenville Avenue, Dallas,
DOI: 10.1161/01.RES.0000258877.57836.d2 published online Jan 25, 2007; Circ. Res.Catherine C. Hedrick
David T. Bolick, Angela M. Whetzel, Marcus Skaflen, Tracy Deem, Jianyi Lee and Monocyte/Endothelial Interactions in Aorta
Absence of the G Protein-Coupled Receptor G2A In Mice Promotes
http://circres.ahajournals.org/cgi/content/full/01.RES.0000258877.57836.d2/DC1Data Supplement (unedited) at:
http://circres.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circres.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation Research is online at
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Absence of the G Protein–Coupled Receptor G2A In MicePromotes Monocyte/Endothelial Interactions in Aorta
David T. Bolick, Angela M. Whetzel, Marcus Skaflen, Tracy Deem, Jianyi Lee, Catherine C. Hedrick
Abstract—The G protein–coupled receptor G2A is highly expressed on macrophages and lymphocytes and has beenlocalized to atherosclerotic plaques. We examined the role of G2A in modulating monocyte/endothelial interactions inthe vessel wall. We measured adhesion of WEHI 78/24 monocytes to aortas of C57BL/6 (B6) and G2A-deficient(G2A�/�) mice using an ex vivo adhesion assay. G2A�/� mice had 10-fold elevations in adhesion of monocytes to aortas.Injection of GFP-expressing wild-type macrophages into B6 and G2A�/� mice in vivo showed increased macrophageaccumulation in the aortic wall of G2A�/� mice. We isolated aortic endothelial cells (ECs) from B6 and G2A�/� miceand found a 2-fold increase in intercellular adhesion molecule-1 and endothelial selectin surface expression on G2A�/�
ECs using flow cytometry. Using ELISA, we found a 3-fold increase in interleukin-6 and monocyte chemoattractantprotein-1 production by G2A�/� ECs compared with B6 ECs. We found a dramatic increase in nuclear localization ofthe p65 subunit of nuclear factor �B in G2A�/� ECs. Transfection of G2A into G2A�/� ECs to restore normal expressionlevels reduced p65 nuclear localization to 35%. Restoration of G2A expression in G2A�/� ECs significantly reducedintercellular adhesion molecule-1 and endothelial selectin surface expression and reduced monocyte chemoattractantprotein-1 and interleukin-6 production. Restoring G2A to G2A�/� ECs reduced monocyte adhesion by 80% comparedwith G2A�/� ECs in a flow chamber assay. Absence of G2A in endothelium results in proinflammatory signaling andincreased monocyte/endothelial interactions in the aortic wall. Thus, endothelial G2A expression may aid in preventionof vascular inflammation and atherosclerosis. (Circ Res. 2007;100:0-0.)
Key Words: G protein–coupled receptor � NF�B � endothelium � ICAM-1
Akey early event in atherosclerosis is the increasedinteraction of monocytes with endothelial cells in the
vessel wall.1 Monocytes are the primary inflammatory cellslocalized to human atherosclerotic plaques and play a majorrole in atherosclerotic plaque progression.2 Monocytes rollalong the vascular endothelium and are activated by endothe-lial-derived soluble and/or surface-bound chemokines, result-ing in enhanced affinity of LFA-1 (leukocyte function–associated molecule-1) and VLA-4 (via �4�1) integrins fortheir endothelial receptors, intercellular adhesion molecule(ICAM)-1 and vascular cell adhesion molecule (VCAM)-1,respectively. Interaction of these monocytic integrins withICAM-1 and VCAM-1 on the endothelial cell surface causesthe monocyte to tether and firmly adhere to the endothelium,where it can subsequently transmigrate into the subendothe-lial space.3,4
The G protein–coupled receptor G2A was originally iden-tified as a stress-inducible receptor that was induced in pre-Bcells by the Bcr-Abl oncogene.5 G2A overexpression infibroblasts causes cell cycle arrest at the G2 phase of mitosis(thus the name G2A for G2 Accumulation).5 G2A expressionhas been shown to attenuate Bcr-Abl oncogene–mediated cell
proliferation, whereas mice lacking G2A have increasedsusceptibility to oncogene-induced leukemia.6 G2A was pre-viously reported to be a specific receptor for lysophosphati-dylcholine (LPC),7 but this report was later retracted.8 Over-expression studies suggest that G2A is redistributed in thecell in response to LPC treatment.9 However, Shimizu andcolleagues suggest that LPC antagonizes G2A action.10 Re-cently, evidence suggests a role for G2A as a proton sensor inthe cell.11,12 G2A, as well as other receptors within the OGR1family (TDAG8, GPR4, and OGR1), can respond to changesin extracellular pH,12 although G2A is less responsive to pHchanges compared with other receptor family members.11
Very recently, G2A was reported to be a receptor for9-S-hydroxyeicosadienoic acid (9SHODE), a product of the12/15-lipoxygenase family of enzymes.13 Although these datado not include direct binding, Obinata et al demonstrated thatmicromolar concentrations of 9SHODE induced G2A-dependent intracellular calcium mobilization, guanosine 5�-3-O-(thio)triphosphate binding, and pertussis toxin–sensitiveinhibition of cAMP production.13 Thus, G2A is activated bymultiple biological factors and may even serve differentfunctions based on the specificity of these factors.
Original received July 29, 2006; revision received January 5, 2007; accepted January 12, 2007.From the Robert M. Berne Cardiovascular Research Center (D.T.B., A.M.W., M.S., T.D., J.L., C.C.H.), Division of Cardiovascular Medicine (C.C.H.),
and Department of Pharmacology (C.C.H.), University of Virginia, Charlottesville.Correspondence to Catherine C. Hedrick, PhD, Cardiovascular Research Center, University of Virginia, PO Box 801394, 415 Lane Rd, MR5 Rm G123,
Charlottesville, VA 22908. E-mail [email protected]© 2007 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000258877.57836.d2
1 by on May 18, 2011 circres.ahajournals.orgDownloaded from

G2A is highly expressed on T cells, B cells, and macro-phages, and lower expression levels have been found onendothelium.14 G2A has been localized to atheroscleroticplaques in the vessel wall of mice, suggesting a possible rolefor G2A in modulating atherosclerosis.15 In the current study,we examined the role of G2A in modulating endothelialactivation and monocyte/endothelial interactions in vivo us-ing G2A�/� mice. We report that G2A deficiency results inendothelial activation and increased monocyte adhesion tothe vessel wall. Thus, we provide the first evidence of anantiinflammatory role of the G2A receptor in endothelium toprotect against early events of atherogenesis.
Materials and MethodsDetailed methods and reagents used can be found in the online datasupplement, available at http://circres.ahajournals.org. C57BL/6(B6) mice were purchased from The Jackson Laboratories. G2A�/�
were obtained from Dr Owen Witte (University of California, LosAngeles). B6 or G2A�/� mouse aortas were removed, openedlongitudinally, pinned on agar, and used in an ex vivo monocyteadhesion assay as previously described.16 B6 and G2A�/� mice wereinjected with peritoneal macrophages from CX3CR1-GFP macro-phages to measure trafficking to the aorta. Alternatively, aorticendothelial cells (ECs) were freshly harvested and cultured. Passage2 ECs were used for conventional RT-PCR to measure G2Aexpression, and flow cytometry was performed to measure adhesionmolecule expression. Monocyte adhesion to cultured ECs wasperformed using a flow chamber (Glycotech). Fluorescent micros-copy for nuclear factor �B (NF�B) was performed as described.17
Immunoblotting for G2A and signaling molecules was performed onprotein extracts isolated from cultured ECs.16,17
ResultsG2A�/� Aorta Binds More Monocytes ThanControl Mouse AortaWe directly tested whether monocyte adhesion to aorticendothelium was altered in G2A�/� mice versus control B6mice. Aortas from G2A�/� and B6 mice were isolated andimmediately used in an ex vivo monocyte adhesion assay.
The WEHI 78/24 mouse monocyte cell line used in this assayhas comparable G2A expression levels as primary B6 mousemonocytes (data not shown). Aortas from G2A�/� micebound approximately 10-fold more monocytes than did B6aortas (P�0.005) (Figure 1), suggesting that the aorticendothelium of the G2A�/� mice is highly activated. Wehypothesized that monocyte/macrophage adhesion and accu-mulation in G2A�/� aortas would be similarly increased invivo. To test this, we isolated peritoneal macrophages fromheterozygous CX3CR1-GFP mice that express green fluores-cent protein (GFP) in monocyte/macrophages.18 G2A expres-sion in heterozygous CX3CR1-GFP macrophages was thesame as in B6 macrophages (data not shown). The GFP-expressing macrophages were injected into G2A�/� and B6recipient mice. After 96 hours, we isolated aortas andmeasured GFP expression using both immunohistochemistryand immunoblotting. Aortic root sections from B6 andG2A�/� mice were stained using an anti-GFP antibody andare shown in Figure 2A. GFP expression is significantlyhigher in the vessel wall of G2A�/� mice than B6 mice(Figure 2A). Moreover, homogenized aortas from G2A�/�
mice exhibited greater expression of GFP (Figure 2B) thandid B6 control mice by immunoblotting. These data indicatethat G2A-deficient aortas accumulate more macrophages thanB6 aortas. To determine whether there were differences inresident leukocyte populations in C57Bl/6J and G2A�/�
aortas, we performed whole-aorta flow cytometry analysis, atechnique developed by Galkina et al.19 There were signifi-cantly fewer T lymphocytes (CD3�) and significantly highernumbers of macrophages (CD11b�) in G2A�/� aortas com-pared with B6 aortas (Figure 2C). Thus, using 2 separateapproaches, we have determined that there is greater macro-phage accumulation in the aortic wall of G2A�/� micecompared with B6 control mice.
