Absence of APOBEC-1 mediated mRNA editing in human carcinomas Jobst Greeve* ,1 , Heinrich Lellek 1 , Frank Apostel 1 , Katja Hundoegger 1 , Akbar Barialai 1 , Romy Kirsten 1 , Sybille Welker 1 and Heiner Greten 1 1 Medizinische Kernklinik und Poliklinik, Universita ¨ts-Krankenhaus Eppendorf, Martinistraße 52, D-20246 Hamburg, Germany The transgene expression of the catalytic subunit APOBEC-1 of the apo B mRNA editing enzyme- complex can cause hepatocellular carcinoma in mice and rabbits. It has been proposed that aberrant editing of mRNA may represent a novel oncogenic principle. This investigation aimed to define whether such aberrant hyperediting mediated by APOBEC-1 occurs in human carcinomas. Editing and hyperediting of apo B, NAT1 or NF1 mRNA was not identified in any of 28 resected tumor specimens, including hepatocellular, bile duct, gastric, colorectal, pancreatic adeno- and neuroendo- crine, lung adeno-, medullary thyroid and breast carcinoma, soft tissue sarcoma and neuroblastoma. In most types of carcinoma, significant levels for full-length APOBEC-1 mRNA could not be detected. Low level expression of APOBEC-1 was found in colorectal and gastric carcinoma where most of the APOBEC-1 mRNA is inactivated by alternate splicing. The ‘auxiliary’ components of the apo B mRNA editing enzyme-complex are missing in many tumors including colorectal and gastric carcinoma, but are highly expressed in hepato- cellular, lung adeno- and breast carcinoma all of which lack APOBEC-1. Taken together, either APOBEC-1 or the ‘auxiliary’ components of the apo B mRNA editing enzyme-complex or both are missing in human carcino- mas resulting in the absence of mRNA editing. Currently, there is no evidence that aberrant editing mediated by APOBEC-1 contributes to the tumorigen- esis of natural human carcinomas. Keywords: mRNA editing; APOBEC-1; human carci- noma; apo B; NAT1 Introduction mRNA editing, the specific posttranscriptional base change in mRNA, is a genetic principle that has attracted a lot of attention in recent years (Simpson and Thiemann, 1995; Scott, 1995). Besides the various forms of editing for RNAs encoded in the mitochondria of plants or kimetoplastid protozoa, two paradigms for mRNA editing of nuclear encoded genes have been described, editing of apolipoprotein (apo) B mRNA and editing of mRNAs for certain subunits of glutamate-gated cation channels (GluRs) (Powell et al., 1987; Sommer et al., 1991). Editing of GluR transcripts changes genomically encoded adenosines into in- osines altering amino acid residues at functional important domains of the encoded ion channels (Mass et al., 1997). This editing is mediated by double-stranded RNA-dependent adenosine deami- nases of which so far three dierent forms have been cloned (Maas et al., 1997). Editing from cytidine to uridine at nucleotide position 6666 in the apo B mRNA creates a premature translational stop codon and leads to the generation of the carboxyterminal truncated apo B-48 (Chen et al., 1987; Powell et al., 1987). In humans and many other mammalian species, the apo B mRNA is completely edited in the intestine but remains unedited in the liver (Greeve et al., 1993). Besides in the small intestine, the apo B mRNA is edited in the human stomach, colon and kidney (Teng et al., 1990). In some mammalian species (dog, horse, rat, mouse), also the hepatic apo B mRNA is edited and the hepatic synthesis of apo B-48 results in low plasma concentrations of the atherogenic LDL that are derived exclusively from apo B-100 containing VLDL (Greeve et al., 1993). The importance of hepatic apo B mRNA editing for plasma lipoprotein concentrations is further demonstrated by mice deficient in apo B mRNA editing and in the LDL receptor which have extremely high LDL plasma levels, are very susceptible to atherosclerosis and closely resemble human familial hypercholesterolemia (Powell-Braxton et al., 1998). The editing of apo B mRNA is mediated by a multicomponent enzyme-complex, designated apo B mRNA editing enzyme (Greeve et al., 1991; Smith et al., 1991). The catalytic subunit APOBEC-1 (apo B mRNA editing enzyme catalytic polypeptide 1) is the only component of this enzyme-complex that has been conclusively characterized (Teng et al., 1993). APOBEC-1 is a member of the cytidine deaminase gene family with a novel RNA binding motif and requires additional ‘auxiliary’ components for ecient editing (Navaratnam et al., 1993, 1995; Yamanaka et al., 1995; Anant et al., 1995). APOBEC-1 is the only component of the editing enzyme-complex missing in the human or rabbit liver (Giannoni et al., 1994; Greeve et al., 1996). In mouse and rat, APOBEC-1 is expressed in the liver as the result of a second promoter which leads to tissue-specific exon use and alternate splicing in the 5’ untranslated region of the APOBEC-1 mRNA (Nakamuta et al., 1995; Qian et al., 1997; Greeve et al., 1998). Adenovirus-mediated gene transfer of APOBEC-1 into the liver of normal or LDL-receptor deficient Watanabe heritable hyper- *Correspondence: J Greeve Received 3 March 1999; revised 18 June 1999; accepted 23 June 1999 Oncogene (1999) 18, 6357 – 6366 ª 1999 Stockton Press All rights reserved 0950 – 9232/99 $15.00 http://www.stockton-press.co.uk/onc

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Absence of APOBEC-1 mediated mRNA editing in human carcinomas
Jobst Greeve*,1, Heinrich Lellek1, Frank Apostel1, Katja Hundoegger1, Akbar Barialai1,Romy Kirsten1, Sybille Welker1 and Heiner Greten1
1Medizinische Kernklinik und Poliklinik, UniversitaÈts-Krankenhaus Eppendorf, Martinistraûe 52, D-20246 Hamburg, Germany
The transgene expression of the catalytic subunitAPOBEC-1 of the apo B mRNA editing enzyme-complex can cause hepatocellular carcinoma in miceand rabbits. It has been proposed that aberrant editing ofmRNA may represent a novel oncogenic principle. Thisinvestigation aimed to de®ne whether such aberranthyperediting mediated by APOBEC-1 occurs in humancarcinomas. Editing and hyperediting of apo B, NAT1 orNF1 mRNA was not identi®ed in any of 28 resectedtumor specimens, including hepatocellular, bile duct,gastric, colorectal, pancreatic adeno- and neuroendo-crine, lung adeno-, medullary thyroid and breastcarcinoma, soft tissue sarcoma and neuroblastoma. Inmost types of carcinoma, signi®cant levels for full-lengthAPOBEC-1 mRNA could not be detected. Low levelexpression of APOBEC-1 was found in colorectal andgastric carcinoma where most of the APOBEC-1 mRNAis inactivated by alternate splicing. The `auxiliary'components of the apo B mRNA editing enzyme-complexare missing in many tumors including colorectal andgastric carcinoma, but are highly expressed in hepato-cellular, lung adeno- and breast carcinoma all of whichlack APOBEC-1. Taken together, either APOBEC-1 orthe `auxiliary' components of the apo B mRNA editingenzyme-complex or both are missing in human carcino-mas resulting in the absence of mRNA editing.Currently, there is no evidence that aberrant editingmediated by APOBEC-1 contributes to the tumorigen-esis of natural human carcinomas.
Keywords: mRNA editing; APOBEC-1; human carci-noma; apo B; NAT1
Introduction
mRNA editing, the speci®c posttranscriptional basechange in mRNA, is a genetic principle that hasattracted a lot of attention in recent years (Simpsonand Thiemann, 1995; Scott, 1995). Besides thevarious forms of editing for RNAs encoded in themitochondria of plants or kimetoplastid protozoa,two paradigms for mRNA editing of nuclearencoded genes have been described, editing ofapolipoprotein (apo) B mRNA and editing ofmRNAs for certain subunits of glutamate-gatedcation channels (GluRs) (Powell et al., 1987;
Sommer et al., 1991). Editing of GluR transcriptschanges genomically encoded adenosines into in-osines altering amino acid residues at functionalimportant domains of the encoded ion channels(Mass et al., 1997). This editing is mediated bydouble-stranded RNA-dependent adenosine deami-nases of which so far three di�erent forms havebeen cloned (Maas et al., 1997).
Editing from cytidine to uridine at nucleotideposition 6666 in the apo B mRNA creates apremature translational stop codon and leads tothe generation of the carboxyterminal truncated apoB-48 (Chen et al., 1987; Powell et al., 1987). Inhumans and many other mammalian species, theapo B mRNA is completely edited in the intestinebut remains unedited in the liver (Greeve et al.,1993). Besides in the small intestine, the apo BmRNA is edited in the human stomach, colon andkidney (Teng et al., 1990). In some mammalianspecies (dog, horse, rat, mouse), also the hepaticapo B mRNA is edited and the hepatic synthesis ofapo B-48 results in low plasma concentrations ofthe atherogenic LDL that are derived exclusivelyfrom apo B-100 containing VLDL (Greeve et al.,1993). The importance of hepatic apo B mRNAediting for plasma lipoprotein concentrations isfurther demonstrated by mice de®cient in apo BmRNA editing and in the LDL receptor whichhave extremely high LDL plasma levels, are verysusceptible to atherosclerosis and closely resemblehuman familial hypercholesterolemia (Powell-Braxtonet al., 1998).
The editing of apo B mRNA is mediated by amulticomponent enzyme-complex, designated apo BmRNA editing enzyme (Greeve et al., 1991; Smith etal., 1991). The catalytic subunit APOBEC-1 (apo BmRNA editing enzyme catalytic polypeptide 1) is theonly component of this enzyme-complex that hasbeen conclusively characterized (Teng et al., 1993).APOBEC-1 is a member of the cytidine deaminasegene family with a novel RNA binding motif andrequires additional `auxiliary' components for e�cientediting (Navaratnam et al., 1993, 1995; Yamanaka etal., 1995; Anant et al., 1995). APOBEC-1 is the onlycomponent of the editing enzyme-complex missing inthe human or rabbit liver (Giannoni et al., 1994;Greeve et al., 1996). In mouse and rat, APOBEC-1 isexpressed in the liver as the result of a secondpromoter which leads to tissue-speci®c exon use andalternate splicing in the 5' untranslated region of theAPOBEC-1 mRNA (Nakamuta et al., 1995; Qian etal., 1997; Greeve et al., 1998). Adenovirus-mediatedgene transfer of APOBEC-1 into the liver of normalor LDL-receptor de®cient Watanabe heritable hyper-
*Correspondence: J GreeveReceived 3 March 1999; revised 18 June 1999; accepted 23 June 1999
Oncogene (1999) 18, 6357 ± 6366ã 1999 Stockton Press All rights reserved 0950 ± 9232/99 $15.00
http://www.stockton-press.co.uk/onc

lipidemic (WHHL) rabbits induces editing of thehepatic apo B mRNA and results in a drasticreduction of circulating LDL levels (Greeve et al.,1996).
In APOBEC-1 transgenic animals, the hepaticoverexpression of APOBEC-1 mediated by thepromoter of apolipoprotein E causes hepatocellulardysplasia and carcinomas (Yamanaka et al., 1995).The overexpression of APOBEC-1 leads to addi-tional `hyperediting' of multiple cytidines down-stream of the natural editing site C6666 in the apoB mRNA, and to aberrant `hyperediting' of cytidineresidues in other mRNAs (Yamanaka et al., 1996,1997; Sowden et al., 1996). One of these othermRNAs, NAT1 (novel APOBEC-1 target 1), wasidenti®ed by the `mooring' motif in the apo BmRNA required for editing by APOBEC-1(Yamanaka et al., 1997; Shah et al., 1991). NAT1has homology to the carboxy-terminal portion ofthe eukaryotic translation initiation factor eIF4G,and most likely is a translational repressor(Yamanaka et al., 1997). Hyperediting of NAT1mRNA leads to a loss of NAT1 function, whichwas postulated to contribute to the oncogenesis inthe APOBEC-1 transgenic animals (Yamanaka etal., 1997). Further evidence that mRNA editingmay be involved in tumorigenesis comes fromstudies in malignant neuro®bromas of patients withneuro®bromatosis type 1 (NF1) (Cappione et al.,1997). The mRNA of the NF1 gene has beendemonstrated to be edited from C to U atnucleotide position 2914, creating a premature stoptranslation codon (Skuse et al., 1996). The editingof NF1 mRNA was much higher in neurofibroma-tous tumors as compared to normal tissues and itwas concluded that mRNA editing may contributeto the genetic inactivation of the NF1 gene in NF1malignancies (Cappione et al., 1997). Whether theediting of NF1 is mediated by APOBEC-1 togetherwith novel trans-acting factors remains to beconclusively established (Skuse et al., 1996; Cap-pione et al., 1997). The third line of evidence thatmRNA editing may be involved in oncogenesis isderived from an investigation of the Wilms tumorsusceptibility gene WT1 (Sharma et al., 1994). ThemRNA of WT1 can be edited from U to C atnucleotide position 839, changing codon 280 fromCTC for leucine into CCC for proline (Sharma etal., 1994). The editing of WT1 mRNA is tissue-speci®c, temporally controlled and functionallyimportant as the edited form of WT1 protein isless e�cient in transcriptional repression than theunedited form (Sharma et al., 1994).
This study addressed the question as to whetherAPOBEC-1 mediates potential oncogenic hyperedit-ing in human carcinomas. Out of 28 resected tumorspecimens, no apo B mRNA editing or aberranthyperediting of apo B, NAT1 or NF1 mRNAcould be identi®ed. APOBEC-1 was found to beeither not expressed or only at very low levels. Thecarcinomas which express low levels of APOBEC-1demonstrated a loss of the `auxiliary' componentsrequired for mRNA editing. Thus, there is noevidence that mRNA editing by APOBEC-1represents an oncogenic principle in human carci-nomas.
Results
Expression of APOBEC-1 and apo B mRNA in humancarcinomas
APOBEC-1 mRNA could not be detected by primaryRT ±PCR in any of 28 individual tumor specimens ofprimary or metastatic carcinomas. Nested PCR,however, detected APOBEC-1 mRNA in all colorectalcarcinomas studied, in one out of three hepatocellularcarcinomas, in two out of three gastric carcinomas, inone of two pancreatic adenocarcinomas and in one oftwo lung adenocarcinomas. APOBEC-1 mRNA wasnot detectable by nested RT±PCR in bile duct, breast,medullary thyroid or pancreatic neuroendocrinecarcinoma, and undetectable in two out of the threehepatocellular carcinomas, in one of the three gastriccarcinomas, in one of the two pancreatic and in one ofthe two lung adenocarcinomas.
Primary RT±PCR generated a clearly visibleproduct for apo B mRNA in ®ve colorectalcarcinomas, in all three hepatocellular carcinomas, inboth bile duct carcinomas, in both breast carcinomas,in one of the three gastric carcinomas, in theneuroendocrine carcinoma and in the medullarythyroid carcinoma. Nested RT±PCR identi®ed apo BmRNA in four additional colorectal carcinomas and inthe neuroblastoma. Altogether, 20 out of 28 tumorswere positive for apo B mRNA using primary ornested RT±PCR.
Apo B mRNA editing in human carcinomas
The 20 RT±PCR products for apo B mRNA wereanalysed by primer extension assay for editing at thenatural editing site C6666 (Figure 1). Substantialamount of editing (41%) at this site C6666 could notbe detected in any of the 20 tumors. The apo B RT±PCR products of two colorectal carcinoma werecloned and individual clones were analysed by primerextension assay. All 80 and 83 cDNA clones,respectively, were unedited and contained the genomi-cally encoded C at nucleotide position 6666. The apoB mRNA of two colorectal, two hepatocellular andone gastric carcinoma was ampli®ed from nucleotideposition 6552 to 6864 by RT±PCR and cloned.Twelve individual clones of each tumor weresequenced: No sequence divergence could be detectedin these 60 apo B cDNA clones. Notably, thepotential sites (C6675, C6699, C6707, C6718, C6738, C6743,C6762, C6782, C6783, C6802, C6806, C6807, C6815, C6833, C6840 andC6851) for aberrant hyperediting of hepatic apo BmRNA as identi®ed in the APOBEC-1 transgenicanimals (Yamanaka et al., 1996) were entirelyunedited in these ®ve human carcinomas.
The NAT1 mRNA of the same ®ve tumors wasampli®ed by RT±PCR from nucleotide position2260 ± 2758, cloned and sequenced. This region of theNAT1 mRNA contains 28 presumptive editing sites asidenti®ed in APOBEC-1 transgenic animals (Yamana-ka et al., 1997). At four of these sites, C/U editing cancreate a premature stop translation codon. No C/Ubase heterogeneity at any of these sites could bedetected in a total number of 36 NAT1 cDNA clones,all of which contained the unedited cDNA sequence asreported by Yamanaka et al. (1997).