We hypothesized that increased monocyte/endothelial in-teractions in G2A�/� aortas are caused by G2A deficiency in
Figure 1. Aortas from G2A�/� mice showincreased monocyte adhesion. Top,Monocyte adhesion to intact G2A�/�
aorta. Aortas were harvested fromC57BL/6 (B6) and G2A-deficient (G2A�/�)mice as described in Materials andMethods. Fluorescently labeled mono-cytes were added to aortas for an adhe-sion assay and adherent monocyteswere counted by blinded observers usinga fluorescent microscope. Bottom, *sig-nificantly higher than B6 control,P�0.005 by ANOVA. Data represent themean�SE of 3 counted grids per aortafrom 3 mice per group. Data represent 4independent experiments.
2 Circulation Research March 2, 2007
by on May 18, 2011 circres.ahajournals.orgDownloaded from
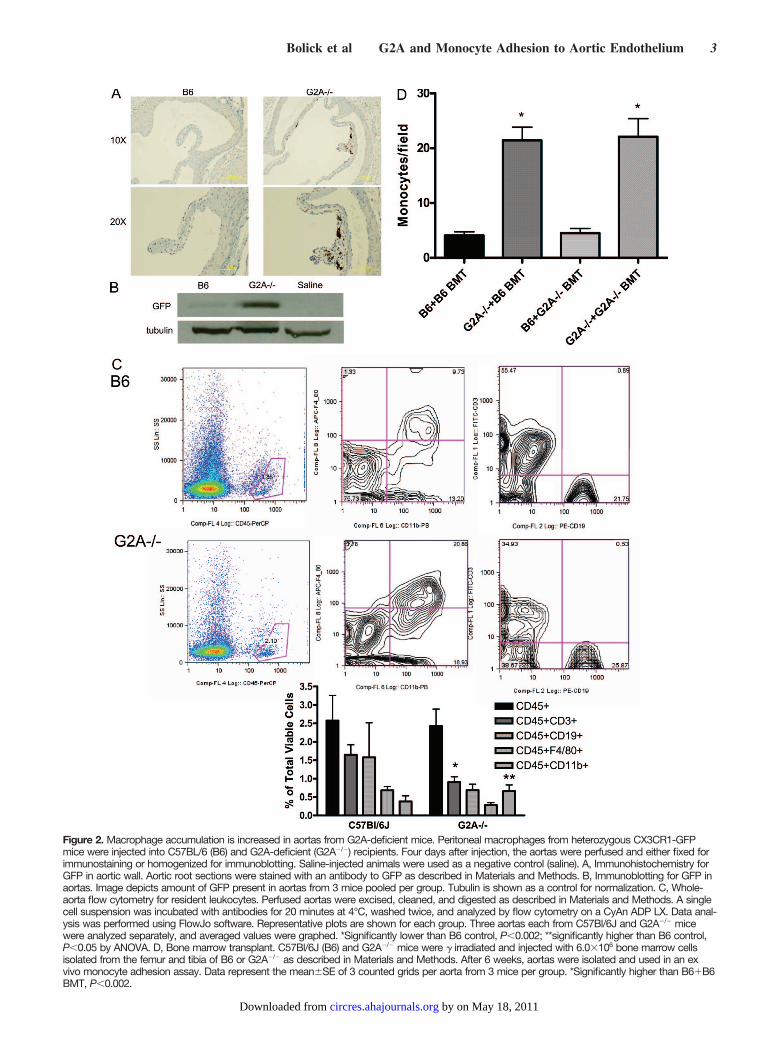
Figure 2. Macrophage accumulation is increased in aortas from G2A-deficient mice. Peritoneal macrophages from heterozygous CX3CR1-GFPmice were injected into C57BL/6 (B6) and G2A-deficient (G2A�/�) recipients. Four days after injection, the aortas were perfused and either fixed forimmunostaining or homogenized for immunoblotting. Saline-injected animals were used as a negative control (saline). A, Immunohistochemistry forGFP in aortic wall. Aortic root sections were stained with an antibody to GFP as described in Materials and Methods. B, Immunoblotting for GFP inaortas. Image depicts amount of GFP present in aortas from 3 mice pooled per group. Tubulin is shown as a control for normalization. C, Whole-aorta flow cytometry for resident leukocytes. Perfused aortas were excised, cleaned, and digested as described in Materials and Methods. A singlecell suspension was incubated with antibodies for 20 minutes at 4°C, washed twice, and analyzed by flow cytometry on a CyAn ADP LX. Data anal-ysis was performed using FlowJo software. Representative plots are shown for each group. Three aortas each from C57Bl/6J and G2A�/� micewere analyzed separately, and averaged values were graphed. *Significantly lower than B6 control, P�0.002; **significantly higher than B6 control,P�0.05 by ANOVA. D, Bone marrow transplant. C57Bl/6J (B6) and G2A�/� mice were � irradiated and injected with 6.0�106 bone marrow cellsisolated from the femur and tibia of B6 or G2A�/� as described in Materials and Methods. After 6 weeks, aortas were isolated and used in an exvivo monocyte adhesion assay. Data represent the mean�SE of 3 counted grids per aorta from 3 mice per group. *Significantly higher than B6�B6BMT, P�0.002.
Bolick et al G2A and Monocyte Adhesion to Aortic Endothelium 3
by on May 18, 2011 circres.ahajournals.orgDownloaded from
the endothelium rather than by G2A deficiency in residentleukocytes. To test this, we performed bone marrow trans-plant studies using bone marrow isolated from both C57Bl/6Jmice and G2A�/� mice injected into �-irradiated B6 andG2A�/� recipient mice. G2A�/� aortas bound significantlymore monocytes in an ex vivo monocyte adhesion assay thanB6 aortas (Figure 2D). This increase in monocyte adhesion toG2A�/� aortas appeared to be independent of whether themice received bone marrow from C57Bl/6J or G2A�/� do-nors. These studies indicate that G2A deficiency in theendothelium is directly responsible for the increased adher-ence and accumulation of monocyte/macrophages to G2A�/�
aortas in vivo.
Expression of Adhesion Molecules andInflammatory Cytokines Is Upregulated inG2A�/� EndotheliumTo understand the mechanisms contributing to enhancedmonocyte adhesion to G2A�/� aortas, we isolated aorticendothelial cells from G2A�/� and control B6 mice. We havesuccessfully isolated and characterized mouse aortic ECs
(MAECs), as reported previously.20 Primary MAECs wereused at passages 2 to 3, and all MAEC experiments wereperformed in DMEM containing 1% heat-inactivated FBS.
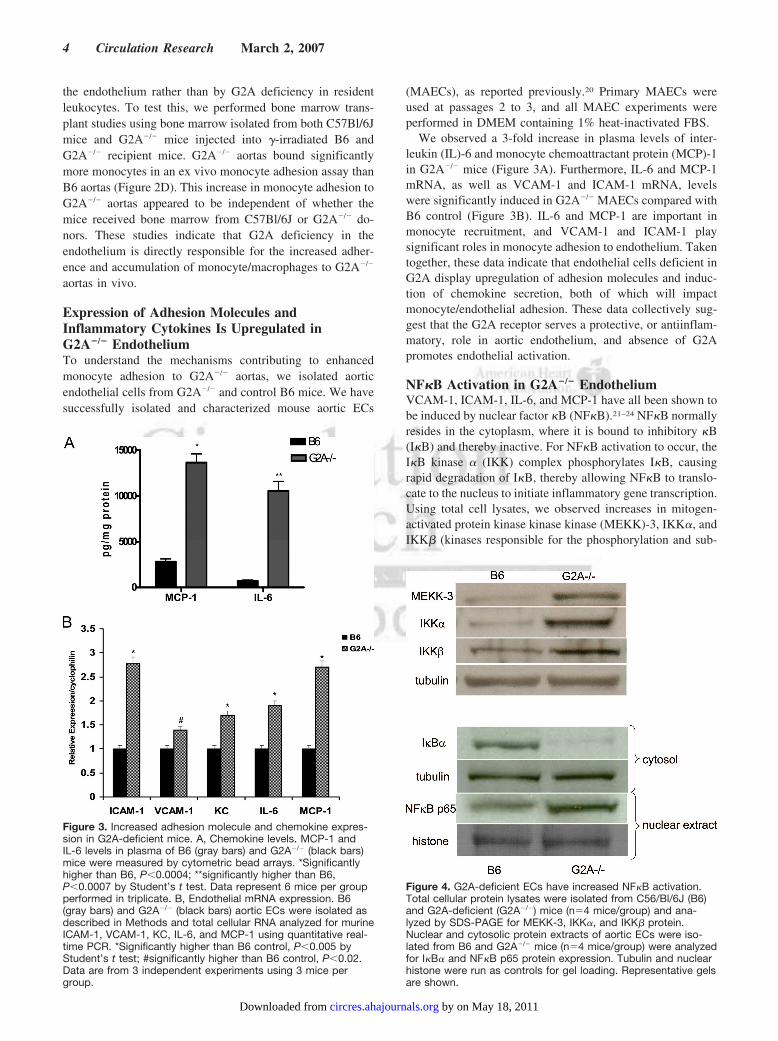
We observed a 3-fold increase in plasma levels of inter-leukin (IL)-6 and monocyte chemoattractant protein (MCP)-1in G2A�/� mice (Figure 3A). Furthermore, IL-6 and MCP-1mRNA, as well as VCAM-1 and ICAM-1 mRNA, levelswere significantly induced in G2A�/� MAECs compared withB6 control (Figure 3B). IL-6 and MCP-1 are important inmonocyte recruitment, and VCAM-1 and ICAM-1 playsignificant roles in monocyte adhesion to endothelium. Takentogether, these data indicate that endothelial cells deficient inG2A display upregulation of adhesion molecules and induc-tion of chemokine secretion, both of which will impactmonocyte/endothelial adhesion. These data collectively sug-gest that the G2A receptor serves a protective, or antiinflam-matory, role in aortic endothelium, and absence of G2Apromotes endothelial activation.
NF�B Activation in G2A�/� EndotheliumVCAM-1, ICAM-1, IL-6, and MCP-1 have all been shown tobe induced by nuclear factor �B (NF�B).21–24 NF�B normallyresides in the cytoplasm, where it is bound to inhibitory �B(I�B) and thereby inactive. For NF�B activation to occur, theI�B kinase � (IKK) complex phosphorylates I�B, causingrapid degradation of I�B, thereby allowing NF�B to translo-cate to the nucleus to initiate inflammatory gene transcription.Using total cell lysates, we observed increases in mitogen-activated protein kinase kinase kinase (MEKK)-3, IKK�, andIKK� (kinases responsible for the phosphorylation and sub-
Figure 3. Increased adhesion molecule and chemokine expres-sion in G2A-deficient mice. A, Chemokine levels. MCP-1 andIL-6 levels in plasma of B6 (gray bars) and G2A�/� (black bars)mice were measured by cytometric bead arrays. *Significantlyhigher than B6, P�0.0004; **significantly higher than B6,P�0.0007 by Student’s t test. Data represent 6 mice per groupperformed in triplicate. B, Endothelial mRNA expression. B6(gray bars) and G2A�/� (black bars) aortic ECs were isolated asdescribed in Methods and total cellular RNA analyzed for murineICAM-1, VCAM-1, KC, IL-6, and MCP-1 using quantitative real-time PCR. *Significantly higher than B6 control, P�0.005 byStudent’s t test; #significantly higher than B6 control, P�0.02.Data are from 3 independent experiments using 3 mice pergroup.
Figure 4. G2A-deficient ECs have increased NF�B activation.Total cellular protein lysates were isolated from C56/Bl/6J (B6)and G2A-deficient (G2A�/�) mice (n�4 mice/group) and ana-lyzed by SDS-PAGE for MEKK-3, IKK�, and IKK� protein.Nuclear and cytosolic protein extracts of aortic ECs were iso-lated from B6 and G2A�/� mice (n�4 mice/group) were analyzedfor I�B� and NF�B p65 protein expression. Tubulin and nuclearhistone were run as controls for gel loading. Representative gelsare shown.
4 Circulation Research March 2, 2007
by on May 18, 2011 circres.ahajournals.orgDownloaded from
sequent degradation of I�B) in G2A�/� ECs compared withB6 (Figure 4). We examined NF�B activation in G2A�/� ECsthrough analysis of cytosolic and nuclear proteins isolatedfrom G2A�/� and B6 ECs. Using immunoblotting for the p65subunit of NF�B and I�B, we observed a significant 55%increase in NF�B p65 expression in the nucleus and acorresponding 60% decrease in I�B� in the cytosol inG2A�/� ECs (Figure 4) compared with B6 ECs. These dataimplicate NF�B as the mediator of the observed activation inG2A�/� ECs.
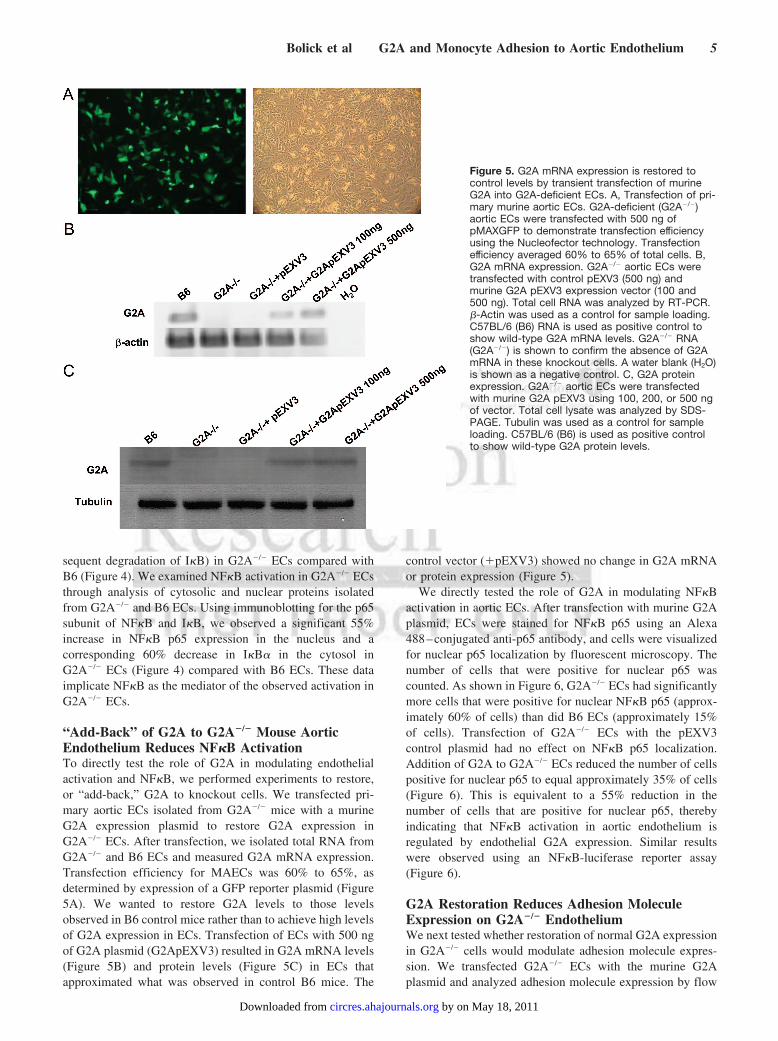
“Add-Back” of G2A to G2A�/� Mouse AorticEndothelium Reduces NF�B ActivationTo directly test the role of G2A in modulating endothelialactivation and NF�B, we performed experiments to restore,or “add-back,” G2A to knockout cells. We transfected pri-mary aortic ECs isolated from G2A�/� mice with a murineG2A expression plasmid to restore G2A expression inG2A�/� ECs. After transfection, we isolated total RNA fromG2A�/� and B6 ECs and measured G2A mRNA expression.Transfection efficiency for MAECs was 60% to 65%, asdetermined by expression of a GFP reporter plasmid (Figure5A). We wanted to restore G2A levels to those levelsobserved in B6 control mice rather than to achieve high levelsof G2A expression in ECs. Transfection of ECs with 500 ngof G2A plasmid (G2ApEXV3) resulted in G2A mRNA levels(Figure 5B) and protein levels (Figure 5C) in ECs thatapproximated what was observed in control B6 mice. The
control vector (�pEXV3) showed no change in G2A mRNAor protein expression (Figure 5).
We directly tested the role of G2A in modulating NF�Bactivation in aortic ECs. After transfection with murine G2Aplasmid, ECs were stained for NF�B p65 using an Alexa488–conjugated anti-p65 antibody, and cells were visualizedfor nuclear p65 localization by fluorescent microscopy. Thenumber of cells that were positive for nuclear p65 wascounted. As shown in Figure 6, G2A�/� ECs had significantlymore cells that were positive for nuclear NF�B p65 (approx-imately 60% of cells) than did B6 ECs (approximately 15%of cells). Transfection of G2A�/� ECs with the pEXV3control plasmid had no effect on NF�B p65 localization.Addition of G2A to G2A�/� ECs reduced the number of cellspositive for nuclear p65 to equal approximately 35% of cells(Figure 6). This is equivalent to a 55% reduction in thenumber of cells that are positive for nuclear p65, therebyindicating that NF�B activation in aortic endothelium isregulated by endothelial G2A expression. Similar resultswere observed using an NF�B-luciferase reporter assay(Figure 6).
G2A Restoration Reduces Adhesion MoleculeExpression on G2A�/� EndotheliumWe next tested whether restoration of normal G2A expressionin G2A�/� cells would modulate adhesion molecule expres-sion. We transfected G2A�/� ECs with the murine G2Aplasmid and analyzed adhesion molecule expression by flow
Figure 5. G2A mRNA expression is restored tocontrol levels by transient transfection of murineG2A into G2A-deficient ECs. A, Transfection of pri-mary murine aortic ECs. G2A-deficient (G2A�/�)aortic ECs were transfected with 500 ng ofpMAXGFP to demonstrate transfection efficiencyusing the Nucleofector technology. Transfectionefficiency averaged 60% to 65% of total cells. B,G2A mRNA expression. G2A�/� aortic ECs weretransfected with control pEXV3 (500 ng) andmurine G2A pEXV3 expression vector (100 and500 ng). Total cell RNA was analyzed by RT-PCR.�-Actin was used as a control for sample loading.C57BL/6 (B6) RNA is used as positive control toshow wild-type G2A mRNA levels. G2A�/� RNA(G2A�/�) is shown to confirm the absence of G2AmRNA in these knockout cells. A water blank (H2O)is shown as a negative control. C, G2A proteinexpression. G2A�/� aortic ECs were transfectedwith murine G2A pEXV3 using 100, 200, or 500 ngof vector. Total cell lysate was analyzed by SDS-PAGE. Tubulin was used as a control for sampleloading. C57BL/6 (B6) is used as positive controlto show wild-type G2A protein levels.