Absence of mRNA editing in human carcinomasJ Greeve et al
6358
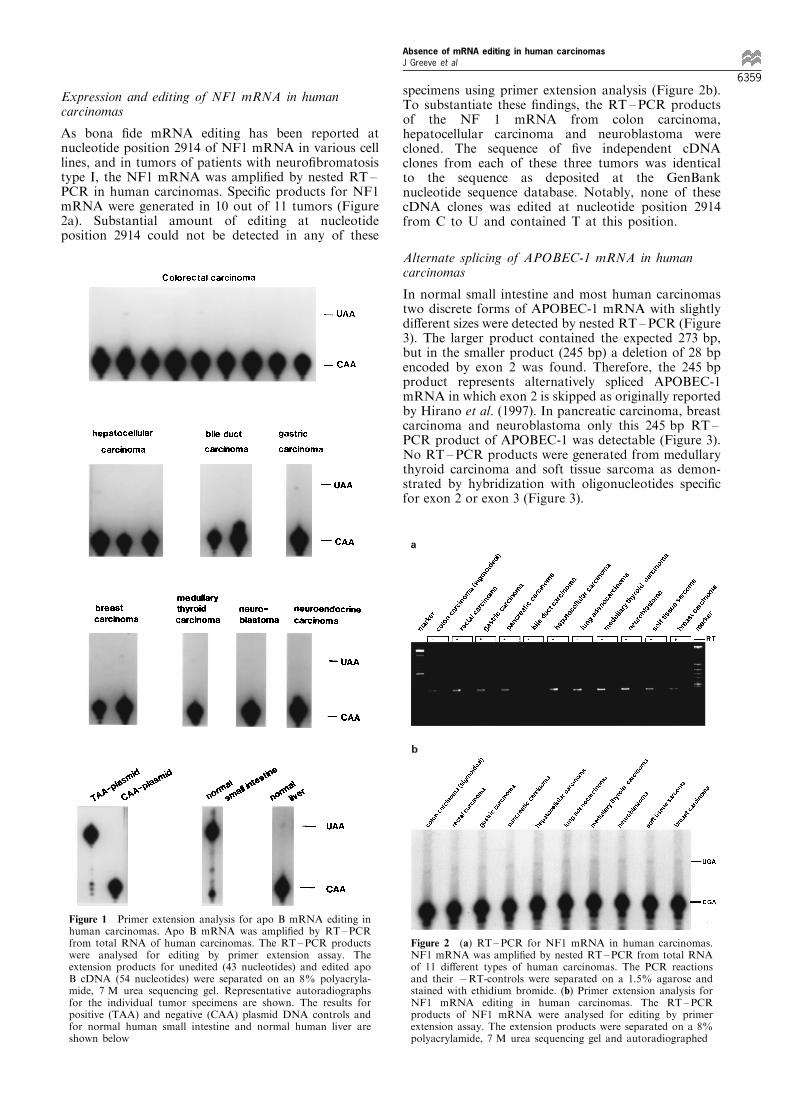
Expression and editing of NF1 mRNA in humancarcinomas
As bona ®de mRNA editing has been reported atnucleotide position 2914 of NF1 mRNA in various celllines, and in tumors of patients with neuro®bromatosistype I, the NF1 mRNA was ampli®ed by nested RT ±PCR in human carcinomas. Speci®c products for NF1mRNA were generated in 10 out of 11 tumors (Figure2a). Substantial amount of editing at nucleotideposition 2914 could not be detected in any of these
specimens using primer extension analysis (Figure 2b).To substantiate these ®ndings, the RT ±PCR productsof the NF 1 mRNA from colon carcinoma,hepatocellular carcinoma and neuroblastoma werecloned. The sequence of ®ve independent cDNAclones from each of these three tumors was identicalto the sequence as deposited at the GenBanknucleotide sequence database. Notably, none of thesecDNA clones was edited at nucleotide position 2914from C to U and contained T at this position.
Alternate splicing of APOBEC-1 mRNA in humancarcinomas
In normal small intestine and most human carcinomastwo discrete forms of APOBEC-1 mRNA with slightlydi�erent sizes were detected by nested RT±PCR (Figure3). The larger product contained the expected 273 bp,but in the smaller product (245 bp) a deletion of 28 bpencoded by exon 2 was found. Therefore, the 245 bpproduct represents alternatively spliced APOBEC-1mRNA in which exon 2 is skipped as originally reportedby Hirano et al. (1997). In pancreatic carcinoma, breastcarcinoma and neuroblastoma only this 245 bp RT±PCR product of APOBEC-1 was detectable (Figure 3).No RT±PCR products were generated from medullarythyroid carcinoma and soft tissue sarcoma as demon-strated by hybridization with oligonucleotides speci®cfor exon 2 or exon 3 (Figure 3).
Figure 1 Primer extension analysis for apo B mRNA editing inhuman carcinomas. Apo B mRNA was ampli®ed by RT±PCRfrom total RNA of human carcinomas. The RT±PCR productswere analysed for editing by primer extension assay. Theextension products for unedited (43 nucleotides) and edited apoB cDNA (54 nucleotides) were separated on an 8% polyacryla-mide, 7 M urea sequencing gel. Representative autoradiographsfor the individual tumor specimens are shown. The results forpositive (TAA) and negative (CAA) plasmid DNA controls andfor normal human small intestine and normal human liver areshown below
a
b
Figure 2 (a) RT±PCR for NF1 mRNA in human carcinomas.NF1 mRNA was ampli®ed by nested RT±PCR from total RNAof 11 di�erent types of human carcinomas. The PCR reactionsand their 7RT-controls were separated on a 1.5% agarose andstained with ethidium bromide. (b) Primer extension analysis forNF1 mRNA editing in human carcinomas. The RT±PCRproducts of NF1 mRNA were analysed for editing by primerextension assay. The extension products were separated on a 8%polyacrylamide, 7 M urea sequencing gel and autoradiographed
Absence of mRNA editing in human carcinomasJ Greeve et al
6359
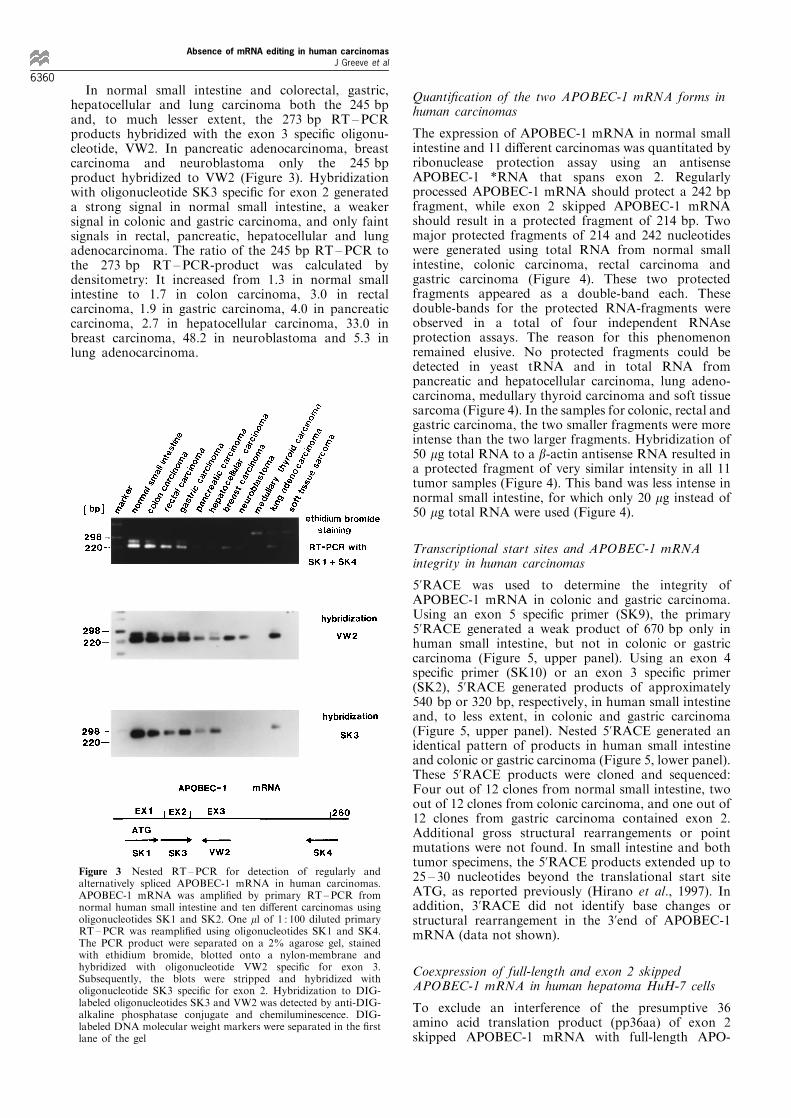
In normal small intestine and colorectal, gastric,hepatocellular and lung carcinoma both the 245 bpand, to much lesser extent, the 273 bp RT ±PCRproducts hybridized with the exon 3 speci®c oligonu-cleotide, VW2. In pancreatic adenocarcinoma, breastcarcinoma and neuroblastoma only the 245 bpproduct hybridized to VW2 (Figure 3). Hybridizationwith oligonucleotide SK3 speci®c for exon 2 generateda strong signal in normal small intestine, a weakersignal in colonic and gastric carcinoma, and only faintsignals in rectal, pancreatic, hepatocellular and lungadenocarcinoma. The ratio of the 245 bp RT±PCR tothe 273 bp RT ±PCR-product was calculated bydensitometry: It increased from 1.3 in normal smallintestine to 1.7 in colon carcinoma, 3.0 in rectalcarcinoma, 1.9 in gastric carcinoma, 4.0 in pancreaticcarcinoma, 2.7 in hepatocellular carcinoma, 33.0 inbreast carcinoma, 48.2 in neuroblastoma and 5.3 inlung adenocarcinoma.