Bolick et al G2A and Monocyte Adhesion to Aortic Endothelium 5
by on May 18, 2011 circres.ahajournals.orgDownloaded from
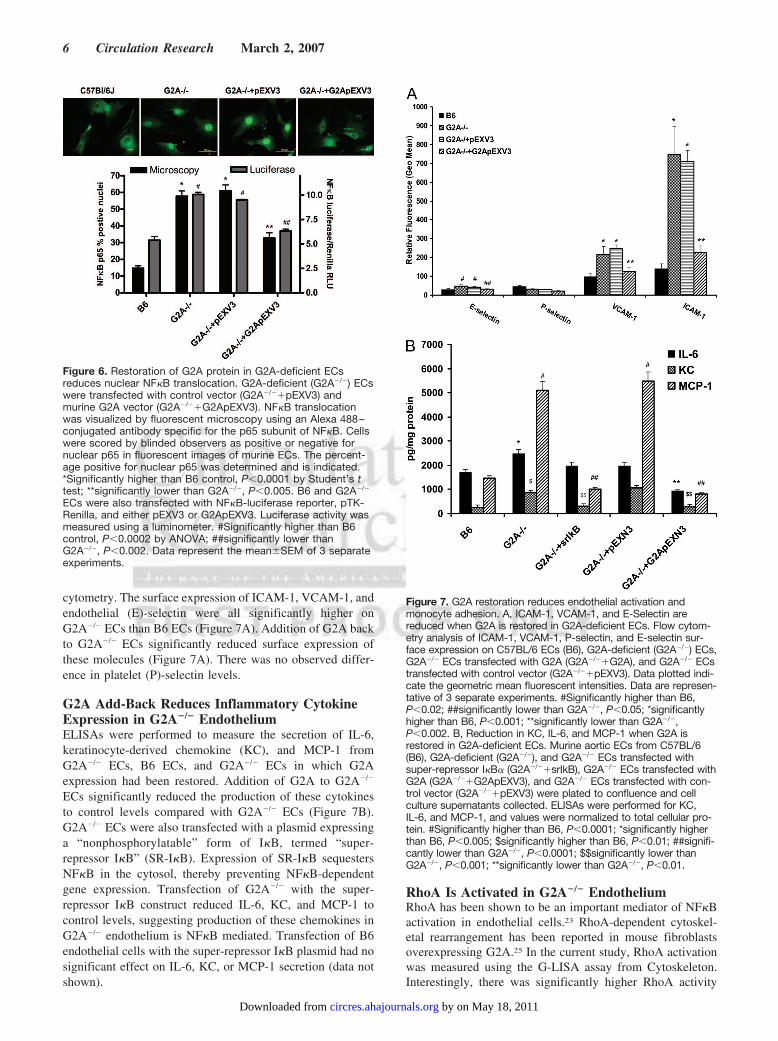
cytometry. The surface expression of ICAM-1, VCAM-1, andendothelial (E)-selectin were all significantly higher onG2A�/� ECs than B6 ECs (Figure 7A). Addition of G2A backto G2A�/� ECs significantly reduced surface expression ofthese molecules (Figure 7A). There was no observed differ-ence in platelet (P)-selectin levels.
G2A Add-Back Reduces Inflammatory CytokineExpression in G2A�/� EndotheliumELISAs were performed to measure the secretion of IL-6,keratinocyte-derived chemokine (KC), and MCP-1 fromG2A�/� ECs, B6 ECs, and G2A�/� ECs in which G2Aexpression had been restored. Addition of G2A to G2A�/�
ECs significantly reduced the production of these cytokinesto control levels compared with G2A�/� ECs (Figure 7B).G2A�/� ECs were also transfected with a plasmid expressinga “nonphosphorylatable” form of I�B, termed “super-repressor I�B” (SR-I�B). Expression of SR-I�B sequestersNF�B in the cytosol, thereby preventing NF�B-dependentgene expression. Transfection of G2A�/� with the super-repressor I�B construct reduced IL-6, KC, and MCP-1 tocontrol levels, suggesting production of these chemokines inG2A�/� endothelium is NF�B mediated. Transfection of B6endothelial cells with the super-repressor I�B plasmid had nosignificant effect on IL-6, KC, or MCP-1 secretion (data notshown).
RhoA Is Activated in G2A�/� EndotheliumRhoA has been shown to be an important mediator of NF�Bactivation in endothelial cells.23 RhoA-dependent cytoskel-etal rearrangement has been reported in mouse fibroblastsoverexpressing G2A.25 In the current study, RhoA activationwas measured using the G-LISA assay from Cytoskeleton.Interestingly, there was significantly higher RhoA activity
Figure 6. Restoration of G2A protein in G2A-deficient ECsreduces nuclear NF�B translocation. G2A-deficient (G2A�/�) ECswere transfected with control vector (G2A�/��pEXV3) andmurine G2A vector (G2A�/��G2ApEXV3). NF�B translocationwas visualized by fluorescent microscopy using an Alexa 488–conjugated antibody specific for the p65 subunit of NF�B. Cellswere scored by blinded observers as positive or negative fornuclear p65 in fluorescent images of murine ECs. The percent-age positive for nuclear p65 was determined and is indicated.*Significantly higher than B6 control, P�0.0001 by Student’s ttest; **significantly lower than G2A�/�, P�0.005. B6 and G2A�/�
ECs were also transfected with NF�B-luciferase reporter, pTK-Renilla, and either pEXV3 or G2ApEXV3. Luciferase activity wasmeasured using a luminometer. #Significantly higher than B6control, P�0.0002 by ANOVA; ##significantly lower thanG2A�/�, P�0.002. Data represent the mean�SEM of 3 separateexperiments.
Figure 7. G2A restoration reduces endothelial activation andmonocyte adhesion. A, ICAM-1, VCAM-1, and E-Selectin arereduced when G2A is restored in G2A-deficient ECs. Flow cytom-etry analysis of ICAM-1, VCAM-1, P-selectin, and E-selectin sur-face expression on C57BL/6 ECs (B6), G2A-deficient (G2A�/�) ECs,G2A�/� ECs transfected with G2A (G2A�/��G2A), and G2A�/� ECstransfected with control vector (G2A�/��pEXV3). Data plotted indi-cate the geometric mean fluorescent intensities. Data are represen-tative of 3 separate experiments. #Significantly higher than B6,P�0.02; ##significantly lower than G2A�/�, P�0.05; *significantlyhigher than B6, P�0.001; **significantly lower than G2A�/�,P�0.002. B, Reduction in KC, IL-6, and MCP-1 when G2A isrestored in G2A-deficient ECs. Murine aortic ECs from C57BL/6(B6), G2A-deficient (G2A�/�), and G2A�/� ECs transfected withsuper-repressor I�B� (G2A�/��srIkB), G2A�/� ECs transfected withG2A (G2A�/��G2ApEXV3), and G2A�/� ECs transfected with con-trol vector (G2A�/��pEXV3) were plated to confluence and cellculture supernatants collected. ELISAs were performed for KC,IL-6, and MCP-1, and values were normalized to total cellular pro-tein. #Significantly higher than B6, P�0.0001; *significantly higherthan B6, P�0.005; $significantly higher than B6, P�0.01; ##signifi-cantly lower than G2A�/�, P�0.0001; $$significantly lower thanG2A�/�, P�0.001; **significantly lower than G2A�/�, P�0.01.
6 Circulation Research March 2, 2007
by on May 18, 2011 circres.ahajournals.orgDownloaded from
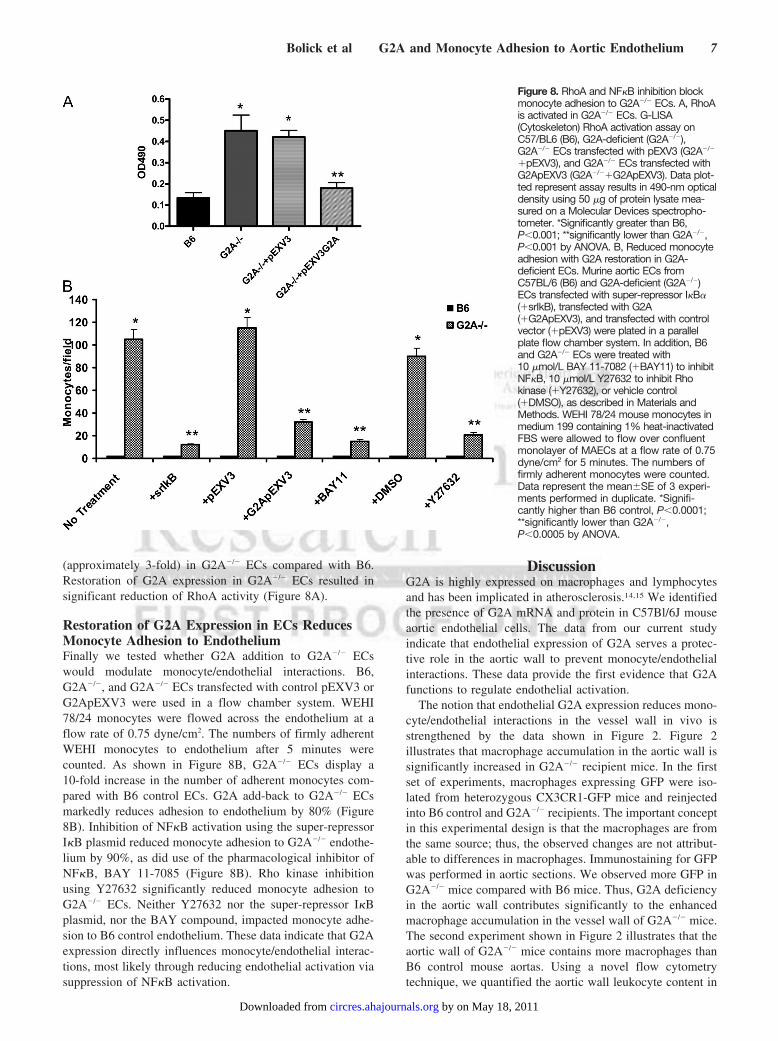
(approximately 3-fold) in G2A�/� ECs compared with B6.Restoration of G2A expression in G2A�/� ECs resulted insignificant reduction of RhoA activity (Figure 8A).