Quanti®cation of the two APOBEC-1 mRNA forms inhuman carcinomas
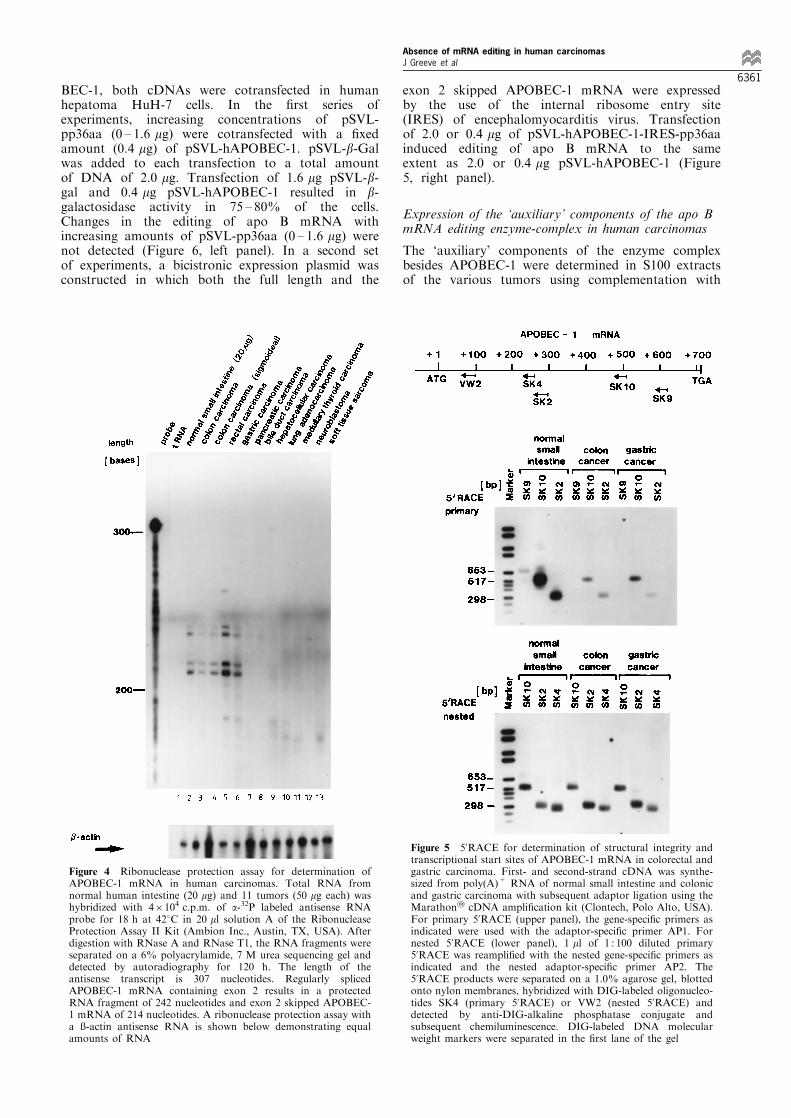
The expression of APOBEC-1 mRNA in normal smallintestine and 11 di�erent carcinomas was quantitated byribonuclease protection assay using an antisenseAPOBEC-1 *RNA that spans exon 2. Regularlyprocessed APOBEC-1 mRNA should protect a 242 bpfragment, while exon 2 skipped APOBEC-1 mRNAshould result in a protected fragment of 214 bp. Twomajor protected fragments of 214 and 242 nucleotideswere generated using total RNA from normal smallintestine, colonic carcinoma, rectal carcinoma andgastric carcinoma (Figure 4). These two protectedfragments appeared as a double-band each. Thesedouble-bands for the protected RNA-fragments wereobserved in a total of four independent RNAseprotection assays. The reason for this phenomenonremained elusive. No protected fragments could bedetected in yeast tRNA and in total RNA frompancreatic and hepatocellular carcinoma, lung adeno-carcinoma, medullary thyroid carcinoma and soft tissuesarcoma (Figure 4). In the samples for colonic, rectal andgastric carcinoma, the two smaller fragments were moreintense than the two larger fragments. Hybridization of50 mg total RNA to a b-actin antisense RNA resulted ina protected fragment of very similar intensity in all 11tumor samples (Figure 4). This band was less intense innormal small intestine, for which only 20 mg instead of50 mg total RNA were used (Figure 4).
Transcriptional start sites and APOBEC-1 mRNAintegrity in human carcinomas
5'RACE was used to determine the integrity ofAPOBEC-1 mRNA in colonic and gastric carcinoma.Using an exon 5 speci®c primer (SK9), the primary5'RACE generated a weak product of 670 bp only inhuman small intestine, but not in colonic or gastriccarcinoma (Figure 5, upper panel). Using an exon 4speci®c primer (SK10) or an exon 3 speci®c primer(SK2), 5'RACE generated products of approximately540 bp or 320 bp, respectively, in human small intestineand, to less extent, in colonic and gastric carcinoma(Figure 5, upper panel). Nested 5'RACE generated anidentical pattern of products in human small intestineand colonic or gastric carcinoma (Figure 5, lower panel).These 5'RACE products were cloned and sequenced:Four out of 12 clones from normal small intestine, twoout of 12 clones from colonic carcinoma, and one out of12 clones from gastric carcinoma contained exon 2.Additional gross structural rearrangements or pointmutations were not found. In small intestine and bothtumor specimens, the 5'RACE products extended up to25 ± 30 nucleotides beyond the translational start siteATG, as reported previously (Hirano et al., 1997). Inaddition, 3'RACE did not identify base changes orstructural rearrangement in the 3'end of APOBEC-1mRNA (data not shown).
Coexpression of full-length and exon 2 skippedAPOBEC-1 mRNA in human hepatoma HuH-7 cells
To exclude an interference of the presumptive 36amino acid translation product (pp36aa) of exon 2skipped APOBEC-1 mRNA with full-length APO-
Figure 3 Nested RT±PCR for detection of regularly andalternatively spliced APOBEC-1 mRNA in human carcinomas.APOBEC-1 mRNA was ampli®ed by primary RT±PCR fromnormal human small intestine and ten di�erent carcinomas usingoligonucleotides SK1 and SK2. One ml of 1 : 100 diluted primaryRT±PCR was reampli®ed using oligonucleotides SK1 and SK4.The PCR product were separated on a 2% agarose gel, stainedwith ethidium bromide, blotted onto a nylon-membrane andhybridized with oligonucleotide VW2 speci®c for exon 3.Subsequently, the blots were stripped and hybridized witholigonucleotide SK3 speci®c for exon 2. Hybridization to DIG-labeled oligonucleotides SK3 and VW2 was detected by anti-DIG-alkaline phosphatase conjugate and chemiluminescence. DIG-labeled DNA molecular weight markers were separated in the ®rstlane of the gel
Absence of mRNA editing in human carcinomasJ Greeve et al
6360
BEC-1, both cDNAs were cotransfected in humanhepatoma HuH-7 cells. In the ®rst series ofexperiments, increasing concentrations of pSVL-pp36aa (0 ± 1.6 mg) were cotransfected with a ®xedamount (0.4 mg) of pSVL-hAPOBEC-1. pSVL-b-Galwas added to each transfection to a total amountof DNA of 2.0 mg. Transfection of 1.6 mg pSVL-b-gal and 0.4 mg pSVL-hAPOBEC-1 resulted in b-galactosidase activity in 75 ± 80% of the cells.Changes in the editing of apo B mRNA withincreasing amounts of pSVL-pp36aa (0 ± 1.6 mg) werenot detected (Figure 6, left panel). In a second setof experiments, a bicistronic expression plasmid wasconstructed in which both the full length and the
exon 2 skipped APOBEC-1 mRNA were expressedby the use of the internal ribosome entry site(IRES) of encephalomyocarditis virus. Transfectionof 2.0 or 0.4 mg of pSVL-hAPOBEC-1-IRES-pp36aainduced editing of apo B mRNA to the sameextent as 2.0 or 0.4 mg pSVL-hAPOBEC-1 (Figure5, right panel).
Expression of the `auxiliary' components of the apo BmRNA editing enzyme-complex in human carcinomas
The `auxiliary' components of the enzyme complexbesides APOBEC-1 were determined in S100 extractsof the various tumors using complementation with
Figure 5 5'RACE for determination of structural integrity andtranscriptional start sites of APOBEC-1 mRNA in colorectal andgastric carcinoma. First- and second-strand cDNA was synthe-sized from poly(A)+ RNA of normal small intestine and colonicand gastric carcinoma with subsequent adaptor ligation using theMarathon2 cDNA ampli®cation kit (Clontech, Polo Alto, USA).For primary 5'RACE (upper panel), the gene-speci®c primers asindicated were used with the adaptor-speci®c primer AP1. Fornested 5'RACE (lower panel), 1 ml of 1 : 100 diluted primary5'RACE was reampli®ed with the nested gene-speci®c primers asindicated and the nested adaptor-speci®c primer AP2. The5'RACE products were separated on a 1.0% agarose gel, blottedonto nylon membranes, hybridized with DIG-labeled oligonucleo-tides SK4 (primary 5'RACE) or VW2 (nested 5'RACE) anddetected by anti-DIG-alkaline phosphatase conjugate andsubsequent chemiluminescence. DIG-labeled DNA molecularweight markers were separated in the ®rst lane of the gel
Figure 4 Ribonuclease protection assay for determination ofAPOBEC-1 mRNA in human carcinomas. Total RNA fromnormal human intestine (20 mg) and 11 tumors (50 mg each) washybridized with 46104 c.p.m. of a-32P labeled antisense RNAprobe for 18 h at 428C in 20 ml solution A of the RibonucleaseProtection Assay II Kit (Ambion Inc., Austin, TX, USA). Afterdigestion with RNase A and RNase T1, the RNA fragments wereseparated on a 6% polyacrylamide, 7 M urea sequencing gel anddetected by autoradiography for 120 h. The length of theantisense transcript is 307 nucleotides. Regularly splicedAPOBEC-1 mRNA containing exon 2 results in a protectedRNA fragment of 242 nucleotides and exon 2 skipped APOBEC-1 mRNA of 214 nucleotides. A ribonuclease protection assay witha û-actin antisense RNA is shown below demonstrating equalamounts of RNA
Absence of mRNA editing in human carcinomasJ Greeve et al
6361

recombinant APOBEC-1. Extracts of baculovirus-infected Sf9 cells expressing APOBEC-1 did notexhibit in vitro editing activity (Figure 7, lane 1).Addition of Hela-cell extracts containing `auxiliary'components but no apo B mRNA editing activityreconstituted apo B mRNA editing activity (Figure 8,lane 2 ± 4). None of the S100 extracts from tendi�erent carcinomas had endogenous apo B mRNAediting activity (Figure 7). After supplementation withrecombinant APOBEC-1, strong in vitro apo BmRNA editing activity was observed in the extractsof the lung adenocarcinoma, the breast carcinomaand of one of the two hepatocellular carcinomas(Figure 7). Less in vitro editing activity wasreconstituted in extracts from the other hepatocellu-lar carcinoma, from the rectal carcinoma and fromthe neuroblastoma (Figure 7). Addition of Sf9 cellextract containing APOBEC-1 did not generate invitro editing activity in extracts of colonic, sigmoideal,gastric and pancreatic carcinoma (Figure 7). Additionof increasing amounts of Hela-cell extracts containingabundant amounts of `auxiliary' components did notreconstitute in vitro apo B mRNA editing activity inany of the tumor extracts, including colorectal andgastric carcinoma (data not shown). Addition ofincreasing amounts of S100 extract from colorectalcarcinomas did not diminish the editing activity ofHela-extracts supplemented with recombinant APO-BEC-1, ruling out that colorectal carcinomas expressspeci®c inhibitors for mRNA editing (data notshown).
Discussion
The observation that the transgenic hepatic expressionof APOBEC-1 can cause hepatocellular carcinomademonstrated that APOBEC-1 can act as a potentoncogene (Yamanaka et al., 1995). This investigationaimed to establish whether aberrant mRNA editingmediated by APOBEC-1 may contribute to naturaltumorigenesis in human patients. Unexpectedly, editingof apo B, NAT1 or NF1 mRNA could not be detectedin 28 human carcinomas studied, and APOBEC-1 wasfound to be either not expressed or only at very lowlevels that were further inactivated by alternatesplicing. The `auxiliary' components of the apo BmRNA editing enzyme-complex besides APOBEC-1are expressed in some, but notably not in gastro-intestinal carcinomas that express APOBEC-1 todetectable levels. Complementation of extracts fromcolorectal or gastric carcinomas, with extracts fromHela-cells containing the `auxiliary' components,demonstrated that in gastrointestinal carcinomas theexpression of APOBEC-1 is not su�cient to promotemRNA editing. Thus, compared to normal humantissues (Teng et al., 1990), mRNA editing mediated byAPOBEC-1 is lost in human carcinomas.
The induction of hepatocellular carcinoma inAPOBEC-1 transgenic animals has been postulated tobe due to aberrant hyperediting of mRNAs for genesthat are involved in the control of cellular growth anddi�erentiation (Yamanaka et al., 1995, 1997). Aberranthyperediting of NAT1, a putative translationalrepressor, may contribute to tumor development inthe APOBEC-1 transgenic animals (Yamanaka et al.,1997). Editing of the natural site in apo B mRNA or ataberrant sites in apo B or NAT 1 mRNA, however,could not be detected in any of the tumors studied.Moreover, editing of C to U at nucleotide position2914 of the NF1 mRNA, a presumptive site forAPOBEC-1 mediated editing in tumors of patients
Figure 7 Complementation of S100 extracts from humancarcinomas with S100 extracts of insect Sf9 cells expressingrecombinant APOBEC-1. S100 extracts (20 mg protein) from tendi�erent human carcinoma specimens were incubated for 2 h at308C with a synthetic apo B RNA in the absence or presence ofinsect Sf9 cell extracts (2 mg protein) containing recombinantAPOBEC-1. As controls (left panel), the Sf9 cell extracts wereincubated with the synthetic apo B RNA in the absence andpresence of Hela-cell nuclear extracts (10 and 20 mg protein,Promega1). The synthetic apo B RNA was analysed for editingby primer extension assay. A representative autoradiograph isshown, the positions of edited and unedited extension productsare indicated
Figure 6 Coexpression of full-length and exon 2 skippedAPOBEC-1 mRNA in human hepatoma HuH-7 cells. Leftpanel: Human hepatoma HuH-7 cells (36105 cells) weretransfected with 0.4 mg of pSVL-hAPOBEC-1 and increasingconcentrations of pSVL-pp36aa (0 ± 1.6 mg) expressing the exon 2skipped form of APOBEC-1. pSVL-b-Gal was added to a totalamount of 2.0 mg DNA. Two days after transfection, the cellularapo B mRNA was analysed for editing by primer extension assay(left panel). Right panel: The bicistronic expression plasmidpSVL-hAPOBEC-1-IRES-pp36aa in which both the full-lengthand the exon 2 skipped APOBEC-1 cDNA are expressed by theuse of the internal ribosome entry site (IRES) of encephalomyo-carditis virus was transfected in human hepatoma HuH-7 cells. Asa control, the same amount of pSVL-hAPOBEC-1 expressingonly full-length human APOBEC-1 was transfected under exactlythe same conditions. Two days after transfection the cellular apoB mRNA was analysed for editing by primer extension assay(right panel)
Absence of mRNA editing in human carcinomasJ Greeve et al
6362

with neuro®bromatosis type 1, or at any other sitebetween nucleotide position 2879 ± 3076 of NF1mRNA could not be found in ten di�erent types ofhuman carcinomas. In hepatocellular and bile ductcarcinoma, in breast carcinoma, in neuroendocrinecarcinoma of the pancreas, in lung adenocarcinoma, inmedullary thyroid carcinoma and in neuroblastoma theabsence of mRNA editing was the consequence of alack of APOBEC-1 expression. In one of the threehepatocellular carcinomas, full-length APOBEC-1mRNA was detected by nested RT ±PCR but not byribonuclease protection assay. This tumor containedconsiderable amounts of the `auxiliary' components ofthe apo B mRNA editing enzyme, but less than 0.25%of its apo B mRNA was edited and hyperediting of apoB or NAT1 mRNA was not found. Similar levels offull-length APOBEC-1 mRNA were found in one ofthe two lung adenocarcinomas that contained highamounts of `auxiliary' components, but no apo BmRNA editing activity. These low levels of full-lengthAPOBEC-1 mRNA do not promote mRNA editingeven in the presence of abundant amounts of `auxiliary'components. Editing of apo B mRNA at the naturalsite C6666 is an excellent functional marker for theexpression of APOBEC-1. At least in carcinomas thatexpress the `auxiliary' components, the lack of apo BmRNA editing proves the absence of active APOBEC-1 and ®rmly excludes its oncogenic potential.