Restoration of G2A Expression in ECs ReducesMonocyte Adhesion to EndotheliumFinally we tested whether G2A addition to G2A�/� ECswould modulate monocyte/endothelial interactions. B6,G2A�/�, and G2A�/� ECs transfected with control pEXV3 orG2ApEXV3 were used in a flow chamber system. WEHI78/24 monocytes were flowed across the endothelium at aflow rate of 0.75 dyne/cm2. The numbers of firmly adherentWEHI monocytes to endothelium after 5 minutes werecounted. As shown in Figure 8B, G2A�/� ECs display a10-fold increase in the number of adherent monocytes com-pared with B6 control ECs. G2A add-back to G2A�/� ECsmarkedly reduces adhesion to endothelium by 80% (Figure8B). Inhibition of NF�B activation using the super-repressorI�B plasmid reduced monocyte adhesion to G2A�/� endothe-lium by 90%, as did use of the pharmacological inhibitor ofNF�B, BAY 11-7085 (Figure 8B). Rho kinase inhibitionusing Y27632 significantly reduced monocyte adhesion toG2A�/� ECs. Neither Y27632 nor the super-repressor I�Bplasmid, nor the BAY compound, impacted monocyte adhe-sion to B6 control endothelium. These data indicate that G2Aexpression directly influences monocyte/endothelial interac-tions, most likely through reducing endothelial activation viasuppression of NF�B activation.
DiscussionG2A is highly expressed on macrophages and lymphocytesand has been implicated in atherosclerosis.14,15 We identifiedthe presence of G2A mRNA and protein in C57Bl/6J mouseaortic endothelial cells. The data from our current studyindicate that endothelial expression of G2A serves a protec-tive role in the aortic wall to prevent monocyte/endothelialinteractions. These data provide the first evidence that G2Afunctions to regulate endothelial activation.
The notion that endothelial G2A expression reduces mono-cyte/endothelial interactions in the vessel wall in vivo isstrengthened by the data shown in Figure 2. Figure 2illustrates that macrophage accumulation in the aortic wall issignificantly increased in G2A�/� recipient mice. In the firstset of experiments, macrophages expressing GFP were iso-lated from heterozygous CX3CR1-GFP mice and reinjectedinto B6 control and G2A�/� recipients. The important conceptin this experimental design is that the macrophages are fromthe same source; thus, the observed changes are not attribut-able to differences in macrophages. Immunostaining for GFPwas performed in aortic sections. We observed more GFP inG2A�/� mice compared with B6 mice. Thus, G2A deficiencyin the aortic wall contributes significantly to the enhancedmacrophage accumulation in the vessel wall of G2A�/� mice.The second experiment shown in Figure 2 illustrates that theaortic wall of G2A�/� mice contains more macrophages thanB6 control mouse aortas. Using a novel flow cytometrytechnique, we quantified the aortic wall leukocyte content in
Figure 8. RhoA and NF�B inhibition blockmonocyte adhesion to G2A�/� ECs. A, RhoAis activated in G2A�/� ECs. G-LISA(Cytoskeleton) RhoA activation assay onC57/BL6 (B6), G2A-deficient (G2A�/�),G2A�/� ECs transfected with pEXV3 (G2A�/�
�pEXV3), and G2A�/� ECs transfected withG2ApEXV3 (G2A�/��G2ApEXV3). Data plot-ted represent assay results in 490-nm opticaldensity using 50 �g of protein lysate mea-sured on a Molecular Devices spectropho-tometer. *Significantly greater than B6,P�0.001; **significantly lower than G2A�/�,P�0.001 by ANOVA. B, Reduced monocyteadhesion with G2A restoration in G2A-deficient ECs. Murine aortic ECs fromC57BL/6 (B6) and G2A-deficient (G2A�/�)ECs transfected with super-repressor I�B�(�srIkB), transfected with G2A(�G2ApEXV3), and transfected with controlvector (�pEXV3) were plated in a parallelplate flow chamber system. In addition, B6and G2A�/� ECs were treated with10 �mol/L BAY 11-7082 (�BAY11) to inhibitNF�B, 10 �mol/L Y27632 to inhibit Rhokinase (�Y27632), or vehicle control(�DMSO), as described in Materials andMethods. WEHI 78/24 mouse monocytes inmedium 199 containing 1% heat-inactivatedFBS were allowed to flow over confluentmonolayer of MAECs at a flow rate of 0.75dyne/cm2 for 5 minutes. The numbers offirmly adherent monocytes were counted.Data represent the mean�SE of 3 experi-ments performed in duplicate. *Signifi-cantly higher than B6 control, P�0.0001;**significantly lower than G2A�/�,P�0.0005 by ANOVA.
Bolick et al G2A and Monocyte Adhesion to Aortic Endothelium 7
by on May 18, 2011 circres.ahajournals.orgDownloaded from

B6 versus G2A�/� mice. We found a significant increase inthe number of macrophages present in the aortic wall ofG2A�/� mice. We found no changes in B-cell content in theaortic wall, yet T-lymphocyte numbers were lower in G2A�/�
aortas than in B6 controls. These data do not rule out potentialdifferences in trafficking of the leukocytes to the aortic wallin the recipient animals, but trafficking of leukocytes to theaortic wall could be dramatically impacted by the endothelialactivation state of the aortas. These data also do not rule outsome contribution of leukocyte activation on the endothelialactivation state in the G2A�/� mice. In an attempt to addresssuch concerns, we performed a series of bone marrowtransplant studies in which we examined monocyte adhesionto the aortic wall as a functional measure of endothelialactivation. In this study, B6 or G2A�/� recipients weretransplanted with either G2A�/� or B6 bone marrow. After 6weeks of recovery on a rodent chow diet, aortas wereharvested from the mice and used in an ex vivo assay tomeasure monocyte adhesion. If endothelial G2A expressionwere of primary importance in regulating monocyte/endothe-lial interactions, then we would have expected to find thatG2A expression in myeloid cells would have no impact onmonocyte adhesion to aorta. The results of the study showedexactly these results (Figure 2D), indicating that G2A expres-sion in endothelium, not in myeloid cells, regulates mono-cyte/endothelial interactions in the aortic wall. The fact thatwe observed decreased T-lymphocyte accumulation in thewall suggests that lymphocyte activation of the endotheliummay not be important in this particular case; however, it ispossible that the lymphocytes that are present in the aorticwall contribute to endothelial activation. Both macrophagesand lymphocytes in the aortic wall may indeed contribute toendothelial activation in the G2A�/� mice through cytokinesecretion; however, our data suggest that the absence of G2Ain endothelium is indeed a direct, significant contributor toendothelial activation.
With regard to signaling pathways in endothelium that maybe regulated by G2A, the most striking evidence is theenhanced NF�B activation observed in G2A�/� mice. Weobserved dramatic changes in nuclear translocation of NF�Band in cytosolic I�B degradation in G2A�/� endothelium(Figures 4 and 6). Restoration of G2A expression strikinglyreduced NF�B activation, reduced endothelial expression ofICAM-1, IL-6, KC, and MCP-1, and dramatically reducedmonocyte adhesion to endothelium. In contrast, Lin and Yereported that overexpression of G2A in HeLa cells increasedNF�B-luciferase production.26 However, these studies wereperformed using overexpression of G2A in a HeLa cell lineand transfection of RGS (regulator of G protein signaling)constructs to block NF�B signaling. Although these overex-pression studies suggest that NF�B can be activated by G2Ain a LPC-independent manner, they do not preclude that thereis ligand-dependent NF�B inhibition occurring in the cell invivo. Perhaps control of NF�B activation by G2A depends onligand availability, or there could be cell-type specific. Ourdata indicate that NF�B regulation by G2A is critical inendothelium. Recently, we observed upregulation of p38mitogen-activated protein kinase and activator protein-1 inG2A�/� ECs using a set of gene arrays (data not shown). The
p38 mitogen-activated protein kinase pathway is linked toNF�B activation.27 Studies are currently ongoing in ourlaboratory to study these additional signaling pathways in-duced by G2A deficiency in aortic endothelium.
Despite conflicting evidence regarding the agonist/antago-nist effects of LPC on G2A,7–10 LPC has remained a focalpoint in the studies of the role of G2A in atherosclerosis.14,15
Although direct binding of LPC to G2A is unlikely,8 there isevidence of indirect action of LPC on G2A cellular distribu-tion.9 Recent studies in lipid metabolism28 suggest there is alink between LPC and arachidonic acid synthesis. The 12/15lipoxygenase pathway converts arachidonic acid into 12-S-hydroxyeicosatetraenoic acid (12-S-HETE), 15-S-HETE, andlipoxin A4, all of which have been implicated in either aproinflammatory or antiinflammatory manner in atheroscle-rosis development.29 –31 Obinata et al identifies9-hydroxyoctadecadienoic acid (9SHODE), and other eico-sanoids, as ligands for G2A,13 suggesting that G2A may be amultiligand receptor. Studies are currently ongoing in thelaboratory to further identify eicosanoid ligands of G2A.
Despite evidence suggesting that G2A is activated bymultiple lipids, there is also evidence to suggest that G2A isconstitutively active in the absence of ligand. When overex-pressed in fibroblasts, G2A causes constitutive RhoA activa-tion through G�13.25 Lin and Ye have reported that G2Astimulates the accumulation of both inositol phosphates andcAMP in the absence of ligand and that G2A differentiallycouples to G�s, G�q, or G�13 depending on whether it isbound to ligand.26 Thus it is also possible that G2A isconstitutively active in the cell, even in the absence of ligand.