The situation in colorectal and gastric carcinoma ismore complex since these tumors express APOBEC-1mRNA to levels that are detectable by ribonucleaseprotection assay but do not contain the `auxiliary'components. No editing of C6666 in the apo B mRNAor hyperediting of additional cytidine residues in theapo B or NAT1 mRNA could be found in thesetumors. These observations are consistent withprevious ®ndings which demonstrated a requirementof the `auxiliary' components for natural editing andhyperediting in the apo B mRNA (Yamanaka et al.,1996, Sowden et al., 1996). Supplementation ofextracts from various colorectal or gastric carcino-mas with nuclear extracts from Hela-cells thatcontain abundant amounts of these `auxiliary'components, failed to restore apo B mRNA editingactivity in vitro. This experiment indicates that incolorectal and gastric carcinomas APOBEC-1 is notsu�ciently expressed to induce mRNA editing, evenif the `auxiliary' components are abundantly present.Therefore, also in these tumors APOBEC-1 appearsto be unable to exert oncogenic e�ects by mRNAediting.
In colorectal and gastric carcinoma, the exon 2skipped mRNA form was prevailing over the regularlyspliced full length APOBEC-1 mRNA. The ratio ofexon 2 skipped to full-length APOBEC-1 mRNA wasincreased in these tumors compared to normal humanintestine. An overexpression of the exon 2 skippedAPOBEC-1 form has recently been reported in humancolon cancer and the expression of this aberranttranslation product was postulated to be associatedwith an altered growth phenotype (Lee et al., 1998).Coexpression of the exon 2 skipped and the full-lengthAPOBEC-1 cDNA excluded that this polypeptide of36 amino acids interferes and possibly inhibits theediting function of APOBEC-1. According to thesedata, this aberrant translation product has no role in
mRNA editing. The alternate splicing of APOBEC-1mRNA appears to be a peculiar form of posttran-scriptional inactivation that contributes to a presump-tive down-regulation of APOBEC-1 in humancarcinomas. In breast carcinoma and neuroblastoma,only the exon 2 skipped form of APOBEC-1 mRNAcould be detected. Whether this is indicative for aspeci®c regulation of APOBEC-1 mRNA splicingremains to be established.
Besides the exon 2 skipped form, no other mRNAvariant of APOBEC-1 could be detected in colonic orgastric carcinoma. Sequencing of 5'RACE and RT±PCR products con®rmed the cDNA sequence ofAPOBEC-1 in colorectal and gastric carcinomaswithout evidence for point mutations. Thus, a gain offunction for APOBEC-1 in these carcinomas by speci®cmutations appears to be highly unlikely. 5'RACEmapped the transcriptional start sites of APOBEC-1mRNA in normal small intestine and colorectal andgastric carcinoma approximately 25 ± 30 nucleotidesupstream from ATG, similarly as reported previously(Hirano et al., 1997). In human small intestine, twominor 5'RACE products were generated whichextended approximately 1.2 kb upstream. Theseproducts could only be identi®ed by hybridizationand were not visible by ethidium bromide staining. It ismost likely, that these two products represent thetranscriptional start sites as determined by Fujino et al.(1998). According to our data, the usage of thesetranscriptional start sites appears to be very minorcompared to the transcriptional start site close toATG.
In humans, only in small intestine the APOBEC-1mRNA is expressed to levels that are clearlydetectable by Northern blotting (Lau et al., 1994). Asurvey in several fetal, postnatal and adult humantissues demonstrated a more widespread tissueexpression of apo B mRNA editing in humans(Teng et al., 1990). Notably, 50 ± 91% of the apo BmRNA were edited in adult human colon, 490% inadult human stomach and approximately 40% inadult human kidney (Teng et al., 1990). Thus, evenlow level expression of APOBEC-1 which is notdetectable by Northern blotting can promote apo BmRNA editing activity. In most human carcinomas,the loss of apo B mRNA editing is due to absent orvery low expression of APOBEC-1. In colorectal andgastric carcinoma the low level expression ofAPOBEC-1 is further silenced by the loss of the`auxiliary' components that are required for editing.The absence of mRNA editing in human carcinomasdoes not appear to be accidental. The expression ofAPOBEC-1 and mRNA editing most probably doesnot provide a growth advantage for tumors. Aderegulation of APOBEC-1 function with aberranthyperediting of new mRNA targets may be detri-mental for tumor growth and therefore may beselected against in human carcinomas. This investiga-tions indicates that APOBEC-1 is not a naturaloncogene. Whether the loss of apo B mRNA editingis a prerequisite or a consequence of carcinogenesisremains to be investigated. The induction ofhepatocellular carcinoma in APOBEC-1 transgenicanimals, however, does not directly relate to naturalhuman pathology and cannot serve as a model forhepatocarcinogenesis in human patients.
Absence of mRNA editing in human carcinomasJ Greeve et al
6363

Materials and methods
Tissue specimens
Tissue specimens of 28 patients who underwent surgicalresections of primary or metastatic tumors in the Departmentof Surgery, University Hospital Eppendorf, Hamburg,Germany, were used for this study, including colorectalcarcinoma (n=10), hepatocellular carcinoma (n=3), bile ductcarcinoma (n=2), gastric adenocarcinoma (n=3), pancreaticadenocarcinoma (n=2), pancreatic neuroendocrine carcino-ma (n=1), breast carcinoma (n=2), lung adenocarcinoma(n=2), medullary thyroid carcinoma (n=1), soft tissuesarcoma (n=1) and neuroblastoma (n=1). After surgicalresection the tumors were immediately examined by apathologist. Homogeneous and viable portions of theresected tumors which were not required for subsequenthistological assessments or for intraoperative histologicalexamination, but were clearly identi®ed as carcinomatouswere snap-frozen in liquid nitrogen within 10 min of excisionand stored in liquid nitrogen until use. The de®nitehistological diagnosis of each tumor was established by theInstitute of Pathology, University Hospital Eppendorf. Allaspects of this study were approved by the ethics committeeof the medical board of Hamburg (Ethik-Kommission derAÈ rztekammer Hamburg).
RT±PCR of apo B mRNA and APOBEC-1 mRNA
Apo B mRNA was ampli®ed from total RNA by RT±PCRusing oligonucleotides apo B2 (CAC GGA TAT GAT AGTGCT CAT CAA GAC, apo B antisense, nt 6786 ± 6760) andapo B1 (CTG ACT GCT CTC ACA AAA AAG TAT AGA,apo B sense, nt 6552 ± 6578) as described (Greeve et al.,1993). Nested PCR (20 cycles, 1 min at 958C, 1 min at 628C,2 min at 728C) was performed using 1 ml of 1 : 100 dilutedprimary +RT±PCR or 7RT-control PCR and theoligonucleotides apo B15 (TGC CAA AAT CAA CTTTAA TGA AAA AC, apo B sense, nt 6614 ± 6639) and apoB16 (A ATC ATG TAA ATC ATA ACT ATC TTT AATATA CTG A, apo B antisense, nt 6708 ± 6674). Primary andnested RT±PCR of apo B mRNA with their respectivecontrols were analysed by electrophoresis on 1.5% agarosegels. APOBEC-1 mRNA was ampli®ed by RT±PCR from5 mg total RNA as described above using the oligonucleotidesSK1 (AGA GAC AGA GCA CCA TGA CTT CTG AGA,APOBEC-1 sense, nt 714 in the 5'UTR to +13) and SK2(AGC ATT CCC AGC AGG GAC TCC AGG ACA,APOBEC-1 antisense, nt 323 ± 297). Nested RT±PCR ofAPOBEC-1 mRNA was performed with 1 ml of 1 : 100 dilutedprimary +RT±PCR and primary 7RT-control PCR andoligonucleotides SK3 (GGT CCT TCA ACC GGT GACCCC ACT CTG, APOBEC-1 sense, nt 16 ± 42) and SK4(ACC AGG TGA TGG AGC AGC TGA TGG ATG,APOBEC-1 antisense, nt 259 ± 233) for 25 cycles (1 min at958C, 1 min at 648C, 2 min at 728C).
Analysis of apo B mRNA editing
The apo B RT±PCR products were analysed for editing byprimer extension assay as described (Greeve et al., 1991,1993). The unedited 43-nt and the edited 54-nt extensionproducts were separated on an 8% polyacrylamid, 7 M ureasequencing gel and analysed by autoradiography andradiophosphorimaging (Greeve et al., 1996).
Analysis of aberrant hyperediting in the mRNA of apo B and ofthe novel APOBEC-1 target 1 (NAT1)
Apo B mRNA was ampli®ed from two colorectal, twohepatocellular and one gastric carcinoma using oligonucleo-tides apo B17 (TCC AAG ATG CAG TAC TAC TTC CAC,apo B antisense, nt 6864 ± 6841) and apo B1. The RT±PCR
products were cloned and individual recombinant clones wereanalysed by primer extension or DNA sequencing (Greeve etal., 1998). The mRNA of NAT1 was ampli®ed by RT±PCRfrom three colonic and two hepatocellular carcinomas usingthe oligonucleotides NAT-S1 (CAT TTC AGA GCT GGTGAG CAT TTC AG, NAT1 sense, nt 2260 ± 2285) and NAT-S2 (CTG CAT TAC TGG CTT GAA AGA TAG T, NAT1antisense, nt 2758 ± 2734). The RT±PCR products werecloned and 12 individual recombinant NAT1 cDNA clonesfrom each of these ®ve tumors were sequenced.
Analysis of NF1 mRNA editing
The mRNA of NF1 was ampli®ed from total RNA by RT±PCR with oligonucleotides NF1 (CCG AGG CAA CAGCTT GGC CAG TAA A, sense, nt 2823 ± 2847) and NF2(CTG AGG GAA ACG CTG GCT AAC CAC CTG G,antisense, nt 3132 ± 3105) for 30 cycles (0.5 min at 958C,0.5 min at 608C, 0.5 min at 728C) using a Perkin Elmer 2400thermocycler. One ml of 1 : 100 diluted primary RT±PCR wasampli®ed by nested PCR with oligonucleotides NF3 (GACATT CTG TTT CAA GGT ATA TGG TGC, sense, nt2853 ± 2879) and NF4 (ATA AAC AGT GGC ACA CACTTC GAA GTT G, antisense, nt 3103 ± 3076) for 25 cyclesusing the conditions as described above. The RT±PCRproducts were analysed for editing by primer extension assayas described (Skuse et al., 1996). In addition, the RT±PCRproducts for NF 1 mRNA were cloned and sequenced.
Analysis of alternate splicing in the APOBEC-1 mRNA
One ml of 1 : 100 diluted primary +RT±PCR of APOBEC-1mRNA was re-ampli®ed by PCR using the oligonucleotidesSK1 and SK4 (ACC AGG TGA TGG AGC AGC TGA TGGATG, APOBEC-1 antisense, nt 259 ± 233). These PCR-products were separated on a 2% agarose gel and stainedwith ethidium bromide. After transfer to nylon membranes, theblots were prehybridized for 1 h at 608C in DIG-Easy Hyb(Boehringer, Mannheim) and hybridized for 2 h at 588C inDIG-Easy Hyb containing 3 pmol digoxigenin (DIG)-labeledoligonucleotide SK 3. Anti-DIG-alkaline phosphatase con-jugate (Boehringer, Mannheim) was applied for detection ofhybridized oligonucleotides by chemiluminescence. For DIG-labeling the oligonucleotides SK3 and VW2 were 3'tailed usingterminal transferase and DIG-dUTP (Boehringer, Mannheim).Quanti®cation of the two PCR products was performed bydensitometry using NIH Image version 1.55.
RNase protection assay of APOBEC-1 mRNA
APOBEC-1 mRNA was ampli®ed from total RNA of humanintestine (Clontech1) by RT±PCR using oligonucleotidesSK4 and SK3. The PCR-product was cloned and sequenced.Radiolabeled APOBEC-1 antisense RNA was synthesized asdescribed (Greeve et al., 1998). It contained a total of 307bases encompassing 242 bases of the APOBEC-1 sequence (nt259 to 17). Total RNA from normal human small intestine(20 mg, Clontech1) and from tumor tissues (50 mg each) thatwere prepared with Tri-Reagent (Molecular Research CenterInc., Cincinnati, Ohio, USA), were coprecipitated with46104 c.p.m. of the a-32P-labeled antisense RNA probe.Hybridization and RNase digestion with the RibonucleaseProtection Assay II Kit (Ambion Inc., Austin, TX, USA) andthe analysis of the protected RNAs in denaturing poly-acrylamide sequencing gels was performed as described(Greeve et al., 1998).
5'RACE of APOBEC-1 mRNA
Poly(A)+ RNA was prepared from total RNA of normalhuman small intestine (Clontech1) and of one colonic and
Absence of mRNA editing in human carcinomasJ Greeve et al
6364

one gastric carcinoma using oligo (dT) spin columns(Oligotex2) (Qiagen, Hilden, Germany). First- and second-strand cDNA synthesis with subsequent adaptor ligation wasperformed with a commercially available kit (Marathon2
cDNA ampli®cation kit, Clontech, Paolo Alto, USA) asdescribed (Greeve et al., 1998). PCR was performed usingAdvantage KlenTaq-1 Polymerase, the adaptor-speci®cprimer AP1 and one of the following APOBEC-1 antisenseoligonucleotides: SK9 (GAT GTG TGG CGG AAT CGTTTG GTA ATG G, nt 665 ± 639), SK10 (TGT GGC CAGTGA GCT TCA TCC, nt 512 ± 485) and SK2. Nested5'RACE was performed with the adaptor-speci®c primerAP2 and the following APOBEC-1 antisense oligonucleo-tides: SK10, SK2 or SK4. The primary and nested 5'RACEproducts were separated on a 1.0% agarose gel, blotted ontonitrocellulose and hybridized with DIG-labeled oligonucleo-tide SK4 (primary 5'RACE) or VW2 (nested 5'RACE). Theprimary and nested 5'RACE products were cloned andsequenced. The 3'end of APOBEC-1 mRNA was ampli®edfrom the cDNA pool by PCR with oligonucleotides SK6(ACA TTT CTA GAT TCT AAA TGG C, antisense,3'UTR, nt +86 to +64) and SK5 (CGC ACT GGA GCTGCA CTG CAT, APOBEC-1 cDNA sequence, nt 533 ± 553),cloned and sequenced.
Generation of mono- and bicistronic expression vectors forexpression of full-length and exon 2 skipped APOBEC-1 cDNA
The exon 2 skipped APOBEC-1 form was ampli®ed witholigonucleotides SK-BspH1 (ACT CAT GAC TTA TGAGAA AGG AGA AG, APOBEC-1 sense spanning the exon1/exon 2 junction plus a BspH1 restriction site at the 5'end)and SK-Spe1 (GCG ACT AGT GAT GGA GCA GCT GATGGA TG, SK4 plus an Spe1 site at the 5'end) and clonedinto the NcoI-Spe sites of the plasmid pBSKS-IRES contain-ing the internal ribosome entry sequence (IRES) of theencephalomyocarditis virus to generate pBSKS-IRES-pp36aa.The full-length human APOBEC-1 cDNA sequence wasampli®ed by RT±PCR from human total intestinal RNA(Clontech1) using the oligonucleotides SK8 (SK1 plus XbaIsite at the 5'end) and SK6 (ACA TTT CTA GAT TCT AAATGG C, antisense, nt 86 ± 65 of the 3' untranslated region ofAPOBEC-1) and cloned into the pGEM-T easy vector. TheXbaI-fragment containing the full-length human APOBEC-1cDNA was cloned into the XbaI site of the expressionplasmid pSVL (Pharmacia) to generate pSVL-hAPOBEC-1.The BamHI-fragment of pBSKS-IRES-pp36aa was insertedinto the BamHI site of pSVL-hAPOBEC-1 in 3'position ofAPOBEC-1 to generate pSVL-hAPOBEC-1-IRES-pp36aa.pSVL-pp36-aa was generated by inserting a SacI ±SacIIfragment from pGEM-T-easy containing the 245 bp RT±PCR of exon 2 skipped APOBEC-1 mRNA into pSVL. The
control construct pSVL-b-gal was generated by inserting a3.2 kb XbaI-fragment of CMV-b-gal (Promega) into the XbaIsite of pSVL. For transsient transfections FuGENE2 6transfection reagent (Boehringer, Mannheim) was used.