In summary, we report that absence of G2A in aorticendothelium promotes endothelial activation and monocyte/endothelial interactions. Absence of G2A expression in en-dothelium promotes NF�B activation and induces expressionof proinflammatory endothelial chemokines and adhesionmolecules. Thus, G2A expression in vascular endotheliummay serve a protective role for prevention of early events ofinflammation and atherosclerosis.
AcknowledgmentsWe thank Dr Owen Witte and Dr Caius Radu (Howard HughesMedical Institute, University of California, Los Angeles) for the giftsof the G2A�/� mice and murine G2A antibody. We thank Dr SteffenJung (Lerner Research Institute, Cleveland Clinic) and Dr Klaus Ley(University of Virginia) for the gift of the CX3CR1-GFP mice. Wethank Dr Janusz Kabarowski (University of Alabama, Birmingham)for the gift of the control and murine G2A pEXV3 plasmids and DrDean Ballard (Vanderbilt University, Nashville, Tenn) for thesuper-repressor I�B plasmid. We thank Leah Cochran for assistancewith mouse genotyping. We also thank Dr Klaus Ley and Dr OwenWitte for helpful discussions and critical review of the manuscript.
Sources of FundingThis work was supported by NIH grant R01 HL071141 (to C.C.H.).
DisclosuresNone.
References1. Witztum JL, Steinberg D. Role of oxidized low density lipoprotein in
atherogenesis. J Clin Invest. 1991;88:1785–1792.
8 Circulation Research March 2, 2007
by on May 18, 2011 circres.ahajournals.orgDownloaded from

2. Gerrity RG. The role of the monocyte in atherogenesis: II. Migration offoam cells from atherosclerotic lesions. Am J Pathol. 1981;103:191–200.
3. Springer TA. Traffic signals on endothelium for lymphocyte recirculationand leukocyte emigration. Annu Rev Physiol. 1995;57:827–872.
4. Berliner JA, Navab M, Fogelman AM, Frank JS, Demer LL, Edwards PA,Watson AD, Lusis AJ. Atherosclerosis: basic mechanisms. Oxidation,inflammation, and genetics. Circulation. 1995;91:2488–2496.
5. Weng Z, Fluckiger AC, Nisitani S, Wahl MI, Le LQ, Hunter CA, FernalAA, Le Beau MM, Witte ON. A DNA damage and stress inducible Gprotein-coupled receptor blocks cells in G2/M. Proc Natl Acad Sci U S A.1998;95:12334–12339.
6. Le LQ, Kabarowski JH, Wong S, Nguyen K, Gambhir SS, Witte ON.Positron emission tomography imaging analysis of G2A as a negativemodifier of lymphoid leukemogenesis initiated by the BCR-ABLoncogene. Cancer Cell. 2002;1:381–391.
7. Kabarowski JH, Zhu K, Le LQ, Witte ON, Xu Y. Lysophosphatidyl-choline as a ligand for the immunoregulatory receptor G2A. Science.2001;293:702–705.
8. Witte ON, Kabarowski JH, Xu Y, Le LQ, Zhu K. Retraction. Science.2005;307:206.
9. Wang L, Radu CG, Yang LV, Bentolila LA, Riedinger M, Witte ON.Lysophosphatidylcholine-induced surface redistribution regulates sig-naling of the murine G protein-coupled receptor G2A. Mol Biol Cell.2005;16:2234–2247.
10. Murakami N, Yokomizo T, Okuno T, Shimizu T. G2A is a proton-sensingG-protein-coupled receptor antagonized by lysophosphatidylcholine.J Biol Chem. 2004;279:42484–42491.
11. Radu CG, Nijagal A, McLaughlin J, Wang L, Witte ON. Differentialproton sensitivity of related G protein-coupled receptors T cell death-associated gene 8 and G2A expressed in immune cells. Proc Natl AcadSci U S A. 2005;102:1632–1637.
12. Tomura H, Mogi C, Sato K, Okajima F. Proton-sensing and lysolipid-sensitive G-protein-coupled receptors: a novel type of multi-functionalreceptors. Cell Signal. 2005;17:1466–1476.
13. Obinata H, Hattori T, Nakane S, Tatei K, Izumi T. Identification of9-hydroxyoctadecadienoic acid and other oxidized free fatty acids asligands of the G protein-coupled receptor G2A. J Biol Chem. 2005;280:40676–40683.
14. Parks BW, Gambill GP, Lusis AJ, Kabarowski JH. Loss of G2A promotesmacrophage accumulation in atherosclerotic lesions of low densitylipoprotein receptor-deficient mice. J Lipid Res. 2005;46:1405–1415.
15. Rikitake Y, Hirata K, Yamashita T, Iwai K, Kobayashi S, Itoh H, OzakiM, Ejiri J, Shiomi M, Inoue N, Kawashima S, Yokoyama M. Expressionof G2A, a receptor for lysophosphatidylcholine, by macrophages inmurine, rabbit, and human atherosclerotic plaques. Arterioscler ThrombVasc Biol. 2002;22:2049–2053.
16. Bolick DT, Srinivasan S, Kim KW, Hatley ME, Clemens JJ, Whetzel A,Ferger N, Macdonald TL, Davis MD, Tsao PS, Lynch KR, Hedrick CC.Sphingosine-1-phosphate prevents tumor necrosis factor-{alpha}-mediated monocyte adhesion to aortic endothelium in mice. ArteriosclerThromb Vasc Biol. 2005;25:976–981.
17. Bolick DT, Srinivasan S, Whetzel A, Fuller LC, Hedrick CC. 12/15Lipoxygenase mediates monocyte adhesion to aortic endothelium in apo-lipoprotein E-deficient mice through activation of RhoA andNF-{kappa}B. Arterioscler Thromb Vasc Biol. 2006;26:1260–1266.
18. Jung S, Aliberti J, Graemmel P, Sunshine MJ, Kreutzberg GW, Sher A,Littman DR. Analysis of fractalkine receptor CX(3)CR1 function by
targeted deletion and green fluorescent protein reporter gene insertion.Mol Cell Biol. 2000;20:4106–4114.
19. Galkina E, Kadl A, Sanders J, Varughese D, Sarembock IJ, Ley K.Lymphocyte recruitment into the aortic wall before and during devel-opment of atherosclerosis is partially L-selectin dependent. J Exp Med.2006;203:1273–1282.
20. Hatley ME, Srinivasan S, Reilly KB, Bolick DT, Hedrick CC. Increasedproduction of 12/15 lipoxygenase eicosanoids accelerates monocyte/en-dothelial interactions in diabetic db/db mice. J Biol Chem. 2003;278:25369–25375.
21. Perona R, Montaner S, Saniger L, Sanchez-Perez I, Bravo R, Lacal JC.Activation of the nuclear factor-kappaB by Rho, CDC42, and Rac-1proteins. Genes Dev. 1997;11:463–475.
22. Read MA, Whitley MZ, Gupta S, Pierce JW, Best J, Davis RJ, Collins T.Tumor necrosis factor alpha-induced E-selectin expression is activated bythe nuclear factor-kappaB and c-JUN N-terminal kinase/p38 mitogen-ac-tivated protein kinase pathways. J Biol Chem. 1997;272:2753–2761.
23. Bolick DT, Orr AW, Whetzel A, Srinivasan S, Hatley ME, Schwartz MA,Hedrick CC. 12/15-Lipoxygenase regulates intercellular adhesionmolecule-1 expression and monocyte adhesion to endothelium throughactivation of RhoA and nuclear factor-kappaB. Arterioscler Thromb VascBiol. 2005;25:2301–2307.
24. Manning AM, Bell FP, Rosenbloom CL, Chosay JG, Simmons CA,Northrup JL, Shebuski RJ, Dunn CJ, Anderson DC. NF-kappa B isactivated during acute inflammation in vivo in association with elevatedendothelial cell adhesion molecule gene expression and leukocyterecruitment. J Inflamm. 1995;45:283–296.
25. Kabarowski JH, Feramisco JD, Le LQ, Gu JL, Luoh SW, Simon MI,Witte ON. Direct genetic demonstration of G alpha 13 coupling to theorphan G protein-coupled receptor G2A leading to RhoA-dependent actinrearrangement. Proc Natl Acad Sci U S A. 2000;97:12109–12114.
26. Lin P, Ye RD. The lysophospholipid receptor G2A activates a specificcombination of G proteins and promotes apoptosis. J Biol Chem. 2003;278:14379–14386.
27. Lopez-Pedrera C, Buendia P, Cuadrado MJ, Siendones E, Aguirre MA,Barbarroja N, Montiel-Duarte C, Torres A, Khamashta M, Velasco F.Antiphospholipid antibodies from patients with the antiphospholipidsyndrome induce monocyte tissue factor expression through the simulta-neous activation of NF-kappaB/Rel proteins via the p38 mitogen-acti-vated protein kinase pathway, and of the MEK-1/ERK pathway. ArthritisRheum. 2006;54:301–311.
28. Yan W, Jenkins CM, Han X, Mancuso DJ, Sims HF, Yang K, Gross RW.The highly selective production of 2-arachidonoyl lysophosphatidyl-choline catalyzed by purified calcium-independent phospholipaseA2gamma: identification of a novel enzymatic mediator for the gen-eration of a key branch point intermediate in eicosanoid signaling. J BiolChem. 2005;280:26669–26679.
29. Cyrus T, Witztum JL, Rader DJ, Tangirala R, Fazio S, Linton MF, FunkCD. Disruption of the 12/15-lipoxygenase gene diminishes atherosclero-sis in apo E-deficient mice. J Clin Invest. 1999;103:1597–1604.
30. Serhan CN, Drazen JM. Antiinflammatory potential of lipoxygenase-derived eicosanoids: a molecular switch at 5 and 15 positions? J ClinInvest. 1997;99:1147–1148.