Expression of APOBEC-1 in insect Sf9 cells
The full-length cDNA of rat APOBEC-1 (Greeve et al., 1996)was cloned into the BamHI site of pBacPAK-His3 transfervector (Clontech1) and cotransfected into Sf9 cells withBsu36I digested BacPAK6 viral DNA. Recombinant baculo-virus was plaque-puri®ed and ampli®ed on Sf9 cells grown inmonolayers. For expression of APOBEC-1 protein, 2.56108
Sf9 cells were infected with the APOBEC-1 recombinantbaculovirus at a MOI of 20 for 80 h. The Sf9 cells wereresuspended in 5 ml bu�er A (20 mM HEPES, pH 8.0, 5 mM
b-mercaptoethanol) and homogenized with 20 strokes of aDounce homogenizer using a tight ®tting pestel. Thehomogenate was adjusted to 100 mM NaCl, 20% glycerol,centrifuged for 30 min at 100 000 g using a SW40 rotor in aBeckman ultracentrifuge and stored at 7808.
In vitro editing reactions using S100 extracts from humancarcinomas
200 mg of tumor tissue was grinded in liquid nitrogen andresuspended in 500 ml of 20 mM HEPES, pH 7.4, 100 mM
KCl, 1 mM DTT, containing leupeptin, pepstatin, aprotinin,benzamidine and phenylmethyl-sulphonyl ¯uoride, each at aconcentration of 10 mg/ml. After centrifugation at 12 000 gfor 10 min, the supernatant was centrifuged for 30 min at100 000 g in a Beckman TS 100 centrifuge and stored at7808C. For in vitro editing reactions, an apo B RNAtranscript of 448 bases was used (Greeve et al., 1991). Twentymg protein of the S100 extract from the tumor tissues wasincubated without and with 2 mg of Sf9 cell-extract contain-ing recombinant APOBEC-1 and analysed for in vitro editing(Greeve et al., 1991). Nuclear extracts of Hela-cells contain-ing high amounts of the `auxiliary' components but no invitro editing activity were obtained from Promega1.
AcknowledgementsThe contribution of Dr S von der Kammer during the earlycharacterization of mRNA editing and APOBEC-1 expres-sion in human carcinomas is gratefully acknowledged. Thiswork was supported by Bundesministerium fuÈ r Bildungund Forschung (BMBF), 01KV9509/0 (to J Greeve) andDeutsche Forschungsgemeinschaft (DFG), SFB 545, A6 (toJ Greeve).
References
Anant S, MacGinnitie AJ and Davidson NO. (1995). J. Biol.Chem., 270, 14762 ± 14767.
Cappione AJ, French BL and Skuse GR. (1997). Am. J.Hum. Genet., 60, 305 ± 312.
Chen SH, Habib G, Yang CY, Gu ZW, Lee BR, Weng SA,Silbermann SR, Cai SJ, Deslypere JP, Rosseneu M, GottoAM, Li WH, and Chan L. (1987). Science, 238, 363 ± 366.
Fujino T, Navaratnam N and Scott J. (1998). Genomics, 47,266 ± 275.
Giannoni F, Bonen DK, Funahashi T, Hadjiagapiou C,Burant CF and Davidson NO. (1994). J. Biol. Chem., 269,5932 ± 5936.
Greeve J, Navaratnam N and Scott J. (1991). Nucl. AcidsRes., 19, 3569 ± 3576.
Greeve J, Altkemper I, Dieterich JH, Greten H and WindlerE. (1993). J. Lipid Res., 34, 1367 ± 1382.
Greeve J, Jona VK, Chowdhury NR, Horwitz MS andChowdhury JR. (1996). J. Lipid Res., 37, 2001 ± 2017.
Greeve J, Axelos D, Welker S, Schipper M and Greten H.(1998). Arterioscl. Thromb. Vasc. Biol., 18, 1079 ± 1092.
Hirano KI, Min J, Funahashi T, Baunoch DA, andDavidson NO. (1997). J. Lipid Res., 38, 847 ± 859.
Lau PP, Zhu HJ, Baldini A, Charnsangavei C and Chan L.(1994). Proc. Natl. Acad. Sci. USA, 91, 8522 ± 8526.
Lee RM, Hirano KI, Ananat S, Baunoch D and DavidsonNO. (1998). Gastroenterology, 115, 1096 ± 1103.
Nakamuta M, Oka K, Krushkal J, Kobayashi K, YamamotoM, Li WH and Chan L. (1995). J. Biol. Chem., 270,13042 ± 13056.
Navaratnam N, Morrison JR, Battacharya S, Patel D,Funahashi T, Giannoni F, Teng BB, Davidson NO andScott J. (1993). J. Biol. Chem., 268, 20709 ± 20712.
Absence of mRNA editing in human carcinomasJ Greeve et al
6365

Navaratnam N, Bhattacharya S, Fu®no T, Patel D, JaumuzAL and Scott J. (1995). Cell, 81, 187 ± 195.
Maas S, Melcher T and Seeburg P. (1997). Curr. Opin. CellBiol., 9, 343 ± 349.
Powell LM, Wallis SC, Pease RJ, Edwards YH, Knott TYand Scott J. (1987). Cell, 50, 831 ± 840.
Powell-Braxton LM, Venianti M, Latvala RD, Hirano KI,Won WB, Ross J, Dybdal N, Zlot CH, Young SG andDavidson NO. (1998). Nature Med., 4, 934 ± 938.
Qian X, Balestra ME and Innerarity TL. (1997). J. Biol.Chem., 272, 18060 ± 18070.
Scott J. (1995). Cell, 81, 833 ± 836.Shah RR, Knott TJ, Legros JE, Navaratnam N, Greeve J
and Scott J. (1991). J. Biol. Chem., 266, 16301 ± 16304.Sharma PM, Bowman M, Madden SL, Rauscher FJ and
Sukumar S. (1994). Genes Dev., 8, 720 ± 731.Smith HC, Kuo SR, Backus JW, Harris SG, Sparks CE and
Sparks JD. (1991). Proc. Natl. Acad. Sci. USA, 88, 1489 ±1493.
Simpson L and Thiemann OH. (1995). Cell, 81, 837 ± 840.
Skuse GR, Cappione AJ, Sowden M, Metheny LJ and SmithHC. (1996). Nucl. Acids Res., 24, 478 ± 486.
Sommer B, KoÈ hler M, Sprengel R and Seeburg PH. (1991).Cell, 67, 11 ± 19.
Sowden M, Hamm JK and Smith HC. (1996). J. Biol. Chem.,271, 3011 ± 3017.
Teng BB, Verp M, Salomon J and Davidson NO. (1990). J.Biol. Chem., 265, 20616 ± 20620.
Teng BB, Burand CF and Davidson NO. (1993). Science,260, 1816 ± 1819.
Yamanaka S, Poksay KS, Balestra ME, Zeng GQ andInnerarity TL. (1994). J. Biol. Chem., 269, 21725 ± 21734.
Yamanaka S, Balestra ME, Ferrell LD, Fan J, Arnold KS,Taylor S, Taylor JM and Innerarity TL. (1995). Proc.Natl. Acad. Sci. USA, 92, 8483 ± 8487.
Yamanaka S, Poksay KS, Driscoll DM and Innerarity TL.(1996). J. Biol. Chem., 271, 11506 ± 11510.
Yamanaka S, Poksay KS, Arnold KS, and Innerarity TL.(1997). Genes Dev., 11, 321 ± 333.
Absence of mRNA editing in human carcinomasJ Greeve et al
6366
Related Documents