31. Shen J, Herderick E, Cornhill JF, Zsigmond E, Kim HS, Kuhn H,Guevara NV, Chan L. Macrophage-mediated 15-lipoxygenase expressionprotects against atherosclerosis development. J Clin Invest. 1996;98:2201–2208.
Bolick et al G2A and Monocyte Adhesion to Aortic Endothelium 9
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
Reagents. WEHI 78/24 monocyte cells were a kind gift of Dr Judy Berliner (UCLA).
Control plasmid, pEXV3, and a plasmid containing the full-length murine G2A cDNA
(G2ApEXV3) were kind gifts of Dr. Janus Kabarowski (University of Alabama
Birmingham). The super-repressor IκB plasmid was obtained from Dr. Julie Reuther
(University of North Carolina). CX3CR1-GFP mice1 were a kind gift from Dr Klaus Ley
(University of Virginia) and Dr Steffen Jung (Cleveland Clinic). Cytometric bead arrays
(#552364) were from BD Biosciences. Antibodies used were: PE-conjugated ICAM-1
(clone 3E2), FITC-conjugated VCAM-1 (clone 429), FITC-conjugated P-selectin (clone
GMP140), and PE-conjugated E-selectin (clone 10E9.6) for flow cytometry (all from BD
Biosciences). Isotype antibodies included rat IgG1 (control for E-selectin antibody), and
rat IgG2b (control for ICAM-1 antibody) and rat IgG1k (control for VCAM-1 antibody).
NFκB p65 antibody (# sc-109), histone, and GFP antibody were from Santa Cruz, and
IKKα/β and IkBα antibodies was from Cell Signaling. Tubulin antibody was obtained
from Sigma. Alexa 488 antibody was from Molecular Probes. BAY 11-7085 was from
BIOMOL Research Laboratories.
Mice: C57BL/6 (B6) mice were purchased from Jackson Laboratories. G2A-deficient
mice were constructed using homologous recombination in 129svJ embryonic stem
cells to disrupt the G2A locus as previously described by Witte and colleagues22. G2A-
deficient mice have been subsequently backcrossed for 12 generations onto a
C57BL/6J background. G2A-deficient mice are healthy and viable, and appear
indistinguishable from littermate controls. B6 and G2A-/- mice were maintained on
rodent chow except for atherosclerosis studies. All animal studies were performed
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
following approved guidelines of the University of Virginia Animal Care and Use
Committee.
Mouse Aorta Isolation and Ex Vivo Adhesion Assay: Aortas were harvested from
the mice, opened up longitudinally, and pinned onto sterile agar. Aortas were
immediately incubated for 30 mins with 1x106 fluorescently-labeled (using Calcein AM
according to manufacturer’s instructions) WEHI 78/24 mouse monocytes in 1% DMEM.
WEHI 78/24 cells are a mouse monocyte cell line that has been fully characterized by
McEvoy and colleagues 3. WEHI 78/24 monocytes are maintained in DMEM +10%
heat-inactivated FBS prior to use. After incubation, unbound monocytes were rinsed
away, and the number of monocytes firmly bound to aorta was counted in 3 consistent
fields using fluorescent microscopy 4. The areas counted were identified within a
10mmx10mm grid, and the same locations per aorta were counted by a blinded
observer.
Isolation of mouse aortic endothelial cells: Aortic EC from B6 and G2A-/- mice were
harvested from mouse aorta under sterile conditions as outlined previously 5. Mouse
endothelial cell cultures were cultured in DMEM containing 15% heat-inactivated FBS,
30µg/ml ECGS, and 50µg/ml heparin. For all in vitro experiments, EC cultures were
switched to DMEM containing 1% heat-inactivated FBS for 24h prior to experiments,
and were maintained in 1% DMEM throughout the experiment. EC cultures were used
in experiments from passages 2 to 3.
Flow chamber studies: We have developed a monocyte adhesion assay under flow
conditions that utilizes primary MAEC and WEHI 78/24 cells. MAEC were cultured to
confluency in 60mm dishes and used in a parallel plate flow chamber (Glycotech).
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
WEHI 78/24 mouse monocytes (1x106 cells/ml were labeled with Calcein-AM (Molecular
Probes) according to manufacturer’s instructions. The WEHI cells were allowed to flow
over the endothelium at a flow rate of 0.75 dynes/cm2. Data was recorded for 5 minutes
using a video recorder as previously described 6;7. The total number of firmly adherent
WEHI monocytes to the endothelium at the end of the 5 minute period were counted by
fluorescent microscopy using a 10mmx10mm eyepiece grid and a 10X objective7.
Triplicate plates were performed for each experimental condition. For treatments
MAEC were either transfected as described below or treated for 4h with 10µM BAY 11-
7085 to inhibit NFκB, 10µM Y27632 to inhibit Rho kinase, or with DMSO as a vehicle
control prior to performing a flow chamber assay.
Immunoblotting for NFκB. MAEC from B6 and G2A-/- mice were harvested and
nuclear and cytosolic extracts were collected using the NE-PER kit (Pierce) according to
manufacturer’s instructions. Immunoblotting was performed as previously described 8.
Anti-NFκB p65 antibody was used at 1:2000 dilution. HRP-conjugated anti-rabbit
secondary antibody (Amersham) was used at 1:4000 dilution.
NFκB translocation studies. MAEC were maintained as described above. NFκB p65
nuclear translocation studies were performed as previously described 9. A minimum of
200 cells were counted for each condition and cells were scored as positive or negative
for nuclear p65.
Transfections. Primary MAEC were transfected using a Nucleofector II (Amaxa
Technologies) using the T23 setting and the basic endothelial transfection kit (Amaxa)
according to manufacturer’s instructions. Transfection rates of MAEC were 60-65% of
cells (see Figure 5A). Control vector pEXV3 and a plasmid containing the full-length
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
murine G2A cDNA (G2ApEXV3) were kind gifts of Dr. Janus Kabarowski (University of
Alabama Birmingham). For NFκB inhibition studies, MAEC were transfected with 500ng
of a ‘nonphosphorylatable’ form of IκBα, termed ‘super-repressor IκB’, plasmid which
was obtained from Dr. Julie Reuther (University of North Carolina). Cells were used in
experiments at 24h after transfection. For luciferase studies, NFκB-luc (Stratagene)
was co-transfected with pEXV3 or G2ApEXV3. pTK-Renilla was also co-transfected for
normalization. Luciferase and renilla expression were analyzed on a Berthold Sirius
luminometer.
Conventional and Quantitative real-time PCR. MAEC were freshly isolated from
aorta and cultured as described above for mRNA measurements. Total cellular RNA
was obtained from MAEC as previously described 10. For measurement of G2A
receptor mRNA abundance, 2 µl cDNA from each experimental group were utilized.
Primer sequences used were as follows: G2A: sense 5’-
GGTGAAACGCAGGTAGTGGT -3’ antisense 5’-CTGGTGGTGGTGTACAGTGC-3’,
and β-actin: sense 5’-CATGTTTGAGACCTTCAACAC-3’ antisense 5’-
CTGCTTGCTGATCCACATCT-3’. Conventional PCR conditions for G2A and β-actin
were as follows: 94°C 2min, followed by 30 cycles of 94°C 30 sec, 55°C 30 sec, 72°C
45 sec, with a final extension time of 5 min at 72°C. Bands were analyzed on a 1.0%
agarose gel in 1XTAE. PCR for β-actin was performed as a control.
For quantitative PCR analysis of chemokine and adhesion molecule mRNA
expression, cDNA was obtained as described above, and was diluted 1:8; 4 µL of this
dilution were used for each PCR reaction. Reagents from the BioRad real-time PCR kit
containing Sybr Green were used for quantitative PCR reactions. Primers for murine
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
ICAM-1 are F: 5’-AGATCACATTCACGGTGCTG-3’ and R: 5’-
CTTCAGAGGCAGGAAACAGG-3’; VCAM-1 are F: 5’-TCAAAGAAAGGGAGACTG-3’
and R: 5’-GCTGGAGAACTTCATTATC-3’; MCP-1 are F: 5’-
ATCCCAATGAGTAGGCTGGA-3’ and R: 5’-TCTGGACCCATTCCTTCTTG-3’; IL-6 are
F: 5’-TCCATCCAGTTGCCTTCTTG-3’ and R: 5’-TTTCTCATTTCCACGATTTCCC-3’;
KC are F: 5’-CTTGAAGGTGTTGCCCTCAG-3’ and R: 5’-
TGGGGACACCTTTTAGCATC-3’. Primers for murine cyclophilin are: F: 5’-
TGGAGAGCACCAAGACAGACA-3’ and R: 5’-TGCCGGAGTCGACAATGAT-3’. The
PCR conditions were: 95C 10 min, 95C 4 min, followed by 40 cycles of 95C 15 secs,
55C 30 secs, 72C 30 secs, followed by a final extension at 81C for 15 secs. Data were
analyzed and presented based upon the relative expression method 11. This formula for
calculation is:
Relative expression = 2-(S∆CT- C∆CT)
where ∆CT is the difference in threshold cycle between the gene of interest (i.e., ICAM-
1) and the housekeeping gene (cyclophilin). In this equation, S= G2A-/- mouse and C=
B6 control mouse.
Whole aorta flow cytometry. FACs analysis was performed on whole aorta as
previously desribed 12. Briefly, perfused aortas were excised, cleaned, and digested in
125 U/ml collagenase type XI, 60 U/ml hyaluronidase type I-s, 60 U/ml DNase1, and
450 U/ml collagenase type I in PBS containing 20 mM Hepes at 37°C for 1 h. A cell
suspension was obtained by forcing the aorta digest through a 70-µm strainer. Cells
were incubated with antibodies for 20 min at 4°C, washed twice, and analyzed by flow
cytometry on a CyAn™ ADP LX. Data analysis was performed using FlowJo software.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
Flow cytometry. MAEC at passage 3 were collected in PBS containing 1% FAFBSA
using Accutase (Fisher). 150,000 cells per sample were analyzed for each antibody.
Cells were incubated for 30 min at 4C with either PE-labeled antibodies for mouse
ICAM-1 and E-selectin or FITC-labeled antibodies for P-selectin and VCAM-1 as
described previously 13. To gate on live cells, 7-AAD (BD Biosciences) was used as a
viability dye according to manufacturer’s instructions. Samples were analyzed at the
University of Virginia Flow Cytometry Core using a Becton Dickinson FACSCalibur™
instrument. Analyses were performed using FL1 or FL2 channel histograms from
which the geometric mean fluorescence intensity (MFI) channel was calculated for each
sample.
Bone marrow transplants. C57Bl6J (B6) and G2A-/- mice were γ-irradiated twice and
injected with 6.0x106 bone marrow cells isolated from the femur and tibia of B6 or G2A-
/- as previously described 14. Irradiation and reconstitution of bone marrow was
confirmed by RTPCR for G2A expression. After 6 weeks, aortas were harvested and
used in the described ex vivo monocyte adhesion assay.
RhoA activation assay. RhoA activation was measured using the G-Lisa kit from
Cytoskeleton according to the manufacturer’s instructions. 50µg EC protein lysate was
used per well and measured in triplicate for each treatment. OD490 was read using a
Molecular Devices spectrophotometer.
Immunoblotting for G2A. Cells were harvested and lysed in lysis buffer containing
20mM TrisHCl (pH7.5), 2mM 2-Mercaptoethanol, 5mM EGTA, 2mM EDTA, and
protease inhibitors. After centrifugation, cytosol extract supernatant was collected and
resulting pellet was resuspended in 1x lysis buffer containing 50mM TrisHCl (pH8.0),
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
150mM NaCl, 1% NP40, 10mM NaF, 2mM Na3VO4, and protease inhibitors. After
centrifugation, membrane extract supernatant was collected. 100 µg of MAEC
membrane lysate protein was analyzed by 4-12% SDS-PAGE in MOPS running buffer
and transferred to nitrocellulose. Pierce Blocker BLOTTO in TBS was used as a
blocking agent. Membranes were probed with a 1:1000 dilution of G2A mouse
monoclonal antibody and a 1:4000 dilution of anti-mouse IgG-HRP (Amersham). Blots
were stripped and reprobed with 1:4000 tubulin and 1:10,000 anti-mouse IgG-HRP
secondary and quantitated using densitometry.
Inflammatory cytokines. Plasma from 6 each of C57Bl/6J and G2A-/- mice was
obtained as described above. Plasma was quantitatively measured for interleukin-6 (IL-
6), interleukin-10 (IL-10), monocyte chemoattractant protein-1 (MCP-1), interferon- γ
(IFN-γ), TNFα, and interleukin-12p70 (IL-12p70) using a Cytometric Bead Array
according to manufacturer’s instructions (BDBiosciences). For MAEC, 24h after
changing media to DMEM + 1% HIFBS, EC supernatant was collected from both mouse
groups. The collected supernatant was quantitatively measured for interleukin-6 (IL-6),
MCP-1, and KC by ELISA (R&DSystems), or IL-6 and MCP-1 by cytometric bead array
(BDBiosciences) according to the manufacturer’s instructions. Triplicate plates were
performed for each measurement and values were normalized to total cellular protein.
Measuring macrophage accumulation in aorta. Twelve CX3CR1-GFP mice were
injected with 2mL 3% thioglycollate media. Macrophages from these mice express GFP
1. Peritoneal macrophages from CX3CR1-GFP mice were collected 5d post-injection in
PBS and were pooled. Macrophages were washed once and resuspended in sterile
saline. Three B6 and 3 G2A-/- recipient mice were injected with 1mL saline containing
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
2x107 CX3CR1-GFP macrophages. At 4d post-injection, mice were euthanized,
perfused with PBS, and aortas harvested. Aortas from each group were pooled,
homogenized, and lysed in RIPA buffer. 100µg aorta protein was analyzed by SDS
PAGE for GFP using 1:1000 anti-GFP antibody and 1:4000 anti-mouse secondary
antibody (Amersham). Blots were stripped and reprobed for tubulin as a loading
control.
To stain for GFP localization in the aortic wall, 5-µm paraffin-embedded aorta
sections from recipient mice described above were cut and processed as previously
described15. Sections were stained with a biotinylated polyclonal anti-GFP antibody
(Vector, BA-0702) using a 1:500 dilution.
Statistical Analyses: Data for all experiments were analyzed using the StatView 6.0
software program. Comparisons between groups were performed using one-way
analysis of variance (ANOVA) methods. Data are graphically represented as mean +
SE, in which each mean consists of 4 experiments performed in triplicate (unless noted
otherwise in the figure legends) using 3-6 mice per group. Comparisons
between groups and tests of interactions were made assuming a two-factor analysis
with the interaction term testing each main effect with the residual error testing the
interaction. All comparisons were made using Fisher’s LSD procedure, so that multiple
comparisons were made at the 0.05 level only if the overall F- test from the ANOVA was
significant at p<0.05.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
Reference List
1. Jung S, Aliberti J, Graemmel P, Sunshine MJ, Kreutzberg GW, Sher A, Littman DR. Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol Cell Biol. 2000;20:4106-4114.
2. Le LQ, Kabarowski JH, Weng Z, Satterthwaite AB, Harvill ET, Jensen ER, Miller JF, Witte ON. Mice lacking the orphan G protein-coupled receptor G2A develop a late-onset autoimmune syndrome. Immunity. 2001;14:561-571.
3. Tsao PS, McEvoy LM, Drexler H, Butcher EC, Cooke JP. Enhanced Endothelial Adhesiveness in Hpyercholesterolemia Is Attenuated by L-Arginine. Circulation. 1994;89:2176-2182.
4. Bolick DT, Srinivasan S, Kim KW, Hatley ME, Clemens JJ, Whetzel A, Ferger N, Macdonald TL, Davis MD, Tsao PS, Lynch KR, Hedrick CC. Sphingosine-1-phosphate prevents tumor necrosis factor-{alpha}-mediated monocyte adhesion to aortic endothelium in mice. Arterioscler Thromb Vasc Biol. 2005;25:976-981.
5. Srinivasan S, Hatley ME, Reilly KB, Danziger EC, Hedrick CC. Modulation of PPAR{alpha} Expression and Inflammatory Interleukin-6 Production by Chronic Glucose Increases Monocyte/Endothelial Adhesion. Arterioscler Thromb Vasc Biol. 2004;851-857.
6. Ramos CL, Huo Y, Jung U, Ghosh S, Manka DR, Sarembock IJ, Ley K. Direct demonstration of P-selectin- and VCAM-1-dependent mononuclear cell rolling in early atherosclerotic lesions of apolipoprotein E- deficient mice. Circ Res. 1999;84:1237-1244.
7. Zakrzewicz A, Grafe M, Terbeek D, Bongrazio M, Auch-Schwelk W, Walzog B, Graf K, Fleck E, Ley K, Gaehtgens P. L-selectin-dependent leukocyte adhesion to microvascular but not to macrovascular endothelial cells of the human coronary system. Blood. 1997;89:3228-3235.
8. Orr AW, Pallero MA, Murphy-Ullrich JE. Thrombospondin stimulates focal adhesion disassembly through Gi- and phosphoinositide 3-kinase-dependent ERK activation. J Biol Chem. 2002;277:20453-20460.
9. Bolick DT, Orr AW, Whetzel A, Srinivasan S, Hatley ME, Schwartz MA, Hedrick CC. 12/15-lipoxygenase regulates intercellular adhesion molecule-1 expression and monocyte adhesion to endothelium through activation of RhoA and nuclear factor-kappaB. Arterioscler Thromb Vasc Biol. 2005;25:2301-2307.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Online Supplement Bolick et al. G2A and Monocyte Adhesion to Aortic Endothelium
10. Bolick DT, Srinivasan S, Whetzel A, Fuller LC, Hedrick CC. 12/15 Lipoxygenase Mediates Monocyte Adhesion to Aortic Endothelium in Apolipoprotein E-Deficient Mice Through Activation of RhoA and NF-{kappa}B. Arterioscler Thromb Vasc Biol. 2006;26:1260-6.
11. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402-408.
12. Galkina E, Kadl A, Sanders J, Varughese D, Sarembock IJ, Ley K. Lymphocyte recruitment into the aortic wall before and during development of atherosclerosis is partially L-selectin dependent. J Exp Med. 2006;203:1273-1282.
13. Hatley ME, Srinivasan S, Reilly KB, Bolick DT, Hedrick CC. Increased Production of 12/15 Lipoxygenase Eicosanoids Accelerates Monocyte/Endothelial Interactions in Diabetic db/db Mice. J Biol Chem. 2003;278:25369-25375.
14. Day YJ, Huang L, McDuffie MJ, Rosin DL, Ye H, Chen JF, Schwarzschild MA, Fink JS, Linden J, Okusa MD. Renal protection from ischemia mediated by A2A adenosine receptors on bone marrow-derived cells. J Clin Invest. 2003;112:883-891.
15. Orr AW, Sanders JM, Bevard M, Coleman E, Sarembock IJ, Schwartz MA. The subendothelial extracellular matrix modulates NF-kappaB activation by flow: a potential role in atherosclerosis. J Cell Biol. 2005;169:191-202.
by on May 18, 2011 circres.ahajournals.orgDownloaded from
Related Documents











